hiv-1 szubtÍpusokra optimalizÁlt, szemÉlyre szabott...
TRANSCRIPT

1
ELTE TTK Biológia Doktori Iskola Doktori Iskola Vezető: Prof. Dr. Erdei Anna, DSc
Klasszikus és molekuláris genetika program Programvezető: Prof. Dr. Orosz László, DSc
HIV-1 SZUBTÍPUSOKRA OPTIMALIZÁLT, SZEMÉLYRE SZABOTT IMMUNTERÁPIA
Doktori értekezés
Somogyi Eszter
Témavezető: Dr. Lisziewicz Julianna, PhD Genetic Immunity Kft., Budapest
Budapest 2012

2
Tartalomjegyzék 1. Rövidítések ................................................................................................................... 4
2. Bevezetés ...................................................................................................................... 6
3. Irodalmi áttekintés és a téma háttere ............................................................................ 8
3.1 A HIV felépítése és életciklusa .................................................................................. 10
3.2 A HIV fertőzés immunológiája .................................................................................. 13
3.3 HIV diverzitás és epidemiológia ................................................................................ 17
3.4 A DermaVir kifejlesztése ........................................................................................... 19
3.4.1 A DermaVir pDNS alapú antigénje ........................................................................... 21
3.4.2 Az antigén DC-be juttatása patogén-szerű szintetikus nanorészecskével ................. 21
3.4.3 DermaPrep orvosi eszköz a nanomedicina bőrön át történő bejuttatására .............. 22
3.4.4 Preklinikai és klinikai eredmények ........................................................................... 23
4. Célkitűzések ............................................................................................................... 26
5. Anyagok és módszerek .............................................................................................. 27
5.1. Nanomedicina készítés ............................................................................................... 27
5.2. Western blot ............................................................................................................... 27
5.3. Immunprecipitáció ..................................................................................................... 27
5.4. Intracelluláris festés ................................................................................................... 28
5.5. VLP+ termelés és tisztítás ........................................................................................... 28
5.6. Transzmissziós elektronmikroszkópia ....................................................................... 28
5.7. Biológiai aktivitás mérés ............................................................................................ 28
5.8. Statisztikai analízis ..................................................................................................... 29
5.9. In silico T sejt epitóp repertoár analízis ..................................................................... 29
5.10. Filogenetikai analízis ................................................................................................. 29
5.11. pDNS konstrukciók készítése .................................................................................... 30
5.11.1. Szekvenciák................................................................................................................. 30
5.11.2. RNS kivonás ............................................................................................................... 30
5.11.3. Oligonukleotidok tervezése és szintézise .................................................................... 30
5.11.4. RT-PCR ...................................................................................................................... 30
5.11.5. Agaróz gélelektroforézis ............................................................................................ 31
5.11.6. DNS tisztítás gélből .................................................................................................... 31
5.11.7. Inzert és pDNS vektor előkészítése irányított ligáláshoz ........................................... 31
5.11.8. Hőkompetens E. coli kompetens sejt készítés ............................................................. 32
5.11.9. Ligálás és transzformálás .......................................................................................... 32
5.11.10. pDNS termelés ............................................................................................................ 32
5.11.11. Az új pDNS-esek azonosságának ellenőrzése ............................................................ 32
6. Eredmények ................................................................................................................ 33
6.1. A DermaVir in vitro jellemzése ................................................................................. 34
6.1.1. A pLWXu1 pDNS által expresszált fehérje repertoár ................................................ 34

3
6.1.2. VLP+ termelés ............................................................................................................ 36
6.1.3. Biológiai aktivitás ...................................................................................................... 37
6.2. A nanomedicina immunológiai potenciáljának in silico modellezése ....................... 40
6.2.1. T sejt epitóp repertoár analízis kidolgozása .............................................................. 40
6.2.2. Abszolút immunológiai potenciál (AIP) ..................................................................... 42
6.2.3. A DermaVir (pLWXu1) immunológiai potenciálja egy személyben modellezve ....... 43
6.2.4. Relatív immunológiai potenciál (RIP) ....................................................................... 44
6.2.5. Nem-B szubtípusú fertőzés esetén a B-szubtípus alapú terápia hatékonyságának in
silico modellezése ....................................................................................................... 46
6.3. Szubtípus-optimalzált pDNS konstrukciók tervezése és klónozása .......................... 49
6.3.1. A vakcinatervezést segítő új alkalmazás: az “immunológiai fa” .............................. 49
6.3.2. Szubtípus-optimalizált termékjelöltek immunológiai szempontú tervezése ............... 52
6.3.3. Az új pDNS-ek elkészítése és a termékjelöltek in silico jellemzése ............................ 53
6.3.4. Az új szubtípus-optimalizált termékjelöltek in vitro tesztelése .................................. 56
6.3.4.1. VLP+ termelés az új termékjelöltek által.................................................................... 56
6.3.4.2. Az új termékjelöltek biológiai aktivitása .................................................................... 59
6.3.5. A nanomedicina termékcsalád in vitro eredményeinek összefoglalása ..................... 59
7. Eredmények megvitatása ........................................................................................... 61
8. Összefoglalás .............................................................................................................. 69
9. Summary .................................................................................................................... 71
10. Publikációk ................................................................................................................. 72
11. Köszönetnyilvánítás ................................................................................................... 73
12. Irodalomjegyzék…………………………………………..…..…………………....74

4
1. Rövidítések
AIDS Szerzett immunhiányos tünetegyüttes (Acquired Immune Deficiency
Syndrome)
AIP Abszolút immunológiai potenciál
ANN Mesterséges neurális hálózat (Artificial Neural Network)
APC Antigén prezentáló sejt (Antigen presenting cell)
ARB Általános relatív kötés (Average Relative Binding)
ART Antiretrovirális terápia
as aminosav
bp bázispár
CA Kapszid
cART kombinált antiretrovirális terápia
CTL Citotoxikus T limfocita (Cytotoxic T lymphocyte)
DC Dendritikus sejt (Dendritic cell)
EMA Európai Gyógyszerengedélyezési Hivatal (European Medicines Agency)
Env Burokfehérje (Envelope)
FDA Amerikai Élelmiszer- és Gyógyszerengedélyezési Hivatal (Food and Drug
Administration)
HIV Emberi immunhiány vírusa (Human Immunodeficiency Virus)
HLA Humán leukocita antigén (Human leukocyte antigen)
IEDB Immunológiai Epitóp Adatbázis (Immune Epitope Database)
IN Integráz
Kbp kilobázispár
LC Langerhans sejt (Langerhans cell)
MA Mátrix
MHC Fő hisztokompatibilitási antigén (Major histocompatibility complex)
NC Nukleokapszid
Nef Negatív faktor
PCR Polimeráz láncreakció (Polymerase Chain Reaction)
pDNS plazmid DNS
Pol Polimeráz
PEIm Polietilénimin-mannóz
Rev Virion expressziós szabályozó (Regulator of virion expression)

5
RIP Relatív immunológiai potenciál
RT Reverz transzkriptáz fehérje
SIV Majom immunhiány vírusa (Simian Immunodeficiency Virus)
SMM Stabilizált mátrix módszer (Stabilized Matrix Method)
SU Felszíni fehérje (surface protein)
Tat transzaktiváló szabályozó fehérje (Transactivating regulatory protein)
TM Transzmembrán fehérje
Vif Virion fertőzési faktor (Virion infectivity factor)
VLP Vírusszerű részecske (Virus-like particle)
VLP+ Komplex vírusszerű részecske
Vpr R-vírusfehérje (Viral protein R)
Vpu U-vírusfehérje (Viral protein U)

6
2. Bevezetés
Ez a doktori munka a Genetic Immunity Kft. termékfejlesztési laboratóriumában
készült.
A HIV (Human Immunodeficiency Virus) fertőzés és a következtében kialakuló AIDS
(Acquired Immunodeficiency Syndrome) világszerte 34 millió fertőzöttet számláló
halálos betegség [1]. Egyelőre nem létezik ellene sem megelőző oltás, sem terápiás
vakcina, noha a vakcina fejlesztés egyik kiemelkedő fontosságú célterülete ez. Az
antiretrovirális gyógyszeres terápia számos mellékhatása, és a gyógyszer rezisztens
vírustörzsek kialakulása miatt sürgető feladat egy működő, és a szervezetre kevésbé
káros vakcina kifejlesztése.
A Genetic Immunity Kft. terápiás nanomedicina kifejlesztését tűzte ki célul a
HIV/AIDS kezelésére. (A nanomedicina a nanotechnológia orvosi alkalmazásának
összefoglaló megnevezése, és ez a kutatási terület többek között betegségek
megelőzésére, kezelésére, és diagnózisára keres megoldásokat [2].) A Genetic
Immunity első termékjelöltje, a DermaVir HIV Tapasz már a klinikai fejlesztés
második fázisában jár. A nanomedicina terápiás alkalmazásának célja de novo T sejtes
immunválasz kiváltása a kezelt beteg szervezetében, mely a HIV-vel fertőzött sejtek
pusztításával csökkenti a vírusterheltséget, ugyanakkor javítja az immunrendszer –ezzel
a fertőzött személy– általános állapotát.
A nanomedicina hatóanyaga egy patogén-szerű nanorészecske, ami HIV fehérjéket
kódoló plazmid DNS-ből (pDNS) és kationos szintetikus polimerből (polietilénimin-
mannóz, PEIm) áll, folyadék formulációban. A nanorészecske oldat célzott, bőrön át
történő “beadását”-, ezzel együtt az epidermiszben lévő antigén bemutató sejtekhez
(Langerhans sejtek, LC) irányítását a szintén saját fejlesztésű DermaPrep orvosi eszköz
teszi lehetővé. Az immunválasz kiváltásában kulcsfontosságú hatóanyag a pDNS, ami
számos HIV fehérjét (antigént) expresszál, az első termékjelölt esetében B szubtípusra
specifikusakat.
A HIV azonban rövid története ellenére az RNS vírusok gyors evolúciója miatt nagy
természetes diverzitással rendelkezik. Kilenc térbeli és időbeli elkülönülést mutató
szubtípusba (és számos rekombináns alcsoportba) sorolható, melyek között 10-35%
genetikai távolság állapítható meg [3].

7
Kérdésként merül fel, hogy az ilyen nagy divergenciát mutató szubtípusokkal fertőzött
betegek esetében mennyire lehet hatékony a B szubtípusra specifikus nanomedicina?
Ezért a doktori munka egyik célkitűzése a B szubtípusra specifikus termékjelölttel
kiváltható immunválasz in silico modellezésének kidolgozása, majd ezzel a
nanomedicina kereszt-reaktivitásának modellezése volt nem-B szubtípusú fertőzések
esetén. A kapott eredmények alapján döntöttünk egy szubtípus-optimalizált
termékcsalád kifejlesztése mellett. A feladat megvalósításához egy immunológiai
szempontú tervezést segítő eszközt is kifejlesztettünk. Ezenkívül fontos célunk volt a
klinikai tesztelés alatt álló DermaVir termékjelölt in vitro jellemzése, ehhez módszerek
fejlesztése, majd végül az új nanomedicina jelöltek klinikai vizsgálatokra alkalmas
minőségének in vitro igazolása.

8
3. Irodalmi áttekintés és a téma háttere
Létezik-e olyan antigén, ami a HIV fertőzés ellen véd? Ez a kérdés kutatók ezreit és
emberek (tíz)millióit foglalkoztatja 25 éve. Eddig senki nem adott egyértelmű választ a
kérdésre. Nincs megelőző védőoltás, mert a HIV számos védőmechanizmusa, valamint
genetikai változatossága miatt az eddigi összes antigén jelölt hatástalannak bizonyult. A
kutatás tovább folyik, de a sikertelenség miatt időközben előtérbe került a terápiás
vakcinák fejlesztése, hogy a világméretű járvány fertőzöttjeinek jobb életkilátásokat és
jobb életminőséget nyújtsanak. Amíg célt nem ér valaki, addig marad az antiretrovirális
terápia (ART), a vírus életciklusának különböző szakaszait gátló közel harmincféle
piacon lévő gyógyszerrel [4]. Az ART legnagyobb hátránya, hogy a vírus RNS
örökítőanyagának rendkívül gyors mutációs képessége miatt hamar kialakulnak a
gyógyszerrezisztens mutánsok. Épp ezért a gyógyszereket hármas kombinációban adják
(cART), és amikor az adott betegben hatástalanná válik, lecserélik egy újabb
gyógyszerkombinációra. Sajnos sok HIV fertőzött eljut odáig, hogy már nincs mire
lecserélni a gyógyszereit.
A terápiás HIV vakcina tervezés esetén is az egyik legnagyobb kihívás a megfelelő
antigén készlet kiválasztása, amely hatékonyan képes HIV-specifikus T sejteket
indukálni, hogy azok sikeresen elpusztíthassák a vírussal fertőzött sejteket a beteg
szervezetében, ezzel fékezve a betegség előrehaladását. Minél szélesebb az antigén
repertoár, annál többféle antigén-specifikus T sejt termelődése indukálható, és annál
hatékonyabb lehet a vakcina [5]. A gyengített vírust tartalmazó vakcinák ezért sikeresek
sok kórokozó elleni védelemben (pl. mumpsz, kanyaró, rubeola). Sőt, már a SIV
(majom immundeficiencia vírus) esetében is kifejlesztettek hosszú távú védettséget
biztosító, gyengített kórokozót tartalmazó oltóanyagot [6]. Ez bíztató eredmény arra
nézve, hogy esetleg emberben is indukálható tartós, a vírus replikációt kordában tartó
immunválasz a HIV ellen. Azonban a gyengített HIV vírus vakcinaként történő
alkalmazása túl sok biztonsági kockázatot rejt [7].
Az elterjedten használt alegység vakcinák ezzel szemben csak egy vagy néhány
antigént tartalmaznak. Ezek újabb alternatívája a VLP (vírusszerű részecske) vakcina,
ami spontán összeszerelődő kapszidfehérjékből, valamint esetenként burokfehérjékből
áll, és nagy előnye, hogy az alkotó fehérjéket természetes konformációban tartalmazza
a leghatékonyabb immunválasz kiváltása érdekében. Már kereskedelmi forgalomban is

9
kaphatók ilyen VLP vakcinák, pl. a magas kockázatú humán papillomavírus törzsek
elleni megelőző vakcinák (Gardasil® és Cervarix™) [8,9]. Eddig azonban a VLP
vakcinák terápiás felhasználása nem volt sikeres (vagy fel sem merült), mivel nem
váltanak ki a terápiás vakcinák hatékonyságához szükséges T sejtes immunválaszt [10].
Néhány kivételes esetben megfigyeltek már mind a fertőzéstől megvéd, mind terápiás
szempontból hatékony immunválaszt HIV-fertőzött betegekben [11,12,13,14]. Ezen
esetek jelzik, hogy a HIV virion jó antigén, ami ellen létezhet hatékony természetes
immunválasz. Épp ezért a széles antigén repertoár és a virion felszíni
struktúrfehérjéinek együttes bevetése immunogénként működő vakcinát
eredményezhet, amit már többféle vakcinafejlesztési elképzelésben is felhasználtak.
Többen is készítettek kémiailag módosított-, vagy replikációs mutáns HIV
vírustörzseket, de a klinikai vizsgálatokra vonatkozó kérelmeiket nem fogadták el a
gyógyszerengedélyezési hatóságok, mert a vakcina-jelöltek nem bizonyultak eléggé
biztonságosnak [15, 16].
Az egyéb vírus-vektor alapú vakcinák fő hátránya, hogy a vektorral szembeni korábbi
védettség miatt nem alkalmazhatók mindenkinél, és terápiás célú (többszöri)
alkalmazásuk is korlátozott a vektor antigének ellen kialakuló immunválasz miatt, amik
közömbösíthetik a vakcina hatását. A Merck STEP klinikai kísérletében például
Adenovírus 5 vírus-vektor alapú megelőző vakcinát használtak magas fertőzési
kockázatú HIV negatív egyének bevonásával. Sajnos azonban a kezelt csoportban
magasabb volt a HIV-fertőzések száma a korábban Adenovírus fertőzésen átesett
személyek vírusvektorral szembeni immunitása miatt, mint a kontroll csoportban [17].
A nem vírus-vektor alapú DNS vakcinákat is azzal a céllal fejlesztik, hogy számos
antigén ellen váltsanak ki velük immunválaszt. Előnyük, hogy egy DNS konstrukció
sok antigént expresszálhat, ezáltal széles repertoárú patogén-specifikus immunválaszt
válthat ki, emellett a körültekintő és többszörös biztonsági módosítások könnyen
bevihetők a DNS-be. Így megszüntethetők az eredeti kórokozó integrációs-, és onkogén
tulajdonságai, valamint megakadályozható, hogy a vakcinaként felhasznált módosított
örökítőanyag replikálódjon, ill. rekombinálódjon vad vírustörzsekkel.
A Genetic Immunity laboratóriumában olyan terápiás HIV vakcina kifejlesztését tűztük
ki célul, ami HIV specifikus T sejtek széles repertoárjának képződését képes kiváltani,
ezáltal a HIV fertőzés ill. az AIDS lefolyását lassítani. A DermaVir terápiás vakcina
pDNS hatóanyaga a tizenöt HIV fehérje közül tizenhármat teljes hosszúságában-, kettőt

10
pedig részlegesen kódol és expresszál a célzott antigén bemutató sejtekben (APC). A
pDNS célba juttatását lehetővé tevő nanorészecske formuláció-, valamint a bőrben lévő
APC-k elérését szolgáló “DermaPrep” bőr előkészítő és kezelési eljárás fontos részét
képezik az alább kifejtett hatásmechanizmus kiváltásának. Előbb azonban tekintsük át
röviden a HIV életciklusát, immunológiáját és epidemiológiáját!
3.1 A HIV felépítése és életciklusa
A HIV (Human Immunodeficiency Virus) a Retroviridae család Lentivirus genusába
tartozik, többek között a majom immunhiány vírusával (SIV) együtt. A 120 nm
átmérőjű burkos RNS vírus lassú lefolyású halálos fertőzést okoz. A HIV genomját két
azonos, egyszálú pozitív irányultságú, 9,5 kilobázis méretű RNS alkotja, ami a
következő vírusalkotó fehérjéket expresszálja (1. ábra) [18]: Gag poliprotein (mely a
vírus érése során tovább hasítódik mátrix-MA, kapszid-CA és nukleokapszid-NC
fehérjékké), Pol poliprotein (melyet saját enzimaktivitása hasít proteáz-PR, reverz
transzkriptáz-RT és integráz-IN fehérjékre), valamint az Env 160 kDa méretű
poliprotein (amit a gazdasejt proteázai gp120 felszíni fehérje alegységre-SU és a pg41
transzmembrán fehérje alegységre-TM hasítanak, és ezek trimerei alkotják a vírus
felszíni gligoproteinjét). A szerkezeti fehérjéken kívül szabályozó fehérjéket is kódol a
genom: Tat és Rev, valamint a Nef, Vif, Vpr és Vpu járulékos fehérjéket [19].
1. ábra. A HIV virion felépítésének sematikus ábrája.
Integráz (Pol-IN)
vírus genom
Kapszid (Gag-CA)Nukleokapszid
(Gag-NC)
Proteáz (Pol-PR)
Transzmembrán
burokfehérje (Env-TM)
Felszíni burokfehérje
(Env-SU)
Reverz transzkriptáz
(Pol-RT)
Vpr
kettős lipidréteg
Mátrix (Gag-MA)

11
A vírus szexuális úton a nyálkahártya mikrosérülésein át, vagy vérrel, ill. vertikálisan
anyáról magzatra, illetve újszülöttre (anyatejjel) terjed [20]. Sikeres fertőzés az után
jöhet létre, hogy a szervezetbe került virionok bejutnak a szaporodásuk helyszínéül
szolgáló immunsejtekbe: elsősorban a CD4 receptorral rendelkező T limfocitákba,
esetleg dendritikus sejtekbe (DC) ill. makrofágokba. Először általában nyugvó CD4+ T
sejteket fertőz a vírus, majd ezekben intenzív reprodukciós ciklussal megsokszorozza a
vírusmennyiséget (ez a fertőzés akut fázisa, amit enyhe mononukleózis-szerű tünetek
kísérhetnek). A T sejt CD4+ receptorán és koreceptorán (CCR5 vagy CXCR4) az Env-
SU felszíni glikoproteinjének segítségével rögzül a virion. Majd konformáció változás
után az Env-TM fehérje utat nyit a sejtbe: a vírus burka a gazdasejt membránjába olvad,
a kapszid pedig bejut a citoplazmába (2.ábra) [21]. Ezután a kapszid is lebomlik, és
szabaddá válik a vírusgenom, megkezdődik a replikáció. Az RT fehérje révén az RNS-
ről RNS-DNS hibrid, majd kettős szálú DNS másolat készül, mely gyűrű
konformációjú preintegrációs komplexként az IN fehérje segítségével a gazdasejt
genomjába integrálódik [22]. A vírus ezen integrálódott formája a provírus, és
amennyiben a fertőzött T sejtből memória T sejt alakul ki, ez a látens fertőzés hosszú
ideig fennmaradhat, a vírus rezervoárját (ill. annak egy részét) alkotva [23,24]. Ha a
gazdasejt aktiválódik ― vagyis mitózis történik, RNS polimeráza kis mennyiségben
mRNS-eket szintetizál az integrálódott HIV DNS-ről, és ezekről korai szabályozó
fehérjék expresszálódnak [25]. Előbbi (“transzaktivátor fehérje”, a transzkripció fő
virális szabályozó fehérjéje) a genom LTR promóterérnek TAR (“Transactivating
Responsive Region”) szabályozó régiójának transzkriptumához kötődve transzkripciós
iniciátorként és elongációs faktorként fokozza az mRNS-ek képződését (3. ábra)
[25,25,26]. A Rev az RRE (“Rev Responsive Element”) szabályozó régióhoz kötődve a
képződött teljes hosszúságú és a részlegesen hasított mRNS-ek sejtplazmába történő
exportját-, ezzel a HIV fehérjék transzlációját segíti elő [27]. A termelődő vírus
fehérjék a teljes hosszúságú mRNS két kópiáját is becsomagolva éretlen HIV
részecskékké szerelődnek össze a sejthártyát is integrálva annak belső oldalán, majd a
folyamat a sejthártya lefűződésével és ezzel a termelődött HIV vírusok sejtből való
kijutásával zárul. Utolsó lépésként transzlációt követő fehérje hasítással a Gag
poliproteinből származó fehérjék is előállítódnak, felveszik végleges térszerkezetüket és
a Gag-CA fehérjékből kialakul a kapszid, ami az érett virion morfológiájának ismérve
[28].

