hoxc4, hoxc5, and hoxc6 expression in primary
TRANSCRIPT

American Journal ofPathology, Vol. 151, No. 4, October 1997Copyright American Society for Investigative Pathologv
HOXC4, HOXC5, and HOXC6 Expression in PrimaryCutaneous Lymphoid Lesions
High Expression of HOXC5 in Anaplastic Large-CellLymphomas
Janet J. Bijl,* Edgar Rieger,tJohan W. van Oostveen,*§Jan M. M. Walboomers,*§ Marja Kreike,*Rein Willemze,t and Chris J. L. M. Meijer*From the Departments ofPathology*, Hematology,5 andDermatology,t Vrije Universiteit Hospital, Amsterdam, The
Netherlands, and the Department ofDermatology,t University ofGraz, Graz, Austria
Homeobox (HOX) genes are involved in the lineage-specific differentiation of bone marrow stem cells.Recently, we reported a largely similar expressionpattern ofHOXC4 and HOXC6 in normal and neoplas-tic cells of the lymphoid lineage. In contrast, HOXC5was specifically expressed in a subset of B-cell non-
Hodgkin's lymphomas (B-NHL) but not in normallymphocytes or lymphoid leukemias. This might sug-
gest a role for HOXC5 in the pathogenesis of theselymphomas. In the present study the expression ofHOXC4, HOXC5, and HOXC6 in primary cutaneouslymphomas was investigated. Using RNA in situ hy-bridization (RISH) and semiquantitative reverse tran-scriptase polymerase chain reaction (RT-PCR), we
found strong expression ofHOXC4 and HOXC6 in all,except one, primary cutaneous lymphomas and allreactive cutaneous lymphoid infitrates. Interest-ingly, a strong expression of HOXC5 in primary ana-
plastic CD30+ large T-cell lymphomas was found.RISH was consistently negative for HOXC5 in all othertypes of primary cutaneous B- and T-cell lymphomas.However, by semiquantitative RT-PCR these lympho-mas showed a weak expression of HOXC5 mRNA.Therefore, we concluded that these lymphomas ex-
press low constitutive levels of HOXC5 mRNA. Fur-thermore, HOXC5 expression was consistently absentin reactive cutaneous lymphoid infiltrates, hyperplas-tic tonsils and lymph nodes, and peripheral bloodlymphocytes either unstimulated or stimulated by a
cocktail of CD3 and CD28 antibodies. As a strongexpression of HOXC5 in primary cutaneous lympho-mas was observed only in anaplastic large T-cell lym-phomas and reactive control tissues lacked HOXC5
expression, these data strongly support a role forHOXC5 in the genesis of anaplastic large-T-cell lym-phomas. (Amj Patbol 1997, 151:1067-1074)
Homeobox (HOX) genes are known to act as transcrip-tion factors,1-5 playing an essential regulatory role in thetemporal and spatial development of the embryo.267Over the last few years evidence has accumulated thatHOX genes are also crucial for the lineage-specific dif-ferentiation of bone marrow stem cells in hematopoie-SiS.8-15 Some genes of the HOXC cluster are found to beexpressed in lymphoid cells.16,17 In particular, an alter-native transcript of HOXC4 has been shown to be ex-
pressed only in lymphoid cells.16 Recently, using a spe-cific semiquantitative reverse transcriptase polymerasechain reaction (RT-PCR), we have shown that HOXC4and HOXC6 expression in the lymphoid lineage in-creases with differentiation.18 In contrast, expression ofHOXC5 could not be detected in normal lymphoid tissuesand leukemias but has been found in the B-cell non-
Hodgkin's lymphomas tested so far (B-NHL; n = 3) and inlymphoma-derived cell lines.16 Therefore, the questionwas raised whether HOXC5 is exclusively expressed insubsets of NHLs and thus might act as a putative onco-
gene in these lymphomas. We used primary cutaneouslymphomas as a test model, because the easy accessi-bility of a wide variety of benign cutaneous lymphoidlesions allowed us to check the HOX expression in non-neoplastic lymphoid cells located in the same body com-
partment. In addition, hyperplastic lymphoid organs were
analyzed for HOX expression. As HOX genes are sub-jected to cross-regulation and upstream regions of theHOXC5 gene have been shown to be a target for regu-lation by HOXC6 and some other homeoproteins,19 theexpression pattern of neighboring cluster genes HOXC4
Supported by the Dutch Cancer Society (grant IKA 90-17). E. Rieger is arecipient of an Erwin Schrbder fellowship of the Fonds zur Forderung derwissenschaftlichen Forschung, Vienna, Austria.
Accepted for publication July 15, 1997.
Address reprint requests to Dr. Chris J.L.M. Meijer, Department ofPathology, Vrije Universiteit Hospital, De Boelelaan 1117, 1081 HV Am-sterdam, The Netherlands.
1067

1068 Bijl et alAJP October 1997, Vol. 151, No. 4
and HOXC6 was also investigated. To exclude the pos-sibility that HOXC5 can be induced in vivo in lympho-cytes, we incubated peripheral blood mononuclear cells(PBMCs) with a cocktail of stimulating agents and testedHOXC4, HOXC5, and HOXC6 expression by RT-PCR atdifferent time intervals. The results show that HOXC5 isstrongly expressed only in primary CD30+ anaplasticlarge T-cell lymphomas (ALCLs), and thus seems to playa role in the pathogenesis of these lymphomas.
