identification of a cis-acting dna antisilencer element which
TRANSCRIPT

MOLECULAR AND CELLULAR BIOLOGY, May 1992, p. 2230-22400270-7306/92/052230-11$02.00/0
Identification of a cis-Acting DNA Antisilencer Element WhichModulates Vimentin Gene Expression
DENISE M. STOVERt AND ZENDRA E. ZEHNER*
Department ofBiochemistry and Molecular Biophysics and The Massey Cancer Center, Medical College ofVirginia/Virginia Commonwealth University, Richmond, Virginia 23298
Received 19 September 1991/Accepted 10 February 1992
Vimentin is a tissue-specific, developmentally regulated member of the intermediate filament protein familynormally expressed in cells of mesenchymal origin. Transcription factors which recognize specific cis-actingelements of the chicken gene include Sp-1 and the 95-kDa silencer protein which binds to a 40-bp silencerelement at -608 (F. X. Farrell, C. M. Sax, and Z. E. Zehner, Mol. Cell. Biol. 10:2349-2358, 1990). In thisstudy, we have identified a region upstream of the silencer element which restores gene activity. This region hasbeen further delineated into two functional subelements of 75 and 260 bp. In transient transfection assays, the75-bp element overrides the silencer effect of pStkCAT by 100%, while the 260-bp element is about half asactive. Neither element affects gene activity when the silencer element is absent. Therefore, these elements donot function as enhancers, but they may serve only to override the silencer element and therefore can be viewedas antisilencers. In addition, the 75-bp element binds a specific 140-kDa protein, as determined by gel mobilityshift assays and Southwestern (DNA-protein) blots, the binding site of which has been delineated to a 10- to17-bp element by DNase I protection experiments. During myogenesis, a direct correlation can be madebetween the binding efficiency of the 140-kDa protein, the silencer protein, and gene activity in vivo. Genesknown to contain a functional silencer element also contain at least one antisilencer element, as determined bysequence identity. Therefore, we have identified an antisilencer element and protein important in thedevelopmental regulation of vimentin gene expression which may be involved in the regulation of other genes.
Mediation of tissue-specific, developmentally regulatedgene expression necessitates a complex interplay betweenmultiple cis-acting DNA elements and trans-acting factors.Vimentin, a member of the intermediate filament family,displays both of these patterns of expression, thereby pro-viding an attractive model with which to study the interplaybetween cis-acting elements and their respective trans-acting factors. Determination of all regulatory elements,their mechanisms of action, and how they contribute to geneactivity is necessary for an understanding of not only of thefrequency of vimentin mRNA transcription in any givenstate, but also of its use as a prototype for other genesexhibiting similar expression patterns.The cellular cytoskeleton is comprised of three filamen-
tous systems, the microtubules (25 nm in diameter), themicrofilaments (6 nm), and the intermediate filaments (11nm). Intermediate-filament proteins (IFPs) are major com-
ponents of the cellular infrastructure which are thought toanchor both the nucleus and organelles and to maintaincellular compartmentalization as well as cellular morphology(16). IFPs are encoded by a large multigene family, membersof which comprise parts of both the cytoskeleton and kary-oskeleton. This large multigene family is divided into distinctsequence types: acidic keratins (type I), neutral-basic kera-tins (type II), desmin, vimentin, and glial fibrillary acidicprotein (type III), neurofilaments (type IV), lamins (type V),and the recently described nestin (type VI) (22, 42). Unlikethe constituent proteins of microtubules (tubulin) and micro-filaments (actin), IFPs are unique in their cell type specificity(42).
* Corresponding author.t Present address: Institute of Cancer and Developmental Biol-
ogy, Syntex Research, Palo Alto, CA 94304.
Vimentin is the type of IFP expressed in cells of mesen-
chymal origin. In addition, it is the only IFP which deviatesfrom its tissue-specific and developmental pattern of expres-sion (12) (for a review of IFPs, see reference 42). In thiscontext, vimentin can be coexpressed with most IFPs andappears transiently in a variety of developing and differen-tiated tissues as well as in tissue culture (12). Evidencesuggests that the expression pattern of vimentin is varied andcomplex (11). For example, during early erythropoiesis,both mammalian and avian erythrocytes contain vimentinfilaments, but as differentiation proceeds, the mature avianerythrocyte continues to contain vimentin filaments whilethe mammalian erythrocyte does not (8, 15). Several tumorcells express vimentin even though their embryonic origin isnot mesenchymal. For example, a non-hormone-induciblebreast cancer cell line (MDA-MB-231) expresses vimentin,whereas a hormone-inducible breast cancer cell line(MCF-7) does not (5, 40). Overall, vimentin's complexexpression pattern suggests an intricate regulatory circuit.
Previously, vimentin expression has been demonstrated tobe induced by serum, platelet-derived growth factor, andphorbol esters (16, 33). The differential expression pattern ofthe chicken vimentin gene was demonstrated to be depen-dent on multiple cis-acting elements and trans-acting factors(33, 36). In this case, basal expression has been attributed toa proximal promoter element (-161 to +1 bp) which isfunctional both in cells which express vimentin (mouse Lcell) and in those which do not (MH1Cl) (36). Within thispromoter element are five GC boxes (at least three of whichare functional in binding Sp-1) (38), an inversely orientedCAAT box, and a poor TATA box. A proximal enhancerelement (-321 to -161 bp) increases expression of a 5'-end-chloramphenicol acetyltransferase (CAT) construct inmouse L cells, arguing for a tissue-specific enhancer element(36, 37). This same proximal enhancer element contributes
2230
Vol. 12, No. 5

VIMENTIN DEVELOPMENTAL GENE EXPRESSION 2231
to the decrease in vimentin expression during myogenesis inseveral muscle cell lines (11, 36). Recently, Farrell et al.have delineated a negative element located 568 bp from thestart site of transcription (11). This 40-bp element binds a
95-kDa protein, as determined by gel mobility shift assays
(GMSAs), Southwestern (DNA-protein; SW) blots, andDNase I footprinting analysis (11).
All vimentin genes studied to date (human, hamster, andchicken) have been shown to contain similar regulatorycircuits consisting of a promoter and proximal enhancerelements (33). Both the human and chicken 5'-flankingsequences contain functional silencing elements, i.e., a
potential negative element at -853 to -795 bp in the human5'-flanking sequence and at -607 to -568 bp in the chickenvimentin gene (see Fig. 7). In the human vimentin gene, an
element residing at -700 bp has been demonstrated to beresponsible for both serum and phorbol ester inducibility.This 48-bp element contains an Sp-1 site flanked by twoAP-1/Jun binding sites. The presence of a distal enhancerelement in the human vimentin gene prompted an investiga-tion for a yet unidentified distal enhancer element in the5'-flanking region of the chicken vimentin gene. Previouswork suggested that this element resided at least 767 nucle-otides upstream of the initiation site for transcription. Thenotion of such a network of cis-acting elements is not novel,as positive and negative elements have been described forseveral other genes, including the alpha I protease inhibitorgene (28) and the chicken lysozyme gene (3).
In this study, a distal element has been localized upstreamof the transcriptional start site. This element is composed oftwo subelements, a 75-bp and a 260-bp element. Both can
overcome vimentin's silencer element. However, the 75-bpelement exhibits stronger regulating activity than does the260-bp element and is the focus of this report. Neitherelement enhances the transcription of the vimentin promoteror a heterologous thymidine kinase promoter element alone.When the 40-bp negative element is present, these elementsserve to override the silencer effect and restore gene activityto presilencer levels. For this reason, they represent a new
type of element which we will refer to as an antisilencerelement. Moreover, increasing copies of the silencer elementrequire a coordinate increase in the number of antisilencerelements in order to restore reporter gene activity. A specificprotein responsible for the activity of the 75-bp element hasbeen identified by GMSAs, SW analysis, and DNase Ifootprinting protection experiments. In addition, the inter-play between the antisilencer and silencer proteins was
investigated during myogenesis, when vimentin is differen-tially expressed. A direct correlation exists between theexpression state of vimentin and the binding activities ofthese two proteins.