2
3: transzkripció, 4: a virális DNS integrációja a gazdasejt genomjába, 5: fehérjeszintézis és új
vírusok összesz
3
starthelyt
ábra bal alsó rész
TAR
2. ábra.
3: transzkripció, 4: a virális DNS integrációja a gazdasejt genomjába, 5: fehérjeszintézis és új
vírusok összesz
3. ábra.
starthelyt
ábra bal alsó rész
TAR
. ábra.
3: transzkripció, 4: a virális DNS integrációja a gazdasejt genomjába, 5: fehérjeszintézis és új
vírusok összesz
. ábra.
starthelyt
ábra bal alsó rész
TAR RNS szürk
. ábra. A HIV életciklusának áttekintése
3: transzkripció, 4: a virális DNS integrációja a gazdasejt genomjába, 5: fehérjeszintézis és új
vírusok összesz
. ábra.
starthelytő
ábra bal alsó rész
RNS szürk
A HIV életciklusának áttekintése
3: transzkripció, 4: a virális DNS integrációja a gazdasejt genomjába, 5: fehérjeszintézis és új
vírusok összesz
. ábra. A HIV
starthelytől közvetlenül
ábra bal alsó rész
RNS szürk
A HIV életciklusának áttekintése
3: transzkripció, 4: a virális DNS integrációja a gazdasejt genomjába, 5: fehérjeszintézis és új
vírusok összeszerel
A HIV
ől közvetlenül
ábra bal alsó rész
RNS szürk
CD4+ T sejt
A HIV életciklusának áttekintése
3: transzkripció, 4: a virális DNS integrációja a gazdasejt genomjába, 5: fehérjeszintézis és új
erelő
A HIV
l közvetlenül
ábra bal alsó részén látható
RNS szürkével jelölt régiójához köt
CD4+ T sejt
A HIV életciklusának áttekintése
3: transzkripció, 4: a virális DNS integrációja a gazdasejt genomjába, 5: fehérjeszintézis és új
erelődése, 6: új vírusok lef
A HIV genom és promóter
l közvetlenül
én látható
ével jelölt régiójához köt
CD4+ T sejt
A HIV életciklusának áttekintése
3: transzkripció, 4: a virális DNS integrációja a gazdasejt genomjába, 5: fehérjeszintézis és új
ődése, 6: új vírusok lef
genom és promóter
l közvetlenül downstream irányban helyezkedik el a TAR szabályozó régió, ami az
én látható
ével jelölt régiójához köt
HIV
CD4+ T sejt
A HIV életciklusának áttekintése
3: transzkripció, 4: a virális DNS integrációja a gazdasejt genomjába, 5: fehérjeszintézis és új
dése, 6: új vírusok lef
genom és promóter
downstream irányban helyezkedik el a TAR szabályozó régió, ami az
én látható „
ével jelölt régiójához köt
HIV virion
CD4+ T sejt
RNS/DNS
Virális
dsDNS
A HIV életciklusának áttekintése
3: transzkripció, 4: a virális DNS integrációja a gazdasejt genomjába, 5: fehérjeszintézis és új
dése, 6: új vírusok lef
genom és promóter
downstream irányban helyezkedik el a TAR szabályozó régió, ami az
„stem
ével jelölt régiójához köt
virion
1
RNS/DNS
Virális
dsDNS
A HIV életciklusának áttekintése
3: transzkripció, 4: a virális DNS integrációja a gazdasejt genomjába, 5: fehérjeszintézis és új
dése, 6: új vírusok lef
genom és promóter
downstream irányban helyezkedik el a TAR szabályozó régió, ami az
stem-loop
ével jelölt régiójához köt
virion
RNS/DNS
Virális
dsDNS
A HIV életciklusának áttekintése
3: transzkripció, 4: a virális DNS integrációja a gazdasejt genomjába, 5: fehérjeszintézis és új
dése, 6: új vírusok lef
genom és promóter
downstream irányban helyezkedik el a TAR szabályozó régió, ami az
loop
ével jelölt régiójához köt
2
RT
A HIV életciklusának áttekintése
3: transzkripció, 4: a virális DNS integrációja a gazdasejt genomjába, 5: fehérjeszintézis és új
dése, 6: új vírusok lefű
genom és promóter
downstream irányban helyezkedik el a TAR szabályozó régió, ami az
loop” struktúrájú RNS
ével jelölt régiójához kötődik.
Vírus
RT
A HIV életciklusának áttekintése. 1: Vírus köt
3: transzkripció, 4: a virális DNS integrációja a gazdasejt genomjába, 5: fehérjeszintézis és új
dése, 6: új vírusok lefűződése.
vázlatos
downstream irányban helyezkedik el a TAR szabályozó régió, ami az
struktúrájú RNS
ével jelölt régiójához kötődik.
Vírus
1: Vírus köt
3: transzkripció, 4: a virális DNS integrációja a gazdasejt genomjába, 5: fehérjeszintézis és új
űződése.
vázlatos
downstream irányban helyezkedik el a TAR szabályozó régió, ami az
struktúrájú RNS
ődik.
3
4
Vírus ssRNS
1: Vírus köt
3: transzkripció, 4: a virális DNS integrációja a gazdasejt genomjába, 5: fehérjeszintézis és új
ődése.
vázlatos
downstream irányban helyezkedik el a TAR szabályozó régió, ami az
struktúrájú RNS
4
ssRNS
m
1: Vírus kötő
3: transzkripció, 4: a virális DNS integrációja a gazdasejt genomjába, 5: fehérjeszintézis és új
vázlatos térképe
downstream irányban helyezkedik el a TAR szabályozó régió, ami az
struktúrájú RNS
mRNS
1: Vírus kötődés és sejtbe jutás, 2: kicsomagolódás,
3: transzkripció, 4: a virális DNS integrációja a gazdasejt genomjába, 5: fehérjeszintézis és új
térképe
downstream irányban helyezkedik el a TAR szabályozó régió, ami az
struktúrájú RNS-
Sejtmag
Provírus
RNS
ődés és sejtbe jutás, 2: kicsomagolódás,
3: transzkripció, 4: a virális DNS integrációja a gazdasejt genomjába, 5: fehérjeszintézis és új
térképe. A promóter
downstream irányban helyezkedik el a TAR szabályozó régió, ami az
-t kódolja.
Sejtmag
Provírus
dés és sejtbe jutás, 2: kicsomagolódás,
3: transzkripció, 4: a virális DNS integrációja a gazdasejt genomjába, 5: fehérjeszintézis és új
. A promóter
downstream irányban helyezkedik el a TAR szabályozó régió, ami az
t kódolja.
Sejtmag
Provírus
dés és sejtbe jutás, 2: kicsomagolódás,
3: transzkripció, 4: a virális DNS integrációja a gazdasejt genomjába, 5: fehérjeszintézis és új
. A promóter
downstream irányban helyezkedik el a TAR szabályozó régió, ami az
t kódolja.
5
dés és sejtbe jutás, 2: kicsomagolódás,
3: transzkripció, 4: a virális DNS integrációja a gazdasejt genomjába, 5: fehérjeszintézis és új
. A promóter
downstream irányban helyezkedik el a TAR szabályozó régió, ami az
t kódolja. A Tat szabályozó fehérje a
Új HIV
6
dés és sejtbe jutás, 2: kicsomagolódás,
3: transzkripció, 4: a virális DNS integrációja a gazdasejt genomjába, 5: fehérjeszintézis és új
. A promóterben a
downstream irányban helyezkedik el a TAR szabályozó régió, ami az
A Tat szabályozó fehérje a
Új HIV
dés és sejtbe jutás, 2: kicsomagolódás,
3: transzkripció, 4: a virális DNS integrációja a gazdasejt genomjába, 5: fehérjeszintézis és új
ben a
downstream irányban helyezkedik el a TAR szabályozó régió, ami az
A Tat szabályozó fehérje a
Új HIV
dés és sejtbe jutás, 2: kicsomagolódás,
3: transzkripció, 4: a virális DNS integrációja a gazdasejt genomjába, 5: fehérjeszintézis és új
ben a
downstream irányban helyezkedik el a TAR szabályozó régió, ami az
A Tat szabályozó fehérje a
dés és sejtbe jutás, 2: kicsomagolódás,
3: transzkripció, 4: a virális DNS integrációja a gazdasejt genomjába, 5: fehérjeszintézis és új
transzkripciós
downstream irányban helyezkedik el a TAR szabályozó régió, ami az
A Tat szabályozó fehérje a
dés és sejtbe jutás, 2: kicsomagolódás,
3: transzkripció, 4: a virális DNS integrációja a gazdasejt genomjába, 5: fehérjeszintézis és új
transzkripciós
downstream irányban helyezkedik el a TAR szabályozó régió, ami az
A Tat szabályozó fehérje a
dés és sejtbe jutás, 2: kicsomagolódás,
3: transzkripció, 4: a virális DNS integrációja a gazdasejt genomjába, 5: fehérjeszintézis és új
transzkripciós
downstream irányban helyezkedik el a TAR szabályozó régió, ami az
A Tat szabályozó fehérje a
12
dés és sejtbe jutás, 2: kicsomagolódás,
3: transzkripció, 4: a virális DNS integrációja a gazdasejt genomjába, 5: fehérjeszintézis és új
transzkripciós
downstream irányban helyezkedik el a TAR szabályozó régió, ami az
A Tat szabályozó fehérje a
12
dés és sejtbe jutás, 2: kicsomagolódás,
3: transzkripció, 4: a virális DNS integrációja a gazdasejt genomjába, 5: fehérjeszintézis és új
transzkripciós
downstream irányban helyezkedik el a TAR szabályozó régió, ami az
A Tat szabályozó fehérje a

13
3.2 A HIV fertőzés immunológiája
A HIV fertőzés immunológiai alapjainak áttekintéséhez a klinikusok számára készített
UpToDate online tudásbázist használom vezérfonalként [29]. A HIV leggyakrabban az
anogenitális nyálkahártyán át fertőz, és az előző bekezdésben leírt módon jut be a CD4
receptorral rendelkező immunsejtekbe. A célsejtbe jutáshoz a gp120 vírusfehérje és a
célsejt CD4 receptorának kapcsolódásán kívül a CCR5 vagy a CXCR4 koreceptor
keresztkötése is szükséges. A vírus koreceptor használata meghatározza a célsejtek
fajtáját: a CCR5 koreceptor segítségével bejutó HIV törzsek makrofág-, ill. DC
tropikusak, a CXCR4 koreceptor pedig a T sejt tropikus HIV bejutásában játszik
szerepet, de vannak olyan vírusok is, amik mindkét koreceptorral képesek kapcsolódni
[30]. Friss (ún. elsődleges) fertőzést általában makrofág/DC tropikus vírus hoz létre. A
leggyakoribb a DC antigénbemutató sejt fertőzése, produktív vagy nem produktív
módon. A nem produktív fertőzés eredményeképp a vírus a DC-SIGN receptorhoz
kapcsolódva bejut a sejtbe, de nem replikálódik, hanem a DC segítségével a
nyirokcsomóba jut és ott CD4+ T sejteket fertőz [31,32]. A vírus két nap múlva már
kimutatható a közeli nyirokcsomókban, további három nap múlva pedig a vérben is
[33]. Ezután a fertőzés szétterjed az egész szervezetben.
A következő, akut fertőzési szakasz néhány hetében többnyire mononukleózis-szerű
tünetekkel kísérve (láz, nyirokcsomó-duzzanat), esetleg tünetmentesen a vírus gyorsan
replikálódik (4. ábra) [34]. Ezzel egy időben a CD4+ T sejt szám csökken, a CD8+ T
sejt szám pedig emelkedik, de mindkettő hamar visszaáll az eredetihez közeli szintre. A
CD4+ T sejt szám csökkenés, valamint a Th17 (szabályozó) T sejt szám csökkenés az
emésztőrendszer mentén lévő nyirokcsomókban lokális immunhiányt okoz, ennek
következtében a bélfal nyálkahártyáján át baktériumok kerülnek a szervezetbe, ami
krónikus immun-aktivációt eredményez [35,36]. Az immun-aktivációban természetesen
szerepet játszik az intenzív HIV replikáció is.

14
4. ábra. A HIV fertőzés tipikus időbeli lefolyása a CD4+ T sejt szám és a vérplazmában
mérhető HIV RNS mennyiségének függvényében.
Az akut HIV fertőzési szakaszban a természetes ölősejtek (NK) közvetlen lízissel, és
ellenanyag-függő citotoxikus mechanizmussal (ADCC) pusztítják a fertőzött sejteket.
Az NK sejtek átmenetet képeznek a veleszületett- és az adaptív immunválasz között,
mivel elősegítik a DC-k érését [37]. A DC professzionális antigénbemutató sejtek
(APC) szerepe kulcsfontosságú a HIV specifikus immunválasz kiváltásában.
(Ugyanakkor ezek a sejtek fertőződnek meg elsőként, amikor a HIV a nyálkahártyán át
bejut a szervezetbe.) Az éretlen DC-k endocitózissal veszik fel a virionokat a
szervezetbe jutásuk helyén, és a közeli nyirokcsomóba vándorolnak, miközben érett
DC-vé differenciálódnak az antigén feldolgozása során. A DC-k antigén bemutató
funkcióját az MHCI (fő hisztokompatibilitási komplex I) és MHCII (fő
hisztokompatibilitási komplex II) molekulákhoz kapcsolt immunogén peptidekkel, azaz
epitópokkal hajtja végre. A 9-11 aminosav (as) hosszúságú epitópot kötő MHCI
molekula CD8+ T sejtek érését (ún. priming-ot) segíti elő, míg az MHCII molekulához
kapcsolt 13-25 as méretű epitópok a CD4+ helper T sejtek érését stimulálják. A HIV
virionok endocitózisa után az exogén (sejten kívülről származó) vírusfehérjék
proteolitikus lízissel peptidekre esnek, és ezek egy része az endocitotikus úton MHCII
molekulákra kötődik, amit a DC-k CD4+ helper T sejteknek mutatnak be (5.ábra) [38].
A sejten belül képződő vírusfehérjék, és kereszt-prezentációval akár a sejten kívülről

15
felvett antigének is képesek az MHCI útvonalon CD8+ citotoxikus T sejtek (CTL)
primingjára [39,40]. (A HIV vírus Nef fehérjéje az antigén bemutatás hatékonyságát
csökkenti, mert az MHCI és MHCII molekulák downregulációjával akadályozza HIV
epitópok megjelenését a sejtfelszínen [41,42].) Az MHCI útvonalon a HIV antigének
proteasomákban történő részleges lebontás után peptidek formájában haladnak át a TAP
transzportfehérjéken (Transporter associated with Antigen Processing) az
endoplazmatikus retikulumba. A TAP molekulán azonban átlagosan csak minden 7.
peptid képes átjutni, és az epitópok száma még ennél is kisebb, mert csak minden 200.
peptid illik bele az MHCI molekula kötőzsebébe, hogy aztán megfelelő erősséggel
kötődjön [43,44].
5. ábra. Az antigén bemutatás útvonalai DC-kben. A HIV fehérjéket a DC-k mindkét útvonalon
képesek feldolgozni: a sejten kívülről származó antigénekből proteolitikus bontással nyert
peptideket MHCII molekulákon mutatják be, ezzel CD4+ T sejtek érését elősegítve. A sejtben
képződő vírusfehérjék, ill. kereszt-prezentációval az endocitózissal felvett antigénekből
származó peptidek pedig MHCI molekulákhoz kapcsolódva CD8+ T sejtek érését segítik elő.
Az MHCI-hez a TAP transzportfehérjén átjutott peptidek egy része tud kapcsolódni.
A DC-k tehát elősegítik a differenciálódó CD4+ helper- és CD8+ T sejtek HIV
specifikus effektor sejtté- ill. memória sejtté érését. Az antigén bemutatás során a
nyirokcsomóban a vírus specifikus MHC-peptid komplexhez kapcsolódni képes T sejt

16
receptorú (TCR) T sejtek aktiválódnak, és gyorsan proliferálódnak, aminek
köszönhetően több nagyságrenddel megnő a mennyiségük. A CD8+ T sejtek emellett
differenciálódnak is (effektor/memória sejtté). Az effektor sejtek antivirális citokineket
termelnek (pl. IFN-γ, β-kemokin, MIP-1α, MIP-1β és RANTES), ill. citolitikus sejtté
differenciálódnak granzim és perforin enzimek termelésével (ez a sejtpopuláció képes a
nyirokszerveket, ill. nyirokkeringést is elhagyni). A citolitikus sejtek azután közvetlen
lízissel képesek elpusztítani a specifitásuknak megfelelő fertőzött sejteket. Az effektor
sejtek kis része további differenciálódás után nyugvó memóriasejtekké alakul, amik
antigén hiányában is hosszú ideig képesek fennmaradni, és újabb antigén stimulusra
azonnali effektor funkciókkal válaszolni. A CD4+ helper T sejtek termelődése az
immunválasz során két szempontból meghatározó: a HIV fertőzés miatt pusztuló CD4+
T sejt állományt pótolják, és a CD8+ T sejtek működését segítik. Utóbbi segítő funkció
több szinten is megnyilvánul, pl. a CD8+ T sejtek megfelelő primingjában az akut
fertőzés során, és a memória CD8+ T sejt populáció kialakulásában. A CD4+ T sejtek
közreműködésével kialakult CD8+ memória T sejtek pedig hatékonyabb másodlagos
effektor populációt képesek termelni [45,46,47]. Noha a vírusfertőzések során az
ellenanyag termelés kulcsfontosságú a fertőzés megakadályozására, HIV esetében ez
nem igaz. A neutralizáló ellenanyagok definíció szerint a kórokozó felszíni fehérjéihez
specifikusan kapcsolódva megakadályozzák, hogy a fertőzés létrejöjjön. A neutralizáló
(és a nem-neutralizáló) HIV specifikus ellenanyagok azonban nem tudnak megfelelő
hatást kifejteni a fertőzés megelőzésére, de még a vírus replikáció fékezésére sem a
krónikus fertőzési szakaszban. Ennek oka, hogy a vírus gyors mutációs rátájának és az
immunszelekciónak köszönhetően hamar megjelennek olyan mutáns vírustörzsek, amik
az ellenanyag kötőhelyeken változva kivédik az ellenanyag kötődését. [48,49].
A fent részletezett specifikus (adaptív) immunválasz megfékezi és visszaszorítja a HIV
további szaporodását: a HIV-specifikus T limfociták hatékonyan csökkentik a szervezet
vírus-terheltségét. A humorális immunválasz eredményeként pedig jelentős ellenanyag
termelés mutatható ki, vagyis megtörténik a szerokonverzió is. A fertőzés akut
szakaszában a legtöbb HIV virion éppen a frissen fertőzött proliferálódó CD4+ T
sejtekben termelődik. Noha az immunrendszer hatékonyan pusztítja ezeket a fertőzött T
sejteket, egy kis részük elég hosszú ideig életben marad ahhoz, hogy visszaalakuljon
nyugvó memória sejtté, és az integrálódott HIV-et (provírus) hordozva a vírus
rezervoárját képezze.

17
A lecsökkent vírusszám, és replikációs aktivitás mellett az immunrendszer hosszú
évekig képes kontrollálni a fertőzést (akár 15 évig), ez a klinikailag tünetmentes látens
fertőzési szakasz. Ezalatt a CD4+ T sejt szám és a DC-k száma is lassan csökken, sőt
egyes nagyon alacsony vírusszámú fertőzöttekben sosem éri el az AIDS tünetegyüttes
kialakulásához vezető mennyiséget. A csökkenés oka elsősorban az, hogy a vírus a
CD4+ T sejteket fertőzi, továbbá a vírusra ható immunszelekció (escape mutánsok
megjelenése), és az aktivált T sejtek apoptózisra való “hajlama” is közrejátszik a
folyamatban.
Amikor a CD4+ sejtek száma a kritikus mennyiséget közelíti (200/ml), a
vírusmennyiség drasztikusan nőni kezd, és járulékos fertőzések tünetei jelennek meg a
betegben, pl. TBC, toxoplazma, gombás fertőzések, és tumoros betegségek. Az AIDS, a
szerzett immunhiányos tünetegyüttes a HIV fertőzés utolsó fázisa, ami néhány éven
belül halállal végződik.
Fontos megjegyezni, hogy az immunválasz minőségét, ezáltal a betegség lefolyásának
sebességét nagyban befolyásolhatja a fertőzött egyén MHC genotípusa. Egyes MHC
allélek (más néven HLA-humán leukocita antigén) nagyon hatékony immunválasszal
egészen alacsonyra csökkentik az akut fázisban a vírusszámot, ezzel lassú betegség
lefolyást tesznek lehetővé. Ezek a protektív allélek, pl. HLA-B57 [50]. Sajnos azonban
léteznek nem-protektív allélek is, pl. HLA-B35, és az ilyen allélekkel rendelkező
betegekben az átlagosnál gyorsabb a betegség lefolyása. HLA [51].
3.3 HIV diverzitás és epidemiológia
A HIV-1 vírus őse valószínűleg a SIV majom immundeficiencia vírus volt, amit
Kamerunban csimpánz vadászok kaphattak el elsőként [52,53,54]. A majom vírusok
filogenetikai vizsgálatai alapján úgy tűnik több független fertőzési esemény is történt,
az első valamikor az 1930-as évek környékén [55]. Ezekből jöhetett létre a HIV-1
három járványcsoportja: a mára globális járványokozó M (main) csoport, az O (outlier)
csoport és az N (non-outlier) csoport [3]. Egy átfogó filogenetikai tanulmány szerint
viszont az O csoport tagjaihoz filogenetikailag legközelebb álló majom vírustörzsek
gorillából származnak [56]. Az N és O csoport tagjai csak az eredetük helyén, Nyugat-
Afrikában terjedtek el, míg az M csoport alább részletezett szubtípusai és rekombináns
csoportjai tehetők felelőssé a globális járványokért (6. ábra. HIV szubtípusok és
rekombináns formák földrajzi elterjedése.) [54].

