importance of serratia protease in the pathogenesis...
TRANSCRIPT

INFECTION AND IMMUNITY, Apr. 1983, p. 113-1190019-9567/83/040113-07$02.00/0Copyright © 1983, American Society for Microbiology
Vol. 40, No. 1
Importance of Serratia Protease in the Pathogenesis ofExperimental Serratia marcescens P'neumonia
DAVID M. LYERLY AND ARNOLD S. KREGER*
Department ofMicrobiology and Immunology, The Bowman Gray School of Medicine of Wake Forest
University, Winston-Salem, North Carolina 27103
Received 26 October 1982/Accepted 5 January 1983
The results of studies to evaluate the possible importance of serratia proteasesin the development of experimental Serratia marcescens pneumonia revealed thefollowing. (i) Administration of a highly purified serratia protease to the lungs ofguinea pigs and mice resulted in extensive pulmonary edema and hemorrhagesimilar to that observed in animals having an experimentally induced, acuteserratia pneumonia. (ii) Guinea pigs subcutaneously vaccinated with the proteasedeveloped low levels of antiprotease antibodies and were partially protectedagainst serratia pneumonia, as demonstrated by a significant increase in survivaltime. Mice intranasally vaccinated with the protease also developed antiproteaseantibodies and were protected against serratia pneumonia, as demonstrated by a
significant increase in survival time and an increase in the number of survivors.(iii) Serratia protease was detected in lung tissue extracts prepared from the lungsof guinea pigs dying of serratia pneumonia. Our findings support the idea thatserratia protease(s) is involved in the pathogenesis of experimental serratiapneumonia.
Serratia marcescens is an important opportu-nistic pathogen capable of producing pneumoniain humans and experimental animals (2, 9, 10,14, 15). Previous findings in our laboratory (8;A. S. Kreger, Abstr. Annu. Meet. Am. Soc.Microbiol., M288, p. 114, 1974) revealed thatintratracheal administration of serratia proteasepreparations elicited lung damage in rabbits, andthese findings suggested the idea that proteaseselaborated by the bacterium may be involved inthe pathogenesis of serratia pneumonia. There-fore, studies were performed, using a highlypurified serratia protease preparation (8), toevaluate the importance of serratia proteases inserratia-induced lung disease. The experimentsincluded (i) studies to compare the gross appear-ance of lung damage elicited by the serratiaprotease and by experimental serratia pneumo-nia, (ii) studies to determine whether vaccina-tion against the serratia protease could elicitactive immunity against experimental serratiapneumonia, and (iii) studies to determine wheth-er the serratia protease could be detected in lungtissues obtained from guinea pigs dying of serra-tia pneumonia. The results presented in thispaper support the idea that serratia protease(s) isinvolved in the pathogenesis of serratia-inducedpneumonia.
MATERIALS AND METHODS
Growth of bacterium and purification of protease. S.marcescens BG, obtained from the culture collectionof the Department of Microbiology and Immunologyof The Bowman Gray School of Medicine, was culti-vated in tryptone-yeast extract-glucose broth, and thebacterial extracellular protease was purified to homo-geneity as previously described (8).
Production of experimental serratia pneumonia andserratia protease-induced lung damage in guinea pigsand mice. Dilutions of bacterial suspensions wereprepared in 0.85% saline from washed cell suspensionsinitially containing 1010 bacteria per ml. Experimentalserratia pneumonia was produced in Hartley strainguinea pigs (400 to 500 g; Perfection Breeders, Doug-lassville, Pa.) by intratracheally injecting samples (0.5ml) of the diluted suspensions into animals whosetracheas were surgically exposed after anesthesia withsodium pentobarbital. After the injection, the incisionwas closed with surgical wound clips. Experimentalserratia pneumonia was produced in Dub:(ICR) mice(25 g; Dominion Laboratories, Dublin, Va.) by apply-ing samples (50 ,ul) of washed, diluted bacterial sus-pensions onto the external nares of mice anesthetizedwith sodium pentobarbital. The mice readily aspiratedthe sample when challenged in this manner.