MATERIALS AND METHODS
Cell culture, DNA transfections, and CAT assays. Mouse Lcells were grown in Dulbecco modified minimal essentialmedium supplemented with 10% fetal calf serum, L-glu-tamine, and antibiotics. Mouse L cells were seeded at 5 x105 cells per 100-mm dish 24 h prior to transfection. Chimericplasmids (15 ,ug) were transfected by the calcium phosphateprecipitation method, with slight modifications (14). Precip-itates were allowed to absorb to the cells for 24 h and thensubjected to a glycerol shock. Twenty-four hours later, cellswere harvested and lysates were obtained by repeatedfreeze-thawing in 250 mM Tris (pH 7.8). Standardizationamong different transfections was achieved by cotransfect-
ing 3 ,ug of pCMV-f-galactosidase to serve as an internalcontrol (29). CAT assays were performed and analyzed bythin-layer chromatography as described by Gorman et al.(13). Experiments were quantitated by excising the radioac-tive product and substrate from silica plates and determiningtheir 14C content by liquid scintillation counting (CAT activ-ity). P-Galactosidase activity was assayed in all cultures asdescribed by Nielsen et al. (29). CAT enzyme activity wasexpressed as picomoles of chloramphenicol acetylated perminute per microgram of protein per unit of 3-galactosidaseactivity. All values reported for mouse L cells are averagesof at least four separate transfections.DNA sequencing. A chicken genomic bacteriophage
Charon 4A (clone 1A) containing the vimentin gene plus 5'-and 3'-flanking sequences was digested with EcoRI-HindIIIto produce a 5' fragment of 837 bp which was subcloned intopUC18 (45). Both strands were sequenced by the method ofSanger et al. (35), with slight modifications, as well as by themethod of Maxam and Gilbert (24). Double-stranded se-quencing via the method of Sanger et al. (35) was performedby using U.S. Biochemical protocols. The universal primer,primer A (U.S. Biochemical), was used to sequence thenoncoding strand, and the reverse primer, M13 reverseprimer (Bethesda Research Laboratoties), was used to se-quence the coding strand. Three other specific primers,5'-GGGAGGTGGTGAGATCTCTG-3', 5'-ATGGGAAGCATCGGAGCCCG-3', and 5'-GATGCTGCACTCTGCACACCG-3', were synthesized to complete the sequence of the837-bp piece of DNA. Sequencing gels consisted of 8%acrylamide and 8 M urea in Tris-boric acid-EDTA buffer.Homologies to known DNA sequences present in GenBankwere analyzed by the Genetics Computer Group (Madison,Wis.) DNA sequence analysis program.
Plasmid constructions. The expression vector p8CAT is aderivative of pEMBL8 (9) in which the Fl origin of replica-tion has been removed and the bacterial CAT gene has beeninserted. pcV-767 was cloned into the multicloning site ofp8CAT as previously described (36) and includes 823 nucle-otides from -767 to +74 (with the initiation start site at +1)of the chicken vimentin gene. The 837-bp 5'-flanking se-quence was subcloned into pcV-767 to produce pcV-1604,which contains 1,678 nucleotides from the 5'-flanking regionof the chicken vimentin gene. A series of restriction frag-ments from the 837-bp sequence were subcloned blunt into aunique HindIII site upstream of the silencer element fused tothe heterologous herpes simplex virus thymidine kinasepromoter in the expression vector ptkCAT (11, 26). Tworestriction fragments, EcoRI-AvaII (75 bp) and HaeIII-RsaI(260 bp), were then subcloned blunt into a unique HindIIIsite of the vector ptkCAT minus the chicken vimentinsilencer element. Multiple copies of the 75-bp element weresubcloned into the analogous site in front of p2StkCAT andp4StkCAT. The orientation of each subclone was verified byDNA sequencing (35). Footprints and GMSAs were per-formed with the 837-bp fragment in pUC18.
Preparation of nuclear extracts. Nuclear extracts wereprepared from mouse L cells, HeLa cells, and varioustissues of the chicken embryo (11) as previously describedby Dignam et al. (10). HeLa cells were grown to a density of5 x 105 cells per ml in Spinner flasks. Nuclear extracts weretypically obtained from 3 liters of cells. Extracts werefractionated by ammonium sulfate precipitation. The proteinof interest was found in the 30 to 70% ammonium sulfatefraction and was desalted on a G-75 column (Sigma ChemicalCo.) equilibrated in buffer D (minus KCl). Fractipns werecollected, and those containing protein were pooled as
VOL. 12, 1992

2232 STOVER AND ZEHNER
determined by UV absorbance at A280. Crude extracts notsubjected to partial purification were adjusted to a finalconcentration of 0.1 M KCI. Further purification was per-
formed as described below. Extracts were applied to a
phosphocellulose column (3 by 5 cm; Whatman Inc.) equil-ibrated with buffer D minus KCl. The column was washedwith 3 column bed volumes of buffer D minus KCl. Proteinwas eluted from the column with a step gradient of buffer Dcontaining 0.1, 0.2, 0.3, 0.4, and 1.5 M KCl, respectively.Eluted fractions were precipitated with ammonium sulfateand desalted with Centricon 30 concentrators (Amicon) inbuffer D. All extracts were stored at -70°C for up to 6months.GMSAs. A radiolabeled fragment (1 to 2 ng) containing the
region -1604 to -1529 of the 5'-flanking region of thechicken vimentin gene was incubated with 0 to 20 ,g ofcrude nuclear extract or column fractions plus 1 ,ug ofpoly(dI-dC) in a final buffer concentration of 25 mM Tris (pH7.5), 6.25 mM MgCl2, 0.5 M EDTA, 50 mM KCl, 0.5 Mdithiothreitol, and 10% glycerol in 25 RI. Reaction mixtureswere incubated on ice for 15 min and then for 2 min at 22°C.Samples were loaded onto a 5% nondenaturing gel preparedin 0.25 x Tris-boric acid-EDTA. Electrophoresis was carriedout for 90 min at 10 V/cm. Gels were dried and placed on
XAR film overnight for visualization. Shifted bands were
analyzed and quantitated with a densitometer (tungstenlamp, 555 nm).DNase I footprinting. The aforementioned radiolabeled
DNA fragment (20 to 25 fmol) containing the 75-bp elementwas incubated in a 25-,ul solution containing crude nuclearextract or 0.3 M KCl column fractions in a final concentra-tion of 25 mM Tris (pH 7.5), 6.25 mM MgCl2, 0.5 mMEDTA, 50 mM KC1, 0.5 M dithiothreitol, and 10% glycerol.These reaction mixtures were incubated on ice for 15 minand then for 2 min at 22°C. Following incubation, 50 ,u of a
5 mM CaCl2-10 mM MgCl2 solution was added, and thereaction was initiated with the addition of 2.5 [lI of freshlydiluted DNase I (5 ,ug/ml; Sigma) for 1 min. Reactions were
terminated by the addition of 645 RI of 100% ethanol, 3 ,ug oftRNA, and 50 RI of saturated ammonium sulfate. The DNAwas precipitated and loaded onto a 6% polyacrylamide-8 Murea denaturing gel in Tris-boric acid-EDTA buffer andplaced on XAR film for visualization.SW blotting. SW blots were performed as described by
Singh et al. (39). A 50-,ug portion of nuclear extract or
column fraction was diluted 1:1 with sample resuspensionbuffer (2% sodium dodecyl sulfate [SDS], 100 mM Tris [pH7.5], 280 mM 2-mercaptoethanol, 20% glycerol, 0.002%pyronin Y), boiled for 5 min, and loaded onto an SDS-10%denaturing gel in a buffer of 50 mM Tris, 400 mM glycine,and 0.1% SDS. Electrophoresis was carried out at a constantcurrent of 25 mA. The gel was incubated for 30 min in geltransfer buffer (25 mM Tris, 190 mM glycine, 20% methanol)and then transferred to nitrocellulose. The transfer of pro-teins was carried out on a Bio-Rad transfer cell apparatus at0.5 A for 2 h in gel transfer buffer. Transfer of proteins wasmonitored by the use of prestained molecular weight stan-dards (Sigma). The nitrocellulose filter was blocked for 1 h atroom temperature in a solution containing 5% Carnationnonfat dry milk, TNED (50 mM Tris [pH 7.5], 50 mM NaCl,0.1 mM EDTA), and 1 mM dithiothreitol for 2 to 5 min andthen placed in a sealed plastic bag containing 20 ml of TNEDand 106 cpm of probe (either a radiolabeled 40-bp silencerelement or a radiolabeled 75-bp antisilencer element) per ml.The nitrocellulose filter was washed two times with TNED
zoo RI AP-1pcV-1604 5 -GAATTCATGCCAAGAGCACATATGGCAGCACFGACAACATATGAAAATTATATCCAA-3'
Avail5' -AACAAGAGGGAACGAGGTCCTGCTTCTTTACCAGAACAAACACCTCCCTGAATTTCAGGG-5'
5' -GCAGAGCATTGTCGGAGTGAAGGATGGCAGTTCGACATGCGTGCCATAAAGGTTTTGTGG-3'
Has III SPfr5' -AGCATAAACAAACTGTGGGCCATTAAGAGAAGCAGATGGAGCCAAGTGCACCTTTGG A-31
--P-2 JLluXS '-;CTCTGTGACAGCTCCCGGTGTGCAGAGTGCAGCATCTTCCCTCGGGGAGCCGC-3'
SP-15' -TTACT GTGCTTTTTGCCTGTGTTCCTTTACTTATCATTGCCATCAGACTT-3'
5 -TAAAC ACCACGGCC CACCGCCAGACAACCAAAAATCTCCTGCTAC-3'
Rsa I5' -TTCCTTAATAAATATACGGGCTCCGATGCTTCCCATAAGGTACAACACCCGAAGGCAATA-3'
rAP-25' -GCCAACCCTACTGTGAGCCAACCATACTGGIGAACATCACCTCACTCCTTGTGA-3'
OCT5' -TATCTTGCAGATAfTGCATGTCAGTCACTTTTAGAGCAAGGAAGAAAATGGTGT-3'
5 -CACCAATC&GGTGTGTCAGAGTGGGATTGCTCCCTGGGCGAAcE3rGGGCTTTACG-3'
5' -CTCCTAAGGCAGAGATCTCACCACCTCCCTTACACCACTCTGCATTCTCGCTGAAAAGTG-3'
AP-2 SP-1S5|dGTGTGGGGG+GTTTTTCACTGCTGCTCAGGCTTTT:AAAGTTATGCCACTGCTA-3 t
3E=1 se-l llKind III5' -TTGGTGCCTCTGTCTCGCTGc~ GCATAGCGCAGTTGCAGTAGAAGCTT-3' pcV-767
FIG. 1. Nucleotide sequence of an 837-bp fragment from the5'-flanking region of the chicken vimentin gene. This fragment wasobtained from a chicken genomic Charon 4A bacteriophage (clone1A) and cloned upstream of pcV-767 to produce pcV-1604. Shown isthe nucleotide sequence of the 837-bp fragment as determined by themethod of Sanger et al. (35), with slight modifications (see Materialsand Methods). Common sequences were identified by sequenceidentity to other known consensus binding sites and are marked asfollows: AP-1, SP-1, AP-2, and OCT. Restriction enzymes used togenerate various 5'-end-StkCAT constructs as described in Materi-als and Methods are noted.