18
A HIV-2 kórokozó őse szintén majom SIV, a Cercocebus atys fajt fertőző vírus, de a
HIV-2 járványtani jelentősége a HIV-1 N és O csoportjához hasonlóan kis területekre
korlátozódik [57]. Ezért a továbbiakban a HIV rövidítés alatt jellemzően a HIV-1 M
csoportját értem.
Rövid múltja ellenére a HIV nagy természetes diverzitással rendelkezik, és számos,
térbeli és időbeli elkülönülést mutató kisebb genetikai csoportba, szubtípusba sorolható.
Gyors evolúciójának legfőbb oka, hogy a HIV reverz transzkriptáz enzimének nincs
proofreading hibajavító aktivitása, így a magas, kb. 3,4x10-5
mutáció/bázispár/replikációs ciklus mutációs ráta eredményeként milliós nagyságrendű
új vírusvariáns képződik naponta(!) minden fertőzött személyben [58]. Ilyen ütemű
evolúció mellett mára a szubtípusok genetikai diverzitása 10-35% [3]. Jelenleg 9
szubtípusba sorolhatók az M csoportba tartozó izolátumok, melyeket A-K betűkkel
jelölnek, ill. vannak még kisebb genetikailag elkülönülő csoportok: szub-szubtípusok,
valamint rekombináns alcsoportok, amik kialakulásához különböző szubtípusokkal való
egyidejű fertőzés, rekombináció, és a rekombináns sikeres terjedése szükséges (6. ábra)
[54]. A HIV-1 nevezéktani ajánlás definíciója szerint ilyen rekombinánsok (circulating
recombinant form, CRF) akkor jegyezhetők be, ha legalább három személyből,
járványtani kapcsolatot nem mutató esetekben izolálják a stabilizálódott rekombináns
vírust [59].
6. ábra. HIV szubtípusok és rekombináns formák földrajzi elterjedése.
B és CRF01_AE
CRF02_AG és más rekombináns formák
A, B és AB rekombináns forma
B és BF rekombináns forma
B, C és BC rekombináns forma
F, G, H, J, K, CRF_01 és más rekombináns formák
Nincs elegendő adat
A
B
C
D

19
3.4 A DermaVir kifejlesztése
A DermaVir nanomedicina kifejlesztésének ötletét egy HIV-fertőzött beteg különleges
kórtörténete adta 1999-ben [14,60]. A betegnél korai diagnózist követően kezdték meg
az antiretrovirális terápiát, amit a szokásos módon szakítottak meg ún. kezelési
szünetekkel (a kezelési szünetekben az immunrendszer ugyanis új erőre kap). Két ilyen
kezelési szünet után az ilyenkor szintén szokásos mérsékelt vírusszint emelkedés volt
megfigyelhető. Aztán a beteg egy időre abbahagyta a terápiát, és a következő klinikai
ellenőrzés során kiderült, hogy vírusszintje nem emelkedett. Vagyis a beteg szervezete
olyan hatékony immunválaszt produkált, ami kordában tudta tartani a fertőzést. A
vírusszint ezután még évekig alacsony maradt antiretrovirális kezelés nélkül, és csak
azután emelkedett újból mérhető szintre! Ez volt az első klinikai megfigyelés, ami azt
mutatta, hogy létezhet hatékony immunválasz a HIV ellen. Az eset rámutatott, hogy a
fertőzés korai szakaszában a T sejtes immunválasz megfelelően erős ahhoz, hogy a
vírusszámot a mérési határ alá szorítsa. A megfigyelést egy majomkísérlettel is sikerült
igazolni: primer fertőzést követően beiktatott kezelési szünet után a T sejtes
immunválasz sikeresen megfékezte, és visszaszorította a SIV vírus szaporodását [61]. A
fertőzés hosszú távú természetes kontrollját azóta sokan leírták az “elit kontroller” HIV
fertőzöttek esetében, akiknél a szabad vírusszám tartósan nagyon alacsony (50 kópia/ml
alatti) [62].
A termékfejlesztés irányát az ekkor megfogalmazott kérdés határozta meg: hogyan
lehetne olyan HIV-specifikus T sejtes immunválaszt generálni, ami hatékonyan
pusztítja a HIV-vel fertőzött sejteket, és ezzel a tünetmentes fázisban megfékezi a
betegség előrehaladását? Mivel a T sejtek kórokozó-specifikussá érését
antigénbemutató sejtek irányítják, ezért HIV antigének DC-kbe juttatásával lehetne
megoldani a feladatot. A kifejlesztett DermaVir termék lényegében a természetes
immunválasz biztonságos utánzása, az alábbi módon. A hatóanyag pDNS, ami a
biztonsági módosítások mellett a lehető legtöbb HIV antigént kódolja, hogy minél
hatékonyabb (széles specifitású) T sejt választ tudjon kiváltani. A pDNS DC-k általi
felvételét és az antigén expressziót egy szintetikus patogén-szerű nanorészecske
formuláció segíti [63]. A nanorészecskék DC-khez irányítása pedig egy szintén saját
fejlesztésű technológiával történik: a DermaPrep kezelési eljárással a nanorészecske
oldat a bőrben lévő DC populációhoz, az LC-khez juttatható. A DermaVir kezelés
hatásmechanizmusát a 7. ábrán mutatom be röviden.

20
7. ábra. A DermaVir kezelés hatásmechanizmusának áttekintése. A bőr előkészítése a során a
nanomedicina penetrációját akadályozó felső elhalt sejtrétegeket (stratum corneum) kell
eltávolítani dörzsöléssel. Ugyanez a lépés aktiválja, és a kezelt területre vonzza az LC-ket
enyhe bőrpír kíséretében. Ezután zsebet formáló, szélein leragasztható 80 cm2-es tapasz(oka)t
helyezünk a bőrre, és az így képződő “tartályba” juttatjuk a nanorészecskéket tartalmazó
oldatot. Az odasereglő aktivált LC-k felveszik a mannozilált felszínű, patogént imitáló
nanorészecskéket és a nyirokcsomóba vándorolnak, ahol az időközben a pDNS-ről termelődő
fehérje antigéneket feldolgozzák. A feldolgozott antigénekből származó epitópokat ezután
éretlen T sejteknek mutatják be (priming), amit az immár HIV specifikus CD4+ és CD8+ T
sejtek proliferációja-, és a szervezetben való szétterjedése és követ. A kezelés hatására
termelődött T sejtek (effektorok és nagy proliferációs kapacitású memória sejtek) a HIV-vel
fertőzött sejteket pusztítják.
A DermaVir kezelés fő lépései megegyeznek a kórokozók elleni T sejtes immunválasz
természetes lefolyásával: a kezelést végző személy a bőr felső elszarusodott rétegét
dörzsöléssel távolítja el. Ezzel megszünteti a fizikai akadályt, ami a nanomedicina vizes
oldatának felszívódását akadályozná, valamint ez a “veszély-jelzés” odavonzza és
aktiválja az epidermisz antigénbemutató sejtjeit. A felületre ezután egy 80 cm2-es,
szélein leragasztható tapasz kerül, ami a bőrfelületen tartja az ezután egy tű nélküli
fecskendővel a tapasz alá juttatott nanomedicinát. A felszívódás során az LC-k
találkoznak a mannozilált felszínű szintetikus nanorészecskékkel, és endocitózissal
felveszik azokat, mintha kórokozók volnának. A nanorészecske “belsejében” lévő
NYIROKCSOMÓ

21
pDNS-t a szintetikus burok megvédi az enzimatikus bontástól, így a pDNS-ről
expresszálódhat a kódolt 15 antigén, majd megtörténhet az antigén bemutatás. (Az
antigén bemutatás lehetséges útvonalait a disszertáció új eredményei alapján az
“Eredmények megvitatása” c. fejezetben fejtem ki.) Eközben az LC-k a közeli
nyirokcsomókba vándorolnak, ahol CD4+ T sejtek-, és CD8+ T sejtek érését
stimulálják a pDNS-ről származó HIV-specifikus epitópok bemutatásával.
Végeredményül HIV-specifikus effektor sejtek és memória T sejtek termelődnek, amik
képesek a HIV-vel fertőzött sejtek elpusztítására [64].
3.5 A DermaVir pDNS alapú antigénje
A DermaVir hatóanyaga, a pLWXu1 pDNS a széles specifitású T sejtes immunválasz
kiváltása érdekében úgy lett tervezve, hogy a tizenöt HIV fehérje közül tizenhármat
módosítás nélkül expresszáljon, valamint az IN és Nef fehérjék működésképtelen N-
terminális részét is kódolja. Több biztonsági módosítás gátolja meg az integrációt,
reverz transzkripciót és replikációt. A tizenhárom fehérje módosítás nélküli kódolása
elméletileg azt is lehetővé teszi, hogy belőlük komplex vírusszerű részecskék (VLP+)
képződjenek. Az eredeti HIV promóter (5’ LTR) biztosítja a pDNS által kódolt fehérjék
expresszióját antigénbemutató sejtekben (lásd 3. ábra) [25]. A Nef fehérje működésének
kiiktatása az immunogenitás növelésének előnyével is jár, mert a Nef az MHCI és
MHCII molekulák downregulációjával akadályozza HIV epitópok megjelenését a
sejtfelszínen a természetes fertőzés során, amivel meggátolja, hogy az immunrendszer
megtalálja a fertőzött sejteket [41,42,65].
3.6 Az antigén DC-be juttatása patogén-szerű szintetikus nanorészecskével
Annak érdekében, hogy a pDNS hatékonyan jusson be a DC-kbe, és ott a kódolt
antigéneket expresszálja, egy szintetikus nanorészecske formulációt fejlesztettünk ki. A
polietilénimin-mannóz (PEIm) a pDNS-sel a vírusok mérettartományába eső 70-300 nm
átmérőjű gömbszerű nanorészecskéket alkot, amit a DC-k a kórokozókhoz hasonlóan
felvesznek és aktiválódnak [63,66,67]. Ebben belül helyezkedik el a pDNS, amit
beburkol a mannozilált, pozitív töltésű polimer (8. ábra). A nanorészecske endocitózisa
után a pDNS-t a polimer burok megvédi az enzimatikus bontástól, és lehetővé teszi,
hogy a pDNS eljusson a sejtmagba és megtörténjen az antigének expressziója. A pDNS
védelmének “finomhangolását” a nanorészecskét stabilizáló kötések száma határozza

22
meg a pDNS foszfátcsoport oxigénje és a PEIm másodlagos amin nitrogénje közt. A
nanorészecske stabilitása tehát szorosan összefügg a pDNS antigén-expressziós
hatékonyságával (biológiai aktivitásával), mert az endoszómában védenie kell a pDNS-
t, majd a sejtmag közelébe jutva “szétesnie” [66,68].
A szintetikus nanorészecskék használatának előnye tehát, hogy segíti a pDNS-be kódolt
antigének DC-kbe jutását, expresszióját, és azután az endogén antigének bemutatási
útvonalán haladhat az antigének feldolgozása. Továbbá a nanorészecskék terápiás
alkalmazásával kiküszöbölhetjük az általánosan használt vírusvektorokkal szemben a
többszöri alkalmazás miatt kialakuló rezisztenciát.
8. ábra. A DermaVir szintetikus nanorészecske egyszerűsített modellje (keresztmetszet).
pDNS: plazmid DNS, PEIm: polietilénimin-mannóz.
3.7 DermaPrep orvosi eszköz a nanomedicina bőrön át történő bejuttatására
Az antigéneket pDNS-ben kódoljuk, a nanorészecske formuláció segít a pDNS DC
célsejtekbe juttatásában és az antigének expressziójában ill. bemutatásában, végső soron
a HIV-specifikus T sejtek termelésében. De hogyan kerül a DC-k közelébe a
nanorészecske oldat? Ennek a lépésnek a megoldása a szintén saját fejlesztésű
DermaPrep orvosi eszközzel lehetséges. Az eszköz alkalmazása magában foglal egy bőr
előkészítő eljárást: egy durva szivaccsal eltávolításra kerül a bőr külső-, fizikai
védelmet biztosító stratum corneum rétege, és ez veszély-jelzésként odavonzza és
aktiválja az LC (DC) sejteket (7. ábra) [69,70]. A bőrre ragasztott féligáteresztő
anyagból készült tapasz alá juttatott nanomedicina immár könnyen felszívódik, és az
LC-k kórokozók után kutatva hatékonyan fel is veszik a patogén-szerű
nanorészecskéket. A DermaPrep kezelés lényegében a természetes immunválasz
folyamatait segíti elő, ugyanis az aktivált LC-k a veszély-jelzés hatására egy 900-1800
sejt/mm2 hálózatot hoznak létre közvetlenül a bőrfelszín alatt [71]. Vagyis körülbelül 8
pDNS
PEIm

23
millió LC-t állít szolgálatba a 80 cm2 felületen végzett kezelés. Miután pedig az LC-k
felvették a DermaVir nanorészecskéit, a nyirokcsomóba vándorolnak, hogy a pDNS-
ben kódolt antigénekre specifikus T sejtek termelését irányítsák. A DermaPrep eszköz
tehát végső soron a nanomedicinát a bőrön át a nyirokcsomóba juttatja. A DermaPrep
orvosi eszköz európai forgalmazási engedélyét (CE jelzés) 2009-ben szereztük meg.
3.8 Preklinikai és klinikai eredmények
A DermaVir hatásmechanizmusának kísérletes igazolása a humán klinikai kísérletek
előtt ex vivo kísérletekkel, majd előrehaladott állapotú SIV fertőzött rhesusmajmokban
történt a fent ismertetett pDNS konstrukció SIV (és HIV) fehérjéket kódoló változatával
[64,72]. Sikerült igazolni, hogy az ex vivo úton DC-kbe juttatott DermaVir-rel kezelt
sejtek szubkután injektálását követően a pDNS-ben kódolt antigének expresszálódnak a
nyirokcsomóban lévő DC-kben, és, hogy ezek a DC-k HIV-specifikus CD4+ Helper T
sejtek, valamint CD8+ citotoxikus T sejtek termelődését indukálják [67]. Ezután a
mérések in vivo ismétlése következett egér-, és majom modellben, a DermaVir-t
DermaPrep-pel bőrön át bejuttatva. Az in vivo eredmények megerősítették az ex vivo
mérések eredményét, ezzel alátámasztva a DermaPrep kezelési eljárás hatékonyságát
[64]. A következő lépés a humán fázis I kísérletek előtt a tervezett humán kísérlethez
hasonló méréssorozatok elvégzése volt krónikus SIV251 fertőzött rhesusmajmokban. Az
antiretrovirális kezeléssel kombinált (cART) DermaVir kezelések növelték a SIV-
specifikus T sejtek mennyiségét (még a késői stádiumú, AIDS-es majmokban is),
lecsökkentették a vírusszámot, és jelentősen megfékezték a vírus replikációt a kezelés
utáni időszakban is. Ennek eredményeképpen a kezelt állatok életideje 18 hétről 38
hétre növekedett a nem kezelt kontroll csoportéhoz képest [72].
Az első, embereken végzett klinikai kísérlet a budapesti Szent László Kórházban, 2005-
2006 között zajlott [73,74]. Célja a DermaVir kezelés biztonságosságának, és
immunogenitásának vizsgálata volt. Kilenc cART kezelés alatt álló HIV fertőzött
beteget kezeltek DermaVirrel, egy alkalommal, három különböző dózissal: három-
három beteg kapott 0,1 mg DNS-t tartalmazó DermaVir-t két DermaVir tapasszal
bejuttatva, 0,4 mg pDNS-t tartalmazó DermaVir-t négy tapasszal, ill. 0,8 mg pDNS-t
tartalmazó DermaVir-t nyolc tapasszal. Nemcsak a hatóanyag mennyisége különbözött
tehát, hanem a célzott nyirokcsomók (ill. LC-k) száma is. A kezelés után 28 napon át
figyelték az esetleges mellékhatások megjelenését, majd hosszú távon is ellenőrizték:

24
12, 24, 36 és 48 héttel a kezelést követően. A vizsgálat során egyetlen esetben sem
tapasztaltak súlyosabb mellékhatást, a leggyakoribb (négy betegnél) enyhe bőrreakció
volt, dózistól függetlenül. A hatásosság vizsgálata a következő eredményeket hozta: a
cART terápiának köszönhető 50 kópia/ml alatti vírusszint változatlanul alacsony maradt
a DermaVir adását követő 28 napban. A Gag, Tat és Rev antigénekre specifikus (IFNγ-
át és IL2-t termelő) CD4+ és CD8+ effektor T sejtek száma megemelkedett, az alacsony
dózisú csoportban 3-ból 2 kezeltben, és a középső dózisú csoport mindhárom tagjában.
Ezenkívül a magas proliferációs kapacitású HIV-specifikus memória T sejtek száma is
szignifikánsan emelkedett, és még egy évvel később is esetenként detektálható volt
[73,74,75]. Ezek az eredmények azt mutatták, hogy a DermaVir biztonságos, és a
középső dózisban már egyetlen DermaVir immunizálás is kiváltja az elvárt széles
specifitású T sejtes immunválaszt. De a tartós hatás érdekében szükségesnek tűnik a
DermaVir kezelés bizonyos időközönkénti ismétlése.
Az USÁ-ban végzett fázis I/II klinikai kísérletben ezért már többszöri DermaVir
kezelés hatását vizsgálták 24 cART kezelés alatt álló HIV fertőzött betegben [76]. A
betegek egyik csoportja 0,2 mg, ill. 0,4 mg pDNS dózisú DermaVir-t kapott három
alkalommal, a másik csoport 0,4 mg pDNS dózisú DermaVir-t hat alkalommal, a
harmadik csoport pedig placebo-t. A kezelések nem okoztak súlyos mellékhatásokat, és
ismét megemelkedett a HIV-specifikus effektor-, és memória T sejtek száma, a
legjobban a 0,4 mg-mos dózissal kezelt csoportban.
A következő, fázis II klinikai vizsgálat célja a DermaVir biztonságosságának mérése,
valamint az immunogenitásának és antivirális hatásának mérése volt, ezúttal cART
kezelést nem kapó betegekben [77]. 36 HIV-fertőzött egyént kezeltek a háromféle
DermaVir dózis valamelyikével (0,2 mg, 0,4 mg vagy 0,8 mg pDNS/DermaVir), ill.
placebo-val hat-hetenként, összesen négy alkalommal. A kezelt csoportokban
előforduló mellékhatások nem különböztek a placebo csoportétól, csak egy betegnél
fordult elő közepesen súlyos mellékhatás (a háromfokozatú skálán 2-es), de az ő
kezelését sem kellett emiatt felfüggeszteni. A 0,4 mg-mos dózisú csoport megint jobb
immunválaszt mutatott a többinél, és szintén ebben a csoportban a szabad
vírusmennyiség 70%-kal csökkent a placebóhoz képest.
Az eddigi klinikai vizsgálatok eredményei alapján tehát a DermaVir kezelés ismételt
alkalmazása megfelelően biztonságos, és HIV specifikus T sejtes immunválaszt
generál, valamint fékezi a HIV szaporodását.

25
A pLWXu1 pDNS-t tartalmazó DermaVir termékjelölt klinikai vizsgálatai tovább
folytatódnak, és reményeink szerint a termék forgalomba hozatali engedélyével zárul a
fejlesztés. Ezzel párhuzamosan a DermaVir hatásmechanizmusának további,
részletekbe menő jellemzésén is dolgozunk. A munka folyamatosan újabb kutatási
kérdéseket vet fel, amik megválaszolása új fejlesztési irányt is kijelölhet. Így történt ez
doktori munkám esetében is, aminek egy személyre szabható DermaVir terápia, és
hozzá egy szubtípus optimalizált termékcsalád első termékjelöltjeinek elkészítése lett az
eredménye. Lássuk, hogyan!

26
4. Célkitűzések
A DermaVir HIV terápiás nanomedicina in vitro kísérletes jellemzését, és az antigén
repertoár in silico jellemzését, valamint ezekhez vizsgálati módszerek fejlesztését
tűztem ki célul. A DermaVir immunválasz kiváltó képességének in silico modellezési
eredményei alapján ezután indokoltnak láttuk egy HIV szubtípus-optimalizált
DermaVir termékcsalád kifejlesztését, és a termékcsaládon alapuló személyre szabott
HIV immunterápia prototípusának kidolgozását.
Fenti célok eléréséhez az alábbi részfeladatokat állapítottuk meg:
I. A klinikai tesztelés alatt álló DermaVir HIV terápiás nanomedicina kísérletes
jellemzése a hatásmechanizmus egyes részleteinek megismerésére, ill.
alátámasztására. (Eredmények a 6.1. alfejezetben.)
II. Az előbbi célkitűzéshez in vitro vizsgálati módszerek fejlesztése, ill.
optimalizálása és validálása a DermaVir komplex VLP+ termelésének-
valamint biológiai aktivitásának (antigén expressziós potenciáljának)
mérésére. (Eredmények a 6.1.2. és 6.1.3. alfejezetekben.)
III. A DermaVir terápiás nanomedicina pDNS hatóanyaga által expresszált
majdnem teljes proteom immunválasz kiváltó képességének (immunológiai
potenciáljának) in silico modellezésére T sejt epitóp predikción alapuló
módszer kidolgozása. (Eredmények a 6.2.1. alfejezetben.)
IV. A szubtípus-B specifikus HIV szekvenciát tartalmazó DermaVir
immunológiai potenciáljának in silico modellezése, és a nanomedicina
hatásosságának in silico modellezése nem-B szubtípusú HIV-vel
fertőzöttekben. (Eredmények a 6.2.2.-6.2.5. alfejezetekben.)
V. HIV-1 szubtípusokra immunológiailag optimalizált pDNS-ek tervezése, és
ehhez a tervezést segítő eszköz elkészítése, ami a személyre szabható
immunterápia megvalósítását is támogatja. (Eredmények a 6.3.1. és 6.3.2.
alfejezetekben.)
VI. A megtervezett pDNS-ek elkészítése, valamint az elkészített termékjelöltek in
silico jellemzése. (Eredmények a 6.3.3. alfejezetben.)
VII. Az új termékjelöltek in vitro jellemzése. (Eredmények a 6.3.4. alfejezetben.)

27
5. Anyagok és módszerek
5.1. Nanomedicina készítés
1 mg/ml koncentrációjú pDNS-t 6 térfogat egység 10%-os glükóz oldattal higítottunk,
majd 13,6 mM koncentrációjú polietilénimin-mannóz (PEIm) oldatot adtunk az
elegyhez. A nanorészecske képződés 20 perces, szobahőn (23 ± 2ºC) történő inkubálás
alatt történt.
5.2. Western blot
DMEM tápfolyadékban tartott 293T humán sejteket transzfektáltunk 10 µg pDNS-t
tartalmazó DermaVirrel, nem transzfektált sejteket használva negatív kontrollként. A
sejteket 24 órás, 37 °C-on, 5% CO2 tartalom mellett történő inkubálás után lizáltuk, és
szétosztott adagokban felhasználásig -80 °C-on tároltuk. A lizált mintákat, vagy
tisztított VLP+-t (lásd az 5.5. pontban) előre öntött 4-20% gradiens gélben (Bio-Rad)
futtattuk, majd PVDF membránra (Bio-Rad) blottoltuk 1 órán át 300 mA-rel
(omniPAGE Electroblotting System, Cleaver Scientific). A membránt 5% BSA-val
blokkoltuk, és többszöri PBS mosást követően HIV+ humán szérumban (Boston
Biomedica) inkubáltuk 1 órán át. Tween-20-at tartalmazó PBS mosási lépések után a
membránt peroxidázzal konjugált anti-humán IgG-vel (Sigma-Aldrich) inkubáltuk,
majd mostuk, végül az eredményt kolorimetriás eljárással (Amplified Opti-4CN kit,
Bio-Rad) tettük láthatóvá.
5.3. Immunprecipitáció
DMEM tápfolyadékban tartott 293T sejteket transzfektáltunk pLWXu1-gyel, és 24 órán
át 37 °C-on, 5% CO2 tartalom mellett inkubáltuk, majd radioaktívan jelöltük 400
microCi 35S-sel (Perkin Elmer NEG009H) és egy éjszakán át inkubáltuk. Pozitív
kontrollként a pLW fertőzőképes vírust tartalmazó szülői pDNS-t, negatív kontrollként
nem-transzfektált sejteket használtunk. A következő napon a sejteket lizáltuk és
szétosztott adagokban felhasználásig -80 °C-on tároltuk. A lizált minták precipitációját
inaktivált HIV+ humán szérum (Advanced BioScience Laboratories), vagy
monoklonális Nef ellenanyag (NIH AIDS Research and Reference Reagent Program)
hozzáadásával végeztük. Mosást követően a mintákat 12% SDS-PAGE gélben futtattuk.

