identification of potentially virulent strains of haemophilus parasuis using a multiplex pcr for...
TRANSCRIPT

The Veterinary Journal 191 (2012) 213–218
Contents lists available at ScienceDirect
The Veterinary Journal
journal homepage: www.elsevier .com/ locate/ tv j l
Identification of potentially virulent strains of Haemophilus parasuis usinga multiplex PCR for virulence-associated autotransporters (vtaA)
Alex Olvera a,⇑,1, Sonia Pina a,b, Núbia Macedo c, Simone Oliveira c, Virginia Aragon a,b,2, Albert Bensaid a,2
a Centre de Recerca en Sanitat Animal (CReSA), UAB-IRTA, Campus de la Universitat Autònoma de Barcelona, 08193 Bellaterra, Barcelona, Spainb Institut de Recerca i Tecnologia Agroalimentàries (IRTA), Barcelona, Spainc Veterinary Diagnostic Laboratory, College of Veterinary Medicine, University of Minnesota, Saint Paul, MN, USA
a r t i c l e i n f o a b s t r a c t
Article history:Accepted 11 December 2010
Keywords:Haemophilus parasuisvtaAMultilocus sequence typingVirulence
1090-0233/$ - see front matter � 2010 Elsevier Ltd. Adoi:10.1016/j.tvjl.2010.12.014
⇑ Corresponding author. Tel.: +34 93 5814567.E-mail address: [email protected] (A. Olve
1 Present address: Departament de Genètica i dAutònoma de Barcelona, 08193 Bellaterra, Barcelona, S
2 Both authors contributed equally to this work.
Haemophilus parasuis is the aetiological agent of Glässer’s disease and is also a commensal of the upperrespiratory tract of pigs. Trimeric autotransporter (vtaA) genes have been identified in H. parasuis anddivided into three groups on the basis of the translocator domain sequence. In this study, group 3 vtaAgenes were demonstrated by PCR in all 157 H. parasuis isolates tested. Group 1 vtaA genes were associ-ated with virulent strains; 52/54 (96%) group 1 vtaA negative field isolates were isolated from the nasalpassages of healthy animals, whereas no group 1 vtaA negative field isolates were isolated from cases ofGlässer’s disease. There was an association between absence of group 1 vtaA, sensitivity to phagocytosisand serum and classification of isolates into nasal cluster C by multilocus sequence typing. A multiplexPCR was developed for diagnosis of H. parasuis at the species level (group 3 vtaA positive) and to differ-entiate putative non-virulent strains (group 1 vtaA negative). When applied to field samples, the PCR con-firmed a high prevalence of H. parasuis in conventionally farmed pigs and demonstrated that almost halfof the animals carried potentially virulent strains.
� 2010 Elsevier Ltd. All rights reserved.
Introduction
Haemophilus parasuis is a Gram-negative bacterium that causesGlässer’s disease in pigs. Strains of H. parasuis can range fromhighly virulent to non-pathogenic (Rapp-Gabrielson et al., 2006).Multilocus sequence typing (MLST) has demonstrated a geneticlineage associated with polyserositis and another lineage associ-ated with nasal colonisation (Olvera et al., 2006b). The molecularbasis of virulence has not been established, although recent studieshave revealed several putative virulence factors (Jin et al., 2008;Sack and Baltes, 2009). Serum and phagocytosis resistance andinvasion of endothelial cells have been associated with virulenceof H. parasuis (Vanier et al., 2006; Cerdà-Cuéllar and Aragon,2008; Olvera et al., 2009; Aragon et al., 2010a).
H. parasuis possesses virulence associated trimeric autotrans-porters (vtaA), which can be divided into three groups by analysisof their translocator domains (Pina et al., 2009). Comparative geno-mic hybridisation studies have shown that group 3 vtaA is highlyconserved among invasive and non-invasive strains, whilst groups1 and 2 vtaA were detected only in virulent strains (Pina et al., 2009).
ll rights reserved.
ra).e Microbiologia, Universitatpain.
Since it can be difficult to isolate H. parasuis (Rapp-Gabrielsonet al., 2006), PCRs have been designed to improve the diagnosisof this organism (Oliveira et al., 2001; de la Puente Redondoet al., 2003; Jung et al., 2004; Angen et al., 2007; Turni et al.,2010). However, none of the current PCRs differentiates virulentfrom non-virulent strains and the bacterium has to be detectedin systemic lesions to be of diagnostic value. In this study, we eval-uated a multiplex PCR based on vtaA genes for diagnosis of H. para-suis infection and differentiation of non-virulent from potentiallyvirulent isolates.
Materials and methods
Reference bacterial strains
Fifteen serovar reference strains (Kielstein and Rapp-Gabrielson, 1992) and 140field isolates typed by MLST and with clinical records were used in this study (Sup-plementary Table 1). Information about the in vivo virulence of 17 strains was avail-able (Supplementary Table 1). Two field isolates (WB21-4 and IT29205) previouslyused in experimental infections (Aragon et al., 2010b) were also included. The fieldisolates were identified as H. parasuis using biochemical tests (Møller and Kilian,1990; Dousse et al., 2008), confirmed by 16S rRNA gene sequencing (Olvera et al.,2006a). Phylogenetic analysis was performed with a neighbour-joining tree usingmaximum composite likelihood distances and 1000 bootstraps.
