hypoxia inducible factor-1α mediates iron uptake which induces inflammatory response in amoeboid...
TRANSCRIPT

lable at ScienceDirect
Neuropharmacology 77 (2014) 428e440
Contents lists avai
Neuropharmacology
journal homepage: www.elsevier .com/locate/neuropharm
Hypoxia inducible factor-1a mediates iron uptake which inducesinflammatory response in amoeboid microglial cells in developingperiventricular white matter through MAP kinase pathway
Gurugirijha Rathnasamy, Eng-Ang Ling, Charanjit Kaur*
Department of Anatomy, MD10, 4 Medical Drive, Yong Loo Lin School of Medicine, National University of Singapore, Singapore 117594, Singapore
a r t i c l e i n f o
Article history:Received 10 May 2013Received in revised form4 October 2013Accepted 15 October 2013
Keywords:HypoxiaMicrogliaIronCytokinesHIF-1aMAPKsDeferoxamineKC7F2
* Corresponding author. Tel.: þ65 65163209; fax: þE-mail address: [email protected] (C. Kaur).
0028-3908/$ e see front matter � 2013 Elsevier Ltd.http://dx.doi.org/10.1016/j.neuropharm.2013.10.024
a b s t r a c t
Iron accumulation occurs in tissues such as periventricular white matter (PWM) in response to hypoxicinjuries, and microglial cells sequester excess iron following hypoxic exposure. As hypoxia has a role inaltering the expression of proteins involved in iron regulation, this study was aimed at examining theinteraction between hypoxia inducible factor (HIF)-1a and proteins involved in iron transport in micro-glial cells, and evaluating the mechanistic action of deferoxamine and KC7F2 (an inhibitor of HIF-1a)in iron mediated hypoxic injury. Treating the microglial cultures with KC7F2, led to decreased expressionof transferrin receptor and divalent metal transporter-1. Administration of deferoxamine or KC7F2 tohypoxic microglial cells enhanced extracellular signal-regulated kinase (ERK) phosphorylation (p-ERK),but decreased the phosphorylation of p38 (p-p38). The increased p-ERK further phosphorylated the cAMPresponse element-binding protein (p-CREB) which in turn may have resulted in the increased mitogenactivated protein kinase (MAPK) phosphatase 1 (MKP1), known to dephosphorylate MAPKs. Consistentwith the decrease in p-p38, the production of pro-inflammatory cytokines TNF-a and IL-1bwas reduced inhypoxic microglia treated with deferoxamine and SB 202190, an inhibitor for p38. This suggests that theanti-inflammatory effect exhibited by deferoxamine is by inhibition of p-p38 induced inflammationthrough the pERK-pCREB-MKP1 pathway, whereas that of KC7F2 requires further investigation. Thepresent results suggest that HIF-1a may mediate iron accumulation in hypoxic microglia and KC7F2,similar to deferoxamine, might provide limited protection against iron induced PWMD.
� 2013 Elsevier Ltd. All rights reserved.
1. Introduction
Iron remains as an indispensable nutrient required for thenormal development of the brain owing to its vital role in energymetabolism, neurotransmitter synthesis and myelination (Beard,2003). However, any perturbation in iron levels in the brain maylead to neurobehavioral abnormalities (Beard, 2003; Lozoff andGeorgieff, 2006). Hypoxic-ischemic injuries, which remain to bethe major cause of mortality and neurologic morbidity in infants(Chang and Huang, 2006; Scott and Hegyi, 1997), have been re-ported to alter iron levels in the developing brain (Adcock et al.,1996; Palmer et al., 1999). In the perinatal period, the periven-tricular white matter (PWM), peripheral to the lateral ventricles ishighly susceptible to hypoxic-ischemic damage (Follett et al., 2004;Johnston, 1997; Rezaie and Dean, 2002; Volpe, 2003). Pathogenesisof PWM damage (PWMD), which is characterized by axonal
65 67787643.
All rights reserved.
degeneration, oligodendrocyte death, microglial activation andastrocytosis (Ness et al., 2001; Skoff et al., 2001; Takashima et al.,1995), is not yet fully understood due to the complexity in themechanisms leading to such damage. Recently, we reported that,following hypoxic exposure there was increased iron accumulationin PWM in neonatal rats and that the increased ironwas localized inamoeboid microglial cells (AMCs), the resident macrophages of thebrain (Kaur and Ling, 1999; Rathnasamy et al., 2011).
Hypoxic injury results in the activation of transcription complex,hypoxia inducible factor (HIF)-1, which induces the transcription ofa series of genes that aid the survival and adaptation of a cell tohypoxia. HIF-1 consists of a hypoxia inducible HIF-1a subunit andconstitutively expressed HIF-1b subunit (Wang et al., 1995). Thestabilization and translocation of HIF-1a to the nucleus is essentialto form the functional HIF-1 (Huang et al., 1996; Kallio et al., 1997).HIF-1a has been reported to favour transcription of genes involvedin iron homeostasis (Bianchi et al., 1999; Li et al., 2008; Qian et al.,2011; Tacchini et al., 1999). Transcription of proteins involved iniron uptake such as transferrin receptor (TfR) and divalent metaltransporter (DMT)-1 has been reported to bemediated by HIF-1a in

G. Rathnasamy et al. / Neuropharmacology 77 (2014) 428e440 429
various cell types such as Hep3B and HepG2 hepatoma cells, pri-mary astrocytes and neurons (Bianchi et al., 1999; Li et al., 2008;Qian et al., 2011; Tacchini et al., 1999). However, this has not beenestablished in microglia. In addition to the presence of iron regu-latory proteins in microglia reported by us, we further investigatedif HIF-1a would mediate the uptake of iron. The role of HIF-1a wasstudied using KC7F2, a HIF-1a translational inhibitor.
It is well documented that, activated microglia in hypoxic PWMare a source of pro-inflammatory cytokines such tumour necrosisfactor-a (TNF-a) and interleukin-1b (IL-1b) (Deng et al., 2008;Murugan et al., 2011; Zhang et al., 2005), nitric oxide (Muruganet al., 2011) and reactive oxygen species (ROS) (Colton et al.,1996). Consistent with this, iron accumulation in activated micro-glia was demonstrated to result in increased production of ROS,TNF-a and IL-1b (Rathnasamy et al., 2011; Zhang et al., 2006b).These factors secreted bymicroglia have been reported to harm theoligodendrocytes in the hypoxic PWM (Rathnasamy et al., 2011).Deferoxamine, a well-known iron chelator used in clinical practicefor more than 30 years (Selim, 2009), has been proposed as atherapeutic alternative to suppress microglia mediated inflamma-tion in response to iron accumulation (Rathnasamy et al., 2011;Selim, 2009; Zhang et al., 2006b). However, to the best of ourknowledge, the ability of HIF-1a inhibitors to act against ironmediated inflammation has not been investigated. Although, pastinvestigations identified deferoxamine to possess antioxidantproperty (Peeters-Scholte et al., 2003), neuroprotective effects(Sorond et al., 2009) and suppression of microglial activation(Miao et al., 2012; Wu et al., 2011), the mechanism throughwhich deferoxamine modulates the microglial response remainunexplored.
Mitogen activated protein kinases (MAPKs) are a family of pro-teins well known to respond to a wide range of stimuli (Chang andKarin, 2001). The MAPK family of proteins consisting of the extra-cellular signal-regulated kinase (ERK), p38 and c-Jun NH2-terminalkinase (JNK) are regulators of cell survival, proliferation, differen-tiation, apoptosis and inflammation. They do this by activatingvarious transcription factors such as cyclic adenosine mono-phosphate (cAMP) responsive element binding protein (CREB) andactivator protein (AP)-1 (Huang et al., 2007). The finding thatMAPKs mediate the inflammatory pathway in hypoxic microglialcells (Deng et al., 2008) led us to the hypothesis that iron mightmediate cellular changes in hypoxic microglia via the MAPKpathways.
Based on the above, this study was aimed at elucidating the roleof HIF-1a in inducing the expression of iron uptake proteins, TfRand DMT1 and evaluating the role of KC7F2 in rendering protectionagainst iron mediated damage, similar to deferoxamine. The anti-inflammatory activity of deferoxamine and KC7F2 in hypoxicmicroglia as well as the mechanism involved in this was investi-gated with special emphasis on MAPK pathway.
2. Materials and methods
2.1. Animals
One-day-old Wistar rats were used for this study as the development of whitematter in these rats has been reported to be equivalent to the very preterm humanfoetus (Sheldon et al., 1996). A total of 56 rats were used for the in vivo study. Twentyeight rats were subjected to hypoxic exposure by placing them in a multigaschamber (Sanyo Biomedical Electrical; model MCO-18M) filled with 95% N2 and 5%O2 for 2 h. The animals were then allowed to recover under normoxic conditions for3 and 24 h, 3, 7, and 14 d before killing. Another group of 28 rats served as age-matched controls and were placed outside the chamber. For setting up primarycultures of microglia a total of 56 one-day old Wistar rats were used. All the ex-periments were approved by the Institutional Animal Care and Use Committee,National University of Singapore. All efforts were made to minimise animalsuffering, to reduce the number of animals used, and to utilise alternatives to in vivotechniques.
2.2. Primary cell culture: microglial cell culture
Mixed glial cultures were prepared using the cerebral cortices excised from 1-day-old postnatal rat brains, according to the method of Giulian and Baker (1986).In brief, the meningeal layer of the cerebral cortices were removed and made intosingle cell suspension by means of trypsinization. The dissociated cells were re-suspended in DMEM (Sigma-Aldrich) supplemented with 10% foetal calf serum(HyClone/Thermo Fisher Scientific), 10 ml/L antibioticeantimycotic solution (Sigma-Aldrich), 0.1 mM nonessential amino acids, and 1 ml/L insulin and were then seededin a 75 cm2
flask at a density of 1.2� 106 cells/ml of culture medium. The flasks wereplaced in the humidified chamber at 37 �C with 5% CO2 and 95% air. The culturemedium was replaced after 24 h and then on alternate days.
