ĐẠi hỌc quỐc gia hÀ nỘi - hus.vnu.edu.vn (201).pdf · lêi c¶m ¬n lêi ®Çu tiªn, em...
TRANSCRIPT

ĐẠI HỌC QUỐC GIA HÀ NỘI
TRƯỜNG ĐẠI HỌC KHOA HỌC TỰ NHIÊN ----------
Ngô Thị Kim Toán
NGHIÊN CỨU PHÂN LẬP TUYỂN CHỌN CÁC CHỦNG VI SINH VẬT ỨNG DỤNG XỬ LÝ NƯỚC THẢI GIÀU NITƠ, PHOTPHO
LUẬN VĂN THẠC SĨ KHOA HỌC
Hà Nội – 2012

ĐẠI HỌC QUỐC GIA HÀ NỘI
TRƯỜNG ĐẠI HỌC KHOA HỌC TỰ NHIÊN -----------
Ngô Thị Kim Toán
NGHIÊN CỨU PHÂN LẬP TUYỂN CHỌN CÁC CHỦNG VI SINH VẬT ỨNG DỤNG XỬ LÝ NƯỚC THẢI GIÀU NITƠ, PHOTPHO
Chuyên ngành: Sinh học thực nghiệm Mã số: 60 42 30
LUẬN VĂN THẠC SĨ KHOA HỌC
NGƯỜI HƯỚNG DẪN KHOA HỌC: TS. NGUYỄN QUANG HUY
Hà Nội – 2012

Lêi C¶m ¬n
Lêi ®Çu tiªn, em xin göi lêi c¶m ¬n ch©n thµnh ®Õn TS. NguyÔn Quang Huy, ngêi
ThÇy ®· tËn t×nh híng dÉn, chØ b¶o em trong suèt thêi gian häc tËp vµ thùc hiÖn luËn v¨n.
Em xin göi lêi c¶m ¬n ch©n thµnh ®Õn c¸c thÇy c« gi¸o trong Khoa Sinh häc, Trêng
§¹i häc Khoa häc Tù Nhiªn, §HQGHN ®· dµnh t©m huyÕt gi¶ng d¹y, trang bÞ kiÕn thøc cho
chóng em trong suèt qu¸ tr×nh häc tËp vµ thùc hiÖn luËn v¨n.
Trong qu¸ tr×nh häc tËp vµ thùc hiÖn luËn v¨n, em ®· nhËn ®îc rÊt nhiÒu sù gióp ®ì,
hç trî. Em xin göi lêi c¶m ¬n ch©n thµnh ®Õn c¸c thÇy c«, c¸n bé, häc viªn, sinh viªn trong Bé
m«n Sinh lý thùc vËt vµ Hãa sinh, Khoa Sinh häc; Phßng Enzym häc vµ Ph©n tÝch ho¹t tÝnh
sinh häc, Phßng thÝ nghiÖm träng ®iÓm C«ng nghÖ Protein vµ Enzym.
§Ò tµi thùc hiÖn cã sù hç trî kinh phÝ cña ®Ò tµi Nghiªn cøu ph¸t triÓn c«ng nghÖ mµng
sinh häc trong xö lý níc th¶i giµu nit¬ vµ photpho cña Bé C«ng th¬ng
Cuèi cïng, em xin göi lêi c¶m ¬n ®Õn gia ®×nh, b¹n bÌ ®· lu«n ë bªn ®éng viªn, gióp ®ì
em trong suèt thêi gian häc tËp vµ thùc hiÖn luËn v¨n, gióp em trëng thµnh h¬n khi s¾p bíc
®i trªn nh÷ng con ®êng míi.
Hµ Néi, th¸ng 12 n¨m 2012
Häc viªn
Ng« ThÞ Kim To¸n

Luận văn thạc sĩ
Ngô Thị Kim Toán i K19 – Sinh học thực nghiệm
MỤC LỤC
MỞ ĐẦU ................................................................................................................ 1
CHƯƠNG 1: TỔNG QUAN TÀI LIỆU .................................................................. 3
1.1. Tình trạng ô nhiễm môi trường nước hiện nay ở Việt Nam và thế giới ........... 3
1.2. Các phương pháp xử lý ô nhiễm nước thải có chứa hợp chất nitơ, photpho
hiện nay ................................................................................................................... 5
1.2.1. Phương pháp hóa học ........................................................................... 5
1.2.1.1. Xử lý các hợp chất chứa nitơ bằng phương pháp hóa học .............. 5
1.2.1.2. Xử lý các hợp chất photpho bằng phương pháp hóa học. ............... 6
1.2.2. Phương pháp sinh học .......................................................................... 7
1.3. Các vi sinh vật có khả năng chuyển hóa các hợp chất chứa nitơ, photpho trong
xử lý ô nhiễm nước thải. .........................................................................................10
1.3.1. Vi sinh vật có khả năng chuyển hóa các hợp chất chứa nitơ ............... 10
1.3.2. Vi sinh vật có khả năng tích lũy photpho ............................................ 12
1.4. Màng sinh học và ứng dụng của màng sinh học trong việc xử lý ô nhiễm nước
thải giàu nitơ, photpho. ...........................................................................................14
1.4.1. Màng sinh học ................................................................................... 14
1.4.1.1. Định nghĩa về màng sinh học ...................................................... 14
1.4.1.2. Thành phần và quá trình hình thành màng sinh học ..................... 14
1.4.2. Vai trò và ứng dụng của sự hình thành màng sinh học ........................ 18
1.4.2.1. Vai trò của sự hình thành màng sinh học ..................................... 18
1.4.2.2. Ứng dụng của màng sinh học trong xử lý ô nhiễm ....................... 20
CHƯƠNG 2: NGUYÊN LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU .................. 23
2.1. Nguyên liệu ...................................................................................................23
2.2. Hóa chất, thiết bị ...........................................................................................23
2.2.1. Môi trường nuôi cấy .......................................................................... 23
2.2.2. Máy móc thiết bị ................................................................................ 24
2.3. Phương pháp nghiên cứu ...............................................................................25

Luận văn thạc sĩ
Ngô Thị Kim Toán ii K19 – Sinh học thực nghiệm
2.3.1. Phương pháp phân lập vi khuẩn ......................................................... 25
2.3.2. Phương pháp đánh giá khả năng hình thành biofilm ........................... 25
2.3.3. Quan sát cấu trúc biofilm bằng chụp ảnh trên kính hiển vi điện tử quét
(SEM) 26
2.3.4. Ảnh hưởng của các điều kiện môi trường nuôi cấy lên sự hình thành
màng sinh học ................................................................................................ 26
2.3.4.1. Ảnh hưởng của nhiệt độ môi trường nuôi cấy .............................. 26
2.3.4.2. Ảnh hưởng của pH môi trường nuôi cấy ...................................... 27
2.3.5. Phương pháp nhuộm Gram. ............................................................... 27
2.3.6. Phương pháp sử dụng kit APi ............................................................ 28
2.3.7. Phương pháp đánh giá khả năng chuyển hóa các hợp chất nitơ .......... 28
2.3.7.1. Phương pháp phân tích nitơ tổng số............................................. 28
2.3.7.2. Phương pháp phân tích hàm lượng amoni (NH4+) ........................ 29
2.3.7.3. Phương pháp thử khả năng chuyển hóa nitrite ............................. 30
2.3.8. Phương pháp đánh giá khả năng tích lũy photpho .............................. 31
2.3.8.1. Phương pháp phân tích photpho tổng .......................................... 32
2.3.8.2. Phương pháp phân tích hàm lượng Ortho photphate (PO43-) ........ 32
2.3.9. Phương pháp phân loại vi sinh vật dựa trên gen 16S rRNA ................ 33
2.3.10. Phương pháp thống kê sinh học ......................................................... 33
CHƯƠNG 3: KẾT QUẢ VÀ THẢO LUẬN ......................................................... 34
3. 1. Nghiên cứu các chủng vi sinh vật có khả năng chuyển hóa nitơ ....................34
3.1.1. Phân lập chủng vi sinh vật có khả năng chuyển hóa nitơ và có khả năng
hình thành màng sinh học ............................................................................... 34
3.1.1.1. Phân lập các chủng vi sinh vật có khả năng chuyển hóa nitơ ....... 34
3.1.1.2. Khả năng hình thành màng sinh học của các chủng phân lập ....... 36
3.1.2. Khả năng chuyển hóa nitơ .................................................................. 38
3.1.2.1. Khả năng chuyển hóa amoni ....................................................... 38
3.1.2.2. Khả năng chuyển hóa nitrite ........................................................ 39
3. 2. Nghiên cứu các chủng vi sinh vật có khả năng tích lũy photpho. ...................40

Luận văn thạc sĩ
Ngô Thị Kim Toán iii K19 – Sinh học thực nghiệm
3.2.1. Phân lập chủng vi sinh vật có khả năng tích lũy photpho và có khả
năng hình thành màng sinh học. ..................................................................... 40
3.2.1.1. Phân lập các chủng vi sinh vật có khả năng tích lũy photpho ....... 40
3.2.1.2. Khả năng hình thành màng sinh học của các chủng phân lập ....... 41
3.2.2. Khả năng xử lý photpho của các chủng nghiên cứu. ........................... 42
3. 3. Các đặc điểm hình thái, sinh lý, sinh hóa của các chủng nghiên cứu..............43
3.3.1. Khả năng tạo hình thành màng sinh học trên một số giá thể ............... 43
3.3.2. Đặc điểm hình thái của các chủng nghiên cứu .................................... 45
3.3.3. Ảnh hưởng của một số yếu tố môi trường lên sự hình thành màng sinh
học .......................................................................................................... 46
3.3.4. Khả năng chuyển hóa một số chất trong bộ kit APi ............................ 48
3.3.5. Trình tự 16S rRNA và cây phát sinh chủng loại ................................. 50
KẾT LUẬN ........................................................................................................... 54
KIẾN NGHỊ .......................................................................................................... 54
TÀI LIỆU THAM KHẢO ..................................................................................... 55

Luận văn thạc sĩ
Ngô Thị Kim Toán iv K19 – Sinh học thực nghiệm
DANH MỤC BẢNG
Bảng 3.1. Hàm lượng thành phần nitơ và photpho trong mẫu phân tích. .......................... 34
Bảng 3.2. Địa điểm và số lượng các chủng vi sinh vật có khả năng chuyển hóa nitơ ........ 35
Bảng 3.3. Địa điểm và số lượng các chủng vi sinh vật có khả năng tích lũy photpho ........ 41
Bảng 3.4. Một số đặc điểm sinh hoá của các chủng theo kit APi (BioMérieus)................. 48

Luận văn thạc sĩ
Ngô Thị Kim Toán v K19 – Sinh học thực nghiệm
DANH MỤC HÌNH
Hình 1.1. Chu trình nitơ trong tự nhiên ................................................................... 8
Hình 1.2. Các giai đoạn chính của quá trình hình thành một biofilm ...................... 16
Hình 3.1. Hình ảnh các khuẩn lạc phân lập trên môi trường .................................. 35
Hình 3.2. Khả năng hình thành màng sinh học của các chủng trên môi trường
Winogradsky 1 được phân lập từ các mẫu nước thải thu từ bể biogas .................... 36
Hình 3.3. Khả năng hình thành màng sinh học của các chủng trên môi trường
Winogradsky 1 được phân lập từ các mẫu nước thải khu tập trung rác thải ............ 37
Hình 3.4. Khả năng tạo hình thành màng sinh học của các chủng trên môi trường
Winogradsky 2 được phân lập ở bể biogas ............................................................. 37
Hình 3.5. Khả năng chuyển hóa amoni của các chủng nghiên cứu. ........................ 38
Hình 3.6. Khả năng chuyển hóa nitrite của các chủng nghiên cứu .......................... 39
Hình 3.7. Một số khuẩn lạc phân lập trên môi trường AMM .................................. 40
Hình 3.8. Khả năng hình thàng màng sinh học của các chủng vi sinh vật có khả
năng tích lũy photpho ............................................................................................ 41
Hình 3.9. Khả năng tích lũy photpho của các chủng nghiên cứu trong môi trường
với hàm lượng photpho 6mg/l ................................................................................ 42
Hình 3.10. Khả năng tích lũy photpho của các chủng nghiên cứu trong môi trường
hàm lượng photpho18mg/l ..................................................................................... 43
Hình 3.11. Khả năng hình thành biofilm trên giá thể nhựa ống eppendorf. ............ 44
Hình 3.12. Màng biofilm nổi của chủng B11.11, B21.10, B23.2, A4.2. ................. 44
Hình 3.13. Ảnh nhuộm Gram các chủng nghiên cứu ở độ phóng đại 1000 lần. ... 45
Hình 3.14. Cấu trúc hiển vi màng biofilm của chủng nghiên cứu .......................... 45
Hình 3.15. Ảnh hưởng của nhiệt độ lên khả năng hình thành màng sinh học ......... 46
Hình 3.16. Ảnh hưởng của pH môi trường ............................................................ 47
Hình 3.17. Sơ đồ cây phát sinh chủng loại của chủng B11.11 ................................ 50
Hình 3.18. Sơ đồ cây phát sinh chủng loại của chủng B21.10 ................................ 51
Hình 3.19. Sơ đồ cây phát sinh chủng loại của chủng B23.2 .................................. 52
Hình 3.20. Sơ đồ cây phát sinh chủng loại của chủng A4.2 .................................... 53

Luận văn thạc sĩ
Ngô Thị Kim Toán vi K19 – Sinh học thực nghiệm
BẢNG KÝ HIỆU VIẾT TẮT
ADH L-arginine
ADI Axit adipic
AMM Acetate mineral medium
ARA L-arabinose
BOD Biochemical oxygen demand (Nhu cầu oxy sinh học)
BTNMT Bộ Tài nguyên Môi trường
CAP Axit capric
CIT Trisodium citrate
COD Chemical oxygen demand (Nhu cầu oxy hóa học)
EPS Mạng lưới các hợp chất ngoại bào (Extracellular polymeric
substances)
ESC Esculin ferric citrate
GEL Gelatine
GLU D-glucose
GNT Potassium gluconate
HEPES 4-(2-hydroxyethyl)-1- piperazineethanesulfonic
LB Môi trường Luria betani
MAL D-mantose
MAN D-mannitol
MLT Axit malic
MNE D-mannose
NAG N- acetyl- glucosamine
PAC Axit phenyl acetic
PNPG 4-nitrophenyl β D –galactopyranoside
QCVN Quy chuẩn Việt Nam
SEM Ảnh vi điện tử quét (Scanning Electron Microscope)
TRP L- tryptophan
URE Urea

Luận văn thạc sĩ
Ngô Thị Kim Toán 1 K19 – Sinh học thực nghiệm
MỞ ĐẦU
Hiện nay, ô nhiễm môi trường đang là vấn đề được quan tâm của nhiều quốc
gia trên thế giới trong đó có Việt Nam. Ô nhiễm nguồn nước không chỉ ảnh hưởng
đến đời sống con người mà còn ảnh hưởng đến đa dạng sinh học. Tình trạng ô
nhiễm nước thải trong đó có nguyên nhân từ các hợp chất nitơ và photpho đang có
chiều hướng gia tăng trong những năm gần đây cùng với sự phát triển của kinh tế,
xã hội. Để làm giảm mức độ ô nhiễm từ nước thải giàu nitơ và photpho, nhiều
phương pháp đã và đang nghiên cứu bằng cách kết hợp các biện pháp vật lý, hóa
học và sinh học. Xử lý nước thải dựa vào phương pháp hóa học, vật lý thường có
hiệu quả cao, nhanh nhưng chi phí lớn, không mang tính bền vững.
Xử lý nước thải bằng phương pháp sinh học dựa trên cơ sở sử dụng các vi
sinh vật là một phương pháp được quan tâm nghiên cứu. Nhiều nhóm vi sinh vật
trong tự nhiên có khả năng chuyển hóa các hợp chất chứa nitơ và photpho trong
nước thải thành các chất không độc hại với môi trường. Màng sinh học được định
nghĩa là dạng sống tồn tại phổ biến trong tự nhiên và khác biệt với dạng tế bào sống
tự do bởi mạng lưới các hợp chất ngoại bào bao quanh và những thay đổi, biệt hóa
trong tế bào để vi sinh vật thích nghi với môi trường sống. Vi sinh vật hình thành
màng sinh học không chỉ giúp chúng tồn tại và chống chịu được với những điều
kiện bất lợi, tận dụng được nguồn dinh dưỡng của môi trường mà còn thông qua
mối quan hệ hợp tác giữa các loài khác để tăng quá trình phân giải các chất độc hại
trong môi trường.
Việc nghiên cứu xử lý ô nhiễm nguồn nước thải nói chung và nước thải có
hàm lượng nitơ, photpho cao bằng việc sử dụng các vi sinh vật tạo màng sinh học là
hướng nghiên cứu mới mang tính bền vững. Tuy nhiên, hiện nay chưa có nhiều
công trình công bố về kết quả phân lập cũng như khả năng chuyển hóa nitơ,
photpho của các các chủng vi sinh vật có khả năng tạo màng sinh học. Chúng tôi
tiến hành thực hiện đề tài :”Nghiên cứu phân lập tuyển chọn các chủng vi sinh
vật ứng dụng xử lý nước thải giàu Nitơ, Photpho” nhằm mục đích góp phần tìm

Luận văn thạc sĩ
Ngô Thị Kim Toán 2 K19 – Sinh học thực nghiệm
hiểu sự đa dạng về các chủng vi sinh vật tạo màng trong tự nhiên cũng như khả
năng ứng dụng trong xử lý nước ô nhiễm.

Luận văn thạc sĩ
Ngô Thị Kim Toán 3 K19 – Sinh học thực nghiệm
CHƯƠNG 1: TỔNG QUAN TÀI LIỆU
1.1. Tình trạng ô nhiễm môi trường nước hiện nay ở Việt Nam và thế giới
Hiện nay, ô nhiễm môi trường là vấn đề đang được quan tâm không chỉ ở
Việt Nam mà còn ở nhiều quốc gia trên thế giới. Nước thải là một trong những
nguyên nhân gây ô nhiễm môi trường hiện nay. Ô nhiễm nguồn nước không chỉ ảnh
hưởng đến môi trường sống của con người, mà còn ảnh hưởng đến đa dạng sinh
học, đến môi trường sống của các loài động, thực vật trên trái đất.
Theo báo cáo môi trường Quốc gia năm 2010 của Bộ Tài Nguyên và Môi
Trường, từ năm 2007 đến năm 2009, ô nhiễm môi trường nước mặt ở tất cả các chỉ
số đều vượt quá tiêu chuẩn cho phép theo QCVN 08:2008. Các chỉ số COD, BOD
đều vượt quá tiêu chuẩn từ 5 đến 10 lần. Hàm lượng NH4+ trong môi trường nước
mặt của sông Nhuệ, sông Đáy và sông Cầu đều vượt quy chuẩn cho phép QCVN
08:2008/BTNMT cho nước mặt phù hợp với việc bảo tồn động thực vật thủy sinh là
là 0,2 mg/l. Năm 2009, hàm lượng NH4+ trong nước sông Nhuệ đo tại Cự Đà trên
10 mg/l vượt quá tiêu chuẩn 50 lần, sông Đáy đo tại Cầu Hoàng 3 mg/l vượt quá
tiêu chuẩn 15 lần, sông Cầu đo tại Thái Nguyên trên 22 mg/l vượt quá tiêu chuẩn
110 lần [2].
Theo Mulder, lượng hợp chất nitơ trong chuỗi thức ăn là 15 kg/người/năm,
một phần trong đó được con người tiêu thụ, phần lớn được thải ra ngoài môi trường.
Tính theo đầu người, mỗi người thải ra 4,75 kg nitơ một năm. Lượng nitơ trong
nước thải chiếm 30% lượng nitơ tiêu thụ [41]. Nước thải ở các đô thị chủ yếu ở
dạng nitơ hữu cơ và amoni, trong đó 60% ở dạng hữu cơ và 40% ở trạng thái amoni.
Ở Mỹ, hàm lượng nitơ có trong nước thải phụ thuộc vào số dân và lưu lượng nước
thải hằng ngày. Lượng nitơ thải vào nguồn nước trung bình là 16 g/người/ngày.
Hàm lượng và các loại hợp chất chứa nitơ thay đổi trong từng loại nước thải khác
nhau. Hàm lượng nitơ trong nước thải thường dao động trong khoảng 20 đến 85
mg/l trong đó nitơ ở dạng hợp chất hữu cơ trung bình từ 8 đến 35 mg/l, hàm lượng
N-NH3 từ 12 đến 50 mg/l [56].