Materials and Methods
Tissue SamplesFormalin-fixed, paraffin-embedded specimens of primarycutaneous B- and T-cell lymphomas and benign infil-trates of the skin were collected from the archives of theDepartments of Pathology of the Vrije Universiteit Hospi-tal. Lymphomas were classified according to the Euro-pean Organization Research and Treatment of Cancerclassification for primary cutaneous lymphomas.20 TheALCLs used in this study showed CD30 expression andhad large clusters of blast cells, of which many hadtypical anaplastic morphology as described by Suchi etaI21 for lymph nodes and Kaudewitz et a122 for the skin.The group of primary cutaneous lymphomas includedtwo pleomorphic T-cell lymphomas, five mycosis fun-goides (MFs), one immunoblastic T-cell lymphoma, eightanaplastic ALCLs, seven large follicle center cell lympho-mas, and two immunoblastic B-cell lymphomas. In addi-tion, one cutaneous infiltrate of T-cell lympatic leukemiachronic was added. The benign group included six lichenplanus, three eczemas, and two parapsoriasis enplaques. In addition, five hyperplastic tonsils and threereactive lymph nodes were used as controls. Thesespecimens were all analyzed by RNA in situ hybridization(RISH). From nine cutaneous lymphomas (two MFs, threeALCLs, three large follicle center cell lymphomas, andone immunocytoma) and all hyperplastic lymph nodes,frozen material was available for RT-PCR. Freshly col-lected hyperplastic tonsils were divided in three parts.One part was used for isolation of lymphocytes bysqueezing them out of the tissue matrix with phosphate-buffered saline (PBS) over a wire netting, one part wassnap-frozen in liquid nitrogen, and the last part was for-malin-fixed and embedded in paraffin.
RISHA previously described RISH protocol was used. 18'23cDNA fragments of the HOXC4, HOXC5, and HOXC6genes (HOXC4, 437-bp Taql fragment from the region 5'to the homeobox; HOXC5, 271-bp EcoRI-Smal fragmentfrom the region 3' to the homeobox; HOXC6, 531-bp Taqlfragment from the region 3' to the homeobox) were kindlyprovided by Prof. Dr. E. Boncinelli (DIBIT, Istituto Scien-tifico H. San Raffaele, Milan, Italy) and cloned in pGEM-3transcription vectors (Promega, Madison, WI). Theseprobes have been tested in extensive Northern blot stud-
24-26 n-labeled antisense and sense RNAies. Digoxigenintranscripts were generated according to Melton et al,27using digoxigenin-1 1-UTP (Boehringer Mannheim, Mann-heim, Germany) and SP6, T3, or T7 RNA polymerases(Promega). Size, integrity, and relative concentration ofantisense and sense transcripts were checked by dena-turing gel electrophoresis followed by transfer to nylonmembranes and subsequent visualization by anti-digoxi-genin antibodies conjugated to alkaline phosphatase andchemiluminescence.28 Probes were calibrated based onthe number of incorporated digoxigenin-11-UTP. Four-micron paraffin-embedded sections were deparaffinizedin xylene, rehydrated in graded alcohols, treated withLugol's iodine and 3% sodium thiosulfate, and quenchedin 0.3% H202. Prehybridization treatments included 0.2mol/L HCI, 2X SSC at 700C, 10 ,tg/ml proteinase K for 30to 60 minutes, 4% formaldehyde with di-ethylpyrocarbon-ate (0.01%) and acetylation with acetic anhydride. Etha-nol-dried sections were hybridized for 3 hours at 550Cand additionally washed. Digoxigenin was detected by athree-step immunoperoxidase method (mouse anti-digoxigenin antibody, 1:250 (Sigma Chemical Co., St.Louis, MO), followed by incubation with a biotinylated ratanti-mouse Fab fragment, 1:500 (DAKO, Glostrup, Den-mark) and by a streptavidin-biotin complex, 1:200(DAKO)), resulting in visualization by diaminobenzidine(DAB) nickel (Ni) precipitate. Eventually, a silver en-hancement according to Merchenthaler et a129 was per-formed as described previously.30 Sections were lightlycounterstained with hematoxylin or nuclear fast red. Insome cases of tonsillar tissue, a double staining forHOXC6 by RISH and CD20 by immunohistochemistrywas performed. Indirect detection of RISH products us-ing fluorescence-tyramine was followed by an indirectimmunohistochemistry method using streptavidin-cya-nide 3 (Jackson Immunoresearch Laboratories, WestGrove, PA). Sections were counterstained with 4 jug/mI2-diazo-3,3,3-fluoro-propionic acid p-nitrophenyl ester(DAPI; Sigma) and judged using a fluorescence micro-scope (Leica type DMRB, Leica, Rijswijk, The Nether-lands) with a triple filter.
Negative controls included sense probes for eachspecimen and hybridization mix only. Sections of a num-ber of specimens were additionally preincubated with 1.2mg/ml RNAse A (Promega) to test RNA specificity of thesignal. RNA quality of the sections was tested with adigoxigenin-labeled RNA probe specific for human elon-gation factor-la (hEF-1a) mRNA.31 Specimens negativefor hEF-la were not included in the study.
