hsp27regulatesaktactivationandpolymorphonuclear ... leukocyteapoptosisbyscaffoldingmk2toakt...
TRANSCRIPT

Hsp27 Regulates Akt Activation and PolymorphonuclearLeukocyte Apoptosis by Scaffolding MK2 to AktSignal Complex*
Received for publication, December 11, 2006, and in revised form, May 15, 2007 Published, JBC Papers in Press, May 17, 2007, DOI 10.1074/jbc.M611316200
Rui Wu‡1, Hina Kausar‡1, Paul Johnson§, Diego E. Montoya-Durango§, Michael Merchant‡, and Madhavi J. Rane‡§2
From the Departments of ‡Medicine and §Biochemistry and Molecular Biology, University of Louisville, Louisville, Kentucky 40202
We have shown previously that Akt exists in a signal com-plex with p38 MAPK, MAPK-activated protein kinase-2(MK2), and heat shock protein 27 (Hsp27) andMK2 phospho-rylates Akt on Ser-473. Additionally, dissociation of Hsp27from Akt, prior to Akt activation, induced polymorphonu-clear leukocyte (PMN) apoptosis. However, the role of Hsp27in regulating Akt activation was not examined. This studytested the hypothesis that Hsp27 regulates Akt activation andpromotes cell survival by scaffolding MK2 to the Akt signalcomplex. Here we show that loss of Akt/Hsp27 interaction byanti-Hsp27 antibody treatment resulted in loss of Akt/MK2interaction, loss of Akt-Ser-473 phosphorylation, andinduced PMN apoptosis. Transfection of myristoylated Akt(AktCA) in HK-11 cells induced Akt-Ser-473 phosphoryl-ation, activation, and Hsp27-Ser-82 phosphorylation. Co-transfection of AktCA with Hsp27 short interfering RNA,but not scrambled short interfering RNA, silenced Hsp27expression, without altering Akt expression in HK-11 cells.Silencing Hsp27 expression inhibited Akt/MK2 interaction,inhibited Akt phosphorylation and Akt activation, and inducedHK-11 cell death.Deletionmutagenesis studies identified acidiclinker region (amino acids 117–128) onAkt as anHsp27bindingregion. Deletion of amino acids 117–128 on Akt resulted in lossof its interaction with Hsp27 and MK2 but not with Hsp90 asdemonstrated by immunoprecipitation and glutathioneS-transferase pulldown studies. Co-transfection studies demon-strated that constitutively activeMK2 (MK2EE) phosphorylatedAktwt (wild type) on Ser-473 but failed to phosphorylateAkt�117–128mutant in transfixed cells. These studies collectivelydefine a novel role of Hsp27 in regulating Akt activation andcellular apoptosis by mediating interaction between Akt and itsupstream activator MK2.
Apoptosis or programmed cell death is a series of events in acell that leads to its death. Human polymorphonuclear leuko-
cytes (PMN)3 take part in host defense mechanisms againstinfection and inflammatory diseases. Inappropriate termina-tion of PMN activation or failure to remove apoptotic PMNsresults in inflammation. This apoptotic process has been sug-gested to represent an in vivo mechanism limiting oxidant-in-duced tissue injury caused by PMNs at the sites of inflamma-tion. Although PMNs are constitutively committed toapoptosis from the time they enter circulation, the rate of apo-ptosis is not fixed.We reported that interleukin-8, granulocyte-macrophage colony-stimulating factor, LTB4, and bacteriallipopolysaccharide (LPS) delay constitutive PMN apoptosisthrough the activation of the serine/threonine kinase Akt (1, 2).We demonstrated that p38 mitogen-activated protein kinase(MAPK) activity is required for Akt phosphorylation and acti-vation (3). Additionally, we showed thatAkt exists in a signalingmodule with p38 MAPK, MAPK-activated protein kinase-2(MK2), and heat shock protein 27 (Hsp27) (3).Heat shock proteins represent a group of chaperone proteins
that protect the cells against a variety of stresses. Besides beinginvolved in functioning as a chaperone, Hsp27 has also beenshown to regulate stability of the cytoskeleton, cell motility(4–7), and apoptosis (8–13). When overexpressed in tumorcells, Hsp27 increases their tumorigenicity by overexpressingMMP-9 expression and down-regulating Src tyrosine kinaseYes expression (14–16) and protects against apoptotic celldeath triggered by various stimuli, including cytotoxic drugsand ligation of the Fas/Apo-1/CD95 death receptor (17–19).Mice overexpressing Hsp27 were protected from lethal ische-mia/reperfusion injury compared with their negative litter-mates (20). Possiblemechanisms ofHsp27 anti-apoptotic activ-ity are proposed to result from its activity as a molecularchaperone. Hsp27 binds to and inactivates the pro-apoptoticmolecules Smac, caspase 3, caspase 9, and cytochrome c (21–25). Hsp27-mediated suppression of Bid translocation to themitochondria correlates with an inhibition of cytochrome crelease (25). Hsp27 has also been shown to promote survival
* This work was supported by American Heart Association-Scientist Develop-ment Grant 0335278N (to M. J. R.) and National Institutes of Health Grant1R56AI059165-01A2 (to M. J. R.). The costs of publication of this articlewere defrayed in part by the payment of page charges. This article musttherefore be hereby marked “advertisement” in accordance with 18 U.S.C.Section 1734 solely to indicate this fact.
1 Both authors contributed equally to this work.2 To whom correspondence should be addressed: University of Louisville, 570
S. Preston St., Baxter I Bldg. South, 102C, Louisville, KY 40202. Tel.: 502-852-0014; Fax: 502-852-4384; E-mail: [email protected].
3 The abbreviations used are: PMN, polymorphonuclear leukocyte; Hsp27,heat shock protein 27; PDK1, phosphoinositide-dependent kinase-1;PDK2, phosphoinositide-dependent kinase-2; MAPK, mitogen-activatedprotein kinase; MK2, MAPK-activated protein kinase-2; HK-11 cells, humanrenal proximal tubular cells; HEK-293, human embryonic kidney cells;AktCA, c-Myc-tagged myristoylated constitutively active Akt; IEF, isoelec-tric focusing; MES, 4-morpholineethanesulfonic acid; PH, pleckstrin homol-ogy; PMSF, phenylmethylsulfonyl fluoride; fMLP, formylmethionylleucylphe-nylalanine; MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazoliumbromide; FITC, fluorescein isothiocyanate; siRNA, short interfering RNA; BisTris,2-[bis(2-hydroxyethyl)amino]-2-(hydroxymethyl)propane-1,3-diol.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 282, NO. 30, pp. 21598 –21608, July 27, 2007© 2007 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A.
21598 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 30 • JULY 27, 2007
by guest on June 23, 2018http://w
ww
.jbc.org/D
ownloaded from

pathways by modulating IKK complex stability and activity.Parcellier et al. (26) demonstrated that Hsp27 mediates NF-KBactivation and cell survival by promoting the proteasomal deg-radation of polyubiquitinated IKB. Phosphorylated Hsp27 hasbeen shown to bind an adaptor protein Daxx and to inhibitFas-mediated apoptosis (27). Additionally, phosphorylation ofHsp27 has been shown to be required for proper maintenanceof cell adhesion and inhibition of renal epithelial cell apoptosis(28). Furthermore, Sheth et al. (13) showed that introduction ofrecombinant Hsp27 caused delay of PMN apoptosis; however,mechanisms regulating this delay of PMN apoptosis were notdetermined.We recently demonstrated direct protein/protein interac-
tion between Akt/Hsp27 (3, 8). The physical association ofHsp27 with Akt is a critical determinant of PMN survival, asremoval of Hsp27 from the Akt signal module prevented Aktphosphorylation and activation and resulted in acceleratedPMN apoptosis suggesting an important role for Hsp27 in reg-ulating Akt activity (8). MK2 has been shown to bind and phos-phorylate Hsp27 (29), and MK2 is PDK2 for Akt in humanPMNs (3). Recently, Zheng et al. (30) demonstrated thatMK2 isrequired for p38MAPK and Hsp27 interaction; however, asso-ciation ofHsp27 andAktwas not dependent onMK2.Hencewehypothesized that Hsp27 regulates Akt activation and apopto-sis by scaffolding MK2 to the Akt signal complex.Akt contains an N-terminal pleckstrin homology (PH)
domain and a catalytic kinase domain (residues 1–116 and148–411 respectively) linked by a highly acidic linker region(residues 117–147). The C-terminal tail region lies betweenresidues 412 and 480. Phosphoinositides are known to bind thePH domain of Akt and recruit it to the plasma membrane forfull activation by PDK1 and PDK2 (28, 31–33). In the presentstudy we show that amino acids 117–128 within the acidiclinker region of Akt are required for interactionwithHsp27. Anin-frame deletion mutant of Akt (Akt�117–128), lacking theHsp27 binding region, interacts withHsp90 but notwithHsp27and MK2. Disruption of Akt/Hsp27 interaction prevents MK2association with Akt, resulting in loss of MK2-mediated AktSer-473 phosphorylation, activation, and induction of apopto-sis. These studies demonstrate for the first time that Hsp27regulates Akt activation and cellular apoptosis by scaffoldingMK2 to the Akt signal complex.