Luận văn thạc sĩ
Ngô Thị Kim Toán 4 K19 – Sinh học thực nghiệm
Photpho là yếu tố quan trọng trong quá trình trao đổi chất và năng lượng của
sinh vật, khi nồng độ photpho quá cao trong môi trường làm tăng sinh của các loại
tảo gây hiện tượng phú dưỡng. Nồng độ photpho trong nước thải trung bình từ 6
đến 20 mg/l. Việc sử dụng các chất tẩy rửa trong sinh hoạt và sản xuất là một trong
số các nguyên nhân làm tăng hàm lượng photpho trong nước thải. Trong môi trường
nước mặt, nitrate, photphate là hai hợp chất của nitơ và photpho cần thiết cho sự
phát triển của rong, tảo. Hàm lượng photphate trong nguồn nước không ô nhiễm
nhỏ hơn 0,01 mg/l. Giá trị này ở sông Mêkông là nhỏ hơn 0,05 mg/l nhưng ở những
kênh rạch bị ô nhiễm nước thải do sinh hoạt và công nghiệp hàm lượng photphate
có thể lên tới 5 mg/l . Photphate là chất có nhiều trong phân, sản xuất phân lân, thực
phẩm, nước thải của các nhà máy chế biến phân lân, chế biến thủy sản. Theo quy
định của Hà Lan cũng như tiêu chuẩn của Việt Nam, hàm lượng photphate trong
nước uống không được vượt quá 6 mg/l. Theo tiêu chuẩn của cộng đồng chung châu
Âu, trong nước sinh hoạt, hàm lượng photphate không được vượt quá 2,18 mg/l [4].
Nước thải chăn nuôi là một trong những nguyên nhân gây ô nhiễm nguồn
nước. Ô nhiễm nước thải chăn nuôi đặc trưng là ô nhiễm hữu cơ, hàm lượng nitơ,
photpho cao và vi sinh vật gây bệnh, hàm lượng nitơ tổng số nằm trong khoảng từ
512 đến 594 mg/l, trong đó N-NH3 trong nước thải từ 304 đến 471 mg/l, hàm lượng
photpho tổng số dao động trong khoảng từ 13,8 đến 62 mg/l [6].
Ngày nay, cùng với sự phát triển của dân số, rác thải sinh hoạt ngày một gia
tăng. Ở Việt Nam, phương pháp xử lý rác thải chính vẫn là sử dụng các hố chôn lấp.
Nước rỉ rác từ các hố chôn lấp tại khu xử lý rác thải gây ảnh hưởng rất lớn đến đời
sống của người dân xung quanh, gây ô nhiễm nguồn nước mặt và nước ngầm quanh
khu vực. Trong nước thải rỉ rác chứa rất nhiều thành phần độc hại khác nhau trong
đó đặc biệt là hàm lượng chất hữu cơ cao. Tổng hàm lượng nitơ trong nước thải rỉ
rác dao động trong khoảng từ 200 đến 2000 mg/l, hàm lượng amoni cao, trung bình
200 mg/l, trong khi đó tiêu chuẩn cho phép là 0,2 mg/l[6].

Luận văn thạc sĩ
Ngô Thị Kim Toán 5 K19 – Sinh học thực nghiệm
1.2. Các phương pháp xử lý ô nhiễm nước thải có chứa hợp chất nitơ, photpho
hiện nay
Hiện nay có nhiều phương pháp xử lý nước thải được áp dụng như phương
pháp cơ học, phương pháp vật lý, phương pháp hóa học, phương pháp sinh học. Tất
cả các phương pháp xử lý hiện nay đều có những ưu, nhược điểm. Trong thực tế,
quá trình xử lý nước thải cần có sự kết hợp của nhiều phương pháp nhằm nâng cao
hiệu quả và giảm thời gian xử lý. Ví dụ, có thể sử dụng phương pháp cơ học giúp
loại bỏ các chất thải có kích thước lớn ban đầu, sau đó có thể áp dụng các phương
pháp hóa học, sinh học nhằm loại bỏ các chất độc bảo đảm tính bền vững cho môi
trường.
1.2.1. Phương pháp hóa học
Cơ sở của phương pháp hóa học là dựa trên các phản ứng hóa học. Các phản
ứng hóa học được ứng dụng trong xử lý nước thải như phản ứng oxy hóa, phản ứng
trung hòa, phản ứng keo tụ… giữa chất ô nhiễm và các hóa chất bổ sung.
1.2.1.1. Xử lý các hợp chất chứa nitơ bằng phương pháp hóa học
Quá trình xử lý nước thải chứa nitơ dựa trên nguyên tắc hóa học, nước thải
được đưa đến pH trong khoảng từ 10 đến 11 bằng cách thêm vào Ca(OH)2 để tạo
thành NH4OH, khi đó amoni chuyển từ trạng thái lỏng sang khí và sau đó được đưa
ra ngoài không khí qua các tháp làm lạnh [56]. Cheung và cộng sự đã sử dụng
Ca(OH)2 với nồng độ là 10 g/l, sau thời gian xử lý là 24 giờ và ở nhiệt độ từ 20 đến
23oC. Kết quả cho thấy, đã xử lý 65 – 75% NH4+ khi lưu lượng không khí bằng với
môi trường, và 86 – 93% NH4+ khi lưu lượng không khí là 5l/ phút [16]. Ozturk và
cộng sự đã áp dụng phương pháp này, sau 2 giờ, đã xử lý được 72 – 85% lượng
amoni trong nước thải rỉ rác khi bổ sung Ca(OH)2 với hàm lượng là 8 g/l và lưu
lượng không khí là 7,6 l/phút. Tuy nhiên phương pháp này có chi phí xử lý cao vì
đòi hỏi lượng không khí lớn và lượng Ca(OH)2 sau đó phải được xử lý với H2SO4
trước khi thải ra môi trường [48].
Li và cộng sự đã thử nghiệm một phương pháp để loại bỏ amoni trong nước
thải thông qua việc kết tủa amoni dưới dạng (NH4)MgPO4.6H2O khi thêm MgCl2 và

Luận văn thạc sĩ
Ngô Thị Kim Toán 6 K19 – Sinh học thực nghiệm
Na2HPO4 trong quá trình xử lý. Bằng phương pháp này, với tỷ lệ
Mg:NH4:PO4=1:1:1 và pH trong nước từ 8,5 đến 9, nồng độ amoni trong nước thải
rỉ rác giảm từ 5.600 mg/l xuống chỉ còn 110 mg/l trong 15 phút [38]. Phương pháp
này cũng đã được Yangin và cộng sự áp dụng đối với nước thải sinh hoạt, kết quả đã
loại bỏ được 66% lượng amoni trong nước thải và phương pháp này còn có thể ứng
dụng cho việc loại bỏ hợp chất chứa photpho trong nước thải [66]. Một phương
pháp để xử lý nitơ khác là bổ sung thêm clo vào nước thải trong quá trình xử lý. Khi
cho clo vào nước thải, NH3 sẽ phản ứng với clo dưới dạng HOCl để tạo ra các sản
phẩm trung gian là NH2Cl, NHCl2, NCl3. Quá trình xử lý sẽ diễn ra liên tục khi
thêm HOCl vào phản ứng để tạo ra sản phẩm cuối cùng là nitơ phân tử [56]. Quá
trình này diễn ra phụ thuộc rất nhiều vào nhiệt độ, pH, thời gian xử lý và tỷ lệ
HOCl/NH3 [49]. Với tỷ lệ HOCl/NH3 được tính theo mol bằng 1 tại pH trong
khoảng từ 7 đến 8 tất cả NH3 đều chuyển hóa thành NH2Cl. Với tỉ lệ HOCl/NH3
bằng 2 sản phẩm chủ yếu là NHCl2, khi tỉ lệ trên bằng 3 thì sản phẩm xử lý tạo ra
chủ yếu là NCl3 [3].
1.2.1.2. Xử lý các hợp chất photpho bằng phương pháp hóa học.
Photphate là một trong các hợp chất của photpho chiếm tỷ lệ cao trong nước
thải. Quá trình xử lý nước thải có chứa hợp chất photpho bằng phương pháp hóa
học dựa trên nguyên tắc chủ yếu là kết tủa photphate với các ion nhôm, sắt, canxi
tạo ra các muối có độ tan thấp và tách chúng dưới dạng chất rắn [4], [6].
Phương pháp hóa học được sử dụng trước tiên trong xử lý nước thải là sử
dụng kiềm hóa bằng Ca(OH)2. Khi thêm Ca(OH)2 vào nước thải, pH sẽ tăng làm
dịch chuyển cân bằng về PO43-. Tỷ lệ Ca/P nằm trong khoảng 1,33 đến 2,0 và ion
Ca2+ có khả năng loại bỏ photphate do nó tạo với photphate những hợp chất kém
hoà tan. Hydroxy apatit C10(PO4)6(OH)2 không xuất hiện ngay trong quá trình hình
thành dù nó là thành phần ổn định nhất về mặt nhiệt động và kém hòa tan nhất trong
số các kết tủa của photphate canxi. Photphate canxi Ca3(PO10)2 vô định hình là dạng
có cấu trúc tinh thể không ổn định và có độ tan thấp [4].

Luận văn thạc sĩ
Ngô Thị Kim Toán 7 K19 – Sinh học thực nghiệm
Khả năng loại bỏ photphate trong nước thải có hiệu quả cao khi ở pH >10,
đặc biệt khi ở pH từ 10,5 đến 11. Đặc điểm của phương pháp dùng Ca(OH)2 là làm
tăng độ kiềm của nước, thuận lợi cho phản ứng phân hủy sinh học của NH4+, không
đưa anion mới vào nước thải so với cách dùng muối để kết tủa photphate. Ngoài
phương pháp sử dụng Ca(OH)2, loại bỏ phosphate bằng phương pháp hóa học có
thể bằng việc kết tủa sử dụng muối sắt, nhôm, các muối sắt sử dụng là: FeCl3,
FeClSO4, FeSO4 khi bổ sung vào nước thải, sẽ xảy ra quá trình thủy phân tạo ra các
phức chất mang điện tích dương như Fe(OH)2+, Fe(OH)2+ và một số dimer, polymer
tích điện dương. Tại một giá trị pH không đổi, với tỷ lệ Fe3+/P từ 1,4 đến 1,6 có thể
kết tủa hoàn toàn photphate. Nếu tỷ lệ Fe3+/P tăng, sản phẩm kết tủa tạo thành là
Fe(OH)3 tăng lên như vậy kết tủa đó không phải là kết tủa của muối photphate. Do
vậy nếu tỷ lệ Fe3+/P tăng lên thì hiệu suất của quá trình xử lý không thay đổi[55].
Quá trình xử lý nước thải giàu hợp chất nitơ, photpho bằng phương pháp hóa
học chủ yếu là thêm một số chất làm đông tụ, keo tụ chất thải thành các muối kết
tủa. Tuy nhiên, việc tách các muối này ra khỏi nước thải phải dùng đến các màng
lọc: màng nano, màng thẩm thấu ngược, màng thẩm điện tích. Các màng lọc thường
có giá thành cao nên việc áp dụng phương pháp hóa học chưa phù hợp với nhu cầu
ứng dụng trong xử lý nước thải hiện nay. Do vậy, cần có phương pháp khác có khả
năng xử lý tốt ô nhiễm mà chi phí phù hợp [4].
1.2.2. Phương pháp sinh học
Trong môi trường nước, hợp chất nitơ tồn tại chủ yếu ở dạng amoni (NH4+),
nitrate (NO3-), ít hơn ở dạng nitrite (NO2
-) và trong một số hợp chất hữu cơ khác.
Thành phần được xem là bền đối với trường và không gây hiệu quả xấu cho môi
trường là khí nitơ (N2). Nitơ hữu cơ có thể tồn tại trong các sinh vật sống hoặc các
sản phẩm trung gian của quá trình phân hủy các vật chất hữu cơ [53], [54], [70]. Xử
lý nước thải có chứa hợp chất nitơ dựa trên các vi sinh vật có khả năng chuyển hóa
thành các chất không độc như khí N2, trả lại môi trường không khí. Phương pháp
sinh học có những ưu điểm so với các phương pháp vật lý hóa học như: hiệu suất

Luận văn thạc sĩ
Ngô Thị Kim Toán 8 K19 – Sinh học thực nghiệm
khử nitơ cao, sự ổn định và tương đối dễ vận hành, quản lý, chi phí đầu tư hợp lý và
quan trọng cho sự phát triển bền vững, bảo vệ môi trường và hệ sinh thái.
Để xử lý nước thải chứa nitơ theo phương pháp sinh học. Các nghiên cứu
dựa trên cơ sở là trong tự nhiên luôn tồn tại các vi sinh vật có khả năng chuyển hóa
hợp chất nitơ. Quá trình chuyển hóa nitơ trong tự nhiên được trình bày ở hình 1.1
[53], [57].
Các quá trình trong chu trình nitơ chuyển đổi nitơ từ dạng này sang dạng
khác đều được tiến hành bởi các nhóm vi sinh vật khác nhau với mục đích lấy năng
lượng hoặc để tích tụ nitơ thành một dạng cần thiết cho sự phát triển của chúng.
Các dạng nitơ hữu cơ từ nguồn động thực vật sau khi chết được các vi khuẩn amoni
hóa chuyển hóa thành dạng NH4+ ; sau đó NH4
+ được chuyển hóa thành NO2- nhờ vi
khuẩn nitrite hóa; NO2- sinh ra được nhóm sinh vật nitrate hóa chuyển hóa thành
NO3-; cuối cùng nitrate được nhóm sinh vật kỵ khí chuyển thành dạng nitơ phân tử
nhờ quá trình khử nitrate (Hình 1.1) [53], [54], [70].
Hình 1.1. Chu trình nitơ trong tự nhiên [69], [70]

Luận văn thạc sĩ
Ngô Thị Kim Toán 9 K19 – Sinh học thực nghiệm
Xử lý nước thải có chứa các hợp chất photpho bằng phương pháp sinh học
dựa trên khả năng của một số nhóm vi sinh vật tích lũy lượng photpho nhiều hơn
mức cơ thể chúng cần trong điều kiện hiếu khí. Thông thường hàm lượng photpho
trong vi sinh vật chiếm từ 1,5 đến 2,5% khối lượng tế bào khô, một số loài có khả
năng hấp thu cao hơn, từ 6 đến 8%. Vì các hợp chất chứa photpho tồn tại trong
nước thải dưới ba dạng: photphate đơn (PO43-), polyphotphate và hợp chất hữu cơ
chứa photphate, hai hợp chất sau chiếm tỉ lệ lớn trong nước thải. Trong quá trình xử
lý vi sinh, lượng photpho hao hụt từ nước thải duy nhất là lượng được vi sinh vật
hấp thu để xây dựng tế bào. Trong quá trình xử lý hiếu khí, một số loài vi sinh vật
có khả năng hấp thu photphate cao hơn mức bình thường trong tế bào và tồn tại ở
dạng dự trữ [3], [14].
Nghiên cứu Van Bethum và cộng sự, cho thấy photpho trong cơ thể vi sinh
vật được tích lũy dưới dạng chủ yếu là photphate. Trong cơ thể của chúng,
photphate có thể chiếm đến 12% trọng lượng tế bào đối với vi khuẩn có tích lũy
polyphotphate, và với vi khuẩn không tích lũy polyphotphate, chỉ chiếm khoảng
1đến 3% trọng lượng tế bào [63]. Trong điều kiện yếm khí, với sự có mặt của chất
hữu cơ, lượng photphate dư lại được thải ra ngoài cơ thể vi sinh dưới dạng
photphate đơn. Một vài loại tảo cũng có khả năng tích trữ một lượng photphate dư
so với nhu cầu của tế bào [3].
Hiện nay, kết hợp phương pháp sinh học trong xử lý đối với cả nitơ, photpho
trong nước ô nhiễm đang là một hướng nghiên cứu mới. Trong nghiên cứu của
Jorgensen và Pauli, một số chủng vi sinh vật có khả năng tích lũy photpho cũng có
khả năng khử nitrate [33]. Phương pháp kết hợp sử dụng bùn hoạt tính, các hợp
chất trong các quá trình xử lý thiếu khí (anoxic), xử lý hiếu khí (aerobic), xử lý yếm
khí (anaerobic) kết hợp hoặc riêng biệt trong quá trình khử nitơ và photpho [3].

Luận văn thạc sĩ
Ngô Thị Kim Toán 10 K19 – Sinh học thực nghiệm
1.3. Các vi sinh vật có khả năng chuyển hóa các hợp chất chứa nitơ, photpho
trong xử lý ô nhiễm nước thải.
1.3.1. Vi sinh vật có khả năng chuyển hóa các hợp chất chứa nitơ
Trong nước thải có nhiều thành phần khác nhau, bao gồm các hợp chất hữu
cơ và vô cơ. Trong đó các hợp chất chứa nitơ và photpho chiếm tỷ lệ lớn.
Nitrate hóa là quá trình oxi hóa NH4+ thành NO3
-, cung cấp năng lượng cho
vi sinh vật hoạt động. Quá trình oxi hóa này xảy ra cùng với quá trình đồng hóa
CO2. Hầu hết các vi sinh vật tự dưỡng hóa năng vô cơ thuộc loại hiếu khí bắt buộc
đều có khả năng thực hiện quá trình này. Nitrate hóa qua 2 giai đoạn:
Đầu tiên là giai đoạn oxi hóa amoni (NH4+) thành nitrite (NO2
-) bởi một số
đại diện thuộc nhóm vi khuẩn nitrite hóa: Nitrosomonas, Nitrosocystis,
Nitrosococcus, Nitrosolobus... Tất cả chúng khá giống nhau về mặt sinh lý, sinh
hóa, chỉ khác nhau về mặt hình thái học và cấu trúc tế bào. Các đại diện của chi
Nitrosomonas không sinh nội bào tử, tế bào nhỏ bé hình bầu dục. Trên môi trường
lỏng, quá trình phát triển của vi khuẩn thuộc chi Nitrosomonas trải qua một số giai
đoạn và phụ thuộc rất nhiều vào điều kiện môi trường [12], [27].
Giai đoạn 2 của quá trình nitrate hóa oxi hóa nitrite (NO2-) thành nitrate
(NO3-) bởi một số vi khuẩn: Nitrobacter Winogradsky, Nitrospina gracilis,
Nitrococcus mobilis.
NH4+ + 1,5 O2 Nitrosomonas NO2
- + H2O + 2 H+
NO2- + 0,5 O2
Nitrobacter NO3
-
NH4+ + 2 O2 NO3
- + H2O + 2 H+
4 NO3- + 4 H+ + 5 Chữu cơ 5 CO2 + 2 N2 + 2 H2O
Tế bào đặc trưng của nhóm vi khuẩn Nitrobacter trong dịch nuôi là dạng
hình que tròn, hình hạt đậu, hoặc hình trứng, có thể di động hoặc không di động.
Khi điều kiện không thuận lợi chúng có thể liên kết với nhau thành tập đoàn.
Nitrospina gracilis là những trực khuẩn thẳng, thỉnh thoảng có dạng hình cầu,
không di động, và có đặc trưng là liên kết tạo thành tập đoàn. Nitrococcus mobilis
thì có dạng hình tròn, có tiêm mao [5].