2.3. Microglial purification
Twoweeks later, following the procedure described before by Saura et al. (2003)the microglial cells were purified by mild trypsinization (0.05e0.12%) in the pres-ence of 0.2e0.5 mM EDTA. The purified microglial cells adhering to the 75 cm2
flaskwere then detached by trypsinization and seededwith a density of 1�106 cells/flaskfor protein extraction and 2.5 � 105 cells/well in a 24-well plate for immunocyto-chemistry. The plated cells were subjected to various treatments as described belowon the following day.
The rat microglial marker, OX42 [(1:100) (catalog #O-MAS 3706; Harlan-SeraLaboratory)] and nuclear marker, DAPI [(20 mg/ml; Sigma-Aldrich)] were used toassess the purity of microglial cells which was found to be w96%.
2.4. Treatment of microglial culture
For all the experiments, hypoxic induction was performed by placing the pri-mary microglial cultures in a multigas chamber (model 18M; multigas incubator;Sanyo) filled with mixture of gases containing 1% oxygen, 5% CO2 and 94% nitrogenat 37 �C for 4 h. The control cultures were simultaneously placed in an incubatorwith 95% air and 5% CO2 at 37 �C. The cultured cells were divided into 4 groups.
Group-1 cells were used to study the time dependant changes in the expressionof TfR, DMT1 and HIF-1a proteins. The cells were subjected to varying duration ofhypoxia ranging from 2 h to 6 h.
Group-II cells were used to study if HIF-1a indeed regulated the expression ofproteins involved in iron uptake such as the TfR and DMT1. For this, the cells weretreated with HIF-1a translational inhibitor KC7F2 which has been demonstrated todown regulate the genes induced by HIF1a such as carbonic anhydrase IX, matrixmetalloproteinase 2, endothelin 1, and enolase 1 (Narita et al., 2009). The concen-tration of KC7F2 used in this study was 40 mM (Narita et al., 2009).
Group III cells were used to study if MAPK pathway was involved in ironmediated production of inflammatory cytokines in microglia. The ability of defer-oxamine and KC7F2 to attenuate iron mediated inflammation and the mechanisminvolved was simultaneously analysed. The cells in this group were subdivided into6 sets- control (C), controlþ KC7F2 (Cþ K), controlþ deferoxamine (Cþ D), hypoxia(H), hypoxia þ KC7F2 (H þ K) and hypoxia þ deferoxamine (H þ D). The concen-tration of deferoxamine used was 100 mM (Zhang et al., 2006b).
Group IV cells were used to study the downstream processing of MAPK path-ways. While the mechanistic action of ERK1/2 was studied with the help of U0126(MEK1/2 inhibitor), that of p38 and JNK was studied using SB 202190 (a p38 specificinhibitor) and SP600125 (a JNK specific inhibitor), respectively. The microglial cellswere subdivided as control (C), control þ KC7F2 (C þ K), control þ deferoxamine(C þ D), control þ U0126 (C þ U), control þ SB 202190 (C þ P), control þ SP600125(C þ J), hypoxia (H), hypoxia þ KC7F2 (H þ K), hypoxia þ deferoxamine (H þ D),hypoxia þ U0126 (H þ U), hypoxia þ SB 202190 (C þ P) and hypoxia þ SP600125(C þ J). U0126, a MEK1/2 inhibitor, was used at a concentration of 20 mM (Kim et al.,2005) to explore the role of ERK. The concentrations of SB202190 (p38 inhibitor) andSP600125 (JNK inhibitor) used in this study were 5 mM (Markovic et al., 2009) and1 mM (Deng et al., 2008) respectively. In all the experiments, inhibitors and ironchelator were added to the culture medium 30 min prior to hypoxic exposure.
For double immunofluorescence studies the cells were immediately fixed in 4%paraformaldehyde following hypoxic exposure and were then processed asdescribed under double immunofluorescence section.
2.5. Iron assay
The concentration of iron accumulated in control, control þ KC7F2, hypoxia andhypoxia þ KC7F2 groups of cultured microglia was estimated with the help of Ironassay kit (Abcam, Cat. no. ab83366) following the manufacturer’s instruction. Theconcentration of iron in all four groups was calculated using the standard curveobtained.
2.6. Western blotting
Rats subjected to hypoxia were sacrificed at 3 and 24 h, 3, 7, and 14 d (n ¼ 5 ateach time point) after hypoxic exposure along with their age matched controls. ThePWM tissue was dissected out with the help of a dissection microscope from thebrains removed from the hypoxic and control groups of animals. The tissuewas snapfrozen in liquid nitrogen and was stored at �80 �C until protein extraction. The

G. Rathnasamy et al. / Neuropharmacology 77 (2014) 428e440430
tissue samples were homogenized in tissue protein extraction reagent (Pierce/Thermo Fisher Scientific; catalog #78510) containing protease inhibitors. The pro-tein was collected by means of centrifuging the homogenates at 13,000 rpm for20 min at 4 �C. To extract protein from primary microglial cultures mammalianprotein extraction reagent (Pierce/Thermo Fisher Scientific; catalog #78501) wasused. The protein concentration was estimated by Bradford’s method (Bradford,1976) using bovine serum albumin (Sigma-Aldrich) as a standard. Quantified vol-ume of samples containing 40 mg of proteinwere heated to 95 �C for 5 min and wereseparated by SDS-PAGE in 10% SDS gels, in a Mini-Protean 2 apparatus (Bio-RadLaboratories). Protein bands were electroblotted onto 0.45 mm polyvinylindenedifluoride membranes (Bio-Rad) and were blocked with 5% (w/v) nonfat dried milkovernight at 4 �C. The membranes were then incubated with dilutions of primaryantibodies listed in Table 1 followed by incubation with secondary antibodies con-jugated with horseradish peroxidase (Pierce/Thermo Fisher Scientific). Specificbinding was revealed by an enhanced chemiluminescence kit (Pierce/Thermo FisherScientific) following the manufacturer’s instructions.
2.7. Double immunofluorescence
Cellular localization of HIF-1a and DMT1 was carried out in the PWM of ratskilled at 3 days after hypoxic exposure (n¼ 3) along with their agematched controls.The rats were anesthetized using 6% sodium pentobarbital and by means of perfu-sion the brains were fixed with 2% paraformaldehyde in 0.1M phosphate buffer, pH7.4. The brains were removed and post fixed in same fixative for 4 h following whichthey were transferred to phosphate buffer solution containing 15% sucrose at 4 �Covernight. Forty mm thick brain sections containing the PWM were cut using acryostat (Leica Microsystems). To block the endogenous peroxidase activity thesections were blocked for 30 min with 0.3% hydrogen peroxide in methanol. Thesections were further washed with PBS and then incubated with cocktail of primaryantibodies [HIF-1a (Abcam, Cat no. Ab2185)/DMT1 (Santa Cruz Biotechnology, Catno. SC30120) and OX42; 1:100 dilution in PBS] at room temperature overnight.Subsequently the sections were incubated with secondary antibodies [Cy3-conjugated goat anti-rabbit IgG and FITC-conjugated sheep anti-mouse IgG(1:100; Sigma-Aldrich)] and were mounted with a fluorescent mounting medium(DAKO) after being washed in PBS. With the aid of a confocal microscope (FV1000;Olympus), the cellular localization of HIF-1a and DMT1 was assessed. For in vitrostudy primary microglial cells were fixed with 4% paraformaldehyde and the abovementioned procedure was followed. The specificity of the antibodies was confirmedby incubating a few sections with isotype control antibody rabbit anti-rat IgG (PierceBiotechnology).
MAPK signalling in the presence of ironwas analysed by fixing control, hypoxic,and hypoxia þ deferoxamine groups of microglial cultures in 4% paraformaldehyde.The fixed microglial cells were subsequently blocked with 3% normal goat serum for1 h. The cells were then incubatedwith a cocktail of primary antibodies [OX42þ ERK(1:200; Cell signalling technologies, Cat no. 4695), OX42 þ p38 (1:200; Cell sig-nalling technologies, Cat no. 9212)] overnight at 4 �C. Theywere then incubatedwitha mixture of secondary antibodies; Cy3-conjugated goat anti-rabbit IgG and FITC-conjugated sheep anti-mouse IgG (1:100; Sigma-Aldrich) and processed asdescribed above.
2.8. Statistical analysis
The data were presented as mean � SD. Statistical significance of differencesbetween control and hypoxic groupswere calculated using paired Student’s t test forin vivo studies. For in vitro experiments one-way ANOVA followed by a post-hocanalysis by Bonferroni’s Multiple Comparison Test was used to evaluate statisticalsignificance. Statistical significance between the groups was represented as
Table 1List of antibodies used for western blotting.
Antibody Dilution Cat No. Supplier
HIF-1a 1:1000 AB2185 Abcam, Pak Shek Kok, New Territories,Hong Kong
TfR 1:200 MCA155G AbD Serotec, Oxford, UKDMT1 1:1000 SC30120 Santa Cruz Biotechnology Inc., Texas, U.S.Ap-ERK 1:2000 4695 Cell signalling technologies Inc.,
Massachusetts, U.S.Ap-p38 1:500 9212 Cell signalling technologies Inc.,
Massachusetts, U.S.Ap-JNK 1:500 9258 Cell signalling technologies Inc.,
Massachusetts, U.S.Ap-CREB 1:1000 9198 Cell signalling technologies Inc.,
Massachusetts, U.S.Ap-MKP1 1:200 SC370 Santa Cruz Biotechnology Inc., Texas, U.S.ATNFa 1:500 AB1837P Millipore Bioscience, Massachusetts, U.S.AIL-1b 1:500 AB1832P Millipore Bioscience, Massachusetts, U.S.A.B actin 1:10,000 A5441 Sigma Aldrich, St. Louis, MO, U.S.A
*p < 0.05, **p < 0.01 and ***p < 0.001. With respect to cells subjected to hypoxia,statistical significance was represented as #p < 0.05, ##p < 0.01 and ###p < 0.001.