To assess the species specificity of the PCR, 26 non-H. parasuis bacteria weretested at CReSA and 34 at the Minnesota Veterinary Diagnostic Laboratory (MVDL)(Supplementary Table 2). The type strains of Actinobacillus indolicus and Actinobacil-lus porcinus were kindly provided by Øystein Angen (National Veterinary Institute

214 A. Olvera et al. / The Veterinary Journal 191 (2012) 213–218
of the Technical University of Denmark). Actinobacillus suis 97-4918B was kindlyprovided by Marcelo Gottschalk (University of Montreal).
Swabs for isolation of H. parasuis
Swabs were collected at postmortem examination from snatch-farrowed andcolostrum-deprived piglets infected intranasally with approximately 108 colonyforming units (CFU) of six H. parasuis strains (PV1-12, Nagasaki, 9904108,IT29205, CA38-4 and 373/03A) (Aragon et al., 2010b). Animal experiments were ap-proved by the Ethical and Animal Welfare Committee of the Universitat Autònomade Barcelona (Generalitat de Catalunya CEA approval number: 3987). Twenty-fiveduplicate swabs from the nasal cavity (n = 7), thoracic cavity (n = 6), brain (n = 4),abdominal cavity (n = 2), pericardium (n = 2), joints (n = 2), blood (n = 1) and cere-brospinal fluid (CSF, n = 1) of experimentally infected pigs were used in the study.
Nasal swabs from 94 8-week-old piglets from six conventional Spanish farmswith no history of Glässer’s disease were also included (Grau-Roma et al., 2008).Swabs were resuspended in 1 mL phosphate buffered saline (PBS) and vigorouslyvortexed.
Bacterial culture and DNA extraction
Reference strains of H. parasuis and one of each duplicate field swab were platedonto chocolate agar and incubated at 37 �C in 5% CO2. DNA was extracted from bac-terial colonies using the InstaGene Matrix kit (BioRad). DNA was also extracted di-rectly from the second of each duplicate field swab and from blood and CSF usingthe Nucleospin Blood DNA extraction kit (Macherey–Nagel).
PCR for detection of vtaA
PCR primers for amplification of vtaA translocator domains groups 1–3 (PCR1,PCR2 and PCR3, respectively) (Pina et al., 2009) are shown in Table 1. PCR reaction
Table 1Primer pairs used to amplify the three groups of vtaA translocator domains by PCR.
Primers Sequence (50 ? 30) Amplicon (bp)
PCR1 YADAF1 TTTAGGTAAAGATAAGCAAGGAAATCC 406PADHR1 CCACACAAAACCTACCCCTCCTCC
PCR2 YADAF2 AGCTTAATATCTCAGCACAAGGTGC 294PADHR2 CCACTGATAACCTACCCCCACAGAG
PCR3 YADAF3 AATGGTAGCCAGTTGTATAATGTTGC 291PADHR3 CCACTGTAATGCAATACCTGCACC
Virulence + 0 0 0 + + 0ASerovar 8 3 9 7 4 2 11
SW
SW SWMW C
W11
D7 17
W12
W14
H46
b500
W C5 14 74 74 24 40 65bp
400500
300
200500400300300
200200
400500400300
200
Virulence ++ + 0 + ++ 0 0 0BSerovar 1 2 3 4 5 6 7 9
S SS
NaSW
1 D71
SW1
SW1
agasaN 1M
114
7474
124
140
aki
N4
131
00bp
MW
40050000
300
200
Fig. 1. Individual PCR of 10 serovar reference strains tested at Centre de Recerca en SanMinnesota Veterinary Diagnostic Laboratory (MVDL) (B). Amplification products wereidentifier, serovar and virulence classification (Kielstein and Rapp-Gabrielson, 1992) amoderately virulent; ++, highly virulent. More information about the strains can be fou
tubes contained GoTaq buffer (Promega), 1.25 mM MgCl2, 0.4 mM each deoxynucle-otide triphosphate (dNTP), 400 nM each primer, 1 U GoTaq polymerase (Promega)and 50 ng genomic DNA in a final volume of 25 lL. Cycling conditions were5 min at 94 �C, followed by 30 cycles of 45 s at 94 �C, 45 s at 59 �C and 1 min at72 �C, then a final incubation at 72 �C for 7 min.
Annealing temperature and MgCl2 and primer concentrations were optimisedto amplify vtaA translocator domains groups 1 and 3 in a multiplex format(mPCR1/3). Multiplex PCR reaction tubes contained GoTaq buffer, 2 mM MgCl2,0.4 mM each dNTP, 800 nM YADAF1 and PADHR1 primers, 400 nM YADAF3 andPADHR3 primers, 1 U GoTaq polymerase and 10 ng genomic DNA or 2.5 lL DNA ex-tracted from swabs in a final volume of 25 lL. Cycling conditions were 5 min at94 �C, followed by 25 cycles of 45 s at 94 �C, 45 s at 64 �C and 1 min at 72 �C, thena final incubation at 72 �C for 7 min. Amplicons were analysed in 2.5% agarose gels(Seakem SE) and stained with 0.5 lg/mL ethidium bromide. A 100 bp ladder (Invit-rogen) was used to estimate molecular size.