RT-PCRThe RT-PCR protocol has been published in detail previ-ously.18'23 Briefly, total RNA was isolated from 10 serialcryostat sections of frozen specimens of hyperplastictonsils, reactive lymph nodes, and lymphomas using theRNAzol B method (CinnalBiotecx, Houston, TX). To avoidthe possible amplification of HOXC4, HOXC5, andHOXC6 mRNA of epidermal cells in skin localizations oflymphomas and benign infiltrates, the epidermis was re-

HOXC4, HOXC5, and HOXC6 in Cutaneous Lymphoid Lesions 1069AJP October 1997, Vol. 151, No. 4
moved by cutting off the dermis. This was confirmed bylight microscopy of the first and last hematoxylin andeosin (H&E)-stained section of the lesion. Primers andoligonucleotide probes (Perkin Elmer, Oudekerk a/d IJs-sel, The Netherlands) specific for the respective PCRproducts and located outside the homeobox region weredesigned with the PCRPLAN program of the PC/Genesoftware (IntelliGenetics, Mountain View, CA). Primerpairs for HOXC4, P1 BHOXC4 (forward) 5'-CGCCAGTAT-AGCTGCACCAGTTCTC-3', P2BHOXC4 (reverse) 5'-CGATCTCGATCCTTCTCCTTCGGG-3', HOXC5 P3HOXC5(forward) 5'-CAAACTTGTGGGAACTATGGATCGG-3',P4HOXC5 (reverse) 5'-GAGTCTGGTAGCGCGTGTA-ACTGG-3', HOXC6 P5HOXC6 (forward) 5'-TAGTTCT-GAGCAGGGCAGGACTGCG-3', P6HOXC6 (reverse)5'-CCGCTCCGTAGCCGACCCCACTGT-3', and their in-ternal oligomer probes 01 HOXC4 (internal oligomer)for HOXC4 5'-TCCCTCCAGCGCCGCCAGCAAG-3',02HOXC5 (internal oligomer) for HOXC5 5'-CAGCCAT-GTCTACCCCGTGGAGAG-3', and 03HOXC6 (internaloligomer) for HOXC6 5'-CCGCTCCGTAGCCGAC-CCCACTGT-3' give products of 375, 438, and 223 bp,respectively, and span an intron to allow distinction ofamplification products from cDNA and genomic DNA bysize. 19'32 RNA quality was controlled by using primers18to amplify UlA mRNA.33cDNA was synthesized from 1 ,ug of total RNA and
subsequently amplified for 40 cycles at 60°C annealingtemperature (GeneAmp PCR system 9600, Perkin Elmer,Branchburg, NJ). Amplification products were hybridizedwith internal 32P-labeled oligonucleotide probes understringent conditions to prove specificity of amplificationproducts. Omission of avian myeloblastosis virus-RT, ex-cluding amplification of possible pseudogenes, and dis-tilled water were used as negative controls. Additionally,a mature B cell line, JVM-3, was used as a positivecontrol for HOXC4, HOXC5, and HOXC6 amplification.
For a semiquantitative RT-PCR, four samples of a 1:10serial dilution of total RNA were transcribed to cDNAusing the antisense primer for HOXC5 together with thatof UlA. The subsequent PCR was performed for 25, 30,35, or 40 cycles.
Stimulation ofPBMCs
PBMCs were obtained from healthy volunteers. Wholeblood was centrifuged over a Ficoll Hypaque densitygradient (Pharmacia, Uppsala, Sweden). Separated PB-MCs were collected from the interphase, washed twicewith PBS supplemented with 1% bovine serum albumin(Organon Technika, Boxtel, The Netherlands). The cellswere counted and stimulated with a cocktail of anti-CD3and anti-CD28 monoclonal antibodies in the presence ofrecombinant interleukin-2 (rlL-2) as described earli-er.34'35 Cells were collected after a mild trypsinizationstep at 37°C and counted at 0, 1, 2, 4, 20, 48, 72, and 96hours. Total RNA was isolated with the RNAzol B method(Cinna/Biotecx), and RT-PCR was performed using a vol-
ume of RNA solution equivalent to 100,000 cells for eachsingle RT reaction.
Interpretation of ResultsWhen HOXC4, HOXC5, and HOXC6 mRNA was detectedby RISH analysis, it could always be confirmed by RT-PCR analysis. However, sometimes RISH results on rep-resentative lymphoma sections were negative, whereasRT-PCR results were positive, indicating that RT-PCR is amore sensitive technique. When expression was shownby both RT-PCR and RISH, neoplastic lymphoma cellswere considered to express HOXC4, HOXC5, or HOXC6.As reactive lymphocytes do not express HOXC5 mRNA,its expression in lymphomas observed only by RT-PCR,but not by RISH, was considered to be at a low expres-sion level. As reactive lymphoid cells are known to ex-press HOXC4 and HOXC6, lymphomas showing HOXC4and HOXC6 mRNA expression only by RT-PCR, but notby RISH, were considered to express very low levels ofHOXC4 and HOXC6 mRNA or as negative.
