hydrogen peroxide-induced oxidative damage and apoptosis in
TRANSCRIPT

Pharmacological Research, Vol. 41, No. 4, 2000doi:10.1006rphrs.1999.0604, available online at http:rrwww.idealibrary.com on
HYDROGEN PEROXIDE-INDUCED OXIDATIVE DAMAGE ANDAPOPTOSIS IN CEREBELLAR GRANULE CELLS: PROTECTION BY
GINKGO BILOBA EXTRACT
TAOTAO WEI, YUCHENG NI, JINGWU HOU, CHANG CHEN, BAOLU ZHAOand WENJUAN XINU
Institute of Biophysics, Academia Sinica, 15 Datun Road, Chaoyang District, Beijing 100101,PR China
Accepted 23 September 1999
The ability of oxidative stress to induce apoptosis and the protective effects of GinkgoŽ .biloba extract EGb761 against this induction were studied in cultures of rat cerebellar
granule cells. Cells were exposed to oxidative stress by treatment with 50 mM hydrogenperoxideq100 mM ferrous sulphate which generates hydroxyl radicals by Fenton reaction.Both morphological observation and biochemical analysis revealed that H O rFeSO2 2 4treatment induced apoptotic cell death in cerebellar granule cells, which was characterizedby chromatin condensation and DNA fragmentation. During this process, the fluidity of thecell membrane decreased markedly, and the conformation of membrane proteins altered
Žsignificantly. Pretreating cerebellar granule cells with the antioxidant EGb761 Ginkgo.biloba extract effectively attenuated oxidative damage induced by H O rFeSO , and2 2 4
prevented cells from apoptotic cell death. The results suggested that EGb761 might be usedas a potential drug for neuronal diseases associated with the excessive production ofreactive oxygen species. Q 2000 Academic Press
KEY WORDS: apoptosis, cerebellar granule cells, EGb761, lipid peroxidation, ESR spin labelling.
INTRODUCTION
In multicellular organisms homeostasis is main-tained by the dynamic balance between cell prolifer-ation and cell death. Two types of cell death, apop-tosis and necrosis, may occur under certain physio-logical and pathological conditions. Physiologicalapoptosis plays important roles in the development
Ž . w xof the central nervous system CNS 1 , while inap-propriate apoptosis of neuronal cells leads to someneurodegenerative diseases including Alzheimer’s
Ž . Ž .disease AD , Parkinson’s disease PD , amyotrophicŽ .lateral sclerosis ALS , and various forms of cerebel-
w xlar degeneration 2 . In the last decade more andmore evidence has suggested that reactive oxygen
Ž .species ROS play an important role in the cascadeof events leading to neuronal apoptosis associated
w xwith these neuronal diseases 3 . Accordingly, antiox-idants, which protect neuronal cells from ROS-induced apoptosis, may be used as potential thera-peutic agents for these diseases.
U Corresponding author.
1043]6618r00r040427]07r$35.00r0
In this paper, a primary culture of rat cerebellargranule cells, a relatively homogenous and well-
w xcharacterized culture system of neurons 4 , wasused as the experimental model. The cytotoxic effectof oxidative stress and the protective effect ofEGb761 on neuronal cells were examined. The re-sults will help us to understand the process of neuro-nal apoptosis and neurodegeneration morethoroughly, which will lead to selective methods tocontrol them.
MATERIALS AND METHODS
MaterialsWistar rats were purchased from Beijing Medical
Ž .University Beijing, China . Cell culture plastic wareŽwas purchased from Corning Costar Acton, MA,
. Ž .USA . Dulbecco’s modified Eagle medium DMEM ,Ž .foetal bovine serum FBS , cell culture supplements,
Ž .proteinase K, RNase A, and trypsin 1:250 wereŽ .products of Gibco BRL Grand Island, NY, USA .