MATERIALS AND METHODS
Anti-PH domain Akt, anti-Akt, and anti-p38 antisera wereobtained from Cell Signaling Inc. (Beverly, MA). Anti-phos-pho-Ser-473-Akt was obtained from Santa Cruz Biotechnology(Santa Cruz, CA). Anti-MK2 antibody was obtained fromSigma. Mouse isotype control antibody was obtained fromSanta Cruz Biotechnology (Santa Cruz, CA). RecombinantHsp27 and anti-mouse Hsp27 were obtained from StressGenBiotechnologies Corp. (Victoria, British Columbia, Canada).ProteinA-Sepharose and glutathione-Sepharosewere obtainedfrom Pharmingen. Histone H2B was obtained from RocheApplied Science. Recombinant GST-Akt-(1–149), recombi-nant active MK2, and recombinant active catalytic proteinkinase A were obtained from Upstate Biotechnology Inc. (LakePlacid, NY). pUseAktwt (wild type) and pUseAktCA (myristoy-
lated constitutively active Akt) constructs were obtained fromUpstate Biotechnology, Inc. pUseAktwt was shuttled into GSTvector pGEX-4T-2 as described previously (8). pUseAkt�117–128was shuttled into GST vector pGEX-4T-2 as described under“Materials and Methods.” pcDNA3.1-Hsp27-wt was shut-tled into pGEX-5X-2 (GE Healthcare) vector as describedpreviously (8). Constitutively active MK2EE cDNA con-struct was obtained from the Gaestel laboratory (Hannover,Germany).Isolation of PMNs and Culture Conditions—PMNs were iso-
lated from venous blood obtained from healthy volunteers asdescribed previously (3, 8). PMN preparations routinely con-tained �95% PMNs, as determined by morphology, and were�99% viable by trypan blue dye exclusion. PMNs were sus-pended in RPMI 1640 medium supplemented with 10% fetalcalf serum, L-glutamine, penicillin, and streptomycin and incu-bated for the indicated times at 37 °C in 5% CO2.Generation of pUseAkt�117–128 Mutant by Site-directed
Mutagenesis—In-frame deletion of amino acids 117–128 frompUseAktwt cDNA construct was carried out using the Trans-former site-directed mutagenesis kit from BD Biosciencesaccording to the manufacturer’s instructions. The in-framedeletion primer was 5�-AGGCAGGAAGAAGAGTCAGGGG-CTGAAGAG-3�, and the selection primer for pUseAktwt(mutating the KpnI site) was 5�-GTTAAGCTTGAATCCGA-GCTCG-3�. Cloning and mutation were confirmed by DNAsequencing.Subcloning and Purification of GST Beads and Recombinant
Proteins—Akt�117–128 was excised from pUSEAkt�117–128 withrestriction enzymes BamHI/PmeI and ligated into BamHI/SmaIsites of pGEX-4T-2 (GE Healthcare) vector. Generation of GST-Hsp27pGEX-5X-2, GST-AktpGEX-4T-2, was described previ-ously (8). GST-pGEX-4T-2, GST-Hsp27pGEX-5X-2, GST-Akt-pGEX-4T-2, and GST-Akt�117–128pGEX-4T-2 cDNAs weretransformed into Escherichia coli BL21(DE3)PlysS, and theexpression andpurificationofGST,GST-Hsp27,GST-Aktwt, andGST-Akt�117–128 fusion proteins were performed as describedpreviously (8). pRSETA vector was digested with restrictionenzyme EcoRI followed by generation of a blunt end by treat-ment with Klenow enzyme followed by digestion with BamHIrestriction enzyme. This vectorwas then ligated to eitherAktwtor Akt�117–128, which were excised from pUSEAktwt orpUSEAkt�117–128 with restriction enzymes BamHI/PmeI. Allpositive clones were confirmed by DNA sequencing. Expres-sion of pRSET-Aktwt and pRSET-Akt�117–128 plasmids wascarried out in BL21(DE3)pLysS chemically competent E. colicells, and protein was purified using the ProBond purificationsystem (Invitrogen).Immunoblot Analysis—Immunoblotting procedures were
performed as described previously (3, 8). Fifty�g of proteinwassubjected to 10% SDS-PAGE and immunoblot analysis withanti-pAktSer-473 (1:1000, Santa Cruz Biotechnology), anti-Akt(1:1000, Santa Cruz Biotechnology), anti-Hsp27 (StressGen),anti-Ser(P)-82Hsp27 (Cell Signaling), anti-c-Myc (Cell Signal-ing), anti-MK2 (Sigma), and anti-Hsp90 (Santa Cruz Biotech-nology) antisera (3, 8).Gel Filtration Chromatography—The system used for the
size exclusion chromatography experiments consisted of a
Regulation of Akt Activation by Hsp27
JULY 27, 2007 • VOLUME 282 • NUMBER 30 JOURNAL OF BIOLOGICAL CHEMISTRY 21599
by guest on June 23, 2018http://w
ww
.jbc.org/D
ownloaded from

HiPrep 26/60 Sephacryl S-300 HR prepacked chromatographycolumn connected to an AKTA purifier 10 liquid chromatogra-phy system (Amersham Biosciences), equipped with a Frac-900automated fraction collector, and controlled by the UNICORNversion 4.00 software (Amersham Biosciences). Prior to chro-matography, the columnwas equilibrated in 50mMTris-Cl, pH7.4, 1 mM EDTA, 150 mM NaCl, 1.5 mM MgCl2, 5% glycerol,0.5% Triton X-100. For the chromatographic separation of thesamples, control and fMLP (0.3 �M)-stimulated PMNs wereharvested and resuspended inAkt lysis buffer containing 20mMTris-HCl, pH 7.4, 150 mM NaCl, 1% (v/v) Triton X-100, 0.5%(v/v) Nonidet P-40, 1 mM EDTA, 1 mM EGTA, 20 mM sodiumorthovanadate, 10 �M p-nitrophenol phosphate, 20 mM NaF, 5mM PMSF, 21 �g/ml aprotinin, and 5 �g/ml leupeptin. The celllysate was cleared by centrifugation at 14,000 rpm for 15 min.Next, 3–5 ml of total cleared cell lysate was injected onto thecolumn with a manual injection through a 50-ml capacitysuperloop system (Amersham Biosciences). Isocratic elutionwith 2.5 column volumes of the same buffer was performed.Both a constant flow rate of 1.0 ml/min and the absorbanceprofiles atA230nm andA280nmweremonitored during the entirechromatographic procedure. Fractions of 1.0mlwere collected,and 100 �l of every fifth fraction was used for Western blotanalysis. All procedures were performed at 4 °C.Isoelectric Focusing (IEF) Electrophoresis—PMN lysates (25
�g) were subjected to IEF electrophoresis. Proteins were sepa-rated based on their isoelectric point. Precast IEF (NOVEX)gels (Invitrogen) were run according tomanufacturer’s instruc-tions. Gels were fixed in 100ml of 20%methanol solution for 20min. Gel was rinsed in 1� SDS-running buffer for 20 min andtransferred onto nitrocellulosemembrane using semidry trans-fer apparatus (Invitrogen) for 20 min at 20 V. Nitrocellulosemembranes were washed with Krebs� buffer and then immu-noblotted with anti-Hsp27 and anti-Ser(P)-82Hsp27 antisera.Two-dimensional PAGE—Protein lysates (50 �g) were
diluted into urea/thiourea rehydration buffer (Genomic Solu-tions), and proteins were separated by two-dimensional PAGE.IPG strips, pH 3–10 (Invitrogen), were rehydrated overnightwith protein samples. Proteins were separated on the basis oftheir isoelectric point by IEF using the ZOOM IPG Runner(Invitrogen) with a maximal voltage of 2000 V and 50 �A pergel. Following IEF, IPG stripswere incubated twice in equilibra-tion buffer I (6 M urea, 130 mM dithiothreitol, 30% glycerol, 45mM Tris base, 1.6% SDS, 0.002% bromphenol blue; GenomicSolutions) and once in equilibration buffer II (6 M urea, 135mMiodoacetamide, 30% glycerol, 45 mM Tris base, 1.6% SDS,0.002% bromphenol blue; Genomic Solutions) for 10 min.Equilibrated IPG strips were applied to 4–12% BisTris gradientgels (Invitrogen), and proteins were separated in the seconddimension based on their molecular size using NuPAGEMES/SDS buffer (Invitrogen) at 200 V for 40 min. Following electro-phoresis, gels were transferred onto nitrocellulose and immu-noblotted with anti-Hsp27 antibody.GST Pulldown Assay—PMNs (2 � 107) were untreated or
treated with isotype control antibody or Hsp27 antibody for 2 hat 37 °C. The cells were lysed with 200�l of lysis buffer contain-ing 1% (v/v) Nonidet P-40, 10% (v/v) glycerol, 137 mMNaCl, 20mM Tris-HCl, pH 7.4, 1 �g/ml aprotinin, 1 �g/ml leupeptin, 5
mM phenylmethylsulfonyl fluoride, 20 mM NaF, 1 mM sodiumpyrophosphate, 1 mM sodium orthovanadate, and 1% (v/v) Tri-ton X-100. Appropriate GST beads (10 �l) were added to thelysates and incubated at 4 °C for 1 h with shaking. The beadswere washed three times with Krebs buffer, and 15 �l of 2�Laemmli buffer was added to each tube. The samples wereboiled for 3 min and then subjected to 10% SDS-PAGE. Pro-teins were transferred onto nitrocellulose and immunoblottedwith appropriate antibodies.GST PulldownAssay with Recombinant Proteins—Appropri-
ate GST beads (10 �l) were added to 50 �l of kinase buffer (20mM HEPES, 10 mM MgCl2, 10 mM MnCl2) containing 50 ng ofappropriate recombinant protein. The samples were incubatedat 4 °C for 1 h with shaking. The beads were washed three timeswith Krebs buffer, and 15�l of 2� Laemmli buffer was added toeach tube. The samples were boiled for 3 min and then sub-jected to 10% SDS-PAGE and immunoblotting. RecombinantGST-Akt-PH domain (1–149 amino acids) was first conjugatedwith glutathione-Sepharose beads by incubating the two at 4 °Cfor 1 h with shaking. These beads were then incubated with 50ng of recombinant Hsp27 as described above.Cell Culture—HK-11 cells (human renal tubular epithelial
cells) immortalized by transduction with adenovirus 12-SV40were obtained from Dr. Racusen (34). Cells were cultured inDulbecco’s modified Eagle’s medium/Ham’s F-12 (Invitrogen)supplemented with 5% fetal calf serum (Sigma) and penicillin/streptomycin (100 units/ml) (Invitrogen). Fresh growthmedium was added to cells every 3–4 days until confluent.HEK-293 cells were cultured in Dulbecco’s modified Eagle’smedium supplemented with 10% fetal bovine serum and peni-cillin/streptomycin (100 units/ml) (Invitrogen).cDNA Transfections—HK-11or HEK-293 cells were plated
on 6-well trays a day prior to performing transfections, toachieve 60% confluence. On the day of transfection appropriatecellswerewashedwith serum-freeRPMI1640medium.One�gof appropriate cDNA was transfected into these cells usingGenePORTER reagent according to the manufacturer’s proto-col (Gene Therapy Systems). Twenty four hours after transfec-tion cells were lysed in 100 �l of Akt lysis buffer containing 20mM Tris-HCl, pH 7.4, 150 mM NaCl, 1% (v/v) Triton X-100,0.5% (v/v) Nonidet P-40, 1 mM EDTA, 1 mM EGTA, 20 mM
sodiumorthovanadate, 10�M p-nitrophenol phosphate, 20mM
NaF, 5 mM PMSF, 21 �g/ml aprotinin, and 5 �g/ml leupeptin,and proteins concentrations were determined. Protein lysates(50 �g) were subjected to SDS-PAGE and immunoblot analysisor to immunoprecipitation studies.Construction of Hsp27 siRNA—Construction of siRNA
was performed as described previously (35). Hsp27 siRNAwas generated using Silencer� siRNA construction kit(Ambion, Austin, TX). Hsp27 sequence 5�-AAGACCAAG-GATGGCGTGGTG-3� was targeted to generate Hsp27siRNA. The sense and antisense siRNA oligonucleotides usedto generate Hsp27 siRNA were 5�-AACACCACGCCATCCT-TGGTCCCTGTCTC-3� (sense) and 5�-AAGACCAAGGAT-GGCGTGGTGCCTGTCTC-3� (antisense). The Hsp27 siRNAand a scrambled siRNA were transfected by using Gene-PORTER reagent as outlined above.