Sensitivity and specificity of the multiplex PCR
mPCR1/3 sensitivity was determined using serial 5-fold dilutions (starting at10 ng per PCR) of H. parasuis genomic DNA of strains Nagasaki, 264/99, SW114and F9. In addition, an overnight culture of Nagasaki strain was resuspended inPBS at an optical density (OD)660 of 0.2, equivalent to approximately 108 CFU/mL.This suspension was diluted 1:10 and used for DNA extraction using the NucleospinBlood kit (Macherey–Nagel) and the number of CFU was confirmed by chocolateagar plate counts. Serial 10-fold dilutions of this suspension were also used todetermine PCR sensitivity.
The performance of the mPCR1/3 was also evaluated using DNA extracted fromswabs obtained from experimental infections, as described above. To evaluate thespecificity of PCRs 1–3 and the multiplex PCR, BLAST searches (Altschul et al.,1990) using the primer sequences were performed. Also, samples of genomicDNA from 60 non-H. parasuis bacteria that can be isolated from pigs were testedin the multiplex PCR (Supplementary Table 2). The presence of amplifiable DNAin those samples was confirmed by PCR using universal primers for the bacterial16S rRNA gene (Calsamiglia et al., 1999).
Multilocus sequence typing
MLST analysis was performed by sequencing seven gene fragments (NCBI acces-sion numbers DQ781411–DQ782327 and HM242903–HM243119) (Olvera et al.,2006b). The sequence type (ST) of these strains and 131 strains already publishedwere analysed by BURST and unweighted pair group method with arithmetic mean(UPGMA) using START2 software (Jolley et al., 2001).
++ + ++
N
14 15 5
84-2
84-1
Nag2211
1599
gasak
NT13 95 ki C
Group 1Group 1
Group 2Group 2
Group 3
++ 0 ++ ++ ++ +
10 11 12 13 14 15
84-
8484- -159
-221
H4
H3
H4
-179
Multiplex
95
113
465
367
425
975
Group 11G 3Group 3
itat Animal (CReSA) (A) and multiplex PCR of 14 serovar reference strains tested atvisualised in a 2.5% agarose gel stained with 0.5 lg/mL ethidium bromide. Strainre indicated. MW, Molecular weight markers; 0, non-virulent; ±, mildly virulent,nd in Supplementary Table 1.

**A
75
ns
50
*
rain
Str
*25
%
*25
*
Healthy Pneumonia Glässer's
*Healthy
(n=77)Pneumonia
(n=29)Glässer s
(n=44)
B
C
*
s
75*
ains
*
Str 50
%
25 ***
Cluster A Cluster ClusteCluster A(n=48)
Cluster B(n=48)
Cluster C(n=54)
**
7575
ains
50Stra
%S
25
%
*
Cluster A Cl B Cl C
* **
(n=48)Cluster B
(n=48)Cluster C
(n=54)
Fig. 2. Correlation between the presence of vtaA translocator groups 1–3 and theputative virulence of the strains. (A) Association between the clinical origin of thestrains and the presence of the three groups of vtaA translocators. (B) Associationbetween the clinical origin of the strains and MLST classification. (C) Associationbetween vtaA translocators and MLST classification. In (A) and (C), light grey barsindicate isolates positive only for vtaA group 3 (PCR3), dark grey bars indicateisolates that are positive for both vtaA group 2 (PCR2) and vtaA group 3 (PCR3) andblack bars indicate isolates that are positive for all three vtaA translocator groups(PCR1, PCR2 and PCR3). In (B), white bars indicate isolates from healthy animals,bars with horizontal stripes indicate isolates from pigs with pneumonia and barswith oblique stripes indicate isolates from pigs with Glässer’s disease. Eachassociation was significant in the v2 test (P < 0.05). � indicates frequenciesproducing v2 significance by standard residuals (a = 0.01).
A. Olvera et al. / The Veterinary Journal 191 (2012) 213–218 215
Resistance to serum and phagocytosis
Resistance to serum and phagocytosis was determined as described previously(Cerdà-Cuéllar and Aragon, 2008; Olvera et al., 2009). Data already available (32strains for serum resistance and 10 strains for phagocytosis resistance) were com-plemented by assaying additional strains selected to more evenly represent each ofthe three clusters defined by UPGMA analysis of MLST: strains CC2-2, 46080, RU15-4P, CC6-7 and FL1-3 were tested for serum resistance; strains 61/03, 2725, 2620,174, CA38-4, SL3-2, SL7-2, CT-175L, P015/96, 06-3517, ND14-1 and 279/03 weretested for phagocytosis resistance.
Data analysis
Isolates were divided in different categories based on clinical source: (1) appar-ently healthy animals (isolates from nasal cavities); (2) animals with pneumoniawithout Glässer’s disease (isolates from the lungs); and (3) animals with Glässer’sdisease (isolates from the lungs or systemic lesions). The association between clin-ical origin, UPGMA clusters and vtaA translocator domain PCR profiles (n = 150 iso-lates) were evaluated by the v2 test using a significance threshold of P < 0.05. v2
standard residuals (a = 0.010) were calculated using SPSS v15. The association ofserum resistance (n = 37) and phagocytosis resistance (n = 22) with clinical origin,UPGMA clusters and vtaA translocator domain PCR profiles was evaluated usingFisher’s exact test of independence (P < 0.05). Adjusted odds ratios (OR) 95% confi-dence intervals were calculated for two-factor interaction using EpiInfo (CDC) forstrains with profiles positive for PCR1 (n = 54) and isolates from pneumonia or Gläs-ser’s disease (n = 73).