Ž .Agarose, poly-L-lysine, thiazolyl blue MTT , sodiumŽ .dodecyl sulphate SDS , sodium N-lauroylsarcosine,
Q 2000 Academic Press

Pharmacological Research, Vol. 41, No. 4, 2000428
Ž . Žin 500 ml lysis buffer 10 mM Tris, 10 mM EDTA,
Triton X-100, 1,1,3,3-tetraethoxypropane TEP , 5-Ž .doxyl stearic acid 5-doxyl , 16-doxyl stearic acidŽ . Ž .16-doxyl , and 3-maleimido-proxyl 3-mal were pur-Ž .chased from Sigma St. Louis, MO, USA . EGb761
Ž .was a generous gift from IPSEN Paris, France .Other reagents made in China were of analyticalgrade.
Cell culturePrimary cultures of rat cerebellar granule cells
were prepared following procedures described previ-w xously 5 . Briefly, cerebella from 7-day-old Wistar
rats were dissected out, rinsed with HBSS, and dis-sociated by mild trypsinization. Cells were plated on
Ž 6 y1 y1.six-well multidishes 2=10 cells ml , 2 ml wellŽ 6 y1or 24-well multidishes 2.5=10 cells ml , 0.4 ml
y1 .well previously coated with poly-L-lysine. Culturemedium consisted of DMEM supplemented with
Ž . Ž . Ž .KCl 19.6 mM , glutamine 2 mM , HEPES 10 mMŽ .and foetal bovine serum 10%, vrv . Cells were
maintained at 378C in a humidified 5% CO ]95%2air atmosphere. Experiments were carried out 48 hafter plating.
Treatment of cells with H O rFeSO2 2 4Cell culture medium was changed into fresh
Ž .DMEM containing a low concentration of FBS 1%and cells were cultured for 8 h. Then, 50 mM H O2 2and 100 mM FeSO were added into cells from4freshly prepared stock solutions and cells were cul-tured for an indicated time. The antioxidant EGb761was added to cells from freshly prepared stock solu-
Ž y1tion 1 mg ml in DMEM, sterilized upon filtra-.tion 15 min before the treatment with H O r2 2
FeSO .4
Assessment of neurotoxicityw xCell viability was assessed by the MTT assay 6 .
Ž y1 .MTT 0.5 mg ml , final concentration was addedto cells cultured in 24-well multidishes and in-cubated at 378C for 30 min. Then 1 ml of lysis
Ž .solution 10% SDS, 25% DMF, pH 3.5 was addedand the optical density at 570 nm was measured.
Morphological studiesThe ultrastructure of cells was observed by trans-
w xmission electron microscopy 7 . Briefly, cells werefixed with 2.5% glutaraldehyde at 48C for 1 h andpost-fixed with 1% OsO at 48C for 1 h, dehydrated4through a series of graded ethanol solutions, andembedded in resin. Ultrathin sections of sampleswere stained with uranyl acetaterlead citrate andobserved with a transmission electron microscope.
Detection of DNA fragmentationThe laddering pattern of DNA fragmentation was
w xdetected by agarose gel electrophoresis 8 . Briefly,1.2=107 cells were harvested by centrifugation at200 g for 10 min, washed twice with PBS, and lysed
.0.5% sodium N-lauroylsarcosine, pH 8.0 . Aftertreatment with 0.5 mg mly1 proteinase Kq0.5 mgmly1 RNase A at 508C for 3 h, the lysate wasextracted with phenol, and the DNA was precipi-tated with ethanol and ammonium acetate. After
Žbeing dissolved in Tris]EDTA buffer 10 mM Tris, 1.mM EDTA, pH 8.0 , DNA was analysed by 1.5%
agarose gel electrophoresis.The cytosolic DNA fragments were quantified by
w x 7the diphenylamine method 9 . Briefly, 1.2=10 cellsŽwere lysed in 500 ml lysis buffer 10 mM Tris, 10 mM
.EDTA, 0.5% Triton X-100, pH 7.4 on ice for 20min. The lysate was centrifuged at 12,000 g for 20
Ž .min to separate the intact chromatin the pellet andŽ .the cytosolic DNA fragments the supernatant . Both
the pellet and the supernatant were treated with 100ml of 6% perchloric acid at 48C for 30 min. Theprecipitates were sendimented at 12,000 g for 20min. The DNA precipitates were heated at 708C for20 min in 100 ml 6% perchloric acid, and were
Žmixed with 200 ml of diphenylamine solution 1.5%diphenylamine, 1.5% sulphuric acid, and 0.01% ac-
.etaldehyde in glacial acetic acid . After overnightincubation at 308C in the dark, the optical density at600 nm was measured, and the percentage of DNAfragmentation was calculated as the ratio of DNA inthe supernatant to the total DNA.