Regulation of Akt Activation by Hsp27
21600 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 30 • JULY 27, 2007
by guest on June 23, 2018http://w
ww
.jbc.org/D
ownloaded from

Assessment of Cell Viability—Cell viability was measured byassessment of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) reduction (Sigma) as describedpreviously (36, 37). The soluble form of MTT was reduced bymitochondria of live cells, resulting in a water-insoluble salt.Product formation was monitored by reading absorbance at540 nm using a microplate reader.Akt Immunoprecipitation Assays—Appropriate cells were
lysed inAkt lysis buffer containing 20mMTris-HCl, pH 7.4, 150mMNaCl, 1% (v/v) TritonX-100, 0.5% (v/v)Nonidet P-40, 1mMEDTA, 1mMEGTA, 20mM sodiumorthovanadate, 10�M p-ni-trophenol phosphate, 20 mMNaF, 5 mM PMSF, 21 �g/ml apro-tinin, and 5 �g/ml leupeptin. Following centrifugation at15,000 � g for 15 min at 4 °C, cleared lysates were incubatedwith 20 �l of anti-Akt PH domain agarose beads or with mouseisotype control antibody beads as described previously (8).Immunoprecipitated proteins were eluted with 40 �l of 2�Laemmli dye. Samples were boiled for 2 min; beads were pre-cipitated by a quick spin in a picofuge, and 40 �l supernatantcontaining eluted proteins was subjected to SDS-PAGE andimmunoblot analysis.Akt Kinase Assay—HK-11 cells were transfected with pUSE
vector or pUSEAktCA (c-Myc-tagged myristoylated constitu-tively active Akt; Upstate Biotechnology, Inc.) or pUSEAktCAalong with Hsp27siRNA. Transfected lysates were subjected toanti-Akt immunoprecipitation (as described above). Immuno-precipitated Akt was subjected to an in vitro Akt kinase assayusing histone H2B as substrate as described previously (8).Annexin V binding was performed as described previously
(8). Briefly, 1� 106 PMNswerewashed and resuspended in 100�l of RPMI 1640 medium without fetal calf serum. The cellswere prewarmed for 5 min at 37 °C; anti-Hsp27 antibody (10�g) or isotype control antibody (10�g) was then added, and thecellswere further incubated for 2 h at 37 °C. PMNswerewashedonce with RPMI 1640 medium without fetal calf serum andcentrifuged at 400 � g for 2 min. Cells were resuspended inice-cold 1� Binding Buffer (10 mM Hepes/NaOH, pH 7.4, 150mM NaCl, 5 mM KCl, 1 mM MgCl2, 2.5 mM CaCl2) at a concen-tration of 106 cells/0.3 ml. Next 3 �l of annexin conjugateApopNexin FITC was added to the cell suspension and incu-bated at room temperature for 15 min in the dark. The cellswere immediately examined using Zeiss Axiovert 100 micro-scope and LSM 510 software.
RESULTS
Enhanced Akt/Hsp27 Interaction during LPS Delay of PMNApoptosis—Cultured PMNs undergo constitutive apoptosissuch that only about 30% are viable after 24 h. However, treat-ment of PMNs with lipopolysaccharide (LPS) delays constitu-tive PMN apoptosis, such that about 70% are viable after 24 h(2). We determined whether increased PMN viability at 24 h inthe presence of LPS could be attributed to increasedAkt/Hsp27interaction. Freshly isolated PMNs (zero time control) andPMNs in culture for 24 h with or without LPS were lysed.Lysates were subjected to anti-Akt immunoprecipitation andimmunoblotted with anti-Hsp27 antibody. Immunoprecipi-tates were also immunoblotted with anti-Akt to determineequal immunoprecipitation in every condition. Additionally, as
control, zero time PMN lysates were subjected to isotype con-trol antibody immunoprecipitation and immunoblotted withanti-Hsp27 and anti-Akt antisera (Fig. 1, 4th lane). As expected,no Akt or Hsp27 binding was detected in PMN lysates immu-noprecipitated with isotype control antibody (Fig. 1, 4th lane).Akt/Hsp27 interaction was detected in zero time PMN lysatessubjected to anti-Akt immunoprecipitation (Fig. 1, 3rd lane).Akt-Hsp27 association was markedly reduced in PMNs cul-tured for 24 h in the absence of LPS (Fig. 1, 2nd lane); however,this association was maintained in PMNs cultured for 24 h inthe presence of LPS (Fig. 1, 1st lane). These results suggest thereis a need to determine the cause and effect relationship betweenAkt/Hsp27 interaction and PMN viability, suggesting theimportance of Akt/Hsp27 interaction in the regulation of PMNapoptosis during inflammation.Separation of Akt Signal Components in Control and fMLP-
stimulated PMNs—We have previously shown in PMNs thatAkt exists in a complex with Hsp27, MK2, and p38 MAPK andthat p38 MAPK-dependent kinase MK2 regulates Akt activa-tion by phosphorylating Ser-473 on Akt (8). In addition, wedemonstrated that upon fMLP stimulation, Hsp27 dissociatesfrom the Akt signal complex, whereas MK2 and p38 MAPKcontinue to associate with Akt (3, 8). To estimate percentage ofHsp27 or MK2 or p38 MAPK from PMNs that associate withAkt, we subjected control and fMLP-stimulated PMN lysates togel filtration chromatography. Fractions collected were immu-noblotted with anti-Hsp27, anti-MK2, anti-p38 MAPK, andanti-Akt (Fig. 2A). The results demonstrated that Hsp27 elutedin the same fractions asAkt in control PMN lysates. In addition,MK2 and p38MAPK eluted in the same fractions as Hsp27 andAkt; however, both MK2 and p38 were detected in additionalfractions in the absence of Akt and Hsp27. Furthermore, p38MAPK was detected in some fractions in the absence of Akt,Hsp27, andMK2. These results suggest that under control con-ditions, almost 100% of Hsp27 and Akt associate with MK2,whereas a small percentage ofMK2 exists in a complexwith p38MAPK in the absence of Akt andHsp27 (Fig. 2A). These results
FIGURE 1. Enhanced Akt/Hsp27 interaction during LPS delay of apo-ptosis. Lysates from human PMNs freshly isolated or in culture for 24 h inthe presence and absence of LPS (100 ng/ml) were subjected to anti-Aktimmunoprecipitation (IP) and immunoblotted (IB) with anti-Hsp27 anti-body. Lysates from freshly isolated PMNs were also subjected to isotypecontrol immunoprecipitation as control. As a control, immunoprecipitateswere immunoblotted with anti-Akt antibody to demonstrate equalamounts of immunoprecipitations in each condition.