3. Results
3.1. Western blot analysis of HIF-1a and DMT1 in PWM
There were significant differences in the expression of HIF1-aand DMT1 in PWM between control and hypoxic groups. This wasevident from the immunoreactive bands for HIF1-a and DMT1which appeared at 110 kDa and 68 kDa (Fig. 1A) respectively. Whencompared to the controls, the expression of HIF-1a in hypoxic an-imals was significantly up-regulated from 3 h to 7 d (Fig. 1B). Theincrease in expression of HIF-1a in hypoxic group at 14 d was notsignificant when compared to control group. Following hypoxicexposure, the expression of DMT1 was increased until 7d and wassignificantly down-regulated at 14 d. Hence, hypoxic exposureleads to the up-regulation of the biomarkers HIF-1a and DMT1 inthe PWM of neonatal rats.
3.2. Cellular localization of HIF-1a and DMT1
By double immunofluorescence, HIF-1a and DMT1 were foundto be localized in the OX42 labelled AMC in PWM at 3 d followinghypoxic exposure. In hypoxic animals therewas an evident increasein the immunofluorescence of HIF-1a (Fig. 2Aaef) and DMT1(Fig. 2Baef) when compared to the control animals.
3.3. HIF-1a, TfR and DMT1 in microglial cultures
By means of western blotting, the changes in the expression ofHIF-1a, TfR and DMT1 were analysed in the primary microglialcultures. Immunoreactive band for HIF-1a appeared at 110 kDa(Fig. 3A) and when compared to that of the control cells, proteinexpression of HIF-1a was found to be increased significantly inmicroglia exposed to hypoxia for 2 h to 6 h (Fig. 3B). Immunore-active band for TfR appeared at 95 kDa (Fig. 3A). The expression ofTfR was found to be increased significantly in microglial cellssubjected to 4 h of hypoxia (Fig. 3C). The immunoreactive band forDMT1 appeared at 68 kDa. In parallel to that of TfR, expression ofDMT1 (Fig. 3D) was up-regulated in microglia exposed to 4 h ofhypoxia. Further, in microglial cells exposed to 4 h of hypoxia theimmunofluorescence of HIF-1a and DMT1 was found to beenhanced (Fig. 3Eaef, Faef) when compared to that of controlmicroglial cells.
3.4. HIF-1a regulates the expression of TfR and DMT1
The role of HIF-1a in mediating the expression of TfR and DMT1in hypoxic microglia was studied using western blotting. Immu-noreactive bands for TfR and DMT1 appeared at 95 kDa and 68 kDarespectively (Fig. 4A). In control microglial cells treated with KC7F2the expression of TfR was increased but was not significant,whereas expression of DMT1 was similar to that of control. Inmicroglial cells exposed to 4 h of hypoxia the expression of TfR andDMT1 was up-regulated (Fig. 4B, C). However, this increasedexpression of TfR and DMT1 was down-regulated in hypoxicmicroglial cells treated with KC7F2. Consistent with the down-regulation of TfR and DMT1, there was a significant reduction inthe iron levels in hypoxic microglia treated with KC7F2 in com-parison to those not treated with KC7F2 (Fig. 4D). From these re-sults it could be inferred that HIF-1a regulates the expression of TfRand DMT1 and thereby iron accumulation in microglia.

Fig. 1. Western blot of HIF-1a and DMT1 in the PWM of postnatal rats at 3 and 24 h, 3, 7, and 14 d after hypoxic exposure and their corresponding controls. A shows theimmunoreactive bands of HIF-1a (110 kDa), DMT1 (68 kDa), and b-actin (43 kDa). BeC are their corresponding bar graphs showing significant changes in the optical densityfollowing hypoxic exposure (given as mean � SD). The experiment was repeated five times, and a representative blot is shown here. Significant differences in protein levels betweenhypoxic and control groups are expressed as follows: *p < 0.05; **p < 0.01.
G. Rathnasamy et al. / Neuropharmacology 77 (2014) 428e440 431
3.5. Deferoxamine and KC7F2 differentially regulates MAPKsignalling pathway
The differential expression of MAP kinases (ERK, p38 and JNK) inhypoxic microglial cells in response to treatment with deferox-amine (Fig. 5A) and KC7F2 (Fig. 5B) was elucidated using westernblot. Immunoreactive bands for ERK appeared at 44/42 kDa(Fig. 5C). While hypoxia induced a significant increase in p-ERKlevels, treatment with deferoxamine and KC7F2 further accentu-ated the increase in p-ERK (Fig. 5D) when compared to that ofuntreated-hypoxic microglial cultures. The immunoreactive bandsfor p-p38 (Fig. 5C) appeared at 43 kDa. Hypoxia mediated up-regulation of p-p38 was significantly reduced in hypoxic micro-glial cells treated with deferoxamine/KC7F2 (Fig. 5E). The immu-noreactive bands for p-JNK appeared at 46/54 kDa (Fig. 5C) andthere was a significant increase in p-JNK levels in hypoxic microglia(Fig. 5F). However, the change in p-JNK levels on treatment withdeferoxamine/KC7F2 was not significant (Fig. 5F).
Immunofluorescence analysis of p-ERK (Fig. 6Aaei) and p-p38(Fig. 6Baei) in hypoxic microglia too exhibited the same phenom-enon as explained above. When compared to that of hypoxic group,
there was an enhanced immunofluorescence of p-ERK in hypoxicmicroglia treated with deferoxamine whereas that of p-p38 wasdecreased.
3.6. p-ERK mediates p-CREB up-regulation
p-ERK mediated enhanced p-CREB expression was evident withthe use of U0126 on hypoxic microglia. Western blot analysis of p-CREB (Fig. 7A) revealed a significant up-regulation in hypoxic group(Fig. 7B). In parallel to p-ERK levels, protein expression of p-CREBwas enhanced significantly in hypoxic microglia treated withdeferoxamine or KC7F2 (Fig. 7B). The increased expression of p-CREB in hypoxic microglial cultures was significantly down-regu-lated when these cultures were treated with U0126. Collectivelythese results indicate that p-ERK induces the expression of p-CREB.
3.7. p-ERK may induce MKP1 up-regulation
MKP1, which sets up the negative feedback loop to dephos-phorylate the MAP kinases may be induced via ERK pathway.Immunoreactive band for MKP-1 occurred at 40 kDa (Fig. 7A) and
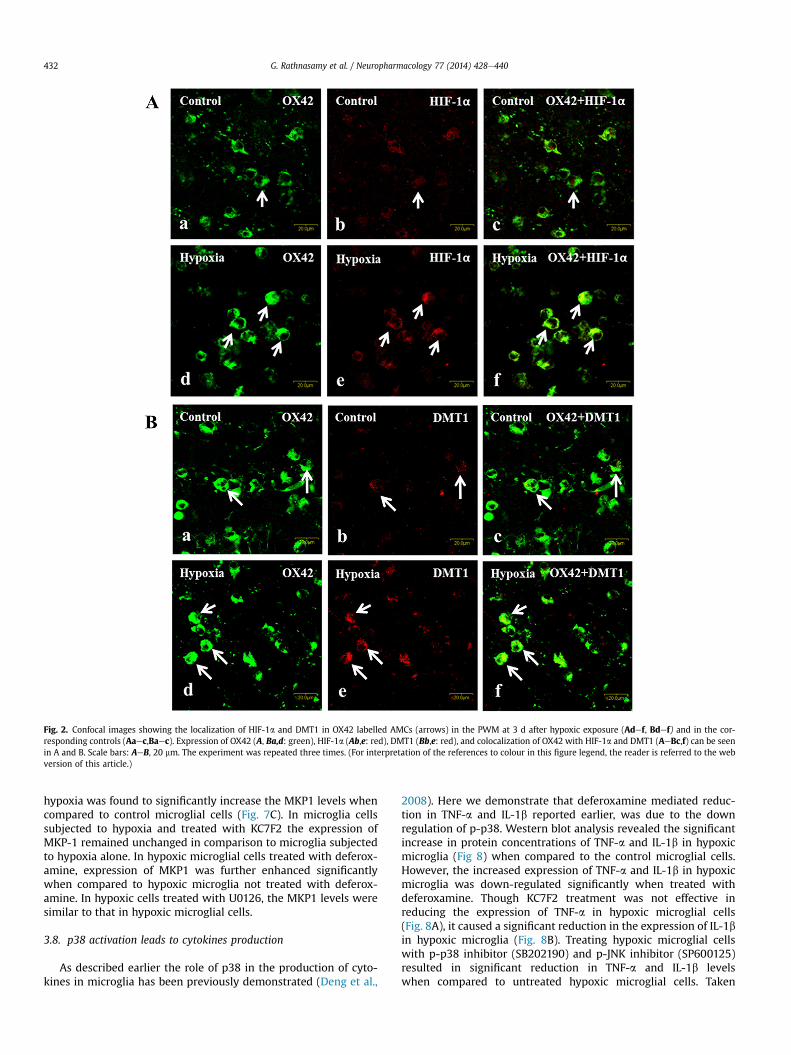
Fig. 2. Confocal images showing the localization of HIF-1a and DMT1 in OX42 labelled AMCs (arrows) in the PWM at 3 d after hypoxic exposure (Adef, Bdef) and in the cor-responding controls (Aaec,Baec). Expression of OX42 (A, Ba,d: green), HIF-1a (Ab,e: red), DMT1 (Bb,e: red), and colocalization of OX42 with HIF-1a and DMT1 (AeBc,f) can be seenin A and B. Scale bars: AeB, 20 mm. The experiment was repeated three times. (For interpretation of the references to colour in this figure legend, the reader is referred to the webversion of this article.)
G. Rathnasamy et al. / Neuropharmacology 77 (2014) 428e440432
hypoxia was found to significantly increase the MKP1 levels whencompared to control microglial cells (Fig. 7C). In microglia cellssubjected to hypoxia and treated with KC7F2 the expression ofMKP-1 remained unchanged in comparison to microglia subjectedto hypoxia alone. In hypoxic microglial cells treated with deferox-amine, expression of MKP1 was further enhanced significantlywhen compared to hypoxic microglia not treated with deferox-amine. In hypoxic cells treated with U0126, the MKP1 levels weresimilar to that in hypoxic microglial cells.