Samples (0.5 ml) of sterile serratia protease prepara-tions (12.5 to 200 ,g/ml in 0.05 M Tris-hydrochloride,pH 7.5) were intratracheally injected into guinea pigsas described above. Mice were challenged, as de-
113
on April 12, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

114 LYERLY AND KREGER
scribed above, with samples (50 p1I) of sterile serratiaprotease preparations (25 to 600 ,ug/ml of 0.05 M Tris-hydrochloride, pH 7.5). Control preparations consist-ed of sterile 0.85% saline buffered with 0.02 M phos-phate (pH 7) (PBS), heat-killed bacteria (15 min at100°C), and heat-inactivated serratia protease (15 minat 100°C).The number of bacteria in the lungs of animals
which died of serratia pneumonia was determined byviable plate counts with dilutions of lung homoge-nates. Guinea pig lungs were homogenized in 10 ml ofPBS in a Sorvall Omni-Mixer. Mouse lungs werehomogenized in 1 ml of PBS with a Ten Broeckhomogenizer.
Production of experimental serratia pneumonia inguinea pigs and mice vaccinated with the serratia prote-ase. Guinea pigs were injected subcutaneously with 0.2ml of an anhydrous oil vaccine (3) containing 0.15 mgof lyophilized serratia protease in incomplete Freundadjuvant, and after 2 weeks they were vaccinatedagain with the same preparation. The animals werebled 1 week after the second injection, and the serawere examined, using the antiprotease antibody fluo-rescent immunoassay described below, for the pres-ence of antibodies against the serratia protease. Oneweek later, the guinea pigs were injected intratracheal-ly with 109 bacteria (2 median [50%] lethal doses[LD50]) and observed for 48 h to note any reduction inthe severity of serratia pneumonia.Mice were anesthetized with sodium pentobarbital
and vaccinated weekly for 3 weeks by administeringserratia protease samples (10 ,ug in 50 RI of PBS) ontothe external nares and allowing the animals to aspiratethe fluid. One week after the final vaccination, lungtissue extracts were obtained from nine of the mice byhomogenizing the lungs in 1 ml of PBS and removingthe insoluble material by centrifugation. Samples ofthe lung tissue extracts were examined, using theantiprotease antibody fluorescent immunoassay de-scribed below, for the presence of antibody against theprotease. The remaining mice were challenged intrana-sally with 3.5 x 10' bacteria (10 LD50) or 7 x 107bacteria (2 LD50) and observed for 48 h to note anyreduction in the severity of the serratia pneumonia.
Fluorescent immunoassay to detect serratia proteaseantibodies in guinea pig sera and mouse lung tissue.Serratia protease was coupled to a commercially avail-able Immunobead reagent (Bio-Rad Laboratories) asrecommended by the manufacturer. Guinea pig seraand mouse lung tissue extracts were obtained fromanimals vaccinated, as described above, with serratiaprotease. Samples (100 ,ul) of these specimens wereincubated with the serratia protease-Immunobead rea-gent, and antibodies bound by the beads were detectedby using fluorescein-labeled conjugates. Rabbit anti-guinea pig gamma globulin-fluorescein conjugate (Nu-tritional Biochemical Corp., Cleveland, Ohio; 40 p.1 ofa 1:2 dilution) was used to detect antibodies in theguinea pig sera. Rabbit anti-mouse immunoglobulin G(IgG)-fluorescein conjugate (Miles-Yeda, Ltd., Elk-hart, Ind.; 40 ,u1 of a 1:10 dilution) and rabbit anti-mouse IgA-fluorescein conjugate (Bionetics Corp.,Kensington, Md.; 40 ,ul of a 1:4 dilution) were used todetect antibodies in the mouse lung tissue extracts.The assay was performed under the conditions de-scribed in a commercially available quantitative immu-nofluorescent assay for human gamma globulin (Bio-
Rad Laboratories). Controls included (i) sera and lungtissue extracts from nonvaccinated animals and (ii)incubation of the serratia protease-Immunobead rea-gent with the conjugate preparations to demonstratethe absence of nonspecific interaction between theconjugate and Immunobead reagent.