for a total of 20 min. The blot was allowed to dry at roomtemperature and placed on XAR film for visualization.
Northern (RNA) analysis. Vimentin mRNA synthesis wasanalyzed during myogenesis in ovo (30) and in various tissueculture cell lines by Northern analysis (6). Total RNA wasisolated by guanidine isothiocyanate extraction. Cells werehomogenized with a Dounce homogenizer (10 strokes with atype B pestle) and, after the addition of a 2% Sarkosylsolution, passed through a 23-gauge needle to shear chromo-somal DNA. The suspensions were loaded into BeckmanQuick Seal tubes, and a 5.7 M CsCl-10 mM EDTA (pH 7.0)solution was layered below each suspension to form a CsClcushion. Samples were pelleted in a Dupont 65.13 rotor for16 to 20 h at 40,000 rpm at 25°C. Total RNA (3 ,ug) was thenanalyzed by using 6.6% formaldehyde-1% agarose gels (23),transferred to nitrocellulose (44), and hybridized with anick-translated human vimentin cDNA probe (a kind giftfrom D. Bloch).
RESULTS
Our goal in this study was to identify putative upstreamelements which can overcome vimentin's silencer elementsin the chicken vimentin gene. Ultimately, the identificationof all cis-acting elements and their cognate trans-actingfactors will be crucial for understanding the complex expres-sion pattern of the vimentin gene.
Analysis of the nucleotide sequence. As a first step in theanalysis of distal DNA elements, the DNA sequence of an837-bp upstream fragment (from -1604 to -767) was deter-
MOL. CELL. BIOL.

VIMENTIN DEVELOPMENTAL GENE EXPRESSION 2233
mined (Fig. 1) and then analyzed for homologies to se-quences of known enhancer elements present in GenBank.Of particular interest was a 7-of-8-bp match to the AP-1consensus sequence. There are other putative AP-1 se-quence identities within this 837-bp fragment, but theirmatch is only 5 of 8 bp or less, and the significance, if any,of these elements is presently under investigation. Since thehuman vimentin gene contains two tandem AP-1 sites whichhave been demonstrated to be important for serum andphorbol ester induction, we were interested in the possibleeffects of this single AP-1 site on expression of the chickenvimentin gene. Restriction sites used to fuse various frag-ments 5' to the CAT reporter gene (p8CAT) are shown.
Transient transfection of 5'-flanking chimeric constructs.To determine which elements were required for vimentingene expression, various 5'-flanking chimeric constructswere generated and transiently transfected into mouse Lcells (Fig. 2A). A proximal enhancer element was discoveredin the first -321 nucleotides from the start site of transcrip-tion which yielded a 59-fold CAT activity over that of thepromoterless p8CAT construct (11). As previously reported,this proximal enhancer element is tissue specific and yieldshigh CAT activity only when transfected into cells of mes-enchymal origin (36, 37). Construct pcV-608 yielded a CATactivity of only fourfold, a 93% drop from the activityobtained with the proximal enhancer element alone. Thesilencer element had been previously localized to a specific40-bp sequence (11), three copies of which have been foundwithin pcV-608, as determined by sequence homology andfunctional CAT assays (unpublished results). This 40-bpelement was also found to bind a silencer protein of 95 kDa,as reported by Farrell et al. (11). When an additional 837-bpfragment is incorporated into pcV-767 to yield pcV-1604,CAT activity increased 32-fold. This represents a 51% in-crease over the activity for the silencer elements withinpcV-608. Therefore, construct pcV-1604 contains an ele-ment(s) capable of overcoming the silencing elements lo-cated in pcV-608.
In defining this upstream element, various restrictionfragments from -1604 to -767 were fused to the 40-bpsilencer element in the heterologous construct ptkCAT.Each construct was transiently transfected into mouse Lcells (Fig. 2B). Construct ptkCAT yielded a CAT activity of46-fold above that of the promoterless construct p8CAT. Aspreviously reported, a single copy of the 40-bp silencerelement (pStkCAT) decreased CAT activity 35% (11). Sim-ilarly, in this study, a 35% drop in CAT activity wasobserved when the 40-bp silencer was present. When theentire 837-bp fragment was fused to the silencer element(p837/pStkCAT), 5/16 of pStkCAT inhibition was relieved(Fig. 2B). Two restriction fragments, a 75-bp element(EcoRl-AvaII) and a HaeIII-HindIII fragment (-1404 to-767) from the 837-bp upstream region, were subcloned 5'to the negative element in pStkCAT. Both of these restric-tion fragments appeared to overcome the silencer effect ofpStkCAT (Fig. 1 and 2B). When the 75-bp element waspresent in either a sense or antisense orientation, CATactivity was restored to 100% of ptkCAT activity, i.e., 45-and 46-fold, respectively. The second restriction fragment, aHaeIII-HindIII fragment (Fig. 1), restored approximately69% (11/16) of pStkCAT inhibition. Further restriction frag-ment constructs defined this second element as a 260-bpelement within the HaeIII-HindIII fragment (data notshown).To assess whether both the 75- and 260-bp elements were
true enhancers, the elements were further subcloned into the
Construct
p8CATpcV-321pcV-608pcV-767
pcV-1604
..
..I
-34L_CM
..
-7:E.161
I_~~B
C.