3.8. p38 activation leads to cytokines production
As described earlier the role of p38 in the production of cyto-kines in microglia has been previously demonstrated (Deng et al.,
2008). Here we demonstrate that deferoxamine mediated reduc-tion in TNF-a and IL-1b reported earlier, was due to the downregulation of p-p38. Western blot analysis revealed the significantincrease in protein concentrations of TNF-a and IL-1b in hypoxicmicroglia (Fig 8) when compared to the control microglial cells.However, the increased expression of TNF-a and IL-1b in hypoxicmicroglia was down-regulated significantly when treated withdeferoxamine. Though KC7F2 treatment was not effective inreducing the expression of TNF-a in hypoxic microglial cells(Fig. 8A), it caused a significant reduction in the expression of IL-1bin hypoxic microglia (Fig. 8B). Treating hypoxic microglial cellswith p-p38 inhibitor (SB202190) and p-JNK inhibitor (SP600125)resulted in significant reduction in TNF-a and IL-1b levelswhen compared to untreated hypoxic microglial cells. Taken
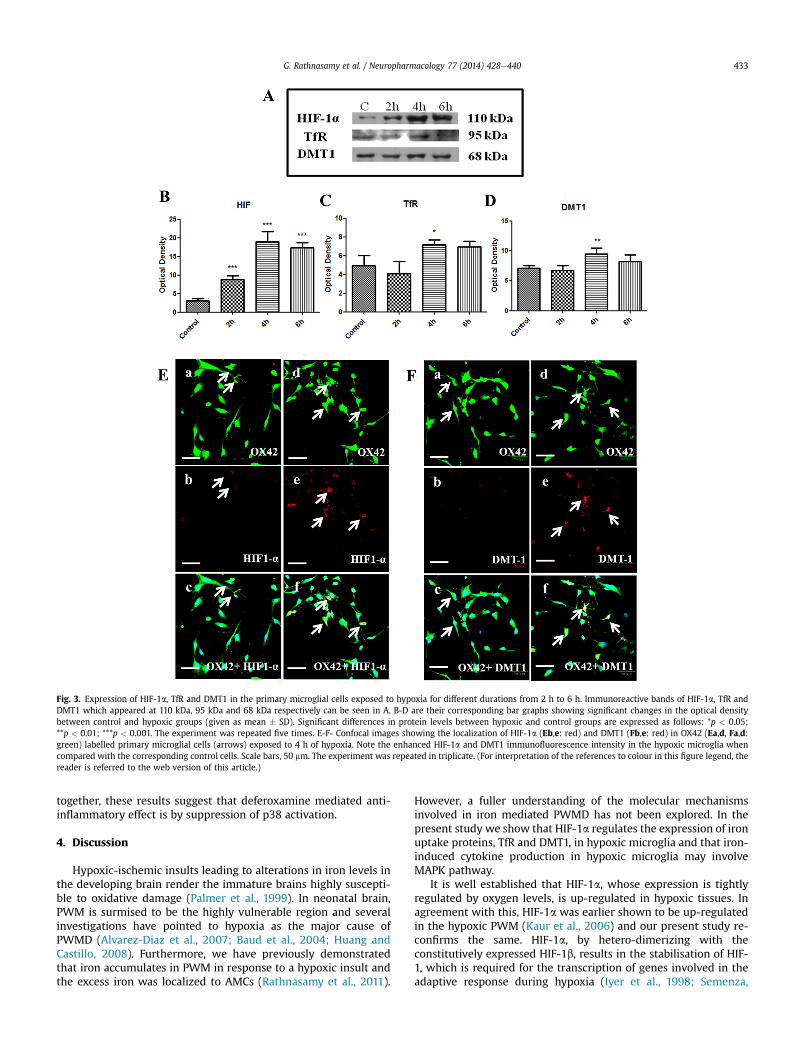
Fig. 3. Expression of HIF-1a, TfR and DMT1 in the primary microglial cells exposed to hypoxia for different durations from 2 h to 6 h. Immunoreactive bands of HIF-1a, TfR andDMT1 which appeared at 110 kDa, 95 kDa and 68 kDa respectively can be seen in A. B-D are their corresponding bar graphs showing significant changes in the optical densitybetween control and hypoxic groups (given as mean � SD). Significant differences in protein levels between hypoxic and control groups are expressed as follows: *p < 0.05;**p < 0.01; ***p < 0.001. The experiment was repeated five times. E-F- Confocal images showing the localization of HIF-1a (Eb,e: red) and DMT1 (Fb,e: red) in OX42 (Ea,d, Fa,d:green) labelled primary microglial cells (arrows) exposed to 4 h of hypoxia. Note the enhanced HIF-1a and DMT1 immunofluorescence intensity in the hypoxic microglia whencompared with the corresponding control cells. Scale bars, 50 mm. The experiment was repeated in triplicate. (For interpretation of the references to colour in this figure legend, thereader is referred to the web version of this article.)
G. Rathnasamy et al. / Neuropharmacology 77 (2014) 428e440 433
together, these results suggest that deferoxamine mediated anti-inflammatory effect is by suppression of p38 activation.
4. Discussion
Hypoxic-ischemic insults leading to alterations in iron levels inthe developing brain render the immature brains highly suscepti-ble to oxidative damage (Palmer et al., 1999). In neonatal brain,PWM is surmised to be the highly vulnerable region and severalinvestigations have pointed to hypoxia as the major cause ofPWMD (Alvarez-Diaz et al., 2007; Baud et al., 2004; Huang andCastillo, 2008). Furthermore, we have previously demonstratedthat iron accumulates in PWM in response to a hypoxic insult andthe excess iron was localized to AMCs (Rathnasamy et al., 2011).
However, a fuller understanding of the molecular mechanismsinvolved in iron mediated PWMD has not been explored. In thepresent study we show that HIF-1a regulates the expression of ironuptake proteins, TfR and DMT1, in hypoxic microglia and that iron-induced cytokine production in hypoxic microglia may involveMAPK pathway.
It is well established that HIF-1a, whose expression is tightlyregulated by oxygen levels, is up-regulated in hypoxic tissues. Inagreement with this, HIF-1a was earlier shown to be up-regulatedin the hypoxic PWM (Kaur et al., 2006) and our present study re-confirms the same. HIF-1a, by hetero-dimerizing with theconstitutively expressed HIF-1b, results in the stabilisation of HIF-1, which is required for the transcription of genes involved in theadaptive response during hypoxia (Iyer et al., 1998; Semenza,

Fig. 4. A-C Western blotting showing the changes in protein levels in TfR and DMT1 when the microglial cultures were treated with KC7F2. A shows the immunoreactive bands forTfR (95 kDa) and DMT1 (68 kDa). B,C are their corresponding bar graphs showing significant differences in all four groups (control (C), control with KC7F2 (C þ K), hypoxia (H) andhypoxia þ KC7F2 (H þ K)). Note the significant reduction in TfR and DMT1 levels in KC7F2 treated hypoxic microglia in comparison to that of untreated hypoxic microglia. Significantdifferences between various groups are expressed as follows: *p < 0.05, **p < 0.01, ***p < 0.001 with respect to control; and #p < 0.05, ##p < 0.01, ###p < 0.001 with respect tohypoxia. The experiment was repeated 5 times. Bar graph in D shows the significant difference in iron accumulation in all four groups of microglial cultures. Note the significantreduction in iron levels in hypoxic microglial group when treated with KC7F2. Significant differences between various groups are expressed as follows: *p < 0.05, **p < 0.01,***p < 0.001 with respect to control; and #p < 0.05, ##p < 0.01, ###p < 0.001 with respect to hypoxia. The experiment was repeated in triplicate.
G. Rathnasamy et al. / Neuropharmacology 77 (2014) 428e440434
1998). Expression of HIF-1a is considered to have a vital role in theneonatal brain injury induced by hypoxia (Fan et al., 2009). Inneonatal rat models of hypoxia-ischemia, inhibition of HIF-1aresulted in the reduction of brain injury (Chen et al., 2009).However, neuron specific deletion of HIF-1a in neonatal rodentshas been reported to exacerbate the injury to the brain due tohypoxic-ischemia (Sheldon et al., 2009). HIF-1a has been sug-gested to be either protective or detrimental, depending on thecell type expressing it (Vangeison et al., 2008). In the presentstudy, the increased expression of HIF-1a was localized in theAMCs of hypoxic PWM. The expression of HIF-1a in hypoxicmicroglia has been recently demonstrated to enhance theexpression of toll like receptor 4, which is implicated in neuro-inflammation (Yao et al., 2013). Besides the above, HIF-1a stabili-zation is reported to be accompanied by transcription of genesinvolved in iron metabolism (Ke and Costa, 2006). Consistent withthis, AMCs in the PWM and cultured microglia, shown here toexpress HIF-1a, were found to accumulate iron in response to ahypoxic insult (Kaur and Ling, 1999; Rathnasamy et al., 2011) andproteins involved in iron acquisition such as the iron regulatoryproteins (IRPs: IRP1 and IRP2) and transferrin receptor (TfR) werefound to be up-regulated (Rathnasamy et al., 2011). However, the
role of HIF-1a in mediating iron uptake in hypoxic microglial cellsremained to be elucidated.