Luận văn thạc sĩ
Ngô Thị Kim Toán 11 K19 – Sinh học thực nghiệm
pH thích hợp cho nhóm vi khuẩn Nitrosomonas là từ 7,8 đến 8, Nitrobacter
là từ 7,3 đến 7,5. Nitrobacter sẽ tăng trưởng chậm hơn ở các mức pH cao đặc trưng
cho các thủy vực nước mặn. Nitrosomonas sống ở những nơi giàu NH3 và các muối
vô cơ như trong bùn đáy ao, nước cống, nước ngọt, các thủy vực bị ô nhiễm chứa
nhiều hợp chất nitơ nhằm tránh ánh sáng. Nitrobacter không có khả năng di động và
cần phải bám vào bề mặt giá thể như đá, cát, hoặc một giá thể sinh học …
Nitrobacter không thể sống trong môi trường khô. Trong môi trường nước, chúng
có thể tồn tại trong khoảng thời gian ngắn ở các điều kiện bất lợi nhờ vào việc sử
dụng các chất dự trữ bên trong tế bào [31].
Oxy hóa amoni bao gồm hai phản ứng kế tiếp nhau nên tốc độ oxy hóa của
quá trình bị khống chế bởi gian đoạn có tốc độ thấp hơn. Tốc độ phát triển của
Nitrosomonas chậm hơn Nitrobacter do đó nồng độ NO2- thấp hơn trong giai đoạn
ổn định. Vì vậy trong quá trình động học người ta chỉ sử dụng các thông số liên
quan đến vi khuẩn Nitrosomonas để đặc trưng cho quá trình oxy hóa amoni [31].
Quá trình chuyển hóa các hợp chất chứa nitơ không chỉ diễn ra do các chi vi
khuẩn nói trên mà trong tự nhiên còn nhiều nhóm vi sinh vật khác cũng có khả năng
chuyển hóa như vậy. Nghiên cứu khả năng chuyển hóa amoni, nitrite, nitrate,
Zhang và cộng sự đã phân lập được chủng vi khuẩn Pseudomonas stutzeri không
chỉ có khả năng chuyển hóa nitrite mà còn có khả năng chuyển hóa amoni. Sau thời
gian 18 giờ, chủng vi khuẩn này đã chuyển hóa được amoni hoàn toàn thành dạng
khí N2 với hiệu suất là 39% [67]. Một Nghiên cứu khác của Zhang và cộng sự về
việc sử dụng vi khuẩn Bacillus methylotrophicus trong xử lý nitơ cho thấy trong
môi trường ở điều kiện pH từ 7 đến 8 và nhiệt độ là 37oC, chủng B.
methylotrophicus đã làm nồng độ NH4+ ban đầu 146,71mg/l giảm xuống 38,29 mg/l
sau 9 ngày nuôi cấy, với tốc độ chuyển hóa là 51,58 mg/l/ngày [68].
Nghiên cứu của Broda cho thấy sự tồn tại của các vi khuẩn gọi là anammox
[15]. Các vi khuẩn này có thể oxy hoá amoni trong điều kiện kị khí (Anaerobic
Amoni Oxidation) hay còn gọi là anammox. Các nhà khoa học Hà Lan và Đức đã
nghiên cứu và phát hiện ra các vi khuẩn này thuộc năm chi gồm Brocadia,

Luận văn thạc sĩ
Ngô Thị Kim Toán 12 K19 – Sinh học thực nghiệm
Kuenenia, Anammoxoglobus, Jettenia và Scalindua, bộ Planctomycetales, ngành
Planctomycetes [42], [52]. Sử dụng kỹ thuật sinh học phân tử cũng như đặc điểm
sinh lý, sinh hóa Schmid cùng cộng sự đã phân loại được vi khuẩn Kuenenia
stuttgartiensis thuộc nhóm vi khuẩn anammox [53]. Ngoài phương pháp xử lý sinh
học kết hợp hai quá trình nitrate hoá và khử nitrate có thể loại bỏ được amoni ra
khỏi nước thải trong thực tế còn tồn tại một số vi khuẩn có khả năng oxy hoá amoni
thành dạng khí N2 sử dụng nitrite được hình thành từ quá trình xử lý thay thế cho
việc phải sử dụng oxy cấp từ nguồn bên ngoài vào [42], [52].
Trong tự nhiên, ngoài các chi Nitrosomonas, Nitrobacter có khả năng chuyển
hóa hợp chất chứa nitơ có trong nước thải là NH4+, NO2
-, NO3- còn có một số chi vi
sinh vật khác như Bacillus, Pseudomonas….
NO3- là sản phẩm cuối cùng của quá trình oxy hóa amoni chưa được coi là
bền vững và còn gây độc cho môi trường nên cần phải chuyển hóa về dạng khí N2.
Vi sinh vật thực hiện quá trình chuyển hóa là các vi sinh vật khử nitrate bao gồm
một số vi sinh vật thuộc các chi như Bacillus, Pseudomonas, Methanomonas,
Thiobacillus [22]. Các vi sinh vật khử nitrate sử dụng oxy hoặc nitrate, nitrite làm
chất oxy hóa để cung cấp năng lượng cho các quá trình sinh hóa.
Quá trình khử nitrate thường được nhận dạng là khử nitrat kị khí, tuy nhiên
diễn biến quá trình sinh hóa không phải là quá trình lên men kị khí mà nó giống quá
trình hô hấp hiếu khí nhưng thay vì sử dụng oxy, vi sinh vật sử dụng nitrate, nitrite.
Để khử nitrate, vi sinh vật cần có chất khử (nitrate là chất oxy hóa), chất khử có thể
là chất hữu cơ hoặc vô cơ như H2, S, Fe2+. Phần lớn vi sinh vật khử nitrate thuộc
loại dị dưỡng, sử dụng nguồn carbon hữu cơ để xây dựng tế bào ngoài phần sử dụng
cho phản ứng khử nitrate. Rất ít vi sinh vật khử nitrate thuộc loại tự dưỡng, ví dụ
Thiobacillus denitrificant sử dụng lưu huỳnh làm chất khử [3], [27].
1.3.2. Vi sinh vật có khả năng tích lũy photpho
Hợp chất photpho trong môi trường nước thải tồn tại trong các dạng:
photpho hữu cơ, muối photphate của các dạng H2PO4-, HPO4
2-, PO43- tan trong
nước, polyphotphate hay còn gọi là photphate trùng ngưng, muối photphate và

Luận văn thạc sĩ
Ngô Thị Kim Toán 13 K19 – Sinh học thực nghiệm
photpho trong tế bào sinh khối. Tất cả các dạng polyphotphate đều có thể chuyển
hóa về dạng orthophotphate (PO43-) trong môi trường nước đặc biệt là trong điều
kiện môi trường axit và ở nhiệt độ cao [4], [6].
Nhiều loài vi sinh vật tham gia vào quá trình hấp thu, tích lũy photpho vào
cơ thể của chúng được quy chung về nhóm vi sinh bio-P mà vi khuẩn thuộc chi
Acinetobacter là chủ yếu. [51], [60].
Trong điều kiện hiếu khí (O2) vi sinh bio-P tích lũy, tổng hợp photphate
trong cơ thể chúng từ photphate đơn tồn tại trong nước thải theo phương trình phản
ứng:
C2H4O2 + 0,16 NH4+ + 1,2 O2 + 0,2 PO4
3- 0,16 C5H7NO2 + 1,2 CO2 +
0,2 (HPO3) + 0,44 OH- + 1,44 H2O.
HPO3 là photphate ở dạng trùng ngưng tồn tại trong cơ thể vi sinh vật. Trong
điều kiện thiếu khí (không có oxy, chỉ có mặt nitrat) quá trình tích lũy photpho xảy
ra:
C2H4O2 + 0,16 NH4+ + 0,2 PO4
3-+0,96 NO3- 0,16 C5H7NO2+ 1,2 CO2
+ 0,2 (HPO3) + 1,4 OH- + 0,48 N2 + 0,96 H2O
Từ phương trình chuyển hóa trên cho thấy, các chủng vi sinh vật có khả năng
tích lũy photpho cũng có khả năng khử nitrate [36], [58].
Đã có nhiều công trình nghiên cứu, phân lập đối với các chủng vi sinh vật có
khả năng tích lũy photpho và khử nitrate trong việc loại bỏ một số hợp chất chứa
nitơ, photpho trong nước thải. Acinetobacter thường được quan tâm bởi khả năng
tích lũy photpho trong cơ thể, ít được biết đến hơn với vai trò khử nitrate. Một số
chi vừa có khả năng tích lũy photpho vừa có khả năng khử nitrate như:
Acinetobacter, Aeromonas, Pseudomonas, Alcaligenes, nhóm Comamonas –
Pseudomonas, nhóm Flavobacterium – Cytophaga, Moraxella, Xanthomonas,
Paracoccus, Bacillus, Corynebacterium và nhiều chủng vi khuẩn Gram dương
khác nữa. Acinetobacter là một trong những chi vi khuẩn được biết đến với khả
năng tích lũy photpho cao [33], [59], [61]. Bao và cộng sự đã tiến hành nghiên cứu
khả năng thu nhận photpho của các vi khuẩn khác nhau trong điều kiện hiếu khí và

Luận văn thạc sĩ
Ngô Thị Kim Toán 14 K19 – Sinh học thực nghiệm
thiếu khí [11], sau 20 giờ nghiên cứu, vi khuẩn thuộc chi Pseudomonas có khả năng
thu nhận đến 14,34 mg/l khi tiến hành ở điều kiện thiếu khí. Các chi
Enterobacteriaceae, Alcaligenes, Staphylococcus và Bacillus trong điều kiện hiếu
khí có khả năng thu nhận photpho lần lượt tương ứng là 8,91 mg/l, 6,43 mg/l, 6,23
mg/l và 4,41 mg/l.
1.4. Màng sinh học và ứng dụng của màng sinh học trong việc xử lý ô nhiễm
nước thải giàu nitơ, photpho.
1.4.1. Màng sinh học
1.4.1.1. Định nghĩa về màng sinh học
Màng sinh học (biofilm) được định nghĩa là một tập hợp của vi sinh vật liên
kết với nhau thông qua mạng lưới polymer ngoại bào [21].
Màng sinh học có thể hình thành do tập hợp các tế bào của cùng một vi sinh
vật hay các vi sinh vật khác nhau. Trong tự nhiên, màng sinh học thường là sự liên
kết của vi khuẩn, nấm, tảo, xạ khuẩn. Trong màng sinh học các tế bào tập hợp thành
các đơn vị cấu trúc là các vi khuẩn lạc. Thành phần này đóng vai trò quan trọng
trong quá trình hình thành màng sinh học đặc biệt là ở giai đoạn đầu bởi nó quy
định đặc tính hình thành màng cho từng loài vi sinh vật, đảm nhiệm chức năng tiết
các hợp chất ngoại bào cũng như có chứa các yếu tố phụ trợ tế bào như lông roi,
lông nhung hỗ trợ cho việc bám dính của các tế bào khác lên bề mặt giá thể [44].
1.4.1.2. Thành phần và quá trình hình thành màng sinh học
Cấu trúc của màng sinh học trong tự nhiên gồm hai thành phần chính là các
tập hợp tế bào vi sinh vật và mạng lưới các chất ngoại bào (Extracellular Polymeric
Substances - EPS). Các tế bào của một hay nhiều loài vi sinh vật khác nhau, bám
dính trên bề mặt nhất định (có thể là hữu sinh hay vô sinh). Các tế bào liên kết với
nhau một cách có trật tự đảm bảo sự trao đổi thông tin liên tục diễn ra giữa các tế
bào. Có thể nói màng sinh học là dạng sống khá phổ biến của nhiều loài vi sinh vật
[37], [39].
Mạng lưới các chất ngoại bào (EPS) bao quanh các tế bào, tạo nên cấu trúc
đặc trưng cho biofilm. Mạng lưới ngoại bào có độ dày từ 0,2 đến 1 µm. Ở một vài

Luận văn thạc sĩ
Ngô Thị Kim Toán 15 K19 – Sinh học thực nghiệm
loài vi khuẩn độ dày của lớp EPS mỏng hơn nằm trong khoảng từ 10 đến 30 nm
[19]. Mạng lưới các chất ngoại bào có vai trò quy định sự sắp xếp tế bào đồng thời
tạo nên những kênh dẫn truyền nước bên trong biofilm nhờ đó mà các chất dinh
dưỡng cũng như nước có thể lưu thông trong biofilm tạo điều kiện cho việc khuếch
tán, phân phối chất dinh dưỡng đến khắp các tế bào vi sinh vật trong biofilm cũng
như loại bỏ đi những chất thải không cần thiết [35].
Về cơ bản màng sinh học được cấu tạo gồm rất nhiều tế bào của cùng một
loài hay từ các loài vi sinh vật khác, khối lượng tế bào vi sinh vật chiếm từ 2 đến
5% tổng khối lượng biofilm. Trong biofilm ngoài thành phần tế bào thì có tới 97%
là nước, 3 đến 6% còn lại là EPS và ion. Một tế bào vi khuẩn tùy thuộc vào điều
kiện môi trường khác nhau có thể hình thành biofilm ở các dạng khác nhau. Thành
phần polymer ngoại bào rất đa dạng tùy loài vi sinh vật, dạng biofilm và điều kiện
hình thành nhưng về cơ bản đều bao gồm các polysaccharide chiếm khoảng từ 40
đến 95%, từ 1 đến 60% là protein, từ 1 đến 10% là axit nucleic, và từ 1 đến 40%
lipit về khối lượng [19], [25]. Các hợp chất này thay đổi theo không gian và thời
gian tồn tại của màng sinh học. Về cơ bản màng sinh học càng dày và thời gian tồn
tại càng lâu thì có hàm lượng EPS càng nhiều.
Mật độ tế bào tập trung cao nhất ở lớp đỉnh của biofilm và giảm dần theo độ
sâu. Trái lại, thành phần EPS lại phong phú hơn ở vùng phía trong biofilm. Thành
phần EPS trong biofilm cũng khác biệt so với ở dạng sống tự do của chính vi khuẩn
đó [19]. EPS có thể chiếm 50% đến 90% của tổng cacbon hữu cơ của các màng sinh
học và có thể coi là thành phần chính của màng sinh học. EPS có thể thay đổi một
vài tính chất hóa học và vật lý, nhưng thành phần chính của nó chủ yếu vẫn gồm các
polysaccharides. Có thể là các polysaccharides trung tính hay là polyanionic là
thành phần chính của EPS vi khuẩn gram âm. Một số vi khuẩn gram dương, thành
phần hóa học của EPS có thể khác nhau và chủ yếu là cation [62].
Dựa trên các phương pháp phân tích di truyền học, sinh học phân tử, cùng
với những phân tích về mặt cấu trúc của màng sinh học, các nhà khoa học đã đưa ra
một mô hình cấu trúc màng sinh học cơ bản [18]. Trong mô hình này, vi khuẩn hình

Luận văn thạc sĩ
Ngô Thị Kim Toán 16 K19 – Sinh học thực nghiệm
thành nên các vi khuẩn lạc và được bao quanh bởi một mạng lưới chất ngoại bào
giúp các thành phần tế bào liên kết với nhau một cách có trật tự đảm bảo sự trao đổi
thông tin liên tục diễn ra giữa các tế bào đồng thời tạo nên những kênh dẫn truyền
dịch ngoại bào bên trong màng sinh vật. Nhờ đó, dịch tế bào có thể đi qua màng
sinh vật tạo điều kiện cho việc khuếch tán, phân phối chất dinh dưỡng đến khắp các
tế bào trong màng cũng như loại bỏ các chất thải. Sự hình thành biofilm là quá trình
phát triển mà trong đó vi khuẩn trải qua những thay đổi trong cách thức tồn tại để
chuyển từ dạng sống đơn bào, riêng rẽ sang dạng tập hợp nhiều tế bào, cố định một
chỗ và có sự sinh trưởng cũng như biệt hóa tế bào khác với dạng sống trôi nổi. Có
năm giai đoạn chính trong quá trình hình thành và phát triển của một biofilm (Hình
1.2) [40].
Hình 1.1. Các giai đoạn chính của quá trình hình thành một biofilm [40]
1. Giai đoạn gắn kết, 2. Hình thành lớp tế bào, 3. Hình thành mạng lưới ngoại bào,
4. Hình thành màng sinh học hoàn chỉnh, 5. Quá trình tách rời
Giai đoạn 1: Gắn kết thuận nghịch lên giá thể
Trong một số điều kiện nhất định và tùy thuộc đặc tính lý hóa, các vi khuẩn
có thể di chuyển hướng đến bề mặt bởi hóa ứng động và hình thành mối tương tác
tạm thời với bề mặt thông qua các lực tương tác yếu như lực Van der Waals, lực hút

Luận văn thạc sĩ
Ngô Thị Kim Toán 17 K19 – Sinh học thực nghiệm
tình điện, liên kết hydro. Nhờ khả năng di chuyển độc lập bằng các cử động co rút
tế bào hay sử dụng các tiêm mao, và khả năng tiết các chất ngoại bào giúp các tế
bào riêng rẽ được bao bọc trong một mạng lưới và bắt đầu sự hình thành màng sinh
vật. Giai đoạn gắn kết thuận nghịch có vai trò quyết định một màng sinh học có thể
được hình thành hay không [8], [46].
Giai đoạn 2: Hình thành lớp tế bào
Khi các tế bào đầu tiên bám dính chặt hơn trên bề mặt giá thể, lúc này các tế
bào sử dụng các chất hữu cơ trên bề mặt giá thể và trong môi trường để sinh trưởng
phát triển tạo nên các vi khuẩn lạc đồng thời cũng trải qua những thay đổi về số
lượng tế bào, số lượng loài cũng như cấu trúc tế bào nhất định. Sản sinh ra các hợp
chất ngoại bào, giúp cho các tế bào bám dính chặt, không thuận nghịch trừ khi có
tác động của các tác nhân vật lý, hóa học. Các tế bào giảm mức độ sinh trưởng, tiêu
giảm các phần phụ trợ tế bào.
Giai đoạn 3: Hình thành mạng lưới ngoại bào
Các hợp chất polymer ngoại bào tiếp tục được tạo ra bởi các tế bào để liên
kết các tế bào một cách có tổ chức đồng thời tạo thành cầu nối giữa các khuẩn lạc.
Chúng cũng có vai trò trong việc thu hút các tế bào sống trôi nổi (có thể là từ nhiều
loài khác nhau) trong môi trường. Kết quả là mật độ tế bào trong một màng sinh
học cũng như lượng các polymer ngoại bào tạo ra tăng lên. Một màng sinh học dần
được hình thành.
Giai đoạn 4: Hoàn thành một màng sinh học hoàn chỉnh
Khi tế bào vi sinh vật bám dính không thuận nghịch lên bề mặt thì quá trình
trưởng thành của màng sinh vật bắt đầu. Các tế bào phân chia và phát triển, hình
thành các cụm tế bào vi khuẩn và mở rộng về không gian, hình thành một cấu trúc
màng sinh học hoàn chỉnh. Từ một phạm vi ban đầu màng sinh học có thể mở rộng
về không gian cũng như độ phức tạp tùy thuộc vào điều kiện môi trường. Một màng
sinh học hoàn chỉnh có cấu trúc giống như tháp hình nấm được bao quanh bởi các
kênh vận chuyển nước có tính thẩm thấu cao tạo điều kiện cho việc vận chuyển chất
dinh dưỡng và oxy vào bên trong màng [28].

Luận văn thạc sĩ
Ngô Thị Kim Toán 18 K19 – Sinh học thực nghiệm
Giai đoạn 5: Quá trình tách rời
Khả năng phát triển của màng sinh học bị giới hạn do nhu cầu dinh dưỡng
của môi trường nuôi cấy và biểu hiện của các phân tử cảm ứng mật độ tế bào. Các
phân tử này được giải phóng ra nhằm đáp ứng với những hạn chế về dinh dưỡng, sự
tích tụ các sản phẩm độc hại và một số nhân tố khác, bao gồm các yếu tố pH, nguồn
cung cấp cacbon, oxy [45]. Trong một số trường hợp, khi màng sinh học đạt đến
khối lượng và một mức cân bằng động tối đa thì các tế bào trong đó sẽ tự tách rời
và cùng với các tế bào của một màng khác hình thành nên các vi khuẩn lạc. Sự phân
hủy các polymer ngoại bào có thể diễn ra trong điều kiện thiếu hụt dinh dưỡng hay
oxy bên trong màng sinh học. Quá trình này liên quan đến sự tăng cường biểu hiện
của các gen mã hóa cho enzyme phân hủy carbohydrate tạo nên các lực liên kết yếu
hơn trong màng sinh học, dẫn đến sự phân tách các tế bào riêng rẽ đồng thời operon
mã hóa cho protein lông roi được tăng cường biểu hiện để chuẩn bị cho các tế bào
sống tự do khi tách rời khỏi màng [43], [45].
1.4.2. Vai trò và ứng dụng của sự hình thành màng sinh học
1.4.2.1. Vai trò của sự hình thành màng sinh học
Sự hình thành màng sinh học mang lại lợi ích cho chính bản thân vi sinh vật.
Trong quá trình hình thành màng sinh học, các tế bào phải trải qua một số thay đổi
về hình thái, đặc tính sinh lý và một trong những thay đổi quan trọng là việc hình
thành mạng lưới các chất ngoại bào bao quanh. Mạng lưới này giúp giữ lại chất hữu
cơ không hòa tan từ môi trường nước xung quanh tạo điều kiện cho vi sinh vật sinh
trưởng, phát triển. Đồng thời nó cũng có vai trò trong việc kiến tạo cấu trúc không
gian 3 chiều đặc trưng cho màng sinh học bằng cách tạo nên một mức độ ổn định,
một sự cân bằng nội môi cho các vi sinh vật.
Bên cạnh đó một vai trò quan trọng của mạng lưới ngoại bào là đem lại khả
năng chống lại các tác nhân kháng khuẩn cho các tế bào sinh sống trong một màng
sinh học. Theo Flemming, vi khuẩn có thể có khả năng kháng đối với các tác nhân
gây hại (chất kháng sinh, chất hoạt động bề mặt...) cao gấp 1000 lần khi gắn kết với
nhau tạo thành màng sinh học so với tế bào sống trôi nổi. Mạng lưới các hợp chất