Measurement of lipid peroxidationLipid peroxidation in cells was measured by
Ž .thiobarbituric acid TBA assay as described previ-w x 7ously 10 . Briefly, 1.2=10 cells were mixed with 0.4
ml of 2.8% trichloroacetic acid and 1 ml of 0.67%thiobarbituric acid, heated at 958C for 1 h, extractedwith butanol, and the optical density of the organiclayer was determined spectrophotometrically. TEPwas used as a standard. The protein concentrationwas determined by the Bradford method.
Spin labellingThe fluidity of the cell membrane was determined
by ESR using 5-doxyl and 16-doxyl as spin labels.The conformation alteration of cell membrane pro-tein sulphydryl groups was determined by ESR using
w x3-mal as spin label 11 . Briefly, cells were washedthree times with PBS, mixed with 10 mM spin labelŽ . Ž .5-doxyl, 16-doxyl or 100 mM spin label 3-mal and
Ž .incubated at 378C for 30 min 5-doxyl, 16-doxyl or 3Ž .h 3-mal . The labelled cells were washed four times
with PBS, and then transferred into quartz capillar-ies for ESR measurement. The ESR spectra were
Ž .recorded at room temperature 298 K by a VarianE-109 spectrometer with measurement conditionsas: X-band, central magnetic field 325 mT, sweepwidth 20 mT, microwave power 20 mW, frequency100 kHz, modulation amplitude 0.2 mT, time con-stant 0.128 s. The methods of calculation for the
Ž .order parameter S , the rotational correlation time
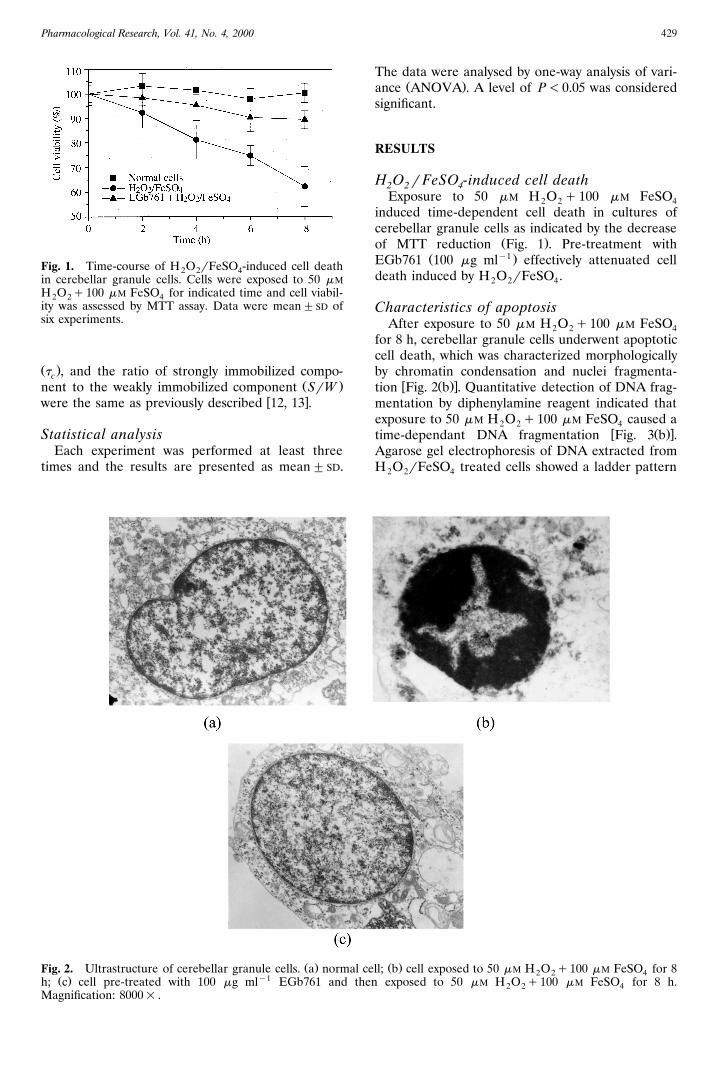
Pharmacological Research, Vol. 41, No. 4, 2000 429
Fig. 1. Time-course of H O rFeSO -induced cell death2 2 4in cerebellar granule cells. Cells were exposed to 50 mMH O q100 mM FeSO for indicated time and cell viabil-2 2 4ity was assessed by MTT assay. Data were mean"SD ofsix experiments.