In the present study, we show that the simultaneous up-regulation of HIF-1 a in AMCs exhibits control over the genesinvolved in iron uptake pathway. TfR and DMT1 are thewell-knownproteins involved in iron uptake in a cell. Owing to the increasedexpression of TfR and DMT1 in response to hypoxic insults theseproteins were identified to be hypoxia inducible (Bianchi et al.,1999; Li et al., 2008; Qian et al., 2011; Tacchini et al., 1999).Consistent with this, herewe demonstrate the increased expressionof TfR and DMT1 along with that of HIF-1a in hypoxic microglialcells. In addition, on treating hypoxic microglial cells with KC7F2the expression of TfR and DMT1 was down-regulated, suggesting arole for HIF-1a in mediating their expression. One possible expla-nation could be the presence of hypoxia responsive element (HRE)in the promoters of TfR and DMT1, which makes them a preferredtarget for HIF-1, (Bianchi et al., 1999; Qian et al., 2011; Tacchini et al.,1999). The binding of HIF-1 to the HRE sequence in the promotersof TfR and DMT1 is demonstrated to enable their transcription(Bianchi et al., 1999; Qian et al., 2011; Tacchini et al., 1999). Whilethe post-transcriptional control by IRPs is evident in leading to theirup-regulation, HIF-1a mediated regulation in the expression of

Fig. 5. A, B shows the chemical structure of iron chelator, deferoxamine and KC7F2, an inhibitor of HIF-1a. CeF: Western blotting showing significant differences in protein levels ofMAP kinases (p-ERK, p-p38 and p-JNK) in various group of primary microglial cells (control (C), control with KC7F2 (C þ K), control þ deferoxamine (C þ D), hypoxia (H),hypoxia þ KC7F2 (H þ K) and hypoxiaþ deferoxamine (Hþ D)). C shows the immunoreactive bands for p-ERK (42/44 kDa), p-p38 (43 kDa) and p-JNK (46/54 kDa). D, E, F show theirrespective bar graphs. Note the significant changes in protein levels for p-ERK and p-p38. Significant differences between the groups are expressed as follows: *p < 0.05, **p < 0.01,***p < 0.001 with respect to control; and #p < 0.05, ##p < 0.01, ###p < 0.001 with respect to hypoxia. The experiment was repeated 5 times.
G. Rathnasamy et al. / Neuropharmacology 77 (2014) 428e440 435
these proteins cannot be excluded. Furthermore, treatment withKC7F2 resulted in reduced iron accumulation in hypoxic microglialcultures in comparison to those not treated with KC7F2. Henceforthour results suggest the direct influence of HIF-1a to increase ironaccumulation in hypoxic microglia.
Activated microglia, being the major source of pro-inflammatory cytokines, are implicated in the progression ofPWMD (Deng et al., 2008). Following hypoxic exposure, microgliaare known to secrete pro-inflammatory cytokines such as TNF-aand IL-1b (Deng et al., 2008) and we, with the aid of deferoxamine,earlier reported that the excess iron accumulated in hypoxic
microglia could have a role in the enhanced production of TNF-aand IL-1b (Rathnasamy et al., 2011). Interestingly, addition of iron,externally in the form of ferrocene, to hypoxic microglial culturessignificantly increased the concentration of TNF-a when comparedto those not treated with ferrocene (Rathnasamy et al., unpub-lished). However, the mechanism through which deferoxamineattenuated microglial reaction had remained unexplained. Basedon our finding that HIF-1a has a role in the expression of iron up-take proteins, TfR and DMT1, and the fact that KC7F2 was able tosignificantly reduce the HIF-1a mediated iron accumulation, wehypothesized that KC7F2 might act against iron induced cytokine
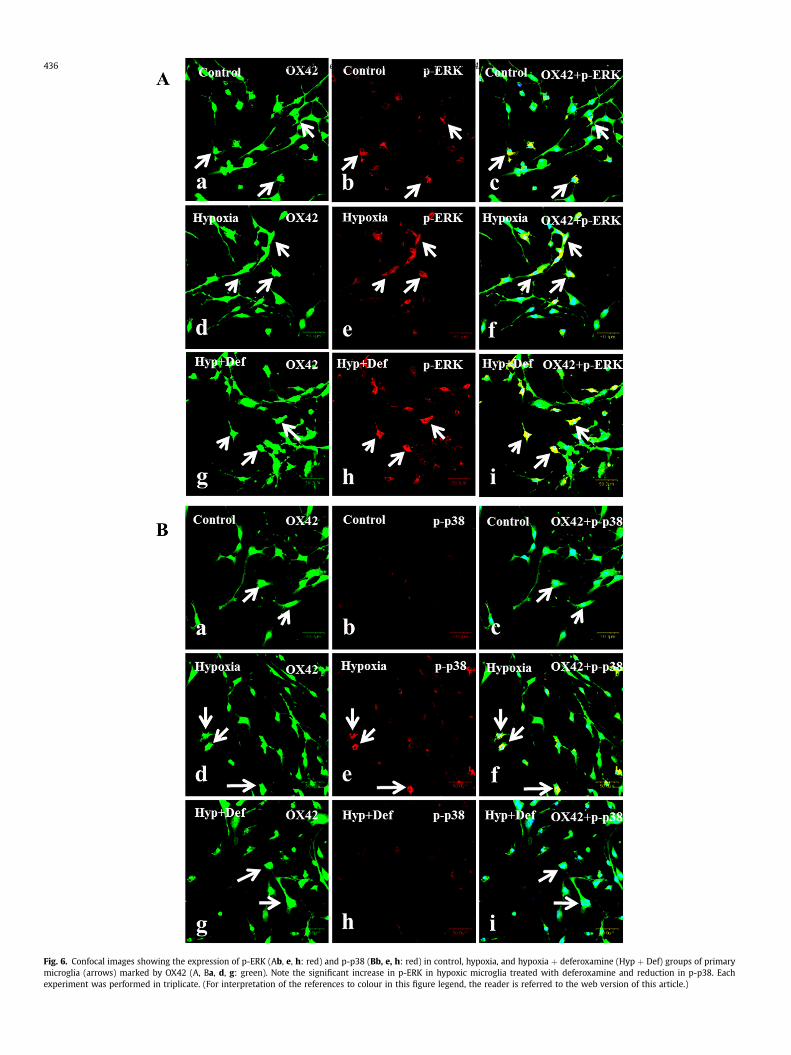
Fig. 6. Confocal images showing the expression of p-ERK (Ab, e, h: red) and p-p38 (Bb, e, h: red) in control, hypoxia, and hypoxia þ deferoxamine (Hyp þ Def) groups of primarymicroglia (arrows) marked by OX42 (A, Ba, d, g: green). Note the significant increase in p-ERK in hypoxic microglia treated with deferoxamine and reduction in p-p38. Eachexperiment was performed in triplicate. (For interpretation of the references to colour in this figure legend, the reader is referred to the web version of this article.)
G. Rathnasamy et al. / Neuropharmacology 77 (2014) 428e440436
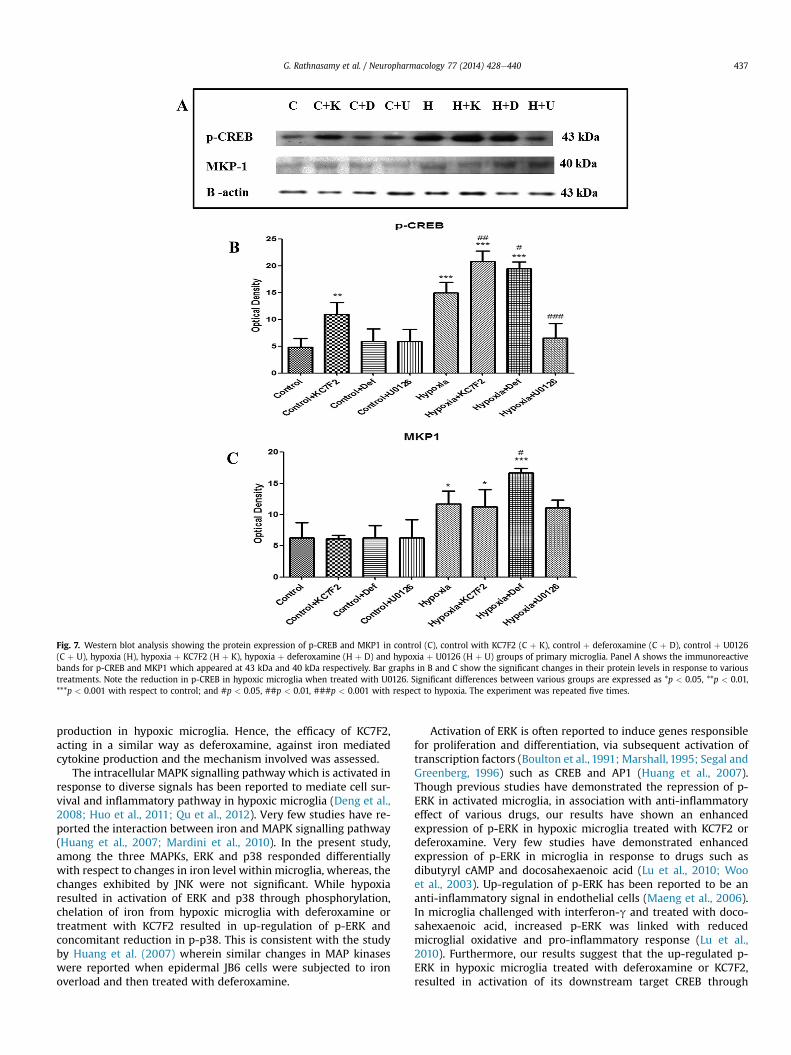
Fig. 7. Western blot analysis showing the protein expression of p-CREB and MKP1 in control (C), control with KC7F2 (C þ K), control þ deferoxamine (C þ D), control þ U0126(C þ U), hypoxia (H), hypoxia þ KC7F2 (H þ K), hypoxia þ deferoxamine (H þ D) and hypoxia þ U0126 (H þ U) groups of primary microglia. Panel A shows the immunoreactivebands for p-CREB and MKP1 which appeared at 43 kDa and 40 kDa respectively. Bar graphs in B and C show the significant changes in their protein levels in response to varioustreatments. Note the reduction in p-CREB in hypoxic microglia when treated with U0126. Significant differences between various groups are expressed as *p < 0.05, **p < 0.01,***p < 0.001 with respect to control; and #p < 0.05, ##p < 0.01, ###p < 0.001 with respect to hypoxia. The experiment was repeated five times.
G. Rathnasamy et al. / Neuropharmacology 77 (2014) 428e440 437
production in hypoxic microglia. Hence, the efficacy of KC7F2,acting in a similar way as deferoxamine, against iron mediatedcytokine production and the mechanism involved was assessed.