Luận văn thạc sĩ
Ngô Thị Kim Toán 19 K19 – Sinh học thực nghiệm
ngoại bào cũng được ghi nhận có khả năng giúp tế bào chống lại tác động của kim
loại nặng, các ion và chất độc, giúp tế bào tránh khỏi rất nhiều yếu tố gây tác động
xấu tới vi sinh vật từ môi trường như tia UV, pH, sốc thẩm thấu và sự khô hạn [23].
Những kênh vận chuyển nước nằm xen kẽ trong cấu trúc của màng sinh học,
giữa các vùng bao quanh vi khuẩn lạc được ví như là một hệ thống tuần hoàn.
Chúng hoạt động hiệu quả trong quá trình trao đổi chất với môi trường xung quanh,
do đó làm tăng hiệu quả trong việc sử dụng nguồn dinh dưỡng cũng như loại bỏ các
sản phẩm chuyển hóa có khả năng gây độc hại. Nhờ vậy quá trình chuyển hóa các
chất trong đó cũng mang những đặc trưng khác so với dạng sống tự do [7].
Mặt khác quá trình hình thàng màng sinh học giúp vi sinh vật tận dụng được
nguồn chất hữu cơ bám dính trên bề mặt giá thể cũng như các cơ chất, chất dinh
dưỡng tạo ra từ các loài vi sinh vật khác sống chung.
Một màng sinh học có thể được hình thành do sự hợp tác cùng chung sống
của nhiều loài vi sinh vật để tạo một cộng đồng có cấu trúc không gian phức tạp.
Các loài vi sinh vật cùng tồn tại trong biofilm thích nghi với những điều kiện về
dinh dưỡng, nồng độ khác nhau tạo nên những “vi ổ sinh thái” trong biofilm. Chẳng
hạn như những vi sinh vật nằm phía ngoài biofilm thích nghi với điều kiện hiếu khí
cao trong khi những loài nằm phía trung tâm biofilm có xu hướng chịu được nồng
độ oxy thấp (vi hiếu khí).
Khả năng thích nghi với nhiều điều kiện dinh dưỡng khác nhau giúp các loài
vi sinh vật tận dụng được nguồn dinh dưỡng từ môi trường đồng thời hỗ trợ lẫn
nhau theo hướng cùng có lợi trong quá trình chuyển hóa vật chất. Mối quan hệ hợp
tác giữa các loài trong biofilm cũng có ảnh hưởng lớn đến chu trình tuần hoàn của
các nguyên tố trong tự nhiên. Hầu hết các quá trình trong tự nhiên đòi hỏi sự phối
hợp của nhiều nhóm vi khuẩn có cơ chế trao đổi chất khác nhau để cùng phân giải
một hợp chất hữu cơ và việc các vi sinh vật thuộc nhiều nhóm khác nhau cùng cư
trú trong biofilm sẽ góp phần thúc đẩy các quá trình này diễn ra nhanh hơn [35].
Quá trình truyền gen ngang đóng vai trò quan trọng trong việc thúc đẩy sự
tiến hóa của các cộng đồng vi sinh vật. Trong đó cơ chế truyền gen phổ biến ở vi

Luận văn thạc sĩ
Ngô Thị Kim Toán 20 K19 – Sinh học thực nghiệm
sinh vật là truyền gen thông qua plasmid và cầu tiếp hợp. Tuy nhiên từ những hiểu
biết rằng hầu hết các vi khuẩn trong tự nhiên định cư dưới dạng biofilm, liên kết với
nhau bởi mạng lưới các chất ngoại bào thì việc tiếp hợp giống như là cơ chế mà nhờ
đó vi khuẩn trong biofilm có thể truyền gen từ tế bào này sang tế bào khác [35].
1.4.2.2. Ứng dụng của màng sinh học trong xử lý ô nhiễm
Màng sinh học tác động đến rất nhiều lĩnh vực trong cuộc sống hàng ngày.
Do vậy nhiều nghiên cứu hiện nay về màng sinh học có ý nghĩa thực tiễn quan trong
và ngày càng thu hút sự quan tâm của nhiều nhà khoa học. Một số ứng dụng cụ thể
của màng sinh học nói riêng và các chủng vi sinh vật tạo màng sinh học nói chung
là xử lý ô nhiễm.
Trong công nghiệp lên men tại các bể lên men là nơi giữ lại sinh khối vi sinh
vật. Thông thường các tế bào ở dạng tự do khó có khả năng được giữ lại trong các
bồn lên men sau mỗi mẻ xử lý. Khi đó để tiếp tục một qui trình mới lại phải bổ sung
thêm một lượng sinh khối nhất định và đợi thời gian để vi sinh vật có thể sinh
trưởng, phát triển tới một nồng độ nhất định mới. Qui trình này gây tốn kém ở khâu
nguyên liệu đầu vào cũng như mất thời gian vận hành. Ngược lại khi đã được bám
giữ trên bề mặt giá thể bằng mạng lưới biofilm sinh khối vi sinh vật có thể được giữ
lại một cách có hiệu quả sau mỗi mẻ xử lý. Những giá thể chất mang có sẵn mạng
lưới biofilm có thể được tái sử dụng ở những lần xử lý tiếp theo mà không phải bổ
sung thêm vi sinh vật cũng như đợi thời gian phát triển [29].
Dầu thô và các sản phẩm từ dầu được loại bỏ bởi các vi khuẩn phân hủy
hydrocarbon. Các vi khuẩn được sử dụng có thể được thả trực tiếp xuống vùng dầu
tràn hoặc có thể thả ở vùng ven bờ mà dầu tràn bị sóng đánh vào. Lý do chính ở đây
là biofilm giúp tăng hiệu quả lọc nước và làm tăng độ kết dính của vi sinh vật với
bề mặt giá thể nơi có dầu tràn. Trong nghiên cứu của Radwan và cộng sự, khi sử
dụng các vi khuẩn thuộc chi Acinetobacter và dùng một lớp phủ làm giá thể cho vi
khuẩn là tảo, kết quả đã làm giảm được 64-98% n-octadecane và khoảng 38-56%
phenanthrene từ môi trường có chứa 0,03% của hydrocarbon sau 2 tuần [50]. Trong
nghiên cứu của Lê Thị Nhi Công cùng cộng sự, đã phân lập từ biển nhóm vi khuẩn

Luận văn thạc sĩ
Ngô Thị Kim Toán 21 K19 – Sinh học thực nghiệm
tạo biofilm và có hoạt tính chuyển hóa các chất hydrocacbon thơm đa vòng như
napthalene, anthracene, pyren [17].
Một trong những ứng dụng của màng sinh học đang được quan tâm liên quan
đến việc làm sạch nguồn nước thải, nguồn nước ngầm bằng công nghệ sinh học.
Ứng dụng này bắt nguồn từ thực tế là bản thân vi sinh vật có khả năng phân hủy các
chất hữu cơ trong môi trường tự nhiên thành các chất vô cơ đơn giản, ít độc. Đã có
nhiều phương pháp xử lý nước thải bao gồm những giai đoạn xử lý mà trong đó
nước thải được lọc qua các biofilm nhằm mục đích tách và đồng hóa các hợp chất
hữu cơ có hại. Một lượng sinh khối lớn các vi sinh vật trong mạng lưới biofilm làm
tăng sự hợp tác trong quá trình trao đổi chất, giúp cho quá trình loại bỏ các chất gây
ô nhiễm trong nước diễn ra hiệu quả hơn so với dạng sống tự do. Quá trình phân
hủy các chất cũng tỏ ra hiệu quả hơn khi thường sản phẩm của chủng này lại là cơ
chất cho một chủng khác trong mạng lưới biofilm, ví dụ trong một mạng lưới
biofilm xử lý nước thải có chứa hợp chất nitơ, ion NH4+ được nhóm Nitrosomonas,
Nitrobacter chuyển hóa thành ion NO3-, rồi tiếp tục được các nhóm vi khuẩn yếm
khí khác sử dụng để cuối cùng tạo thành N2 đi vào khí quyển [37].
Một số nghiên cứu về vi khuẩn anammox có khả năng xử lý nitơ trong nước
thải, đã chỉ ra rằng trong hệ thống các lớp siêu mỏng của lớp màng biofilm của
chủng vi khuẩn Planctomycetes có sự phân bố oxy theo lớp. Các lớp phía trên là
những lớp giàu oxy trong khi các lớp ở phía dưới cùng nằm trong trạng thái kị khí.
Sự phân chia theo lớp màng sinh học sẽ tạo điều kiện thuận lợi trong quá trình ứng
dụng xử lý nước thải giàu hợp chất nitơ vì giai đoạn nitrate hóa là giai đoạn hiếu
khí, giai đoạn khử nitrate là giai đoạn kị khí [52].
Những nghiên cứu về biofilm trong xử lý nước thải có chứa các hợp chất
nitơ và photpho của Boelee và cộng sự, các nhà nghiên cứu đã sử dụng màng sinh
học của vi tảo để thực hiện nghiên cứu này. Kết quả cho thấy, màng sinh học được
thiết kế dựa vào các vi tảo đã xử lý được nitơ là 1.0 g/m2/ngày và photpho là 0.13
g/m2/ngày [13].

Luận văn thạc sĩ
Ngô Thị Kim Toán 22 K19 – Sinh học thực nghiệm
Wellander và cộng sự khi sử dụng một vật liệu bám sinh khối, thả nổi trong
hệ thống xử lý làm giá thể cho các vi sinh vật có khả năng nitrate hóa, đã loại bỏ
được đến 90% lượng nitơ tổng số [64]. Hoilijoki và cộng sự đã nghiên cứu khả năng
nitrate hóa của vi sinh vật thuộc nhóm nitrate hóa. Kết quả cho thấy, quá trình
nitrate hóa chỉ xử lý được 61% amoni khi không có vật liệu bám cho vi sinh vật, và
quá trình nitrate hóa xảy ra hoàn toàn khi có vật liệu bám cho vi sinh vật trong bể
phản ứng bùn hoạt tính [30]. Kết quả này cho thấy, quá trình xử lý nước thải sử
dụng màng sinh học sẽ tăng hiệu quả xử lý khi có mặt vật liệu bám cho vi sinh vật.
Bernet và cộng sự đã nghiên cứu ứng dụng khả năng chuyển hóa nitơ và tạo
màng sinh học của vi sinh vật. Mẫu ban đầu có hàm lượng NH4+ là 250 mg/l, sau 2
ngày, hàm lượng NH4+ giảm xuống chỉ còn 5 mg/l, hiệu quả của quá trình xử lý lên
đến 98% [12].
Kết quả nghiên cứu ứng dụng vi sinh vật có khả năng tạo màng sinh học
trong xử lý ô nhiễm nước thải đặc biệt là nước thải giàu nitơ và photpho hiện nay
chưa nhiều. Các công trình công bố liên quan đến lĩnh vực ứng dụng nghiên cứu
này chưa nhiều, còn thiếu cả về số lượng lẫn chất lượng. Tại Việt Nam tình hình ô
nhiễm nước thải ngày một gia tăng do đó việc cấp thiết là tìm một phương pháp xử
lý hiệu quả là cần thiết. Vì vậy, để phù hợp với mục đích nghiên cứu và ứng dụng
xử lý ô nhiễm tại Việt Nam, chúng tôi đã tiến hành thực hiện đề tài phân lập nghiên
cứu các chủng vi sinh vật có khả năng tạo màng sinh học và có khả năng xử lý nước
thải giàu hợp chất nitơ, photpho với mục tiêu:
Phân lập các chủng có hoạt tính tạo biofilm mạnh đồng thời có khả năng xử
lý nitơ và photpho
Bước đầu nghiên cứu tối ưu các điều kiện cho sự sinh trưởng phát triển của
các chủng vi sinh vật này để có thể áp dụng trong công nghệ xử lý nước thải giàu
nitơ và photpho

Luận văn thạc sĩ
Ngô Thị Kim Toán 23 K19 – Sinh học thực nghiệm
CHƯƠNG 2: NGUYÊN LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Nguyên liệu
Các mẫu nước thải từ bể Biogas tại Vĩnh Yên – Vĩnh Lộc – Thanh Hóa với
vị trí thu mẫu và được ký hiệu là các mẫu số 1,2, 3:
Nước thải lấy từ tầng giữa bể biogas đã ngưng sử dụng: mẫu số 1
Nước thải lấy từ tầng đáy bể biogas đã ngưng sử dụng: mẫu số 2
Nước thải trực tiếp chảy ra từ bể biogas đang sử dụng: mẫu số 3
Mẫu nước thải rỉ rác ở khu tập trung xử lý rác thải Vạn Phúc – Hà Đông –
Hà Nội với hai vị trí thu mẫu là trong và ngoài khu tập trung xử lý rác thải được ký
hiệu là mẫu số 4 và số 5.
Các mẫu sau khi lấy được chuyển về Phòng thí nghiệm Trọng điểm công
nghệ Enzym và Protein, Trường Đại học Khoa học Tự nhiên phân tích.
2.2. Hóa chất, thiết bị
2.2.1. Môi trường nuôi cấy
Môi trường Winogradsky được sử dụng để phân lập vi sinh vật trong quá
trình nitrate hóa.
Môi trường Winogradsky 1 Môi trườngWinogradsky 2
(NH4 )2SO4 2 g NaNO2 1 g
MgSO4 0,5 g MgSO4 0,5 g
NaCl 2 g NaCl 0.3 g
K2HPO4 1 g K2HPO4 1 g
CaCO3 0,001 g NaCO3 1 g
FeSO4 0,4 g FeSO4 0,03 g
Thạch 15 g Thạch 15 g
Nước cất 1 lít Nước cất 1 lít
Môi trường acetate mineral medium (AMM) được sử dụng để phân lập vi
sinh vật có khả năng xử lý photpho:
CH3COONa: 3,68 g

Luận văn thạc sĩ
Ngô Thị Kim Toán 24 K19 – Sinh học thực nghiệm
Na2HPO4: 28,73 mg
NH4Cl: 57,27 mg
MgSO4: 131,82 mg
K2SO4: 26,74 mg
CaCl2. 2H2O: 17,2 mg
HEPES: 12 g
Dung dịch khoáng: 2 ml
Thạch: 15 g
Nước cất: 1 lit
Dung dịch khoáng gồm:
EDTA: 50 g
FeSO4. 7H2O: 5 g
CuSO4. 5H2O: 1,6 g
MnCl2. 4H2O: 5 g
(NH4)6Mo7O24. 4H2O: 1,1 g
H3BO3: 50 mg
KI: 10 mg
CoCl2. 6H2O: 50 mg
Nước cất: 1 lit
Môi trường Luria Betani (LB) (g/l)
Peptone 10 g
Cao nấm men 5 g
NaCl 10 g
Nước cất 1 lit
Các môi trường được khử trùng ở nhiệt độ 121oC trong thời gian 20 phút.
Các hóa chất khác đều đạt độ tinh khiết cho nghiên cứu.
2.2.2. Máy móc thiết bị
Nồi khử trùng (ALP–Nhật).
Máy lắc (Satorius–Đức).

Luận văn thạc sĩ
Ngô Thị Kim Toán 25 K19 – Sinh học thực nghiệm
Box cấy vi sinh vật (Aura vertical–Ý).
Máy đo mật độ quang học (Bionate–Anh).
Máy đo pH (Horiba–Nhật Bản).
Cân Kern (Satorius–Đức).
Cân phân tích (Presica, Ý)
Tủ ấm (Memmert–Đức).
Máy ly tâm sigma 3K30 (Sartorius–Đức).
Tủ sấy (Memmert–Đức).
Máy khuấy từ (IKA–Đức).
Kính hiển vi điện tử quét JSM–5421LV (Nhật Bản).
2.3. Phương pháp nghiên cứu
2.3.1. Phương pháp phân lập vi khuẩn
Dùng các đĩa petri có chứa môi trường Winogradsky và môi trường AMM
đã khử trùng. Pha loãng các mẫu ở các nồng độ 10-1, 10-2, 10-3
, 10-4 và 10-5. Dùng
pipet lấy 100 µl dịch mẫu đã pha loãng cho lên mặt thạch. Dùng que cấy gạt đã vô
trùng gạt đều trên mặt thạch cho đến khi nào mặt thạch khô thì dừng lại. Các mẫu
đã cấy gạt cho vào tủ ấm ở 37oC trong thời gian từ 3 đến 5 ngày.
2.3.2. Phương pháp đánh giá khả năng hình thành biofilm
Nguyên tắc: trong điều kiện dinh dưỡng thích hợp và môi trường nuôi cấy
tĩnh các chủng vi sinh vật hình thành biofilm trên bề mặt giá thể. Phát hiện, quan sát
biofilm bằng cách nhuộm với tím kết tinh – là chất có khả năng bắt màu với tế bào.
Tiến hành thí nghiệm theo phương pháp của O’Toole và cộng sự [46], [47]. Các
chủng vi khuẩn được lắc kích hoạt trong bình tam giác chứa môi trường LB trong
24 giờ ở 37oC sao cho mật độ tế bào tại bước sóng 620 nm (OD620) ở vào khoảng
0,2 đến 0,3. Hút 100 µl dịch nuôi cấy vi khuẩn bổ sung vào 700 µl LB lỏng trong
các ống eppendorf đã khử trùng và ủ trong điều kiện tĩnh ở 37oC.
Sau 24 giờ các dịch nuôi cấy được loại bỏ khỏi các ống eppendorf. Đánh giá
mật độ tế bào sống trôi nổi trong môi trường bằng phương pháp so màu ở bước
sóng 620 nm dịch nuôi cấy vi khuẩn.

Luận văn thạc sĩ
Ngô Thị Kim Toán 26 K19 – Sinh học thực nghiệm
Quan sát khả năng hình thành biofilm: Mỗi ống eppendorf được rửa sạch 2
lần bằng nước cất khử trùng. Bổ sung vào mỗi ống eppendorf 1 ml dung dịch tím
kết tinh 1% và giữ trong 25 phút ở nhiệt độ phòng. Loại bỏ dung dịch nhuộm và
quan sát sự bắt màu của các tế bào bám trên trên thành ống với tím kết tinh.
Đánh giá mật độ tế bào trong biofilm: Sau khi rửa sạch 2 lần bằng nước cất
khử trùng các tinh thể tím bám trên thành eppenodorf đươc hòa tan trong 1 ml
etanol 70%. Mật độ tế bào trong biofilm được xác định bằng cách đo độ hấp thụ tại
bước sóng 570 nm.
2.3.3. Quan sát cấu trúc biofilm bằng chụp ảnh trên kính hiển vi điện tử quét
(SEM)
Chuẩn bị mẫu biofilm nổi: bổ sung dịch nuôi cấy lắc vi khuẩn vào bình tam
giác chứa 20 ml môi trường LB. Nuôi cấy tĩnh 24 giờ ở 37oC.
Gắn mẫu biofilm lên lamen, hơ nhẹ trên ngọn lửa đèn cồn để cố định mẫu.
Rửa nhẹ bằng nước cất khử trùng.
Gắn mẫu lên đế. Mạ phủ mẫu bằng vàng trên máy JFC–1200 trong 5 phút ở
30 mA.
Soi và chụp ảnh trên kính hiển vi quét điện tử JSM–5421LV (Nhật Bản) tại
phòng chụp Hiển vi điện tử quét – Trung Tâm Khoa Học Vật Liệu – Trường
Đại học Khoa học Tự Nhiên.
2.3.4. Ảnh hưởng của các điều kiện môi trường nuôi cấy lên sự hình thành
màng sinh học
Vi sinh vật có khả năng hình thành màng sinh học tốt ở những điều kiện khác
nhau. Nhiệt độ, pH không thích hợp sẽ ảnh hưởng đến sự sinh trưởng và phát triển
của vi sinh vật và khả năng tạo màng sinh học của chúng.
2.3.4.1. Ảnh hưởng của nhiệt độ môi trường nuôi cấy
Đánh giá khả năng hình thành màng sinh học của các chủng vi khuẩn trong
môi trường LB lỏng, nuôi cấy ở các nhiệt độ: 25oC, 30oC, 37oC, 45oC, 50oC, 55oC.
Sau 1 ngày nuôi cấy, tiến hành quan sát, đánh giá khả năng tạo biofilm của chủng vi
khuẩn nghiên cứu.

Luận văn thạc sĩ
Ngô Thị Kim Toán 27 K19 – Sinh học thực nghiệm
2.3.4.2. Ảnh hưởng của pH môi trường nuôi cấy
Đánh giá khả năng hình thành màng sinh học của các chủng vi khuẩn trong
môi trường LB lỏng, nuôi cấy ở các pH: 4, 5, 6, 6.5, 7, 7.5, 8, 9. Sau 1 ngày nuôi
cấy, tiến hành quan sát, đánh giá khả năng hình thành màng sinh học của chủng vi
khuẩn nghiên cứu.
2.3.5. Phương pháp nhuộm Gram.
Nguyên tắc: Dựa vào sự khác biệt giữa thành tế bào vi khuẩn Gram (+) và
Gram (-). Vi khuẩn Gram (+) có peptidoglican hoạt động như một hàng rào thẩm
thấu ngăn cản sự thất thoát của tím kết tinh. Ban đầu vi khuẩn được nhuộm bằng
tím kết tinh sau đó được xử lý bằng iôt để tăng độ giữ màu. Sau đó được tẩy màu
bằng cồn làm co các lỗ của lớp peptidoglican dày lại. Do vậy phức chất tím kết tinh
và iot được giữ lại, vi khuẩn có màu tím. Peptidoglican ở vi khuẩn Gram (-) rất
mỏng, ít liên kết chéo và có lỗ lớn. Sự xử lý bằng cồn có thể loại lipit khỏi thành
Gram (-) đủ để làm tăng hơn kích thước của lỗ. Do vậy, ở bước rửa bằng cồn đã loại
bỏ phức chất màu tím của tím kết tinh-iôt. Khi nhuộm lại bằng safain thì vi khuẩn
có màu hồng [1].
Phương pháp tiến hành:
Tạo vết bôi bằng cách nhỏ một giọt nước lên lam kính sạch, đốt nóng que
cấy trên ngọn lửa đèn cồn, dùng que cấy lấy khuẩn lạc trên đĩa thạch hoặc
trong ống giống, đưa vào giọt nước, cố định vết bôi bằng cách hơ khô trên
ngọn lửa đèn cồn.
Cho một giọt tím kết tinh bao phủ hoàn toàn vết bôi, nhuộm trong thời
gian 1 phút. Sau đó rửa bằng nước cất và thấm khô.
Nhuộm với lưugôn (chứa KI và I2) tương tự như với tím kết tinh.
Rửa bằng cồn trong 30 giây. Thấm khô.
Nhuộm safain trong 2 phút. Rửa bằng nước cất, thấm khô.
Soi trên kính hiển vi quang học có độ phóng đại 100 lần.
Quan sát kết quả và đưa ra kết luận.