Results
HOXC4, HOXC5, and HOXC6 Expression inPrimary Cutaneous B- and T-NHL
Of the 27 primary cutaneous lymphomas tested by RISH,HOXC5 expression was exclusively shown in all (8/8)CD30+ ALCLs (Table 1). The signals were located in thecytoplasm of large anaplastic cells but not in small reac-tive cells (Figure lb). When present, all cells in the over-lying epidermis showed HOXC5 expression in their cyto-plasm (Figure 1 d). All 19 other primary cutaneous B- andT-cell lymphomas did not express HOXC5 by RISH. ByRT-PCR, performed on frozen material of nine primarycutaneous lymphomas, HOXC5 expression was detectedin eight of the nine cases (Table 1 and Figure 2), includ-ing MFs (cases 3 and 7), ALCLs (cases 11, 14, and 16),and three of the four cases of cutaneous follicle centercell lymphomas (cases 22, 23, and 25). To confirm thatthe HOXC5 expression levels detected by RISH analysisare higher in primary cutaneous CD30+ ALCLs than inother histological types of primary cutaneous B- andT-cell lymphomas, which are positive for HOXC5 only byRT-PCR, we performed a semiquantitative RT-PCR ontwo ALCLs (cases 1 1 and 16), two MFs (cases 3 and 7),and one large follicle center cell lymphoma (case 22;data not shown for cases 7, 16, and 22; Figure 3). Theexpression of housekeeping gene UlA in the serial dilu-tions of all lymphomas showed similar patterns, indicat-ing that the amount of input RNA was comparable (Figure3). A weak expression of HOXC5 in both ALCLs wasdetected in the two lowest dilutions after 25 cycles ofPCR, whereas no expression was found for the otherlymphomas. The two MFs and the follicle center celllymphoma showed weak signals in the lowest dilutionafter 30 cycles. At the same number of cell cycles,HOXC5 expression in the two ALCLs was detectable inthe first three dilutions. Detection of HOXC5 after 40

1070 BijI et alAJP October 1997, Vol. 151, No. 4
Table 1. Results of HOXC4, HOXC5, and HOXC6 Expression Detected by RNA In Situ Hybridization and RT-PCR in PrimaryCutaneous Lymphomas, Benign Lymphoid Skin Infiltrates, Hyperplastic Lymphoid Organs, and Peripheral BloodLymphocytes
Case Biopsy site Diagnosis HOXC4 HOXC5 HOXC6
T-cell lymphoma,cutaneous
123*4567*891011*121314*1516*17
B-cell lymphoma,cutaneous181920*2122*23*2425*2627
T-cell infiltrates ofthe skin2829303132333435363738
Reactive lymphoidtissue
3944*45*
46/47*4855*
BackUpper legBackHeadAbdomenHeadArmUpper armArmpitArmpitArmAbdomenAbdomenBackForearmFaceThorax
Lower legLower legNoseEarScapulaHipBackLipLower legBack
ForearmHandThighBackArmLower legShoulderArmUpper armBackBack
TonsilArmpitNeck
Pleomorphic medium/largePleomorphic medium/largeMF tumorMF tumorMF plaqueMF tumorMF plaqueImmunoblasticAnaplasticAnaplasticAnaplasticAnaplasticAnaplasticAnaplasticAnaplasticAnaplasticT-CLL
Follicle centerFollicle centerFollicle centerFollicle centerFollicle centerFollicle centerFollicle centerImmunocytomaImmunoblasticImmunoblastic
Lichen planusLichen planusLichen planusLichen planusLichen planusLichen planusEczemaEczemaEczemaParapsoriasis en plaquesParapsoriasis en plaques
Hyperplastic tonsilHyperplastic lymph nodeHyperplastic lymph nodePBMCs
+, strong staining; ±, weak staining; -, no staining with RISH.*Cases also analyzed with RT-PCR, indicated in parentheses.
cycles of PCR revealed signals in all four dilutions of RNAfrom both ALCLs, whereas HOXC5 expression could bedetected only in the first two or three dilutions of RNAfrom the two MFs and the follicle center cell lymphomasamples (Figure 3).HOXC4 was detected in 25 of 26 cases and HOXC6 in
24 of 26 cases of primary cutaneous lymphomas and in askin localization of a T-cell chronic lymphatic leukemia(Table 1). RT-PCR analysis on nine cutaneous lympho-mas disclosed HOXC4 mRNA in all and HOXC6 mRNAexpression in eight of nine cases.
HOXC4, HOXC5, and HOXC6 Expression inHyperplastic Lymph Nodes, Tonsils, BenignLymphoid Skin Infiltrates, and Peripheral BloodLymphocytes (PBLs)In contrast to the neoplastic cells in some lymphomas, alllymph nodes and lymphocytes isolated from hyperplastictonsils did not express HOXC5 as determined by RISH.Additionally, no HOXC5 expression was found in benignT cell infiltrates of the skin (Table 1). Epithelial cells of the
-+)+
+(+
+
++(++
+
+(++
+(++
+(++
+(++(+)+
+
-+)
-+)
+(++
+(+
+(+)
+(+
+
+(++
+(+
++(++(++
+(++(+
+-+
+
+
+-.+
+
+(++(++(+(+)
+++++++++++
(+) (-)

HOXC4, HOXC5, and HOXC6 in Cutaneous Lymphoid Lesions 1071AJP October 1997, Vol. 151, No. 4
4~~~~~~~~~~~4
-~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
Figure 1. RISH analysis for HOXC5 on two primary cutaneous anaplastic large T-cell lymphomas, using an antisense (b and d) and a sense probe (c and e). H&Estaining of a cutaneous anaplastic T-cell lymphoma is shown in a. Positive signals in the neoplastic cells can be seen as black DAB/Ni precipitates in the cytoplasm(b and d). Note that small reactive cells are negative for HOXC5 (b) and that overlying epithelial cells express HOXC5 mRNA. Magnification, X520.
