identification and partial purification of ankyrin, the high affinity
TRANSCRIPT

THE JOURNAL OF BIOL~CICAL CHEMISTRY Vol. 254, No. 7, Issue of April 10, pp. 2533-2541, 1979 Printed in U.S. A.
Identification and Partial Purification of Ankyrin, the High Affinity Membrane Attachment Site for Human Erythrocyte Spectrin*
(Received for publication, October 18, 1978)
Vann Bennett and Peter J. Stenbuck
From the Department of Molecular Biology, Wellcome Research Laboratories, Research Triangle Park, North Carolina 27709
Antisera have been raised against a 72,000 dalton proteolytic fragment of the membrane attachment site for human erythrocyte spectrin (Bennett, V. (1978) J. Biol. Chem. 253, 2292-2299) which was purified by so- dium dodecyl sulfate-polyacrylamide electrophoresis. Monospecific IgG, prepared by affinity chromatogra- phy with immobilized fragment, was a potent inhibitor of binding of [32P]spectrin to inside out vesicles. The inhibition was primarily due to a reduction in the KD of the interaction. Band 2.1 (a 200,000 Mr protein tightly bound on the inner surface of the membrane) was the only membrane polypeptide which cross-reacted with the antibody, and this reaction was abolished in the presence of the 72,000 dalton fragment. Band 2.1 and the fragment were phosphorylated during incubation of erythrocytes with [32P]Pi, and the 32P-labeled poly- peptides in both cases cross-reacted with anti-fragment IgG. The 32P-labeled fragment was produced from ves- icles prepared from erythrocytes cleaved on their ex- ternal surface with cY-chymotrypsin, which treatment converted phosphorylated Band 3 and PAS-l to frag- ments with M, less than 72,000. Band 2.1 was the only polypeptide in these membranes which was phospho- rylated and of sufficient molecular weight to produce a 72,000 dalton fragment. Selective extraction of Band 4.1 from vesicles with sodium cholate had no effect on binding of [32P]spectrin or on the amount of 72,000 dalton fragment, while extraction of Bands 2.1 and 4.1 tith dilute acid abolished binding of spectrin and pro- duction of the fragment. These data demonstrate that the 72,000 dalton fragment originates from Band 2.1. Direct evidence that spectrin binds to 2.1 in membranes is provided by isolation with anti-fragment IgG of a specific complex of spectrin and 2.1 formed during reassociation of spectrin with inverted vesicles which had a stoichiometry of about 1 mol of 2.l/mol of spec- trin dimer. Furthermore, partially purified 2.1 was a competitive antagonist of binding of [32P]spectrin, with a Ki of 2 pg/ml. It is concluded that Band 2.1 is the protein responsible for the high affinity association of spectrin with the inner surface of the erythrocyte plasma membrane. We have named this protein “an- kyrin” from the Greek ankyra, which means anchor.
Modifications of the fluid mosaic model for membranes (1, 2) have proposed direct connections of membrane-spanning
* The results of these studies were presented in preliminary form at the International Congress of Hematology, July 22, 1978. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduer- tisement” in accordance with 18 USC. Section 1734 solelv to indicate this fact.
cell surface proteins with cytoplasmic structural proteins on the inner surface of the plasma membrane (3, 4). These associations between membrane proteins and intracellular elements may be essential events in translation of perturba- tions at the external surface of the cell into responses such as cell motility (5) and proliferation (3, 4), and redistribution of surface-bound ligands (3, 4). Direct biochemical analysis of the protein components involved in these proposed complexes and of the mechanisms which regulate their association can be achieved, in principle, by measuring reassociation of struc- tural proteins with acceptor sites exposed on inside out mem- brane vesicles. This approach has been employed with human erythrocytes to examine the membrane attachment site for spectrin (6-8) which is a major component of the red cell cytoskeleton.
Spectrin has been radiolabeled with [B2P]Pi in intact eryth- rocytes and purified in a nonaggregated state which can recombine with the membrane (6, 7). Reassociation of [32P]spectrin occurs with inside out vesicles depleted of spec- trin and actin, and exhibits a similar dependence on ionic strength, divalent metal ions, and pH as does association of native spectrin with ghost membranes (6). [“*P]Spectrin binds with a KO in the range of lo-” to 10m7 M to a single class of protein sites tightly bound on the cytoplasmic surface of the membrane and present in approximately 100,000 copies per cell (6). A 72,000 dalton proteolytic fragment of this attach- ment site, which retains ability to bind spectrin and to com- pete for membrane-binding sites, has been purified and char- acterized (8). The membrane polypeptide which is the precur- sor for the 72,000 dalton fragment was not identified, although Band 3l and the sialoglycoproteins were excluded. This paper reports identification of Band 2.1 as the origin of the 72,000 dalton fragment using antibodies raised against the fragment cut from sodium dodecyl sulfate-polyacrylamide gels and with two additional types of experiments. It is also demonstrated that spectrin binds to 2.1 in inside out vesicles, and that partially purified 2.1 is a competitive antagonist of binding of [32P]spectrin.
EXPERIMENTAL PROCEDURES
Materials
[R’PlOrthophosphoric acid, 50 mCi/ml in 0.02 N HCl, was from New England Nuclear. cy-Chymotrypsin (70 units/mg) was from Worthington Biochemicals, 5’-ATP, adenosine, PMSF,” dithiothrei- tol, benzylpenicillin, sodium salt (1625 units/mg) were from Sigma. Acrylamide, N,N’-methylenebisacrylamide, ammonium persulfate, N,N,N’,N’-tetramethylenediamine, SDS, and Coomassie brilliant blue were from Bio-Rad. Freund’s complete and incomplete adjuvant
’ Nomenclature for erythrocyte proteins according to Steck (9). ’ The abbreviations used are: SDS, sodium dodecyl sulfate; PMSF,
phenylmethylsulfonyl fluoride; PAS, periodic acid-Schiff.
2533
by guest on March 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

2534 Identification of Membrane Attachment Site for Spectrin
were purchased from Grand Island Biological Co., Sepharose (Cl)4B and Protein A-Sepharose (Cl)4B (2 mg of protein per ml of packed gel) were from Pharmacia, Triton X-100 was from Packard, and DEAE-cellulose (DE52) was from Whatman. Cholic acid, sodium salt, was purchased from Sigma and was recrystallized from acetone and adsorbed with DE52 cellulose before use. The latter step is necessary to remove uv-absorbing contaminants which interfere with DEAE- cellulose chromatography. PMSF was routinely dissolved in dimeth- ylsulfoxide at a concentration of 200 mg/ml prior to dilution.