Luận văn thạc sĩ
Ngô Thị Kim Toán 28 K19 – Sinh học thực nghiệm
2.3.6. Phương pháp sử dụng kit APi
Để kiểm tra khả năng đồng hóa các hợp chất hữu cơ điển hình của các chủng
vi khuẩn, chúng tôi đã tiến hành thử với Kit thử APi 20NE của hãng BioMérieus.
Hóa chất và thuốc thử cần thiết được cung cấp cùng với bộ kit.
Kết quả được ghi lại và phân loại với phần mềm của hãng BioMérieus .
2.3.7. Phương pháp đánh giá khả năng chuyển hóa các hợp chất nitơ
Mỗi chủng được nuôi lắc ở 37°C trong các khoảng thời gian từ 0, 5, 10, 15,
20 ngày. Môi trường nuôi lắc là dung dịch môi trường Winogradsky có bổ sung 1%
pepton [34].
2.3.7.1. Phương pháp phân tích nitơ tổng số
Nguyên tắc của việc phân tích nitơ tổng số là chuyển toàn bộ nitơ ở cả dạng
vô cơ và hữu cơ về dạng nitrate nhờ chất oxi hóa mạnh sau đó tiến hành so màu ở
bước sóng 220 nm [9].
Phương pháp làm như sau:
Xử lý mẫu
Lấy 50 ml mẫu vào bình chịu nhiệt 100 ml
Thêm 10 ml dd A (dd A : 4 g NaOH + 3 g K2S2O8)/100ml nước cất.
Nút chặt và đun trong nồi khử trùng ở 120°C trong 30 phút.
Sau đó lấy 30 ml mẫu vào bình định mức 50 ml, lọc loại bỏ vẩn đục.
Thêm 6,5 ml HCl (HCl:H2O = 1:15 về thể tích).
Lắc và định mức tới 50 ml bằng nước khử ion.
Lập đường chuẩn
Chuẩn bị đường chuẩn gồm 4 điểm bằng bình định mức 50ml
Dung dịch gốc 100 mg/l: hòa tan 0,722 g KNO3 bằng nước cất, sau đó
định mức lên 1000 ml, đựng trong chai màu nâu, bảo quản trong tủ lạnh.
Dung dịch chuẩn 10 mg/l bằng cách pha loãng dung dịch gốc ra 10 lần
Cách tiến hành: lấy thể tích dung dịch chuẩn theo tỷ lệ vào 4 bình định
mức, thêm vào mỗi bình 0,5 ml dung dịch HCl, định mức tới 50 ml bằng
nước cất, lắc đều. Sau đó đo ở bước sóng 220 nm

Luận văn thạc sĩ
Ngô Thị Kim Toán 29 K19 – Sinh học thực nghiệm
Với mẫu sau khi thực hiện bước xử lý mẫu xong cũng tiến hành tương tự
như bước lập đường chuẩn
Tính toán kết quả
mgN- T = mgN- T đo 2
Chú ý: giới hạn của phép đo là < 3 mg/l, nếu hàm lượng nitơ trong mẫu
nằm quá giới hạn thì phải pha loãng mẫu .
2.3.7.2. Phương pháp phân tích hàm lượng amoni (NH4+)
Khả năng chuyển hóa amoni của các chủng vi khuẩn được phân tích dựa trên
nồng độ amoni trong môi trường theo các khoảng thời gian nuôi.
Nguyên tắc:
Phản ứng của NH4+ và hypochloride với sự có mặt của xúc tác phenol tạo
thành hợp chất indophenol blue. So màu ở bước sóng 640 nm [9].
Chuẩn bị thuốc thử:
Hypochloride: 15 g NaOH khan + 500 ml NaClO 0.1% sau đó định mức
đến 1000ml bằng nước khử ion.
Dung dịch phenol- sodium nitroprusside: 5 g phenol (C6H5OH) + 0.025 g
nitroprusside (Na2[Fe(CN)5.NO].2H2O), định mức đến 500 ml bằng nước khử
ion.
Dung dịch chuẩn:
Dung dịch chuẩn được lấy từ NH4Cl tinh khiết sấy ở 105°C trong 3 giờ, đẻ
nguội trong bình hút ẩm. Cân 3,819 g NH4Cl, hòa tan và định mức đến 1000 ml
bằng nước khử ion. Dung dịch có nồng độ N- NH4+ là 1 g/l. Chuẩn bị dung dịch có
nồng độ 1 mg/l bằng cách pha loãng dung dịch gốc.
Lập đường chuẩn: tương ứng với các nồng độ 0; 0,2; 0,4; 0,6; 0,8; 1 mg/l.
Lấy 5 ml mẫu vào ống thủy tinh.
Thêm 3 ml thuốc thử phenol- sodium nitroprusside.
Lắc trên máy rung 30 giây.
Thêm 3 ml hypochloride.
Lắc trên máy rung 30 giây.

Luận văn thạc sĩ
Ngô Thị Kim Toán 30 K19 – Sinh học thực nghiệm
Điều nhiệt ở 37°C trong 25 phút.
Đo ở bước sóng 640 nm.
Với mẫu cũng làm các bước tương tự như lập đường chuẩn.
2.3.7.3. Phương pháp thử khả năng chuyển hóa nitrite
Khả năng chuyển hóa nitrite (NO2-) thành nitrate (NO3
-) của các chủng được
đánh giá thông qua sự giảm nồng độ nitrite trong môi trường nuôi cấy.
Nguyên tắc:
Phản ứng giữa nitrite và hỗn hợp sulfanylamide với naphtylendiamine tạo
phức màu hồng ở pH 2. So màu ở bước sóng 540 nm [9].
Chuẩn bị thuốc thử:
Dung dịch sunfanylamide: cân 0,6 g NH2C6H4SO2NH2 hòa tan trong 50 ml
nước cất nóng, làm lạnh, thêm 40 ml HCl đặc và định mức đến 100 ml.
Dung dịch naphtylendiamine dihydrochloride: cân 0.1 g
C10H7NHCH2NH2.2HCl hòa tan trong 100 ml nước cất, đựng trong chai màu
nâu, bảo quản 4oC.
Lập đường chuẩn:
Chuẩn bị các ống thí nghiệm.
Hòa tan 1,23 g NaNO2 trong 1000 ml nước cất. Dung dịch gốc có nồng độ
N- NO2 là 250 mg/l.
Chuẩn bị dung dịch chuẩn có nồng độ 1 mg/l bằng cách pha loãng dd gốc.
Lập các nồng độ khác nhau từ 0; 0,02; 0,04; 0,06; 0,08; 0,1 mg/l.
Lấy 10 ml mỗi nồng độ vào các ống nghiệm khác nhau.
Thêm 0,2 ml NH2C6H4SO2NH2.
Trộn đều và giữ yên trong 5 phút.
Thêm 0,2 ml C10H7NHCH2NH2.2HCl.
Trộn đều và giữ yên trong 10 phút.
So màu ở bước sóng 540 nm.
Với các dung dịch mẫu làm tương tự như lập đường chuẩn.

Luận văn thạc sĩ
Ngô Thị Kim Toán 31 K19 – Sinh học thực nghiệm
2.3.8. Phương pháp đánh giá khả năng xử lý photpho
Khả năng tích lũy photpho của các chủng được đánh giá thông qua sự giảm
nồng độ của PO4- trong môi trường nuôi cấy theo thời gian.
Chuẩn bị thuốc thử:
Chuẩn bị dung dịch (NH4)6Mo6
Hòa tan 7,27 g amonium molydat (NH4)6Mo7O2.4H2O (sấy ở 105oC trong 3
giờ) vào 150 ml nước cất.
Hòa tan 0,0945 g Kaly antimon tactrat (C8H8O12K2.Sb2O2.H2O) trong 50ml
nước cất.
96 ml H2SO4 đặc vào bình chứa 600 ml nước cất sau đó làm lạnh đến nhiệt
độ phòng.
Trộn các dung dịch trên với nhau
Thêm vào 10 g amonium amidosulfat (NH4OSO2NH2) vào hỗn hợp dung
dịch trên
Định mức đến 1000 ml
Chuẩn bị dung dịch C6H8O6: Hòa tan 1.8 g C6H8O6 trong 25 ml nước, sử dụng
trong vòng 2 tuần.
Lập đường chuẩn
Chuẩn bị dãy các dung dịch gồm 5 điểm vào 5 ống nghiệm có chia vạch
Dung dịch gốc 100 mg/l: hòa tan 0,4394g KH2PO4 trong 100 ml nước cất.
Dung dịch chuẩn: pha loãng dung dịch gốc 20 lần ta được dung dịch chuẩn 5
mg/l. Lấy dung dịch chuẩn theo tỷ lệ nồng độ sau:
Nồng độ (mg/l) 0 0,2 0,4 0,6 0,8 1
Vdd chuẩn (ml) 0 2 4 6 8 10
Sau khi tiến hành lấy dung dịch chuẩn và lấy mẫu vào ống nghiệm định mức
tới 40 ml
Thêm 1 ml dung dịch C6H8O6
Thêm 5 ml dung dịch (NH4)6Mo6
Định mức tới 50 ml

Luận văn thạc sĩ
Ngô Thị Kim Toán 32 K19 – Sinh học thực nghiệm
Điều nhiệt trong khoảng từ 30 đến 40oC trong thời gian từ 10 đến 20 phút
Đo mẫu tại bước sóng 710 nm
2.3.8.1. Phương pháp phân tích photpho tổng
Nguyên tắc: Amonium molydat và kali antimon tatrat phản ứng với PO43-
trong môi trường axit trung bình tạo thành axit photpho molipdic có màu xanh đậm
đo màu ở bước sóng 710 nm trên máy quang phổ UV – VIS [9].
Xử lý mẫu:
Lấy 10 ml mẫu vào bình có nút 100 ml
Thêm 5 ml K2S2O8 (4 g K2S2O8 trong 100 ml nước)
Thêm 1 ml H2SO4 30%
Đun ở 120oC trong 30 phút
Sau đó chuyển toàn bộ mẫu đã đun vào trong bình định mức tới 50 ml bằng
nước cất. Lấy 10 ml mẫu này cho vào bình định mức 50 ml
Thêm 5 ml dung dịch (NH4)6Mo6 vào ống và chuẩn về 40 ml
Thêm 1 ml dung dịch C6H8O6
Định mức tới 50 ml bằng nước cất.
Điều nhiệt trong khoảng từ 30 đến 40oC trong thời gian từ 10 đến 20 phút
Đo mẫu tại bước sóng 710 nm
Mẫu đối chứng được làm tương tự để đối chiếu lại kết quả.
Tính toán kết quả
C= Cđo 50/10 50/10 (mg/l)
Trong đó: C: nồng độ dung dịch mẫu thực (mg/l)
Cđo: nồng độ dung dịch mẫu đo từ ống nghiệm (mg/l).
2.3.8.2. Phương pháp phân tích hàm lượng Ortho photphate (PO43-)
Lấy 10 ml mẫu cho vào bình định mức 50 ml
Thêm 5 ml dung dịch (NH4)6Mo6 vào ống và chuẩn về 40 ml
Thêm 1 ml dung dịch C6H8O6
Định mức tới 50 ml bằng nước cất.
Điều nhiệt trong khoảng từ 30 đến 40oC trong thời gian từ 10 đến 20 phút

Luận văn thạc sĩ
Ngô Thị Kim Toán 33 K19 – Sinh học thực nghiệm
Đo tại bước sóng 710 nm
Mẫu đối chứng được làm tương tự để đối chiếu lại kết quả.
C= Cđo 50/10 (mg/l)
Trong đó: C: nồng độ dung dịch mẫu thực (mg/l)
Cđo: nồng độ dung dịch mẫu đo từ ống nghiệm (mg/l).
2.3.9. Phương pháp phân loại vi sinh vật dựa trên gen 16S rRNA
Các chủng vi sinh vật có khả năng xử lý nitơ và photpho được phân tích
trình tự gen 16S rRNA để xác định cây phân loại.
Trình tự gen đoạn 16S rRNA được đọc trên máy đọc trình tự tự động ABI
PRISM 3100 Avant (Hoa Kỳ) tại Viện Vi sinh vật và Công nghệ sinh học.
Kết quả phân tích trình tự 16S rRNA được so sánh với trình tự đoạn gen
tương tưng của các loài trong ngân hàng gen quốc tế bằng phương pháp Clustal X.
Cây phát sinh chủng loại được xây dựng dựa trên phần mềm njplot.
2.3.10. Phương pháp thống kê sinh học
Các kết quả thu được được xử lý bằng thông kê sinh học thông qua các phần
mềm Excel, Microsoft Word để đảm bảo độ tin cậy.

Luận văn thạc sĩ
Ngô Thị Kim Toán 34 K19 – Sinh học thực nghiệm
CHƯƠNG 3: KẾT QUẢ VÀ THẢO LUẬN
3. 1. Nghiên cứu các chủng vi sinh vật có khả năng chuyển hóa nitơ.
3.1.1. Phân lập chủng vi sinh vật có khả năng chuyển hóa nitơ và có khả năng
hình thành màng sinh học
3.1.1.1. Phân lập các chủng vi sinh vật có khả năng chuyển hóa nitơ
Các mẫu sau khi đưa về phòng thí nghiệm, chúng tôi tiến hành phân tích hàm
lượng nitơ và photpho ban đầu có trong các mẫu. Kết quả thu được như sau:
Bảng 3.1. Hàm lượng thành phần nitơ và photpho trong mẫu phân tích.
Mẫu
Hàm lượng các hợp chất nitơ trong
mẫu phân lập (mg/l)
Hàm lượng
photpho tổng
(mg/l) Nitơ tổng NH4+ NO2
-
1 140,74 ± 2,1 71,58 ± 1,43 5,31 ± 0,032 16,97 ± 1,02
2 111,93 ± 1,68 78,42 ± 2,35 1,44 ± 0,009 18,76 ± 1,13
3 83,12 ± 1,25 68,26 ± 1,36 1,01 ± 0,006 46,05 ± 2,76
4 71,60 ± 0,71 65,14 ± 1,30 2,59 ± 0,016 10,68 ± 0,64
5 100,41 ±1,50 75,10 ± 1,50 7,33 ± 0,044 31,33 ± 1,88
Kết quả phân tích mẫu ban đầu cho thấy, các mẫu ban đầu đều có hàm lượng
nitơ và photpho cao hơn nhiều so với mức trung bình và quy định cho phép đối với
nước thải (Bảng 3.1).
Từ những mẫu thu được, chúng tôi đã phân lập được 65 chủng vi sinh vật có
khả năng phát triển trên môi trường Winogradsky. Trong đó, 41 chủng có khả năng
phát triển trên môi trường Winogradsky 1, 24 chủng có khả năng phát triển trên môi
trường Winogradsky 2. Kết quả phân lập ở bảng 3.2 cho thấy trên hai môi trường
Winogradsky phân lập được các chủng từ nước thải bể biogas nhiều hơn so với các
mẫu lấy ở khu tập trung rác thải. Số lượng các chủng thu được ở tầng giữa bể
biogas đang sử dụng và đã không sử dụng nhiều hơn ở tầng đáy bể biogas đã ngưng
sử dụng. Kết quả này cũng phù hợp với nhiều nghiên cứu cho rằng các chủng vi
khuẩn có khả năng chuyển hóa NH4+ và NO2
- là những vi khuẩn hiếu khí [69], [71].

Luận văn thạc sĩ
Ngô Thị Kim Toán 35 K19 – Sinh học thực nghiệm
Bảng 3.2. Địa điểm và số lượng các chủng vi sinh vật có khả năng chuyển hóa nitơ
Địa điểm lấy mẫu
Số chủng phân lập
Ký hiệu chủng Môi trường
W1
Môi trường
W2
Nước thải
bể biogas
Tầng giữa bể biogas
đã ngưng sử dụng 11 15
Từ B11.1 đến B11.11
Từ B21.1 đến B21.15
Tầng đáy bể biogas đã
ngưng sử dụng 4 0 Từ B12.1 đến B12.4
Tầng giữa bể biogas
đang sử dụng 13 9
Từ B13.1 đến B13.13
Từ B23.1 đến B23.9
Nước thải
khu tập
trung rác
thải
Ngoài bãi rác 5 0 VP1.1, VP1.4, VP1.5,
VP1.8, VP1.9,
Trong bãi rác 8 0
VP2.2, VP2.3, VP2.4,
VP2.5, VP2.7, VP2.9,
VP2.10, VP2.11
Các chủng phân lập trên môi trường Winogradsky 1 (Hình 3.1A) có đặc
điểm chủ yếu là khuẩn lạc tế bào tròn hay có hình tia, có màu trắng đục hay vàng.
Các chủng phân lập trên môi trường Winogradsky 2 khuẩn lạc chủ yếu giống trên
môi trường Winogradsky 1, có hình tia hay tròn, màu trắng đục (Hình 3.1B).
Hình 3.1. Hình ảnh các khuẩn lạc phân lập trên môi trường Winogradsky 1(A) và
Winogradsky 2 (B)

Luận văn thạc sĩ
Ngô Thị Kim Toán 36 K19 – Sinh học thực nghiệm
3.1.1.2. Khả năng hình thành màng sinh học của các chủng phân lập
Bằng phương pháp nhuộm với tím kết tinh bước đầu đã chúng tôi đánh giá
được khả năng hình thành màng sinh học của 28 chủng vi khuẩn phân lập trên môi
trường Winograky 1 từ mẫu nước thải ở bể biogas. Kết quả cho thấy hầu hết các vi
khuẩn phân lập được đều có khả năng hình thành biofilm, trong đó các chủng
B11.8, B11.11 và B13.1 có khả năng hình thành màng sinh học tốt nhất với số OD
đo ở bước sóng 570 nm lần lượt tương ứng là 1,824; 2,775 và 2,235 (Hình 3.2).
Hình 3.2. Khả năng hình thành màng sinh học của các chủng trên môi trường
Winogradsky 1 được phân lập từ các mẫu nước thải thu từ bể biogas
Khi tiến hành đánh giá 13 mẫu thu được ở khu tập trung rác thải Vạn Phúc
bằng phương pháp tương tự, chúng tôi thu được hai chủng VP1.1 và VP2.11 là hai
chủng có chỉ số OD đo ở bước sóng 570 nm cao hơn các chủng còn lại với số đọc
tưng ứng là 4,65 và 3,24 (Hình 3.3).
Từ những kết quả thu được như trên, chúng tôi đã tuyển chọn được 5 chủng
có khả năng hình thành màng sinh học tốt nhất và có khả năng phát triển trên môi
trường Winogradsky 1 là : VP1.1, VP2.11, B11.8, B11.11, B13.1.