1072 Bijl et alAJP October 1997, Vol. 151, No. 4
Normal tissues T-NHL B-NHLI --I I
I 11 i'1
.C'a ' 00
9 -j J'L mL<X M -a InI I~~~~~
HOXC4
HOXC5
HOXC6
UlA
-375bp
-438bp
-223bp
-221bp
Figure 2. Representative results from three to seven independent experi-ments of RT-PCR analysis showing expression of HOXC4, HOXC5, andHOXC6 in normal lymphoid tissues (lanes 1 to 3), T-NHL (lanes 4 to 8), andB-NHL (lanes 9 and 10). MF samples correspond with cases 3 and 7 of Table1. ALCLs correspond with cases 11, 14, and 16, and the follicle center celllymphomas (cb/ml diffuse) correspond with cases 22 and 23 of Table 1.Lymphoid B-cell line JVM-3 is used as a positive control for HOXC4, HOXC5,and HOXC6 and water (blanco) and omission of reverse transcriptase in allRT reactions (always negative, data not shown) as negative controls. ThesnRNP UlA gene is used as a control for the quality and the quantity of totalRNA used. Blots hybridized with specific 32P-labeled oligomer probes wereexposed for 24 hours.
skin and tonsil, however, showed HOXC5 expression andserved as an internal control for the hybridization condi-tions of the HOXC5 probe. Also by RT-PCR, HOXC5mRNA expression was not detected in hyperplasticlymph nodes, tonsillar lymphocytes, and PBMCs (Figure2).
Using RISH, HOXC4 and HOXC6 mRNA expressionwas found in all hyperplastic tonsils and lymph nodes.
,- 8o- 8 a o rl 8 X-
IV co oiC m v- o o enI I
ALCL 11
MF 3
ALCL 11
MF 3
30 cydes
40ys
i
HOXC5 UIAFigure 3. Results of a semiquantitative RT-PCR for HOXC5 (left panels) andcontrol gene UlA (right panels) on a serial dilution of total RNA (1, 0.1, 0.01,and 0.001 jig) from an ALCL (case 11) and a MF (case 3) sample. PCR resultsof 30 cycles (upper panels) and 40 cycles (lower panels) are shown. Notethat signals for HOXC5 in the ALCL sample are visible after fewer PCR cyclesand at higher dilutions than those in the MF sample.
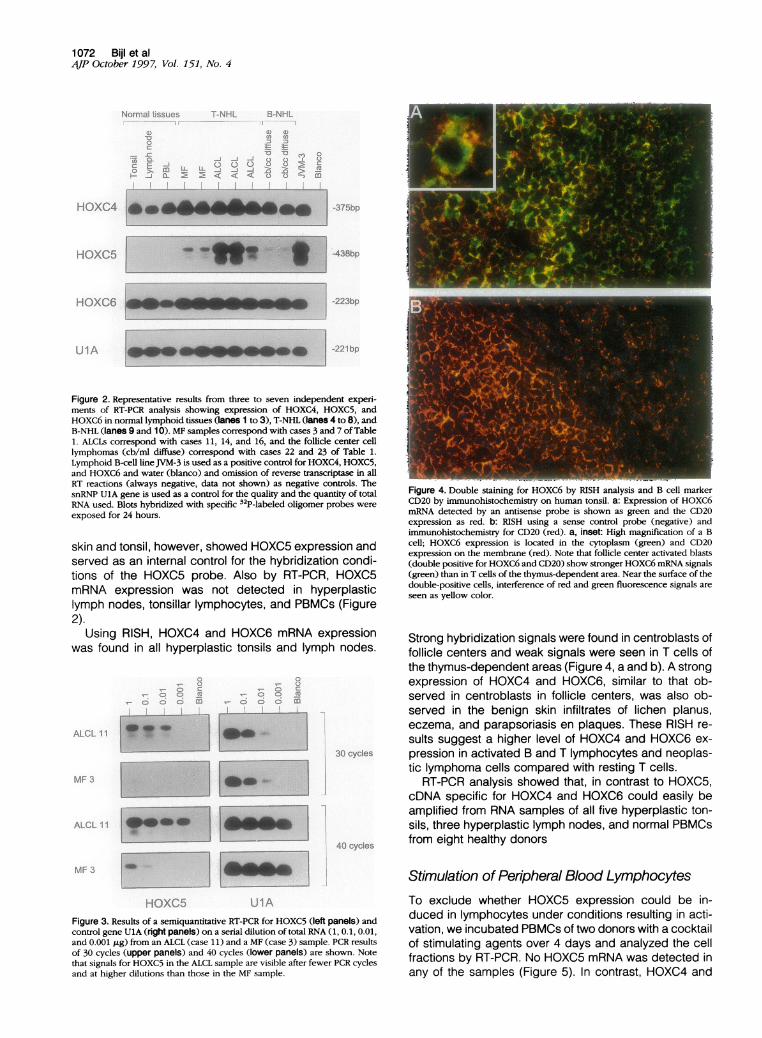
Figure 4. Double staining for HOXC6 by RISH analysis and B cell markerCD20 by immunohistochemistry on human tonsil. a: Expression of HOXC6mRNA detected by an antisense probe is shown as green and the CD20expression as red. b: RISH using a sense control probe (negative) andimmunohistochemistry for CD20 (red). a, inset: High magnification of a Bcell; HOXC6 expression is located in the cytoplasm (green) and CD20expression on the membrane (red). Note that follicle center activated blasts(double positive for HOXC6 and CD20) show stronger HOXC6 mRNA signals(green) than in T cells of the thymus-dependent area. Near the surface of thedouble-positive cells, interference of red and green fluorescence signals areseen as yellow color.
Strong hybridization signals were found in centroblasts offollicle centers and weak signals were seen in T cells ofthe thymus-dependent areas (Figure 4, a and b). A strongexpression of HOXC4 and HOXC6, similar to that ob-served in centroblasts in follicle centers, was also ob-served in the benign skin infiltrates of lichen planus,eczema, and parapsoriasis en plaques. These RISH re-sults suggest a higher level of HOXC4 and HOXC6 ex-pression in activated B and T lymphocytes and neoplas-tic lymphoma cells compared with resting T cells.