Results
Correlation between detection of vtaA translocator domain groups byPCR and clinical origin
On the basis of phylogenetic analysis of the 16S rRNA gene, the140 field isolates of H. parasuis were grouped with H. parasuis ref-erence strains (Supplementary Fig. 1). All strains were positive inPCR3, indicating that this assay was able to detect H. parasuis (Sup-plementary Fig. 2). To determine whether there was an associationbetween vtaA translocator groups and H. parasuis virulence, wecompared the profile of presence/absence of the three differentvtaA translocator groups with the clinical records of each strain.Results for serovar reference strains obtained at CReSA and MVDLare shown in Fig. 1.
Non-virulent reference strains 131, SW114 and D74 and mildlyvirulent strain C5 were positive only for vtaA translocator group 3,while highly virulent reference strains N4, Nagasaki, H367, H425,84-17975 and 84-22113, moderately virulent reference strainsSW140 and 84-15995 and mildly virulent strain SW124 were posi-tive for vtaA translocator groups 1 and 3. Non-virulent referencestrains 174 and H465 were positive in PCR1, PCR2 and PCR3.
All field strains from cases of Glässer’s disease and most of thefield isolates from pigs with pneumonia were positive in all threePCRs. However, 25/77 (32.5%) nasal isolates were also positive inthe three PCRs (Fig. 2A). Five of these 25 strains were isolated fromfarms with Glässer’s disease outbreaks, two from farms vaccinatingagainst H. parasuis and two from farms with late weaning (at28 days). Strains positive in PCR1 had a higher probability of beingisolated from diseased animals (pigs with pneumonia or Glässer’sdisease) than from healthy animals (OR 9.76–285.19). Conversely,there was an association between PCR1-negative strains and nasalisolation, with 52/54 (96%) of the isolates negative for group 1 vtaAbeing isolated from nasal swabs of healthy animals and none fromanimals with Glässer’s disease. The two non-nasal isolates (279/03,03/05) negative for group 1 vtaA were isolated from cases ofpneumonia.
Relationship between MLST UPGMA clusters and vtaA translocatorgroups
A total of 122 STs were defined among 150 strains. Using a re-laxed BURST criterion (strains sharing five identical loci were as-
signed to the same group), the strains were classified into 18groups, while 75 isolates remained as ungrouped individualstrains. Searching for more distant relationships by UPGMA, threemain clusters were defined (Supplementary Fig. 2): isolates in clus-ter A were associated with Glässer’s disease; cluster B included na-sal isolates and isolates from diseased animals (includingpneumonic cases) without a clear association; and cluster C wasassociated with nasal isolates from healthy animals (Fig. 2B). Fur-thermore, 95.8% of the strains in cluster A were grouped in thesame branch in the 16S rRNA gene phylogeny (SupplementaryFig. 1).
When compared, MLST classification and vtaA group PCR pro-files showed an association between the classification of the strains
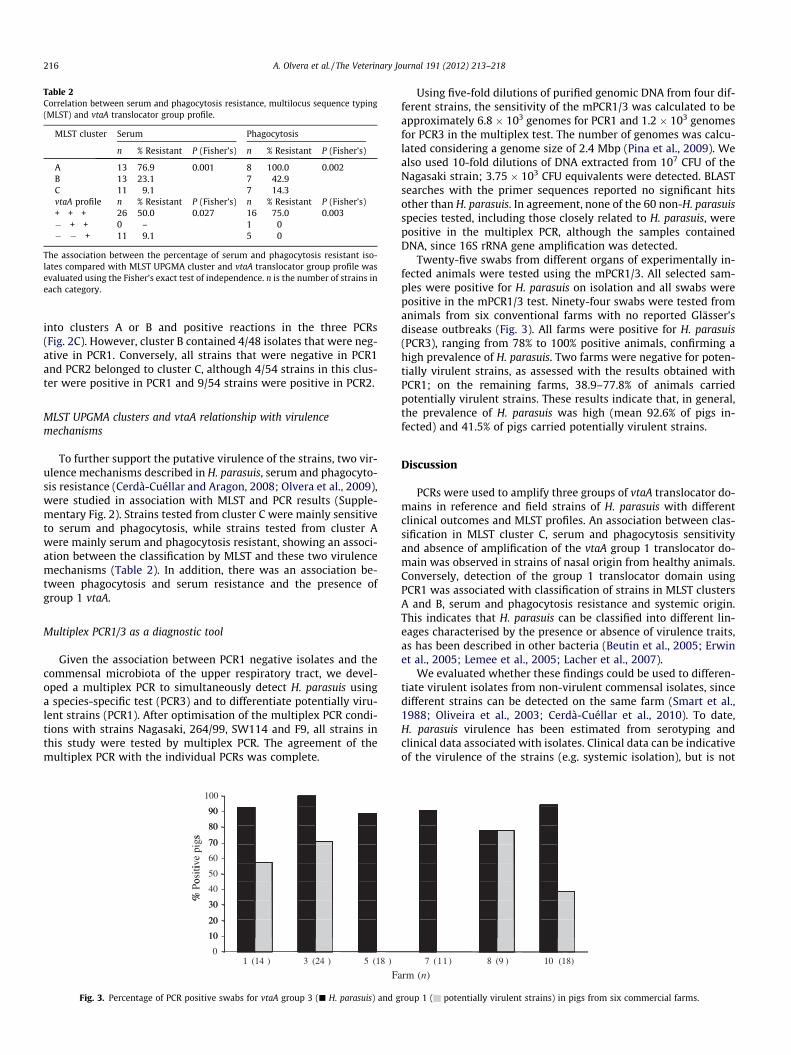
Table 2Correlation between serum and phagocytosis resistance, multilocus sequence typing(MLST) and vtaA translocator group profile.