28
5.4. Intracelluláris festés
293T sejteket transzfektáltunk 10 µg pLWXu1-gyel vagy a pozitív kontroll pLW
pDNS-sel, és 48 órás inkubálás után monoklonális egér ellenanyagokkal (NIH AIDS
Research and Reference Reagent Program ill. Affinity BioReagents) jelöltük, majd
EPICS XL-MCL áramlási citofluoriméterrel vizsgáltuk (Coulter). Minden mérést 50000
adatponttal végeztünk.
5.5. VLP+ termelés és tisztítás
293T sejteket transzfektáltunk DermaVirrel, majd 24 órával a transzfekció után
leszívtuk a VLP+-t tartalmazó felülúszót. A felülúszót 10 ml 20%-os szukróz oldatra
pipettáztuk és a VLP+-ket centrifugálással szűrtük át a szukróz oldaton (50000 g, 3 órás
centrifugálás). A pelletet 1/20 térfogatarányú PBS (pH 7,4) oldatban szuszpendáltuk, és
a tisztított VLP+-ket felhasználásig -80 °C-on tároltuk.
5.6. Transzmissziós elektronmikroszkópia
293T sejteket transzfektáltunk DermaVirrel, és 24 órával a transzfekció után a leszívtuk
a felülúszót, a sejteket PBS-sel mostuk, majd 5% glutaraldehidet tartalmazó, 0.1 M
cacodylate pufferben fixáltuk. A fixált sejteket Dr. Kovács Attila Lajos segítségével az
ELTE Anatómiai, Sejt- és Fejlődésbiológiai Tanszékén beágyaztuk, metszettük, és
elektronmikroszkóppal (JEM100CX II) vizsgáltuk, az alábbiak szerint: a rögzített
sejteket 0.1 M cacodylate pufferrel mostuk, majd 50 °C-os 1,5%-os agarra rétegeztük.
A mintákat cacodylate OsO4 oldattal utó-fixáltuk, “en-block” festettük nem pufferelt
1% uranil-acetát oldattal, végül etanollal és propilén-oxiddal dehidratáltuk. A
beágyazáshoz DPM-30-at tartalmazó TAAB kitet használtunk (TAAB Laboratories
Equipment). Az ultravékony metszeteket lead-citráttal festettük.
5.7. Biológiai aktivitás mérés
DMEM tápfolyadékban tartott 293T humán sejteket transzfektáltunk DermaVirrel. 24
órás, 37 °C-on, 5% CO2 tartalom mellett történő inkubálás után a felülúszót leszívtuk,
és HIV p24 antigén ELISÁ-val (Zeptometrix, Retrotek), a gyártó előírása szerint
meghatároztuk a plazmid DNS által expresszált p24 koncentrációt. A biológiai
aktivitást mérő módszerünket az európai gyógyszerengedélyezési hatóság (International

29
Conference on Harmonisation of Technical Requirements for Registration of
Pharmaceuticals for Human use) ICHQ2(R1) ajánlása alapján végeztük [78]. A
validálás célja az volt, hogy az ajánlásban javasolt paraméterek meghatározásával
igazoljuk, módszerünk alkalmas a DermaVir biológiai aktivitásának (azaz antigén
expressziós képességének) mennyiségi meghatározására. A validált biológiai aktivitást
mérő módszer használatát az európai és amerikai gyógyszerengedélyezési hatóságok
(EMA, FDA) írják elő. Minden esetben öt párhuzamossal végeztük a méréseket
adatpontonként, valamint legalább két alkalommal ismételtük a mérést.
5.8. Statisztikai analízis
Az eredmények szignifikanciájának meghatározása Student’s t-teszttel történt, és
minden esetben megadtuk a P-értéket.
5.9. In silico T sejt epitóp repertoár analízis
Az MHCI és MHCII T sejt epitóp predikciókat az “Immune Epitope Database” (IEDB)
epitóp adatbázis felhasználásával végeztük [79]. Az MHCI epitóp predikciókhoz
mesterséges neurális hálózaton (Artificial Neural Network, ANN) alapuló predikciós
módszert-, míg MHCII epitóp predikciókhoz az úgynevezett Konszenzus módszert
használtuk [80,81,82]. Csak a nagy affinitású MHCI epitópokat vizsgáltuk, melyek 50
nM-nál kisebb koncentrációjú fél-maximális gátlási koncentrációval jellemezhetők [83].
Az MHCI epitóp predikciós paraméterek az alábbiak voltak: humán MHCI forrás faj;
összes, ill. kiválasztott MHCI allélek; peptid hossz: 9 aminosav; eredményhalmaz
szűkítése: csak az IC50=50 nM alatti epitópok; ANN predikciós módszer. Az MHCII
epitóp predikciókat a következő paraméterekkel végeztük: konszenzus predikciós
módszer; összes, ill. kiválasztott MHCII allélek, eredményhalmaz szűkítése: 90 vagy 50
konszenzus százalékos rangsor (Consensus Percentile Rank) ill. magasabb értékkel
rendelkező epitópok.
5.10. Filogenetikai analízis
A Los Alamos HIV Database-ből származó szekvenciák Clustal-W illesztését követően
(BioEdit 7.0.9.0) Neighbor-joining módszerrel Kimura két paraméteres korrekciós
modelljét használva filogenetikai fákat generáltunk (MEGA 4.1) [84,85]. A fák
topológiájának megbízhatóságát 1000 Bootstrap ismétléssel ellenőriztük [86].

30
5.11. pDNS konstrukciók készítése
5.11.1. Szekvenciák
A következő, szubtípus-optimalizált pDNS konstrukciókat készítettük el: pLWXu-AB,
pLWXu-BC, pLWXu-C, pLWXu-BF. A felhasznált szekvenciák kétféle forrásból
származtak: vírus RNS izolálást követő RT-PCR-rel HIV+ vérplazma mintákból
amplifikáltuk (pLWXu-BC, pLWXu-C, pLWXu-BF pDNS-ek esetében), ill. a “Los
Alamos HIV Database”-ben található HIV szekvenciák alapján tervezett gént
szintetizáltattunk (CRF AB gag gén, génbanki szám: AF414006).
5.11.2. RNS kivonás
Az RNS kivonást “QIAamp viral RNA Mini Kit”-tel (Qiagen) végeztük mintánként 140
µl vérplazmából, a gyártó előírása szerint.
5.11.3. Oligonukleotidok tervezése és szintézise
Az oligonukleotidokat génbanki, szubtípus-azonos HIV izolátumok megfelelő
génszakaszainak szekvenciái alapján terveztük OLIGO 4.0 szoftver segítségével. Az
oligonukleotidokat az Invitrogen (RT-PCR primerek) ill. az Eurofins MWG Operon
(szekvenáló primerek) állította elő.
5.11.4. RT-PCR
A szubtípus optimalizáláshoz két HIV genomi szakaszt amplifikáltunk, melyek a (1.)
gag és részleges pol ill. (2.) env-tat-rev-vpu géneket fedik le. A felhasznált primereket
az 1. táblázatban foglaltuk össze. Az egy lépéses RT-PCR reakciókat a SuperScript III
One-step RT-PCR System kittel (Invitrogen) végeztük, 50 µl reakcióelegyben, ami 25
µl 2x “reakció mix” puffert, 10 µl tisztított HIV RNS-t, mindkét oligonukleotidból 10
µM-t, 1U SuperScript III/Taq enzimet, 40U RNase-out enzimet, és nukleáz-mentes
vizet tartalmazott. A PCR reakció paraméterei az alábbiak voltak: 60 °C 30 min, 30x(94
°C 15 s, 55-58 °C 30 s, 68 °C 3 min), 68 °C 5 min.

31
1. táblázat. A HIV RT-PCR reakciókhoz felhasznált primerek.
5.11.5. Agaróz gélelektroforézis
0,8%-os, 0,5 µg/ml etídium-bromiot tartalmazó agaróz gélt és 1xTAE puffert
(Invitrogen) használtunk. Futtatási paraméterek: 100V, 45 perc ill. PCR termék
tisztítása esetén 60V, 2 óra. A gélek értékeléséhez az ImageJ (NIH) szoftvert
használtuk.
5.11.6. DNS tisztítás gélből
A kívánt 2,3 kbp (gag gén) ill. 3 kbp (env-tat-rev-vpu gének) méretű RT-PCR
terméknek megfelelő sávot, ill. az emésztett pDNS vektort (10, ill. 9,3 kbp) 0,8%
agaróz gélben való futtatás után kivágtuk, majd “Genelute Gel Extraction Kit”-tel
(Omega) tisztítottuk, a gyártó előírása szerint. A termékeket precipitáltuk 2,5x térfogat
abszolút etanol és 1/10 térfogat 3M nátrium-acetát hozzáadásával. 1 órás -80 °C-os
inkubálás után 70%-os jéghideg etanollal mostuk, majd szárítás után a pelletet 20 µl
DNáz-mentes vízben visszaoldottuk.
5.11.7. Inzert és pDNS vektor előkészítése irányított ligáláshoz
A tisztított RT-PCR termékeket (inzert) és a pLWXu1 plazmidot (vektor) restrikciós
enzimes emésztéssel készítettük elő a ligáláshoz. A Gag poliprotein klónozásához a
pLWXu1 pDNS-t és a megfelelő gag gént tartalmazó inzertet NarI és BstZ17I
restrikciós enzimekkel (New England BioLabs) emésztettük a gyártó által biztosított 1x
NEBuffer4 pufferben, egész éjszakán át 37 °C-on inkubálva. Az env-tat-rev-vpu gének
klónozásához a pLWXu1 pDNS-t és a megfelelő inzertet NcoI és BlpI restrikciós
enzimekkel (New England BioLabs) emésztettük a gyártó által biztosított NEBuffer4
pufferben egész éjszakán át 37 °C-on inkubálva. Az emésztett pLWXu1 pDNS vektort
az előző bekezdésben leírt módon gélből kivágtuk és tisztítottuk. Az emésztett inzertet
is precipitáltuk a fentiek szerint, és 20 µl DNáz-mentes vízben visszaoldottuk.
Primer azonosító Szekvencia Termék
cDNA1 AGA GAC CCA GTA CAA GC cDNS
Gag-F CTG GTA ACT AGA GAT CCC TCA GA1. PCR termék
Gag-R TTG TTT ATA CTA GGT ATG GTG
Env-F TGG CTC CAT GGC TTA GGA CAA TAT ATC TAT G2. PCR termék
Env-R CTA CTC CCT CTG CTG CTG GCT CAG CT

32
5.11.8. Hőkompetens E. coli kompetens sejt készítés
E.coli Max efficiency Stbl2 kompetens sejt (Invitrogen) felhasználásával saját
kompetenssejt- bankot (WCB) készítettünk az E. coli rázott lombikos fermentálását
követő kétszeri CaCl2-os kezeléssel 0 °C-on (víz-jég elegyben). A kompetens sejtet
felhasználásig (legfeljebb egy hétig) 0 °C-on tároltuk.
5.11.9. Ligálás és transzformálás
A pDNS vektort és az inzertet 1:5 mólarányban adtuk a ligálási reakció elegyhez, 4 U
T4 DNS ligáz (Invitrogen) és 5x ligáz puffer mellett 20 µl-es végtérfogatban, majd
egész éjszakán át szobahőn inkubáltuk. 50 µl E. coli kompetens sejthez 5 µl-t ligálási
elegyet adtunk, majd 30 perces jégen inkubálás, 45mp 42 °C-os vízfürdőben történő
hősokkolás, és újabb 1 perces jégen történő inkubálás után 250 µl LB tápfolyadékot
(Gibco) mértünk a sejtekhez. 90 percen át inkubáltuk 37 °C-on, 250 fordulat/min
keverés mellett, ezután LB-agar-Kanamicin (50 µg/ml) táptalajra szélesztettük, és 30
°C-on egy éjszakán át inkubáltuk. A transzformáns baktérium telepeket 4 °C-on
tároltuk.
5.11.10. pDNS termelés
A baktérium telepeket LB-Kanamicin (50 µg/ml) tápfolyadékba oltottuk, és 16 órán át
30 °C-on, 250 fordulat/min keverés mellett szelektíven növesztettük, majd izoláltuk az
így felszaporított pDNS-t. A pDNS izolálást E.Z.N.A. Miniprep Kittel (BioBasics)
végeztük a gyártó előírása (Plasmid Miniprep Protocol II) szerint, 1.5 ml baktérium
szuszpenzió feldolgozásával, átlagosan 50 µg hozammal. A nagyobb mennyiségű pDNS
termelést Qiagen Plasmid Maxi Kittel (QIAGEN) végeztük, a gyártó előírása szerint,
500 ml baktérium szuszpenzió feldolgozásával és átlagosan 500 µg hozammal. A pDNS
oldat koncentrációját az oldat 260 nm-en mért abszorbanciájának spektrofotometriás
meghatározása után a DNS koncentráció= A260× 50×(hígítási faktor) képlettel
számoltuk ki, és a pDNS oldat koncentrációját 1 mg/ml-re hígítottuk.
5.11.11. Az új pDNS-esek azonosságának ellenőrzése
A tisztított pDNS-eket a ligáláshoz használt enzimpárral (NarI-BstZ17I ill. NcoI-BlpI)
emésztettük. A reakcióelegy 1 µg pDNS-t, 1x NEBuffer4 puffert, 1-1 U restrikciós
enzimet és nukleáz-mentes vizet tartalmazott 20 µl végtérfogatban. 37 ºC-on 2 órás
inkubáció után a mintákat 0,8%-os agaróz gélben futtattuk. A megfelelő méretű DNS
fragmentumokat tartalmazó mintákat szekvenáltattuk (Eurofins MWG Operon).

33
6. Eredmények
A DermaVir hatóanyagát egy fertőzőképes, B szubtípusú HIV izolátumból kiindulva
fejlesztette ki korábban munkacsoportunk. [87,88,89]. A laboratóriumi dolgozóból
izolált vírus klónozása után (pLW) többszörös mutációkkal módosították a szekvenciát,
hogy a replikációt, ezen belül a reverz transzkripciót és az integrációt gátolják (9. ábra).
A reverz transzkripció a 3’LTR deléciójával blokkolható, és ezzel megakadályozható a
módosított DNS vad vírusból származó DNS-sel vagy retrotranszpozonokkal történő
rekombinációja ill. komplementációja is. Továbbá a HIV IN fehérjéje a 3’LTR régióhoz
kötődve irányítja a virális DNS integrációját. A deléció tehát egyszerre több folyamatot
is akadályoz. Az IN fehérje génjének többszörös mutációja (stop kodonok és 7 bp
deléció) további biztosíték az integráció gátlására [90]. Ezek a biztonsági módosítások
két fehérjét tesznek működésképtelenné: Pol-RT és Nef. Mindemellett a többi fehérje
módosítás nélküli kódolása lehetővé teszi, hogy a tizenöt HIV fehérje közül 13
természetes konformációban termelődjön, valamint ezek a fehérjék komplex vírusszerű
részecskékké (VLP+) álljanak össze. A Nef mutációját a fehérje kódoló régiójával
átfedésben lévő 3’ LTR deléciója okozza. De a Nef fehérje működésének kiiktatása az
immunogenitás növelésének előnyével is járhat, mert a Nef az MHCI és MHCI
downregulációjával akadályozza HIV epitópok megjelenését a sejtfelszínen a
természetes fertőzés során, amivel meggátolja, hogy az immunrendszer megtalálja a
fertőzött sejteket [41,42]. Az eredeti HIV promóter (5’ LTR) biztosítja a pDNS által
kódolt fehérjék expresszióját antigénbemutató sejtekben (lásd 3. ábra) [24,25]. A HIV
promóter használata új megközelítés a vakcinatervezésben, pedig a Tat jelenlétében
erősebb transzkripciós aktivitása van, mint az adenovírus promóternek, vagy a CMV
promóternek [25]. Ezenkívül talán az is előnyös lehet az immunválasz kiváltása
szempontjából, hogy az eredeti promóterről a természetes vírusfehérje termeléshez
hasonló mennyiségben, ill. sorrendben történik az átírás [91].

34
9. ábra. A DermaVir nanomedicina aktív hatóanyagának, a pLWXu1 pDNS-nek a (linearizált)
térképe. Biztonsági módosítások a pDNS HIV-specifikus részében: az integráz géntermék
inaktiválására két stop kodon, valamint egy 7 bp hosszúságú deléció, mely további alternatív
stop kodonokat eredményez a deléciótól downstream irányban az int génben. Hosszú deléció a
nef gén 3’ végén, mely csonka Nef átírását eredményezi. 3’ LTR teljes deléció, ami
megakadályozza a reverz transzkripciót. ■: biztonsági módosítások helye a pDNS-ben.
A pLWXu1 pDNS-ben kódolt fehérje repertoár jelenti a nanomedicina antigén
készletét. A fehérjetermelés kísérletes jellemzése ezért a nanomedicina hatásának
igazolása szempontjából, valamint biztonságossága szempontjából is nagyon fontos. Az
alábbiakban az erre irányuló in vitro kísérletek eredményét mutatom be. Azután pedig
az antigén-készlet immunválasz-kiváltó képességének (immunológiai potenciáljának) in
silico modellezése következik.
6.1. A DermaVir in vitro jellemzése
6.1.1. A pLWXu1 pDNS által expresszált fehérje repertoár
A pLWXu1 által expresszált fehérje (antigén) repertoár termelését Western blottal
igazolta munkacsoportunk (10. ábra). A legnagyobb mennyiségben a gag gén termékei
(p17-MA, p24-CA, p55-Gag poliprotein, amely tartalmazza a p7-NC fehérjét is), Env-
SU (gp120) és az Env-TM (gp41) expresszálódtak. A Pol fehérjéket: p66-RT plusz
76007541
CAGGAA TTT GGA ATT CCC TAG CGG TAA CCA-------GAGTAGTAGAATCTATGAATAA-AGAATTAAQ E F G I P stopstop stop stop stop stop stopdeletion
Inaktivált integráz gén Csonka nef génDel 12141-12309
Nef
11689 12309
12141
Inaktivált reverz transzkripcióDel 12141-12611
NF-κBNF-ATc
11978 12611
R U5
TATASP1
12141
Gag5’ LTR
NefEnv
TatRev
Prot RT RNáz IN
Pol
3’ LTR12252
1

35
RNáz, ill. RT (p51) kisebb koncentrációban detektáltuk. A teljes hosszúságú IN és Nef
fehérjék hiányát egyrészt szekvenálással, másrészt immunprecipitációval igazoltuk,
amihez a pLW fertőzőképes HIV-et expresszáló pDNS-t használtuk pozitív kontrollként
(ez a pDNS volt a pLWXu1 elkészítésének kiindulási alapja is; 10.B és C ábra) [92].
10. ábra. A pLWXu pDNS által expresszált fehérjék. A: western blot, HIV+ humán szérum
használatával elsődleges ellenanyagként. M: fehérje marker, Neg: negatív kontroll, nem
transzfektált sejtek; a-gp120 (Env-SU), b-p66 (RT plusz RNase), c-p55 (Gag poliprotein, benne
p17, p24 és p7), d-p51 (Pol-RT), e-gp41 (Env-TM), f-p24 (Gag-CA), g-p17 (Gag-MA). B: A
teljes hosszúságú Integráz termelődésének hiányát igazoló immunprecipitáció (csillaggal
jelölve). Az extra csíkok a HIV+ szérum használata miatt megjelenő többi HIV specifikus
fehérjének felelnek meg. C: A teljes hosszúságú Nef termelődésének hiányát igazoló
immunprecipitáció (csillaggal jelölve). D: pLWXu1-gyel transzfektált 293T sejtek
intracelluláris festése FITC-cel jelölt p24, p55, gp41, gp120, Rev és Tat ellenanyagok
használatával, áramlási citofluorimetriával mérve.
Mivel a többi szabályozó fehérjét sem western blottal, sem immunprecipitációval nem
sikerült kimutatni, egy társ-munkacsoport intracelluláris festéssel igazolta azok
termelődését. Ezzel sikerült bizonyítani a Rev (0,7% pLWXu1 esetében, ill. 1,3% a
kontroll pLW esetében) ill. Tat (0,2% pLWXu1 esetében, ill. 0,5% a kontroll pLW
esetében) fehérjék expresszióját (16.D ábra). Továbbá a p24, p55, gp41 és gp120