Detection of serratia protease in lung tissues fromguinea pigs having serratia pneumonia. One group of 10guinea pigs were injected intratracheally with 109bacteria. A second group of guinea pigs, serving ascontrol animals, were injected intratracheally withPBS. Immediately after the guinea pigs died or weresacrificed, the lungs were excised and lavaged with 20ml of PBS. Lung tissue extracts were prepared fromthe lavaged lungs by homogenizing the lungs in 10 mlof PBS and removing the insoluble material by centrif-ugation. Lung lavage fluids and tissue extracts werelyophilized, and samples (2 mg in 100 p.) were exam-ined as described below.
(i) Protease activity against fluorescein-labeled casein.Protease activity was assayed with a casein substratepreparation obtained by labeling vitamin-free, solublecasein (Teklad Test Diets, Madison, Wis.) with fluo-rescein, using a procedure previously described forpreparing fluorescein-labeled IgG (6). The reactionmixture, containing 0.5 ml of an aqueous solution offluorescein-labeled casein (5 mg/ml), 0.5 ml of 0.2 MTris-hydrochloride (pH 7.5), 0.5 ml of glass-distilledwater, and 100 p.l of sample, was incubated for 10 minat 37°C. The reaction was stopped by the addition of3.5 ml of 5% trichloroacetic acid, and the precipitatewas removed by centrifugation and filtration throughpaper. The pH of the solution containing the fluoresce-in-labeled, trichloroacetic acid-soluble peptides wastitrated to pH 7.5 with 1 N NaOH, and the fluores-cence was measured as previously described.
(ii) Immunologically specific assay for serratia prote-ase. A double-antibody fluorescent immunoassay spe-cific for the serratia protease was developed usingmonospecific antiserum raised against the enzyme.The monospecific antiserum was prepared by injectingNew Zealand white rabbits subcutaneously, every 2weeks, with 0.4 ml of an anhydrous oil vaccine (3)containing 1 mg of the lyophilized serratia protease inincomplete Freund adjuvant. Rabbits were bled asepti-cally for serum samples 7 days after the fourth injec-tion, and acceptable precipitating antibody titers weredemonstrated by using Ouchterlony double immuno-diffusion (acceptable antibody titers were noted byprecipitin line formation with 0.00078 to 0.00156%antigen). Rabbits were exsanguinated 1 week later,and the sera were pooled and lyophilized. The mono-specificity of the antiserum was examined by crossedimmunoelectrophoresis using methods previously de-scribed (8). Briefly, samples (18 p.1 containing 200 p.gof a crude serratia protease preparation [8]) weresubjected to electrophoresis in the first dimension,followed by electrophoresis in the second dimensionagainst antiserum prepared against crude serratia pro-tease preparations (8) or antiserum prepared, as de-scribed above, against the highly purified serratiaprotease. The monospecificity of the antiserum wasdetermined by comparing the number of precipitinarcs obtained to those obtained with the antiseraagainst the crude and highly purified serratia proteasepreparations. The IgG fraction of the monospecificantiserum was obtained by chromatography on a col-
INFECT. IMMUN.
on April 12, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

SERRATIA PROTEASE AND SERRATIA PNEUMONIA 115
3001
w
z0 200ILnw
0
L-iw
! 100J
w
n
IP'O.O5)
(P'co.05l
lP'O.O5,
ip- .5
m
A B C
FIG. 1. Detection of serratia protease antibodies invaccinated animals. Samples obtained from nonvac-cinated (E) and vaccinated (0) animals were incubat-ed with Immunobeads coated with serratia protease,and antibodies bound by the beads were detected withfluorescein-labeled anti-immunoglobulin. (A) Analysisof sera from guinea pigs subcutaneously vaccinatedwith serratia protease. Antiprotease antibodies weredetected with anti-guinea pig immunoglobulin-fluores-cein conjugate. (B) IgG analysis of lung tissue extractsfrom mice intranasally vaccinated with serratia prote-ase. Antiprotease antibodies were detected with anti-mouse IgG-fluorescein conjugate. (C) IgA analysis oflung tissue extracts from mice intranasally/ vaccinatedwith serratia protease. Antiprotease antibodies weredetected with anti-mouse IgA-fluorescein conjugate.Significant differences are shown.