Methods
Preparation of Antisera-The 72,000 dalton proteolytic fragment was prepared by DEAE-cellulose chromatography as described (8), and then purified in 1.2-mg aliquots by preparative SDS electropho- resis on 6% polyacrylamide slab gels (3 mm x 10 cm x 20 cm) which were fixed in 10% acetic acid, 25% isopropyl alcohol and lightly stained with Coomassie blue. The band containing the 72,000 dalton material was cut out, neutralized with multiple washes in 50 mM Tris/HCl, pH 7.8, and homogenized in this buffer (IO ml per gel) with a Brinkmann Polytron for 30 s at 6000 rpm. The suspension was then mixed with 0.5 volume of complete or incomplete Freund’s adjuvant and homog- enized just prior to use. Five New Zealand White male rabbits (3 to 4 kg) were each injected subcutaneously with 4 ml of antigen in complete Freund’s adjuvant at multiple sites along the sides, followed by similar injections at 3-week intervals with antigen in incomplete adjuvant. The titer of the sera was monitored according to ability of purified IgG to inhibit binding of [“‘Plspectrin to inside out vesicles, and became maximal after the third injection in three rabbits. The other two animals never responded and were not used.
Purification of IgG-Pooled immune or preimmune sera (50 ml) were mixed with 12.5 g of ammonium sulfate at O”C, centrifuged (20 min, 6,000 x g), and the resulting pellet washed once with 30 volumes of 1.75 M ammonium sulfate. The washed pellet was resuspended with 20 ml of 70 mM sodium acetate, pH 5, and dialyzed against this buffer in order to precipitate euglobulins and some lipoprotein contaminants (10). The solution was centrifuged (30 min, 20,000 X g) and the supernatant adsorbed batch-wise with 5 ml of DE52 cellulose equili- brated in the acetate buffer. This step removes IgM, with essentially no loss of IgG or IgA (10). The material at this stage still contains heat-resistant protease activity which cleaves Band 2.1 in the pres- ence of PMSF and EDTA, and it was necessary to purify IgG further by affinity chromatography on a 2-ml Protein A-Sepharose (Cl)4B column (2 mg/ml of Protein A, 15 mg/ml of capacity for IgG) (11). Following application of sample, the column was washed at 24°C with 50 ml of 2 M urea, 1% (v/v) Triton X-100, and 0.2 M glycine, followed by 20 ml of 10 mM sodium acetate, pH 5. The adsorbed protein was then eluted at 4°C with 1 M acetic acid and rapidly neutralized with ammonium hydroxide. The protein was precipitated with ammonium sulfate at 50% saturation, and the pellet redissolved at a concentration of 1 mg/ml and dialyzed against 0.1 M NaCl, 10 mM sodium phosphate, 1 mM EDTA, and 1 InM NaNa.
Affinity Purification of Anti- 72,000 Dalton Fragment ZgG
72,000 dalton fragment prepared by DEAE-cellulose chromatog- raphy (8) was coupled to Sepharose (Cl)4B at a substitution of 1 mg of protein per ml of gel using the benzoquinone reaction at pH 8 (12), and the reaction was stopped after 60 min at 24°C by addition of 0.5 M glycine. Approximately 500 mg of immunoglobulin prepared as described above, or an equivalent amount of antiserum, was applied at 4°C to a 3-ml affinity column. The column was washed successively at 4°C with 250 ml of 0.25 M NaCl, 10 mM sodium phosphate, pH 7.5, 100 ml of 2 M urea, 1% (v/v) Triton X-100, 0.2 M glycine pH 6.5, and 25 ml of 10 mM sodium acetate, pH 5. The adsorbed protein was eluted with 1 M acetic acid, neutralized with ammonium hydroxide, and precipitated with solid ammonium sulfate at 50% saturation. The pellet was redissolved at a concentration of 1 mg/ml in the same buffer described above for purified IgG, and stored frozen at -20°C.
Procedures-Erythrocyte ghosts from freshly drawn blood were prepared by hypotonic lysis in 7.5 mM sodium phosphate, pH 7.5, as described (6). [32P]Spectrin (1500 to 5000 cpm/pg) was isolated from erythrocytes labeled with [32P]Pi (2 to 6 Ci/mmol) and purified to homogeneity by preparative rate zonal sedimentation on linear su- crose gradients (6, 7). Spectrin-depleted inside out vesicles were prepared by incubating erythrocyte ghosts for 30 min at 37°C in a low ionic strength buffer as described (6), except that the buffer was modified to 0.25 mM NaEDTA, 50 pg/ml of PMSF, pH 8, in order to minimize proteolysis. The membranes were stored at 0°C at a con-
centration of 1 to 2 mg/ml of membrane protein in 7.5 mM sodium phosphate, 0.5 mM NaEDTA, 0.5 mM dithiothreitol, and 0.5 mM NaN3, pH 7.5. The vesicles retain 1 to 4% of the original spectrin which may be removed by sedimentation through a 10% (w/v) dextran barrier gradient (6). The sedimentation step has been omitted in this report with no detectable change in the properties of [:‘2P]spectrin binding.
Binding of [.‘“P]spectrin to inside out vesicles was measured essen- tially as described (6). Briefly, [32P]spectrin, (20 to 100 pg/ml) was incubated for 60 to 90 min at 0°C in polystyrene tubes in a 200. to 225-d volume containing 50 to 300 pg/ml of membrane protein, 100 mM KC1 or NaCl, 7.5 mM sodium phosphate, 0.4 IIIM dithiothreitol, and 4% (w/v) sucrose, pH 7.5. Free and membrane-bound [““Plspec- trin were separated by layering the samples over 0.2 ml of 20% (w/v) sucrose in 0.4-ml polyethylene microfuge tubes followed by centrifu- gation (20 min, 20,000 x g). The tubes were frozen, and the tips containing membrane pellets, and occasionally the tops with free [“‘Plspectrin, were placed in glass vials with 5 ml of Bray’s solution and the radioactivity determined. Control samples with heat-dena- tured (5 min at 60°C) spectrin were tested at each concentration of [“‘Plspectrin and with each preparation of vesicles to provide an estimate of the amount of nonspecific binding (6). Such binding was less than 15% of the values for untreated spectrin and was routinely subtracted. The values were determined in duplicate with a range of f3 to 6%.
Membrane protein was estimated by the method of Lowry et al. (13), with bovine serum albumin as a standard, while spectrin concen- trations were determined by absorbance at 280 nm, assuming an A& ,,,,, of 10.1 (14). SDS-polyacrylamide electrophoresis was per- formed essentially by the method of Fairbanks et al. (15). Samples containing detergents or salt were prepared for analysis by precipi- tation of protein with an equal volume of ice-cold 20% (w/v) trichlo- roacetic acid dissolved in methanol, followed by centrifugation (20 min, 6000 x g). The pellets were washed once with cold methanol, neutralized with 20 ~1 of 17 N ammonium hydroxide, and lyophilized.