Regulation of Akt Activation by Hsp27
JULY 27, 2007 • VOLUME 282 • NUMBER 30 JOURNAL OF BIOLOGICAL CHEMISTRY 21601
by guest on June 23, 2018http://w
ww
.jbc.org/D
ownloaded from

were confirmed by performing Akt immunoprecipitations andimmunoblotting with anti-Hsp27 and anti-MK2 antisera (datanot shown). In fMLP-stimulated PMNs, we demonstrated thatHsp27 eluted out prior to Akt (Fig. 2B). Thus, only a smallfraction of Hsp27 associates with Akt, whereas a large amountof Hsp27 was not in a complex with Akt in fMLP-stimulatedPMNs. MK2 and p38 MAPK continue to associate with Aktafter fMLP stimulation.Phosphorylation Induces an Acidic Shift in Hsp27 pI without
Altering ItsMolecular Size inHumanPMNs—Theoligomeriza-tion status of Hsp27 in human PMNs has not been demon-strated to date. To determine whether Hsp27 existed at differ-ent isoelectric points by the virtue of its phosphorylation,control and fMLP-stimulated PMN lysates were subjected toIEF gel electrophoresis and immunoblotting with anti-Hsp27antibody. The results demonstrated two species of Hsp27 withdifferent isoelectric points in both control and fMLP-stimu-lated PMN lysates. However, the intensity of the more acidicform of Hsp27 was enhanced in fMLP-stimulated lysate com-paredwith control lysate (Fig. 3A, left panel). Immunoblot anal-ysis with anti-Ser(P)-82Hsp27 antibody confirmed that theincrease in the acidic form of Hsp27 correlated with enhancedHsp27 Ser-82 phosphorylation (Fig. 3A, right panel). Havingdemonstrated that two forms of Hsp27 exist at different iso-electric points, we next sought to determine the size of thesetwo forms ofHsp27. To accomplish this goal, PMN lysates weresubjected to two-dimensional PAGE. PMN proteins were sep-arated in the first dimension on pI and separated on the seconddimension by the virtue of theirmolecular weight or size. These
two-dimensional polyacrylamide gels were subsequently sub-jected to immunoblotting with anti-Hsp27 antibody. Resultsdemonstrated two species of Hsp27; however, both these spe-cies were identical in size (Fig. 3B). Collectively, IEF and two-dimensional PAGE Hsp27 immunoblot analysis of control andfMLP-stimulated PMNs lysates demonstrated that phospho-rylation ofHsp27 in PMNs induced a shift in pIwithout alteringsize of Hsp27. Thus, under these current experimental condi-tions, Hsp27 does not exist as an oligomeric protein in PMNs.Hsp27 Depletion Induces PMN Apoptosis by Inhibiting Akt
Association with Its Upstream Kinase MK2—We have previ-ously shown that loss of Akt/Hsp27 association in human
FIGURE 2. Separation of Akt signal components in control and fMLP-stim-ulated PMNs. Human PMN lysates generated from control (A) and 0. 3 �M
fMLP-treated (B) PMNs were subjected to gel filtration chromatography.Eluted protein fractions were collected as described under “Materials andMethods.” Protein fractions were subjected to SDS-PAGE and immunoblot-ting with anti-Akt, anti-Hsp27, anti-MK2, and anti-p38 MAPK antisera. Resultsshow that Akt and Hsp27 elute out in the same fractions in control PMNlysates, although a majority of Hsp27 elutes out prior to Akt in fMLP-treatedPMN lysates, suggesting its dissociation from Akt. MK2 and p38 MAPK asso-ciate with Akt in the presence and absence of fMLP.
FIGURE 3. Phosphorylation induces an acidic shift in Hsp27 pI withoutaltering its molecular size in human PMNs. A, human PMN lysates gener-ated from control (C) and 5-min fMLP (F5) (0.3 �M)- treated PMNs were sub-jected to IEF electrophoresis and immunoblotted (IB) with anti-Hsp27 andanti-Ser(P)-82Hsp27 antisera as described under “Materials and Methods.” IEFimmunoblotting demonstrated two species of Hsp27 with different isoelec-tric points in both control and fMLP-stimulated PMN lysates. Increased abun-dance of the more acidic species of Hsp27 was seen in fMLP-treated lysates.Immunoblot analysis with anti-Ser(P)-82Hsp27 antibody confirmed that theincrease in the acidic form of Hsp27 correlated with enhanced Hsp27-Ser82phosphorylation. B, control and 5-min fMLP (0.3 �M)-treated PMN lysateswere subjected to two-dimensional PAGE (2D-PAGE) immunoblot analysiswith anti-Hsp27 antibody. Results demonstrated two species of Hsp27 asseen in IEF gels; however, both these species were of identical size. Collec-tively, these results demonstrate that phosphorylation of Hsp27 induced anacidic pI shift of Hsp27 without altering its size in human PMNs.
Regulation of Akt Activation by Hsp27
21602 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 30 • JULY 27, 2007
by guest on June 23, 2018http://w
ww
.jbc.org/D
ownloaded from
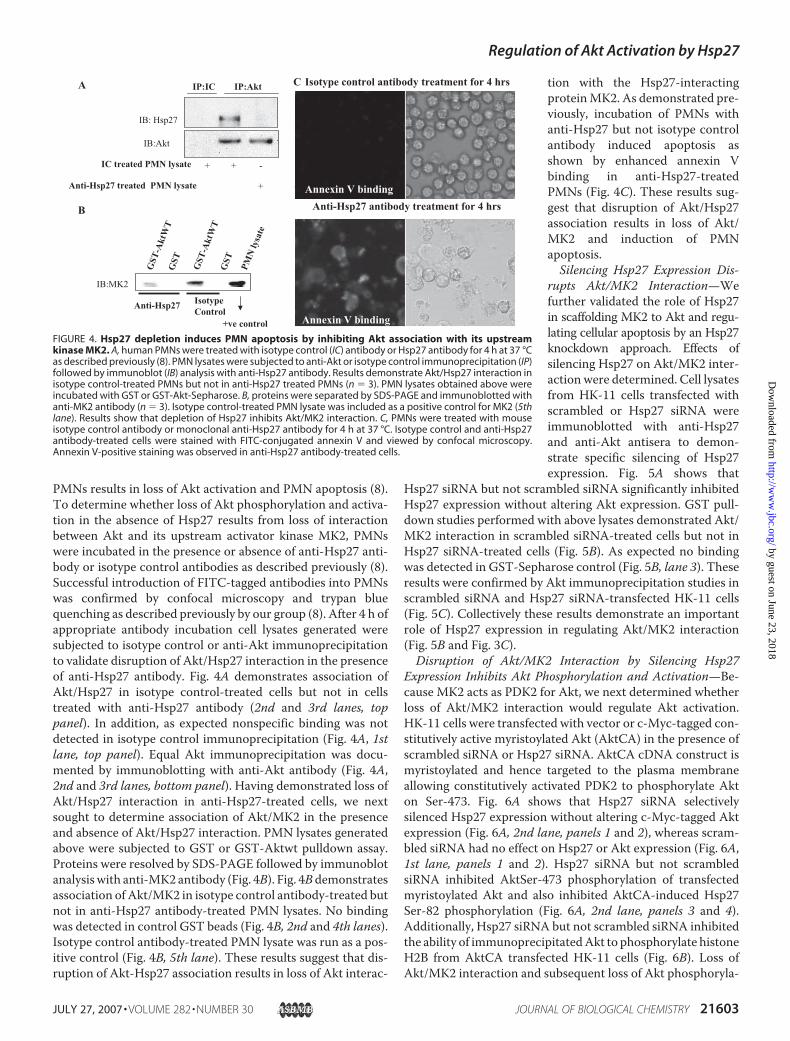
PMNs results in loss of Akt activation and PMN apoptosis (8).To determine whether loss of Akt phosphorylation and activa-tion in the absence of Hsp27 results from loss of interactionbetween Akt and its upstream activator kinase MK2, PMNswere incubated in the presence or absence of anti-Hsp27 anti-body or isotype control antibodies as described previously (8).Successful introduction of FITC-tagged antibodies into PMNswas confirmed by confocal microscopy and trypan bluequenching as described previously by our group (8). After 4 h ofappropriate antibody incubation cell lysates generated weresubjected to isotype control or anti-Akt immunoprecipitationto validate disruption of Akt/Hsp27 interaction in the presenceof anti-Hsp27 antibody. Fig. 4A demonstrates association ofAkt/Hsp27 in isotype control-treated cells but not in cellstreated with anti-Hsp27 antibody (2nd and 3rd lanes, toppanel). In addition, as expected nonspecific binding was notdetected in isotype control immunoprecipitation (Fig. 4A, 1stlane, top panel). Equal Akt immunoprecipitation was docu-mented by immunoblotting with anti-Akt antibody (Fig. 4A,2nd and 3rd lanes, bottom panel). Having demonstrated loss ofAkt/Hsp27 interaction in anti-Hsp27-treated cells, we nextsought to determine association of Akt/MK2 in the presenceand absence of Akt/Hsp27 interaction. PMN lysates generatedabove were subjected to GST or GST-Aktwt pulldown assay.Proteins were resolved by SDS-PAGE followed by immunoblotanalysiswith anti-MK2 antibody (Fig. 4B). Fig. 4Bdemonstratesassociation of Akt/MK2 in isotype control antibody-treated butnot in anti-Hsp27 antibody-treated PMN lysates. No bindingwas detected in control GST beads (Fig. 4B, 2nd and 4th lanes).Isotype control antibody-treated PMN lysate was run as a pos-itive control (Fig. 4B, 5th lane). These results suggest that dis-ruption of Akt-Hsp27 association results in loss of Akt interac-
tion with the Hsp27-interactingproteinMK2. As demonstrated pre-viously, incubation of PMNs withanti-Hsp27 but not isotype controlantibody induced apoptosis asshown by enhanced annexin Vbinding in anti-Hsp27-treatedPMNs (Fig. 4C). These results sug-gest that disruption of Akt/Hsp27association results in loss of Akt/MK2 and induction of PMNapoptosis.Silencing Hsp27 Expression Dis-
rupts Akt/MK2 Interaction—Wefurther validated the role of Hsp27in scaffolding MK2 to Akt and regu-lating cellular apoptosis by an Hsp27knockdown approach. Effects ofsilencing Hsp27 on Akt/MK2 inter-action were determined. Cell lysatesfrom HK-11 cells transfected withscrambled or Hsp27 siRNA wereimmunoblotted with anti-Hsp27and anti-Akt antisera to demon-strate specific silencing of Hsp27expression. Fig. 5A shows that
Hsp27 siRNA but not scrambled siRNA significantly inhibitedHsp27 expression without altering Akt expression. GST pull-down studies performed with above lysates demonstrated Akt/MK2 interaction in scrambled siRNA-treated cells but not inHsp27 siRNA-treated cells (Fig. 5B). As expected no bindingwas detected in GST-Sepharose control (Fig. 5B, lane 3). Theseresults were confirmed by Akt immunoprecipitation studies inscrambled siRNA and Hsp27 siRNA-transfected HK-11 cells(Fig. 5C). Collectively these results demonstrate an importantrole of Hsp27 expression in regulating Akt/MK2 interaction(Fig. 5B and Fig. 3C).Disruption of Akt/MK2 Interaction by Silencing Hsp27
Expression Inhibits Akt Phosphorylation and Activation—Be-cause MK2 acts as PDK2 for Akt, we next determined whetherloss of Akt/MK2 interaction would regulate Akt activation.HK-11 cells were transfected with vector or c-Myc-tagged con-stitutively active myristoylated Akt (AktCA) in the presence ofscrambled siRNA or Hsp27 siRNA. AktCA cDNA construct ismyristoylated and hence targeted to the plasma membraneallowing constitutively activated PDK2 to phosphorylate Akton Ser-473. Fig. 6A shows that Hsp27 siRNA selectivelysilenced Hsp27 expression without altering c-Myc-tagged Aktexpression (Fig. 6A, 2nd lane, panels 1 and 2), whereas scram-bled siRNA had no effect on Hsp27 or Akt expression (Fig. 6A,1st lane, panels 1 and 2). Hsp27 siRNA but not scrambledsiRNA inhibited AktSer-473 phosphorylation of transfectedmyristoylated Akt and also inhibited AktCA-induced Hsp27Ser-82 phosphorylation (Fig. 6A, 2nd lane, panels 3 and 4).Additionally, Hsp27 siRNA but not scrambled siRNA inhibitedthe ability of immunoprecipitatedAkt to phosphorylate histoneH2B from AktCA transfected HK-11 cells (Fig. 6B). Loss ofAkt/MK2 interaction and subsequent loss of Akt phosphoryla-
FIGURE 4. Hsp27 depletion induces PMN apoptosis by inhibiting Akt association with its upstreamkinase MK2. A, human PMNs were treated with isotype control (IC) antibody or Hsp27 antibody for 4 h at 37 °Cas described previously (8). PMN lysates were subjected to anti-Akt or isotype control immunoprecipitation (IP)followed by immunoblot (IB) analysis with anti-Hsp27 antibody. Results demonstrate Akt/Hsp27 interaction inisotype control-treated PMNs but not in anti-Hsp27 treated PMNs (n � 3). PMN lysates obtained above wereincubated with GST or GST-Akt-Sepharose. B, proteins were separated by SDS-PAGE and immunoblotted withanti-MK2 antibody (n � 3). Isotype control-treated PMN lysate was included as a positive control for MK2 (5thlane). Results show that depletion of Hsp27 inhibits Akt/MK2 interaction. C, PMNs were treated with mouseisotype control antibody or monoclonal anti-Hsp27 antibody for 4 h at 37 °C. Isotype control and anti-Hsp27antibody-treated cells were stained with FITC-conjugated annexin V and viewed by confocal microscopy.Annexin V-positive staining was observed in anti-Hsp27 antibody-treated cells.