umn ofDEAE Affi-Gel Blue (Bio-Rad Laboratories) asrecommended by the manufacturer. A portion of thepurified IgG was coupled to the Immunobead reagent,as recommended by the manufacturer, for the prepara-tion of antiprotease IgG-Immunobead reagent, andpart of the IgG preparation was used to prepareantiprotease IgG-fluorescein conjugate, using previ-ously described procedures (6). The 495 nm/280 nmabsorbance ratio of the conjugate was ca. 0.5.The immunoassays were performed by incubating
samples (2 mg in 100 ,ul) of lavage fluids and lung tissueextracts with antiprotease IgG-Immunobead reagent.The beads were washed, and antiprotease IgG-fluores-cein conjugate (100 ,ug in 40 ,ul) was used to detectserratia protease bound by the beads. The assay wasperformed under the conditions described in a com-mercially available quantitative immunofluorescentassay for human gamma globulin (Bio-Rad Labora-tories).
Statistical analysis of data. Median lethal doses
(LD50) were determined by the method of Reed andMuench (13). The production of serratia proteaseantibodies by guinea pigs and mice was analyzed(Student's t test) by comparing fluorescence measure-
ments in vaccinated versus nonvaccinated animals.Significant differences in the survival of vaccinatedversus nonvaccinated animals were determined byusing the chi-square distribution method with theYates correction for continuity. The presence of serra-
tia protease in guinea pig lungs was analyzed (Stu-dent's t test) by comparing fluorescence measure-
ments in animals receiving viable bacteria versusanimals receiving PBS.
RESULTS AND DISCUSSIONPrevious investigators have reported that S.
marcescens is an opportunistic pathogen capa-ble of producing pneumonia in humans andexperimental animals (2, 9, 10, 14, 15); however,the mechanisms by which the bacterium pro-duces lung disease have not been determined.The present study was undertaken to evaluatethe importance of serratia protease(s) in thepathogenesis of lethal, experimental serratiapneumonia.The purpose of the first series of experiments
was to compare the gross appearance of lungdamage produced by experimental serratiapneumonia with lung damage observed after theintrapulmonary administration of serratia prote-ase. Postmortem examination of the lungs ofguinea pigs which were intratracheally injectedwith 2 LD50 of bacteria (109 bacteria) revealededema, brownish discoloration of the lung sur-face, and confluent and focal pulmonary hemor-rhage 12 to 48 h postinfection. Mice whichaspirated suspensions of the bacterium also de-veloped lethal serratia pneumonia within 24 hpostchallenge, and intranasal challenge with 2LD50 of bacteria (7 x 107 bacteria) resulted inextensive hemorrhage similar to that observed inthe guinea pig model. In both animal models, thenumber of bacteria recovered from the lungswas ca. 10-fold greater than the initial challenge
TABLE 1. Active immunization of guinea pigsagainst serratia pneumonia by vaccination with
serratia proteaseaNo. dead/no. injected
Treatment16 h 24 h 36 h 48 h
Control' 5/12 6/12 7/12 7/12Actively 1/14d 3/14 3/14 5/14immunizedca Results were observed after the intratracheal in-
jection of 109 bacteria (2 LD50)-I Guinea pigs were subcutaneously injected with
incomplete Freund adjuvant.c Guinea pigs were subcutaneously vaccinated with
the serratia protease.d p < 0.05.
UK I 9%I I
VOL. 40, 1983
on April 12, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

116 LYERLY AND KREGER
TABLE 2. Active immunization of mice againstserratia pneumonia by vaccination with serratia
proteaseaNo. dead/no. challenged
Treatment Challengeb10h 24h 48h
Controlc 10 LD5o 16/18 17/18 18/18Actively 10 LD50 6/16'e 14/16 16/16immunizedd
Controlc 2 LD50 11/21 18/21 21/21Actively 2 LD50 1/17e 6/17e 7/17eimmunizedda Results were observed after the intranasal admin-
istration of bacteria.b Challenge doses of 10 LD50 and 2 LD50 represent
ca. 3.5 x 108 and ca. 7 x 107 bacteria, respectively.Mice were administered PBS intranasally.