Ž .t , and the ratio of strongly immobilized compo-cŽ .nent to the weakly immobilized component SrW
w xwere the same as previously described 12, 13 .
Statistical analysisEach experiment was performed at least three
times and the results are presented as mean"SD.
Ž .Fig. 2. Ultrastructure of cerebellar granule cells. a normalŽ . y1h; c cell pre-treated with 100 mg ml EGb761 and t
Magnification: 8000= .
The data were analysed by one-way analysis of vari-Ž .ance ANOVA . A level of P-0.05 was considered
significant.
RESULTS
H O rFeSO -induced cell death2 2 4Exposure to 50 mM H O q100 mM FeSO2 2 4
induced time-dependent cell death in cultures ofcerebellar granule cells as indicated by the decrease
Ž .of MTT reduction Fig. 1 . Pre-treatment withŽ y1 .EGb761 100 mg ml effectively attenuated cell
death induced by H O rFeSO .2 2 4
Characteristics of apoptosisAfter exposure to 50 mM H O q100 mM FeSO2 2 4
for 8 h, cerebellar granule cells underwent apoptoticcell death, which was characterized morphologicallyby chromatin condensation and nuclei fragmenta-
w Ž .xtion Fig. 2 b . Quantitative detection of DNA frag-mentation by diphenylamine reagent indicated thatexposure to 50 mM H O q100 mM FeSO caused a2 2 4
w Ž .xtime-dependant DNA fragmentation Fig. 3 b .Agarose gel electrophoresis of DNA extracted fromH O rFeSO treated cells showed a ladder pattern2 2 4
Ž .cell; b cell exposed to 50 mM H O q100 mM FeSO for 82 2 4hen exposed to 50 mM H O q100 mM FeSO for 8 h.2 2 4
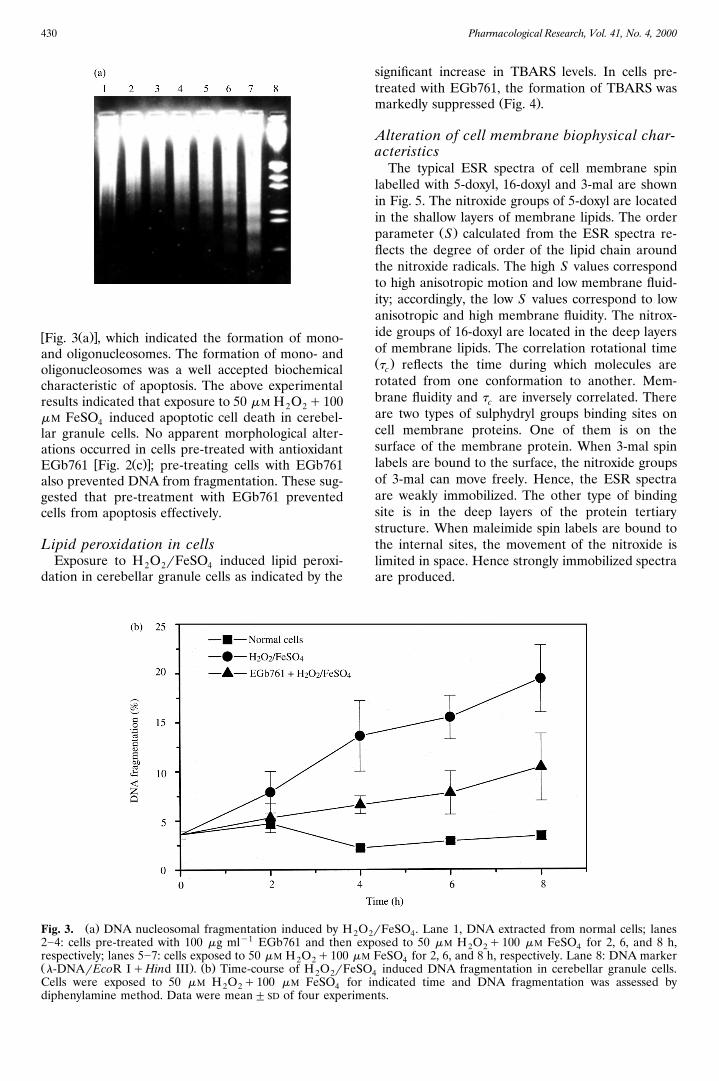
Pharmacological Research, Vol. 41, No. 4, 2000430
w Ž .xFig. 3 a , which indicated the formation of mono-and oligonucleosomes. The formation of mono- andoligonucleosomes was a well accepted biochemicalcharacteristic of apoptosis. The above experimentalresults indicated that exposure to 50 mM H O q1002 2mM FeSO induced apoptotic cell death in cerebel-4lar granule cells. No apparent morphological alter-ations occurred in cells pre-treated with antioxidant
w Ž .xEGb761 Fig. 2 c ; pre-treating cells with EGb761also prevented DNA from fragmentation. These sug-gested that pre-treatment with EGb761 preventedcells from apoptosis effectively.