Mouse L celsFOLD (pmollb-al)
1
59'4)±254(9)± 24(9)± 2
32()± 2
Construct Mouse L cellsFOLD (pmollb-al)
p8CATptkCATpStkCATp837 IStkCAT
P75F IStkCATp75 R IStkCATpHae-H3 RIStkCAT
Construct
1
46(8) ±16
30(14)±1335() ±1045(s) ±846(5s ±9
41(9 ±9
Mouse L cellsFOLD (pmollb-gal)
p8CAT 1
3 ptkCAT 46(8) +16
PP75FItkCAT 42(6) + 7
E3{CAT p75RItkCAT 39(6) + 9
G 31 P26ORItkCAT 45(4) + 6
= g P75FI260 F ItkCAT 37(4) +11
FIG. 2. Transient transfections of various 5'-flanking chimericconstructs into mouse L cells. CAT activity is expressed as pico-moles of ['4C]chloramphenicol acetylated per minute per microgramof protein per unit of P-galactosidase (b-gal) activity. Fold denotesthe induction of CAT activity relative to the promoterless p8CATvector ± standard deviation. The superscript represents the numberof assays performed with each construct. The subscripts F and Rdenotes fragment placement in the sense (F) or antisense (R)orientation as found in the chicken vimentin gene. (A) Transfectionof the various chicken vimentin 5'-flanking deletion constructs; (B)transfection of pStkCAT constructs including the silencer elementplus various restriction fragments or the entire 837-bp element (seeFig. 1) fused 5' to the silencer element; (C) restriction fragmentsfrom Fig. 1 fused to the ptkCAT vector without the silencer element.The 260-bp element extends from the HaeIII site at -1404 to a Rsasite at -1144. HSV tk, herpes simplex virus thymidine kinasepromoter.
ptkCAT vector in which the silencer element is absent.When a single copy of the 75-bp element was cloned 5' to thepromoter in either a sense or antisense orientation, there wasno increase in CAT activity (Fig. 2C). Similarly, multiplecopies of the 75-bp element did not increase CAT activityabove that of ptkCAT. In fact, increasing copies of the 75-bpelement appeared to slightly repress CAT activity (data notshown). The 260-bp element also did not activate geneactivity. Next, the 75- and 260-bp elements were subclonedtogether to assess whether they could function in concert.
VOL. 12, 1992
2234 STOVER AND ZEHNER
(.X
CL0
40~
~i~E
1 J~[
p pp
FIG. 3. In vivo effects of the silencer and antisilencer elementson CAT gene activity. Multiple copies (zero, one, two, or four) ofthe silencer element (S) minus or plus increasing copies of the 75-bpantisilencer element (checkered oval) are fused to the thymidinekinase promoter (arrow) in front of the CAT gene (C). Results areexpressed as percent activity of ptkCAT and are compiled from 6 to14 transient transfection assays in mouse L cells. The standarddeviation is shown as a line above each bar.
No increase in CAT activity was obtained; instead, de-creased activity was observed in this chimeric construct(Fig. 2C). It appears that the 75- and 260-bp elements do notfunction as true enhancers, since they do not enhancetranscription but rather function only when the silencerelement is present. Therefore, we will refer to these ele-ments as antisilencers.
Previously, we showed that increasing copies of the si-lencer element resulted in increased silencing; i.e., twocopies yielded a 66% reduction, whereas four copies reducedCAT activity 90% (11). Next, we determined whether mul-tiple copies of the 75-bp antisilencer element were requiredto overcome the additive negative effect of multiple silencerelements (Fig. 3). As before, one copy of the silencerelement reduced transcription 35%, and the addition of asingle 75-bp element completely restored CAT activity.Again, no enhancement of CAT activity was noted with the75-bp element alone. However, with increased copies (eithertwo or four) of the silencer element, multiple copies of theantisilencer element were required to restore reporter geneactivity.The 75-bp element binds a specific protein(s). Transfection
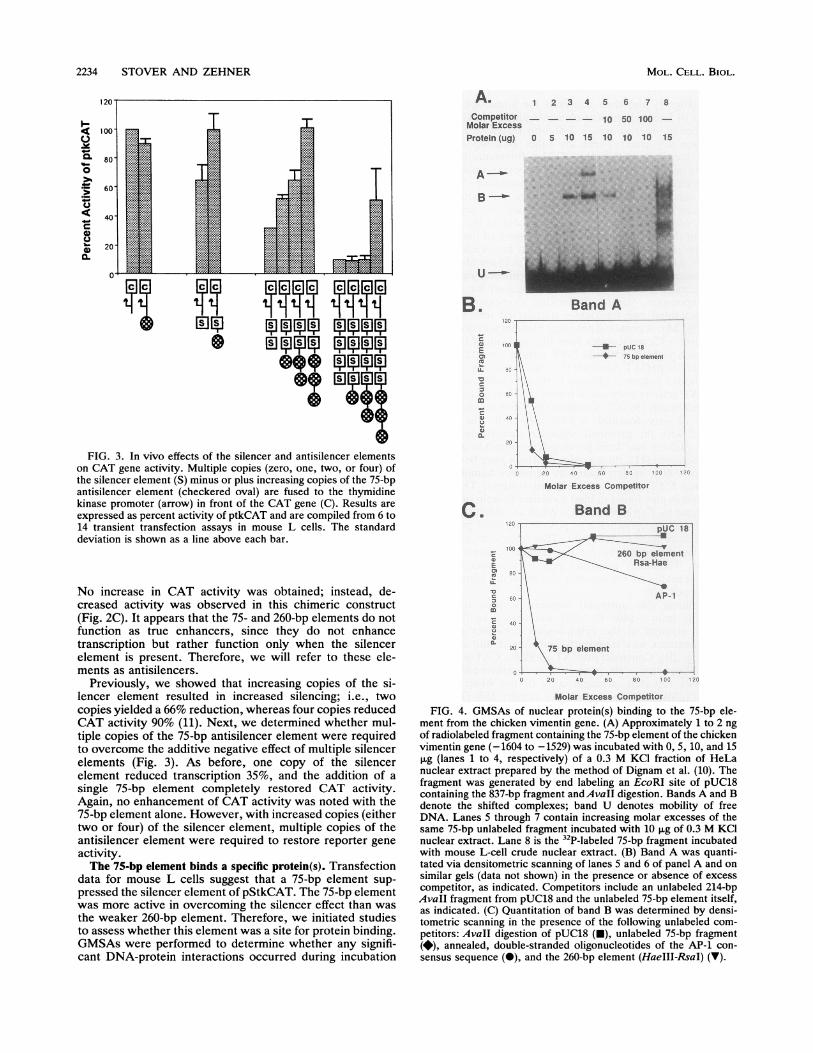
data for mouse L cells suggest that a 75-bp element sup-pressed the silencer element of pStkCAT. The 75-bp elementwas more active in overcoming the silencer effect than wasthe weaker 260-bp element. Therefore, we initiated studiesto assess whether this element was a site for protein binding.GMSAs were performed to determine whether any signifi-cant DNA-protein interactions occurred during incubation
A.CompetitorMolar ExcessProtein (ug)
A P
B
U
Be20
ICH
o <,oIC-<
Eil kc... I.,
m- i
O e i
0
C.
'en)ECD
u
20
1 2 3 4 5 6 7 8
- - - - 10 50 100 -
0 5 10 15 10 10 10 15
Band A
* puc 185---* 5bp element
Molar Excess Competitor
Band B
Molar Excess CompetitorFIG. 4. GMSAs of nuclear protein(s) binding to the 75-bp ele-
ment from the chicken vimentin gene. (A) Approximately 1 to 2 ngof radiolabeled fragment containing the 75-bp element of the chickenvimentin gene (-1604 to -1529) was incubated with 0, 5, 10, and 15,ug (lanes 1 to 4, respectively) of a 0.3 M KCI fraction of HeLanuclear extract prepared by the method of Dignam et al. (10). Thefragment was generated by end labeling an EcoRI site of pUC18containing the 837-bp fragment andAvaIl digestion. Bands A and Bdenote the shifted complexes; band U denotes mobility of freeDNA. Lanes 5 through 7 contain increasing molar excesses of thesame 75-bp unlabeled fragment incubated with 10 ,ug of 0.3 M KCInuclear extract. Lane 8 is the 32P-labeled 75-bp fragment incubatedwith mouse L-cell crude nuclear extract. (B) Band A was quanti-tated via densitometric scanning of lanes 5 and 6 of panel A and onsimilar gels (data not shown) in the presence or absence of excesscompetitor, as indicated. Competitors include an unlabeled 214-bpAvaIl fragment from pUC18 and the unlabeled 75-bp element itself,as indicated. (C) Quantitation of band B was determined by densi-tometric scanning in the presence of the following unlabeled com-petitors: AvaII digestion of pUC18 (-), unlabeled 75-bp fragment(*), annealed, double-stranded oligonucleotides of the AP-1 con-sensus sequence (0), and the 260-bp element (HaeIII-RsaI) (v).
MOL. CELL. BIOL.