36
struktúrfehérjék termelődését a pLWXu1-ről ez a módszer is megerősítette. A Vif, Vpr
és Vpu fehérjék létét közvetett módon, szekvenálással igazoltuk. Eredményeink a
szekvenálási adatokkal együtt alátámasztják a pDNS-ben kódolt tizenhárom intakt
fehérje termelődését, valamint a két módosított génről a natív fehérje termelődésének
hiányát.
6.1.2. VLP+ termelés
Mivel a pLWXu1 tizenhárom teljes hosszúságú HIV fehérjét expresszál, feltételeztük,
hogy ezek komplex vírusszerű részecskévé (VLP+) képesek összeállni. A “+” index a
komplex VLP megkülönböztetésére szolgál a hagyományos, általában egy ill. kétfajta
struktúrfehérjéből spontán összeszerelődő VLP-ktől [93]. A pLWXu1 VLP+
termelésének igazolására humán 293T sejteket transzfektáltunk pLWXu1 pDNS-t
tartalmazó nanomedicinával, és egy napos inkubálást követően transzmissziós
elektronmikroszkóppal vizsgáltuk a mintákat (11.A ábra). A felvételeken jól láthatók a
vad típusú HIV-re jellemző fenotípusú VLP+ részecskék, melyek az érett virionhoz
hasonló mag (core) és burok (envelop) struktúrákat tartalmaznak. Megfigyeltük
továbbá, hogy a VLP+ részecskék a sejten kívüli térbe, vagy intracitoplazmatikus
vakuolumokba fűződnek le, és a virion érési fázisaihoz hasonló stádiumok különíthetők
el [92]. A VLP+ összetett felépítését tisztított VLP+ felhasználásával végzett western
blot is igazolta (11.B ábra).
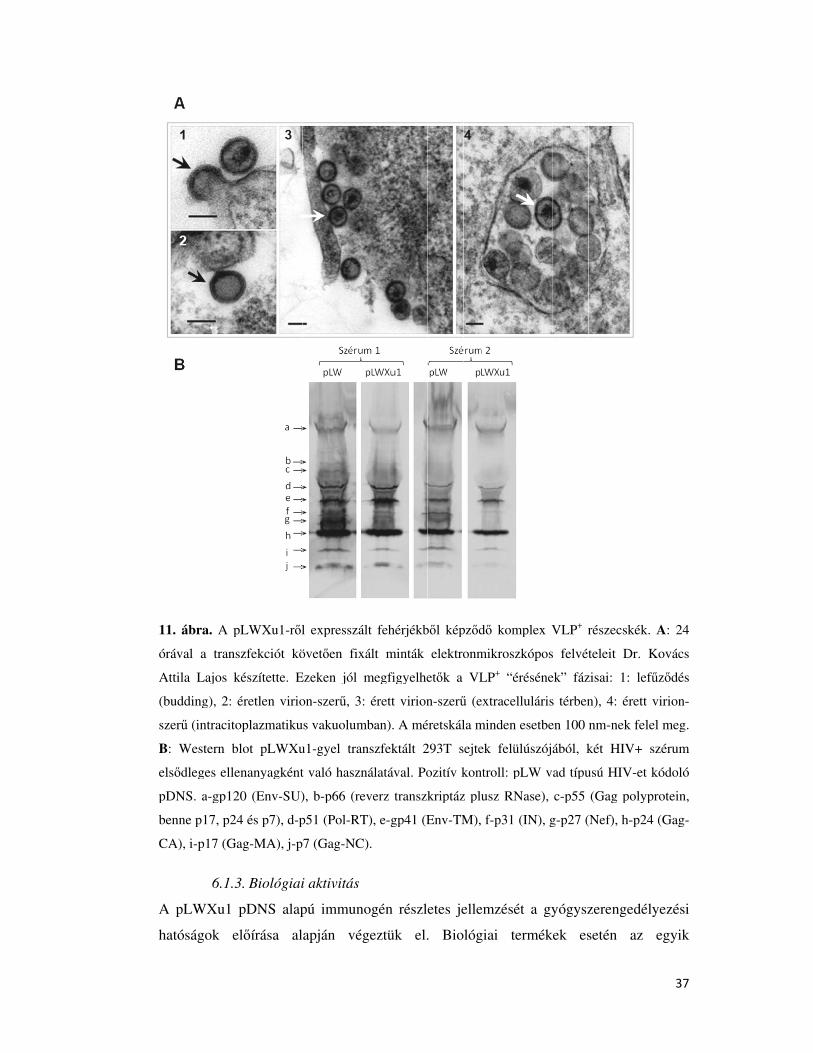
11
órával a transzfekciót követ
Attila Lajos készítette. Ezeken jól megfigyelhet
(budding), 2:
szer
B
els
pDNS. a
benne p17, p24 és p7), d
CA), i
A pLWXu1 pDNS alapú immunogén részletes jellemzését a
hatóságok el
11. ábra
órával a transzfekciót követ
Attila Lajos készítette. Ezeken jól megfigyelhet
(budding), 2:
szerű
B: Western blot pLWXu1
első
pDNS. a
benne p17, p24 és p7), d
CA), i
A pLWXu1 pDNS alapú immunogén részletes jellemzését a
hatóságok el
. ábra
órával a transzfekciót követ
Attila Lajos készítette. Ezeken jól megfigyelhet
(budding), 2:
szerű (intracitoplazmatikus vakuolumban). A méretskála minden esetben 100 nm
: Western blot pLWXu1
elsődleges ellenanyagként való használatával. Pozitív kontroll: pLW vad típusú HIV
pDNS. a
benne p17, p24 és p7), d
CA), i-
A pLWXu1 pDNS alapú immunogén részletes jellemzését a
hatóságok el
. ábra.
órával a transzfekciót követ
Attila Lajos készítette. Ezeken jól megfigyelhet
(budding), 2:
ű (intracitoplazmatikus vakuolumban). A méretskála minden esetben 100 nm
: Western blot pLWXu1
ődleges ellenanyagként való használatával. Pozitív kontroll: pLW vad típusú HIV
pDNS. a-gp120 (Env
benne p17, p24 és p7), d
-p17 (Gag
6.1.3.
A pLWXu1 pDNS alapú immunogén részletes jellemzését a
hatóságok el
A pLWXu1
órával a transzfekciót követ
Attila Lajos készítette. Ezeken jól megfigyelhet
(budding), 2:
(intracitoplazmatikus vakuolumban). A méretskála minden esetben 100 nm
: Western blot pLWXu1
dleges ellenanyagként való használatával. Pozitív kontroll: pLW vad típusú HIV
gp120 (Env
benne p17, p24 és p7), d
p17 (Gag
6.1.3.
A pLWXu1 pDNS alapú immunogén részletes jellemzését a
hatóságok el
A pLWXu1
órával a transzfekciót követ
Attila Lajos készítette. Ezeken jól megfigyelhet
(budding), 2: éretlen virion
(intracitoplazmatikus vakuolumban). A méretskála minden esetben 100 nm
: Western blot pLWXu1
dleges ellenanyagként való használatával. Pozitív kontroll: pLW vad típusú HIV
gp120 (Env
benne p17, p24 és p7), d
p17 (Gag
6.1.3. Biológiai aktivitás
A pLWXu1 pDNS alapú immunogén részletes jellemzését a
hatóságok előírása alapján végeztük el. Biológiai termékek esetén az egyik
A pLWXu1
órával a transzfekciót követ
Attila Lajos készítette. Ezeken jól megfigyelhet
éretlen virion
(intracitoplazmatikus vakuolumban). A méretskála minden esetben 100 nm
: Western blot pLWXu1
dleges ellenanyagként való használatával. Pozitív kontroll: pLW vad típusú HIV
gp120 (Env
benne p17, p24 és p7), d
p17 (Gag-MA), j
Biológiai aktivitás
A pLWXu1 pDNS alapú immunogén részletes jellemzését a
hatóságok előírása alapján végeztük el. Biológiai termékek esetén az egyik
A pLWXu1
órával a transzfekciót követ
Attila Lajos készítette. Ezeken jól megfigyelhet
éretlen virion
(intracitoplazmatikus vakuolumban). A méretskála minden esetben 100 nm
: Western blot pLWXu1
dleges ellenanyagként való használatával. Pozitív kontroll: pLW vad típusú HIV
gp120 (Env-SU), b
benne p17, p24 és p7), d
MA), j
Biológiai aktivitás
A pLWXu1 pDNS alapú immunogén részletes jellemzését a
őírása alapján végeztük el. Biológiai termékek esetén az egyik
A pLWXu1-ről e
órával a transzfekciót követ
Attila Lajos készítette. Ezeken jól megfigyelhet
éretlen virion
(intracitoplazmatikus vakuolumban). A méretskála minden esetben 100 nm
: Western blot pLWXu1
dleges ellenanyagként való használatával. Pozitív kontroll: pLW vad típusú HIV
SU), b
benne p17, p24 és p7), d-p51 (Pol
MA), j-p7 (Gag
Biológiai aktivitás
A pLWXu1 pDNS alapú immunogén részletes jellemzését a
írása alapján végeztük el. Biológiai termékek esetén az egyik
ről expresszált fehérjékb
órával a transzfekciót követő
Attila Lajos készítette. Ezeken jól megfigyelhet
éretlen virion-szer
(intracitoplazmatikus vakuolumban). A méretskála minden esetben 100 nm
: Western blot pLWXu1-gyel transzfektált 293T sejtek felülúszójából, két HIV
dleges ellenanyagként való használatával. Pozitív kontroll: pLW vad típusú HIV
SU), b-
p51 (Pol
p7 (Gag
Biológiai aktivitás
A pLWXu1 pDNS alapú immunogén részletes jellemzését a
írása alapján végeztük el. Biológiai termékek esetén az egyik
xpresszált fehérjékb
órával a transzfekciót követően fixált minták elektronmikroszkópos felvételeit Dr. Kovács
Attila Lajos készítette. Ezeken jól megfigyelhet
szerű
(intracitoplazmatikus vakuolumban). A méretskála minden esetben 100 nm
gyel transzfektált 293T sejtek felülúszójából, két HIV
dleges ellenanyagként való használatával. Pozitív kontroll: pLW vad típusú HIV
-p66 (reverz transzkriptáz plusz RNase), c
p51 (Pol
p7 (Gag
Biológiai aktivitás
A pLWXu1 pDNS alapú immunogén részletes jellemzését a
írása alapján végeztük el. Biológiai termékek esetén az egyik
xpresszált fehérjékb
ően fixált minták elektronmikroszkópos felvételeit Dr. Kovács
Attila Lajos készítette. Ezeken jól megfigyelhet
szerű, 3: érett virion
(intracitoplazmatikus vakuolumban). A méretskála minden esetben 100 nm
gyel transzfektált 293T sejtek felülúszójából, két HIV
dleges ellenanyagként való használatával. Pozitív kontroll: pLW vad típusú HIV
p66 (reverz transzkriptáz plusz RNase), c
p51 (Pol-RT), e
p7 (Gag-NC).
Biológiai aktivitás
A pLWXu1 pDNS alapú immunogén részletes jellemzését a
írása alapján végeztük el. Biológiai termékek esetén az egyik
xpresszált fehérjékb
en fixált minták elektronmikroszkópos felvételeit Dr. Kovács
Attila Lajos készítette. Ezeken jól megfigyelhet
ű, 3: érett virion
(intracitoplazmatikus vakuolumban). A méretskála minden esetben 100 nm
gyel transzfektált 293T sejtek felülúszójából, két HIV
dleges ellenanyagként való használatával. Pozitív kontroll: pLW vad típusú HIV
p66 (reverz transzkriptáz plusz RNase), c
RT), e
NC).
Biológiai aktivitás
A pLWXu1 pDNS alapú immunogén részletes jellemzését a
írása alapján végeztük el. Biológiai termékek esetén az egyik
xpresszált fehérjékb
en fixált minták elektronmikroszkópos felvételeit Dr. Kovács
Attila Lajos készítette. Ezeken jól megfigyelhet
, 3: érett virion
(intracitoplazmatikus vakuolumban). A méretskála minden esetben 100 nm
gyel transzfektált 293T sejtek felülúszójából, két HIV
dleges ellenanyagként való használatával. Pozitív kontroll: pLW vad típusú HIV
p66 (reverz transzkriptáz plusz RNase), c
RT), e-gp41 (Env
A pLWXu1 pDNS alapú immunogén részletes jellemzését a
írása alapján végeztük el. Biológiai termékek esetén az egyik
xpresszált fehérjékb
en fixált minták elektronmikroszkópos felvételeit Dr. Kovács
Attila Lajos készítette. Ezeken jól megfigyelhet
, 3: érett virion
(intracitoplazmatikus vakuolumban). A méretskála minden esetben 100 nm
gyel transzfektált 293T sejtek felülúszójából, két HIV
dleges ellenanyagként való használatával. Pozitív kontroll: pLW vad típusú HIV
p66 (reverz transzkriptáz plusz RNase), c
gp41 (Env
A pLWXu1 pDNS alapú immunogén részletes jellemzését a
írása alapján végeztük el. Biológiai termékek esetén az egyik
xpresszált fehérjékbő
en fixált minták elektronmikroszkópos felvételeit Dr. Kovács
Attila Lajos készítette. Ezeken jól megfigyelhet
, 3: érett virion
(intracitoplazmatikus vakuolumban). A méretskála minden esetben 100 nm
gyel transzfektált 293T sejtek felülúszójából, két HIV
dleges ellenanyagként való használatával. Pozitív kontroll: pLW vad típusú HIV
p66 (reverz transzkriptáz plusz RNase), c
gp41 (Env
A pLWXu1 pDNS alapú immunogén részletes jellemzését a
írása alapján végeztük el. Biológiai termékek esetén az egyik
xpresszált fehérjékből képz
en fixált minták elektronmikroszkópos felvételeit Dr. Kovács
Attila Lajos készítette. Ezeken jól megfigyelhetők a VLP
, 3: érett virion-szer
(intracitoplazmatikus vakuolumban). A méretskála minden esetben 100 nm
gyel transzfektált 293T sejtek felülúszójából, két HIV
dleges ellenanyagként való használatával. Pozitív kontroll: pLW vad típusú HIV
p66 (reverz transzkriptáz plusz RNase), c
gp41 (Env
A pLWXu1 pDNS alapú immunogén részletes jellemzését a
írása alapján végeztük el. Biológiai termékek esetén az egyik
ől képz
en fixált minták elektronmikroszkópos felvételeit Dr. Kovács
Attila Lajos készítette. Ezeken jól megfigyelhetők a VLP
szerű
(intracitoplazmatikus vakuolumban). A méretskála minden esetben 100 nm
gyel transzfektált 293T sejtek felülúszójából, két HIV
dleges ellenanyagként való használatával. Pozitív kontroll: pLW vad típusú HIV
p66 (reverz transzkriptáz plusz RNase), c
gp41 (Env-TM), f
A pLWXu1 pDNS alapú immunogén részletes jellemzését a
írása alapján végeztük el. Biológiai termékek esetén az egyik
l képződő
en fixált minták elektronmikroszkópos felvételeit Dr. Kovács
ők a VLP
szerű (extracelluláris térben), 4: érett virion
(intracitoplazmatikus vakuolumban). A méretskála minden esetben 100 nm
gyel transzfektált 293T sejtek felülúszójából, két HIV
dleges ellenanyagként való használatával. Pozitív kontroll: pLW vad típusú HIV
p66 (reverz transzkriptáz plusz RNase), c
TM), f
A pLWXu1 pDNS alapú immunogén részletes jellemzését a
írása alapján végeztük el. Biológiai termékek esetén az egyik
ődő komplex VLP
en fixált minták elektronmikroszkópos felvételeit Dr. Kovács
k a VLP+
ű (extracelluláris térben), 4: érett virion
(intracitoplazmatikus vakuolumban). A méretskála minden esetben 100 nm
gyel transzfektált 293T sejtek felülúszójából, két HIV
dleges ellenanyagként való használatával. Pozitív kontroll: pLW vad típusú HIV
p66 (reverz transzkriptáz plusz RNase), c
TM), f-p31 (I
A pLWXu1 pDNS alapú immunogén részletes jellemzését a
írása alapján végeztük el. Biológiai termékek esetén az egyik
ő komplex VLP
en fixált minták elektronmikroszkópos felvételeit Dr. Kovács + “érésének” fázisai: 1: lef
(extracelluláris térben), 4: érett virion
(intracitoplazmatikus vakuolumban). A méretskála minden esetben 100 nm
gyel transzfektált 293T sejtek felülúszójából, két HIV
dleges ellenanyagként való használatával. Pozitív kontroll: pLW vad típusú HIV
p66 (reverz transzkriptáz plusz RNase), c
p31 (I
A pLWXu1 pDNS alapú immunogén részletes jellemzését a
írása alapján végeztük el. Biológiai termékek esetén az egyik
komplex VLP
en fixált minták elektronmikroszkópos felvételeit Dr. Kovács
“érésének” fázisai: 1: lef
(extracelluláris térben), 4: érett virion
(intracitoplazmatikus vakuolumban). A méretskála minden esetben 100 nm
gyel transzfektált 293T sejtek felülúszójából, két HIV
dleges ellenanyagként való használatával. Pozitív kontroll: pLW vad típusú HIV
p66 (reverz transzkriptáz plusz RNase), c
p31 (IN), g
A pLWXu1 pDNS alapú immunogén részletes jellemzését a
írása alapján végeztük el. Biológiai termékek esetén az egyik
komplex VLP
en fixált minták elektronmikroszkópos felvételeit Dr. Kovács
“érésének” fázisai: 1: lef
(extracelluláris térben), 4: érett virion
(intracitoplazmatikus vakuolumban). A méretskála minden esetben 100 nm
gyel transzfektált 293T sejtek felülúszójából, két HIV
dleges ellenanyagként való használatával. Pozitív kontroll: pLW vad típusú HIV
p66 (reverz transzkriptáz plusz RNase), c
N), g
A pLWXu1 pDNS alapú immunogén részletes jellemzését a gyógyszerengedélyezési
írása alapján végeztük el. Biológiai termékek esetén az egyik
komplex VLP
en fixált minták elektronmikroszkópos felvételeit Dr. Kovács
“érésének” fázisai: 1: lef
(extracelluláris térben), 4: érett virion
(intracitoplazmatikus vakuolumban). A méretskála minden esetben 100 nm
gyel transzfektált 293T sejtek felülúszójából, két HIV
dleges ellenanyagként való használatával. Pozitív kontroll: pLW vad típusú HIV
p66 (reverz transzkriptáz plusz RNase), c-p55 (Gag polyprotein,
N), g-p27 (Nef), h
gyógyszerengedélyezési
írása alapján végeztük el. Biológiai termékek esetén az egyik
komplex VLP+ részecskék.
en fixált minták elektronmikroszkópos felvételeit Dr. Kovács
“érésének” fázisai: 1: lef
(extracelluláris térben), 4: érett virion
(intracitoplazmatikus vakuolumban). A méretskála minden esetben 100 nm
gyel transzfektált 293T sejtek felülúszójából, két HIV
dleges ellenanyagként való használatával. Pozitív kontroll: pLW vad típusú HIV
p55 (Gag polyprotein,
p27 (Nef), h
gyógyszerengedélyezési
írása alapján végeztük el. Biológiai termékek esetén az egyik
részecskék.
en fixált minták elektronmikroszkópos felvételeit Dr. Kovács
“érésének” fázisai: 1: lef
(extracelluláris térben), 4: érett virion
(intracitoplazmatikus vakuolumban). A méretskála minden esetben 100 nm-
gyel transzfektált 293T sejtek felülúszójából, két HIV
dleges ellenanyagként való használatával. Pozitív kontroll: pLW vad típusú HIV
p55 (Gag polyprotein,
p27 (Nef), h
gyógyszerengedélyezési
írása alapján végeztük el. Biológiai termékek esetén az egyik
részecskék.
en fixált minták elektronmikroszkópos felvételeit Dr. Kovács
“érésének” fázisai: 1: lef
(extracelluláris térben), 4: érett virion
-nek felel meg.
gyel transzfektált 293T sejtek felülúszójából, két HIV
dleges ellenanyagként való használatával. Pozitív kontroll: pLW vad típusú HIV
p55 (Gag polyprotein,
p27 (Nef), h
gyógyszerengedélyezési
írása alapján végeztük el. Biológiai termékek esetén az egyik
részecskék.
en fixált minták elektronmikroszkópos felvételeit Dr. Kovács
“érésének” fázisai: 1: lef
(extracelluláris térben), 4: érett virion
nek felel meg.
gyel transzfektált 293T sejtek felülúszójából, két HIV+ szérum
dleges ellenanyagként való használatával. Pozitív kontroll: pLW vad típusú HIV-et kódoló
p55 (Gag polyprotein,
p27 (Nef), h-p24 (Gag
gyógyszerengedélyezési
írása alapján végeztük el. Biológiai termékek esetén az egyik
részecskék. A
en fixált minták elektronmikroszkópos felvételeit Dr. Kovács
“érésének” fázisai: 1: lefűző
(extracelluláris térben), 4: érett virion
nek felel meg.
+ szérum
et kódoló
p55 (Gag polyprotein,
p24 (Gag
gyógyszerengedélyezési
írása alapján végeztük el. Biológiai termékek esetén az egyik
37
A: 24
en fixált minták elektronmikroszkópos felvételeit Dr. Kovács
“érésének” fázisai: 1: lefűződés
(extracelluláris térben), 4: érett virion
nek felel meg.
+ szérum
et kódoló
p55 (Gag polyprotein,
p24 (Gag
gyógyszerengedélyezési
írása alapján végeztük el. Biológiai termékek esetén az egyik
37
: 24
en fixált minták elektronmikroszkópos felvételeit Dr. Kovács
ű ődés
(extracelluláris térben), 4: érett virion-
nek felel meg.
+ szérum
et kódoló
p55 (Gag polyprotein,
p24 (Gag-
gyógyszerengedélyezési
írása alapján végeztük el. Biológiai termékek esetén az egyik

38
kulcsfontosságú paraméter a biológiai aktivitás, azaz esetünkben az antigén expressziós
hatékonyság meghatározása. A biológiai aktivitás in vitro meghatározására egy olyan
módszert optimalizáltunk és validáltunk, mely humán sejtvonal transzfekcióját
követően a felülúszóba szekretált HIV specifikus fehérje, a p24 (Gag-CA) mennyiségi
mérésével történik.
A p24 fehérje a pLWXu1-ről a HIV virionhoz hasonló módon, csak a transzkripció
késői szakaszában termelődik, és expressziójához a transzkripció korai szakaszában már
átíródó szabályozó fehérjék szükségesek. Azt találtuk, hogy a transzfektált 293T sejtek
felülúszójában található p24 mennyiségének 92.1%-a VLP+ részecskék alkotóelemeként
van jelen (12. ábra). Ezért a p24 mennyiségi kimutatásával mérhető a pDNS
konstrukcióban kódolt fehérjekészlet-, valamint a VLP+ expressziója is, azaz a pDNS
biológiai aktivitása [92].
12. ábra. A pLWXu1 biológiai aktivitása. A nanomedicinával transzfektált 293T sejtek által
termelt összes p24 92,1%-a (62,7 ng/ml) VLP+ alkotóelemként van jelen a transzfektált sejtek
felülúszójában (*P<0,001).
Az optimalizált (sztenderdizált) módszert az európai ICH Q2(R1) analitikai módszerek
validálására vonatkozó irányelvek alapján validáltuk [78]. A validálás lényege, hogy
igazoljuk a mérési módszer alkalmasságát a tervezett célra: a DermaVir vizsgálati
anyag biológiai aktivitásának in vitro meghatározására a pDNS hatóanyag által
expresszált p24 antigén mennyiségi mérésével. Ehhez a módszer következő
paramétereit kell meghatározni: pontosság (accuracy), ismételhetőség (precision)
(napok közti variabilitás: különböző napokon ugyanaz a személy végzi el a mérést;
0
10
20
30
40
50
60
70
80
90
Összes p24 Nem-VLP+ alkotó p24
Bio
lóg
iai a
ktiv
itás
(p2
4 n
g/m
l)
*

39
minták közti variabilitás: két személy ugyanazon vizsgálati anyagból származó
mintákkal végzi el a mérést; mintán belüli variabilitás: egy személy a vizsgálati anyag
5 különböző adagjából vett 3-3 mintával végzi el a mérést; mérést végző személyek közti
variabilitás: azonos napon többen is elvégzik a mérést), specifikusság (a módszer
specifikus a kívánt paraméter meghatározására, ebben az esetben a DermaVir
formuláció p24 fehérje expresszióját képes mérni), mérési tartomány, kimutatási határ
és a mennyiségi meghatározásra alkalmas alsó méréshatár, linearitás (koncentráció
függés, vagyis annak igazolása, hogy a módszer a mérési tartományban kvantitatív).
Minden mérés során 5 párhuzamossal dolgoztunk. A kívánt paramétereket
meghatároztunk a pontosság kivételével, amelyben egy minősített sztenderd anyaghoz
kell hasonlítani a mintánkat, csakhogy a DermaVir esetében nincs ilyen összehasonlító
anyag. Eredményeink összefoglaló számadatai a következők: ismételhetőség (precision,
n=49): [p24] = 45,19 ng/ml; CV% (variációs együttható, az adatcsoport átlagtól való
eltérése osztva az átlaggal, százalékban kifejezve) = 14,12 %. Specifikusság:
DermaVir[p24] = 43,80 ng/ml, CV (%) = 4,4 %. A formuláció nélküli csupasz pDNS
esetében a kimutatási határ alatti eredményeket kaptunk. Szintén a méréshatár alatti
eredményt kaptunk a p24 fehérjét expresszálni nem képes, aspecifikus pDNS-t
tartalmazó nanorészecske formuláció esetében is. Ezzel igazoltuk, hogy módszerünk a
DermaVir formuláció biológiai aktivitásának mérésére specifikus. Mérési tartomány:
0,5-0,8 µg pDNS-t tartalmazó DermaVir. Linearitás: a mérési tartományon belül 7
adatpontra határoztuk meg, ahol a 0,7 µg pDNS/DermaVir az alapértelmezett vizsgálati
mennyiség, és ennek 70-120%-át jelenti a többi vizsgálati pont. A linearitás mérés
adataira illesztett egyenes egyenlete: y=0,472x-22,636, korrelációs együtthatója
R2=0,83, ez megfelel az R2>0,8 elfogadási kritériumnak. A kimutatási határ a vizsgálati
anyag kalibrációs egyeneséből (linearitás adatok) és a negatív kontroll mintákra kapott
p24 adatok átlagtól való eltérése alapján (SD%=0,78) 16,60 pg/ml-nek adódott.
A validálás során minden előírt paramétert meghatároztunk, valamint igazoltuk a
módszer alkalmasságát a DermaVir biológiai aktivitásának pontos, specifikus és
reprodukálható mennyiségi meghatározására.