RESULTS
Effect of Anti-72,000 Dalton Fragment IgG on [“‘P]Spec- trin Binding to Vesicles-Following injection of the 72,000 dalton fragment purified by preparative SDS electrophoresis (see “Methods”), some rabbits developed IgG capable of in-
hibiting reassociation of [“2P]spectrin with inside out vesicles. The IgG purified further by affinity chromatography with immobilized 72,000 dalton fragment (see “Methods”) ex- hibited a 50- to loo-fold increase in specific activity and was a potent inhibitor of [““Plspectrin binding (Fig. 1). Approxi- mately 20 pg/ml of immune immunoglobulin was sufficient to abolish totally reassociation of spectrin, while preimmune IgG had a negligible influence (Fig. 1). The inhibition by the immune IgG was primarily due to a decrease in the affinity of the interaction of spectrin with the membrane; 10 pg/ml of IgG reduced the Keq by about 65%, while only a 15% fall was observed in the number of available binding sites (Fig. 1).
The inhibition of binding of spectrin was due to interaction of the antibody with the membrane, since vesicles preincu- bated with IgG and subsequently washed still exhibited a reduced association with spectrin. Furthermore, the antibody exhibited no cross-reactivity with spectrin (Fig. 2). The inhi- bition of binding by IgG cannot be explained by proteolysis of vesicles during the preincubation, since analysis of these mem- branes revealed no visible alteration in their SDS-electropho- retie profile. Proteolysis is a serious problem, however, in crude IgG preparations, and it was necessary to purify the antibody by affinity chromatography with immobilized Pro- tein A or fragment.
Identification of the Membrane Polypeptide Precursor of the 72,000 Dalton Fragment-The fact that affinity-purified antibody directed against SDS-purified 72,000 dalton frag- ment is capable of blocking reassociation of spectrin with the membrane, and that the fragment is itself a potent competitive antagonist and binds to spectrin in solution (8), makes it
by guest on March 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Identification of Membrane Attachment Site for Spectrin 2535
I I I 5 IO 15
W. pg/ml
FIG. 1. Effect of preimmune and affinity-purified anti-72,000 dal- ton fragment IgG on binding of [32P]spectrin to spectrin-depleted inside out vesicles. Left, binding of [“‘Plspectrin in the presence of increasing concentrations of IgG. Right, Scatchard plot of [3’P]spec- trin binding in the presence of 10 pg/ml of preimmune or immune IgG. Inside out vesicles (12 pg of membrane protein) were preincu- bated for 3 h at 0°C in a 100~~1 volume either in the presence of various concentrations of preimmune (o--O) or immune (M) IgG (left) or in the presence of 20 rug/ml of preimmune (M) or immune (M) IgG (right) in a buffer containing 0.1 M NaCl, 10 mM sodium phosphate, 1 InM EDTA, and 20 pg/ml of PMSF, pH 7.5. [32P]Spectrin (100 ~1,250O cpm/pg) was then added to
highly probable that this polypeptide is the membrane attach- ment site for spectrin. Elucidation of the parent polypeptide for this fragment should, therefore, provide identification of the anchorage protein. This has been achieved by three dif- ferent methods, all of which lead to the same conclusion.
Determination of Cross-reactiue Polypeptides in Erythro- cyte Membranes-The target protein for the affinity-purified anti-72,000 dalton fragment was identified by incubating in- side out vesicles with antibody followed by solubilization of the membranes with Triton X-100 and adsorption of the antigen. antibody complexes with immobilized Protein A (Fig. 2). The Protein Aa IgG . antigen complex is stable in the pres- ence of 2 M urea and 1% Triton X-100, which permits effective washing of the columns to remove proteins not bound directly to the antibody. Analysis of the membrane proteins bound under these conditions revealed only a 200,000 dalton poly- peptide (Band 2.1) and a polypeptide of about 190,000 daltons in significant amounts (Fig. 2). The 190,000 dalton protein was present in variable amounts in different vesicle preparations and probably is a proteolytic fragment of 2.1. Band 2.1 was not adsorbed to the column when vesicles were reacted with preimmune IgG, and the binding of 2.1 and the high molecular weight fragment by immune IgG was inhibited by the presence of the 72,000 dalton fragment (Fig. 2). Spectrin was not adsorbed following incubation with antibody under identical conditions (Fig. 2). No cross-reaction of antibody with sialo- glycoproteins was detected using vesicles prepared from eryth- rocytes labeled in their sialic acid residues by periodate oxi- dation followed by reduction with [3H]borohydride (17) (not shown).
“‘P-labeling of the 72,000 Dalton Fragment-The Band 2.1 polypeptide is known to be phosphorylated in membranes in a salt and cyclic AMP-dependent manner (18), and also to be phosphorylated in intact erythrocytes incubated with [3’P]Pi (7). It was, therefore, of interest to determine if the
[“P] SPECTRIN BOUND,pg/mg MEMBRANE PROTEIN
give a final concentration of 24 yg/ml (left) or various concentrations from 12 to 96 ag/ml (right), and the incubation continued for 60 min at 0°C. Membrane-bound spectrin was then determined (see “Meth- ods”). The data in the right panel are plotted according to the Scatchard equation, B/F = nK - BK (16), where B is micrograms of spectrin bound per mg of membrane protein, F is the concentration of unbound spectrin (micrograms per ml), n is the total number of binding sites (micrograms per mg), and K is the equilibrium constant (milliliters per ag). N is decreased in the presence of immune IgG from 113 to 95 pg of spectrin per mg of membrane protein, while K is decreased from 0.08 to 0.03 ml/ag, respectively.
72,000 dalton fragment also contains phosphate. SDS-poly- acrylamide electrophoresis of spectrin-depleted inside out ves- icles prepared from cells labeled with [“‘PIPi revealed exten- sive incorporation of 32P in the region of Bands 2.1 and 3, while Bands 4.1 and 4.2 contained virtually no phosphate (Fig. 3). A phosphorylated protein, which has been identified pre- viously as PAS-l (19), was also observed migrating slightly ahead of Band 3. Labeled 2.1 and the high molecular weight polypeptide fragments were the only proteins adsorbed by immune IgG, while essentially no radioactive polypeptides were obtained with preimmune IgG (Fig. 4).
Limited digestion of these labeled vesicles with a-chymo- trypsin completely removed Band 2.1, partially reduced Band 3, and had little effect on PAS-l (Fig. 3). Water-soluble 32P- labeled polypeptides were released with molecular weights of about 100,000, 72,000, and 45,000. The labeled 72,000 dalton fragment cross-reacted with anti-fragment IgG, while the 45,000 dalton proteins did not (Fig. 4). In addition, the 100,000 dalton phosphorylated bands also cross-reacted, although these are only faintly visible in the final photographs (Fig. 4). These bands disappeared with more prolonged digestion, and most likely were incompletely digested precursors of the 72,000 dalton fragment.