VIMENTIN DEVELOPMENTAL GENE EXPRESSION 2235
with soluble nuclear extracts from mouse L or HeLa cells(Fig. 4A). The two nuclear extracts yielded a common band(B) (Fig. 4A, lanes 3 to 5 compared with lane 8). Since HeLacells are a reliable source for nuclear extracts and have beenused previously for transcription factor purifications, wechose to perform most of our in vitro binding studies withHeLa nuclear extracts. HeLa cells express vimentin, asmonitored by Northern analysis, and contain the silencerprotein as well as other proteins which bind vimentin'sproximal enhancer and promoter elements, as we havepreviously shown by in vitro binding assays and functionaltransfection studies (11, 36).
Since several proteins were observed to bind to the 75-bpelement in crude HeLa nuclear extracts (data not shown) ormouse L-cell extracts (Fig. 4A, lane 8), ammonium sulfatefractionation was performed as described in Materials andMethods. The precipitate from the 30 to 70% ammoniumsulfate fraction was desalted by using a G-75 matrix andloaded onto a phosphocellulose column. Putative DNA-binding proteins were eluted from the column by increasingthe salt concentration and assayed for binding activity viaGMSAs. The optimal complex formation occurred with the0.3 M KCI fraction (data not shown). Incubation of the HeLa0.3 M KCI fraction with the 75-bp element yielded twodistinct complexes, bands A and B (Fig. 4A). Band A wasdetermined to be nonspecific, since it competed with both anunrelated DNA fragment (not containing the 75-bp element)and a 75-bp fragment (Fig. 4B). Also, band A was notpresent in the crude nuclear extracts from mouse L cells(Fig. 4, lane 8). Band B was determined to be specific, sinceaddition of an unrelated DNA fragment (either a fragmentfrom pUC18 or the 260-bp RsaI-HaeIII fragment from vi-mentin's 5' end) did not compete with the 75-bp element,while incubation with the unlabeled 75-bp element did com-pete for binding (Fig. 4C). Band B was found both inpartially purified HeLa and crude mouse L-cell nuclearextracts (Fig. 4A, lanes 3 to 5 compared with lane 8).The human distal enhancer element consists of two tan-
dem AP-1 binding sites, and the 75-bp chicken elementcontains one copy of the AP-1 consensus sequence (7-of-8-bp match). Therefore, the 75-bp fragment was competedfor with an excess of the AP-1 consensus sequence todetermine whether binding was due to an AP-1 like pro-tein(s). Two oligonucleotides which contained the consensussequence for AP-1 [(G/C)TGA(C/G)TCA] and its comple-ment were synthesized. In GMSAs, a fragment containingthe annealed double-stranded AP-1 binding site was incu-bated with the 32P-labeled 75-bp element (Fig. 4C). Nocompetition was exhibited at a 20 M excess; at a 100 Mexcess, however, there was a 30% decease in complexformation of band B. Under these conditions, AP-1 doesseem to compete slightly with the 75-bp element but only ata high molar excess, the physiological relevance of which isunknown. The question of whether or not the 75-bp elementfunctions through an AP-1-like protein is fully resolved withthe SW analysis and footprint protection experiments dis-cussed below (Fig. 5 and 6).To determine whether the 75- and 260-bp antisilencer
elements function through the same protein(s), incubationwith the unlabeled 260-bp element (RsaI-HaeIII fragment)was performed in GMSAs (Fig. 4C). Band B showed nocompetition at even a 100 M excess with the unlabeled260-bp fragment. This result suggests that the distal 75- and260-bp elements do not function through the same protein(s).
Identification of a putative binding site in the 75-bp elementby DNase I footprinting. DNase I footprinting experiments
were conducted to determine which nucleotides are involvedin DNA-protein binding. A 32P-labeled DNA fragment con-taining the 75-bp element was incubated with both crude and30 to 70% ammonium sulfate fractions of HeLa cell nuclearextracts. Binding was allowed to continue for 15 min on ice,a freshly diluted solution of DNase I was added, anddigestion was stopped after 1 min (Fig. 5). A specificDNA-protein complex showed protection from -1604 to-1587, a 17-bp region on the noncoding strand (Fig. 5A). Ahypersensitive site was noted within the consensus AP-1binding site (Fig. 5A, arrow). Binding of a protein(s) to theAP-1 consensus sequence seems unlikely, since no protec-tion was observed within this region. Maximal bindingoccurs in the crude nuclear extracts at 40 ,ug, whereasmaximal binding required twice that amount of protein in the30 to 70% ammonium sulfate fraction. A comparison be-tween the DNase protection patterns in the crude fractionversus the 30 to 70% ammonium sulfate fraction suggeststhat there may be an additional protein(s) which favors theDNA-protein formation in the crude fraction.On the coding strand, a 10-bp element beginning at -1597
to -1587 was protected from DNase I digestion. A poorcleavage pattern was obtained when the 75-bp fragment waslabeled on the sense compared with the antisense strand(Fig. SA). However, the cleavage of two bands at positions-1597 and -1587 was protected. Although the footprintexhibited by this strand is weak, protection is assumed tooccur between these two bands. Again, a hypersensitivitysite occurs at -1598 within the AP-1 site as well as at theadenine residue at -1598 immediately adjacent to the pro-tein-protected region. Once again, better protection oc-curred with 40 ,ug of the crude HeLa nuclear extract,whereas comparable protection required 80 ,ug of the ammo-nium sulfate fraction. In summary, these data suggest that invitro a protein binds to a smaller 10- to 17-bp element withinthe 75-bp element not inclusive of the AP-1 consensussequence. The sequence 5'-ATTCATGCCAAGAGCA-3'was protected in these experiments.The 75-bp element binds a 140-kDa protein. To determine
the molecular size of the protein which binds to the 75-bpelement, SW analysis was performed. Previously, weshowed by this method that the silencer element in thechicken vimentin gene bound a 95-kDa protein (11). Proteinsfrom the 0.3 M KCI fraction were separated on an SDS-10%denaturing polyacrylamide gel and immobilized onto nitro-cellulose. The presence of a specific protein was analyzed byprobing with the DNA binding site. The 75-bp elementbound a protein from HeLa cell extracts of approximately140 kDa compared with molecular size standards (Fig. 6A).Nuclear extracts from various cell lines were used to
assess the presence of the 140-kDa protein (Fig. 6A). Com-parisons were made by preparing nuclear extracts fromequivalent cells and performing SW blots with equalamounts of total nuclear protein. The analysis of equivalentamounts of nuclear protein per lane was verified by densit-ometric scanning of Coommasie blue-stained, SDS-10%denaturing polyacrylamide gels. Little variation in proteinbands was detected in the various nuclear extracts. Previ-ously, SW blots of nuclear and cytosolic extracts verifiedthat the majority of the 95- and 140-kDa proteins werepresent in the nuclear fraction (unpublished results). BothHeLa and mouse L-cell nuclear extracts contained the140-kDa protein (Fig. 6A). This 140-kDa protein appeared tobe active in binding the 75-bp element in a tissue-specificfashion. Fibroblasts such as mouse L and Swiss 3T3 cellswhich express vimentin exhibit a good hybridization signal
VOL. 12, 1992

2236 STOVER AND ZEHNER
Crude
A+G 0 20 40 A+G
B.
Crude
A+G 0 20 40 A+G
30 - 70 % AS
A+G 0 20 40 80 0
Ap-I
..... .....
rrsDU!