MLST cluster Serum Phagocytosis
n % Resistant P (Fisher’s) n % Resistant P (Fisher’s)
A 13 76.9 0.001 8 100.0 0.002B 13 23.1 7 42.9C 11 9.1 7 14.3vtaA profile n % Resistant P (Fisher’s) n % Resistant P (Fisher’s)+ + + 26 50.0 0.027 16 75.0 0.003� + + 0 – 1 0� � + 11 9.1 5 0
The association between the percentage of serum and phagocytosis resistant iso-lates compared with MLST UPGMA cluster and vtaA translocator group profile wasevaluated using the Fisher’s exact test of independence. n is the number of strains ineach category.
216 A. Olvera et al. / The Veterinary Journal 191 (2012) 213–218
into clusters A or B and positive reactions in the three PCRs(Fig. 2C). However, cluster B contained 4/48 isolates that were neg-ative in PCR1. Conversely, all strains that were negative in PCR1and PCR2 belonged to cluster C, although 4/54 strains in this clus-ter were positive in PCR1 and 9/54 strains were positive in PCR2.
MLST UPGMA clusters and vtaA relationship with virulencemechanisms
To further support the putative virulence of the strains, two vir-ulence mechanisms described in H. parasuis, serum and phagocyto-sis resistance (Cerdà-Cuéllar and Aragon, 2008; Olvera et al., 2009),were studied in association with MLST and PCR results (Supple-mentary Fig. 2). Strains tested from cluster C were mainly sensitiveto serum and phagocytosis, while strains tested from cluster Awere mainly serum and phagocytosis resistant, showing an associ-ation between the classification by MLST and these two virulencemechanisms (Table 2). In addition, there was an association be-tween phagocytosis and serum resistance and the presence ofgroup 1 vtaA.
Multiplex PCR1/3 as a diagnostic tool
Given the association between PCR1 negative isolates and thecommensal microbiota of the upper respiratory tract, we devel-oped a multiplex PCR to simultaneously detect H. parasuis usinga species-specific test (PCR3) and to differentiate potentially viru-lent strains (PCR1). After optimisation of the multiplex PCR condi-tions with strains Nagasaki, 264/99, SW114 and F9, all strains inthis study were tested by multiplex PCR. The agreement of themultiplex PCR with the individual PCRs was complete.
90
100
80
90
70
80
s
60
70
pigs
50
ive
30
40
ositi
20
30
% P
o
10
20
%
0
10
1 (14 ) 3 (24 ) 5 (18 )
Fa
Fig. 3. Percentage of PCR positive swabs for vtaA group 3 (j H. parasuis) and g
Using five-fold dilutions of purified genomic DNA from four dif-ferent strains, the sensitivity of the mPCR1/3 was calculated to beapproximately 6.8 � 103 genomes for PCR1 and 1.2 � 103 genomesfor PCR3 in the multiplex test. The number of genomes was calcu-lated considering a genome size of 2.4 Mbp (Pina et al., 2009). Wealso used 10-fold dilutions of DNA extracted from 107 CFU of theNagasaki strain; 3.75 � 103 CFU equivalents were detected. BLASTsearches with the primer sequences reported no significant hitsother than H. parasuis. In agreement, none of the 60 non-H. parasuisspecies tested, including those closely related to H. parasuis, werepositive in the multiplex PCR, although the samples containedDNA, since 16S rRNA gene amplification was detected.
Twenty-five swabs from different organs of experimentally in-fected animals were tested using the mPCR1/3. All selected sam-ples were positive for H. parasuis on isolation and all swabs werepositive in the mPCR1/3 test. Ninety-four swabs were tested fromanimals from six conventional farms with no reported Glässer’sdisease outbreaks (Fig. 3). All farms were positive for H. parasuis(PCR3), ranging from 78% to 100% positive animals, confirming ahigh prevalence of H. parasuis. Two farms were negative for poten-tially virulent strains, as assessed with the results obtained withPCR1; on the remaining farms, 38.9–77.8% of animals carriedpotentially virulent strains. These results indicate that, in general,the prevalence of H. parasuis was high (mean 92.6% of pigs in-fected) and 41.5% of pigs carried potentially virulent strains.
Discussion
PCRs were used to amplify three groups of vtaA translocator do-mains in reference and field strains of H. parasuis with differentclinical outcomes and MLST profiles. An association between clas-sification in MLST cluster C, serum and phagocytosis sensitivityand absence of amplification of the vtaA group 1 translocator do-main was observed in strains of nasal origin from healthy animals.Conversely, detection of the group 1 translocator domain usingPCR1 was associated with classification of strains in MLST clustersA and B, serum and phagocytosis resistance and systemic origin.This indicates that H. parasuis can be classified into different lin-eages characterised by the presence or absence of virulence traits,as has been described in other bacteria (Beutin et al., 2005; Erwinet al., 2005; Lemee et al., 2005; Lacher et al., 2007).
We evaluated whether these findings could be used to differen-tiate virulent isolates from non-virulent commensal isolates, sincedifferent strains can be detected on the same farm (Smart et al.,1988; Oliveira et al., 2003; Cerdà-Cuéllar et al., 2010). To date,H. parasuis virulence has been estimated from serotyping andclinical data associated with isolates. Clinical data can be indicativeof the virulence of the strains (e.g. systemic isolation), but is not
7 (11 ) 8 (9 ) 10 (18)
rm (n)
roup 1 ( potentially virulent strains) in pigs from six commercial farms.