RT-PCR analysis showed that, in contrast to HOXC5,cDNA specific for HOXC4 and HOXC6 could easily beamplified from RNA samples of all five hyperplastic ton-sils, three hyperplastic lymph nodes, and normal PBMCsfrom eight healthy donors
Stimulation of Peripheral Blood LymphocytesTo exclude whether HOXC5 expression could be in-duced in lymphocytes under conditions resulting in acti-vation, we incubated PBMCs of two donors with a cocktailof stimulating agents over 4 days and analyzed the cellfractions by RT-PCR. No HOXC5 mRNA was detected inany of the samples (Figure 5). In contrast, HOXC4 and
-

HOXC4, HOXC5, and HOXC6 in Cutaneous Lymphoid Lesions 1073AJP October 1997, Vol. 151, No. 4
0 1 2 4 20 48 72 96 JVM-3
HOXC4
HOXC5
UlA
Figure 5. RT-PCR analysis of HOXC4, HOXC5, and control gene snRNP UlAexpression on unstimulated PBMCs (t = 0) and PBMCs incubated withanti-CD3/CD28 monoclonal antibodies and rIL-2 for 1, 2, 4, 20, 48, 72, and 96hours. Each PCR was performed on cDNA equivalent to 25,000 cells. B cellline JVM-3 was used as a positive control for HOXC4 and HOXC5 expression.
HOXC6 showed stronger RT-PCR signals in stimulatedthan in unstimulated PBMCs.
DiscussionIn this study we have shown by RISH that HOXC5 mRNAis expressed in all primary cutaneous CD30+ ALCLs butnot in other primary cutaneous B- and T-cell lymphomasor in reactive lymphoid skin infiltrates, hyperplastic lymphnodes, and tonsils.
By RT-PCR, HOXC5 expression was also found inother subsets of primary cutaneous B- and T-cell lympho-mas, but in addition to RISH, control lymphoid tissuesremained negative.
By semiquantitative RT-PCR analysis using varying cellcycles and different RNA dilutions we could confirmstrong expression of HOXC5 mRNA in CD30+ ALCLscompared with weak expression of HOXC5 mRNA inother primary cutaneous B- and T-cell lymphomas. Thelow expression levels of HOXC5 mRNA can be due to alow constitutive expression in the neoplastic cells. How-ever, although sections before and after those cut forRNA isolation were shown to be free from epithelial cellsand adnexal structures by light microscopy, we cannotcompletely exclude that these low expression levels ofHOXC5 by RT-PCR were derived from contaminatingepithelial cells, which were found to express HOXC5constitutively.When lymphomas are ranked according to their matu-
ration, ALCLs are considered to be the most matureones. The high expression of HOXC5 in these lymphomasis in agreement with increasing levels of HOXC5 mRNA inB- and T-cell lines with a higher degree of maturation.18These strongly HOXC5-expressing cell lines includedalso the CD30+ cell line Karpas 299.
The neoplastic cells in these ALCLs strongly expressCD30 and other activation markers (ie, IL-2 receptor,transferrin receptor (CD71), and HLA-DR). As these ana-plastic cells are associated with activation, PBLs were
stimulated with a cocktail of monoclonal antibodies to seewhether HOXC5 could be induced in these cells. How-ever, in contrast to the stronger signals for HOXC4 andHOXC6 in stimulated PBLs, HOXC5 expression was notinducible under these conditions or with either phytohe-magglutinin or interferon-y,18 indicating no relation withactivation. These observations suggest that HOXC5, dueto its overexpression, might play a role in the pathogen-esis of CD30+ ALCLs. Overexpression of HOXC5 mightbe caused by DNA amplifications or translocations in-volving region of the HOXC5 gene, or might be causedby deregulation of gene expression. However, amplifica-tions or translocations in the area where the HOXC clusteris designated, ie, chromosome 12q13.3,36 have to ourknowledge not been reported. Neither has the deregula-tion of HOXC gene expression in lymphomas been de-scribed.
In this respect, it is interesting to know what the func-tion of HOXC5 might be. Until now, no target genes ofHOXC5 have been identified. Some other HOX genes, ie,HOXD9, HOXC6, and HOXD3, have been shown to beable to regulate the expression of adhesion mole-cules.37-39 For example, HOXC6, the neighboring geneof HOXC5, can bind to promoter sequences in the N-CAM gene and is able to up-regulate the expression ofN-CAM protein.37 Moreover, by sequence homologysearches, we found that the DNA target motifs for HOXC6protein are also present in sequences of genes encodingother adhesion molecules such as VLA4 (CD49d/CD29),ICAM-1 (CD54), L-selectin (CD62L), and CD44,18 all ofwhich can be expressed on lymphoid cells. By regulatingthe expression of these morphoregulatory proteins, HOXgenes might influence the spreading and tissue position-ing of lymphomas. As HOXC5 is expressed only in sometissue-localized neoplastic equivalents of the lymphoidlineage (ie, some types of NHL) and not in the normalcells of the lymphoid lineage,18 we suggest that alsoHOXC5 plays a role in the adhesion molecule expression.