External cleavage of erythrocytes with o-chymotrypsin con- verts Band 3 to a 55,000 dalton membrane-bound fragment (20-22), and degrades PAS-l (20). The phosphorylated 72,000 dalton fragment was released in equivalent amounts from such externally cleaved and control vesicles even though in cleaved vesicles the 32P label in Band 3 was almost completely relocated to a 55,000 dalton position and the radioactivity had disappeared from the PAS-l region (Fig. 3). Examination of these gels leads to the conclusion that the 72,000 dalton fragment cannot be derived from Bands 4.2 and 4.1, which are not labeled, or from Band 3 and PAS-l. Band 2.1 is the only polypeptide which is sufficiently labeled and of appropriate
by guest on March 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

2536 Identification of Membrane Attachment Site for Spectrin
ABC I. i P i P i P
3 4.1.
4.2a
6-
-( HI2
-H
-L
spectrin vesicles vesicles
fraljfnent FIG. 2. Immunoadsorption with anti-72,000 dalton fragment IgG
or with preimmune IgG of spectrin, and of polypeptide components of spectrin-depleted vesicles incubated alone and in the presence of the 72,000 dalton fragment. Purified spectrin (53 pg), spectrin-de- pleted vesicles (1 mg), or vesicles (1 mg) and 72,000 dalton fragment (80 ag) were incubated for 3 h at 0°C in a l-ml volume containing 40 ag of either immune (i) or preimmune (p) IgG and 0.1 M NaCl, 10 mM
sodium phosphate, 1 mM EDTA, and 50 ag/ml of PMSF, pH 7.5. Triton X-100 (1.5% (v/v), final concentration) was added to each tube and the samples incubated for 15 min at 24°C and then applied to 0.5-ml columns of Protein A-Sepharose (C1)4B. The columns were each washed successively with 30 ml of 0.15 M NaCl, 10 mM sodium phosphate, 1% Triton X-100, pH 7.5,50 ml of 2 M urea, 0.15 M glycine, 1% Triton X-100, pH 6.8,15 ml of 5 mM sodium phosphate, and 0.02% Triton X-100, pH 7.5. IgG and adsorbed proteins were then eluted by addition of 1.8 ml of 1 M acetic acid and 0.02% Triton X-100, and the effluents were neutralized with ammonium hydroxide, lyophilized, and reconstituted with 0.08 ml of HzO. Twenty-microliter samples were then analyzed by SDS electrophoresis on a 7% polyacrylamide slab gel. The samples used as antigens were run in Lanes A (spectrin), B (spectrin-depleted vesicles), and C (72,000 dalton fragment). L refers to IgG light chain, H to heavy chain, and (Hh to heavy chain dimer.
molecular weight to generate the 72,000 dalton fragment and the other cross-reacting phosphorylated bands. (Fig. 4).
A 100,000 dalton membrane-bound polypeptide appeared in Coomassie blue-stained gels of externally chymotrypsin- cleaved vesicles only after the limited digestion which pro- duced the 72,000 dalton fragment and cleaves Band 2.1 (Fig. 3). The most likely precursor for this new band is 2.1, which is the only polypeptide of appropriate molecular weight pres- ent in sufficient amounts. Densitometry of these gels indicated that the Coomassie blue staining of the 100,000 dalton band was about one-half of that of the sum of 2.1 and the 190,000 dalton probable proteolytic derivative of 2.1. This is the amount of staining predicted if 2.1 is cleaved to a 100,000 dalton portion which is retained on the membrane. The 100,000 dalton fragment was not phosphorylated (Fig. 3), and, therefore, cannot be a precursor of the 72,000 dalton peptide. The membrane-bound fragment lacks ability to bind spectrin since almost all of the high affinity binding sites are absent following limited digestion (8).
Selective Extraction of Bands 4.1 and 2.1-The 72,000
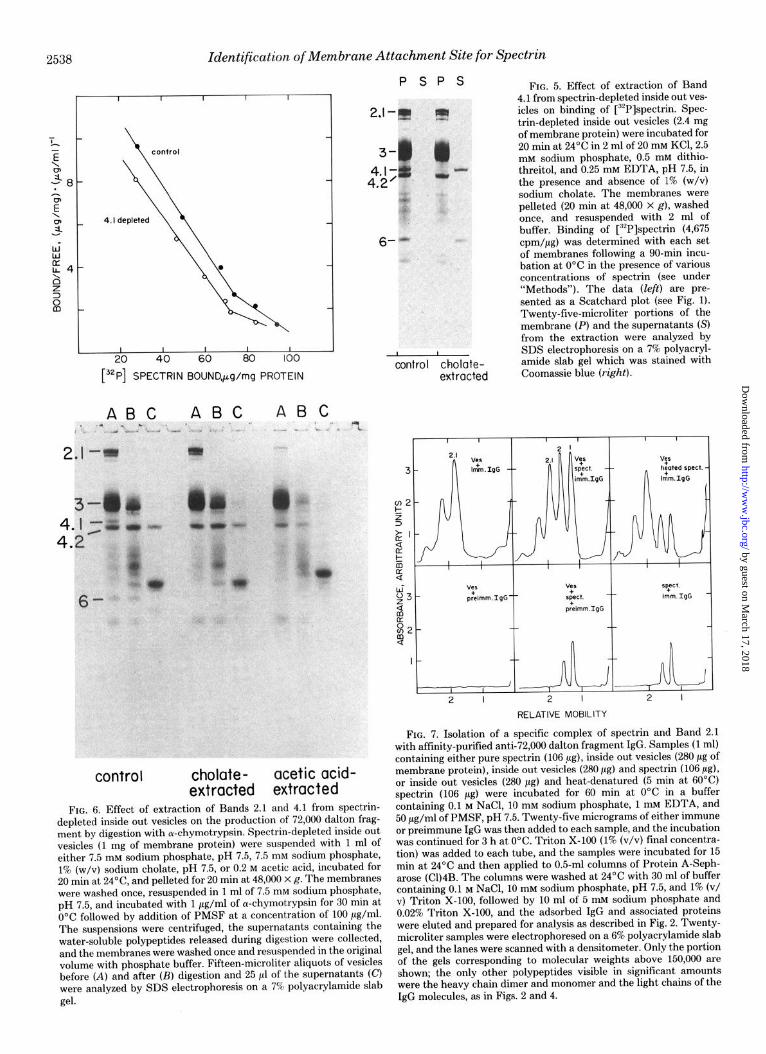
dalton fragment constitutes approximately 2% of the total erythrocyte membrane protein (8), and, therefore, should be derived from a visible band on Coomassie blue-stained SDS gels. The only polypeptides of sufficient molecular weight and stain intensity to be the precursor of the 72,000 dalton frag- ment are Bands 4.2, 4.1, 3 and possibly other polypeptides in this region, and 2.1. Band 3 can be excluded since vesicles from externally cleaved cells which lack 100,000 dalton ma- terial still generate the fragment (8) (Fig. 3). Band 4.2 is also unlikely since this polypeptide is undigested under conditions where maximal amounts of fragment are produced (8). Con- ditions have been developed for release of Band 4.1 by extrac- tion of inside out vesicles with sodium cholate (Fig. 5). These 4.1-depleted vesicles exhibited essentially no difference in the number of spectrin-binding sites, or in the KD of the spectrin- membrane interaction (Fig. 5). Furthermore, release of 72,000 dalton fragment was undiminished from 4.1-depleted mem- branes (Fig. 6).