A. Olvera et al. / The Veterinary Journal 191 (2012) 213–218 217
conclusive, since multiple factors contribute to the development ofGlässer’s disease. This is especially problematic for lung isolates,since this organ is potentially exposed to upper respiratory tractcommensal microbiota.
Serotyping also has limitations due to the existence of non-typeable strains and virulence variations within some serovars.For example, serotype 2 strain Bakos A9 (Kielstein and Rapp-Gabri-elson, 1992), serotype 14 strain H792 (Bak and Riising, 2002) andserotype 4 strain 9904108 (Aragon et al., 2010b) appear to benon-virulent. Furthermore, serotype 2 strain SW124 and serotype4 strain SW140 were not able to reproduce disease when inocu-lated intranasally (Nielsen, 1993) instead of the intraperitonealroute used previously (Kielstein and Rapp-Gabrielson, 1992). Con-versely, ‘non-virulent’ serovars 7 and 11 have been isolated fromlesions of animals with Glässer’s disease (Angen et al., 2004; Caiet al., 2005; Rubies et al., 1999) and disease was reproduced in 1/6 animals challenged intranasally with serovar 7 strain 373/03A(Aragon et al., 2010b). Interestingly, the non-virulent referencestrain of this serovar (strain 174) and isolate 373/03A were positivein the group 1 PCR.
Since 96% of isolates that were negative in PCR1 came from thenasal cavities of healthy animals and since no PCR1 negative iso-lates were identified from cases of Glässer’s disease, this suggeststhat group 1 vtaA could be a necessary, but not sufficient, factorfor H. parasuis virulence; necessary, because there were no PCR1negative isolates from Glässer’s disease affected animals, but notsufficient, because some PCR1 positive nasal isolates from healthyanimals were identified.
Although an isolate positive for vtaA1 can be either virulent ornon-virulent, it has a greater probability of being virulent. Non-vir-ulent reference strains 174 and H465, as well as moderate andmildly virulent reference strains SW140 and SW124, were positivein the three vtaA PCRs, but these strains did not induce diseaseafter intranasal inoculation (Nielsen, 1993; Amano et al., 1994,1996). Conversely, intraperitoneal inoculation of group 1 negativestrain C5 resulted only in mild clinical signs and lesions (Kielsteinand Rapp-Gabrielson, 1992); however, no experimental intranasalinoculation of this strain has been undertaken. Moreover, 32.5% ofnasal isolates were positive in the three vtaA translocator PCRs.
The existence of isolates positive for group 1 in nasal samplesfrom healthy animals could be explained by the existence of carrieranimals immunologically protected against virulent strains (Bakand Riising, 2002; Solano-Aguilar et al., 1999; Martin de la Fuenteet al., 2009) or animals at an early (subclinical) stage of infection.Several reference virulence strains (N4, SW124, SW140, H555) iso-lated from the nasal cavities of healthy animals were able to repro-duce Glässer’s disease experimentally (Rapp-Gabrielson et al.,2006).
These results are compatible with the predicted function ofVtaA in adhesion. However, it should be taken into account thatthe functionality of trimeric autotransporters is given by the pas-senger domain (Linke et al., 2006), while the PCRs described heredetect the translocator domains. Therefore, further studies are re-quired to clarify the role of these molecules in the pathogenesisof H. parasuis.
Since the absence of the group 1 translocator domain could bean indicator for non-virulence, a multiplex PCR was evaluated foridentification of H. parasuis strains. The mPCR1/3 test was foundto be species-specific, but its sensitivity was low. The multiplexPCR showed a sensitivity of 3.75 x 103 CFU, whereas Oliveiraet al. (2001) reported a sensitivity of 102 CFU for their 16S rRNAgene based test. This PCR was used by Angen et al. (2007) with asensitivity of 0.5 CFU (10 CFU/mL) or 102 16S rRNA geneamplicons.
In addition, we evaluated the mPCR1/3 test in swabs from dif-ferent organs of experimentally infected animals and the results
were consistent with bacterial isolation. Animals were also testedfrom commercial farms with no reported current outbreak of Gläs-ser’s disease. As expected, the prevalence of H. parasuis was high in8-week-old piglets, but the distribution of animals carrying bacte-ria bearing group 1 vtaA translocators varied between farms, con-firming the complex epidemiology of H. parasuis. Interestingly,nasal swabs from animals from two farms with no Glässer’s diseaserecords were negative in PCR1.
Conclusions
Isolation of H. parasuis from the nasal cavity of pigs was associ-ated with the absence of group 1 vtaA translocators, MLST cluster-ing and the lack of recognised virulence mechanisms (serum andphagocytosis resistance). A multiplex PCR amplifying group 1 andgroup 3 vtaA translocator domains was used to detect H. parasuisand to identify putative non-virulent strains. This multiplex PCRcould be used to prevent introduction of pigs with potentially vir-ulent H. parasuis strains onto farms without Glässer’s disease.However, full validation of the multiplex PCR, mainly its predictivevalue for non-virulent isolates, requires further inter-laboratorytesting.
Conflict of interest statement
None of the authors of this paper has a financial or personalrelationship with other people or organisations that could inappro-priately influence or bias the content of the paper.