Regulation of Akt Activation by Hsp27
JULY 27, 2007 • VOLUME 282 • NUMBER 30 JOURNAL OF BIOLOGICAL CHEMISTRY 21603
by guest on June 23, 2018http://w
ww
.jbc.org/D
ownloaded from

tion and activation in the presence of Hsp27 siRNA suggest acritical role for Hsp27 in the regulation of Akt activation.Disruption of Akt/MK2 Interaction by Silencing Hsp27
Expression Induces HK-11 Cell Death—Silencing Hsp27expression inhibits Akt/MK2 interaction and Akt activation.Thus, we determined effects of silencing Hsp27 expression onHK-11 cell viability.HK-11 cells alone or HK-11 cells in the presence of Gene-
PORTER reagent, scrambled siRNA, or Hsp27 siRNA wereincubated at 37 °C in 5% CO2 for 24 h, and cells were subjectedto MTT cell viability assay. Hsp27 siRNA but not scrambledsiRNA significantly inhibited Hsp27 without altering Aktexpression (Fig. 7, bottom panel). Additionally, Hsp27 siRNAbut not scrambled siRNA significantly decreased HK-11 cellviability. GenePORTER reagent by itself had no effect on cellviability (Fig. 7, top panel). These results suggest that Hsp27 is asurvival protein, and its regulation of HK-11 survival may bemediated by scaffoldingMK2 to Akt, allowing Akt activation tooccur, thereby promoting cell survival.Akt PH Domain Is Not Required for Interaction with Hsp27—
Wehave demonstrated previously that Hsp27 directly interactswith Akt (8). To identify Hsp27-binding site(s) on Akt, wedetermined the ability of GST-Akt-(1–149)-Sepharose, which
contains theAkt PHdomain (1–116 amino acids) and the acidiclinker region (117–149) to interact with recombinant Hsp27.Recombinant GST-Akt-(1–149) was conjugated to glutathi-one-Sepharose as described under “Materials and Methods.”Fig. 8A demonstrates thatGST-Akt-(1–149) (2nd lane), but notglutathione-Sepharose (1st lane), specifically interacts withrecombinantHsp27 suggesting a role for the PHdomain and/orthe acidic linker region to interact with Hsp27. RecombinantGST-Akt-(1–149) and recombinantHsp27were run as positivecontrols (Fig. 8A, 3rd and 4th lanes). To determine the role ofAkt PH domain in interacting with Hsp27, recombinantAkt�PH (mutant lacking N-terminal PH domain or 1–116amino acids) was precipitated with GST or GST-Hsp27. Fig. 8Bdemonstrates interaction of recombinant Akt�PH with GST-Hsp27 but not with GST-Sepharose. Recombinant Akt�PH wasrun as positive control (Fig. 8B, lane 3). These results suggestthat Akt-PH domain is not required for its interaction withHsp27.
FIGURE 5. Silencing Hsp27 expression disrupts Akt/MK2 interaction.HK-11 cells were transfected with scrambled or Hsp27 siRNA. A, cell lysatesfrom transfected cells were immunoblotted (IB) with anti-Hsp27 and anti-Akt antisera to demonstrate specific silencing of Hsp27 expression (n � 3).B, above lysates were subjected to GST or GST-Aktwt pulldown assay todetermine Akt/MK2 interaction in the presence of scrambled or Hsp27siRNA (n � 3). C, above lysates were also subjected to mouse anti-Aktimmunoprecipitation (IP) and immunoblotted with rabbit polyclonal anti-MK2 and rabbit polyclonal anti-Akt antisera (n � 3). The results demon-strate loss of Hsp27 expression abrogates Akt/MK2 interaction.
FIGURE 6. Disruption of Akt/MK2 interaction by silencing Hsp27 expres-sion inhibits Akt phosphorylation and activation. Human renal tubularcells (HK-11) were transfected with c-Myc-tagged myristoylated Akt (AktCA)construct with scrambled or Hsp27 siRNA. A, transfected cell lysates weresubjected to immunoblot analysis with anti-Hsp27 (panel 1), anti-c-Myc(panel 2), anti-phosphoAktSer-473 (panel 3), and anti-phosphoHsp27Ser-82(panel 4) antisera (n � 3). Results show that transfection of Hsp27 inhibitsHsp27 expression without altering expression of c-Myc-tagged Akt. Addition-ally, AktSer-473 phosphorylation and Hsp27Ser-82 phosphorylation wereinhibited in the presence of Hsp27 siRNA but not in the presence of scram-bled siRNA. B, HK-11 cells were transfected with vector (1st lane) or AktCAalong with scrambled siRNA (2nd lane) or AktCA along with Hsp27siRNA (3rdlane). Lysates were immunoprecipitated with anti-Akt antibody and sub-jected to an in vitro Akt kinase assay using histone H2B as substrate (n � 3).Autoradiograph demonstrates that transfection of AktCA induces increasedAkt-mediated histone H2B phosphorylation in the presence of scrambledsiRNA but not in the presence of Hsp27siRNA. These results demonstrate thatsilencing Hsp27 expression inhibits Akt phosphorylation and activation.