Extraction of vesicles with dilute acetic acid abolishes bind- ing of spectrin (6), and removed Bands 4.1 and 2.1 (Fig. 6). The 72,000 dalton fragment was not produced during digestion of these vesicles, although the 45,000 dalton fragment of Band 3 was released in unaltered amounts (22) (Fig. 6). It is of interest that the 100,000 dalton polypeptide which appeared following digestion of externally cleaved vesicles (Fig. 3) also was abolished following acid extraction (not shown).
Isolation of a Complex of Spectrin and 2.1-Anti-72,000 dalton fragment IgG reduced the affinity of spectrin binding to membranes, but had only a small effect on the number of sites (Fig. 1). This antibody may, therefore, in principle, be used to evaluate directly a possible interaction between 2.1 and spectrin in membranes. Reassociation of spectrin at a saturating concentration with inside out vesicles followed by addition of antibody, solubilization of the membranes, and chromatography on Protein A-Sepharose did co-elute spectrin and 2.1 (Fig. 7). The interaction of spectrin with anti-fragment IgG was reduced by 70% with heat-denatured spectrin, and by 90% when spectrin and immune antibody were reacted in the absence of vesicles, or when spectrin and vesicles were reacted with preimmune antibody (Fig. 7). Integration of the area under Bands 1 and 2 of spectrin and 2.1 with its degradation product, gives an approximate ratio of 1 mol of spectrin dimer per mol of 2.1 (Fig. 7). This value for the stoichiometry of the reconstituted complex depends on the assumptions that spec- trin and 2.1 stain equally with Coomassie blue, that spectrin does not dissociate during chromatography, and that the antibody does not interfere significantly with the binding. Reassociated spectrin dissociates quite slowly with the ionic strength and temperature employed in these experiments. Furthermore, the addition of IgG has little influence on bind- ing of spectrin in the presence of saturating concentrations of spectrin once the membrane.spectrin complex has formed (not shown).
Partial Purification of 2.1-It was important to determine if isolated 2.1 could inhibit reassociation of [32P]spectrin. Initial efforts to extract this protein in an active form with chaotropic agents were unsuccessful. Instead, spectrin-de- pleted inside out vesicles have been solubilized with Triton X-100, with addition of PMSF and EDTA to prevent prote- olysis. The Triton X-100 was exchanged for sodium cholate during DEAE-cellulose chromatography which also achieved a substantial purification of 2.1 (Fig. 8). It was possible to assay fractions for ability to compete for binding of spectrin to membranes in the presence of cholate, since low levels of this detergent have no effect on binding, but are sufficient to maintain the solubilized proteins in solution. Some detergent
by guest on March 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Identification of Membrane Attachment Site for Spectrin 2537
A B C D E F , -
4. I- 4.2’
FIG. 3. Release of “P-labeled 72,008 dalton fragment by limited a-chymotryptic digestion of spectrin-depleted inside out vesicles which were prepared from 32P-labeled erythrocytes that were un- treated, or had been digested externally with cu-chymotrypsin. Three milliliters of packed, washed erythrocytes were incubated for 150 min at 37°C in the presence or absence of 100 ag/ml of cY-chymotrypsin in 20 ml of Krebs-Ringer bicarbonate buffer supplemented with 200 units/ml of penicillin G and 10 mM glucose. The cells were washed three times in 150 mM NaCl, and then labeled with [“‘PIP, during a 15-h incubation at 37°C in the presence of 5 ml of a buffer containing 135 mM NaCl, 20 mM NaHC03,4 mM KCl, 2.5 mM MgC12,l mM CaC12, 10 mM glucose, 200 units/ml of penicillin G, 50 pg/ml of PMSF, and 0.5 mru sodium [32P]phosphate (4 Ci/mmol), pH 7.5. The 32P-labeled erythrocytes were washed, and spectrin-depleted inside out vesicles
is >ip i.2 >i p
3- PAS- I’
R Lipid-
FIG. 4. Immunoadsorption with anti-72,000 dalton fragment IgG or with preimmune IgG of polypeptide components of “‘P-labeled spectrin-depleted vesicles and of the water-soluble polypeptides re- leased from these vesicles by digestion with a-chymotrypsin. Spectrin- depleted inside out vesicles were prepared from 32P-labeled erythro- cytes, and a portion of these were digested with a-chymotrypsin and the supernatants collected (see Fig. 3). Undigested vesicles (VES)
ABC DEF
PAS-:-
P. Lipid-
were prepared (see “Methods”) and resuspended in 7.5 mM sodium phosphate, pH 7.5, at a concentration of 2 mg/ml of membrane protein. Membranes from untreated (A) and digested (D) cells were incubated with 1 ag/ml of a-chymotrypsin for 30 min at 0°C followed by addition of PMSF (100 pg/ml) and were then pelleted for 30 min at 48,000 x g. The supernatants containing solubilized polypeptides were collected, and the digested vesicles (B, E) washed once and then resuspended in the original volume. Fifteen-microliter aliquota of vesicles (A, B, D, E) and 25-d portions of the supernatants (C, F) were analyzed by electrophoresis on a 7.5% polyacrylamide slab gel. The gel was stained with Coomassie blue (left), dried, and the 32P- labeled polypeptides visualized by autoradiography (right). The ar- row indicates the position of a new band appearing in the externally cleaved inside out vesicles following digestion on their inner surface.
c3 Ei p
(3 iii p
and the polypeptides released during digestion (DIG) were incubated for 2 h ,at 0°C with either immune (i) or preimmune (p) IgG and treated as described in Fig. 2. The material eluted from the Protein A-Sepharose (C1)4B columns was analyzed by SDS electrophoresis on 7.5% polyacrylamide gels, which were stained with Coomassie blue (left), dried, and the 32P-labeled polypeptides visualized by autora- diography (right).
by guest on March 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Identification of Membrane Attachment Site for Spectrin
I I I I I
PSPS
2.1-j@ tz”
FIG. 5. Effect of extraction of Band 4.1 from spectrin-depleted inside out ves- icles on binding of [32P]spectrin. Spec- trin-depleted inside out vesicles (2.4 mg of membrane protein) were incubated for 20 min at 24’C in 2 ml of 20 mM KCl, 2.5 mM sodium phosphate, 0.5 mM dithio- threitol, and 0.25 mru EDTA, pH 7.5, in the presence and absence of 1% (w/v) sodium cholate. The membranes were pelleted (20 min at 48,000 X g), washed once, and resuspended with 2 ml of buffer. Binding of [32P]spectrin (4,675 cpm/pg) was determined with each set of membranes following a 90-min incu- bation at 0°C in the presence of various concentrations of spectrin (see under “Methods”). The data (left) are pre- sented as a Scatchard plot (see Fig. 1). Twenty-five-microliter portions of the membrane (P) and the supernatanta (S) from the extraction were analyzed by SDS electrophoresis on a 7% polyacryl- amide slab gel which was stained with Coomassie blue (right).