40
6.2. A nanomedicina immunológiai potenciáljának in silico modellezése
6.2.1. T sejt epitóp repertoár analízis kidolgozása
Jelen fejezetben az in silico immunológiai potenciál modellezésének kidolgozásáról
lesz szó: hogyan választottuk ki a T sejt epitóp predikciós módszereket és a felhasznált
HIV szekvencia adatbázist. Ezt követően a kiválasztott módszerek megbízhatóságának
tesztelését, és az általunk bevezetett kvantitatív és kvalitatív mérőszámok: az abszolút
immunológiai potenciál (AIP) és a relatív immunológiai potenciál (RIP) definícióját és
alkalmazását mutatom be a DermaVir nanomedicina pDNS hatóanyagának (pLWXu1)
példáján. Ezután pedig a nanomedicina keresztvédő képességének modellezését
különböző szubtípusú fertőzésekre. A személyre szabott immunterápia tervezése is
ugyanezen T sejt predikciós módszeren alapul, ami alapján kifejlesztettünk egy
immunológiai hasonlósági csoportokat megjelenítő új vakcinatervező “eszközt”, az
immunológiai fá-t. Utóbbi lényegét és alkalmazását a 6.3. fejezetben részletezem.
Mivel lényegében a T sejt epitópok határozzák meg egy T sejtes immunválaszt kiváltó
terápiás vakcina immunológiai karakterét, célul tűztük ki a pLWXu1-ről termelődő
fehérje antigének elméleti T sejt epitóp repertoárjának mennyiségi és minőségi
vizsgálatát bioinformatikai eszközökkel. Az analízishez a legmegbízhatóbb módszerek
és adatbázis kiválasztására törekedtünk. A legnagyobb és leghomogénebb adatokat
tartalmazó T sejt epitóp adatbázist, az “Immune Epitope Database”-t (IEDB)
használtuk. A legmegbízhatóbb in silico epitóp predikciós módszer kiválasztásához
összehasonlítottuk a stabilizált mátrix módszer (Stabilized Matrix Method, SMM), az
átlagos relatív kötésen alapuló módszer (Average Relative Binding, ARB) és a
mesterséges neurális hálózat alapú módszer (Artificial Neural Network, ANN)
megbízhatóságát. Ehhez a HXB2 HIV-1 referencia törzs struktúrfehérjéit választottuk
(GenBank azonosító: NC_001802), és az IEDB adatbázis segítségével, mindhárom
módszerrel prediktáltuk a nagy affinitású MHCI epitópokat, majd összehasonlítottuk
azokat a “HIV Molecular Immunology” című kiadványban publikált, kísérletesen
igazolt epitópokkal [83]. (A nagy affinitású epitópok definíció szerint 50 nM-nál kisebb
fél-maximális gátlási koncentrációval – IC50 értékkel – jellemezhetők.)
A megbízhatóságot a kísérletes adatokkal korrekt egyezést mutató (teljesen egyező-,
plusz az egy as eltérést tartalmazó) epitópok százalékban kifejezett arányaként adtuk
meg. Az ANN predikciós módszer bizonyult a legmegbízhatóbbnak, az Env-TM (gp41)

41
fehérje esetén például a prediktált epitópok 74,5%-a mutatott korrekt egyezést a
kísérletesen igazolt, valós epitópokkal, míg az SMM 66,4%-os, az ARB módszer pedig
57,6%-os predikciós megbízhatóságot eredményezett.
Ezután meghatároztuk az ANN MHCI predikciós módszer megbízhatóságát a HXB2
összes struktúrfehérjéjének felhasználásával, ugyanarra az allélre. A legalacsonyabb,
70,5%-os megbízhatósági értéket a gp120 eredményezte. Az MHCII epitóp predikciós
“Konszenzus módszert” publikált validálási eredmények alapján választottuk [81]. A
továbbiakban tehát az ANN predikciós módszerrel, valamint a Konszenzus módszerrel
prediktáltuk az MHCI és MHCII epitópokat mind a tizenöt pLWXu1 által kódolt teljes
vagy csonka fehérjére [92]. Ezen predikciók, és a szintén ebben a fejezetben részletezett
keresztvédettség elemzések elvégzése után az addig általunk prediktált összes epitóp
felhasználásával ismét elvégeztük az ANN ill. Konszenzus predikciós módszerek
megbízhatóságának vizsgálatát, most már az összes prediktálható allélre (57 MHCI és
63 MHCII allélre). Eredményül 97,7%-os megbízhatóságot kaptunk az MHCI
predikciós módszerre, és 95,1%-os megbízhatóságot az MHCII predikciós módszerre
(2. Táblázat). Ez azt jelenti, hogy a kísérletesen igazolt 1480 CD8+ T sejt specifikus
MHCI epitóp 97,7%-át prediktáltuk pontos-, vagy korrekt egyezéssel. Az MHCII
esetében pedig a 615 kísérletesen igazolt CD4+ T sejt specifikus MHCII epitóp 95,1%-
át sikerült pontos-, ill. korrekt egyezéssel prediktálnunk.
2. táblázat. Az MHCI és MHCII epitóp predikciós módszerek megbízhatóságának vizsgálata
kísérletesen igazolt epitópok segítségével.
Predikcióink alapján a pLWXu1 által expresszált fehérjék összesen 933 nagy affinitású
(CD8+ T sejtek érését indukáló) MHCI epitópot, és 2330 nagy affinitású (CD4+ T
sejtes választ eredményező) MHCII epitópot tartalmaznak az IEDB adatbázisban
elérhető összes gyakori humán MHCI allélre (n=57) valamint MHCII allélre (n=63).
MHCI MHCII
Megengedett eltérések száma az epitópban 1 1
Adathalmaz szűkítése (erős epitópok <50 nM <50
Kísérletesen igazolt epitópok száma* 1480 615
Predikcióval „megtalált„ epitópok száma 1436 585
A predikció megbízhatósága 97,7% 95,1%
*HIV Molecular Immunology 2008

42
Fontos megjegyezni, hogy ez az epitóp repertoár nem egy adott egyénben kifejeződő
epitópokat jelenti, hanem a teljes humán populációban elméletileg képződő HIV epitóp
repertoárt. Vagyis ez az összegzett érték a pLWXu1 által expresszált fehérje készlet
elméleti epitóp mennyiségét adja meg, amely T sejtes immunválasz kiváltását
indukálhatja a humán populációban.
6.2.2. Abszolút immunológiai potenciál (AIP)
Az immunológiai potenciál részletesebb jellemzéséhez bevezettük az abszolút
immunológiai potenciál (AIP) fogalmát, ami megmutatja, hogyan oszlik meg az egyes
fehérjékre specifikus epitópok mennyisége a teljes proteom immunológiai potenciálján
belül. Az AIP tehát minden fehérjének megadja az antigén-specifikus T sejtes
immunválaszban képviselt elméleti részarányát. Az AIP-t az összes gyakori IEDB
adatbázisban található MHCI és MHCII allélre prediktáltuk.
Az alább részletezett eredményeink rámutattak a strukturális és szabályozó fehérjék
egyidejű kódolásának fontosságára a széles spektrumú immunválasz kiváltására
tervezett HIV vakcinánkban. A pLWXu1 által expresszált fehérjék a következő MHCI-
AIP sorrendet eredményezték: Env-TM (22,9%), Pol-RT (15,0%) és Env-SU (13,7%),
valamint Gag-CA (7,6%), Rev (6,6%), Vif (6,5%), Nef (5,6%), Vpu (4,2%), Gag-MA
(3,8%), Pol-RNáz (3,8%), Pol-IN (3,4%), Vpr (3,0%), Pol-Proteáz (2,0%), Tat (1,3%)
és Gag-NC (0,5%) (13. ábra). Az MHCII specifikus AIP pedig ezt a sorrendet mutatta:
Env-SU > Tat > Pol-RT > Gag-NC > Vif > Gag-CA > Gag-MA > Env-TM > Rev >
Nef > Vpr > Pol-IN > Pol-RNáz > Vpu > Pol-Proteáz. 17,3%-tól 2,0%-ig terjedő
értékekkel. Eredményeink megmutatták, milyen jelentős különbségek vannak az egyes
HIV fehérjék CD4+ ill. CD8+ T sejt indukáló képességében: az MHCI-AIP megoszlása
a struktúrfehérjék és szabályozófehérjék között 72,8% ill. 27,2%, az MHCII-AIP
esetében pedig 63,6% ill. 36,4%. A legmagasabb AIP részarányt a Gag-CA, Pol-RT,
Vif, Rev, Env-SU és Env-TM fehérjék képviselik a pLWXu1 expresszált fehérjéi közül.
A legnagyobb MHCI-AIP és MHCII-AIP értékek közti különbséget a Tat és Gag-NC
fehérhéknél találtuk. Ezek nagyon kis aktivitást mutatnak a CD8+ T sejtek
indukálására, viszont nagyon immunogénnek adódnak a CD4+ helper T sejtek
indukálására nézve. (Az Env-SU ötször hosszabb fehérje, mégis a Tat szinte
ugyanakkora részt tesz ki az MHCII-AIP-ből (15,0 %-ot), mint az Env-SU (17,3%).)

43
13. ábra. A pLWXu1 pDNS gyógyszerhatóanyag által expresszált antigén készlet abszolút
immunológiai potenciálja (AIP) MHCI és MHCII T sejt epitópok predikciója alapján. A
pLWXu1 által expresszált proteom AIP értékének az egyes fehérjék közti megoszlása
százalékban kifejezve: az MHCI molekulákon bemutatott epitópok által indukálható CD8+
specifikus T sejtek mennyisége (fekete oszlop), ill. az MHCII molekulákon bemutatott
epitópokkal indukálható CD4+ T sejtek mennyisége (szürke oszlop). A csillaggal jelölt fehérjék
nem teljes hosszúságúak; a fehérjék hossza (as) a fehérjék rövidítése mellett zárójelben
szerepel.
6.2.3. A DermaVir (pLWXu1) immunológiai potenciálja egy személyben
modellezve
A teljes populációra vonatkoztatható eredmények után lássuk, milyen egy hipotetikus
genetikai HLA háttérrel rendelkező személy AIP-je. A legáltalánosabb HLA alléleket is
hordozó hipotetikus személy genotípusa legyen a következő: HLA A*0250, 3001; HLA
B*0702, 0801 MHCI specifikus allélek, valamint DP A1*0103, 0201; DP B1*0201,
0501; DQ A1*0101, 0301; DQ B1*0501, 0302; DR A1*0101, 0101; DR B1*0101,
0102 MHCII specifikus allélek. (HLA C alléleket még nem tartalmazott az IEDB
adatbázis a vizsgálat idején, ezért nem szerepelnek a hipotetikus genotípusban sem.)
Az MHCI-AIP eredmények hasonló trendet mutattak egy személy esetén, mint a
minden fenti, populáció szintű predikció esetében. Csak három fehérje: Pol-RNase, Vpu
és a Pol-IN sorrendje módosult minimálisan (± 1 hellyel). Ezzel szemben az MHCII-
AIP nagyobb eltéréseket eredményezett a modell személy esetében: az Env-TM > Tat >
5.6
22.9
13.7
4.2
6.6
1.3
3.0
6.5
3.4
3.8
15.0
2.0
0.5
7.6
3.8
4.4
5.9
17.3
2.3
4.8
15.0
3.0
7.0
3.0
2.7
11.0
1.7
8.8
6.9
6.4
-30 -20 -10 0 10 20 30
Nef* (150)
TM (345)
SU (508)
Vpu (81)
Rev (115)
Tat (101)
Vpr (96)
Vif (192)
IN* (142)
RNase (120)
RT (440)
Prot. (99)
NC (55)
CA (231)
MA (132)
Env
Pol
Gag
pLW
Xu
1 á
ltal
exp
ress
zált
feh
érj
ék
Abszolút immunológiai potenciál (%)
30 20 10
MHCI MHCII

44
Pol-RT > Gag-NC > Env-TM > Gag-CA > Vif > Rev > Gag-MA > Vpr > Nef > Pol-IN
> Vpu > Pol-Proteáz > Pol-RNáz antigén sorrendet kaptuk [92]. Fontos megjegyezni,
hogy az egyes MHC allélek hozzájárulása immunológiai potenciálhoz nagyon eltérő, pl.
az MHCI epitópok mennyisége a különböző alléleken a következőképpen alakult:
A*0250-re 78 epitóp, A*3001-re 43 epitóp, A*0702-re 9 epitóp míg az A*0801 allélre
mindössze 1 nagy affinitású epitóp volt prediktálható. Tehát az immunválasz minősége
függ az egyén genetikai MHC hátterétől.
Ezek az eredmények jelzik, hogy módszerünk alkalmas lehet a DermaVirre (vagy más
antigénre) specifikus T sejtes immunválasz modellezésére minden személyben,
amennyiben a HLA genotípusa ismert. Ezért úgy gondoljuk a módszer segítséget
nyújthat a klinikai kísérletek immunogenitási eredményeinek értelmezése során is,
mivel prediktálhatjuk az immunogén immunválaszban mérhető hatását.
6.2.4. Relatív immunológiai potenciál (RIP)
Az egyes fehérjék T sejtes választ kiváltó potenciáljának további jellemzése céljából
definiáltuk a relatív immunológiai potenciált (RIP), amely az adott fehérjére specifikus
adat. A RIP megmutatja, hogy egy fehérjében mennyi nagy affinitású epitóp található
az elméletileg lehetséges epitópmennyiséghez képest, vagyis MHCI esetén a modell
szerinti 9 as hosszúságú, MHCII esetén 15 as hosszúságú peptidek maximális számához
képest. Mivel az MHCI epitópok hossza 9 as a predikcióinkban, az MHCII epitópok
hossza pedig 15 as, ezért az elméleti epitóp szám MHCI esetén a fehérje hossza mínusz
8, MHCII esetén a fehérje hossza mínusz 14. A RIP értékét a prediktált nagy affinitású
epitópok számának, valamint az elméleti epitópok számának hányadosa adja,
százalékban kifejezve. A RIP tehát a fehérje epitópsűrűségének mérőszáma.
Az MHCI-RIP az Env-TM struktúrfehérje (214/337; 63,5%) és a Rev szabályozó
fehérje (62/107; 57,9%) esetében volt a legmagasabb, de a Gag-CA (71/124; 57,3%), és
a Vpu is magas relatív immunológiai potenciált eredményezett (39/73; 53,4%) (14.
ábra). A legalacsonyabb értékek a Tat (12/93; 12,9%) és Gag-NC fehérjékre (5/47;
10,6%) adódtak.
A legnagyobb MHCII-RIP értékeket a Gag-NC (204/41; 497,6%) és Tat (349/87;
401,1%) fehérjékre számoltuk, ami összecseng ezen fehérjék magas MHCII-AIP
részarányával. 100%-nál magasabb RIP értéket akkor kaphatunk, ha a vizsgált fehérje
bizonyos mértékben “MHC-független” erős epitópokat is tartalmaz, vagyis ha egy

45
epitóp több MHCII allélre is prediktálható (ekkor az ismétlések is beleszámítandók az
összes prediktált epitóp mennyiségbe). A Gag-CA szintén magas RIP-pel jellemezhető
(160/118; 135,6%), ezután pedig a Rev > Vif > Vpr > Env-SU > Vpu > Nef > Gag-MA
> Pol-RT > Pol-RNáz > Pol-IN > Pol-Proteáz > Env-TM fehérjék következnek a sorban
[92].
Az MHCI-RIP és MHCII-RIP értékek tehát nagyban különböznek, ami szintén jelzi a
sokféle fehérje antigént tartalmazó pLWXu1 pDNS alapú vakcina immunológiai
potenciálját széles specifitású CD4+ és CD8+ T sejtes immunválasz kiváltására.
14. ábra. A pLWXu1 pDNS gyógyszerhatóanyag által expresszált antigén készlet relatív
immunológiai potenciálja (RIP) MHCI és MHCII T sejt epitópok predikciója alapján. A
pLWXu1 expresszált antigén készletének RIP értéke, vagyis az egyes fehérjéken belüli epitóp
sűrűség ábrázolása: az MHCI specifikus epitópok (fekete oszlop) aránya az adott fehérjében
elméletileg lehetséges 9 as hosszú epitópok mennyiséghez képest (fehér oszlop), ill. az MHCII
specifikus epitópok aránya az adott fehérjében elméletileg lehetséges 15 as hosszú epitópok
mennyiséghez képest (fehér oszlop). A csillaggal jelölt fehérjék nem teljes hosszúságúak; a
fehérjék hossza (as) a fehérjék rövidítése mellett zárójelben szerepel.
-600 -400 -200 0 200 400 600
Nef* (150)
TM (345)
SU (508)
Vpu (81)
Rev (115)
Tat (101)
Vpr (96)
Vif (192)
IN* (142)
RNase (120)
RT (440)
Prot. (99)
NC (55)
CA (132)
MA (231)
Env
Pol
Gag
pL
WX
u1
de
rive
dp
rote
ins
600 400 200
Number of theoretical and predicted epitopes
-600 -400 -200 0 200 400 600
Nef* (150)
TM (345)
SU (508)
Vpu (81)
Rev (115)
Tat (101)
Vpr (96)
Vif (192)
IN* (142)
RNase (120)
RT (440)
Prot. (99)
NC (55)
CA (231)
MA (132)
Env
Pol
Gag
pL
WX
u1
álta
l e
xpre
sszá
ltfe
hé
rjé
k
Elméleti- és prediktált epitópok száma
600 400 200
MHCI MHCII

46
6.2.5. Nem-B szubtípusú fertőzés esetén a B-szubtípus alapú terápia
hatékonyságának in silico modellezése
A fent ismertetett in silico T sejt epitóp predikciós módszer segítségével modelleztük,
hogy a B szubtípusra specifikus T sejtes immunválasz mennyire lehet hatékony nem B-
szubtípusú HIV fertőzés esetén.
Mivel az Afrikában (Szaharától délre) domináló C szubtípusú HIV a legjelentősebb
járványokozó bolygónkon, részletes összehasonlításukat ezzel a szubtípussal mutatom
be. Először egyetlen gyakori MHCI specifikus HLA allélre, részletesen vizsgáljuk meg
a keresztvédettséget a közös epitópok számba vételével! A HLA A*0250 allélre
vonatkozó eredményeinket a 3. táblázatban foglaltam össze. A nanomedicina által
expresszált fehérjekészlet 104 nagy affinitású epitópja közül B szubtípusú (vagyis
szubtípus-azonos) fertőzés esetén 93 teljesen megegyezik, vagyis a prediktált pLWXu1
epitópok 89%-a hatékony CD8+ T sejtes válasz indukálására lehet képes, amennyiben a
kezelt beteg az elemzett B szubtípusú HIV-vel fertőzött (3.A táblázat).
C szubtípusú fertőzés esetén azonban sokkal alacsonyabb a nanomedicinából származó
epitópok, és a fertőzést okozó HIV-ből származó epitópok közti egyezés mértéke:
mindössze 13 egyező epitópot találtunk, amik a Gag, Pol és Env poliproteinekre
korlátozódtak (3.B táblázat). Ez mérsékelt keresztvédettségre utal, más szóval a B
szubtípusú fehérjéket expresszáló vakcina alacsony hatékonyságát jelzi C szubtípusú
HIV-vel fertőzött betegben.
3. táblázat. A B szubtípus specifikus pLWXu1 nanomedicina hatékonyságának modellezése C
szubtípusú HIV-vel fertőzött betegben, T sejt epitóp predikcióval az MHCI specifikus HLA
A*0250 allélre. A: Összefoglaló táblázat a nanomedicina hatékonyságára egy szubtípus-azonos
(génbanki azonosító: NC_001802), valamint C szubtípusú (AB254141) fertőzés esetén. *A
teljesen megegyező epitópok alapján számoltuk ki a keresztvédettség mértékét, melyet a
pLWXu1 összes epitópjának százalékában adtuk meg. B: A megegyező epitópok eloszlása a
proteomban. (Az epitóp szekvenciákat követő számok az epitóp fehérjén belüli pozícióját
jelölik. Az egyező epitópokat +, az eltérőeket – szimbólum jelöli.)
A
A fertőzés szubtípusa pLWXu1 epitópok száma B C
A*0250 MHCI allélre egyező/Σ epitóp egyező/Σ epitóp 104 93/107 13/92
Keresztvédettség* (%) 89 13