Luận văn thạc sĩ
Ngô Thị Kim Toán 37 K19 – Sinh học thực nghiệm
Hình 3.3. Khả năng hình thành màng sinh học của các chủng trên môi trường
Winogradsky 1 được phân lập từ các mẫu nước thải khu tập trung rác thải
Hình 3.4. Khả năng tạo hình thành màng sinh học của các chủng trên môi trường
Winogradsky 2 được phân lập ở bể biogas
Đối với các chủng phân lập được trên môi trường Winogradsky 2, chúng tôi
cũng tiến hành các thí nghiệm tương tự để đánh giá khả năng hình thành biofilm.
Kết quả cho thấy, trong 24 chủng thu được, 3 chủng B21.1, B21.10 và B23.2 có khả

Luận văn thạc sĩ
Ngô Thị Kim Toán 38 K19 – Sinh học thực nghiệm
năng tạo màng biofilm tốt nhất. Chúng tôi lựa chọn 3 chủng này cho các nghiên cứu
tiếp theo để phân tích khả năng chuyển hóa NO2- (Hình 3.4).
3.1.2. Khả năng chuyển hóa nitơ
8 chủng vi sinh vật có khả năng hình thành màng sinh học tốt nhất được
chúng tôi chọn để thử hoạt tính chuyển hóa các hợp chất nitơ. Trong đó 5 chủng
B11.8, B11.11, B13.1, VP1.1 và VP2.11 được thử khả năng chuyển hóa amoni, 3
chủng B21.1, B21.10 và B23.2 được thử khả năng chuyển hóa nitrite.
3.1.2.1. Khả năng chuyển hóa amoni
Khả năng chuyển hóa amoni được đánh giá thông qua hàm lượng NH4+ trong
mẫu nghiên cứu theo thời gian. Chủng vi sinh vật nào có khả năng chuyển hóa
amoni thì hàm lượng NH4+ sẽ giảm đi trong quá trình nghiên cứu. 5 chủng B11.8,
B11.11, B13.1, VP1.1 và VP2.11 được nuôi lắc trong môi trường Winogradsky 1,
sau mỗi khoảng thời gian nghiên cứu, lấy mẫu phân tích.
Hình 3.5. Khả năng chuyển hóa amoni của các chủng nghiên cứu.
Với hàm lượng amoni trong mẫu ban đẫu là 450 mg/l. Kết quả (Hình 3.5)
cho thấy, sau 5 ngày, bốn chủng B11.8, B13.1, VP1.1 và VP2.11 nồng độ NH4+
giảm đi không nhiều, trong khi đó, với chủng B11.11, nồng độ NH4+ giảm 58,93%
so với mẫu ban đầu. Nồng độ NH4+ tiếp tục giảm đi, sau 10, 15 ngày, hàm lượng
trong mẫu của chủng B11.8 giảm đi 12,2%, B13.1 giảm đi 28,53%, VP1.1 giảm đi

Luận văn thạc sĩ
Ngô Thị Kim Toán 39 K19 – Sinh học thực nghiệm
14,44% và VP2.11 giảm đi 10,52%. Sau 20 ngày, chủng B11.11 hàm lượng NH4+
có trong mẫu đã giảm đi 85,21%. Trong khi đó, 4 chủng còn lại hàm lượng NH4+
trong dung dịch chỉ giảm đi vào khoảng từ 20-50% so với mẫu ban đầu. Từ kết quả
trên cho thấy chủng B11.11 có khả năng chuyển hóa amoni tốt nhất. Do vậy, chúng
tôi đã lựa chọn chủng B11.11 cho các nghiên cứu tiếp theo của mình.
3.1.2.2. Khả năng chuyển hóa nitrite
Với các chủng vi sinh vật phân lập trên môi trường Winogradsky 2, chúng tôi
cũng tiến hành các thí nghiệm tương tự. Khả năng chuyển hóa nitrite được đánh giá
thông qua hàm lượng NO2- giảm theo thời gian nghiên cứu. 3 chủng B21.1, B21.10,
B23.2 được nuôi lắc trong môi trường Winogradsky 2.
Hình 3.6. Khả năng chuyển hóa nitrite của các chủng nghiên cứu
Với hàm lượng nitrite trong mẫu ban đầu là 80 mg/l. Kết quả hình 3.6 cho
thấy cả ba chủng nghiên cứu đều có khả năng chuyển hóa nitrite, sau 5 ngày, nồng
độ NO2- trong dịch nuôi lắc của chủng B21.10 giảm đi 75.23% so với mẫu ban đầu,
tương ứng, hai chủng B23.2 và B21.1 là 69.07% và 7.26%. Chủng B21.1 có khả
năng chuyển hóa NO2- nhưng khả năng chuyển hóa chậm hơn, sau 20 ngày hàm
lượng NO2- trong dung dịch giảm đi 75% so với mẫu ban đầu trong khi hai chủng
B21.10 và B23.2 đã chuyển hóa gần như hoàn toàn lượng NO2- có trong mẫu lắc.

Luận văn thạc sĩ
Ngô Thị Kim Toán 40 K19 – Sinh học thực nghiệm
Kết quả, sau 20 ngày hai chủng B21.10 và B23.2 đã chuyển hóa được trên 97%
lượng NO2- có trong mẫu. Từ kết quả này, chúng tôi đã lựa chọn ra hai chủng
B21.10 và B23.2 cho các thí nghiệm nghiên cứu tiếp theo.
3. 2. Nghiên cứu các chủng vi sinh vật có khả năng xử lý photpho.
3.2.1. Phân lập chủng vi sinh vật có khả năng xử lý photpho và có khả năng
hình thành màng sinh học.
3.2.1.1. Phân lập các chủng vi sinh vật có khả năng xử lý photpho
Mẫu nước thải từ bể biogas và nước thải khu xử lý rác thải khi phân lập trên
môi trường AMM thu được các khuẩn lạc chủ yếu màu trắng đục, hơi vàng, khuẩn
lạc tròn (Hình 3.7). Dựa vào kích thước, màu sắc và hình dạng, chúng tôi đã thu
được 21 chủng với ký hiệu như bảng 3.3.
Hình 3.7. Một số khuẩn lạc phân lập trên môi trường AMM: mẫu khu vực biogas
(A) và khu tập trung rác thải (B)
Kết quả phân lập ở bảng 3.3 cho thấy, câc chủng phân lập ở tầng giữa bể
biogas cao hơn so với tầng đáy, trong khu vực tập trung rác thải cao hơn so với bên
ngoài khu vực.

Luận văn thạc sĩ
Ngô Thị Kim Toán 41 K19 – Sinh học thực nghiệm
Bảng 3.3. Địa điểm và số lượng các chủng vi sinh vật có khả năng tích lũy photpho
Địa điểm thu mẫu Mẫu thu được Ký hiệu
Nước thải từ bể biogas 13 chủng
Từ A1.1 dến A1.8
Từ A2.1 đến A2.2
Từ A3.1 đến A3.3
Nước thải từ khu tập trung
rác thải 8 chủng
Từ A4.1 đến A4.3
Từ A5.1 đến A5.5
3.2.1.2. Khả năng hình thành màng sinh học của các chủng phân lập
Tiến hành các thí nghiệm tương tự về xác định khả năng hình thành màng
sinh học của các chủng phân lập từ môi trường AMM. Kết quả trên hình cho thấy,
chủng A4.2, A5.1, A5.2, A5.3 là bốn chủng có khả năng hình thành màng sinh học
tốt nhất (Hình 3.8). Do vậy chúng tôi đã lựa chọn 4 chủng này cho các nghiên cứu
tiếp theo.
Hình 3.8. Khả năng hình thàng màng sinh học của các chủng vi sinh vật có khả
năng tích lũy photpho

Luận văn thạc sĩ
Ngô Thị Kim Toán 42 K19 – Sinh học thực nghiệm
3.2.2. Khả năng xử lý photpho của các chủng nghiên cứu.
Đánh giá khả năng giảm hàm lượng photpho, chúng tôi đánh giá thông qua
hàm lượng photphate tích lũy trong vi sinh vật. Chúng tôi tiến hành nuôi cấy lắc các
chủng nghiên cứu trong môi trường AMM trong thời gian 10 ngày ở các điều kiện
tối ưu về pH và nhiệt độ. Để đánh giá khả năng tích lũy photpho của vi sinh vật,
chúng tôi phân tích, đánh giá thông qua hàm lượng photphate còn lại trong môi
trường.
Hình 3.9. Khả năng xử lý photpho của các chủng nghiên cứu trong môi trường với
hàm lượng photpho 6mg/l
Kết quả hình 3.9 cho thấy, với hàm lượng photpho trong dung dịch AMM
ban đầu là 6mg/l, các chủng vi sinh vật đều có khả năng tích lũy photpho, sau 10
ngày, lượng photpho trong dung dịch nuôi lắc gần như không còn được phát hiện.
Các chủng A5.1, A5.2 và A5.3 sau 5 ngày đã không còn phát hiện thấy photphate có
trong dung dịch nuôi cấy, tương ứng với chủng A4.2 là 7 ngày.
Từ kết quả như vậy, chúng tôi đã tiến hành tăng hàm lượng photphate trong
môi trường nuôi cấy lên 18mg/l (Hình 3.10). Kết quả phân tích cho thấy, sau 5
ngày, hàm lượng photphate trong dịch nuôi của 2 chủng A4.2 và A5.1 giảm đi cao
nhất trong khoảng 25%. Hàm lượng photphate tiếp tục giảm sau 5, 7 và 10 ngày.

Luận văn thạc sĩ
Ngô Thị Kim Toán 43 K19 – Sinh học thực nghiệm
Sau 10 ngày, chủng A4.2 có hàm lượng photphate giảm đi lớn nhất, lượng
photphate giảm đi là 39,32% so với dịch mẫu ban đầu. Các chủng A5.1, A5.2 và
A5.3 hàm lượng photphate giảm đi trong dịch mẫu lần lượt là 34,88%, 21,78% và
17,95%. Từ các kết quả này, chúng tôi đã lựa chọn chủng A4.2 cho các nghiên cứu
tiếp theo. Đây là những kết quả bước đầu cho nghiên cứu, ứng dụng màng biofilm
vào xử lý nước thải giàu hợp chất nitơ, photpho từ các chủng vi sinh vật phân lập tại
Việt Nam.
Hình 3.10. Khả năng xử lý photpho của các chủng nghiên cứu trong môi trường
hàm lượng photpho18mg/l
3. 3. Các đặc điểm hình thái, sinh lý, sinh hóa của các chủng nghiên cứu
3.3.1. Khả năng tạo hình thành màng sinh học trên một số giá thể
Bốn chủng có khả năng chuyển hóa hợp chất chứa nitơ và photpho tốt nhất là
B11.11, B21.10, B23.2 và A4.2 được chúng tôi xác định khả năng tạo biofilm trên
một số giá thể, chất mang.
Để xác định khả năng hình thành màng sinh học trên giá thể nhựa (ống
eppendorf) chúng tôi dựa vào phương pháp nhuộm tím kết tinh.
Kết quả nhuộm tím kết tinh trên ống eppendorf cho thấy cả 4 chủng đều tạo
biofilm tốt trên giá thể nhựa, trong đó chủng B11.11 có khả năng tạo biofilm tốt
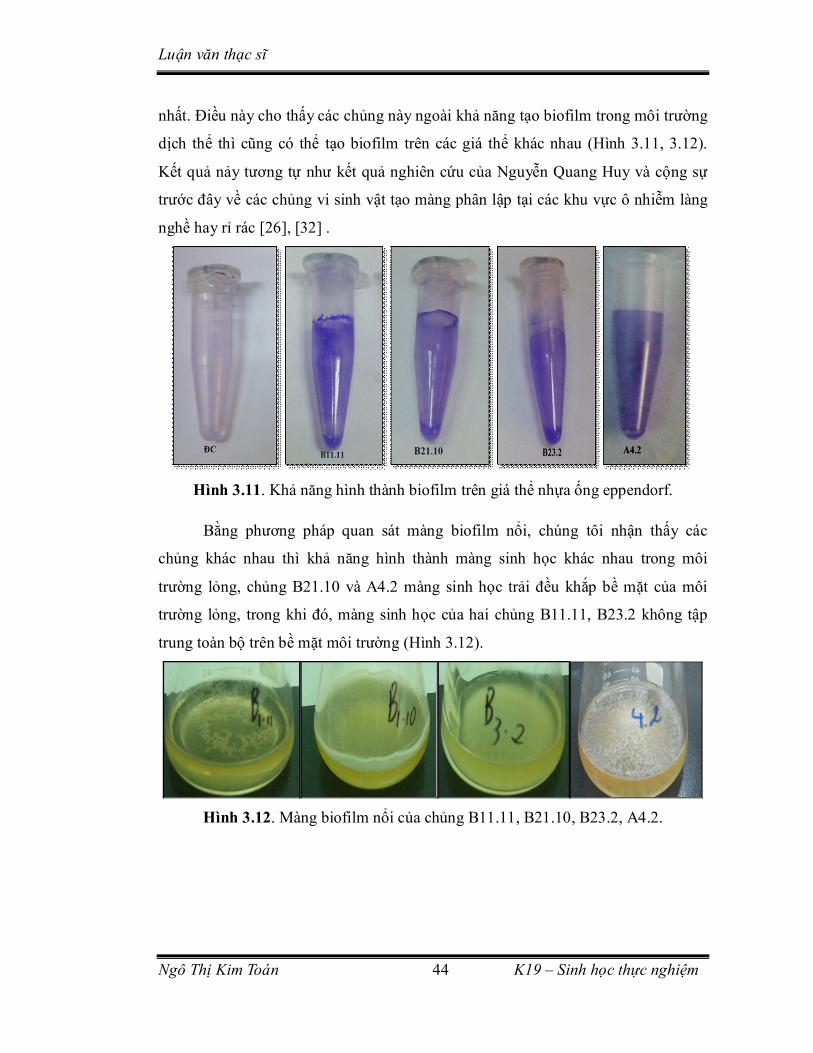
Luận văn thạc sĩ
Ngô Thị Kim Toán 44 K19 – Sinh học thực nghiệm
nhất. Điều này cho thấy các chủng này ngoài khả năng tạo biofilm trong môi trường
dịch thể thì cũng có thể tạo biofilm trên các giá thể khác nhau (Hình 3.11, 3.12).
Kết quả nảy tương tự như kết quả nghiên cứu của Nguyễn Quang Huy và cộng sự
trước đây về các chủng vi sinh vật tạo màng phân lập tại các khu vực ô nhiễm làng
nghề hay rỉ rác [26], [32] .
Hình 3.11. Khả năng hình thành biofilm trên giá thể nhựa ống eppendorf.
Bằng phương pháp quan sát màng biofilm nổi, chúng tôi nhận thấy các
chủng khác nhau thì khả năng hình thành màng sinh học khác nhau trong môi
trường lỏng, chủng B21.10 và A4.2 màng sinh học trải đều khắp bề mặt của môi
trường lỏng, trong khi đó, màng sinh học của hai chủng B11.11, B23.2 không tập
trung toàn bộ trên bề mặt môi trường (Hình 3.12).
Hình 3.12. Màng biofilm nổi của chủng B11.11, B21.10, B23.2, A4.2.

Luận văn thạc sĩ
Ngô Thị Kim Toán 45 K19 – Sinh học thực nghiệm
3.3.2. Đặc điểm hình thái của các chủng nghiên cứu
Để bước đầu nghiên cứu, phân loại các chủng có khả năng xử lý nước thải
giàu nitơ, photpho, chúng tôi tiến hành nhuộm Gram và quan sát dưới kính hiển vi
quang học có độ phóng đại 1000 lần.
Hình 3.13. Ảnh nhuộm Gram các chủng nghiên cứu ở độ phóng đại 1000 lần.
Quan sát dưới kính hiển vi quang học ở độ phóng đại 1000 lần, chúng tôi
thấy, các chủng nghiên cứu đều có hình que, xếp thành chuỗi (Hình 3.13).
Chúng tôi cũng tiến hành quan sát hình thái màng sinh học và tế bào vi
khuẩn qua kính hiển vi điện tử quét JSM – 5421LV ở độ phóng đại 10000 lần.
Hình 3.14. Cấu trúc hiển vi màng biofilm của chủng nghiên cứu ở độ phóng đại
10000 lần
Khi quan sát dưới kính hiển vi điện tử quét ở độ phóng đại 10000 lần, nhận
thấy các tế bào có hình dạng khác biệt nhau rất rõ và màng sinh học tạo thành cũng
có sự khác biệt. Chủng B11.11 có màng sinh học dày, lớp EPS ngoại bào bao xung
quanh tế bào, khi thu nhận thì nhận thấy lớp màng khá nhầy, rất dai, lớp màng
không đồng đều trên bề mặt, màu vàng nhạt. Chủng B21.10 lớp màng sinh học

Luận văn thạc sĩ
Ngô Thị Kim Toán 46 K19 – Sinh học thực nghiệm
tương đối đồng nhất, không gian ba chiều phức tạp, nhiều lớp cuộn vào nhau, khi
thu nhận thì biofilm dễ bị cắt nhỏ hơn của chủng B11.11, có màu trắng đục. Chủng
B23.2, màng sinh học hình thành lớp rất mỏng, trong suốt, sần sùi. Chủng A4.2 các
tế bào xếp sít nhau, hình trực khuẩn, khi thu nhận biofilm dễ bị phân mảnh, đứt gẫy,
màu trắng đục (Hình 3.14).
3.3.3. Ảnh hưởng của một số yếu tố môi trường lên sự hình thành màng sinh
học
Quá trình hình thành màng sinh học trải qua nhiều giai đoạn và chịu ảnh
hưởng của nhiều yếu tố môi trường. Trong đó nhiệt độ và pH là hai yếu tố ảnh
hưởng đến sự phát triển của vi sinh vật lớn nhất.
Nhiệt độ là một trong những yếu tố quan trọng trong sự phát triển của vi sinh
vật, sự hình thành màng sinh học, yếu tố nhiệt độ đóng một vai trò không thể thiếu
trong sự hình thành màng. Khi tiến hành nghiên cứu màng sinh học ở các điều kiện
nhiệt độ khác nhau của vi khuẩn Salmonella enteritidis, Giaouris và cộng sự đã chỉ
ra khả năng hình thành màng sinh học tốt nhất của vi khuẩn này khá thấp, chỉ là
20oC [24]. Trong nghiên cứu của Di Bonaventura cùng cộng sự, vi khuẩn
Stenotrophomonas maltophilia có khả năng tạo màng sinh học tốt nhất ở 32oC [20].
Những nghiên cứu này cho thấy mỗi chủng vi sinh vật có khả năng tạo màng sinh
học tốt nhất ở các nhiệt độ khác nhau.
Hình 3.15. Ảnh hưởng của nhiệt độ lên khả năng hình thành màng sinh học

Luận văn thạc sĩ
Ngô Thị Kim Toán 47 K19 – Sinh học thực nghiệm
Kết quả thử khả năng hình thành màng sinh học ở các điều kiện nhiệt độ
khác nhau (Hình 3.15) cho thấy, hai chủng B11.11 và B21.10 có khả năng hình
thành màng sinh học tốt nhất ở điều kiện nhiệt độ là 37oC, chủng B23.2 có khả năng
hình thành màng sinh học tốt trong dải nhiệt độ khá rộng từ 37 đến 50oC, điều kiện
nhiệt độ tốt nhất cho sự hình thành màng sinh học của chủng A4.2 là 50oC.
Khi nghiên cứu các điều kiện cho vi sinh vật phát triển thì một trong những
yếu tố được quan tâm là giá trị pH. Ở các pH khác nhau, mức độ sinh trưởng của vi
sinh vật cũng rất khác nhau và điều này có thể ảnh hưởng đến sự phát triển của vi
sinh vật và hình thành màng sinh học của bản thân chúng. Vi sinh vật chỉ có thể
phát triển trongmột khoảng pH nhất định, khi pH môi trường nằm ngoài khoảng cho
phép, vi sinh vật sẽ phát triển yếu đi hoặc có thể bị ức chế hoàn toàn. Các chủng vi
sinh vật khác nhau có khoảng pH phát triển khác nhau do đó ảnh hưởng tới khả
năng tạo màng sinh học.
Hình 3.16. Ảnh hưởng của pH môi trường lên khả năng hình thành màng sinh học
Kết quả thử khả năng tạo màng của các chủng nghiên cứu cho thấy, hầu hết
các chủng đều có khả năng tạo màng sinh học tốt ở điều kiện pH kiềm khoảng 7,5,
chủng A4.2 có khả năng tạo màng tốt ở pH 7, chủng B21.10 có khả năng tạo màng
sinh học tốt ở pH 8 (Hình 3.16). Theo nghiên cứu của Wulff và cộng sự [65] đối với
sự phát triển và hình thành màng sinh học của vi khuẩn Xylella fastidiosa nhận thấy

Luận văn thạc sĩ
Ngô Thị Kim Toán 48 K19 – Sinh học thực nghiệm
rằng sự hình thành màng sinh học chịu ảnh hưởng lớn của giá trị pH, với vi khuẩn
này, khả năng hình thành màng sinh học tốt nhất ở pH 6.8, và bị ức chế hình thành
màng ở pH dưới 6.6. Kết quả này phù hợp với nghiên cứu, các chủng phân lập từ
nước thải bể biogas và khu tập trung xử lý rác thải thường phát triển tốt ở pH trong
khoảng từ 7 đến 8.5 [10].
3.3.4. Khả năng chuyển hóa một số chất trong bộ kit APi
Để nghiên cứu một số tính chất về mặt sinh hóa, chúng tôi đã sử dụng kit thử
APi. Các chất trong bộ kit dùng để kiểm tra khả năng sử dụng các vi sinh vật là các
nguồn cacbon khác nhau (D-glucose, L-arabinose, D- mannose, D- mannitol, N-
acetyl- glucosamine…), urea, L-tryptophan, L-arginine, gelatine…Kết quả được
trình bày ở bảng 3.4:
Bảng 3.4. Một số đặc điểm sinh hoá của các chủng theo kit APi (BioMérieus)
Ký hiệu chất
kiểm tra Các chất chuyển hóa
Các chủng vi sinh vật nghiên cứu
B11.11 B21.10 B23.2 A4.2
NO3 KNO3 + + + +
TRP L- tryptophan - - - -
ADH L- arginine + - + +
URE Urea - - + -
ESC Esculin ferric citrate + + + +
GEL Gelatine + + - +
PNPG 4-nitrophenyl β D –
galactopyranoside + + + +
GLU D- glucose + + + +
ARA L- arabinose - + - +
MNE D- mannose + + + +