In contrast to HOXC5, by RISH, HOXC4 and HOXC6genes were found to be expressed in almost all primarycutaneous B- and T-NHLs, reactive cutaneous lymphoidinfiltrates, and hyperplastic tonsils and lymph nodes.These results are consistent with the earlier findings thatHOXC4 and HOXC6 are expressed from early to maturestages of lymphoid differentiation.18 The similar expres-sion pattern of HOXC4 and HOXC6 in anaplastic T-celllymphomas compared with the other lymphomas andlymphoid cells argues against a direct regulation ofHOXC5 by these genes in lymphoid cells.
In conclusion, our results suggest that the HOXC5gene is involved in the pathogenesis of primary cutane-ous CD30+ anaplastic T-cell lymphomas. Due to its rolein the formation of the body plan in embryogenesis, thequestion is raised whether HOXC5 expression is re-stricted to certain types of lymphomas or whether expres-sion is related to the site of origin of a lymphoma. There-fore, HOXC5 gene expression studies in lymphomasoriginated from different body sites are currently underinvestigation.

1074 Bijl et alAJP October 1997, Vol. 151, No. 4
AcknowledgmentsWe thank Prof. Dr. E. Boncinelli for providing the HOXcDNA clones. We are indebted to Drs. A. van den Brule,M. N. Jiwa, and J. Oudejans, Mrs. A. Horstman, and Mr.W. Vos, who have contributed to this work by stimulatingdiscussions, advice, and technical support.
References
1. Han K, Levine MS, Manley JL: Synergistic activation and repression oftranscription by Drosophila homeobox proteins. Cell 1989, 56:573-583
2. McGinnis W, Krumlauf R: Homeobox genes and axial patterning. Cell1992, 68:283-302
3. Schier AF, Gehring WJ: Functional specificity of the homeodomainprotein fushi tarazu: the role of DNA-binding specificity in vivo. ProcNatl Acad Sci USA 1993, 90:1450-1454
4. Gehring WJ, Qian YO, Billeter M, Furukubo-Tokunaga K, Schier A,Resendez-Perez D, Affolter M, Otting G, Wuthrich K: Homeodomain-DNA recognition. Cell 1994, 78:211-223
5. Pellerin I, Schnabel C, Catron KM, Abate C: Hox proteins have dif-ferent affinities for a consensus DNA site that correlate with thepositions of their genes on the hox cluster. Mol Cell Biol 1994, 14:4532-4545
6. Hunt P, Krumlauf R: Hox codes and positional specification in verte-brate embryonic axes. Annu Rev Cell Biol 1992, 8:227-256
7. Krumlauf R: Hox genes in vertebrate development. Cell 1994, 78:191-201
8. Magli MC, Barba P, Celetti A, De VG, Cillo C, Boncinelli E: Coordinateregulation of HOX genes in human hematopoietic cells. Proc NatIAcad Sci USA 1991, 88:6348-6352
9. Shen WF, Detmer K, Mathews CH, Hack FM, Morgan DA, Largman C,Lawrence HJ: Modulation of homeobox gene expression alters thephenotype of human hematopoietic cell lines. EMBO J 1992, 11:983-989
10. Blatt C, Lotem J, Sachs L: Inhibition of specific pathways of myeloidcell differentiation by an activated Hox-2.4 homeobox gene. CellGrowth Differ 1992, 3:671-676
11. Deguchi Y, Kirschenbaum A, Kehrl JH: A diverged homeobox gene isinvolved in the proliferation and lineage commitment of human he-matopoietic progenitors and highly expressed in acute myelogenousleukemia. Blood 1992, 79:2841-2848
12. Lawrence HJ, Largman C: Homeobox genes in normal hematopoiesisand leukemia. Blood 1992, 80:2445-2453
13. Kehrl JH: Homeobox genes in hematopoiesis. Crit Rev Oncol Hema-tol 1994, 16:145-156
14. Sauvageau G, Lansdorp PM, Eaves CJ, Hogge DE, Dragowska WH,Reid DS, Largman C, Lawrence HJ, Humphries RK: Differential ex-pression of homeobox genes in functionally distinct CD34+ subpopu-lations of human bone marrow cells. Proc Natl Acad Sci USA 1994,91:12223-12227
15. Lawrence HJ, Sauvageau G, Humphries RK, Largman C: The role ofHOX homeobox genes in normal and leukemic hematopoiesis. StemCells 1996, 14:281-291
16. Lawrence HJ, Stage KM, Mathews CHE, Detmer K, Scibienski R,MacKenzie M, Migliaccio E, Boncinelli E, Largman C: Expression ofHOX C homeobox genes in lymphoid cells. Cell Growth Differ 1993,4:665-669
17. Inamori K, Takeshita K, Chiba S, Yazaki Y, Hirai H: Identification ofhomeobox genes expressed in human T-lymphocytes. Biochem Bio-phys Res Commun 1993, 196:203-208
18. Bijl JJ, van Oostveen JW, Kreike M, Rieger E, Raaij-Helmer LMH,Walboomers JMM, Corte G, Boncinelli E, van den Brule AJC, MeijerCJLM: Expression of HOXC4, HOXC5, and HOXC6 in human lym-phoid cell lines, leukemias, and benign and malignant lymphoidtissue. Blood 1996, 87:1737-1745
19. Arcioni L, Simeone A, Guazzi 5, Zappavigna V, Boncinelli E, MavilioF: The upstream region of the human homeobox gene HOX3D is atarget for regulation by retinoic acid and HOX homeoproteins. EMBOJ 1992, 11:265-277