The intracellular MAPK signalling pathway which is activated inresponse to diverse signals has been reported to mediate cell sur-vival and inflammatory pathway in hypoxic microglia (Deng et al.,2008; Huo et al., 2011; Qu et al., 2012). Very few studies have re-ported the interaction between iron and MAPK signalling pathway(Huang et al., 2007; Mardini et al., 2010). In the present study,among the three MAPKs, ERK and p38 responded differentiallywith respect to changes in iron level within microglia, whereas, thechanges exhibited by JNK were not significant. While hypoxiaresulted in activation of ERK and p38 through phosphorylation,chelation of iron from hypoxic microglia with deferoxamine ortreatment with KC7F2 resulted in up-regulation of p-ERK andconcomitant reduction in p-p38. This is consistent with the studyby Huang et al. (2007) wherein similar changes in MAP kinaseswere reported when epidermal JB6 cells were subjected to ironoverload and then treated with deferoxamine.
Activation of ERK is often reported to induce genes responsiblefor proliferation and differentiation, via subsequent activation oftranscription factors (Boulton et al., 1991; Marshall, 1995; Segal andGreenberg, 1996) such as CREB and AP1 (Huang et al., 2007).Though previous studies have demonstrated the repression of p-ERK in activated microglia, in association with anti-inflammatoryeffect of various drugs, our results have shown an enhancedexpression of p-ERK in hypoxic microglia treated with KC7F2 ordeferoxamine. Very few studies have demonstrated enhancedexpression of p-ERK in microglia in response to drugs such asdibutyryl cAMP and docosahexaenoic acid (Lu et al., 2010; Wooet al., 2003). Up-regulation of p-ERK has been reported to be ananti-inflammatory signal in endothelial cells (Maeng et al., 2006).In microglia challenged with interferon-g and treated with doco-sahexaenoic acid, increased p-ERK was linked with reducedmicroglial oxidative and pro-inflammatory response (Lu et al.,2010). Furthermore, our results suggest that the up-regulated p-ERK in hypoxic microglia treated with deferoxamine or KC7F2,resulted in activation of its downstream target CREB through
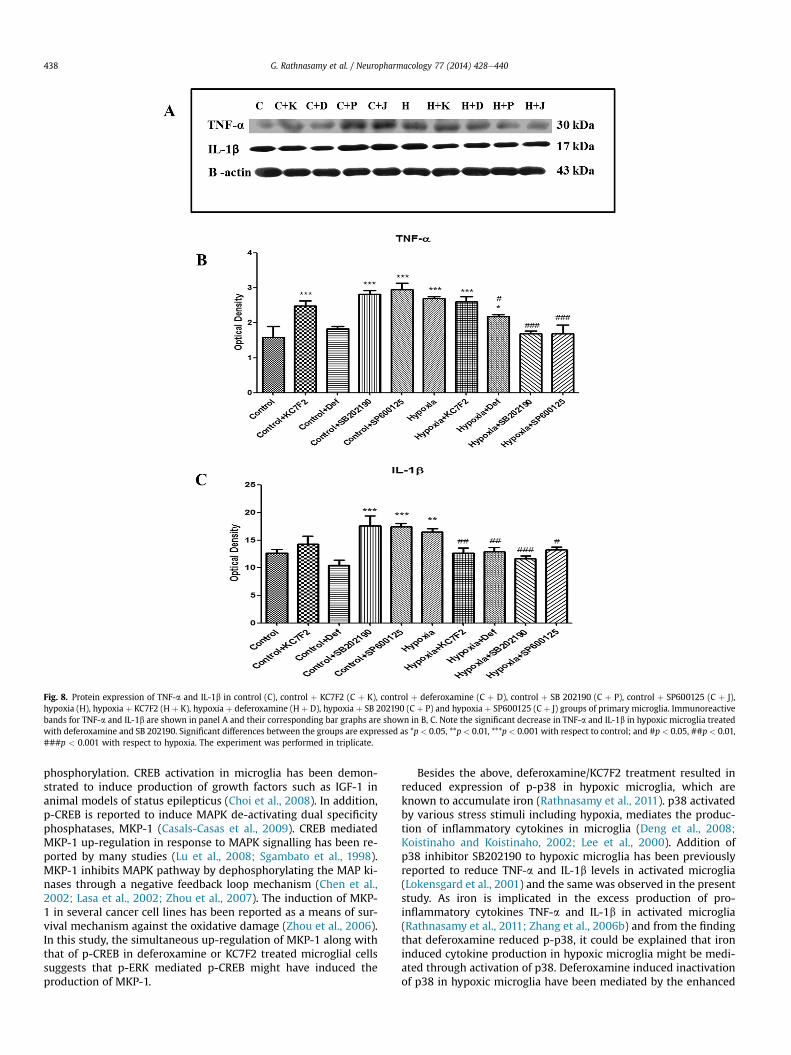
Fig. 8. Protein expression of TNF-a and IL-1b in control (C), control þ KC7F2 (C þ K), control þ deferoxamine (C þ D), control þ SB 202190 (C þ P), control þ SP600125 (C þ J),hypoxia (H), hypoxia þ KC7F2 (H þ K), hypoxia þ deferoxamine (H þ D), hypoxia þ SB 202190 (C þ P) and hypoxia þ SP600125 (C þ J) groups of primary microglia. Immunoreactivebands for TNF-a and IL-1b are shown in panel A and their corresponding bar graphs are shown in B, C. Note the significant decrease in TNF-a and IL-1b in hypoxic microglia treatedwith deferoxamine and SB 202190. Significant differences between the groups are expressed as *p < 0.05, **p < 0.01, ***p < 0.001 with respect to control; and #p < 0.05, ##p < 0.01,###p < 0.001 with respect to hypoxia. The experiment was performed in triplicate.
G. Rathnasamy et al. / Neuropharmacology 77 (2014) 428e440438
phosphorylation. CREB activation in microglia has been demon-strated to induce production of growth factors such as IGF-1 inanimal models of status epilepticus (Choi et al., 2008). In addition,p-CREB is reported to induce MAPK de-activating dual specificityphosphatases, MKP-1 (Casals-Casas et al., 2009). CREB mediatedMKP-1 up-regulation in response to MAPK signalling has been re-ported by many studies (Lu et al., 2008; Sgambato et al., 1998).MKP-1 inhibits MAPK pathway by dephosphorylating the MAP ki-nases through a negative feedback loop mechanism (Chen et al.,2002; Lasa et al., 2002; Zhou et al., 2007). The induction of MKP-1 in several cancer cell lines has been reported as a means of sur-vival mechanism against the oxidative damage (Zhou et al., 2006).In this study, the simultaneous up-regulation of MKP-1 along withthat of p-CREB in deferoxamine or KC7F2 treated microglial cellssuggests that p-ERK mediated p-CREB might have induced theproduction of MKP-1.
Besides the above, deferoxamine/KC7F2 treatment resulted inreduced expression of p-p38 in hypoxic microglia, which areknown to accumulate iron (Rathnasamy et al., 2011). p38 activatedby various stress stimuli including hypoxia, mediates the produc-tion of inflammatory cytokines in microglia (Deng et al., 2008;Koistinaho and Koistinaho, 2002; Lee et al., 2000). Addition ofp38 inhibitor SB202190 to hypoxic microglia has been previouslyreported to reduce TNF-a and IL-1b levels in activated microglia(Lokensgard et al., 2001) and the same was observed in the presentstudy. As iron is implicated in the excess production of pro-inflammatory cytokines TNF-a and IL-1b in activated microglia(Rathnasamy et al., 2011; Zhang et al., 2006b) and from the findingthat deferoxamine reduced p-p38, it could be explained that ironinduced cytokine production in hypoxic microglia might be medi-ated through activation of p38. Deferoxamine induced inactivationof p38 in hypoxic microglia have been mediated by the enhanced

G. Rathnasamy et al. / Neuropharmacology 77 (2014) 428e440 439
expression ofMKP-1, as shown in the present study. MKP-1 induceddephosphorylation of p38 in microglia has been reported by manystudies and this was attributed to the reduced inflammation (Huoet al., 2011; Ndong et al., 2012). Although, KC7F2 resulted inreduced expression of p-p38, the expression of MKP-1 remainedunchanged in hypoxic microglia treated with KC7F2 whencompared to those not treated with KC7F2. The reduction in p-p38in hypoxic microglia when treated with KC7F2 could have beenmediated either by inhibiting the upstream molecules such asMKK3/MKK6 or through the increased expression of p-CREB whichinhibits MKK3-MKK6-p38 complex formation, which is essentialfor p38 activation, as previously demonstrated by Zhang et al.(2006a). Henceforth, additional studies are required to elucidatethe mechanism by which KC7F2 suppresses p-p38. Concomitant top-p38 suppression, there was a significant reduction in theexpression of IL-1b in hypoxic microglia treated with KC7F2.Although KC7F2 was able to reduce the iron accumulation in hyp-oxic microglial cultures, its efficacy against iron induced damageand the signalling mechanisms involved requires furtherinvestigation.
The present results have shown that besides the post-transcriptional control exhibited by IRPs there exists a transcrip-tional control by HIF-1a on the proteins involved in iron uptake.While our previous investigation has led to the identification ofprotective role of deferoxamine in inhibiting iron induced pro-inflammatory cytokines secretion by hypoxic microglia, here wehave demonstrated that this was possible due to inhibition ofphosphorylation of p38. p-ERK mediated p-CREB up-regulationmight have led to the enhanced production of MKP-1, which isknown to deactivate the MAP kinases by dephosphorylating them.Although, KC7F2 treatment of hypoxic microglia resulted inincreased p-CREB when compared to hypoxic microglia not treatedwith KC7F2, the increase in MKP1 was not significant suggesting anincidental role of KC7F2 in rendering protection against ironinduced damage and warrants further investigation.
Acknowledgements
This study was supported by the research grant (R-181-000-120-213) from the National Medical Research Council, Singapore.There is no conflict of interest among the authors.