I I 1 I I
20 40 60 80 100
[“‘P] SPECTRIN EOUND+g/mg PROTEIN
6 - *.R.
control cholote- extracted
ABC ABC ABC
control cholate- acetic acid- extracted extracted
FIG. 6. Effect of extraction of Bands 2.1 and 4.1 from spectrin- depleted inside out vesicles on the production of 72,000 dalton frag- ment by digestion with cu-chymotrypsin. Spectrin-depleted inside out vesicles (1 mg of membrane protein) were suspended with 1 ml of either 7.5 mM sodium phosphate, pH 7.5, 7.5 mM sodium phosphate, 1% (w/v) sodium cholate, pH 7.5, or 0.2 M acetic acid, incubated for 20 min at 24”C, and pelleted for 20 min at 48,000 X g. The membranes were washed once, resuspended in 1 ml of 7.5 mM sodium phosphate, pH 7.5, and incubated with 1 pg/ml of cY-chymotrypsin for 30 min at 0°C followed by addition of PMSF at a concentration of 100 pg/ml. The suspensions were centrifuged, the supernatants containing the water-soluble polypeptides released during digestion were collected, and the membranes were washed once and resuspended in the original volume with phosphate buffer. Fifteen-microliter aliquots of vesicles before (A) and after (B) digestion and 25 al of the supernatants (C) were analyzed by SDS electrophoresis on a 7% polyacrylamide slab gel.
,-
>-
I -
I- L
I I I I
“es ves &lmm.IgG-- &t. -
spp. Imm.IgG -
&imm.IgG
RELATIVE MOBILITY
FIG. 7. Isolation of a specific complex of spectrin and Band 2.1 with affinity-purified anti-72,000 dalton fragment IgG. Samples (1 ml) containing either pure spectrin (106 ug), inside out vesicles (280 pg of membrane protein), inside out vesicles (280 pg) and spectrin (106 ,ug), or inside out vesicles (280 pg) and heat-denatured (5 min at 60°C) spectrin (106 pg) were incubated for 60 min at 0°C in a buffer containing 0.1 M NaCl, 10 mM sodium phosphate, 1 mu EDTA, and 50 pgg/ml of PMSF, pH 7.5. Twenty-five micrograms of either immune or preimmune IgG was then added to each sample, and the incubation was continued for 3 h at 0°C. Triton X-100 (1% (v/v) final concentra- tion) was added to each tube, and the samples were incubated for 15 min at 24’C and then applied to 0.5~ml columns of Protein A-Seph- arose (C1)4B. The columns were washed at 24°C with 30 ml of buffer containing 0.1 M NaCl, 10 mM sodium phosphate, pH 7.5, and 1% (v/ v) Triton X-100, followed by 10 ml of 5 mu sodium phosphate and 0.02% Triton X-100, and the adsorbed IgG and associated proteins were eluted and prepared for analysis as described in Fig. 2. Twenty- microliter samples were electrophoresed on a 6% polyacrylamide slab gel, and the lanes were scanned with a densitometer. Only the portion of the gels corresponding to molecular weights above 150,000 are shown; the only other polypeptides visible in significant amounts were the heavy chain dimer and monomer and the light chains of the IgG molecules, as in Figs. 2 and 4.
by guest on March 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Identification of Membrane Attachment Site for Spectrin 2539
is necessary, however, since activity in binding assays disap- peared following removal of cholate by dialysis. A large peak of protein containing 4.1 and some 2.1, which had a small amount of inhibition activity (Fig. 8), was eluted with 0.1 M salt. Bands 2.1, some 4.1, and 4.2 appeared along with inhibi- tory activity at about 0.2 to 0.35 M salt. Band 3 and the sialoglycoproteins were not eluted, even at 0.5 M salt, and probably were precipitated on the column in the absence of Triton X-100. Rechromatography of Peak 3 in Fig. 8 on DEAE-cellulose yielded over 90% recovery of protein, and removed all major bands except 2.1 and 4.2. There were two poorly separated peaks of inhibitory activity which had iden- tical composition in SDS gels; the basis for their difference in specific activity is not known.
The partially purified 2.1 was a potent competitive antag- onist of spectrin binding with a Ki of 2 pg/ml (Fig. 9). The activity was quite labile, and usually disappeared within 2 days of eluting protein from the column. This behavior may be due to slow aggregation of protein, since solutions became faintly turbid within several days. The unstable nature of the inhibitory activity has prevented further purification of 2.1 in a functional form.
It thus seems likely that the Band 2.1 cannot exist in a native form in aqueous solution and is truly a membrane protein. The 100,000 dalton domain of Band 2.1 (see above) is probably the membrane-bound portion of this protein, since the 72,000 dalton fragment is freely soluble in water.
DISCUSSION
Several lines of evidence presented in this report indicate that Band 2.1 is the membrane protein responsible for the high affinity binding of spectrin to the cytoplasmic surface of erythrocyte ghosts. The key to identification of 2.1 was the previous purification of a 72,000 dalton fragment, released during digestion of inside out vesicles (8). This polypeptide is a competitive antagonist (Kz = 10M7 M) of reassociation of spectrin with membranes, forms a specific complex with spec- trin in solution, and comprises about 2% of the erythrocyte membrane protein (8). These properties led to the conclusion that this fragment is the spectrin-binding portion of the an- chorage protein. This function of the 72,000 dalton fragment is further supported by the fact that affinity-purified IgG raised against fragment cut from SDS-polyacrylamide gels is a potent inhibitor of [32P]spectrin binding (Fig. 1). The mon- ospecific antibodies cross-react only with Band 2.1 among the membrane proteins, and the binding to Band 2.1 is blocked by the 72,000 dalton fragment (Fig. 2).
Band 2.1 and the fragment are phosphorylated following incubation of erythrocytes with [32P]Pi, and the 32P-labeled polypeptides in both cases cross-react with anti-fragment IgG (Figs. 3 and 4). 32P-labeled 72,000 dalton fragment and -100,000 dalton cross-reacting polypeptides are produced from vesicles prepared from erythrocytes cleaved on their external surface with a-chymotrypsin when 32P-labeled Band
8 16 24 32 40 48 FRACTION NUMBER
ABCDl234 I I I I , - - -____.