47
B
Fehérje pLWXu1 B szubtípus C szubtípus Fehérje pLWXu1 B szubtípus
16 15 / 16 5 / 20 9 8 / 9
GLLETSEGC + + FLYQSTHLP 21-29 -
ILGQLQPSL + - RLVNGSLAL 54-62 +
SLYNTVATL + + LIWDDLRSL 62-70 +
TLYCVHQRI + - RLRDLLLIV 77-85 +
TLNAWVKVV 151-159 + - LLLIVTRIV 81-89 +
ALSEGATPQ 174-182 + + LIVTRIVEL 83-91 +
DLNTMLNTV 183-191 + + LLGRRGWEA 91-99 +
AMQMLKETI 197-205 + - ALKYWWNLL 99-107 +
AEWDRVHPV 210-218 + - LLQYWSQEL 106-114 +
IILGLNKIV 266-274 + + 4 3 / 4
RMYSPTSIL 275-283 + - ALVVAIIIA +
WMTETLLVQ 316-324 + - VVAIIIAIV +
EMMTACQGV 345-353 + - IIAIVVWSI +
VLAEAMSQV 362-370 + - ALAEMGVEM -
AMSQVTNSA 366-374 + - 37 32 / 37
SLFGNDPSS 491-499 - - MLLGMLMIC 20-28 +
24 23 / 27 4 / 18 QMHEDIISL 103-111 +
ITLWQRPLV 59-67 + + KLTPLCVSL 121-129 +
LLTQIGCTL 145-153 + - TLTSCNTSV 192-200 +
ALVEICTEM 188-196 + - VITQACPKV 200-208 +
DVGDAYFSV 265-273 + + SLAEEEVVI 264-272 +
YQYMDDLYV 336-344 + + NLTDNVKTI 276-284 -
YMDDLYVGS 338-346 + + IIVQLNQSV 284-292 -
HLLRWGLTT 363-371 + - TLKQIASKL 341-349 +
ELHPDKWTV 388-396 + - QIINMWQEV 422-430 -
TVNDIQKLV 408-416 + - ALFLGFLGA 517-525 +
KLVGKLNWA 414-422 + - FLGAAGSTM 522-530 +
KLLRGTKAL 436-444 + - TMGAASMTL 529-537 +
LLRGTKALT 437-445 + - QLLSGIVQQ 543-551 +
ALTEVIPLT 443-451 + - LLSGIVQQQ 544-552 +
ELAENREIL 457-465 + - AIEAQQHLL 558-566 +
ILKEPVHGV 464-472 + - LLQLTVWGI 565-573 +
QLTEAVQKI 522-530 + - QLQARILAV 575-583 +
YQLEKEPIV 582-590 + - SLWNWFNIT 668-676 +
IVGAETFYV 589-597 + - NITNWLWYI 674-682 +
YLALQDSGL 638-646 + - WLWYIKIFI 678-686 -
ALQDSGLEV 640-648 + - YIKIFIMIV 681-689 -
IVTDSQYAL 650-658 + - FIMIVGGLV 685-693 +
LVNQIIEQL 672-680 + - MIVGGLVGL 687-695 +
YLAWVPAHK 687-695 + - GLRIVFAVL 694-702 +
HLEGKVILV 782-790 + - IVFAVLSIV 697-705 +
6 6 / 6 0 / 5 AVLSIVNRV 700-708 +
WQVMIVWQV 5-13 + - RLVNGSLAL 747-755 +
RIRTWKSLV 17-25 + - LIWDDLRSL 755-763 +
SLVKHHMYV 23-31 + - RLRDLLLIV 770-778 +
LVITTYWGL 64-72 + - LLLIVTRIV 774-782 +
ELADQLIHL 101-109 + - LIVTRIVEL 776-784 +
KIKPPLPSV 158-166 + - LLGRRGWEA 784-792 +
5 4 / 5 0 / 5 ALKYWWNLL 792-800 +
LLEELKNEA 22-30 + - LLQYWSQEL 799-807 +
WLHGLGQHI 38-46 + - SLLNATAIA 813-821 +
GLGQHIYET 41-49 + - AVAEGTDRV 821-829 +
AIIRILQQL 59-67 + - 3 2 / 3
QLLFTHFRI 66-74 - - LLRPMTYKA 75-83 -
GLEGLIHSQ 96-104 +
ILDLWIYHT 109-117 +
Nef
Vp
r
En
v (p
oli
pro
tein
)V
pu
Rev
Gag
(p
oli
pro
tein
)P
ol
(po
lip
rote
in)
Vif
Tat 0 0 0

48
A pLWXu1 pDNS alapú nanomedicina hatékonyságának elemzését nem B-szubtípusú
fertőzöttben ezután kiterjesztettük az összes prediktálható gyakori MHCI allélre (n=57),
valamint MHCII allélre is (n=63). A CD8+ T sejtes immunválasz elméleti hatékonysága
58%-osnak adódott szubtípus-azonos pácienst feltételezve, míg a C szubtípusú
fertőzöttben mindössze 26% volt a megegyező epitópok aránya a nanomedicinából
származó epitópokkal (4. táblázat). Modellünkbe bevettünk egy harmadik elterjedt
szubtípust is, az A szubtípust. Ennél szintén alacsony, 24%-os epitóp-egyezést találtunk
a nanomedicinából származó MHCI epitópokkal. A CD4+ T sejtes immunválasz
esetében szintén korlátozott mennyiségben találtunk egyező epitópokat: 47%-ot az
egyező szubtípusú fertőzés esetén, 15%-ot a C szubtípusú fertőzés esetén, ill. 14%-ot A
szubtípusú fertőzés esetén. Noha modellünkben csak egy-egy szubtípus-specifikus
szekvenciát elemeztünk, eredményeink megerősítik a fenti, egyetlen allél vizsgálatával
tett megállapításunkat, azaz a nanomedicina eltérő szubtípusú fertőzés esetén
lényegesen kisebb hatékonyságú lehet. A természetes felülfertőződéshez hasonlít ez a
megfigyelés, aminek során a HIV fertőzött beteg egy újabb HIV vírussal is fertőződik.
A felülfertőzés vírusszint emelkedést okoz, mert az alacsony kereszt-reaktivitás miatt az
addig hatékony immunválasz nem működik az új vírussal szemben [94,95].
4. táblázat. A B-szubtípus specifikus pLWXu1 nanomedicina hatékonyságának modellezése
szubtípus-azonos (génbanki azonosító: NC_001802), C szubtípusú (AB254141) és A szubtípusú
(AB253428) HIV-vel fertőzés esetén, T sejt epitóp predikcióval az összes prediktálható gyakori
MHCI és MHCII allélre. A keresztvédettséget a pLWXu1 összes epitópjának százalékában
adtuk meg.
A fertőzés szubtípusa
pLWXu1 B C A epitópok száma
57 MHCI allélre egyező/Σ epitóp egyező/Σ epitóp egyező/Σ epitóp
991 578/1164 257/1051 242/964
Keresztvédettség (%) 58 26 24
epitópok száma 63 MHCII allélre
egyező/Σ epitóp egyező/Σ epitóp egyező/Σ epitóp
2046 966/1636 297/2102 280/2261
Keresztvédettség (%) 47 15 14
Eredményeink azt sugallják, hogy a teljes HIV-fertőzött humán populáció számára
akkor tudunk hatékony terápiás vakcinát (nanomedicinát) nyújtani, ha a vírus genetikai
változatosságát, pontosabban a genetikai változatosságból származtatható immunválaszt

49
befolyásoló különbségeket is figyelembe véve szubtípus-optimalizált termékeket
fejlesztünk.
6.3. Szubtípus-optimalzált pDNS konstrukciók tervezése és klónozása
6.3.1. A vakcinatervezést segítő új alkalmazás: az “immunológiai fa”
Mivel a HIV genetikai változatossága (szubtípusok) a megfertőzött személy antigén-
feldolgozásért felelős genetikai változatosságával (MHC vagy HLA genotípus) együtt
határozza meg a kialakuló immunválasz minőségét, indokolt mindkét paramétert
figyelembe venni az immunogén tervezés során. Ezért a hagyományos HIV diverzitás
genetikai rokonsági viszonyainak vizsgálata helyett kidolgoztunk egy immunológiai
hasonlóságokon alapuló vírus csoportosítást. Módszerünk fehérje aminosav sorrend
helyett immunológiailag releváns T sejt epitópok alapján illeszti a szekvenciákat, és a
Neighbor joining filogenetikai fa készítő algoritmussal analóg módon kapcsolja össze
az epitópok alapján leghasonlóbb szekvenciákat, ill. szekvencia csoportokat [84].
Eredményül a hasonló immunválaszt kiváltani képes HIV szekvenciák csoportjait
kapjuk. Lássunk egy példát a genetikai hasonlóság-alapú fa és az immunológiai fa
információtartalma közti különbség bemutatására! Az elemzéshez egy jól izolált
epidémia, az ausztrál járvány génbankban található összes teljes HIV szekvenciáját
használtuk fel. A 15. ábrán a legkonzervatívabb HIV fehérje, a Pol poliprotein
aminosav alapú illesztése után készített filogenetikai fa, és a gyakori MHCI allélekre
prediktált CD4+ T sejt epitópok illesztése után készített immunológiai fa látható.
Külcsoportnak három, főként a Távol-Keleten elterjedt CRF_AG izolátumot
választottunk. A filogenetikai fán az elágazási pontokon a bootstrap értékek a
megbízhatóságot jelző mérőszámok (1000 ismétléssel generált fa hány százalékában
szerepel az adott elágazás). Az immunológiai fa elágazási pontjain a számok két
izolátum, ill. izolátum-csoport közötti korrekt egyezést mutató epitópok (megegyező
plusz egyetlen aminosavban különböző) számát adják meg. Tehát minél több a korrekt
egyezést mutató epitóp, annál nagyobb az immunológiai, pontosabban immunválaszbeli
hasonlóság két szekvencia között. Éppen ezért a közös ágon lévő szekvenciák jobb
immunológiai keresztreakciót adhatnak egymással, mint a csoporton kívüli
szekvenciákkal.

50
15. ábra. Példa a filogenetikai fa és az immunológiai fa információtartalmának különbségére, ausztrál HIV izolátumok Pol poliproteinjének vizsgálatával.
Filogenetikai fa: Clustal-W illesztés, Neighbor-Joining algoritmus, 1000 Bootstrap ismétlés. Immunológiai fa: prediktált MHCI epitópok sorozatának
(epitom) illesztése; a számok a vonatkozó két taxon közti korrekt egyezést mutató (megegyező plusz egy as-ban eltérő) epitópok mennyiségét jelentik.
Filogenetikai fa Immunológiai fa

51
A CRF_AG külcsoport filogenetikailag jól elkülönül a szubtípus-B csoporttól, és az
immunológiai fán is egy klasztert alkot, noha két B szubtípusú szekvencia
immunológiailag kilóg a csoportból. Közülük az egyik éppen a pLWXu1 által
expresszált Pol poliprotein, de ennek a nagyobb immunológiai távolságát az
magyarázhatja, hogy a biztonsági módosítások miatt rövidebb fehérjét hasonlítottunk a
többi teljes hosszúságú poliproteinhez. A fák topológiájában azonban több különbség is
látható, például a filogenetikai fán bekeretezett három szekvencia filogenetikai
távolsága lényegében elhanyagolható, míg az immunológiai fán a DQ676879
azonosítójú szekvencia távolabb helyezkedik el a filogenetikailag közelebbi DQ676877
és DQ676878-as szekvenciáktól, és több közös epitópot tartalmaz a DQ676880-as
szekvenciával, mint az előbbiekkel. Ehhez hasonló különbségeket a két fa között több
helyen is megfigyelhetünk még.
Noha az eltérések viszonylag kismértékűek, mégis rávilágítanak arra, hogy a
filogenetikai csoportok nem feltétlenül egyeznek a szekvenciák immunológiai
csoportosítással előálló klasztereivel. Ugyanis az immunológiai fa az immunválaszt
meghatározó epitópok szempontjából súlyozza a szekvenciák közti távolságokat,
minőségileg más eredményt adva. Az immunológiai-, ill. filogenetikai fák
összehasonlítását a többi HIV fehérjére is elvégeztük, és minden esetben az előző
példához hasonló szintű minőségi különbségeket találtunk köztük. Az immunológiai
fákat elkészítettük MHCI és MHCII allélekre is, amelyek minden fehérje esetén
kongruensek voltak: a módszer azonos immunológiai hasonlósági viszonyokat
eredményezett mind a CD8+ T sejtes immunválasz, mind a CD4+ T sejtes immunválasz
szempontjából.
Vizsgálataink alapján úgy véljük, hogy az immunológiai fa hatékony eszköz lehet az új
vakcinajelöltek (immunogének) fejlesztéséhez.
A továbbiakban az immunológiai fa alapján történő pDNS immunogén tervezést
mutatom be a szubtípus-optimalizált új nanomedicina hatóanyagok példáján.

52
6.3.2. Szubtípus-optimalizált termékjelöltek immunológiai szempontú tervezése
Az új nanomedicina termékjelöltek tervezése előtt kiválasztottuk a célzott járványokat.
A leendő termékcsalád első tagjai az alábbi szubtípusokba ill. cirkuláló rekombináns
formákhoz tartoznak: a legnagyobb járványokat okozó C szubtípus, a Kínában és a
Távol-Keleten gyakori BC rekombináns forma (CRF_07 ill. CRF_08), a Dél-
Amerikában jellemző BF rekombináns típusok (CRF_39 ill. CRF_40), és a Kelet-
Európában, főként Ukrajnában az utóbbi évtizedben gyorsan terjedő AB rekombináns
szubtípus (5. táblázat) [96].
5. táblázat. HIV-1 szubtípus-optimalizált pDNS konstrukciók, és a célzott járványok helye
valamint mérete.
Szubtípus / CRF Elterjedési terület Fertőzöttek száma* pDNS név C szubtípus Afrika (Szaharától délre) 22,5 millió pLWXu-C
CRF_BC (07 és 08) Kína 3,3 millió pLWXu-BC CRF_BF (12 és 39) D-Amerika 1,6 millió pLWXu-BF
CRF_AB (03) K-Európa 10 000 pLWXu-AB * A régió összes HIV fertőzöttjének száma, amin belül a megadott szubtípus, ill. CRF-ek dominálnak.
Ezt követően T sejtes immunválasz alapú immunológiai hasonlósági fákat készítettünk
teljes HIV proteomokra a kiválasztott járványok izolátumainak felhasználásával.
Célunk az volt, hogy az immunológiai fa csoportjaiba sorolható konstrukciókat
tervezzünk. Ugyanakkor kulcsfontosságú feladat volt a konstrukciók biztonsági
módosítása, a klinikai tesztelésre hatóságilag engedélyezett (pLWXu1) pDNS
biztonsági profiljának megőrzésével. Ezért a pLWXu1 pDNS Pol poliprotein génjét
(melynek 3’ végéhez közeli IN tartalmaz többszörös biztonsági módosításokat),
valamint a Nef gén 3’ végén csonkított kódoló darabját változtatás nélkül megőriztük a
szubtípus-optimalizált konstrukciókban. Vagyis az immunológiai optimalizálást a fent
említett HIV-specifikus szekvenciákon kívül eső régiókra kellett korlátoznunk. A
biztonsági és immunológiai optimalizálási szempontoknak megfelelő új pDNS
szekvenciákat a 16. ábrán látható térképek szerint terveztük meg. A CRF-ek esetében
természetesen az eredeti szubtípusok rekombinációs mintázatát is figyelembe vettük
[97].

53
16. ábra. A négy új pDNS HIV-specifikus részének térképe. A szubtípus-B eredetű
szekvenciák és a biztonsági módosítások a pLWXu1 pDNS-ből származnak.
Az immunológiai szekvencia optimalizálást a következőképpen valósítottuk meg: a
pLWXu-AB termékjelölt esetében az immunológiai fa CRF_03 AB klaszteréből
kiválasztott izolátum alapján terveztük a pDNS szekvenciát (16. ábra). Azon
konstrukcióknál, amelyeket frissen izolált HIV törzsek klónozásával készítettünk
(pLWXu-BC, pLWXu-BF és pLWXu-C), az immunológiai fát használtuk fel a
megfelelő epidemiológiai egységek meghatározásához, majd a pDNS-ek készítéséhez.
6.3.3. Az új pDNS-ek elkészítése és a termékjelöltek in silico jellemzése
Ezután elkészítettük az új pDNS termékjelölteket. A HIV-fertőzött betegek
vérszérumából RNS izolálás után RT-PCR-rel amplifikáltuk a vírus örökítőanyagát. A
pLWXu-AB konstrukcióhoz az AB rekombinánsra optimalizált DNS-t
szintetizáltattunk. A pLWXu1 “szülői” pDNS biztonsági módosításokat is tartalmazó
részeit változtatás nélkül megtartottuk, míg a szubtípus-specifikus DNS szakaszokat a
pLWXu1 DNS megfelelő szakaszai helyére illesztettük irányított klónozással. A
restrikciós enzimes gyors ellenőrzést követően az új pDNS-eket megszekvenáltattuk.
Az elkészített pDNS konstrukciók szekvenciáit felhasználva azt is ellenőriztük, hogy
tényleg az immunológiai fa megfelelő csoportjába tartoznak-e. A 17. ábrán az
pLWXu-C
pLWXu-BF
pLWXu-AB
LTR LTR
NefGag
LTR
Env
Tat
Rev
RT RNáz IN
Pol
Prot
LTR LTR
Nef
LTR
Env
Tat
Rev
GagRT RNáz IN
Pol
Prot
Env
Tat
Rev
Gag
LTR LTR
Nef
LTR
Prot RT RNáz IN
Pol
Szubtípus- B: ; C: ; F: ; AB: ; biztonsági módosítások:
pLWXu-BCLTR LTR
NefGag
LTR
Env
Tat
Rev
RT RNáz IN
Pol
Prot

54
immunológiai szekvencia-optimalizálás célfehérjéi közül a két legjelentősebb
poliprotein, a Gag és Env immunológiai fáit mutatjuk be, melyek a szubtípusok és
CRF-ek csoportspecifikus referencia szekvenciákat is tartalmazó válogatásai. A
szubtípus-optimalizált szekvenciák minden esetben a megfelelő (tervezett)
immunológiai klaszterbe sorolódtak.
17. ábra. Az immunológiai szekvencia-optimalizálás eredménye: az új pDNS immunogén
jelöltek a tervezett immunológiai klaszterekbe tartoznak mind a Gag poliprotein-, mind pedig az
Env poliprotein immunológiai fáján. (A termékjelölteket aláhúzással emeltük ki a fákon.)
A munkát az új termékjelöltek in silico jellemzésével folytattuk: a pDNS-ek által kódolt
fehérjék immunológiai potenciálját modelleztük különböző szubtípusú és CRF-ű
fertőzések esetén. Annak érdekében, hogy a modell reprezentatív legyen, minden
gyakori HLA allélre elvégeztük a T sejt epitóp predikciót, amit az IEDB adatbázis
tartalmaz (57 MHC I allél, 63 MHC II allél, 2010-es allélkészlet alapján). Ezáltal
populációs szintű immunválaszokat kaptunk, amik sosem egyetlen egyénre
Env poliprotein
A
B
C
pLWXu-BF
pLWXu-ABpLWXu1 (B)
pLWXu-C
BF
A
B
C
AB
pLWXu-C
pLWXu-AB
pLWXu1 (B)
BF pLWXu-BF
pLWXu-BC
pLWXu-BC
Gag poliprotein

55
specifikusak, viszont általánosan jellemezhetik a nanomedicina-jelöltek
használhatóságát különböző szubtípusokba tartozó járványokban.
Eredményeinket a 6. táblázatban rendszereztük. Vizsgálatunkat a következő
szempontok szerint végeztük: Az A, B, C és F szubtípusok esetén a Los Alamos HIV
Database teljes genom szekvenciáiból készítettünk egy-egy reprezentatív szekvencia
gyűjteményt, 1 szekvencia / év / izolálás országa / szubtípus kiválasztással. Ez
átlagosan 50 szekvenciát jelent szubtípusonként. A CRF-ek esetében minden teljes
genom szekvenciát felhasználtunk az elemzéshez. A csoportokon belül a Los Alamos
Database szerinti referencia szekvenciát választottuk ki összehasonlító szekvenciának
(génbanki azonosítók: AB253428, NC_001802, AF286224, AF077336, AF193277,
AY008716, EU735534). Ezután a megfelelő referencia szekvencia proteomja alapján
prediktált T sejt epitóp készlettel hasonlítottuk össze a csoporton belüli összes proteom
epitóp készletét, valamint az összes szubtípus-optimalizált pDNS által termelt fehérje
repertoár epitóp-készletét. A referencia szekvencia összes prediktált MHC I ill. MHC II
epitópjának számát adtuk meg 100%-nak. (Mivel ez a szám szekvenciánként eltérő
lehet, a táblázat utolsó sorában megadtuk, hogy az adott összehasonlító szekvencia
esetén 1% mennyi epitópot jelent.) Majd kiszámoltuk a csoportátlagokat (szubtípus ill.
CRF csoportátlag), amely az egyező epitópok átlagos mennyisége egy csoporton belül.
Ez az érték adja meg a szigorúan vett keresztvédettséget csoporton belül, noha a
kizárólag nagy, ill. közepes affinitású epitópok vizsgálata miatt ezek a számok
valószínűleg kissé alábecsült értékek. Ezenkívül a csoportátlag egy elvi maximumot, ill.
viszonyítási értéket is kijelöl, hiszen a HIV szekvenciákkal a legerősebb kereszt-
reaktivitást mutató új termékjelölteknél sem várunk a csoportátlagnál nagyobb
számokat. Az 6. táblázatból kiolvashatjuk tehát, hogy a vizsgált szubtípusú ill. CRF-ű
csoportba eső fertőzések esetén mely szubtípus-optimalizált pDNS konstrukciók
nyújtanák a legjobb védelmet. Egy kivétellel a konstrukciók az azonos szubtípusú/CRF-
ű fertőzéssel szemben adták a legmagasabb prediktált keresztvédettséget. A
csoportspecifikus átlaggal összemérhető eredményeket kaptunk, vagyis a legjobb
immunválaszt adó termékjelölt megközelíti a csoporton belüli átlagos
keresztvédettséget. A CRF_BC fertőzési modellünkben a vizsgált HIV törzs nem az
azonos CRF-ű termékjelölttel adta a legjobb immunválaszt, hanem a pLWXu1-gyel.
Vagyis immunológiailag a pLWXu1 hatóanyagú nanomedicina közelebb áll a CRF_BC
referencia izolátumhoz, mint a pLWXu_BC. Ez az eredmény megerősíti a szubtípus-

56
optimalizált termékekből álló termékcsalád hasznosságát, kiegészítve azzal a
megállapítással, hogy a jövőben a leghatékonyabb nanomedicina kiválasztásához a
fertőzést okozó vírustörzs szekvenciáját érdemes analizálni és megnézni, hogy
ténylegesen melyik termékjelölttel adja a legjobb keresztvédettséget. Tehát a páciens
fertőzést okozó HIV vírusának-, és a T sejt epitópjait meghatározó HLA genotípusának
ismeretében személyre szabott in silico analízist végezhetünk, és személyre szabott
ajánlást tehetünk a betegben legjobb hatékonyságot mutató immunogénre ill.
immunterápiára.
6. táblázat. Az új nanomedicina termékjelöltek immunológiai hatásosságának modellezése
különböző HIV fertőzések esetén. Az immunológiailag legközelebbi, vagyis a fertőzést okozó
HIV törzzsel legtöbb közös epitópot tartalmazó termékjelölteket szürkével emeltük ki.
DermaVir
szubtípus (fertőzésé) CRF (fertőzésé) A B C F AB BC BF Szubtípus/CRF összehasonlító szekvenciával
egyező MHC I/MHC II epitópok (%) pLWXu1 (B) 25/12 80/59 26/16 23/10 37/20 27/20 22/14 pLWXu-AB 28/14 69/46 27/12 24/12 43/30 23/15 22/11 pLWXu-BC 28/13 37/47 30/17 22/12 29/13 27/15 24/13 pLWXu-C 26/12 26/18 31/17 24/10 28/12 27/15 25/10
pLWXu-BF 26/11 48/35 28/16 29/16 33/14 29/14 30/18 Szubtípus ill. CRF csoportátlag 30/15 37/16 37/18 36/23 70/62 38/38 30/16
1% epitóp egyezés mennyi epitópot jelent (MHC I/MHC II)
10/22 7/16 10/21 10/21 9/21 10/21 10/22
6.3.4. Az új szubtípus-optimalizált termékjelöltek in vitro tesztelése
Az új termékjelöltek minőségének ellenőrzése során a klinikai DermaVirt használtuk
referencia anyagként. Az új pDNS konstrukciók mindegyike a pLWXu1 pDNS HIV-
specifikus részeinek cseréjével készült, az összes biztonsági módosítást változtatás
nélkül megőrizve. Ezt szekvenálással ellenőriztük minden esetben. Ezért az új
nanomedicina jelöltek esetén elegendő volt az antigén expressziót jellemezni a VLP+
termelés és a biológiai aktivitás in vitro mérésével.
6.3.4.1. VLP+ termelés az új termékjelöltek által
Az új termékjelöltek VLP+ termelésének igazolására a pLWXu1-nél ismertetett módon
a pDNS-ekkel transzfektált 293T sejteket transzmissziós elektronmikroszkóppal
vizsgáltunk (18.A ábra). Minden új termékjelölt esetében jól láthatók a vad típusú HIV-
re jellemző fenotípusú VLP+ részecskék, melyek az érett virionhoz hasonló mag (core)

57
és burok (envelop) struktúrákat tartalmaznak, és a sejten kívüli térbe, valamint sejten
belüli vakuolumokba fűződnek le.
A VLP+ összetett felépítését tisztított VLP+ felhasználásával végzett western blottal is
megerősítettük (18.B ábra). Ugyanezen módszerrel igazoltuk a pDNS-ekben kódolt
fehérje repertoár VLP+-alkotó részének expresszióját. A pLWXu-BF és pLWXu-BC
esetén a p17-nek megfelelő sáv nem látható, amit a felhasznált VLP+ minta alacsonyabb
koncentrációja okozhat. (A p17 expresszióját egyértelműen igazolja a VLP+
termelődése, hiszen a VLP+ core részét a p17 fehérjék határolják, lásd 18.A ábra.)