* =,l=ee ..aSii/9 _d: *__-.4Z
.. .!_,
S.e82n_-Fs *w
i :.| t.6
;4siit. _
sveRXI x5i
I ....|0I .; ".N N| .:t =
. _= .-_ Siense
30 - 70 %HE ASFA+Gl 0 20 40 80 0
A.l.m.:
*'
.;L_
a:p-X.s a - ~Ap-
31 CTTAAGTACGGTTCTCGTATACCGTCGTGAGTGTpTTGTA-5lwV_1fj -IL !co-I Ap-IpcV-1I875' -GAA'CA'GCCAAGA(iCA|'A'l'A'l'(; GCA(;(CAC'R;ACACAAACAT' 3'
pcV-1604 Ap-IpcV-1597 pcV-1587
FIG. 5. DNase I footprints of the 75-bp element with crude and fractionated HeLa cell nuclear extract. A 24-fmol radiolabeled portion ofthe 75-bp element was incubated with increasing amounts of both crude and 30 to 70% ammonium sulfate fractions of HeLa cell nuclearextract, as noted. Reaction mixtures were digested with 2.5 ,A of DNase I (freshly diluted to 5 ILg/plA) and analyzed on an 8% polyacrylamidedenaturing gel. The A+G sequencing ladder was generated by the method of Maxam and Gilbert (24). Antisense (A) and sense (B) strandsare shown, and nucleotide positions are as indicated. The nucleotides protected from DNase I are boxed in the sequences at the bottom. Inpanel B, the unlabeled arrow denotes a hypersensitivity site immediately adjacent to the protected region. Hypersensitivity sites present inthe consensus AP-1 sequence are demarcated with arrows.
for the 140-kDa protein. Chicken crystalline lens extractsfrom day 14 embryos also contained the 140-kDa protein(Fig. 6A), while no evidence for the silencer protein has beendetected in this extract (unpublished observation). Previ-ously, it has been established that vimentin is expressed inthe chicken lens despite its epithelial origin (37).The 140-kDa protein may be developmentally regulated
during myogenesis. The interplay between the 140- and95-kDa proteins was investigated in a developmental study.Previously, we and others have shown that the expression ofvimentin mRNA decreases during myogenesis (4, 11). Fur-thermore, Farrell et al. have correlated this decrease invimentin mRNA to an increase in binding of the 95-kDasilencer protein (11). In this previous study, chicken eggswere incubated from fertilization to 9 days posthatch. Nu-clear extracts and mRNA were prepared from various mus-cle tissues at days 10 to 11 and day 17 in ovo and 1 dayposthatch as described previously (11). These same nuclearextracts were probed with the 75-bp element via SW blots,and the binding activity of the 140-kDa protein decreasedduring myogenesis (Fig. 6A). Both Northern and SW blotswere scanned via densitometry, and the data for gizzardmuscle are collated in Fig. 6B. As previously reported, the91% decrease in vimentin mRNA levels (Fig. 6B, inset)correlated with a 90% increase in the binding activity of thesilencer protein (11). However, the binding activity of the140-kDa antisilencer protein in the same extract diminished51%. At 3 days posthatch, there was a weak signal for the
140-kDa protein in skeletal muscle (Fig. 6A), in which astrong signal for the 95-kDa silencer protein was reported byFarrell et al. (11).
DISCUSSION
Multiple positive and negative factors have been shown byseveral laboratories (11, 31, 33, 37, 45) to contribute tovimentin's unique expression pattern. We have been study-ing the elements required for vimentin expression in mesen-chymal cells such as fibroblasts, in which expression is high,and in muscle, in which gene expression is turned off duringmyogenesis. Previously, we have detected various promoterand proximal enhancer elements in addition to a silencerelement, all of which contribute to vimentin's expressionpattern (11, 36). We have detected a 95-kDa protein whichbinds to the silencer element and is present in many celltypes, including fibroblasts, in which vimentin is expressed.This finding led us to believe that a positive element(s) andfactor(s) may exist (upstream of pcV-767 in the chickenvimentin gene) which are capable of overcoming this nega-tive element, thereby permitting the vimentin gene to bedifferentially expressed. Here, we have sought to definethese elements and how their activities may contribute tovimentin's differential expression pattern.
Sequence analysis of the chicken vimentin 5'-flankingsequence -1604 to -767 (Fig. 1) reveals several homologiesto known consensus sequences: AP-1, Sp-1, OCT-1, and
A.Antisense
mini
4w
8 .s
"._._
m
:.
so =to
MOL. CELL. BIOL.
pCV-im_W%

VIMENTIN DEVELOPMENTAL GENE EXPRESSION 2237
A.11 Ia S%xiss 3T.f (Cgjard
M louse 1. Lens 17d14d17d 3 1)
..
MW (k Da)
-180
-116
- 89
- 58
-48
-36
B.
>ll,
--
;~l-
/'C
~; ;z
i 7
In - IIX das- 14 dav 17 - IS dav
FIG. 6. SW analysis of nuclear extracts isolated from variouscell lines and tissues. All nuclear extracts were probed with a32P-labeled 75-bp element from -1604 to -1529. (A) A 50- g portionof various nuclear protein extracts was separated on a 10% dena-turing polyacrylamide gel and transferred to nitrocellulose filters asdescribed in Materials and Methods. Mouse L cells, Swiss 3T3 cells,and HeLa cells were obtained from tissue culture; other extractswere obtained from various chicken tissues at various stages ofdevelopment as described previously (10, 11). d days during in ovodevelopment; D, days posthatch. (B) Densitometric scans to deter-mine the binding activity of the 140-kDa protein and the 95-kDasilencer protein (11). Vimentin mRNA expression was quantitated inthe 11-day embryo and 1 day posthatch (adult) samples by Northernanalysis (inset).
AP-2 (2, 21, 43, 45). To identify which cis-acting sequenceswere required for restoring gene activity, we fused variousDNA fragments from the 837-bp region to pStkCAT. Adetailed analysis of this 837-bp region revealed two newdistal elements, a 75- and a 260-bp element, both capable ofovercoming the silencer element to various degrees (Fig. 2).The stronger 75-bp element restores gene activity by 100%,while the weaker 260-bp element yields a 69% increase.Curiously, the entire 837-bp fragment fused to pStkCATdoes not restore gene activity to 100% (Fig. 2B). Similarly,pcV-1604 produces CAT activity only 32-fold above that ofp8CAT, whereas pcV-321 produces a 59-fold increase (Fig.2A). These data suggest that there may be some othersilencer element(s) present within the region -1529 to -767.If so, they would be distinct from the original 40-bp silencerelement located at -568, as little if any sequence homologycan be found. Studies are under way to determine whethersuch a putative negative element(s) exists.When either the 75-bp or the weaker 260-bp element is
fused to ptkCAT minus the silencer element, there is noobserved increase in CAT activity (Fig. 2C). To determinewhether there was any interplay between the 75- and 260-bpelements, we investigated whether these two elements could
cooperate more effectively together than the entire 837-bpfragment fused to ptkCAT. However, this construct yieldedno transcription above that of ptkCAT alone (Fig. 2C). Weinterpret this result to mean that the distance separatingthese elements is crucial. To address this question, we arenow using polymerase chain reaction techniques to con-struct specific deletion mutants plus and minus the 75- and260-bp elements within the context of the chicken vimentingene. Typically, synergistic activity occurs when enhancersare spaced close together (23), but in this case, placing twotandem copies of the 75-bp element or the 75-bp plus the260-bp element did not display any synergism (Fig. 2C). Thisfinding suggests that the protein which recognizes the 75-bpelement may interact directly with the silencer elementand/or its protein to reactivate transcription.
It appears that these two elements are not in themselvestrue enhancers but rather function to overcome the silencerelement of the chicken vimentin gene. These elements do notact in a synergistic manner, they do not activate transcrip-tion of a heterologous promoter, and their activity dependson the presence of another element. It has recently beenreported that the GAGA factor, which binds a GA-richsequence of the Drosophila Ultrabithorax and Kruppelgenes, is by itself a weak repressor (7). Since this elementwas found to overcome the repression of a general repressor,Hi, and could activate transcription only when the Hiprotein was present, it was termed an antirepressor (7).Because the 75- and 260-bp elements selectively restoredgene activity only when a specific 40-bp silencer element waspresent, we chose to call these elements antisilencer ele-ments.
Interestingly, increasing copies of the silencer elementrequire a corresponding increase in the number of 75-bpantisilencer elements, implying a possible equimolar associ-ation of trans-acting factors and DNA elements (Fig. 3). Thisis particularly relevant for the chicken vimentin gene, inwhich three such silencer elements are found. The ability ofeach copy to repress transcription is now under study. Atthis time, we can say that all three copies are active but todifferent degrees. Within pcV-1604 there is only one copy ofthe 75-bp element, which may account for pcV-1604 beingonly one-half as active as pcV-321. This has prompted us tobegin analyzing the DNA upstream of -1604 in order todetermine whether there are any additional copies of the75-bp element which could promote even higher gene activ-ity.The human vimentin gene contains a 48-bp element (-713
to -664) which consists of an Sp-1 site flanked by two AP-1sites. This element was demonstrated to confer serum andphorbol ester inducibility on the human vimentin gene. Inaddition, this element served as an enhancer by increasingCAT activity independent of its relative position and orien-tation (33). Since the chicken vimentin gene also contains asingle AP-1 site (7-of-8-bp match) within the 75-bp antisi-lencer element (-1574 to -1566), we were interested in thepossible effects of this AP-1 site in the regulation of thechicken vimentin gene. We observed several differencesbetween the chicken distal antisilencer elements and thehuman distal enhancer. The chicken vimentin gene containstwo subregions, a 75-bp and a 260-bp element, both of whichwere capable of overcoming the chicken vimentin silencer tovarious degrees (Fig. 2). Our results suggest that the AP-1site is not involved in antisilencer activity for several rea-sons. First, GMSAs show that a specific protein binds to the75-bp element, which does not compete with either theconsensus AP-1 binding site or the 260-bp element (Fig. 4C).