d Mice were intranasally vaccinated with the serra-tia protease.eP < 0.005.
dose. In addition, S. marcescens was culturedfrom the blood of dying or dead animals, thusindicating that the death of these animals mayhave resulted from septicemia as well as fromextensive pulmonary damage. Guinea pigs intra-tracheally injected with sublethal doses (10 to100 ,ug) of serratia protease had confluent andfocal pulmonary hemorrhage by 24 h postinjec-tion, which was similar to that observed duringexperimental serratia pneumonia. Injection ofless than 10 ,ug of protease caused mottling andbrownish discoloration of the lungs. The mini-mum lethal intranasal dose of serratia proteasefor mice (12 to 24 h postinjection) was 25 to 30Rig; this dose caused confluent pulmonary hem-orrhage similar to that observed in mice whichdied after intranasal challenge with the viablebacterium. The lipopolysaccharide endotoxin ofS. marcescens is known to cause corneal tissuedamage (1); therefore, to investigate the possi-bility that serratia lipopolysaccharide caused thedamage observed during experimental serratiapneumonia, we challenged guinea pigs and micewith an amount of heat-killed bacteria whichwas 20-fold greater than the LD50 of the viablebacterium. The heat-killed bacteria were notlethal for the animals and caused only minordiscoloration of the lungs. The gross pathologi-cal findings that we observed in our experimen-tal serratia pneumonia models and after theadministration of serratia protease are similar tothose previously reported by Meltz and Grieco(10), who noted pulmonary tissue necrosis andhemorrhage, both by gross observation and bylight microscopy, in a human who died of serra-tia pneumonia.The second series of studies were performed
to determine whether guinea pigs vaccinated
subcutaneously or mice vaccinated intranasallywith serratia protease were actively immunizedagainst serratia pneumonia. After vaccination,serum samples from the guinea pigs and lungtissue extracts from the mice were examined forantiprotease antibodies, using an antiproteaseantibody fluorescent immunoassay. Guinea pigsvaccinated with the serratia protease producedlow levels of antiprotease antibodies (Fig. 1).Mice vaccinated with serratia protease also pro-duced antibodies against the enzyme, and, byusing specific anti-immunoglobulin-fluoresceinconjugates, we determined that the mouse anti-protease antibodies consisted of IgG and IgA(Fig. 1). Although the IgG may have been ofhematogenous origin and, therefore, may havebeen released into the extract during the grind-ing of the lung tissue, the presence of IgAantibodies indicated that antiprotease antibodieswere present on the mucosal surface of the lung.Vaccinated guinea pigs challenged with 2 LD50of bacteria survived significantly longer thannonvaccinated animals (Table 1). Vaccinatedmice also showed a significant increase in sur-vival time, as well as a significant increase in thenumber of survivors (Table 2). Our results aresimilar to those of Homma et al. (4), whoreported that mink vaccinated subcutaneouslywith Formalin-inactivated preparations of Pseu-domonas aeruginosa proteases produced anti-
w 20zw
w
D-JIL 1Ow
-Jw
20 40SERRATIA PROTEASE (ng)
FIG. 2. Standard curve for protease assay withfluorescein-labeled casein substrate. Samples of serra-tia protease were incubated with fluorescein-labeledcasein for 10 min at 37°C, and the release of fluoresce-in-labeled, trichloroacetic acid-soluble peptides wasmeasured with a spectrophotofluorometer. The cor-rected relative fluorescence was determined by sub-tracting the fluorescence of supernatant fluids ob-tained from incubation mixtures without enzyme fromthe fluorescence of supernatant fluids obtained fromincubation mixtures containing protease.