Lipid peroxidation in cellsExposure to H O rFeSO induced lipid peroxi-2 2 4
dation in cerebellar granule cells as indicated by the
Ž .Fig. 3. a DNA nucleosomal fragmentation induced by H O22]4: cells pre-treated with 100 mg mly1 EGb761 and then erespectively; lanes 5]7: cells exposed to 50 mM H O q100 m2 2Ž . Ž .l-DNArEcoR IqHind III . b Time-course of H O rFeS2 2Cells were exposed to 50 mM H O q100 mM FeSO for2 2 4diphenylamine method. Data were mean"SD of four experim
significant increase in TBARS levels. In cells pre-treated with EGb761, the formation of TBARS was
Ž .markedly suppressed Fig. 4 .
Alteration of cell membrane biophysical char-acteristics
The typical ESR spectra of cell membrane spinlabelled with 5-doxyl, 16-doxyl and 3-mal are shownin Fig. 5. The nitroxide groups of 5-doxyl are locatedin the shallow layers of membrane lipids. The order
Ž .parameter S calculated from the ESR spectra re-flects the degree of order of the lipid chain aroundthe nitroxide radicals. The high S values correspondto high anisotropic motion and low membrane fluid-ity; accordingly, the low S values correspond to lowanisotropic and high membrane fluidity. The nitrox-ide groups of 16-doxyl are located in the deep layersof membrane lipids. The correlation rotational timeŽ .t reflects the time during which molecules arec
rotated from one conformation to another. Mem-brane fluidity and t are inversely correlated. Therec
are two types of sulphydryl groups binding sites oncell membrane proteins. One of them is on thesurface of the membrane protein. When 3-mal spinlabels are bound to the surface, the nitroxide groupsof 3-mal can move freely. Hence, the ESR spectraare weakly immobilized. The other type of bindingsite is in the deep layers of the protein tertiarystructure. When maleimide spin labels are bound tothe internal sites, the movement of the nitroxide islimited in space. Hence strongly immobilized spectraare produced.
rFeSO . Lane 1, DNA extracted from normal cells; lanes2 4xposed to 50 mM H O q100 mM FeSO for 2, 6, and 8 h,2 2 4M FeSO for 2, 6, and 8 h, respectively. Lane 8: DNA marker4O induced DNA fragmentation in cerebellar granule cells.4indicated time and DNA fragmentation was assessed by
ents.