i-
-1
VOL. 12, 1992

2238 STOVER AND ZEHNER
This finding suggests that the two subregions functionthrough separate proteins which are not AP-1-like proteins.Second, footprinting experiments further support the exist-ence of a protein which binds to the 75-bp element. Protec-tion was observed on both the sense and antisense strands,not inclusive of the AP-1 site (Fig. 5), although DNaseI-hypersensitive sites were observed on both strands withinthe AP-1 sequence. Third, SW blots indicate that the molec-ular size of the 75-bp antisilencer protein is 140 kDa (Fig. 6).The AP-1/Jun-like proteins usually consist of a family ofproteins ranging in size from 36 to 52 kDa (26). From theseexperiments, we conclude that the antisilencer effect of the75-bp element does not depend on an AP-1 like protein(s).Furthermore, this is not the same element as described forthe human vimentin gene, for which the 48-bp element wasrequired to induce vimentin's response to phorbol ester andserum. We conclude that in the chicken vimentin gene, the75-bp antisilencer element is the fibroblast-specific elementcapable of overcoming chicken vimentin's silencer element,thereby permitting vimentin to be expressed in cells ofmesenchymal origin in the presence of the 95-kDa silencerprotein. Whether or not this element is the phorbol esterand/or serum-inducible element has yet to be determined.We are currently conducting experiments to delineate thelocation of this element.The 140-kDa protein which recognizes the 75-bp antisi-
lencer element has been found in HeLa cells, mouse L cells,Swiss 3T3 cells, and various muscle tissues during develop-ment (Fig. 6). During gizzard or skeletal myogenesis, vimen-tin mRNA has been shown to decrease by 91% as thechicken embryo develops to an adult (Fig. 6B, inset). Farrellet al. (11) have noted an inverse relationship in the level ofvimentin mRNA and the binding activity of the silencerprotein, which increased 90% in gizzard muscle from day 10to day 18 in ovo. We observe a direct correlation in thedecrease of vimentin mRNA and the binding efficiency of the140-kDa protein in these same nuclear extracts. As vimentinmRNA decreases during myogenesis, the apparent bindingefficiency of the 140-kDa antisilencer protein also decreasedby 51%. This finding suggests that the interplay betweenthese two proteins is important for vimentin's developmentalregulation, as the binding activity of these regulatory pro-teins correlates with the expression of the chicken vimentingene.From these studies, it appears that vimentin mRNA levels
can be directly correlated to the binding activity of regula-tory factors which recognize specific sequences in the 5'-flanking sequence of the chicken vimentin gene. The mech-anism by which these multiple elements act in concert iscomplex. Adding to this complexity is the fact that the140-kDa antisilencer element appears to function only wheqthe silencer element is present. Several models may accountfor the differential binding activity of the 95-kDa silencerprotein and antisilencer binding protein. Recently, it wasreported that in the glucagon gene, a proximal promoterelement (Gl) was necessary for two distal enhancerlikeelements (G2 and G3) to function (32). When G1 was mutatedand nonfunctional, both G2 and G3 could not activate tran-scription. The authors hypothesized that the proximal G1element was a relay protein allowing for close interactionbetween these upstream elements and the transcriptionalmachinery. When the distal elements, G2 and G3, weremoved close to the promoter, G1 was no longer needed, forthese two elements could enhance transcription directly.The involvement of the 95-kDa silencer protein within the
_D CMCKEN-1597 -608 - 321 -160
-362 -353-713 -476D-322-225-882 -853- 713 -476 -322 -22
0-i
- 1724
1 HIVLTR
-249
-77s t ~~~-77*RATGROWVTHHOfRMONE
LYSOZYME-m
da a RATINSULIN 1tw^-2300 - 1444
FIG. 7. Comparison of the 5'-flanking regions of several eukary-otic genes which contain both silencer elements (shaded box labeledS) and sequence identities to the 75-bp antisilencer element (check-ered oval). Other elements of interest are as follows: the chickenvimentin proximal promoter region (P), proximal enhancer elements(E), and distal 260-bp element (260). The relative positions of thehuman vimentin tandem AP-1 sites (48 bp) and other negativeelements (N) (33) are as noted. HIV LTR represents the humanimmunodeficiency virus type 1 long terminal repeat. The arrowdenotes the transcriptional start point.
chicken vimentin gene may also be a type of relayingmechanism, the details of which are unknown at this time.A number of other possibilities could account for how the
distal antisilencer element(s) override vimentin's silencerelement. It is possible that the silencer protein serves toblock the requisite protein-protein interactions required fortranscriptional activation. In this capacity, the antisilencerelement may serve to modulate silencer-transcriptional ma-chinery interactions. Alternatively, the silencer elementitself may be involved in looping DNA in some nonfunc-tional constraint which is relieved by antisilencer binding. Itis also possible that auxiliary proteins are important inregulating silencer and antisilencer interactions. It is inter-esting that in footprinting experiments, crude nuclear ex-tracts exhibit better protection at lower amounts of proteinthan do the ammonium sulfate-precipitated fractions. Thisfinding suggests the involvement of a yet unidentified acces-sory protein similar to the increased enhancement of tran-scription when the oncogene fos protein is heterodimerizedto thejun protein (1). Furthermore, the complex expressionpattern of the chicken vimentin gene may not rely solely onone mechanism but rather require a combination of thesesuggested models. To determine which model plays a role invimentin expression, a more detailed study will be required.
Several other eukaryotic genes have been shown to con-tain sequence homologies with vimentin's silencer element.Further sequence analysis shows that in every case, thesegenes also contain the 75-bp antisilencer element and some-times more than one copy (Fig. 7). For example, the humanvimentin gene contains a functional silencer element as wellas sequence identity to the 75-bp antisilencer element. Twocopies of the antisilencer element are found upstream in thehuman immunodeficiency virus long terminal repeat gene
MOL. CELL. BIOL.
.222
m Aak
-150 -119
ISPik"

VIMENTIN DEVELOPMENTAL GENE EXPRESSION 2239
(34), both 5' and 3' to the silencer element in the rat growthhormone gene (20), and upstream of both the chickenlysozyme (3, 41) and rat insulin 1 (19) genes. It appears thatthe distance and orientation between the silencer and 75-bpelement are variable. The coexistence of the 75-bp antisi-lencer and silencer elements in the aforementioned eukary-otic genes suggests that this element is important for theregulation of other genes in addition to vimentin. Experi-ments are in progress to elucidate the mechanism by whichthe 75-bp antisilencer element overcomes the silencer ele-ment.
ACKNOWLEDGMENT
This work was supported by Public Health Service grant HL45422to Z.E.Z.
REFERENCES1. Abate, C., D. Luk, E. Gagne, R. G. Roeder, and T. Curran.
1990. Fos and Jun cooperate in transcriptional regulation viaheterologous activation domains. Mol. Cell. Biol. 10:5532-5535.
2. Angel, P., M. Imagawa, R. Chui, B. Stein, R. J. Imbra, H. J.Rahmsdorf, C. Jonat, P. Herrlich, and M. Karin. 1987. Phorbolester-inducible genes contain a common cis element recognizedby a TPA-modulated trans-acting factor. Cell 49:729-739.
3. Baniahmad, A., M. Muller, C. Steiner, and R. Renkawita. 1987.Activity of two different silencer elements of the chickenlysozyme gene can be compensated by enhancer elements.EMBO J. 6:2297-2303.
4. Capetanaki, Y. G., J. Ngai, and E. Lazarides. 1984. Regulationof the expression of genes coding for the intermediate filamentsubunits vimentin, desmin, and glial fibrillary acidic protein, p.415-434. In G. G. Borisy, D. W. Cleveland, and D. B. Murphy(ed.), Molecular biology of the cytoskeleton. Cold Spring Har-bor Laboratory, Cold Spring Harbor, N.Y.
5. Cattoretti, G., S. Andreola, C. Clemente, L. Damato, and F.Rilke. 1988. Vimentin and p53 expression on epidermal growthfactor receptor-positive estrogen receptor-negative carcinoma.Br. J. Cancer 57:353-357.