Luận văn thạc sĩ
Ngô Thị Kim Toán 49 K19 – Sinh học thực nghiệm
Chú thích (-): phản ứng âm tính, (+) phản ứng dương tính
Với kí hiệu dấu (-) biểu thị cho phản ứng âm tính, dấu (+) biểu thị cho phản
ứng dương tính với sự chuyển hóa các hợp chất hữu cơ. Kết quả cho thấy, các
chủng B11.11, B21.1, B23.2 và A4.2 có thể sử dụng nhiều cơ chất khác nhau, đặc
biệt chủng A4.2 và B23.2 có khả năng sử dụng hầu hết các loại đường, chủng
B11.11 không có khả năng sử dụng L – arabinose, chủng B21.10 không sử dụng
được L – arabinose, N – acetyl – glucosamine và D – mantose (Bảng 3.4). Khi tiến
hành phân tích trên phần mềm của BioMérieus, chủng B23.2 có khả năng thuộc chi
Pseudomonas.
Với khả năng sử dụng các loại đường khác nhau, các chủng vi sinh vật này
có khả năng sử dụng nhiều nguồn cacbon thay thế khác nhau, việc thay thế nguồn
cacbon từ vật liệu có chi phí và giá thành cao sang có chi phí phù hợp sẽ giúp cho
quá trình đưa sản phẩm vào ứng dụng rộng rãi hơn. Các chủng nghiên cứu xử lý
nước thải có chứa nitơ đều có khả năng khử NO3- cho thấy phù hợp với nghiên cứu,
chủng A4.2 là chủng có khả năng xử lý photpho, tuy nhiên, kết quả trên bảng cho
thấy, chủng này cũng có khả năng khử NO3-, theo nghiên cứu của Jorgensen và
Pauli cho rằng các chủng có khả năng tích lũy photphate cũng có khả năng chuyển
hóa nitơ [33].
MAN D- mannitol + + + +
NAG N- acetyl- glucosamine + + - +
MAL D- mantose + + - +
GNT Potassium gluconate - - + +
CAP Axit capric - - + -
ADI Axit Adipic - - - -
MLT Axit Malic + - + +
CIT Trisodium Citrate - - + +
PAC Axit phenyl acetic - + - -

Luận văn thạc sĩ
Ngô Thị Kim Toán 50 K19 – Sinh học thực nghiệm
3.3.5. Trình tự 16S rRNA và cây phát sinh chủng loại
Chúng tôi tiếp tục tiến hành giải trình tự gen mã hóa 16S rARN. Kết quả giải
trình tự gen 16S rARN được so sánh với các trình tự gen 16S rARN trong ngân
hàng gen quốc tế dựa vào phần mềm Clustal X. Trên cơ sở đó, chúng tôi xây dựng
cây chủng loại phát sinh dựa vào phần mềm njplot
Trình tự gen mã hóa 16S rARN của chủng B11.11 tương đồng 99,9 %
(1399/1400 bp) với đoạn gen 16S rARN của vi khuẩn Bacillus
licheniformis_X68416; tương đồng 99,6 % (1394/1400 bp) với đoạn gen 16S rARN
của vi khuẩn Bacillus sonorensis_AF302118; tương đồng 99,4 % (1391/1400 bp)
với đoạn 16S rARN của vi khuẩn Bacillus aerius_AJ831843 (Hình 3.17). Theo
nghiên cứu của Kim và cộng sự, cùng với hai chủng vi khuẩn Bacillus subtilis và
Bacillus cereus, Bacillus licheniformis là một trong các chủng thuộc chi Bacillus có
khả năng nitrate hóa và khử nitrate trong điều kiện hiếu khí, và lượng amoni đã
chuyển sang khí N2 là 33% sau 4 giờ nuôi cấy [34].
Hình 3.17. Sơ đồ cây phát sinh chủng loại của chủng B11.11
Staphylococcus aureus_X68417 Bacillus aerophilus_AJ831844 Bacillus stratosphericus_AJ831841 Bacillus altitudinis_AJ831842
99
Bacillus pumilus_AY876289 Bacillus safensis_AF234854
99
100Bacillus aerius_AJ831843
Bacillus licheniformis_X68416 B11.11
100
Bacillus sonorensis_AF302118
63
Bacillus atrophaeus_AB021181 Bacillus siamensis_GQ281299
Bacillus amyloliquefaciens_AB255669 Bacillus methylotrophicus_EU194897
57
Bacillus vallismortis_AB021198 Bacillus mojavensis_AB021191 Bacillus subtilis_AB042061
Bacillus subtilis subsp spizizenii_AF
54
78
88
52
81
100
97
100
99
0.01

Luận văn thạc sĩ
Ngô Thị Kim Toán 51 K19 – Sinh học thực nghiệm
Hình 3.18. Sơ đồ cây phát sinh chủng loại của chủng B21.10
Kết quả giải trình tự gen 16S rARN của chủng B21.10 tương đồng 99,8%
(1397/1400 bp) với đoạn gen 16S rARN của vi khuẩn Bacillus
amyloliquefaciens_AB255669 và Bacillus methylotrophicus_EU194897; tương
đồng 99,6% (1395/1400 bp) với đoạn gen 16S rARN của vi khuẩn Bacillus
vallismortis_AB021198; tương đồng 99,6% (1394/1400 bp) với đoạn gen 16S
rARN của vi khuẩn Bacillus siamensis_GQ281299 (Hình 3.18). Trong nghiên cứu
của Zhang cùng cộng sự, Bacillus methylotrophicus có khả năng chuyển hóa cả
NH4+ và NO2
- thành khí N2O với hiệu suất tương ứng là 51,58 và 5,81 mg/l/ngày
[68].
Staphylococcus aureus_X68417 Bacillus aerophilus_AJ831844 Bacillus stratosphericus_AJ831841 Bacillus altitudinis_AJ831842
99
Bacillus pumilus_AY876289 Bacillus safensis_AF234854
99
100Bacillus aerius_AJ831843
Bacillus licheniformis_X68416 Bacillus sonorensis_AF302118
100
Bacillus atrophaeus_AB021181 Bacillus mojavensis_AB021191
Bacillus subtilis subsp spizizenii_AF074970 Bacillus subtilis_AB042061
77
Bacillus vallismortis_AB021198
Bacillus amyloliquefaciens_AB255669 Bacillus siamensis_GQ281299
Bacillus methylotrophicus_EU194897
B21.10
50
8089
79
6880
100
97
100
99
0.01

Luận văn thạc sĩ
Ngô Thị Kim Toán 52 K19 – Sinh học thực nghiệm
Hình 3.19. Sơ đồ cây phát sinh chủng loại của chủng B23.2
Kết quả giải rình tự gen 16S rARN của chủng B23.2 tương đồng 97,8%
(1370/1401 bp) với đoạn gen 16S rARN của vi khuẩn Pseudomonas
pseudoalcaligenes_Z76666; tương đồng 97,6% (1368/1401 bp) với đoạn gen 16S
rARN của vi khuẩn Pseudomonas mendocina_D84016 (Hình 3.19). Kết quả này
cho thấy, chủng B23.2 có thể thuộc chi Pseudomonas. Đây là kết quả được công bố
đầu tiên về vi khuẩn Pseudomonas pseudoalcaligenes có khả năng chuyển hóa
nitrite trong môi trường so với nghiên cứu của Zhang và cộng sự về khả năng
chuyển hóa của chủng Pseudomonas stutzeri có khả năng chuyển hóa nitrite [67].
Azorhizophilus paspali_AJ308318 Azotobacter beijerinckii_AJ308319
Azotobacter chroococcum_AB175653 Pseudomonas aeruginosa_X06684
Pseudomonas otitidis_AY953147 Pseudomonas resinovorans_Z76668
73
Serpens flexibilis_GU269546 Pseudomonas tuomuerensis_DQ868767
Pseudomonas stutzeri_AF094748 Pseudomonas xanthomarina_AB176954
Pseudomonas oleovorans subsp. lubricanti_DQ842018 Pseudomonas alcalophila_AB030583
Pseudomonas pseudoalcaligenes_Z76666
100
Pseudomonas mendocina_D84016
B23.2
66
76
71
50
100
57
92
68
93
89
0.01

Luận văn thạc sĩ
Ngô Thị Kim Toán 53 K19 – Sinh học thực nghiệm
.
Hình 3.20. Sơ đồ cây phát sinh chủng loại của chủng A4.2
Trình tự gen 16S rARN của chủng A4.2 tương đồng 99.9% (1411/1413 bp)
với đoạn gen 16S rARN của vi khuẩn Bacillus licheniformis_X68416 khi so sánh
với các trình tự gen 16S rARN của các vi sinh vật trong ngân hàng gen quốc tế
(Hình 3.20). Điều này cho chúng tôi kết luận: chủng A4.2 thuộc chi Bacillus. Đây là
công bố đầu tiên ở Việt Nam, chủng vi khuẩn Bacillus licheniformis có khả năng xử
lý photpho trong nước thải bằng việc tích lũy vào cơ thể vi sinh vật, một phần cung
cấp năng lượng cho các quá trình sinh hóa và trao đổi chất.
Staphylococcus aureus_X68417 Bacillus cibi_AY550276 Bacillus indicus_AJ583158
Bacillus idriensis_AY904033
100
Bacillus isabeliae_AM503357 100
Bacillus aerophilus_AJ831844 Bacillus altitudinis_AJ831842 Bacillus stratosphericus_AJ831841
99
Bacillus safensis_AF234854 Bacillus pumilus_AY876289
99
97
Bacillus aerius_AJ831843 Bacillus licheniformis_X68416 A4.2 100
Bacillus sonorensis_AF302118 100
Bacillus atrophaeus_AB021181 Bacillus malacitensis_AY603656
Bacillus axarquiensis_AY603657 Bacillus mojavensis_AB021191
59
Bacillus subtilis_AB042061
77
Bacillus vallismortis_AB021198 Bacillus nematotocita_AY820954
Bacillus velezensis_AY603658 Bacillus siamensis_GQ281299
Bacillus amyloliquefaciens_NR041455 Bacillus methylotrophicus_EU194897
6866
62
81
5971
100
100
100
99
68
89
0.01

Luận văn thạc sĩ
Ngô Thị Kim Toán 54 K19 – Sinh học thực nghiệm
KẾT LUẬN
1. Đã phân lập và được 65 chủng vi sinh vật trên môi trường Winogradsky và
21 chủng trên môi trường AMM từ các mẫu nước thải ở Thanh Hóa và Hà Nội.
2. Đã tuyển chọn được 4 chủng vi sinh vật có khả năng hình thành màng sinh
học và có khả năng xử lý nitơ và photpho tốt nhất là các chủng có ký hiệu B11.11,
B21.10, B23.2 và A4.2. Các chủng có khả năng hình thành màng sinh học ở nhiệt
độ trong khoảng từ 37 đến 50oC, pH từ 7 đến 8.
3. Đã phân loại và xác định được 3 chủng có khả năng xử lý nitơ tốt nhất,
trong đó, chủng B11.11 có khả năng chuyển hóa 85.21% NH4+ sau 20 ngày nuôi
cấy, chủng B21.10 chuyển hóa 97,28% và B23.2 chuyển hóa 97,14% lượng NO2-
sau 20 ngày nuôi cấy.
4. Đã phân loại và xác định được chủng A4.2 có khả năng xử lý photpho tốt
nhất, sau 10 ngày, hàm lượng photpho trong môi trường giảm đi 39.32% tương ứng
với môi trường có hàm lượng photpho là 18 mg/l.
5. Dựa trên trình tự gen 16S rARN, đặc điểm khuẩn lạc, tế bào, các đặc điểm
sinh lý, sinh hóa, chủng B11.11 và A4.2 được xác định gần với loài Bacillus
licheniformis, chủng B21.10 được xác định gần với loài Bacillus amyloliquefaciens,
chủng B23.2 xác định gần với loài Pseudomonas pseudoalcaligenes.
KIẾN NGHỊ
1. Thay đổi hàm lượng nitơ và photpho phù hợp với nghiên cứu. Kiểm tra khả
năng chuyển hóa các chất trong môi trường và sự hình thành màng sinh học khi
thay đổi hàm lượng.
2. Nghiên cứu, tối ưu hóa điều kiện chuyển hóa các hợp chất nitơ và photpho,
nghiên cứu và lựa chọn giá thể, chất mang phù hợp cho sự hình thành màng sinh
học của các chủng nghiên cứu.

Luận văn thạc sĩ
Ngô Thị Kim Toán 55 K19 – Sinh học thực nghiệm
TÀI LIỆU THAM KHẢO
Tiếng Việt
1. Kiểu Hữu Ảnh (2006), Giáo trình vi sinh vật học, phần 1, NXB Đại học
Quốc Gia Hà Nội
2. Bộ Tài Nguyên và Môi trường (2010), Báo cáo môi trường Quốc gia 2010,
Hà Nội.
3. Lê Văn Cát (2007), Xử lý nước thải giàu hợp chất nitơ và photpho, NXB
Khoa học tự nhiên và Công nghệ.
4. Lương Đức Phẩm (2003), Công nghệ xử lý nước thải bằng biện pháp sinh
học, NXB Giáo dục.
5. Nguyễn Hoài Hương (2009), Giáo trình thực hành vi sinh ứng dụng, NXB
Đại học Quốc Gia TPHCM.
6. Nguyễn Văn Phước (2007), Xử lý nước thải sinh hoạt và công nghiệp bằng
phương pháp sinh học, NXB Xây dựng.
Tiếng Anh
7. Anderson I.C., Poth M., Homstead J., and Burdige D. (1993), “A comparison
of NO and N2O production by the autotrophic nitrifier Nitrosomonas
europaea and the heterotrophic nitrifier Alcaligenes faecalis”, Applied and
Environmental Microbiology, 59 (11), pp. 3525-3533.
8. Annachhatre A.P. and Bhamidimarri S.M.R. (1992), “Microbial attachment
and growth in fixed-film reactors: Process startup considerations”,
Biotechnology Advances, 10 (1), pp. 69-91.
9. APHA (2001), Standard Methods for the Examination of Water and
Wastewate., 20th edition, American Public Health Association, Washington,
DC.
10. Asgari M.J., Safavi K., and Mortazaeinezahad F. (2011), “Landfill biogas
production process”, International Conference on Food Engineering and
Biotechnology, 9, pp. 208-212

Luận văn thạc sĩ
Ngô Thị Kim Toán 56 K19 – Sinh học thực nghiệm
11. Bao L.-L., Li D., Li X.K., Huang R.X., Zhang J., Yang L., and Xia G.Q.
(2007), “Phosphorus accumulation by bacteria isolated from a continuous-
flow two-sludge system”, Journal of Environmental Sciences, 19 (4), pp.
391-395.
12. Bernet N., Dangcong P., Delgenès J., and Moletta R. (2001), “Nitrification at
low oxygen concentration in biofilm reactor”, Journal of Environmental
Engineering, 127 (3), pp. 266-271.
13. Boelee N.C., Temmink H., Janssen M., Buisman C.J.N., and Wijffels R.H.
(2011), “Nitrogen and phosphorus removal from municipal wastewater
effluent using microalgal biofilms”, Water Research, 45 (18), pp. 5925-5933.
14. Boyd C.E. and Tucker C.S. (1998), Pond Aquaculture Water Quality
Management, Kluwer Acad. Publ.
15. Broda E. (1977), “Two kinds of lithotrophs missing in nature”, Zeitschrift für
allgemeine Mikrobiologie, 17 (6), pp. 491-493.
16. Cheung K.C., Chu L.M., and Wong M.H. (1997), “Ammonia stripping as a
pretreatment for landfill leachate”, Water, Air, and Soil Pollution, 94 (1-2),
pp. 209-221.
17. Cong L.T.N., Huyen H.T., and Minh N.N. (2012), “Phenol degradation of
biofilm formed by mixing - marine bacteria”, VNU. Journal of Science, 28
(2S), pp. 75-81.
18. Costerton J.W., Lewandowski Z., Caldwell D.E., Korber D.R., and Lappin-
Scott H.M. (1995), “Microbial biofilms”, Annual Review of Microbiology,
49, pp. 711-745.
19. Czaczyk K. and Myszka K. (2007), “Biosynthesis of extracellular polymeric
substances (EPS) and its role in microbial biofilm formation”, Polish Journal
of Environmental Studies, 16 (6), pp. 799-806.
20. Di Bonaventura G., Stepanovic S., Picciani C., Pompilio A., and Piccolomini
R. (2007), “Effect of environmental factors on biofilm formation by clinical

Luận văn thạc sĩ
Ngô Thị Kim Toán 57 K19 – Sinh học thực nghiệm
Stenotrophomonas maltophilia isolates”, Folia Microbiologica., 52 (1), pp.
86-90.
21. Donlan R.M. (2002), “Biofilms: microbial life on surfaces”, Emerging
Infectious Diseases Journal, 8 (9), pp. 881-890.
22. Federation W.E. (1998), Biological and chemical systems for nutrient
removal, Water Environment Federation, Alexandria, VA.
23. Flemming H.-C. (1993), “Biofilms and Environmental Protection”, Water
Science & Technology, 27 (7-8), pp. 1-10.
24. Giaouris E., Chorianopoulos N., and Nychas G.J.E. (2005), “Effect of
temperature, pH, and water activity on biofilm formation by Salmonella
enterica enteritidis PT4 on stainless steel surfaces as indicated by the bead
vortexing method and conductance measurements”, Journal of Food
Protection, 68 (10), pp. 2149-2154.
25. Gilbert P., Das J., and Foley I. (1997), “Biofilm susceptibility to
antimicrobials”, Advances in Dental Research, 11 (1), pp. 160-167.
26. Hang T.T. and Huy N.Q. (2011), “Isolate biofilm forming Bacillus strains
from contamination site in trade villages in Viet Nam”, VNU. Journal of
Science, 27 (2S), pp. 157-162.
27. Henze M., Harremoes P., Jansen J.C., and Arvin E. (2001), Wastewater
Treatment: Biological and Chemical Processes, Springer.
28. Heydorn A., Nielsen A.T., Hentzer M., Sternberg C., Givskov M., Ersboll
B.K., and Molin S. (2000), “Quantification of biofilm structures by the novel
computer program COMSTAT”, Microbiology (Reading, England), 146
(10), pp. 2395-2407.
29. Ho K.L., Pometto A.L., and Hinz P.N. (1997), “Optimization of L-(+)-lactic
acid production by ring and disc plastic composite supports through
repeated-batch biofilm fermentation”, Applied and Environmental
Microbiology, 63 (7), pp. 2533-2542.

Luận văn thạc sĩ
Ngô Thị Kim Toán 58 K19 – Sinh học thực nghiệm
30. Hoilijoki T.H., Kettunen R.H., and Rintala J.A. (2000), “Nitrification of
anaerobically pretreated municipal landfill leachate at low temperature”,
Water Research, 34 (5), pp. 1435-1446.
31. Hunik J.H., Van Den Hoogen M.P., De Boer W., Smit M., and Tramper J.
(1993), “Quantitative determination of the spatial distribution of
Nitrosomonas europaea and Nitrobacter agilis cells immobilized in kappa-
carrageenan gel beads by a specific Ffuorescent-antibody labelling
technique”, Applied and Environmental Microbiology, 59 (6), pp. 1951-
1954.
32. Huy N.Q., Lien N.T.P., and Hang T.T. (2011), “Characterization of biofilm-
forming bacteria isolated from soil in Viet Nam”, VNU. Journal of Science,
27 (2S), pp. 187-193.
33. Jørgensen K.S. and Pauli A.S.L. (1995), “Polyphosphate accumulation
among denitrifying bacteria in activated sludge”, Anaerobe, 1 (3), pp. 161-
168.
34. Kim J.K., Park K.J., Cho K.S., Nam S.-W., Park T.-J., and Bajpai R. (2005),
“Aerobic nitrification–denitrification by heterotrophic Bacillus strains”,
Bioresource Technology, 96 (17), pp. 1897-1906.
35. Kokare C.R.C., Khopade A.N., and Mahadik K. (2009), “Biofilm :
importance and applications”, Indian Journal of Biotechnology, 18, pp. 159-
168.
36. Lacko N., Drysdale G.D., and Bux F. (2003), “Anoxic phosphorus removal
by denitrifying heterotrophic bacteria”, Water science and technology : a
journal of the International Association on Water Pollution Research, 47
(11), pp. 17-22.
37. Lazarova V. and Manem J. (1995), “Biofilm characterization and activity
analysis in water and wastewater treatment”, Water Research, 29 (10), pp.
2227-2245.

Luận văn thạc sĩ
Ngô Thị Kim Toán 59 K19 – Sinh học thực nghiệm
38. Li X.Z., Zhao Q.L., and Hao X.D. (1999), “Ammonium removal from
landfill leachate by chemical precipitation”, Waste Management, 19 (6), pp.
409-415.
39. Lopez D., Vlamakis H., and Kolter R. (2010), “Biofilms”, Cold Spring
Harbor Perspectives in Biology, 2 (7), pp. a000398.
40. Monroe D. (2007), “Looking for chinks in the armor of bacterial biofilms”,
PLoS Biology, 5 (11).
41. Mulder A. (2003), “The quest for sustainable nitrogen removal
technologies”, Water science and technology : a journal of the International
Association on Water Pollution Research, 48 (1), pp. 67-75.
42. Mulder A., Van De Graaf A.A., Robertson L.A., and Kuenen J.G. (1995),
“Anaerobic ammonium oxidation discovered in a denitrifying fluidized bed
reactor”, FEMS Microbiology Ecology, 16 (3), pp. 177-183.
43. Nadell C.D., Xavier J.B., Levin S.A., and Foster K.R. (2008), “The evolution
of quorum sensing in bacterial biofilms”, PLoS biology, 6 (1), pp. e14.
44. O'toole G., Kaplan H.B., and Kolter R. (2000), “Biofilm formation as
microbial development”, Annual Review of Microbiology, 54, pp. 49-79.
45. O'toole G.A., Gibbs K.A., Hager P.W., Phibbs P.V., Jr., and Kolter R.
(2000), “The global carbon metabolism regulator Crc is a component of a
signal transduction pathway required for biofilm development by
Pseudomonas aeruginosa”, Journal of Bacteriology, 182 (2), pp. 425-431.
46. O'toole G.A. and Kolter R. (1998), “Flagellar and twitching motility are
necessary for Pseudomonas aeruginosa biofilm development”, Molecular
Microbiology, 30 (2), pp. 295-304.
47. O'toole G.A. and Kolter R. (1998), “Initiation of biofilm formation in
Pseudomonas fluorescens WCS365 proceeds via multiple, convergent
signalling pathways: a genetic analysis”, Molecular Microbiology, 28 (3),
pp. 449-461.