Regulation of Akt Activation by Hsp27
21604 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 30 • JULY 27, 2007
by guest on June 23, 2018http://w
ww
.jbc.org/D
ownloaded from

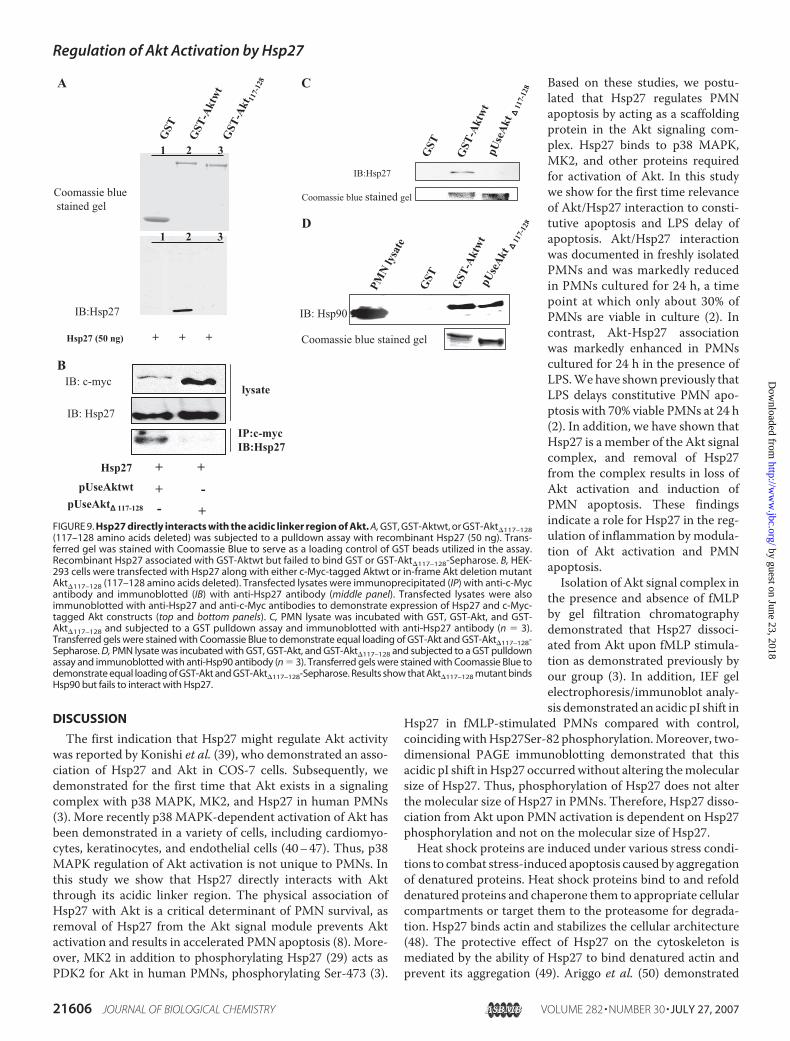
Hsp27 Directly Interacts with the Acidic Linker Region of Akt—In-framedeletionmutant ofAktwas generated such that aminoacids 117–128 from the acidic linker region ofAktwere deleted.In a GST pulldown assay recombinant Hsp27 was precipitatedby GST-Aktwt but not by GST or GST-Akt�117–128-Sepharose(Fig. 9A, bottom panel). Transferred gel was stained with Coo-massie Blue to serve as a loading control for GST beads utilizedin the assay. Additionally, HEK-293 cells were co-transfectedwith c-Myc-tagged pUseAktwt or pUseAkt�117–128 along withHsp27 cDNA. After transfection cells were lysed, and lysateswere subjected to anti-c-Myc immunoprecipitation and immu-noblotted with anti-Hsp27 antibody. Fig. 9B (bottom panel)demonstrates interaction between Aktwt and Hsp27, whereasno interaction is detected between Akt�117–128 and Hsp27. Todemonstrate overexpression of c-Myc-tagged Akt constructsand Hsp27, total lysates were immunoblotted with anti-c-Mycand anti-Hsp27 antisera (Fig. 9B, top and middle panel). Fur-thermore, these results were confirmed in human PMNs byperforming a GST pulldown assay. GST-Akt but not GST orGST-Akt�117–128 interacted with Hsp27 from PMN lysates(Fig. 9C). Transferred gels were stained with Coomassie Blue todemonstrate equal loading of GST-Akt and GST-Akt�117–128-Sepharose. These results collectively indicate that amino acids117–128 in the acidic linker region ofAkt are required for inter-action with Hsp27.We next determined the ability of GST, GST-Aktwt, and
GST-Akt�117–128 to interact with Hsp90 from PMN lysates.Hsp90 has been shown previously to directly interact withAkt via amino acids 229–309 (38). To confirm that deletionof 117–128 amino acids from Akt did not alter protein confor-mation and its ability to interactwithHsp90, we subjectedGST,GST-Akt, and GST-Akt�117–128 to a GST pulldown assay withPMN lysate. Fig. 9D demonstrates that as expected both GST-Aktwt and GST-Akt�117–128 but not GST interacted withHsp90 from PMN lysates. PMN lysate run as positive controlimmunoreacted with anti-Hsp90 antibody (Fig. 9D, lane 1).Transferred gels were stained with Coomassie Blue to demon-strate equal loading of GST-Akt and GST-Akt�117–128-Sepha-rose. These results collectively indicate that Hsp27 interactswith Akt via amino acids 117–128, and the deletion of thisregion does not severely alter its conformation and binding toits known binding protein Hsp90.MK2 Fails to Interact with Akt�117–128 and Phosphorylate It
on Ser-473 ex Vivo—Having disrupted Akt/Hsp27 interactionby deleting Hsp27 binding region on Akt, we next determinedthe ability ofGST,GST-Aktwt, andGST-Akt�117–128 to precip-itate MK2 from PMN lysates. Fig. 10A shows that GST-Aktwt,but not GST or GST-Akt�117–128, interacted with MK2 fromPMN lysate. Transferred gels were stained with CoomassieBlue to demonstrate equal loading of GST-Akt and GST-Akt�117–128-Sepharose. Having shown loss of MK2 binding toAkt�117–128, we next determined the ability of constitutivelyactivatedMK2 (MK2EE), a knownupstream activator of Akt, tophosphorylate Aktwt and Akt�117–128 in transfected HEK-293cells. Fig. 10B shows thatMK2EEphosphorylatesAktwt on Ser-473 but fails to phosphorylate Akt�117–128. These results estab-lish a novel role for Hsp27 in regulating Akt activation by scaf-folding MK2 to Akt signal complex.
FIGURE 8. Akt PH domain is not required for interaction with Hsp27.A, recombinant Hsp27 was incubated with glutathione-Sepharose (Seph, 1stlane), glutathione-Sepharose coupled to recombinant GST-Akt-(1–149) (2ndlane), and subjected to GST-pulldown assay and immunoblotted (IB) withanti-Hsp27 and anti-Akt antisera (n � 3). Recombinant GST-Akt-(1–149) andrecombinant Hsp27 alone were run as positive controls. Results show that Akt(1–149 amino acids) mediates interaction with recombinant Hsp27. B, recom-binant Akt�PH (PH domain 1–116 amino acids deleted from N terminus)mutant was incubated with GST (lane 1) or GST-Hsp27-Sepharose (lane 2) andimmunoblotted with anti-Akt antibody (n � 3). Recombinant Akt�PH was runas positive control (lane 3). Results show that PH domain of Akt is not requiredfor interaction with Hsp27.
FIGURE 7. Disruption of Akt/MK2 interaction by silencing Hsp27expression induces HK-11 cell death. HK-11 cells were treated withGenePORTER (transfection reagent) or transfected with scrambled siRNAor Hsp27 siRNA. Transfected cells were subjected to MTT cell viabilityassay. MTT reduction was determined as a measure of HK-11 cell viabilityin the presence of GenePORTER, scrambled siRNA, or Hsp27 siRNA. Resultswere expressed as % viability relative to cell alone (100%). Silencing Hsp27expression induced significant cell death (*, p � 0.05 versus cell alone, n �4). IB, immunoblot.
Regulation of Akt Activation by Hsp27
JULY 27, 2007 • VOLUME 282 • NUMBER 30 JOURNAL OF BIOLOGICAL CHEMISTRY 21605
by guest on June 23, 2018http://w
ww
.jbc.org/D
ownloaded from
DISCUSSION
The first indication that Hsp27 might regulate Akt activitywas reported by Konishi et al. (39), who demonstrated an asso-ciation of Hsp27 and Akt in COS-7 cells. Subsequently, wedemonstrated for the first time that Akt exists in a signalingcomplex with p38 MAPK, MK2, and Hsp27 in human PMNs(3). More recently p38 MAPK-dependent activation of Akt hasbeen demonstrated in a variety of cells, including cardiomyo-cytes, keratinocytes, and endothelial cells (40–47). Thus, p38MAPK regulation of Akt activation is not unique to PMNs. Inthis study we show that Hsp27 directly interacts with Aktthrough its acidic linker region. The physical association ofHsp27 with Akt is a critical determinant of PMN survival, asremoval of Hsp27 from the Akt signal module prevents Aktactivation and results in accelerated PMN apoptosis (8). More-over, MK2 in addition to phosphorylating Hsp27 (29) acts asPDK2 for Akt in human PMNs, phosphorylating Ser-473 (3).
Based on these studies, we postu-lated that Hsp27 regulates PMNapoptosis by acting as a scaffoldingprotein in the Akt signaling com-plex. Hsp27 binds to p38 MAPK,MK2, and other proteins requiredfor activation of Akt. In this studywe show for the first time relevanceof Akt/Hsp27 interaction to consti-tutive apoptosis and LPS delay ofapoptosis. Akt/Hsp27 interactionwas documented in freshly isolatedPMNs and was markedly reducedin PMNs cultured for 24 h, a timepoint at which only about 30% ofPMNs are viable in culture (2). Incontrast, Akt-Hsp27 associationwas markedly enhanced in PMNscultured for 24 h in the presence ofLPS.Wehave shownpreviously thatLPS delays constitutive PMN apo-ptosis with 70% viable PMNs at 24 h(2). In addition, we have shown thatHsp27 is a member of the Akt signalcomplex, and removal of Hsp27from the complex results in loss ofAkt activation and induction ofPMN apoptosis. These findingsindicate a role for Hsp27 in the reg-ulation of inflammation by modula-tion of Akt activation and PMNapoptosis.Isolation of Akt signal complex in
the presence and absence of fMLPby gel filtration chromatographydemonstrated that Hsp27 dissoci-ated from Akt upon fMLP stimula-tion as demonstrated previously byour group (3). In addition, IEF gelelectrophoresis/immunoblot analy-sis demonstrated an acidic pI shift in
Hsp27 in fMLP-stimulated PMNs compared with control,coincidingwithHsp27Ser-82 phosphorylation.Moreover, two-dimensional PAGE immunoblotting demonstrated that thisacidic pI shift inHsp27 occurredwithout altering themolecularsize of Hsp27. Thus, phosphorylation of Hsp27 does not alterthe molecular size of Hsp27 in PMNs. Therefore, Hsp27 disso-ciation from Akt upon PMN activation is dependent on Hsp27phosphorylation and not on the molecular size of Hsp27.Heat shock proteins are induced under various stress condi-
tions to combat stress-induced apoptosis caused by aggregationof denatured proteins. Heat shock proteins bind to and refolddenatured proteins and chaperone them to appropriate cellularcompartments or target them to the proteasome for degrada-tion. Hsp27 binds actin and stabilizes the cellular architecture(48). The protective effect of Hsp27 on the cytoskeleton ismediated by the ability of Hsp27 to bind denatured actin andprevent its aggregation (49). Ariggo et al. (50) demonstrated
FIGURE 9. Hsp27 directly interacts with the acidic linker region of Akt. A, GST, GST-Aktwt, or GST-Akt�117–128(117–128 amino acids deleted) was subjected to a pulldown assay with recombinant Hsp27 (50 ng). Trans-ferred gel was stained with Coomassie Blue to serve as a loading control of GST beads utilized in the assay.Recombinant Hsp27 associated with GST-Aktwt but failed to bind GST or GST-Akt�117–128-Sepharose. B, HEK-293 cells were transfected with Hsp27 along with either c-Myc-tagged Aktwt or in-frame Akt deletion mutantAkt�117–128 (117–128 amino acids deleted). Transfected lysates were immunoprecipitated (IP) with anti-c-Mycantibody and immunoblotted (IB) with anti-Hsp27 antibody (middle panel). Transfected lysates were alsoimmunoblotted with anti-Hsp27 and anti-c-Myc antibodies to demonstrate expression of Hsp27 and c-Myc-tagged Akt constructs (top and bottom panels). C, PMN lysate was incubated with GST, GST-Akt, and GST-Akt�117–128 and subjected to a GST pulldown assay and immunoblotted with anti-Hsp27 antibody (n � 3).Transferred gels were stained with Coomassie Blue to demonstrate equal loading of GST-Akt and GST-Akt�117–128-Sepharose. D, PMN lysate was incubated with GST, GST-Akt, and GST-Akt�117–128 and subjected to a GST pulldownassay and immunoblotted with anti-Hsp90 antibody (n � 3). Transferred gels were stained with Coomassie Blue todemonstrate equal loading of GST-Akt and GST-Akt�117–128-Sepharose. Results show that Akt�117–128 mutant bindsHsp90 but fails to interact with Hsp27.