Acknowledgements
The authors would like to thank J. Segalés and L.L. Grau-Romafor providing farm samples, M. Nofraries for helping us with thestatistical analysis and Núria Galofré for technical support. Thiswork has been supported by Grant AGL2007-60432 from the Min-isterio de Ciencia e Innovación of the Spanish Government.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.tvjl.2010.12.014.
References
Altschul, S.F., Gish, W., Miller, W., Myers, E.W., Lipman, D.J., 1990. Basic localalignment search tool. Journal of Molecular Biology 215, 403–410.
Amano, H., Shibata, M., Kajio, N., Morozumi, T., 1994. Pathologic observations ofpigs intranasally inoculated with serovar 1, 4 and 5 of Haemophilus parasuisusing immunoperoxidase method. Journal of Veterinary Medicine Science 56,639–644.
Amano, H., Shibata, M., Kajio, N., Morozumi, T., 1996. Pathogenicity of Haemophilusparasuis serovars 4 and 5 in contact-exposed pigs. Journal of VeterinaryMedicine Science 58, 559–561.
Angen, Ø., Svensmark, B., Mittal, K.R., 2004. Serological characterization of DanishHaemophilus parasuis isolates. Veterinary Microbiology 103, 255–258.
Angen, Ø., Oliveira, S., Ahrens, P., Svensmark, B., Leser, T.D., 2007. Development of animproved species specific PCR test for detection of Haemophilus parasuis.Veterinary Microbiology 119, 266–276.
Aragon, V., Bouchet, B., Gottschalk, M., 2010a. Invasion of endothelial cells bysystemic and nasal strains of Haemophilus parasuis. The Veterinary Journal 186,264–267.
Aragon, V., Cerdà-Cuéllar, M., Fraile, L., Mombarg, M., Nofrarías, M., Olvera, A., Sibila,M., Solanes, D., Segalés, J., 2010b. Correlation between clinico-pathologicaloutcome and typing of Haemophilus parasuis field strains. VeterinaryMicrobiology 142, 387–393.
Bak, H., Riising, H.J., 2002. Protection of vaccinated pigs against experimentalinfections with homologous and heterologous Haemophilus parasuis. VeterinaryRecord 151, 502–505.
Beutin, L., Kaulfuss, S., Herold, S., Oswald, E., Schmidt, H., 2005. Genetic analysis ofenteropathogenic and enterohemorrhagic Escherichia coli serogroup O103strains by molecular typing of virulence and housekeeping genes and pulsed-field gel electrophoresis. Journal of Clinical Microbiology 43, 1552–1563.

218 A. Olvera et al. / The Veterinary Journal 191 (2012) 213–218
Cai, X., Chen, H., Blackall, P.J., Yin, Z., Wang, L., Liu, Z., Jin, M., 2005. Serologicalcharacterization of Haemophilus parasuis isolates from China. VeterinaryMicrobiology 111, 231–236.
Calsamiglia, M., Pijoan, C., Solano, G., Rapp-Gabrielson, V., 1999. Development of anoligonucleotide-specific capture plate hybridization assay for detection ofHaemophilus parasuis. Journal of Veterinary Diagnostic Investigation 11, 140–145.
Cerdà-Cuéllar, M., Aragon, V., 2008. Serum-resistance in Haemophilus parasuis isassociated with systemic disease in swine. The Veterinary Journal 175, 384–389.
Cerdà-Cuéllar, M., Naranjo, J.F., Verge, A., Nofrarias, M., Cortey, M., Olvera, A.,Segalés, J., Aragon, V., 2010. Sow vaccination modulates the colonization ofpiglets by Haemophilus parasuis. Veterinary Microbiology 145, 315–320.
de la Puente Redondo, V.A., Navas Mendez, J., Garcia del Blanco, N., Ladron Boronat,N., Gutierrez Martin, C.B., Rodriguez Ferri, E.F., 2003. Typing of Haemophilusparasuis strains by PCR-RFLP analysis of the tbpA gene. Veterinary Microbiology92, 253–262.
Dousse, F., Thomann, A., Brodard, I., Korczak, B.M., Schlatter, Y., Kuhnert, P., Miserez,R., Frey, J., 2008. Routine phenotypic identification of bacterial species of thefamily Pasteurellaceae isolated from animals. Journal of Veterinary DiagnosticInvestigation 20, 716–724.
Erwin, A.L., Nelson, K.L., Mhlanga-Mutangadura, T., Bonthuis, P.J., Geelhood, J.L.,Morlin, G., Unrath, W.C., Campos, J., Crook, D.W., Farley, M.M., Henderson, F.W.,Jacobs, R.F., Muhlemann, K., Satola, S.W., van Alphen, L., Golomb, M., Smith, A.L.,2005. Characterization of genetic and phenotypic diversity of invasivenontypeable Haemophilus influenzae. Infection and Immunity 73, 5853–5863.
Grau-Roma, L., Crisci, E., Sibila, M., Lopez-Soria, S., Nofrarias, M., Cortey, M., Fraile, L.,Olvera, A., Segales, J., 2008. A proposal on porcine circovirus type 2 (PCV2)genotype definition and their relation with postweaning multisystemic wastingsyndrome (PMWS) occurrence. Veterinary Microbiology 128, 23–35.