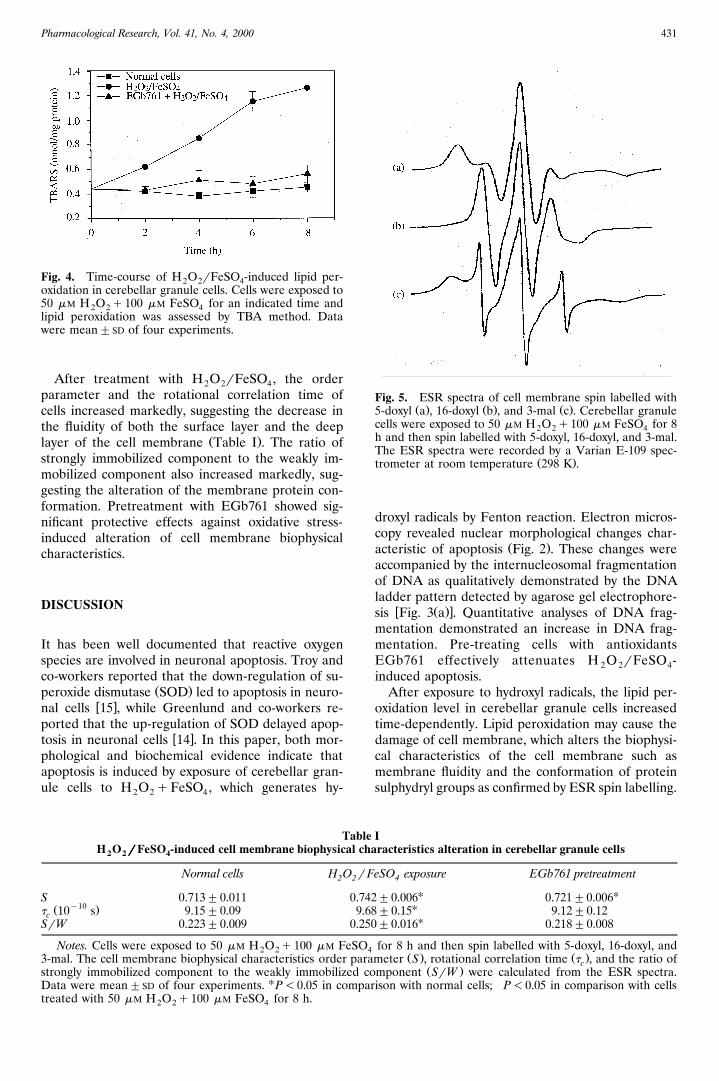
Pharmacological Research, Vol. 41, No. 4, 2000 431
Fig. 4. Time-course of H O rFeSO -induced lipid per-2 2 4oxidation in cerebellar granule cells. Cells were exposed to50 mM H O q100 mM FeSO for an indicated time and2 2 4lipid peroxidation was assessed by TBA method. Datawere mean"SD of four experiments.
After treatment with H O rFeSO , the order2 2 4parameter and the rotational correlation time ofcells increased markedly, suggesting the decrease inthe fluidity of both the surface layer and the deep
Ž .layer of the cell membrane Table I . The ratio ofstrongly immobilized component to the weakly im-mobilized component also increased markedly, sug-gesting the alteration of the membrane protein con-formation. Pretreatment with EGb761 showed sig-nificant protective effects against oxidative stress-induced alteration of cell membrane biophysicalcharacteristics.