20. VWillemze R, Kerl H, Stery W, Berti E, Cerroni L, Chimanti 5, Diaz-Perez
JL, Geerts ML, Goos M, Knobler R, Ralfkiaer E, Santucci M, Smith N,Wechsler J, van Vloten WH, Meijer CJLM: EORTC classification forprimary cutaneous lymphomas: a proposal from the Cutaneous Lym-phoma Study Group of the European Organization for Research andTreatment of Cancer. Blood 1997, 90:354-371
21. Suchi T, Lennert K, Tu L-Y, Kikuchi M, Sato E, Stansfeld AG, FellerAC: Histopathology and immunohistochemistry of peripheral T-celllymphomas: a proposal for their classification. J Clin Pathol 1987,40:995-1015
22. Kaudewitz P, Stein H, Dallenbach F, Eckert F, Bieber K, Burg G,Braun-Falco 0: Primary and secondary cutaneous Ki-1 (CD30+) ana-plastic large cell lymphomas. Am J Pathol 1989, 135:359-367
23. Rieger E, Bijl JJ, van Oostveen JW, Soyer HP, Oudejans CBM, Jiwa NM,Walboomers JMM, Meijer CJLM: Expression of the homeobox geneHOXC4 in keratinocytes of normal skin and epithelial skin tumorsis correlated with differentiation. J Invest Dermatol 1994, 103:341-346
24. Cillo C, Barba P, Freschi G, Bucciarelli G, Magli MC, Boncinelli E:HOX gene expression in normal and neoplastic human kidney. Int JCancer 1992, 51:892-897
25. Celetti A, Barba P, Cillo C, Rotoli B, Boncinelli E, Magli MC: Charac-teristic patterns of HOX gene expression in different types of humanleukemia. Int J Cancer 1993, 53:237-244
26. De Vita G, Barba P, Odartchenko N, Givel J-C, Freschi G, BucciarelliG, Magli MC, Boncinelli E, Cillo C: Expression of homeobox-contain-ing genes in primary and metastatic colorectal cancer. Eur J Cancer1993, 29A:887-893
27. Melton DA, Krieg PA, Rebagliati MR, Maniatis T, Zinn K, Green MR:Efficient in vitro synthesis of biologically active RNA and RNA hybrid-ization probes from plasmids containing a bacteriophage SP6 pro-moter. Nucleic Acids Res 1984, 12:7035-7056
28. Bijl JJ, Rieger E, van Oostveen JW, Meijer CJLM, Oudejans CBM,Walboomers JMM: Quantification of biotinylated RNA probes for insitu hybridization using chemiluminescence. Histochemistry 1994,102:77-82
29. Merchenthaler I, Stankovics J, Gallyas F: A highly sensitive one-stepmethod for silver intensification of the nickel-diaminobenzidine endprod-uct of peroxidase reaction. J Histochem Cytochem 1989, 37:1563-1565
30. Mullink H, Vos W, Jiwa M, Horstman A, van der Valk P, WalboomersJMM, Meijer CJLM: Application and comparison of silver intensifica-tion methods for the diaminobenzidine and diaminobenzidine-nickelendproduct of the peroxidation reaction in immunohistochemistry andin situ hybridization. J Histochem Cytochem 1992, 40:495-504
31. Brands JHGM, Maassen JA, van Hemert FJ, Amons R, Moller W: Theprimary structure of the a subunit of human elongation factor. I.Structural aspects of guanidine-nucleotide-binding sites. Eur J Bio-chem 1986, 155:167-171
32. Simeone A, Pannese M, Acampora D, D'Esposito M, Boncinelli E: Atleast three human homeoboxes on chromosome 12 belong to thesame transcription unit. Nucleic Acids Res 1988, 16:5379-5390
33. Sillekens PTG, Habets WJ, Beijer RP, van Venrooij WJ: cDNA cloningof the human Ul snRNA-associated A protein: extensive homologybetween Ul- and U2-specific proteins. EMBO J 1987, 6:38-41
34. van Lier RAW, Brouwer M, de Jong R, Groot M, de Groot E, AardenLA: Functional properties of the human T-cell differentiation antigenCD28. Leucocyte Typing IV. Edited by Knapp W, Dorken B, Gilks WR,Rieber EP, Schmidt RE, Stein H, von dem Borne AEGK. Oxford,Oxford University Press, 1989, pp 353-355
35. Nijhuis EWP, van de Wiel-van Kemenade E, Figdor CG, van Lier RAW:Activation and expansion of tumour-infiltrating lymphocytes by anti-CD3 and anti-CD28 monoclonal antibodies. Cancer Immunol Immu-nother 1990, 32:245-250
36. Apiou F, Flagiello D, Cillo C, Malfoy B, Poupon MF, Dutrillaux B: Finemapping of human HOX gene clusters. Cytogenet Cell Genet 1996,73:114-115
37. Jones FS, Holst BD, Minowa 0, De REM, Edelman GM: Binding andtranscriptional activation of the promoter for the neural cell adhesionrpolecule by HoxC6 (Hox-3.3). Proc Natl Acad Sci USA 1993, 90:6557-6561
38. Taniguchi Y, Komatsu N, Moriuchi T: Overexpression of the HOX4A(HOXD3) homeobox gene in human erythroleukemia HEL cells resultsin altered adhesive properties. Blood 1995, 85:2786-2794
39. Goomer RS, Holst BD, Wood IC, Jones FS, Edelman GM: Regulationin vitro of an L-CAM enhancer by hcmeobox genes HoxD9 andHNF-1. Proc Natl Acad Sci USA 1994, 91 :7985-7989