Luận văn thạc sĩ
Ngô Thị Kim Toán 60 K19 – Sinh học thực nghiệm
48. Ozturk I., Altinbas M., Koyuncu I., Arikan O., and Gomec-Yangin C.
(2003), “Advanced physico-chemical treatment experiences on young
municipal landfill leachates”, Waste Management, 23 (5), pp. 441-446.
49. Pressley T.A., Bishop D.F., and Roan S.G. (1972), “Ammonia-nitrogen
removal by breakpoint chlorination”, Environmental Science & Technology,
6 (7), pp. 622-628.
50. Radwan S.S., Al-Hasan R.H., Salamah S., and Al-Dabbous S. (2002),
“Bioremediation of oily sea water by bacteria immobilized in biofilms
coating macroalgae”, International Biodeterioration & Biodegradation,
50 (1), pp. 55-59.
51. Rieger L., Koch G., Kühni M., Gujer W., and Siegrist H. (2001), “The eawag
bio-p module for activated sludge model no. 3”, Water Research, 35 (16),
pp. 3887-3903.
52. Schmid M.C., Maas B., Dapena A., Pas-Schoonen K.V.D, Vossenberg
J.V.D, Kartal B., Niftrik L.V, Schmidt I., Cirpus I., Kuenen J. G., Wagner
M., Damsté J.S.S., Kuypers M., Revsbech N. P, Mendez R., Jetten M. S., and
Strous M. (2005), “Biomarkers for in situ detection of anaerobic ammonium-
oxidizing (anammox) bacteria”, Applied and Environmental Microbiology,
71 (4), pp. 1677-1684.
53. Schmidt I. and Bock E. (1997), “Anaerobic ammonia oxidation with nitrogen
dioxide by Nitrosomonas eutropha”, Archives of Microbiology, 167 (2-3),
pp. 106-111.
54. Schmidt I. and Bock E. (1998), “Anaerobic ammonia oxidation by cell-free
extracts of Nitrosomonas eutropha”, Antonie Van Leeuwenhoek, 73 (3), pp.
271-278.
55. Sedlak R.I. (1991), Phosphorus and Nitrogen Removal From Municipal
Wastewater: Principles and Practice, Lewis Publication.
56. Sedlak R.I. (1991), Phosphorus and Nitrogen Removal from Municipal
Wastewater: Principles and Practice, 2nd Edition, CRC Press, English.

Luận văn thạc sĩ
Ngô Thị Kim Toán 61 K19 – Sinh học thực nghiệm
57. Sharma B. and Ahlert R.C. (1977), “ Nitrification and nitrogen removal”,
Water Research, 11, pp. 897-925.
58. Shoji T., Satoh H., and Mino T. (2003), “Quantitative estimation of the role
of denitrifying phosphate accumulating organisms in nutrient removal”,
Water science and technology : a journal of the International Association on
Water Pollution Research, 47 (11), pp. 23-29.
59. Sidat M, Bux F., and Kasan H. (1999), “Polyphosphate accumulation by
bacteria isolated from activated sludge”, Water South Africa, 25 (2), pp. 175-
180.
60. Siegrist H., Rieger L., Koch G., Kuhni M., and Gujer W. (2002), “The eawag
bio-P module for activated sludge model No. 3”, Water science and
technology : a journal of the International Association on Water Pollution
Research, 45 (6), pp. 61-76.
61. Streichan M., Golecki J.R., and Schön G. (1990), “Polyphosphate-
accumulating bacteria from sewage plants with different proceses for
biological phosphorus removal”, FEMS Microbiology Letters, 73 (2), pp.
113-124.
62. Sutherland I. (2001), “Biofilm exopolysaccharides: a strong and sticky
framework”, Microbiology (Reading, England), 147 (Pt 1), pp. 3-9.
63. Van Benthum W.A.J., Van Loosdrecht M.D.M., and Heijnen J.J. (1997),
“Control of heterotrophic layer formation on nitrifying biofilms in a biofilm
airlift suspension reactor”, Biotechnology and Bioengineering, 53 (4), pp.
397-405.
64. Welander U., Henrysson T., and Welander T. (1998), “Biological nitrogen
removal from municipal landfill leachate in a pilot scale suspended carrier
biofilm process”, Water Research, 32 (5), pp. 1564-1570.
65. Wulff N.A., Mariano A.G., Gaurivaud P., De Almeida Souza L.C., Virgilio
A.C., and Monteiro P.B. (2008), “Influence of culture medium pH on

Luận văn thạc sĩ
Ngô Thị Kim Toán 62 K19 – Sinh học thực nghiệm
growth, aggregation, and biofilm formation of Xylella fastidiosa”, Current
Microbiology, 57 (2), pp. 127-132.
66. Yangin C., Yilmaz S., Altinbas M., and Ozturk I. (2002), “A new process for
the combined treatment of municipal wastewaters and landfill leachates in
coastal areas”, Water science and technology : a journal of the International
Association on Water Pollution Research, 46 (8), pp. 111-118.
67. Zhang J., Wu P., Hao B., and Yu Z. (2011), “Heterotrophic nitrification and
aerobic denitrification by the bacterium Pseudomonas stutzeri YZN-001”,
Bioresource Technology, 102 (21), pp. 9866-9869.
68. Zhang Q.-L., Liu Y., Ai G.-M., Miao L.-L., Zheng H.-Y., and Liu Z.-P.
(2012), “The characteristics of a novel heterotrophic nitrification–aerobic
denitrification bacterium, Bacillus methylotrophicus strain L7”, Bioresource
Technology, 108 (0), pp. 35-44.
Website
69. http://en.wikipedia.org/wiki/Nitrogen_cycle
70. http://www.visionlearning.com/library/module_viewer.php?mid=98
71. http://www.thewaterplanetcompany.com/docs/WPC_Nitrification and
Denitrification.pdf

Luận văn thạc sĩ
Ngô Thị Kim Toán K19 – Sinh học thực nghiệm
PHỤ LỤC
1. Trình tự gen mã hóa 16S rARN của vi khuẩn B11.11 được đọc trên máy
đọc trình tự tự động ABI PRISM 3100 Avant (Hoa Kỳ) .
GTGCCTAATACATGCAAGTCGAGCGGACCGACGGGAGCTTGCTCCCTTA
GGTCAGCGGCGGACGGGTGAGTAACACGTGGGTAACCTGCCTGTAAGA
CTGGGATAACTCCGGAAACCGGGGCTAATACCGGATGCTTGATTGAACC
GCATGGTTCAATCATAAAAGGTGGCTTTTAGCTACCACTTACAGATGGA
CCCGCGGCGCATTAGCTAGTTGGTGAGGTAACGGCTCACCAAGGCGACG
ATGCGTAGCCGACCTGAGAGGGTGATCGGCCACACTGGGACTGAGACA
CGGCCCAGACTCCTACGGGAGGCAGCAGTAGGGAATCTTCCGCAATGG
ACGAAAGTCTGACGGAGCAACGCCGCGTGAGTGATGAAGGTTTTCGGAT
CGTAAAACTCTGTTGTTAGGGAAGAACAAGTACCGTTCGAATAGGGCGG
CACCTTGACGGTACCTAACCAGAAAGCCACGGCTAACTACGTGCCAGCA
GCCGCGGTAATACGTAGGTGGCAAGCGTTGTCCGGAATTATTGGGCGTA
AAGCGCGCGCAGGCGGTTTCTTAAGTCTGATGTGAAAGCCCCCGGCTCA
ACCGGGGAGGGTCATTGGAAACTGGGGAACTTGAGTGCAGAAGAGGAG
AGTGGAATTCCACGTGTAGCGGTGAAATGCGTAGAGATGTGGAGGAAC
ACCAGTGGCGAAGGCGACTCTCTGGTCTGTAACTGACGCTGAGGCGCGA
AAGCGTGGGGAGCGAACAGGATTAGATACCCTGGTAGTCCACGCCGTA
AACGATGAGTGCTAAGTGTTAGAGGGTTTCCGCCCTTTAGTGCTGCAGC
AAACGCATTAAGCACTCCGCCTGGGGAGTACGGTCGCAAGACTGAAACT
CAAAGGAATTGACGGGGGCCCGCACAAGCGGTGGAGCATGTGGTTTAA
TTCGAAGCAACGCGAAGAACCTTACCAGGTCTTGACATCCTCTGACAAC
CCTAGAGATAGGGCTTCCCCTTCGGGGGCAGAGTGACAGGTGGTGCATG
GTTGTCGTCAGCTCGTGTCGTGAGATGTTGGGTTAAGTCCCGCAACGAG
CGCAACCCTTGATCTTAGTTGCCAGCATTCAGTTGGGCACTCTAAGGTG
ACTGCCGGTGACAAACCGGAGGAAGGTGGGGATGACGTCAAATCATCA
TGCCCCTTATGACCTGGGCTACACACGTGCTACAATGGGCAGAACAAAG

Luận văn thạc sĩ
Ngô Thị Kim Toán K19 – Sinh học thực nghiệm
GGCAGCGAAGCCGCGAGGCTAAGCCAATCCCACAAATCTGTTCTCAGTT
CGGATCGCAGTCTGCAACTCGACTGCGTGAAGCTGGAATCGCTAGTAAT
CGCGGATCAGCATGCCGCGGTGAATACGTTCCCGGGCCTTGTACACACC
GCCCGTCACACCACGAGAGTTTGTAACACCCGAAGTCGGTGAGGTAACC
TTTTGGAGCCAGCCGCCGAAGGTGGGACAGA
2. Trình tự gen mã hóa 16S rARN của chủng B21.10 được đọc trên máy đọc
trình tự tự động ABI PRISM 3100 Avant (Hoa Kỳ).
AGGACGAACGCTGGCGGCGTGCCTAATACATGCAAGTCGAGCGGACAG
ATGGGAGCTTGCTCCCTGATGTTAGCGGCGGACGGGTGAGTAACACGTG
GGTAACCTGCCTGTAAGACTGGGATAACTCCGGGAAACCGGGGCTAATA
CCGGATGGTTGTCTGAACCGCATGGTTCAGACATAAAAGGTGGCTTCGG
CTACCACTTACAGATGGACCCGCGGCGCATTAGCTAGTTGGTGAGGTAA
CGGCTCACCAAGGCGACGATGCGTAGCCGACCTGAGAGGGTGATCGGC
CACACTGGGACTGAGACACGGCCCAGACTCCTACGGGAGGCAGCAGTA
GGGAATCTTCCGCAATGGACGAAAGTCTGACGGAGCAACGCCGCGTGA
GTGATGAAGGTTTTCGGATCGTAAAGCTCTGTTGTTAGGGAAGAACAAG
TGCCGTTCAAATAGGGCGGCACCTTGACGGTACCTAACCAGAAAGCCAC
GGCTAACTACGTGCCAGCAGCCGCGGTAATACGTAGGTGGCAAGCGTTG
TCCGGAATTATTGGGCGTAAAGGGCTCGCAGGCGGTTTCTTAAGTCTGA
TGTGAAAGCCCCCGGCTCAACCGGGGAGGGTCATTGGAAACTGGGGAA
CTTGAGTGCAGAAGAGGAGAGTGGAATTCCACGTGTAGCGGTGAAATG
CGTAGAGATGTGGAGGAACACCAGTGGCGAAGGCGACTCTCTGGTCTGT
AACTGACGCTGAGGAGCGAAAGCGTGGGGAGCGAACAGGATTAGATAC
CCTGGTAGTCCACGCCGTAAACGATGAGTGCTAAGTGTTAGGGGGTTTC
CGCCCCTTAGTGCTGCAGCTAACGCATTAAGCACTCCGCCTGGGGAGTA
CGGTCGCAAGACTGAAACTCAAAGGAATTGACGGGGGCCCGCACAAGC
GGTGGAGCATGTGGTTTAATTCGAAGCAACGCGAAGAACCTTACCAGGT
CTTGACATCCTCTGACAATCCTAGAGATAGGACGTCCCCTTCGGGGGCA
GAGTGACAGGTGGTGCATGGTTGTCGTCAGCTCGTGTCGTGAGATGTTG

Luận văn thạc sĩ
Ngô Thị Kim Toán K19 – Sinh học thực nghiệm
GGTTAAGTCCCGCAACGAGCGCAACCCTTGATCTTAGTTGCCAGCATTC
AGTTGGGCACTCTAAGGTGACTGCCGGTGACAAACCGGAGGAAGGTGG
GGATGACGTCAAATCATCATGCCCCTTATGACCTGGGCTACACACGTGC
TACAATGGACAGAACAAAGGGCAGCGAAACCGCGAGGTTAAGCCAATC
CCACAAATCTGTTCTCAGTTCGGATCGCAGTCTGCAACTCGACTGCGTG
AAGCTGGAATCGCTAGTAATCGCGGATCAGCATGCCGCGGTGAATACGT
TCCCGGGCCTTGTACACACCGCCCGTCACACCACGAGAGTTTGTAACAC
CCGAAGTCGGTGAGGTAACCTTTTAGGAGCCAGCCGCCGAAGGTGGGA
CAGA
3. Trình tự gen mã hóa 16S rARN của chủng B23.2 được đọc trên máy đọc
trình tự tự động ABI PRISM 3100 Avant (Hoa Kỳ).
AGATTGAACGCTGGCGGCAGGCCTAACACATGCAAGTCGAGCGGATGA
AGGGAGCTTGCTCCTGGATTTAGCGGCGGACGGGTGAGTAATGCCTAGG
AATCTGCCTGGTAGTGGGGGATAACGTTCCGAAAGGAACGCTAATACCG
CGTACGTCCTACGGGAGAAAGCAGGGGACCTTCGGGCCTTGCGCTATCA
GATGAGCCTAGGTCGGATTAGCTAGTTGGTGAGGTAATGGCTCACCAAG
GCGACGATCCGTAACTGGTCTGAGAGGATGATCAGTCACACTGGAACTG
AGACACGGTCCAGACTCCTACGGGAGGCAGCAGTGGGGAATATTGGAC
AATGGGCGAAAGCCTGATCCAGCCATGCCGCGTGTGTGAAGAAGGTCTT
CGGATTGTAAAGCACTTTAAGTTGGGAGGAAGGGTTGTACGTTAATACC
GTGCAATTTTGACGTTACCGACAGAATAAGCACCGGCTAACTTCGTGCC
AGCAGCCGCGGTAATACGAAGGGTGCAAGCGTTAATCGGAATTACTGG
GCGTAAAGCGCGCGTAGGTGGTTCAGTAAGTTGGAAGTGAAATCCCCGG
GCTCAACCTGGGAACTGCTTTCAAAACTGCTGAGCTAGAGTACGGTAGA
GGGTAGTGGAATTTCCTGTGTAGCGGTGAAATGCGTAGATATAGGAAGG
AACACCAGTGGCGAAGGCGACTACCTGGACTGATACTGACACTGAGGT
GCGAAAGCGTGGGGAGCAAACAGGATTAGATACCCTGGTAGTCCACGC
CGTAAACGATGTCAACTAGCCGTTGGGATCCTTGAGATCTTAGTGGCGC
AGCTAACGCATTAAGTTGACCGCCTGGGGAGTACGGCCGCAAGGTTAAA

Luận văn thạc sĩ
Ngô Thị Kim Toán K19 – Sinh học thực nghiệm
ACTCAAATGAATTGACGGGGGCCCGCACAAGCGGTGGAGCATGTGGTTT
AATTCGAAGCAACGCGAAGAACCTTACCTGGCCTTGACATGCTGAGAAC
TTTCCAGAGATGGATTGGTGCCTTCGGGAACTCAGACACAGGTGCTGCA
TGGCTGTCGTCAGCTCGTGTCGTGAGATGTTGGGTTAAGTCCCGTAACG
AGCGCAACCCTTGTCCTTAGTTACCAGCACCTCGGGTGGGCACTCTAAG
GAGACTGCCGGTGACAAACCGGAGGAAGGTGGGGATGACGTCAAGTCA
TCATGGCCCTTACGGCCAGGGCTACACACGTGCTACAATGGTCGGTACA
AAGGGTTGCCAAGCCGCGAGGTGGAGCTAATCCCATAAAACCGATCGT
AGTCCGGATCGCAGTCTGCAACTCGACTGCGTGAAGTCGGAATCGCTAG
TAATCGTGAATCAGAATGTCACGGTGAATACGTTCCCGGGCCTTGTACA
CACCGCCCGTCACACCATGGGAGTGGGTTGCTCCAGAAGTAGCTAGTCT
AACCTTCGGGGGGACGGTACCACGGAG
4. Trình tự gen mã hóa 16S rARN của chủng A4.2 được đọc trên máy đọc
trình tự tự động ABI PRISM 3100 Avant (Hoa Kỳ).
CTCTGCTCAGGACGAACGCCGGCGGCGTGCCTAATACATGCAAGTCGAG
CGGACCGACGGGAGCTTGCTCCCTTAGGTTAGTGGCGGACGGGTGAGTA
ACACGTGGGTAACCTGCCTGTAAGATTGGGATAACTCCGGGAAACCGGG
GCTAATACCGGATGCTTGATTGAACCGCATGGTTCAATCATAAAAGGTG
GCTTTTAGCTACCACTTACAGATGGACCCGCGGCGCATTAGCTAGTTGG
TGAGGTAACGGCTCACCAAGGCGACGATGCGTAGCCGACCTGAGAGGG
TGATCGGCCACACTGGGACTGAGACACGGCCCAGACTCCTACGGGAGG
CAGCAGTAGGGAATCTTCCGCAATGGACGAAAGTCTGACGGAGCAACG
CCGCGTGAGTGATGAAGGTTTTCGGATCGTAAAACTCTGTTGTTAGGGA
AGAACAAGTACCGTTCGAATAGGGCGGCACCTTGACGGTACCTAACCAG
AAAGCCACGGCTAACTACGTGCCAGCAGCCGCGGTAATACGTAGGTGG
CAAGCGTTGTCCGGAATTATTGGGCGTAAAGCGCGCGCAGGCGGTTTCT
TAAGTCTGATGTGAAAGCCCCCGGCTCAACCGGGGAGGGTCATTGGAAA
CTGGGGAACTTGAGTGCAGAAGAGGAGAGTGGAATTCCACGTGTAGCG
GTGAAATGCGTAGAGATGTGGAGGAACACCAGTGGCGAAGGCGACTCT

Luận văn thạc sĩ
Ngô Thị Kim Toán K19 – Sinh học thực nghiệm
CTGGTCTGTAACTGACGCTGAGGCGCGAAAGCGTGGGGAGCGAACAGG
ATTAGATACCCTGGTAGTCCACGCCGTAAACGATGAGTGCTAAGTGTTA
GAGGGTTTCCGCCCTTTAGTGCTGCAGCAAACGCATTAAGCACTCCGCC
TGGGGAGTACGGTCGCAAGACTGAAACTCAAAGGAATTGACGGGGGCC
CGCACAAGCGGTGGAGCATGTGGTTTAATTCGAAGCAACGCGAAGAAC
CTTACCAGGTCTTGACATCCTCTGACAACCCTAGAGATAGGGCTTCCCCT
TCGGGGGCAGAGTGACAGGTGGTGCATGGTTGTCGTCAGCTCGTGTCGT
GAGATGTTGGGTTAAGTCCCGCAACGAGCGCAACCCTTGATCTTAGTTG
CCAGCATTCAGTTGGGCACTCTAAGGTGACTGCCGGTGACAAACCGGAG
GAAGGTGGGGATGACGTCAAATCATCATGCCCCTTATGACCTGGGCTAC
ACACGTGCTACAATGGGCAGAACAAAGGGCAGCGAAGCCGCGAGGCTA
AGCCAATCCCACAAATCTGTTCTCAGTTCGGATCGCAGTCTGCAACTCG
ACTGCGTGAAGCTGGAATCGCTAGTAATCGCGGATCAGCATGCCGCGGT
GAATACGTTCCCGGGCCTTGTACACACCGCCCGTCACACCACGAGAGTT
TGTAACACCCGAAGTCGGTGAGGTAACCTTTGGAGCCAGCCGCCGAAGT
GGACAGAATGACTCA