INFECT. IMMUN.
on April 12, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

SERRATIA PROTEASE AND SERRATIA PNEUMONIA 117
A
3001-
LLLI-
+' 200w
0
zw
LI)
w
o 1000
-j
lLw
!a n-i
icCKx
(P.C0.05)
(P.c0.05t
Us Im -i
30 - B(P.c 0.05)
0 om"
LUNG LUNGLAVAGE TISSUEFLUID EXTRACTS
FIG. 3. Detection of serratia protease in tof guinea pigs dying from experimental serratmonia. Control (E) and test (0) guinea piintratracheally injected with PBS and 2 LD50na (109 bacteria), respectively. Lung lavage flilung tissue extracts were prepared immediatthe death or exsanguination of the animals, aples (2 mg) were examined as described in theDetection of in vivo protease activity againstcein-labeled casein. (B) Detection of in vivoprotease by double-antibody fluorescent imsay. Significant differences are shown.
protease antibodies and were protectedacute, lethal, hemorrhagic pseudomonamonia.Our third series of experiments were d
to determine whether serratia protease edetected in lung lavage fluids and lunextracts from guinea pigs with expeiserratia pneumonia, thus demonstratingratia protease is produced in situ duringdevelopment. In our first group of stu4developed a protease assay which utili2rescein-labeled casein as the substrate.geneous preparations of serratia proteaed a dose-dependent response in this as,2), and, by using a fluorescein-labeled sithe sensitivity of the assay was increathat 10 to 20 ng of serratia proteasecould be detected. We found that lunfluids and lung tissue extracts from gui
dying of serratia pneumonia contained levels ofprotease activity significantly higher than thosein lung samples from guinea pigs injected withPBS (Fig. 3A). In view of the fact, however, thatthe protease assay used in the first group ofstudies was not specific for serratia protease,and that the elevated levels of activity we ob-served may have been due, at least in part, toendogenous proteases released during the dis-ease by infiltrating macrophages and polymor-phonuclear leukocytes, the second group ofstudies were done with a double-antibody fluo-rescent immunoassay utilizing IgG from antise-rum specific for the serratia protease (Fig. 4).Homogeneous preparations of serratia proteaseelicited a dose-dependent response in the assay(Fig. 5), with 50 to 60 ng of protease being thelowest detectable amount. Although Miyata etal. (12) previously described a double-antibodyradioimmunoassay capable of detecting smallamounts (1 to 10 ng) of serratia protease, the
A
-he lungstia pneu-igs wereof bacte-luids and /eiyw aner ,/.eiy atterand sam-text. (A)t fluores-serratia
kmunoas-
[againstiS pneu-
lesignedcould beIg tissuerimentalthat ser-disease
dies, wezed fluo-. Homo-,se elicit-say (Fig.ubstrate,sed suchactivity
kg lavageinea pigs
B
Fkle
FIG. 4. Demonstration of the monospecificity ofantiserum against serratia protease by crossed immu-noelectrophoresis. Sample wells in both (A) and (B)contained 200 ,ug of a crude serratia protease prepara-tion. (A) Second dimension of gel contained 0.5 ml ofantiserum against crude serratia protease preparation.At least 10 to 11 immunoprecipitin arcs are visible. (B)Second dimension of gel contained 0.5 ml of antiserumagainst purified serratia protease preparation. Onlyone immunoprecipitin arc is visible.