Jin, H., Wan, Y., Zhou, R., Li, L., Luo, R., Zhang, S., Hu, J., Langford, P.R., Chen, H., 2008.Identification of gene transcribed by Haemophilus parasuis in necrotic porcinelung through the selective capture of transcribed sequences (SCOTS).Environmental Microbiology 10, 3326–3336.
Jolley, K.A., Feil, E.J., Chan, M.S., Maiden, M.C., 2001. Sequence type analysis andrecombinational tests (START). Bioinformatics 17, 1230–1231.
Jung, K., Ha, Y., Kim, S.H., Chae, C., 2004. Development of polymerase chain reactionand comparison with in situ hybridization for the detection of Haemophilusparasuis in formalin-fixed, paraffin-embedded tissues. Journal of VeterinaryMedicine Science 66, 841–845.
Kielstein, P., Rapp-Gabrielson, V.J., 1992. Designation of 15 serovars of Haemophilusparasuis on the basis of immunodiffusion using heat-stable antigen extracts.Journal of Clinical Microbiology 30, 862–865.
Lacher, D.W., Steinsland, H., Blank, T.E., Donnenberg, M.S., Whittam, T.S., 2007.Molecular evolution of typical enteropathogenic Escherichia coli: clonal analysisby multilocus sequence typing and virulence gene allelic profiling. Journal ofBacteriology 189, 342–350.
Lemee, L., Bourgeois, I., Ruffin, E., Collignon, A., Lemeland, J.F., Pons, J.L., 2005.Multilocus sequence analysis and comparative evolution of virulence-associated genes and housekeeping genes of Clostridium difficile. Microbiology151, 3171–3180.
Linke, D., Riess, T., Autenrieth, I.B., Lupas, A., Kempf, V.A., 2006. Trimericautotransporter adhesins: variable structure, common function. Trends inMicrobiology 14, 264–270.
Martin de la Fuente, A.J., Gutierrez Martin, C.B., Perez Martinez, C., Garcia Iglesias,M.J., Tejerina, F., Rodriguez Ferri, E.F., 2009. Effect of different vaccineformulations on the development of Glässer’s disease induced in pigs byexperimental Haemophilus parasuis infection. Journal of Comparative Pathology140, 169–176.
Møller, K., Kilian, M., 1990. V factor-dependent members of the familyPasteurellaceae in the porcine upper respiratory tract. Journal of ClinicalMicrobiology 28, 2711–2716.
Nielsen, R., 1993. Pathogenicity and immunity studies of Haemophilus parasuisserotypes. Acta Veterinaria Scandinavica 34, 193–198.
Oliveira, S., Galina, L., Pijoan, C., 2001. Development of a PCR test to diagnoseHaemophilus parasuis infections. Journal of Veterinary Diagnostic Investigation13, 495–501.
Oliveira, S., Blackall, P.J., Pijoan, C., 2003. Characterization of the diversity ofHaemophilus parasuis field isolates by use of serotyping and genotyping.American Journal of Veterinary Research 64, 435–442.
Olvera, A., Calsamiglia, M., Aragon, V., 2006a. Genotypic diversity of Haemophilusparasuis field strains. Applied and Environmental Microbiology 72, 3984–3992.
Olvera, A., Cerda-Cuellar, M., Aragon, V., 2006b. Study of the population structure ofHaemophilus parasuis by multilocus sequence typing. Microbiology 152, 3683–3690.
Olvera, A., Ballester, M., Nofrarias, M., Sibila, M., Aragon, V., 2009. Differences inphagocytosis susceptibility in Haemophilus parasuis strains. Veterinary Research40, 24.
Pina, S., Olvera, A., Barcelo, A., Bensaid, A., 2009. Trimeric autotransporters ofHaemophilus parasuis: generation of an extensive passenger domain repertoirespecific for pathogenic strains. Journal of Bacteriology 191, 576–587.
Rapp-Gabrielson, V., Oliveira, S., Pijoan, C., 2006. Haemophilus parasuis. In: Straw,B.E., Zimmerman, J.J., D’Allaire, S., Taylor, D.J. (Eds.), Diseases of Swine. IowaState University Press, Iowa, IO, USA, p. 1153.
Rubies, X., Kielstein, P., Costa, L., Riera, P., Artigas, C., Espuna, E., 1999. Prevalence ofHaemophilus parasuis serovars isolated in Spain from 1993 to 1997. VeterinaryMicrobiology 66, 245–248.
Sack, M., Baltes, N., 2009. Identification of novel potential virulence-associatedfactors in Haemophilus parasuis. Veterinary Microbiology 136, 382–386.
Smart, N.L., Miniats, O.P., MacInnes, J.I., 1988. Analysis of Haemophilus parasuisisolates from southern Ontario swine by restriction endonucleasefingerprinting. Canadian Journal of Veterinary Research 52, 319–324.
Solano-Aguilar, G.I., Pijoan, C., Rapp-Gabrielson, V., Collins, J., Carvalho, L.F.,Winkelman, N., 1999. Protective role of maternal antibodies againstHaemophilus parasuis infection. American Journal Veterinary Research 60, 81–87.
Turni, C., Pyke, M., Blackall, P.J., 2010. Validation of a real-time PCR for Haemophilusparasuis. Journal of Applied Microbiology 108, 1323–1331.
Vanier, G., Szczotka, A., Friedl, P., Lacouture, S., Jacques, M., Gottschalk, M., 2006.Haemophilus parasuis invades porcine brain microvascular endothelial cells.Microbiology 152, 135–142.