References
Adcock, L.M., Yamashita, Y., GoddardFinegold, J., Smith, C.V., 1996. Cerebral hypoxia-ischemia increases microsomal iron in newborn piglets. Metab. Brain Dis. 11,359e367.
Alvarez-Diaz, A., Hilario, E., de Cerio, F.G., Valls-i-Soler, A., Alvarez-Diaz, F.J., 2007.Hypoxic-ischemic injury in the immature brainekey vascular and cellularplayers. Neonatology 92, 227e235.
Baud, O., Daire, J.L., Dalmaz, Y., Fontaine, R.H., Krueger, R.C., Sebag, G., Evrard, P.,Gressens, P., Verney, C., 2004. Gestational hypoxia induces white matter dam-age in neonatal rats: a new model of periventricular leukomalacia. Brain Pathol.14, 1e10.
Beard, J., 2003. Iron deficiency alters brain development and functioning. J. Nutr.133, 1468Se1472S.
Bianchi, L., Tacchini, L., Cairo, G., 1999. HIF-1-mediated activation of transferrinreceptor gene transcription by iron chelation. Nucleic Acids Res. 27, 4223e4227.
Boulton, T.G., Nye, S.H., Robbins, D.J., Ip, N.Y., Radziejewska, E., Morgenbesser, S.D.,DePinho, R.A., Panayotatos, N., Cobb, M.H., Yancopoulos, G.D., 1991. ERKs: afamily of protein-serine/threonine kinases that are activated and tyrosinephosphorylated in response to insulin and NGF. Cell 65, 663e675.
Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of micro-gram quantities of protein utilizing the principle of protein-dye binding. Anal.Biochem. 72, 248e254.
Casals-Casas, C., Alvarez, E., Serra, M., de la Torre, C., Farrera, C., Sanchez-Tillo, E.,Caelles, C., Lloberas, J., Celada, A., 2009. CREB and AP-1 activation regulatesMKP-1 induction by LPS or M-CSF and their kinetics correlate with macrophageactivation versus proliferation. Eur. J. Immunol. 39, 1902e1913.
Chang, L., Karin, M., 2001. Mammalian MAP kinase signalling cascades. Nature 410,37e40.
Chang, Y.C., Huang, C.C., 2006. Perinatal brain injury and regulation of transcription.Curr. Opin. Neurol. 19, 141e147.
Chen, C., Hu, Q., Yan, J., Yang, X., Shi, X., Lei, J., Chen, L., Huang, H., Han, J., Zhang, J.H.,Zhou, C., 2009. Early inhibition of HIF-1alpha with small interfering RNA re-duces ischemic-reperfused brain injury in rats. Neurobiol. Dis. 33, 509e517.
Chen, P., Li, J., Barnes, J., Kokkonen, G.C., Lee, J.C., Liu, Y., 2002. Restraint of proin-flammatory cytokine biosynthesis by mitogen-activated protein kinasephosphatase-1 in lipopolysaccharide-stimulated macrophages. J. Immunol. 169,6408e6416.
Choi, Y.S., Cho, H.Y., Hoyt, K.R., Naegele, J.R., Obrietan, K., 2008. IGF-1 receptor-mediated ERK/MAPK signaling couples status epilepticus to progenitor cellproliferation in the subgranular layer of the dentate gyrus. Glia 56, 791e800.
Colton, C., Wilt, S., Gilbert, D., Chernyshev, O., Snell, J., Dubois-Dalcq, M., 1996.Species differences in the generation of reactive oxygen species by microglia.Mol. Chem. Neuropathol. 28, 15e20.
Deng, Y., Lu, J., Sivakumar, V., Ling, E.A., Kaur, C., 2008. Amoeboid microglia in theperiventricular white matter induce oligodendrocyte damage through expres-sion of proinflammatory cytokines via MAP kinase signaling pathway in hyp-oxic neonatal rats. Brain Pathol. 18, 387e400.
Fan, X., Heijnen, C.J., van der Kooij, M.A., Groenendaal, F., van Bel, F., 2009. Therole and regulation of hypoxia-inducible factor-1alpha expression in braindevelopment and neonatal hypoxic-ischemic brain injury. Brain Res. Rev. 62,99e108.
Follett, P.L., Deng, W., Dai, W., Talos, D.M., Massillon, L.J., Rosenberg, P.A., Volpe, J.J.,Jensen, F.E., 2004. Glutamate receptor-mediated oligodendrocyte toxicity inperiventricular leukomalacia: a protective role for topiramate. J. Neurosci. 24,4412e4420.
Giulian, D., Baker, T.J., 1986. Characterization of ameboid microglia isolated fromdeveloping mammalian brain. J. Neurosci. 6, 2163e2178.
Huang, B.Y., Castillo, M., 2008. Hypoxic-ischemic brain injury: imaging findingsfrom birth to adulthood. Radiographics 28, 417e439 quiz 617.
Huang, L.E., Arany, Z., Livingston, D.M., Bunn, H.F., 1996. Activation of hypoxia-inducible transcription factor depends primarily upon redox-sensitive stabili-zation of its alpha subunit. J. Biol. Chem. 271, 32253e32259.
Huang, X., Dai, J., Huang, C., Zhang, Q., Bhanot, O., Pelle, E., 2007. Deferoxaminesynergistically enhances iron-mediated AP-1 activation: a showcase of theinterplay between extracellular-signal-regulated kinase and tyrosine phos-phatase. Free Radic. Res. 41, 1135e1142.
Huo, Y., Rangarajan, P., Ling, E.A., Dheen, S.T., 2011. Dexamethasone inhibits theNox-dependent ROS production via suppression of MKP-1-dependent MAPKpathways in activated microglia. BMC Neurosci. 12, 49.
Iyer, N.V., Kotch, L.E., Agani, F., Leung, S.W., Laughner, E., Wenger, R.H.,Gassmann, M., Gearhart, J.D., Lawler, A.M., Yu, A.Y., Semenza, G.L., 1998. Cellularand developmental control of O2 homeostasis by hypoxia-inducible factor 1alpha. Genes Dev. 12, 149e162.
Johnston, M.V., 1997. Hypoxic and ischemic disorders of infants and children. Lec-ture for 38th Meeting of Japanese Society of Child Neurology, Tokyo, Japan, July1996. Brain Dev. 19, 235e239.
Kallio, P.J., Pongratz, I., Gradin, K., McGuire, J., Poellinger, L., 1997. Activation ofhypoxia-inducible factor 1alpha: posttranscriptional regulation and confor-mational change by recruitment of the Arnt transcription factor. Proc. Natl.Acad. Sci. U. S. A. 94, 5667e5672.
Kaur, C., Ling, E.A., 1999. Increased expression of transferrin receptors and iron inamoeboid microglial cells in postnatal rats following an exposure to hypoxia.Neurosci. Lett. 262, 183e186.
Kaur, C., Sivakumar, V., Ang, L.S., Sundaresan, A., 2006. Hypoxic damage to theperiventricular white matter in neonatal brain: role of vascular endothelialgrowth factor, nitric oxide and excitotoxicity. J. Neurochem. 98, 1200e1216.
Ke, Q., Costa, M., 2006. Hypoxia-inducible factor-1 (HIF-1). Mol. Pharmacol. 70,1469e1480.
Kim, Y.S., Kim, S.S., Cho, J.J., Choi, D.H., Hwang, O., Shin, D.H., Chun, H.S., Beal, M.F.,Joh, T.H., 2005. Matrix metalloproteinase-3: a novel signaling proteinase fromapoptotic neuronal cells that activates microglia. J. Neurosci. 25, 3701e3711.
Koistinaho, M., Koistinaho, J., 2002. Role of p38 and p44/42 mitogen-activatedprotein kinases in microglia. Glia 40, 175e183.
Lasa, M., Abraham, S.M., Boucheron, C., Saklatvala, J., Clark, A.R., 2002. Dexameth-asone causes sustained expression of mitogen-activated protein kinase (MAPK)phosphatase 1 and phosphatase-mediated inhibition of MAPK p38. Mol. CellBiol. 22, 7802e7811.
Lee, Y.B., Schrader, J.W., Kim, S.U., 2000. p38 map kinase regulates TNF-alpha pro-duction in human astrocytes and microglia by multiple mechanisms. Cytokine12, 874e880.
Li, Z., Lai, Z., Ya, K., Fang, D., Ho, Y.W., Lei, Y., Ming, Q.Z., 2008. Correlation betweenthe expression of divalent metal transporter 1 and the content of hypoxia-inducible factor-1 in hypoxic HepG2 cells. J. Cell Mol. Med. 12, 569e579.
Lokensgard, J.R., Hu, S., Sheng, W., vanOijen, M., Cox, D., Cheeran, M.C.,Peterson, P.K., 2001. Robust expression of TNF-alpha, IL-1beta, RANTES, and IP-10 by human microglial cells during nonproductive infection with herpessimplex virus. J. Neurovirol. 7, 208e219.
Lozoff, B., Georgieff, M.K., 2006. Iron deficiency and brain development. Semin.Pediatr. Neurol. 13, 158e165.
Lu, D.Y., Tsao, Y.Y., Leung, Y.M., Su, K.P., 2010. Docosahexaenoic acid suppressesneuroinflammatory responses and induces heme oxygenase-1 expression inBV-2 microglia: implications of antidepressant effects for omega-3 fatty acids.Neuropsychopharmacology 35, 2238e2248.

G. Rathnasamy et al. / Neuropharmacology 77 (2014) 428e440440
Lu, T.C., Wang, Z., Feng, X., Chuang, P., Fang, W., Chen, Y., Neves, S., Maayan, A.,Xiong, H., Liu, Y., Iyengar, R., Klotman, P.E., He, J.C., 2008. Retinoic acid utilizesCREB and USF1 in a transcriptional feed-forward loop in order to stimulateMKP1 expression in human immunodeficiency virus-infected podocytes. Mol.Cell Biol. 28, 5785e5794.