25
8 16 24 32 FRACTION NUMBER
FIG. 8. Partial purification of Band 2.1 and replacement of Triton X-100 by sodium cholate by DE52 cellulose chromatography of Triton X-NO-solubilized spectrin-depleted inside out vesicles. Packed eryth- rocyte ghosts (100 ml) were resuspended in 1.2 liters of 0.1 mM EDTA and 100 pg/ml of PMSF, pH 8, incubated for 30 min at 37°C to dissociate spectrin, and pelleted (90 min, 20,OCQ X g) in a GSA rotor. The resulting vesicles were washed once with 1 liter of 0.1 mM EDTA, pH 8, and 7.5 mM sodium phosphate, pH 7.5, resuspended at a concentration of 2 mg/ml of membrane protein in 115 ml of buffer containing 1.4% (v/v) Triton X-100, 7.5 mu sodium phosphate, 0.2 mM EDTA, 0.5 mM dithiothreitol, and 50 pg/ml of PMSF, pH 7.5, and incubated for 30 min at 0°C. The suspension was pelleted (90 min, 100,000 x g in an SW 27 rotor), and the supernatant (1.8 mg/ml of protein) applied at 4°C to a column (1 x 10 cm) of DE52 cellulose equilibrated with 1% (w/v) sodium cholate, 7.5 mM sodium phosphate, ---. .-- . . . --~ -. 0.2 mM E;U’I’A, and 0.5 mM dithiothreitol, pH 7.5. The column was
washed with 700 ml of equilibration buffer, and then eluted (4-ml fractions, 50 ml/h) with 0.1 M KC1 dissolved in this buffer, followed by a 200~ml linear gradient of 0.1 to 0.6 M KCl. The portions were monitored for A280 (M), and for inhibition of binding of [3*P]spectrin (1,000 cpm/M/ml) to vesicles (180 &ml) (O-----O) (left). Samples 20 to 30 were pooled, dialyzed for 12 h against equili- bration buffer, and reapplied to a 5-ml DE52 column which was eluted with 0.1 M KC1 followed by a 150~ml linear gradient of 0.1 M to 0.6 M
KC1 (middle). Samples from starting vesicles (A), the pellet (B) and the supernatant (CJ from the Triton X-100 extraction, the unabsorbed material from the first DE52 column (D), and pooled fractions 1 to 4 were analyzed by electrophoresis on a 6% polyacrylamide slab gel which was stained with Coomassie blue (right). A companion gel was also stained for sialoglycoproteins with the periodic acid-Schiff reac- tion, and no staining was observed in the eluted fractions (not shown).
by guest on March 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

2540 Identification of Membrane Attachment Site for Spectrin
I I I I I I I I I
2 0 2 4 PROTEIN,pg/ml
FIG. 9. Dixon plot (23) of inhibition of binding of [““Plspectrin to inverted vesicles in the presence of increasing concentrations of partially purified Band 2.1 with 14 (O--O), 32 (H), and 71 (A-A) pg/ml of [32P]spectrin. Binding of [32P]spectrin (1800 cpm/ ag) to inside out vesicles (85 pg/ml) was determined (see “Methods”) in the presence of various concentrations of partially purified Band 2.1 (Fig. 8B, Fractions 13 to 15).
3 is converted to a 55,000 dalton fragment and [32P]PAS-1 is no longer evident (Fig. 3). Band 2.1 is the only polypeptide in these membranes which is phosphorylated and of sufficient molecular weight to produce these fragments. A further dem- onstration that the fragment is derived from 2.1 is that selec- tive extraction of Band 2.1, but not Band 4.1, abolished production of the 72,000 dalton fragment and destroys binding of [32P]spectrin (6) (Fig. 6). The same conclusion that Band 2.1 is the origin of the fragment has been reached by compar- ison of peptide maps of the fragment with those of Bands 1, 2; 2.1, 3, 4.1, and 4.2 (36).
Direct evidence that spectrin binds to 2.1 in membranes is provided by isolation of a specific complex of spectrin, 2.1, and anti-fragment antibody following reassociation of spectrin with inverted vesicles and addition of IgG (Fig. 7). Further- more, partially purified 2.1 containing Band 4.2 as the major contaminant (Fig. 8) is a competitive antagonist of binding of [32P]spectrin to vesicles, with a Ki of about 2 ,sg/ml (Fig. 9).
Band 2.1 and the 190,000 dalton fragment constitute about 6% of the material on Coomassie blue-stained gels of spectrin- depleted vesicles (data not shown), while spectrin comprises about 25% of the staining in ghosts (9,14). Spectrin, therefore, is present in a ratio of about 2 mol of dimer (Band 1 and 2) per mol of 2.1. It is conceivable that each 2.1 polypeptide contains two high affinity binding sites for spectrin. This possibility is not likely, however, since the 100,000 dalton membrane-bound domain of 2.1 (see “Results”) has no binding capacity, and the stoichiometry of spectrin binding to the 72,000 dalton fragment has been determined previously to be about 1 mol of spectrin dimer per mol of fragment (8). Fur- thermore, the complex of spectrin and 2.1 isolated with anti- fragment IgG contains only 1 mol of spectrin dimer per mol of 2.1 (Fig. 7). These data suggest that in erythrocyte ghosts, spectrin present in excess over the number of membrane attachment sites is bound through association with other spectrin molecules to form tetramers or by lower affinity (& > 10m6 M) interaction with another membrane protein(s) (or both). Strong evidence that spectrin exists as a tetramer in
uiuo is provided by the fact that stable, tetrameric spectrin has been isolated from bovine and human ghosts by elution at low temperature, and is the predominant form of spectrin under these conditions (24, 25). Estimates of the number of binding sites for dimeric (8 S) [32P]spectrin in vesicles have routinely been in the range of 100 to 140 fig of spectrin per mg of membrane protein (Figs. 1 and 5), which is lower than the amount of spectrin in ghosts. These values are subject to problems in accurate determination of membrane protein and of the absorbance coefficient for spectrin, but are consistent with binding of spectrin as a dimer to the amount of 2.1 present in the membrane. It is, therefore, reasonable to assume that in erythrocytes spectrin tetramer (i.e. (Bands 1 + 2)~) is bound to a single high affinity membrane attachment site containing Band 2.1.
It is important to emphasize that association of spectrin with the membrane with a & greater than 10m6 M would not be detectable in these studies due to the low concentrations of spectrin employed in the binding assays. However, the actual local concentration of spectrin in erythrocytes may exceed 10 mg/ml or 10m5 M in spectrin tetramer.3 Thus, spec- trin may interact in a meaningful way with proteins other than 2.1, or with membrane lipids, although the KD would be in the range of 10m6 to 10m5 M.