VOL. 40, 1983
on April 12, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
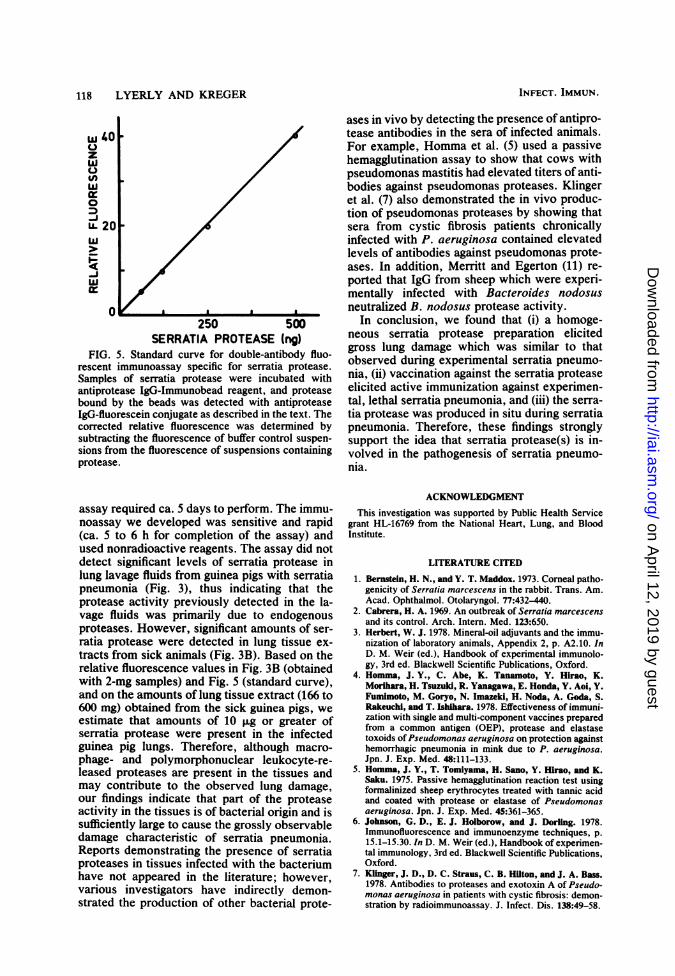
118 LYERLY AND KREGER
W 4Uz
0
"-20
uO20 . .
0
250 500SERRATIA PROTEASE (ng)
FIG. 5. Standard curve for double-antibody fluo-rescent immunoassay specific for serratia protease.Samples of serratia protease were incubated withantiprotease IgG-Immunobead reagent, and proteasebound by the beads was detected with antiproteaseIgG-fluorescein conjugate as described in the text. Thecorrected relative fluorescence was determined bysubtracting the fluorescence of buffer control suspen-sions from the fluorescence of suspensions containingprotease.
assay required ca. 5 days to perform. The immu-noassay we developed was sensitive and rapid(ca. 5 to 6 h for completion of the assay) andused nonradioactive reagents. The assay did notdetect significant levels of serratia protease inlung lavage fluids from guinea pigs with serratiapneumonia (Fig. 3), thus indicating that theprotease activity previously detected in the la-vage fluids was primarily due to endogenousproteases. However, significant amounts of ser-ratia protease were detected in lung tissue ex-tracts from sick animals (Fig. 3B). Based on therelative fluorescence values in Fig. 3B (obtainedwith 2-mg samples) and Fig. 5 (standard curve),and on the amounts of lung tissue extract (166 to600 mg) obtained from the sick guinea pigs, weestimate that amounts of 10 F.g or greater ofserratia protease were present in the infectedguinea pig lungs. Therefore, although macro-phage- and polymorphonuclear leukocyte-re-leased proteases are present in the tissues andmay contribute to the observed lung damage,our findings indicate that part of the proteaseactivity in the tissues is of bacterial origin and issufficiently large to cause the grossly observabledamage characteristic of serratia pneumonia.Reports demonstrating the presence of serratiaproteases in tissues infected with the bacteriumhave not appeared in the literature; however,various investigators have indirectly demon-strated the production of other bacterial prote-
ases in vivo by detecting the presence of antipro-tease antibodies in the sera of infected animals.For example, Homma et al. (5) used a passivehemagglutination assay to show that cows withpseudomonas mastitis had elevated titers of anti-bodies against pseudomonas proteases. Klingeret al. (7) also demonstrated the in vivo produc-tion of pseudomonas proteases by showing thatsera from cystic fibrosis patients chronicallyinfected with P. aeruginosa contained elevatedlevels of antibodies against pseudomonas prote-ases. In addition, Merritt and Egerton (11) re-ported that IgG from sheep which were experi-mentally infected with Bacteroides nodosusneutralized B. nodosus protease activity.
In conclusion, we found that (i) a homoge-neous serratia protease preparation elicitedgross lung damage which was similar to thatobserved during experimental serratia pneumo-nia, (ii) vaccination against the serratia proteaseelicited active immunization against experimen-tal, lethal serratia pneumonia, and (iii) the serra-tia protease was produced in situ during serratiapneumonia. Therefore, these findings stronglysupport the idea that serratia protease(s) is in-volved in the pathogenesis of serratia pneumo-nia.