Maeng, Y.S., Min, J.K., Kim, J.H., Yamagishi, A., Mochizuki, N., Kwon, J.Y., Park, Y.W.,Kim, Y.M., Kwon, Y.G., 2006. ERK is an anti-inflammatory signal that suppressesexpression of NF-kappaB-dependent inflammatory genes by inhibiting IKKactivity in endothelial cells. Cell Signal. 18, 994e1005.
Mardini, L., Gasiorek, J., Derjuga, A., Carriere, L., Schranzhofer, M., Paw, B.H.,Ponka, P., Blank, V., 2010. Antagonistic roles of the ERK and p38 MAPK signallingpathways in globin expression, haem biosynthesis and iron uptake. Biochem. J.432, 145e151.
Markovic, D.S., Vinnakota, K., Chirasani, S., Synowitz, M., Raguet, H., Stock, K.,Sliwa, M., Lehmann, S., Kalin, R., van Rooijen, N., Holmbeck, K., Heppner, F.L.,Kiwit, J., Matyash, V., Lehnardt, S., Kaminska, B., Glass, R., Kettenmann, H., 2009.Gliomas induce and exploit microglial MT1-MMP expression for tumorexpansion. Proc. Natl. Acad. Sci. U. S. A. 106, 12530e12535.
Marshall, C.J., 1995. Specificity of receptor tyrosine kinase signaling: transientversus sustained extracellular signal-regulated kinase activation. Cell 80, 179e185.
Miao, X.Y., Liu, X.B., Yue, Q., Qiu, N., Huang, W.D., Wang, J.J., Xu, Y.G., Zhang, Y.L.,Yang, J., Chen, X.L., 2012. Deferoxamine suppresses microglia activation andprotects against secondary neural injury after intracerebral hemorrhage in rats.Nan Fang Yi Ke Da Xue Xue Bao (J. South. Med. Univ.) 32, 970e975.
Murugan, M., Sivakumar, V., Lu, J., Ling, E.A., Kaur, C., 2011. Expression of N-methylD-aspartate receptor subunits in amoeboid microglia mediates production ofnitric oxide via NF-kappaB signaling pathway and oligodendrocyte cell death inhypoxic postnatal rats. Glia 59, 521e539.
Narita, T., Yin, S., Gelin, C.F., Moreno, C.S., Yepes, M., Nicolaou, K.C., Van Meir, E.G.,2009. Identification of a novel small molecule HIF-1alpha translation inhibitor.Clin. Cancer Res. 15, 6128e6136.
Ndong, C., Landry, R.P., DeLeo, J.A., Romero-Sandoval, E.A., 2012. Mitogen activatedprotein kinase phosphatase-1 prevents the development of tactile sensitivity ina rodent model of neuropathic pain. Mol. Pain 8, 34.
Ness, J.K., Romanko, M.J., Rothstein, R.P., Wood, T.L., Levison, S.W., 2001. Perinatalhypoxia-ischemia induces apoptotic and excitotoxic death of periventricularwhite matter oligodendrocyte progenitors. Dev. Neurosci. 23, 203e208.
Palmer, C., Menzies, S.L., Roberts, R.L., Pavlick, G., Connor, J.R., 1999. Changes in ironhistochemistry after hypoxic-ischemic brain injury in the neonatal rat.J. Neurosci. Res. 56, 60e71.
Peeters-Scholte, C., Braun, K., Koster, J., Kops, N., Blomgren, K., Buonocore, G.,van Buul-Offers, S., Hagberg, H., Nicolay, K., van Bel, F., Groenendaal, F.,2003. Effects of allopurinol and deferoxamine on reperfusion injury of thebrain in newborn piglets after neonatal hypoxia-ischemia. Pediatr. Res. 54,516e522.
Qian, Z.M., Wu, X.M., Fan, M., Yang, L., Du, F., Yung, W.H., Ke, Y., 2011. Divalent metaltransporter 1 is a hypoxia-inducible gene. J. Cell Physiol. 226, 1596e1603.
Qu, W.S., Tian, D.S., Guo, Z.B., Fang, J., Zhang, Q., Yu, Z.Y., Xie, M.J., Zhang, H.Q.,Lu, J.G., Wang, W., 2012. Inhibition of EGFR/MAPK signaling reduces microglialinflammatory response and the associated secondary damage in rats afterspinal cord injury. J. Neuroinflam. 9, 178.
Rathnasamy, G., Ling, E.A., Kaur, C., 2011. Iron and iron regulatory proteins inamoeboid microglial cells are linked to oligodendrocyte death in hypoxicneonatal rat periventricular white matter through production of proin-flammatory cytokines and reactive oxygen/nitrogen species. J. Neurosci. 31,17982e17995.
Rezaie, P., Dean, A., 2002. Periventricular leukomalacia, inflammation and whitematter lesions within the developing nervous system. Neuropathology 22,106e132.
Saura, J., Tusell, J.M., Serratosa, J., 2003. High-yield isolation of murine microglia bymild trypsinization. Glia 44, 183e189.
Scott, R.J., Hegyi, L., 1997. Cell death in perinatal hypoxic-ischaemic brain injury.Neuropathol. Appl. Neurobiol. 23, 307e314.
Segal, R.A., Greenberg, M.E., 1996. Intracellular signaling pathways activated byneurotrophic factors. Annu. Rev. Neurosci. 19, 463e489.
Selim, M., 2009. Deferoxamine mesylate: a new hope for intracerebral hemorrhage:from bench to clinical trials. Stroke 40, S90eS91.
Semenza, G.L., 1998. Hypoxia-inducible factor 1: master regulator of O2 homeo-stasis. Curr. Opin. Genet. Dev. 8, 588e594.
Sgambato, V., Pages, C., Rogard, M., Besson, M.J., Caboche, J., 1998. Extracellularsignal-regulated kinase (ERK) controls immediate early gene induction oncorticostriatal stimulation. J. Neurosci. 18, 8814e8825.
Sheldon, R.A., Chuai, J., Ferriero, D.M., 1996. A rat model for hypoxic-ischemic braindamage in very premature infants. Biol. Neonate 69, 327e341.
Sheldon, R.A., Osredkar, D., Lee, C.L., Jiang, X., Mu, D., Ferriero, D.M., 2009. HIF-1alpha-deficient mice have increased brain injury after neonatal hypoxia-ischemia. Dev. Neurosci. 31, 452e458.
Skoff, R.P., Bessert, D.A., Barks, J.D., Song, D., Cerghet, M., Silverstein, F.S., 2001.Hypoxic-ischemic injury results in acute disruption of myelin gene expressionand death of oligodendroglial precursors in neonatal mice. Int. J. Dev. Neurosci.19, 197e208.
Sorond, F.A., Shaffer, M.L., Kung, A.L., Lipsitz, L.A., 2009. Desferroxamine infusionincreases cerebral blood flow: a potential association with hypoxia-induciblefactor-1. Clin. Sci. 116, 771e779.
Tacchini, L., Bianchi, L., Bernelli-Zazzera, A., Cairo, G., 1999. Transferrin receptorinduction by hypoxia. HIF-1-mediated transcriptional activation and cell-specific post-transcriptional regulation. J. Biol. Chem. 274, 24142e24146.
Takashima, S., Iida, K., Deguchi, K., 1995. Periventricular leukomalacia, glial devel-opment and myelination. Early Hum. Dev. 43, 177e184.
Vangeison, G., Carr, D., Federoff, H.J., Rempe, D.A., 2008. The good, the bad, and thecell type-specific roles of hypoxia inducible factor-1 alpha in neurons and as-trocytes. J. Neurosci. 28, 1988e1993.
Volpe, J.J., 2003. Cerebral white matter injury of the premature infant e morecommon than you think. Pediatrics 112, 176e180.
Wang, G.L., Jiang, B.H., Semenza, G.L., 1995. Effect of altered redox states onexpression and DNA-binding activity of hypoxia-inducible factor 1. Biochem.Biophys. Res. Commun. 212, 550e556.
Woo, M.S., Jang, P.G., Park, J.S., Kim, W.K., Joh, T.H., Kim, H.S., 2003. Selectivemodulation of lipopolysaccharide-stimulated cytokine expression and mitogen-activated protein kinase pathways by dibutyryl-cAMP in BV2 microglial cells.Brain Research. Mol. Brain Res. 113, 86e96.
Wu, H., Wu, T., Xu, X., Wang, J., 2011. Iron toxicity in mice with collagenase-inducedintracerebral hemorrhage. J. Cereb. Blood Flow Metab. 31, 1243e1250.
Yao, L., Kan, E.M., Lu, J., Hao, A., Dheen, S.T., Kaur, C., Ling, E.A., 2013. Toll-like re-ceptor 4 mediates microglial activation and production of inflammatory me-diators in neonatal rat brain following hypoxia: role of TLR4 in hypoxicmicroglia. J. Neuroinflam. 10, 23.
Zhang, J., Bui, T.N., Xiang, J., Lin, A., 2006a. Cyclic AMP inhibits p38 activation viaCREB-induced dynein light chain. Mol. Cell Biol. 26, 1223e1234.
Zhang, X., Haaf, M., Todorich, B., Grosstephan, E., Schieremberg, H., Surguladze, N.,Connor, J.R., 2005. Cytokine toxicity to oligodendrocyte precursors is mediatedby iron. Glia 52, 199e208.
Zhang, X., Surguladze, N., Slagle-Webb, B., Cozzi, A., Connor, J.R., 2006b. Cellular ironstatus influences the functional relationship between microglia and oligoden-drocytes. Glia 54, 795e804.
Zhou, J.Y., Liu, Y., Wu, G.S., 2006. The role of mitogen-activated protein kinasephosphatase-1 in oxidative damage-induced cell death. Cancer Res. 66, 4888e4894.
Zhou, Y., Ling, E.A., Dheen, S.T., 2007. Dexamethasone suppresses monocyte che-moattractant protein-1 production via mitogen activated protein kinasephosphatase-1 dependent inhibition of Jun N-terminal kinase and p38mitogen-activated protein kinase in activated rat microglia. J. Neurochem. 102,667e678.