Functionally important associations of cytoplasmic struc- tural proteins with the plasma membrane have been proposed (3-5), and direct evidence has been presented for binding of actin (26, 27) and intermediate filament protein (28) with membrane proteins. 2.1 is the fist example, as far as we are aware, of a membrane protein identified in detail as the anchorage site for a cytoskeletal protein. We propose that in recognition of this function, 2.1 be named ankyrin (derived from the Greek ankyra, n. anchor). Ankyrin itself is not a transmembrane protein (9), but probably does interact with integral proteins. Evidence for these additional interactions of ankyrin is provided by reports that perturbations of spectrin altered the topography of intramembrane particles (29-30) and sialoglycoproteins (31), and by the localization of spectrin in patterns coincident with those of the intramembrane par- ticles (32). Furthermore, incubation of ghosts with the 72,000 dalton fragment has been shown to increase the rate of lateral mobility of fluorescent-labeled integral membrane proteins (33), presumably due to competition by the fragment for spectrin-binding sites, thereby causing release of ankyrin with its associated proteins. Ankyrin thus may be involved in direct or indirect association with multiple membrane proteins, and may provide the common intermediate for linkage of integral membrane proteins with cytoskeletal elements. A protein of this type has been proposed to explain co-capping of receptors for various, unrelated ligands (34), and may be involved in the co-aggregation of receptors for insulin, epidermal growth fac- tor, and (u-2 macroglobulin (35). Ankyrin contains, in addition to the water-soluble 72,000 dalton portion, a 100,000 dalton membrane-bound fragment (see “Results”). It is this large fragment which most likely contributes the hydrophobic prop- erties of ankyrin (see “Results”), and may be the portion of the molecule responsible for interactions with integral mem- brane proteins.
Future areas of interest will be the physical properties of ankyrin, identification of the polypeptides associated with this protein, and investigation of the possible regulatory signifi- cance of the cyclic AMP-dependent phosphorylation of an- kyrin (18). It will also be important to determine if ankyrin or related proteins are present in cells other than the erythrocyte.
3 Calculated assuming spectrin is confined to within 0.1 pm of the plasma membrane, that the concentration of spectrin in packed cells is 1 mg/ml, and that the erythrocyte is a sphere with a radius of 4 pm.
by guest on March 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Identification of Membrane Attachment Site for Spectrin 2541
Achnowledgments-Interesting discussions with Drs. J. Niedel, N. Biophys. Res. Commun. 48,242-251 Sahyoun, and M. Hebden are gratefully acknowledged. 18. Fairbanks, G., and Avruch, J. (1974) Biochemistry 13, 5514-5521
19. Shapiro, D. L., and Marchesi, V. T. (1977) J. Biol. Chem. 252,
REFERENCES
1. Singer, S. J., and Nicolson, G. L. (1972) Science 175, 720-731 2. Gitler, C. (1972) Annu. Rev. Biophys. Bioeng. 1, 51-92 3. Edelman, G. M. (1976) Science 192,218-226 4. Nicolson, G. L. (1976) Biochim. Biophys. Acta 457, 57-108 5. Korn, E. D. (1976) in Cell Motility (Cold Spring Harbor Confer-
ence on Ceil Proliferation) (Goldman. R., Pollard, T.. and Rosenbaum, J., eds) Vol. 13, pp. 623-630, Cold Spring Harbor Laboratory, Cold Spring Harbor, N. Y.
6. Bennett, V., and Branton, D. (1977) J. Biol. Chem. 252, 2753- 2763
7. Bennett, V. (1977) Life Sci. 21, 433-440 8. Bennett. V. (1978) J. Biol. Chem. 253.2292-2299 9. Steck, T. L. (1974) J. Cell Biol. 62, l-‘19
10. Axelsen, N. H., Kroll, J., and Wecke, B., eds (1973) A Manual of Quantitative Immunoelectrophoresis, pp. 162-164, Universitets for laget, Oslo
11. Hjelm, H., Hjelm, K., and Sjoquist, J. (1972) FEBS Lett. 28, 73- 76
12. Brandt, J., Andersson, L.-O., and Porath, J. (1975) Biochim. Biophys. Acta 386, 196-202
13. Lowry, 0. H., Rosebrough, N. J., Farr, A. L., and Randall, R. J. (1951) J. Biol. Chem. 193,265-275
14. Clarke, M. (1971) Biochem. Biophys. Res. Commun. 45, 1063- 1070
15. Fairbanks, G., Steck, T. L., and Wallach, D. F. H. (1971) Bio- chemistry 10,2606-2617
16. Scatchard, G. (1949) Ann. N. Y. Acad. Sci. 51,660-672 17. Blumenfeld, 0. O., Gallop, P. M., and Liao, T. H. (1972) Biochem.
20.
21.
22. Steck, T. L., Ramos, B., and Strapazon, E. (1976) Biochemistry 15, 1154-1161
23. Dixon, M. (1953) Biochem. J. 55,170-171 24. Ralston, G. B. (1976) Biochim. Biophys. Acta 445, 163-172 25. Ralston. G. B.. Dunbar. J.. and White. M. (1977) Biochim. Bio-
26. 27. 28.
29. 30.
31.
32.
33.
34.
35.
36.
508-519 Steck, T. L. (1972) in Membrane Research (Fox, C. F., ed) p. 71,
Academic Press, New York Cobantchik, Z. I., and Rothstein, A. (1974) J. Membr. Biol. 15,
207
phys.‘Acta 491,354-348’ Koch, G., and Smith, M. (1978) Nature 273, 274-278 Flanagan, J., and Koch, G. (1978) Nature 273,278-281 Carter, W. R., and Hakomori, S. (1978) J. Biol. Chem. 253,2867-
2874 Elgsaeter, A., and Branton, D. (1974) J. Cell Biol. 63, 1018-1036 Elgsaeter, A., Shotton, D. M., and Branton, D. (1976) Biochim.
Biophys. Acta 426, 101-122 Nicolson, G. M., and Painter, R. G. (1973) J. Cell Biol. 59, 395-
406 Shotton, D., Thompson, K., Wofsy, L., and Branton, D. (1978) J.
Cell Biol. 76, 512 Fowler, V., and Bennett, V. (1978) J. Supramol. Struct. 8, 215-
221 Ash, J. F., Louvard, D., and Singer, S. J. (1977) Proc. Natl. Acad.
Sci. U. S. A. 74,5584-5588 Maxfield, F. R., Schlessinger, J., Schecter, Y., Pastan, I., and
Willingham, M. (1978) Cell 14,805-811 Luna, E. J., Kidd, G. H., and Branton, D. (1979) J. Biol. Chem.
254,2526-2532
by guest on March 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

V Bennett and P J Stenbuckattachment site for human erythrocyte spectrin.
Identification and partial purification of ankyrin, the high affinity membrane
1979, 254:2533-2541.J. Biol. Chem.
http://www.jbc.org/content/254/7/2533.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/254/7/2533.citation.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on March 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from