DISCUSSION
It has been well documented that reactive oxygenspecies are involved in neuronal apoptosis. Troy andco-workers reported that the down-regulation of su-
Ž .peroxide dismutase SOD led to apoptosis in neuro-w xnal cells 15 , while Greenlund and co-workers re-
ported that the up-regulation of SOD delayed apop-w xtosis in neuronal cells 14 . In this paper, both mor-
phological and biochemical evidence indicate thatapoptosis is induced by exposure of cerebellar gran-ule cells to H O qFeSO , which generates hy-2 2 4
TabH O rrrrrFeSO -induced cell membrane biophysical c2 2 4
Normal cells H O r2 2
S 0.713"0.011 0.7y10Ž .t 10 s 9.15"0.09 9c
SrW 0.223"0.009 0.2
Notes. Cells were exposed to 50 mM H O q100 mM FeSO2 23-mal. The cell membrane biophysical characteristics order pastrongly immobilized component to the weakly immobilizedData were mean"SD of four experiments. UP-0.05 in comptreated with 50 mM H O q100 mM FeSO for 8 h.2 2 4
Fig. 5. ESR spectra of cell membrane spin labelled withŽ . Ž . Ž .5-doxyl a , 16-doxyl b , and 3-mal c . Cerebellar granule
cells were exposed to 50 mM H O q100 mM FeSO for 82 2 4h and then spin labelled with 5-doxyl, 16-doxyl, and 3-mal.The ESR spectra were recorded by a Varian E-109 spec-
Ž .trometer at room temperature 298 K .
droxyl radicals by Fenton reaction. Electron micros-copy revealed nuclear morphological changes char-
Ž .acteristic of apoptosis Fig. 2 . These changes wereaccompanied by the internucleosomal fragmentationof DNA as qualitatively demonstrated by the DNAladder pattern detected by agarose gel electrophore-
w Ž .xsis Fig. 3 a . Quantitative analyses of DNA frag-mentation demonstrated an increase in DNA frag-mentation. Pre-treating cells with antioxidantsEGb761 effectively attenuates H O rFeSO -2 2 4induced apoptosis.
After exposure to hydroxyl radicals, the lipid per-oxidation level in cerebellar granule cells increasedtime-dependently. Lipid peroxidation may cause thedamage of cell membrane, which alters the biophysi-cal characteristics of the cell membrane such asmembrane fluidity and the conformation of proteinsulphydryl groups as confirmed by ESR spin labelling.
le Iharacteristics alteration in cerebellar granule cells
FeSO exposure EGb761 pretreatment4
U U42"0.006 0.721"0.006U.68"0.15 9.12"0.12†
U50"0.016 0.218"0.008†
for 8 h and then spin labelled with 5-doxyl, 16-doxyl, and4Ž . Ž .rameter S , rotational correlation time t , and the ratio ofc
Ž .component SrW were calculated from the ESR spectra.arison with normal cells; †P-0.05 in comparison with cells

Pharmacological Research, Vol. 41, No. 4, 2000432
Fig. 6. Chemical structures of typical flavonoids in EGb761.
Lipid peroxidation may also damage the receptors ated from H O and FeSO . The protective effect of
w xand ion channels on the cell membrane 16 , whichmay result in more serious consequences includingcalcium influx, and cause cell death finally. On theother hand, unpublished data of our laboratory de-monstrated that H O rFeSO treatment could2 2 4down-regulate the anti-apoptotic gene bcl-2 and up-regulate the apoptotic gene p53. H O r2 2FeSO might induce apoptosis in neuronal cells by4regulating the expression of these apoptosis-relatedgenes.
Pre-treating cells with EGb761 attenuates oxida-tive stress-induced alteration of cell membrane bio-physical characteristics and prevented the cells fromapoptosis effectively. EGb761 is a complex mixturew x17 , which contains flavonoid and non-flavonoidcomponents. The main chemical structures thatflavonoid components possess are aromatic rings
Ž .and double bonds Fig. 6 , so flavonoid componentsprefer to react with hydroxyl radicals to form anadditional product, and thus scavenge the hydroxylradicals directly. On the other hand, flavonoid com-ponents of EGb761 also possess phenolic hydroxylgroups, which may chelate the ferrous cation anddecrease the formation of hydroxyl radicals gener-
2 2 4EGb761 on apoptosis may be the effect of bothscavenging of hydroxyl radicals directly and chelat-ing iron to inhibit Fenton reaction indirectly. On theother hand, EGb761 could up-regulate the activityof SOD, the key antioxidant enzyme in the brainw x18 , and thus enhance the antioxidant capacity ofthe neuronal system.