6. Chirgwin, J. M., A. E. Przybyla, R. J. MacDonald, and W. J.Rutter. 1979. Isolation of biologically active ribonucleic acidfrom sources enriched in ribonuclease. Biochemistry 18:5294-5299.
7. Croston, G. E., L. A. Kerrigan, L. M. Lira, D. R. Marshak, andJ. T. Kadonaga. 1991. Sequence-specific antirepression of his-tone Hi-mediated inhibition of basal RNA polymerase II tran-scription. Science 261:643-649.
8. Dellagi, K., W. Vainchenker, G. Vinci, D. Paulin, and J. C.Brouet. 1983. Alteration of vimentin intermediate filamentexpression during differentiation of human hemopoietic cells.EMBO J. 2:1509-1514.
9. Dente, L., G. Cesareni, and R. Cortese. 1983. pEMBL: a new
family of single stranded plasmids. Nucleic Acids Res. 6:1645-1655.
10. Dignam, J. D., R. M. Lebovitz, and R. G. Roeder. 1983.Accurate transcription initiation by RNA polymerase II in asoluble extract from isolated mammalian nuclei. Nucleic AcidsRes. 11:1475-1489.
11. Farrell, F. X., C. M. Sax, and Z. E. Zehner. 1990. A negativeelement involved in vimentin gene expression. Mol. Cell. Biol.10:2349-2358.
12. Franke, W. W., E. Schmid, M. Osborn, and K. Weber. 1978.Different intermediate sized filaments distinguished by immuno-fluorescence microscopy. Proc. Natl. Acad. Sci. USA 75:5034-5038.
13. Gorman, C., L. Moffat, and B. H. Howard. 1982. Recombinantgenomes which express chloramphenicol acetyltransferase inmammalian cells. Mol. Cell. Biol. 2:1044-1054.
14. Graham, F. L., and A. J. Van der Eb. 1973. A new technique forthe assay of infectivity of human adenovirus 5 DNA. Virology52:456-467.
15. Granger, B. L., E. A. Repasky, and E. Lazarides. 1982. Synemin
and vimentin are components of intermediate filaments in avianerythrocytes. J. Cell Biol. 92:299-312.
16. Kacmarek, L., B. Calabretta, and R. Baserga. 1985. Expressionof cell-cycle-dependent genes in phytohemagglutinin-stimulatedhuman lymphocytes. Proc. Natl. Acad. Sci. USA 82:5375-5379.
17. Klymkowsky, M. W., J. B. Bachant, and A. Domingo. 1989.Functions of intermediate filaments. Cell Motil. Cytoskel. 14:309-331.
18. Kuruc, N., and W. W. Franke. 1988. Transient coexpression ofdesmin and cytokeratins 8 and 18 in developing myocardial cellsof some vertebrate species. Differentiation 38:177-193.
19. Laimonis, L., M. Holmgren-Konig, and G. Khoury. 1986. Tran-scriptional "silencer" element in rat repetitive sequences asso-ciated with the rat insulin 1 gene locus. Proc. Natl. Acad. Sci.USA 83:3151-3155.
20. Larsen, P. R., J. W. Harney, and D. D. Moore. 1987. Repressionmediated cell-type-specific expression of the rat growth hor-mone gene. Proc. Natl. Acad. Sci. USA 83:8283-8287.
21. Lee, W., P. Mitchell, and R. Tjian. 1987. Purified transcriptionfactor AP-1 interacts with TPA-inducible enhancer elements.Cell 49:741-752.
22. Lendahl, V., L. B. Zimmerman, and D. G. McCay. 1990. CNSstem cells express a new class of intermediate filament protein.Cell 60:585-595.
23. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecularcloning: a laboratory manual. Cold Spring Harbor Laboratory,Cold Spring Harbor, N.Y.
24. Maxam, A. M., and W. Gilbert. 1980. Sequencing end-labeledDNA with base-specific chemical cleavages. Methods Enzymol.65:499-560.
25. McKnight, S. L., and R. Kingsbury. 1982. Transcriptionalcontrol signals of a eukaryotic protein coding gene. Science217:316-324.
26. Miksicek, R., A. Heber, W. Schmid, U. Danesch, G. Posseckert,M. Beato, and G. Shutz. 1986. Glucocorticoid responsiveness ofthe transcriptional enhancer of Moloney murine sarcoma virus.Cell 46:283-290.
27. Mitchell, P. M., and R. Tjian. 1989. Transcriptional regulation inmammalian cells by sequence-specific DNA binding proteins.Science 245:371-378.
28. Montgomery, K. T., J. Tardiff, L. M. Reid, and K. S. Krauter.1990. Negative and positive cis-acting elements control theexpression of murine alpha1-protease inhibitor genes. Mol. Cell.Biol. 10:2625-2637.
29. Nielsen, D. A., J. Chou, A. J. Mackrell, M. J. Casadaban, andD. F. Steiner. 1983. Expression of a preproinsulin-beta-galac-tosidase gene fusion in mammalian cells. Proc. Natl. Acad. Sci.USA 80:5198-5202.
30. Paterson, B. M., and B. Roberts. 1981. Structural gene identifi-cation utilizing eukaryotic cell-free translational systems, p.417-437. In J. G. Chirikjian and T. S. Papas (ed.), Structuralanalysis of nucleic acids, vol. 2. Elsevier North-Holland, Am-sterdam.
31. Paulin, D. 1989. Expression of the genes coding for the humanintermediate filament proteins. Pathol. Biol. 37:277-282.
32. Philippe, J., and S. Rochat. 1991. Strict distance requirement fortranscriptional activation by two regulatory elements of theglucagon gene. DNA Cell Biol. 10:119-124.
33. Rittling, S. R., L. Coutinho, T. Amram, and M. Kolbe. 1989.Ap-1/jun binding sites mediate serum inducibility of the humanvimentin promoter. Nucleic Acids Res. 17:1619-1632.
34. Rosen, C. A., J. G. Sodroski, and W. A. Haseltine. 1985. Thelocation of cis-acting regulatory sequences in the human T celllymphotrophic virus type III (HIV-III/LAV) long terminal re-peat. Cell 41:813-823.
35. Sanger, F., S. Nicklen, and A. R. Coulson. 1977. DNA sequenc-ing with chain-terminating inhibitors. Proc. Natl. Acad. Sci.USA 74:5463-5467.
36. Sax, C. M., F. X. Farrell, J. A. Tobian, and Z. E. Zehner. 1988.Multiple elements are required for expression of an intermediatefilament gene. Nucleic Acids Res. 16:8057-8076.
37. Sax, C. M., F. X. Farrell, Z. E. Zehner, and J. Piatigorsky. 1990.Regulation of vimentin gene expression in the ocular lens. Dev.
VOL. 12, 1992

2240 STOVER AND ZEHNER
Biol. 139:56-64.38. Schnitzer, J., W. W. Franke, and M. Schachner. 1985. Immuno-
cytochemical demonstration of vimentin in astrocytes andependymal cells of developing an adult mouse nervous system.J. Cell Biol. 90:435-447.
39. Singh, H., J. H. Lebowitz, A. S. Baldwin, and P. A. Sharp. 1988.Molecular cloning of an enhancer binding protein: isolation byscreening of an expression library with a recognition site DNA.Cell 52:415-423.
40. Sommers, C. L., D. Walker-Jones, S. E. Heckford, and P.Worland. 1989. Vimentin rather than keratin expression in somehormone-independent breast cancer cell lines and in oncogene-transformed mammary epithelial cells. Cancer Res. 49:4258-4263.
41. Steiner, C., M. Muller, A. Baniahmad, and R. Renkawitz. 1987.
MOL. CELL. BIOL.
Lysozyme gene activity in chicken macrophages is controlledby positive and negative regulatory elements. Nucleic AcidsRes. 15:4163-4177.
42. Steinert, P. M., and D. R. Roop. 1988. Molecular and cellularbiology of intermediate filaments. Annu. Rev. Biochem. 57:593-625.
43. Strum, P. M., G. Das, and W. Herr. 1988. The ubiquitousoctamer-binding protein Oct-1 contains a POU domain with ahomeo box domain. Genes Dev. 2:1582-1599.
44. Thomas, P. S. 1980. Hybridization of denatured RNA and smallDNA fragments transferred to nitro cellulose. Proc. Natl. Acad.Sci. USA 77:5201-5205.
45. Zehner, Z. E., X. Li, B. A. Paterson, and C. M. Sax. 1987. Thechicken vimentin gene. J. Biol. Chem. 262:8112-8120.