ACKNOWLEDGMENT
This investigation was supported by Public Health Servicegrant HL-16769 from the National Heart, Lung, and BloodInstitute.
LITERATURE CITED
1. Bernstein, H. N., and Y. T. Maddox. 1973. Corneal patho-genicity of Serratia marcescens in the rabbit. Trans. Am.Acad. Ophthalmol. Otolaryngol. 77:432-440.
2. Cabrera, H. A. 1969. An outbreak of Serratia marcescensand its control. Arch. Intern. Med. 123:650.
3. Herbert, W. J. 1978. Mineral-oil adjuvants and the immu-nization of laboratory animals, Appendix 2, p. A2.10. InD. M. Weir (ed.), Handbook of experimental immunolo-gy, 3rd ed. Blackwell Scientific Publications, Oxford.
4. Homma, J. Y., C. Abe, K. Tanamoto, Y. Hirao, K.Morihara, H. Tsuzuki, R. Yanagawa, E. Honda, Y. Aol, Y.Fumimoto, M. Goryo, N. Imazeki, H. Noda, A. Goda, S.Rakeuchl, and T. Ishihara. 1978. Effectiveness of immuni-zation with single and multi-component vaccines preparedfrom a common antigen (OEP), protease and elastasetoxoids ofPseudomonas aeruginosa on protection againsthemorrhagic pneumonia in mink due to P. aeruginosa.Jpn. J. Exp. Med. 48:111-133.
5. Homma, J. Y., T. Tomiyama, H. Sano, Y. Hirao, and K.Saku. 1975. Passive hemagglutination reaction test usingformalinized sheep erythrocytes treated with tannic acidand coated with protease or elastase of Pseudomonasaeruginosa. Jpn. J. Exp. Med. 45:361-365.
6. Johnson, G. D., E. J. Holborow, and J. Dorling. 1978.Immunofluorescence and immunoenzyme techniques, p.15.1-15.30. In D. M. Weir (ed.), Handbook of experimen-tal immunology, 3rd ed. Blackwell Scientific Publications,Oxford.
7. Klinger, J. D., D. C. Straus, C. B. Hilton, and J. A. Bass.1978. Antibodies to proteases and exotoxin A of Pseudo-monas aeruginosa in patients with cystic fibrosis: demon-stration by radioimmunoassay. J. Infect. Dis. 138:49-58.
INFECT. IMMUN.
on April 12, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

SERRATIA PROTEASE AND SERRATIA PNEUMONIA 119
8. Lyerly, D., and A. Kreger. 1979. Purification and charac-terization of a Serratia marcescens metalloprotease. In-fect. Immun. 24:411-421.
9. McCormack, R. C., and C. M. Kunin. 1966. Control of a
single source nursery epidemic due to Serratia marces-
cens. Pediatrics 37:750.10. Meltz, D. J., and M. H. Grieco. 1973. Characteristics of
Serratia marcescens pneumonia. Arch. Intern. Med.132:359-364.
11. Merritt, G. C., and J. R. Egerton. 1978. IgG, and IgG2immunoglobulins to Bacteroides (Fusiformis) nodosusprotease in infected and immunized sheep. Infect. Im-mun. 22:1-4.
12. Miyata, K., M. Tsuda, and K. Tomoda. 1980. Determina-tion of Serratia protease by radioimmunoassay. Anal.Biochem. 101:332-338.
13. Reed, L. J., and H. Muench. 1938. A simple method ofestimating fifty per cent endpoints. Am. J. Hyg. 27:493-497.
14. Sanders, C. V., Jr., J. P. Luby, W. G. Johanson, Jr.,J. A. Barnett, and J. P. Sanford. 1970. Serratia marces-
cens infections from inhalation therapy medications: no-socomial outbreak. Am. Intern. Med. 73:15-21.
15. Wollstein, M., and S. J. Meltzer. 1913. The lesions pro-duced by intra-bronchial insufflation of B. prodigiosus.Science 38:452-453.
VOL. 40, 1983
on April 12, 2019 by guest
http://iai.asm.org/
Dow
nloaded from