In conclusion, exposure of cerebellar granule cellsto oxidative stress triggers apoptosis, and pre-treat-ment with EGb761 could effectively have preventedcells from apoptosis.
REFERENCES
1. Henderson CE. Programmed cell death in the devel-oping nervous system. Neuron 1996; 17: 579]85.
2. Thompson CB. Apoptosis in the pathogenesis andtreatment of disease. Science 1995; 267: 1456]62.
3. Jacobson MD. Reactive oxygen species and pro-grammed cell death. Trends Biochem Sci 1996; 21:83]6.
4. Dudek H, Datta SR, Franke TF, Birnbaum MJ, YaoR, Cooper GM, Segel RA, Kaplan DR, GreenbergME. Regulation of neuronal survival by the serine-

Pharmacological Research, Vol. 41, No. 4, 2000 433
threonine protein kinase Akt. Science 1997; 275: 12. Xin WJ, Zhao BL, Zhang JZ. Studies on the property
661]5.5. Resink A, Hack N, Boer GJ, Balazs R. Growthconditions differentially modulate the vulnerability ofdeveloping cerebellar granule cells to excitatoryamino acids. Brain Res 1994; 655: 222]32.
6. Wei TT, Chen C, Zhao BL, Xin WJ, Mori A. EPC-K1attenuates peroxynitrite-induced apoptosis in cere-bellar granule cells. Biochem Mol Biol Int 1998; 46:89]97.
7. Watt JA, Pike CJ, Walencewicz-Wasserman AJ, Cot-man CW. Ultrastructural analysis of b-amyloid-in-duced apoptosis in cultured hippo-campal neurons.Brain Res 1994; 661: 147]56.
8. Palluy O, Rigaud M. Nitric oxide induces culturedcortical neuron apoptosis. Neurosci Lett 1996; 208:1]4.
9. Yabuki M, Inai Y, Yoshioka T, Hamazaki K, YasudaT, Inoue M, Utsumi K. Oxygen-dependent fragmen-tation of cellular DNA by nitric oxide. Free Radic Res1997; 26: 245]55.
10. Salgo MG, Pryor WA. Trolox inhibits peroxynitrite-mediated oxidative stress and apoptosis in rat thymo-cytes. Arch Biochem Biophys 1996; 333: 482]8.
11. Shi HL, Zhao BL, Xin WJ. Scavenging effects ofbaicalin on free radicals and its protection on ery-throcyte membrane from free radical injury. BiochemMol Biol Int 1995; 35: 981]94.
of sulfhydryl binding site on the lung normal andcancer cell membrane of Chinese hamster with
( )maleimide spin labels. Sci Sin Series B 1984; 27:1008]14.
13. Zhao BL, Zhang QC, Zhang JZ, Xin WJ. Studies onthe membrane fluidity of the lung cell of Chinesehamster with fatty acid spin labels. Kexue Tongbao1983; 28: 392]6.
14. Greenlund LJS, Deckwerth TL, Johnson EM Jr. Su-peroxide dismutase delays neuronal apoptosis: a rolefor reactive oxygen species in programmed cell death.Neuron 1995; 14: 303]15.
15. Troy CM, Derossi D, Prochiantz A, Greene LA,Shelanski ML. Downregulation of CurZn superoxidedismutase leads to cell death via the nitric oxide]per-oxynitrite pathway. J Neurosci 1996; 16: 253]61.
16. Mattson MP. Modification of ion homeostasis bylipid peroxidation: roles in neuronal degenerationand adaptive plasticity. Trends Neurosci 1998; 21:53]7.
Ž .17. DeFeudis FV. Ginkgo biloba extract EGb761 : phar-macological activities and clinical applications. Paris:Elsevier, 1991.
18. Seif-El-Nasr M, El-Fattah AA. Lipid peroxide, phos-pholipids, glutathione levels and superoxide dismu-tase activity in rat brain after ischemia: effect ofginkgo biloba extract. Pharmacol Res 1995; 32: 273]8.