Regulation of Akt Activation by Hsp27
21606 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 30 • JULY 27, 2007
by guest on June 23, 2018http://w
ww
.jbc.org/D
ownloaded from

that Hsp27 inhibited apoptosis by maintaining the redox equi-librium of the cell. In addition, overexpression of Hsp27 inhib-ited oxidant stress-induced cellular damage by increasing levelsof cellular glutathione (51). Overexpression of various heatshock proteins has been shown to induce a survival response.Contrary to most heat shock proteins, Hsp60 (52, 53) has beenshown to induce apoptosis by interaction with caspase-3,whereas Hsp27 (21–25), Hsp70 (54–56), and Hsp90 (57) havebeen shown to inhibit apoptosis by binding to and inactivatingspecific components of the apoptosis machinery. In generalheat shock protein-induced protection from stress was shownto act at the level of the apoptosome by preventing downstreamactivation of caspases. This study defines a novel role of Hsp27in the regulation of cellular apoptosis by acting as a scaffoldingprotein.We have recently demonstrated that Hsp27 exists in a com-
plex with Akt, p38 MAPK, MK2, and Hsp27, and MK2 acts asPDK2 for Akt (3). In addition, we demonstrated that MK2-binding protein Hsp27 dissociated from Akt signal complexafter Akt activation (3). However, disruption of Akt/Hsp27interaction prior to Akt activation resulted in loss of fMLP-stimulated Akt activation and induced PMN apoptosis (8).These studies indicated the importance of Akt/Hsp27 interac-tion to Akt activation and PMN survival. Recently Zheng et al.(30) demonstrated that MK2 was required for p38 MAPK/Hsp27 interaction. However, MK2 was not required for asso-ciation of Hsp27 with Akt. Given that Akt/Hsp27 interactionwas not regulated by MK2 and that MK2 serves as PDK2 for
Akt, we hypothesized that Hsp27 modulated interactionswithin the Akt-Hsp27-MK2 signal complex by acting as ascaffolding protein. This study defined a novel scaffoldingrole of Hsp27 in the regulation of Akt activation and cellularapoptosis.Disruption of Akt/Hsp27 interaction by depleting Hsp27
fromhuman PMNs resulted in loss of Akt interactionwith bothHsp27 and MK2 and induction of PMN apoptosis. BecauseMK2 is anHsp27-binding protein, we next determinedwhetherdepletion of Hsp27 nonspecifically removedMK2 from the Aktsignal complex. To address this question, siRNA technologywas utilized to specifically silence Hsp27 expression fromHK-11 cells to determine effects of silencing Hsp27 on Akt/MK2 interaction andHK-11 cell viability. Hsp27 siRNAbut notscrambled siRNA-transfected HK-11 cells demonstrated amarked decrease in Hsp27 expression, loss of Akt/MK2 inter-action, and induction of HK-11 cell death. These results sug-gested a role for Hsp27 in regulating Akt/MK2 interaction andAkt activation. Cell death induced by silencing Hsp27 was notentirely surprising as Schepers et al. (58) have shown thatsilencing Hsp27 expression resulted in a 2-fold increase inVP-17-induced apoptosis in acute myeloid leukemia cells.However, induction of cell death by modulating Akt/MK2interaction was a novel observation. To determine moredirectly the role of Hsp27 in regulating Akt activation, HK-11cells were transfected with myristoylated-c-Myc-tagged Akt(AktCA) in the presence and absence of Hsp27 siRNA. Immu-noblotting studies demonstrated that loss of Hsp27 expressioninhibited AktSer-473 phosphorylation and activation. Thesestudies clearly identified a role for Hsp27 in regulating Akt acti-vation. Hsp27 negatively regulates protein kinase C� activityand inhibits cell apoptosis (59); however, we show for the firsttime that Hsp27 positively regulates Akt activation and pro-motes cell survival.We have demonstrated direct protein/protein interaction
betweenHsp27 andAkt (3, 8). This study identified amino acids117–128 in the acidic linker region of Akt to mediate interac-tion with Hsp27. An in-frame Akt deletion mutant (Akt�117–128)interacted with Hsp90� but failed to interact with Hsp27 andMK2. This finding is in accordance with the literature, whichdemonstrated that Akt interacted with Hsp90� via amino acids229–309 (38).These studies suggested that loss of Akt�117–128 interaction
with MK2 and Hsp27 was not because of altered conformationof the mutant. The importance of Hsp27 in regulating Akt/MK2 interaction and AktSer473 phosphorylation was docu-mented by co-transfecting HEK-293 cells with constitutivelyactiveMK2 (MK2EE) andHsp27alongwithAktwtorAkt�117–128.MK2EE phosphorylated Aktwt on Ser473 as expected; however,it failed to phosphorylate Akt�117–128 on Ser473, a critical siteknown to regulate Akt activation (60). Furthermore, the phys-iologic consequence of loss of Akt/MK2 interaction and loss ofSer473 phosphorylation resulted in cellular apoptosis. Thesestudies collectively define a novel role of Hsp27 in regulatingAkt activation and apoptosis by mediating interaction betweenAkt and its upstream activator MK2.
FIGURE 10. MK2 fails to interact with Akt�117–128 and phosphorylate it onSer-473 ex vivo. A, PMN lysate was incubated with GST, GST-Akt, and GST-Akt�117–128 and subjected to a GST pulldown assay and immunoblotted withanti-MK2 antibody (n � 3). Transferred gels were stained with Coomassie Blueto demonstrate equal loading of GST-Akt and GST-Akt�117–128-Sepharose.Results show that Akt�117–128 mutant fails to interact with MK2 from PMNlysates. B, HEK-293 cells were transfected with vector or co-transfected withHsp27 and MK2EE (constitutively active MK2) along with either Aktwt orAkt�117–128. Transfected lysates were immunoblotted with anti-Hsp27, anti-MK2, anti-c-Myc, and anti-pAktS473 antisera. Results show that active MK2fails to phosphorylate Akt�117–128 ex vivo in HEK-293 cells.
Regulation of Akt Activation by Hsp27
JULY 27, 2007 • VOLUME 282 • NUMBER 30 JOURNAL OF BIOLOGICAL CHEMISTRY 21607
by guest on June 23, 2018http://w
ww
.jbc.org/D
ownloaded from

REFERENCES1. Klein, J. B., Rane,M. J., Scherzer, J. A., Coxon, P. Y., Kettritz, R.,Mathiesen,
J. M., Buridi, A., and McLeish, K. R. (2000) J. Immunol. 164, 4286–42912. Klein, J. B., Buridi, A., Coxon, P. Y., Rane, M. J., Manning, T., Kettritz, R.,
and McLeish, K. R. (2001) Cell. Signal. 13, 335–3433. Rane, M. J., Coxon, P. Y., Powell, D. W., Webster, R., Klein, J. B., Ping, P.,
Pierce, W., and McLeish, K. R. (2001) J. Biol. Chem. 276, 3517–35234. Okamoto, C. T. (1999) Am. J. Physiol. 277, C1029–C10315. Huot, J., Houle, F., Rousseau, S., Deschesnes, R. G., Shah, G. M., and
Landry, J. (1998) J. Cell Biol. 143, 1361–13736. Piotrowicz, R. S., Hickey, E., and Levin, E. G. (1998) FASEB J. 12,
1481–14907. Rousseau, S., Houle, F., Landry, J., and Huot, J. (1997) Oncogene 15,
2169–21778. Rane, M. J., Pan, Y., Singh, S., Powell, D. W., Wu, R., Cummins, T., Chen,
Q., McLeish, K. R., and Klein, J. B. (2003) J. Biol. Chem. 278, 27828–278359. Venkatakrishnan, C. D., Tewari, A. K., Moldovan, L., Cardounel, A. J.,
Zweier, J. L., Kuppusamy, P., and Ilangovan, G. (2006) Am. J. Physiol. 291,H2680–H2691
10. Oya-Ito, T., Liu, B. F., and Nagaraj, R. H. (2006) J. Cell Biochem. 99,279–291
11. Rocchi, P., Beraldi, E., Ettinger, S., Fazli, L., Vessella, R. L., Nelson, C., andGleave, M. (2005) Cancer Res. 65, 11083–11093
12. Son, G. H., Geum, D., Chung, S., Park, E., Lee, K. H., Choi, S., and Kim, K.(2005) Biochem. Biophys. Res. Commun. 338, 1751–1758
13. Sheth, K., De, A., Nolan, B., Friel, J., Duffy, A., Ricciardi, R., Miller-Grazi-ano, C., and Bankey, P. (2001) J. Surg. Res. 99, 129–133
14. Hansen, R. K., Parra, I., Hilsenbeck, S. G., Himelstein, B., and Fuqua, S. A.(2001) Biochem. Biophys. Res. Commun. 282, 186–193
15. Garrido, C., Fromentin, A., Bonnotte, B., Favre, N., Moutet, M., Arrigo,A. P., Mehlen, P., and Solary, E. (1998) Cancer Res. 58, 5495–5499
16. Bruey, J. M., Paul, C., Fromentin, A., Hilpert, S., Arrigo, A. P., Solary, E.,and Garrido, C. (2000) Oncogene 19, 4855–4863
17. Garrido, C., Ottavi, P., Fromentin, A., Hammann, A., Arrigo, A. P.,Chauffert, B., and Mehlen, P. (1997) Cancer Res. 57, 2661–2667
18. Garrido, C., Mehlen, P., Fromentin, A., Hammann, A., Assem,M., Arrigo,A. P., and Chauffert, B. (1996) Eur. J. Biochem. 237, 653–659
19. Mehlen, P., Schulze-Osthoff, K., and Arrigo, A. P. (1996) J. Biol. Chem.271, 16510–16514
20. Efthymiou, C. A., Mocanu, M. M., de Belleroche, J., Wells, D. J., Latch-mann, D. S., and Yellon, D. M. (2004) Basic Res. Cardiol. 99, 392–394
21. Chauhan, D., Li, G., Hideshima, T., Podar, K., Mitsiades, C., Mitsiades, N.,Catley, L., Tai, Y. T., Hayashi, T., Shringarpure, R., Burger, R., Munshi, N.,Ohtake, Y., Saxena, S., and Anderson, K. C. (2003) Blood 102, 3379–3386
22. Pandey, P., Farber, R., Nakazawa, A., Kumar, S., Bharti, A., Nalin, C.,Weichselbaum, R., Kufe, D., and Kharbanda, S. (2000) Oncogene 19,1975–1981
23. Garrido, C., Bruey, J. M., Fromentin, A., Hammann, A., Arrigo, A. P., andSolary, E. (1999) FASEB J. 13, 2061–2070
24. Bruey, J. M., Ducasse, C., Bonniaud, P., Ravagnan, L., Susin, S. A., Diaz-Latoud, C., Gurbuxani, S., Arrigo, A. P., Kroemer, G., Solary, E., and Gar-rido, C. (2000) Nat. Cell Biol. 2, 645–652
25. Paul, C., Manero, F., Gonin, S., Kretz-Remy, C., Virot, S., and Arrigo, A. P.(2002)Mol. Cell. Biol. 22, 816–834
26. Parcellier, A., Schmitt, E., Gurbuxani, S., Seigneurin-Berny, D., Pance, A.,Chantome, A., Plenchette, S., Khochbin, S., Solary, E., and Garrido, C.(2003)Mol. Cell. Biol. 23, 5790–5802
27. Charette, S. J., Lavoie, J. N., Lambert, H., and Landry, J. (2000) Mol. Cell.Biol. 20, 7602–7612
28. de Graauw,M., Tijdens, I., Cramer, R., Corless, S., Timms, J. F., and van deWater, B. (2005) J. Biol. Chem. 280, 29885–29898
29. Stokoe, D., Engel, K., Campbell, D. G., Cohen, P., and Gaestel, M. (1992)FEBS Lett. 313, 307–313
30. Zheng, C., Lin, Z., Zhao, Z. J., Yang, Y., Niu, H., and Shen, X. (2006) J. Biol.Chem. 281, 37215–37226
31. Chan, T. O., and Tsichlis, P. N. (2001) Sci. STKE 2001, PE132. Franke, T. F., Yang, S. I., Chan, T. O., Datta, K., Kazlauskas, A., Morrison,
D. K., Kaplan, D. R., and Tsichlis, P. N. (1995) Cell 81, 727–73633. Alessi, D. R., James, S. R., Downes, C. P., Holmes, A. B., Gaffney, P. R.,
Reese, C. B., and Cohen, P. (1997) Curr. Biol. 7, 261–26934. Racusen, L. C., Monteil, C., Sgrignoli, A., Lucskay, M., Marouillat, S.,
Rhim, J. G., and Morin, J. P. (1997) J. Lab. Clin. Med. 129, 318–32935. Khundmiri, S. J., Weinman, E. J., Steplock, D., Cole, J., Ahmad, A., Bau-
mann, P. D., Barati, M., Rane, M. J., and Lederer, E. (2005) J. Am. Soc.Nephrol. 16, 2598–2607
36. Zhang, S. X., Gozal, D., Sachleben, L. R., Jr., Rane, M., Klein, J. B., andGozal, E. (2003) FASEB J. 17, 1709–1711
37. Gozal, E., Sachleben, L. R., Jr., Rane, M. J., Vega, C., and Gozal, D. (2005)Am. J. Physiol. 288, C535–C542
38. Sato, S., Fujita, N., and Tsuruo, T. (2000) Proc. Natl. Acad. Sci. U. S. A. 97,10832–10837
39. Konishi, H.,Matsuzaki, H., Tanaka,M., Takemura, Y., Kuroda, S., Ono, Y.,and Kikkawa, U. (1997) FEBS Lett. 410, 493–498
40. Zhang, Q. S., Maddock, D. A., Chen, J. P., Heo, S., Chiu, C., Lai, D., Souza,K., Mehta, S., and Wan, Y. S. (2001) Int. J. Oncol. 19, 1057–1061
41. Souza, K., Maddock, D. A., Zhang, Q., Chen, J., Chiu, C., Mehta, S., andWan, Y. (2001)Mol. Med. 7, 767–772
42. Baudhuin, L. M., Cristina, K. L., Lu, J., and Xu, Y. (2002)Mol. Pharmacol.62, 660–671
43. Shiue, H., Musch, M. W., Wang, Y., Chang, E. B., and Turner, J. R. (2005)J. Biol. Chem. 280, 1688–1695
44. Cabane, C., Coldefy, A. S., Yeow, K., and Derijard, B. (2004) Cell. Signal.16, 1405–1415
45. Cappellini, A., Tazzari, P. L., Mantovani, I., Billi, A. M., Tassi, C., Ricci, F.,Conte, R., and Martelli, A. M. (2005) Apoptosis 10, 141–152
46. Nakajima, K., Hirade, K., Ishisaki, A., Matsuno, H., Suga, H., Kanno, Y.,Shu, E., Kitajima, Y., Katagiri, Y., and Kozawa, O. (2005) Life Sci. 77,96–107
47. Morissette,M. R., Cook, S. A., Foo, S.,McKoy,G., Ashida,N., Novikov,M.,Scherrer-Crosbie, M., Li, L., Matsui, T., Brooks, G., and Rosenzweig, A.(2006) Circ. Res. 99, 15–24
48. Lavoie, J. N., Lambert, H., Hickey, E., Weber, L. A., and Landry, J. (1995)Mol. Cell. Biol. 15, 505–516
49. Pivovarova, A. V., Mikhailova, V. V., Chernik, I. S., Chebotareva, N. A.,Levitsky, D. I., and Gusev, N. B. (2005) Biochem. Biophys. Res. Commun.331, 1548–1553
50. Arrigo, A. P., Virot, S., Chaufour, S., Firdaus, W., Kretz-Remy, C., andDiaz-Latoud, C. (2005) Antioxid. Redox. Signal. 7, 414–422
51. Mehlen, P., Kretz-Remy, C., Preville, X., and Arrigo, A. P. (1996) EMBO J.15, 2695–2706
52. Xanthoudakis, S., Roy, S., Rasper, D.,Hennessey, T., Aubin, Y., Cassady, R.,Tawa, P., Ruel, R., Rosen, A., and Nicholson, D. W. (1999) EMBO J. 18,2049–2056
53. Samali, A., Cai, J., Zhivotovsky, B., Jones, D. P., and Orrenius, S. (1999)EMBO J. 18, 2040–2048
54. Beere, H. M., Wolf, B. B., Cain, K., Kuwana, T., Tailor, P., Morimoto, R. I.,Cohen, G., and Green, D. R. (2000) Nat. Cell Biol. 2, 469–475
55. Li, C. Y., Lee, J. S., Ko, Y. G., Kim, J. I., and Seo, J. S. (2000) J. Biol. Chem.275, 25665–25671
56. Saleh, A., Srinivasula, S. M., Balkir, L., Robbins, P. D., and Alnemri, E. S.(2000) Nat. Cell Biol. 2, 476–483
57. Pandey, P., Saleh, A., Nakazawa, A., Kumar, S., Srinivasula, S. M., Kumar,V.,Weichselbaum, R., Nalin, C., Alnemri, E. S., and Kufe, D. (2000) EMBOJ. 19, 4310–4322
58. Schepers, H., Geugien,M., van der Toorn,M., Bryantsev, A. L., Kampinga,H. H., Eggen, B. J., and Vellenga, E. (2005) Exp. Hematol. 33, 660–670
59. Lee, Y. J., Lee, D. H., Cho, C. K., Bae, S., Jhon, G. J., Lee, S. J., Soh, J.W., andLee, Y. S. (2005) J. Biol. Chem. 280, 18108–18119
60. Andjelkovic, M., Jakubowicz, T., Cron, P., Ming, X. F., Han, J. W., andHemmings, B. A. (1996) Proc. Natl. Acad. Sci. U. S. A. 93, 5699–5704
Regulation of Akt Activation by Hsp27
21608 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 30 • JULY 27, 2007
by guest on June 23, 2018http://w
ww
.jbc.org/D
ownloaded from

and Madhavi J. RaneRui Wu, Hina Kausar, Paul Johnson, Diego E. Montoya-Durango, Michael Merchant
Scaffolding MK2 to Akt Signal ComplexHsp27 Regulates Akt Activation and Polymorphonuclear Leukocyte Apoptosis by
doi: 10.1074/jbc.M611316200 originally published online May 17, 20072007, 282:21598-21608.J. Biol. Chem.
10.1074/jbc.M611316200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/282/30/21598.full.html#ref-list-1
This article cites 59 references, 29 of which can be accessed free at
by guest on June 23, 2018http://w
ww
.jbc.org/D
ownloaded from