58
18. ábra. Az új pDNS-ekről expresszált fehérjékből képződő komplex VLP+ részecskék. A: a
VLP+ “érésének” különböző fázisai az új pDNS-ek esetében (intracelluláris- és extracelluláris
lefűződés, éretlen virion-szerű fázis, érett virion-szerű fázis). A méretskála minden esetben 100
nm-nek felel meg. A fixált minták elektronmikroszkópos felvételeit Dr. Kovács Attila Lajos
készítette. B: Western blot pLWXu1-gyel transzfektált 293T sejtek felülúszójából, HIV+
szérum-keverék elsődleges ellenanyagként való használatával. Pozitív kontroll: pLWXu1
“szülői” pDNS. a-gp120 (Env-SU), b-p66 (reverz transzkriptáz plusz RNáz), c-p55 (Gag
poliprotein, benne p17, p24 és p7), d-p51 (Pol-RT), e-gp41 (Env-TM), f-p24 (Gag-CA), g-p17
(Gag-MA), h-p7 (Gag-NC).
A
B
pLWXu-C
pLWXu-BF
pLWXu-AB pLWXu-BC
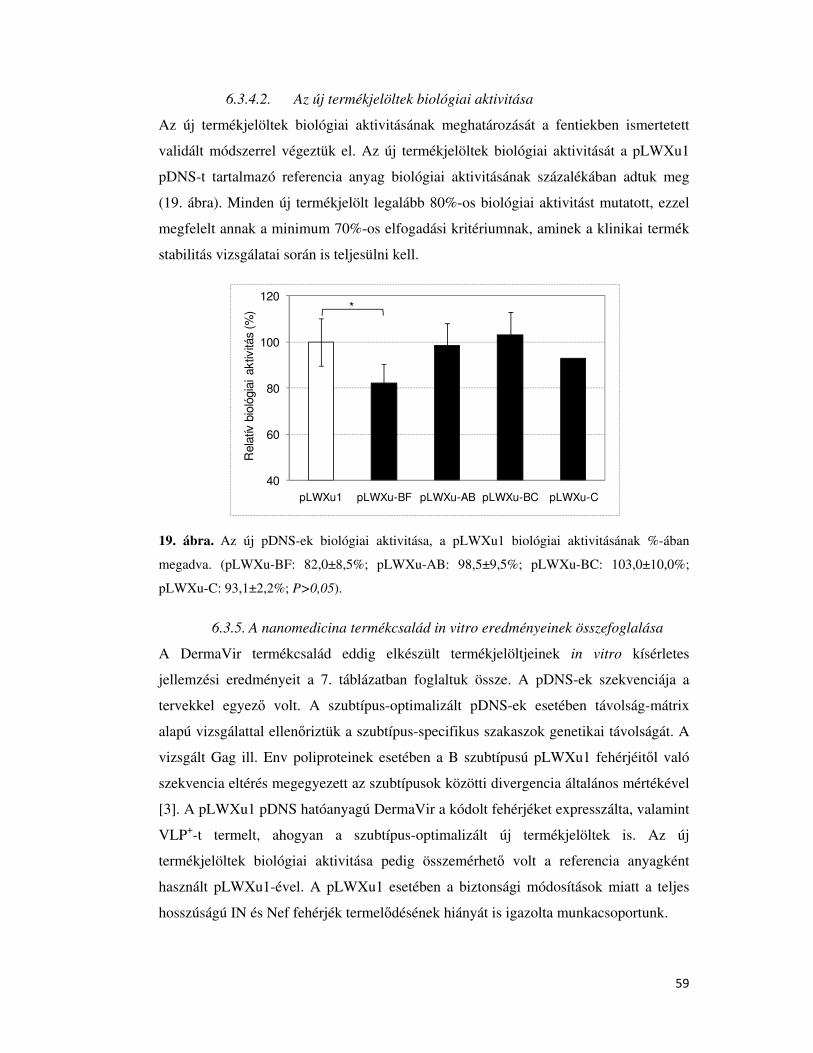
59
6.3.4.2. Az új termékjelöltek biológiai aktivitása
Az új termékjelöltek biológiai aktivitásának meghatározását a fentiekben ismertetett
validált módszerrel végeztük el. Az új termékjelöltek biológiai aktivitását a pLWXu1
pDNS-t tartalmazó referencia anyag biológiai aktivitásának százalékában adtuk meg
(19. ábra). Minden új termékjelölt legalább 80%-os biológiai aktivitást mutatott, ezzel
megfelelt annak a minimum 70%-os elfogadási kritériumnak, aminek a klinikai termék
stabilitás vizsgálatai során is teljesülni kell.
19. ábra. Az új pDNS-ek biológiai aktivitása, a pLWXu1 biológiai aktivitásának %-ában
megadva. (pLWXu-BF: 82,0±8,5%; pLWXu-AB: 98,5±9,5%; pLWXu-BC: 103,0±10,0%;
pLWXu-C: 93,1±2,2%; P>0,05).
6.3.5. A nanomedicina termékcsalád in vitro eredményeinek összefoglalása
A DermaVir termékcsalád eddig elkészült termékjelöltjeinek in vitro kísérletes
jellemzési eredményeit a 7. táblázatban foglaltuk össze. A pDNS-ek szekvenciája a
tervekkel egyező volt. A szubtípus-optimalizált pDNS-ek esetében távolság-mátrix
alapú vizsgálattal ellenőriztük a szubtípus-specifikus szakaszok genetikai távolságát. A
vizsgált Gag ill. Env poliproteinek esetében a B szubtípusú pLWXu1 fehérjéitől való
szekvencia eltérés megegyezett az szubtípusok közötti divergencia általános mértékével
[3]. A pLWXu1 pDNS hatóanyagú DermaVir a kódolt fehérjéket expresszálta, valamint
VLP+-t termelt, ahogyan a szubtípus-optimalizált új termékjelöltek is. Az új
termékjelöltek biológiai aktivitása pedig összemérhető volt a referencia anyagként
használt pLWXu1-ével. A pLWXu1 esetében a biztonsági módosítások miatt a teljes
hosszúságú IN és Nef fehérjék termelődésének hiányát is igazolta munkacsoportunk.
40
60
80
100
120
pLWXu1 pLWXu-BF pLWXu-AB pLWXu-BC pLWXu-C
Rel
atív
bio
lógi
ai a
ktiv
itás
(%) *

60
7. táblázat. A nanomedicina termékcsalád kísérletes (in vitro) jellemzésének összefoglalása.
Jellemzett tulajdonság
(vizsgálati módszer)pLWXu1 (B) pLWXu-AB pLWXu-BC pLWXu-C pLWXu-BF
Azonosság meghatározás és szekvencia alapú fehérje
különbségek(Szekvenálás, távolság mtx.)
pDNS tervvel egyezőpDNS tervvel
egyezőGag: 15%
pDNS tervvel egyező
Env: 31%
pDNS tervvel egyező
Gag: 20 %Env: 31%
DNS tervvel egyező
Env: 26%
VLP+ termelés
(TEM)VLP+ kimutatható VLP+ kimutatható VLP+ kimutatható VLP+ kimutatható VLP+ kimutatható
Antigén termelés (Western Blot)
gp120 (Env-SU), p66 (RT+RNáz), p55 (Gag poliprot.), p51
(Pol-RT), gp41 (Env-TM), p24 (Gag-CA), p17 (Gag-MA)
kimutatható
A várt fehérjék kimutathatók
A várt fehérjék kimutathatók, p17
csak Gagpoliproteinalkotóként
A várt fehérjék kimutathatók
A várt fehérjék kimutathatók, p17
csak Gagpoliproteinalkotóként
Relatív biológiai aktivitás
(transzfekció, p24 ELISA)100% 99% 103% 93% 82%
Antigén termelés(Intracelluláris festés, FACS)
p24, p55, gp41, gp120, Rev, Tat
kimutathatóNem mértük Nem mértük Nem mértük Nem mértük
Biztonsági módosítások(Immunprecipitáció)
IN, Nef módosított fehérjék nem mutathatók ki
Nem mértük Nem mértük Nem mértük Nem mértük

61
7. Eredmények megvitatása
Az immunológiai szempontból szubtípus-optimalizált HIV immunterápia egy új
lehetőség a HIV fertőzöttek kezelésére úgy, hogy a kezelt beteg vírusának és az
immunválasz minőségét befolyásoló HLA típusának figyelembe vételével a neki
leginkább megfelelő termék választható ki a szubtípus-optimalizált termékcsaládból. A
termékcsalád első tagjait a DermaVir HIV Tapasz fázis II klinikai kísérleti fejlesztési
fázisban járó nanomedicina alapján terveztük és készítettük. Ehhez felhasználtuk az
eddig kifejlesztett technológiákat, hiszen az új termékjelöltekben csak az antigéneket
kódoló pDNS-ek különböznek. A termékcsalád kifejlesztésének szükségességét
támogató in silico immunológiai potenciál predikciók és kereszt-védettségi
modellezések, valamint az immunogén tervezésére kifejlesztett módszereink egészítik
ki az eddigi technológiákat. Miért, és hogyan érhetünk el eredményt ezzel a
termékcsaláddal? A klinikai DermaVir termékjelölt in vitro jellemzésének új
eredményei, a nanomedicina termékcsalád in silico és in vitro tesztelésének eddigi
adatai, valamint a HIV/AIDS elleni vakcinafejlesztés tanulságai alapján tekintsük át a
lehetséges válaszokat!
A kutatók a természetes immunológiai kontrollal rendelkező HIV fertőzöttek (“elit
kontrollerek”) tanulmányozásával próbálják megtalálni a legjobb antigént, egyelőre
sikertelenül [98,99,100]. A HIV fertőzöttek szervezetében termelődnek ugyan vírus
neutralizáló ellenanyagok, de ezeket eddig nem sikerült eredményesen felhasználni
vakcina hatóanyagként. A legújabb vizsgálatok mutatnak rá az eredménytelenség
okaira. A neutralizáló ellenanyag molekulák a HIV felszínén lévő Env glikoproteinek
trimereihez kapcsolódnak. Úgy tűnik, hogy a hatékony neutralizálás feltétele a HIV
esetében is a pl. influenza vírusnál sikeres antigén keresztkötés az ellenanyag által.
Csakhogy a HIV felszínén kevés keresztköthető fehérje trimer található, egymástól
távol. Ezért itt az ún. polireaktív ellenanyagok lehetnek hatékonyak, amik a trimeren
belüli keresztkötést teszik lehetővé, ezzel stabilizálva az ellenanyag-antigén kötést
[101]. Sajnos azonban még az ilyen polireaktív ellenanyagok is csak korlátosan
működnek [102]. A HIV diverzitás, a vírusra ható immunszelekció és a vírus többféle
védőmechanizmusa gördít leküzdhetetlennek tűnő akadályokat a megelőző vakcina
kifejlesztése elé. A terápiás vakcinafejlesztés esetében is több antigén használata, és
ezzel a minél szélesebb skálájú immunválasz kiváltása látszik célravezetőnek az egyedi
antigének használata helyett [103].

62
A HIV vakcinafejlesztés tehát a sok antigénes hatóanyagok fejlesztése irányába halad
[104]. Ennek a feladatnak a megoldásában egyre inkább előtérbe kerül a
bioinformatikai modellezés, mint a tervezés idő- és költséghatékonyságát növelő
eszköz.
Az in silico T sejt epitóp predikciót mi is felhasználtuk a doktori munka több lépéséhez:
(1) a B-szubtípusú DermaVir pDNS-e (pLWXu1) által expresszált antigének
immunológiai potenciáljának modellezéséhez; (2) a DermaVir hatékonyságának
modellezéséhez nem-B szubtípusú fertőzöttek esetében a keresztvédettség
vizsgálatával; (3) szubtípus-optimalizált DermaVir termékcsalád pDNS-einek
immunológiai alapú tervezéséhez; (4) az új termékjelöltek immunológiai potenciáljának
(kereszt-reaktivitásának) modellezéséhez szubtípus-azonos ill. eltérő szubtípusú
fertőzések esetén.
A bioinformatikai modellezési munkához először kiválasztottunk egy in silico T sejt
epitóp predikciós módszert mind a CD8+ T sejtekre specifikus MHCI-, mind pedig a
CD8+ T sejtekre specifikus epitópok prediktálására. A módszerek megbízhatóságát
kísérletesen igazolt epitópok adatbázisának segítségével határoztuk meg: a nagy ill.
közepes affinitású T sejt epitópok prediktálásának megbízhatósága 97,1%-ot
eredményezett MHCI epitópok esetén, míg 97,7%-ot MHCII epitópok predikciója
esetén, vagyis a kísérletesen igazolt epitópok 97,1%-át, ill. 97,7%-át teljes- ill. korrekt
egyezéssel (1 as eltérést megengedve) tudtuk prediktálni (2.táblázat). Ezzel az eljárással
modelleztük azután a pLWXu1 pDNS-ben kódolt antigén készlet, vagyis a
nanomedicina várható immunológiai hatását. Több, mint 100 eltérő HLA allélre
prediktáltuk a nagy-, és közepes affinitású T sejt epitópokat. Eredményeink alapján a
nanomedicina jelölt által expresszált 15 fehérjéből álló antigén repertoárja széles
spektrumú CD4+ és CD8+ specifikus immunválasz kiváltására képes. Ez az
irodalomban eddig közölt HIV vakcinajelöltek közül a legszélesebb HIV epitóp
repertoárt jelenti [105,106].
Az abszolút immunológiai potenciál (AIP) és relatív immunológiai potenciál (RIP)
analízisünk megerősítette mind a struktúrfehérjék, mind pedig a szabályozó fehérjék
kódolásának fontosságát a T sejtes immunválaszt célzó nanomedicina pDNS-ében. A
pLWXu1 pDNS által kódolt fehérje repertoár AIP értéke mennyiségileg prediktálja a
hatóanyag által indukálható CD4+ és CD8+ specifikus immunválaszt. Ez alapján
minden HIV fehérje szerepet játszik a CD4+ ill. CD8+ T sejtek különböző mértékű

63
aktiválásában, de csak az Env-SU, Pol-RT, Gag-CA Vif és Rev fehérjék aktiválják
hatékonyan mindkettőt. A Tat és Gag-NC a CD4+ T sejt válasz kiváltására nagyon
magas, míg a CD8+ T sejtes válasz kiváltására szerény potenciált mutatott. Klinikai
kutatások eredményei is megerősítették, hogy a széles HIV epitóp repertoár felismerése
fordítottan korrelál a vírus terheltséggel, noha a szerepet játszó fehérjék immunválasz
szempontú sorrendje természetesen függ az egyén HLA hátterétől és érdekes módon a
betegség stádiumától is [107,108]. A széles epitóp repertoárnak köszönhetően a T sejtek
képesek felismerni és elpusztítani a HIV-vel fertőzött sejteket (a látensen fertőzöttek
kivételével), de lehet, hogy a korai fehérjékre specifikus T sejtek (pl. Vif és Rev)
szerepe is meghatározó a HIV replikáció visszaszorításában.
A RIP mérőszámmal a fehérjén belüli nagy epitópok relatív sűrűségét adhatjuk meg, a
fehérje elméleti (9, ill. 15 as hosszú) epitóp mennyiséghez képest. (Az elméleti epitóp
mennyiség a protein hossz mínusz 8 az MHCI RIP predikció esetén, ill. protein hossz
mínusz 14 az MHCII RIP esetén.) A RIP hasonlít egy már korábban bevezetett
mérőszámhoz: a SIR (“size of immune repertoire”, immunológiai repertoár mérete) a
fehérjében prediktált epitóp mennyiséget hasonlítja egy ugyanolyan mennyiségű
elméleti nonamert tartalmazó, és hasonló aminosav eloszlású fehérjében várt epitóp
mennyiséghez [109]. RIP analízisünk alapján a Gag és Env proteinek méltán
szerepelnek sok fejlesztés alatt álló HIV vakcina antigénjeként, hiszen magas MHCI-,
és MHCII RIP értékkel rendelkeznek. Szintén magas MHCI-RIP-et kaptunk a Rev és
Vpu fehérjék esetében, és a Tat fehérje MHCII-RIP értéke is kiemelkedő volt, jelezvén
a korai szabályozó fehérjék magas immunológiai potenciálját, ill. támogatva ezek HIV
vakcina antigénként való felhasználását.
Immunogén-tervezési koncepciónk némiképp hasonlít a mozaik vakcinatervezési
módszerhez, melynek lényege, hogy az antigént mesterségesen dúsítják fel epitóp-
gazdag szakaszokkal, in silico evolúciót modellezve [110]. A felhasznált valódi
kórokozó szekvenciák egy csoportját in silico “rekombináltatják”, majd minden ciklus
után meghatározott paraméterek figyelembevételével kiválogatják a legtöbb epitópot
tartalmazó (átfedő epitópokat is tartalmazó) szekvenciákat, és az eredeti szekvencia
csoport epitóp-szegény szekvenciáit ezekre cserélik. Bizonyos számú ciklus után
végeredményül egy, a valóságban nem létező mozaik szekvenciát kapnak, amit aztán
vakcina antigénként tesztelnek.

64
A DermaVir nanomedicinában az epitóp repertoár kiterjesztésének a sok fehérje
kódolásán kívül egy másik módja is van: az egyszerű antigén expresszión kívül
komplex vírus-szerű részecskék (VLP+) is termelődnek. A DermaVir kísérletes
jellemzése során igazoltuk, hogy a pDNS-ben kódolt közel teljes proteomból komplex
VLP+ részecskék is képződnek. Tehát természetes konformációban expresszálja a HIV
antigéneket, ami a természetes HIV fertőzéssel egyező antigén feldolgozást és
immunválasz kiváltását eredményezheti. Megjegyzendő, hogy a gyógyszerjelölt ezen
potenciálját akár megelőző vakcina kifejlesztéséhez is fel lehetne használni, hiszen a
natív térszerkezetű felszíni fehérjék nagy hatékonyságú, polireaktív neutralizáló
ellenanyagok termelését is kiválthatnák [101]. Másrészt a VLP+ termelés a CD4+ helper
T sejtek indukcióját is elősegíti, ami pedig a CD8+ T sejtek érését (priming-ot),
valamint a hatékony immunválaszban kulcsszerepű memóriasejtek termelődését
támogatja [45,46,47].
Kísérletesen kimutattuk a DermaVir által expresszált antigén készlet termelődését,
valamint igazoltuk, hogy ezek a HIV fehérjék a vad típusú HIV-hez hasonló struktúrájú
komplex VLP+ részecskékké állnak össze. A vírus érési fázisainak megfelelő
struktúrájú VLP+-k kimutathatók, azonban ezek biztonságosak, és nem fertőzőképesek
(utóbbi eredmény nem része a dolgozatnak [92]). A VLP+ intracelluláris-, és
extracelluláris térbe egyaránt ürül, akárcsak a vad HIV virionja [111].
A DermaVir pDNS-éről termelődő antigének (fehérjék és VLP+) az antigénbemutató
sejt általi feldolgozását követően kerülhetnek MHCI és MHCII molekulákra. A 20.
ábrán foglaltuk össze a nanomedicina pDNS immunogénjének lehetséges antigén
expressziós és antigén prezentációs útvonalait [92]. A pLWXu1 pDNS-ről
expresszálódó antigén készlet mindkét antigén feldolgozási útvonalon képes
immunválaszt kiváltani: az endogén antigén-bemutatási út a pDNS-ről az
antigénbemutató sejtben termelődő fehérjék feldolgozásával történik (20. ábra, “a”
útvonal). Az expresszált fehérjékből spontán összeszerelődő VLP+ részecskék sejten
belüli vezikulumokba és a sejten kívüli térbe is juthatnak. Mindkét esetben az exogén
antigének feldolgozási útvonalán halad a folyamat, ami a VLP+-ből származó MHCII
epitópok bemutatásával HIV-specifikus CD4+ helper T sejtes immunválasz kiváltását
eredményezi (20. ábra “b” és “c”).

65
20. ábra. HIV-specifikus T sejtes immunválasz kiváltásának lehetséges útvonalai a DermaVir
pDNS hatóanyagáról termelődő antigénekkel. A nanomedicinát felvevő antigénbemutató
sejtben a pDNS-ről HIV fehérjék termelődnek: a) az intracelluláris fehérjék feldolgozásának
útvonalán halad tovább a folyamat, ami MHC I molekulákon való epitópok bemutatását, ezzel
HIV-specifikus CD8+ T sejtek indukcióját eredményezi. b) Az expresszált fehérjékből VLP+
részecskék képződnek, melyek sejten belüli vezikulumokba szekretálódnak, és az antigén
feldolgozás során képződő epitópok MHC II molekulákon bemutatva HIV-specifikus CD4+ T
sejtek képződését indukálják. c) VLP+ részecskék a sejten kívüli térbe is szekretálódhatnak,
amik kis eséllyel egy újabb antigénbemutató sejtben az antigén bemutatás CD4+ T sejt választ
eredményező útján haladhatnak. Mivel a VLP+ nem képes replikálódni, a folyamat az antigén
feldolgozással véget ér.
Az összetett VLP-t expresszáló HIV vakcina ötlete egyébként nem egyedülálló, bár a
szintén klinikai tesztelés alatt álló másik immunogén DNS csak a HIV fehérjék kb.
50%-át expresszálja, és az éretlen vírushoz hasonló morfológiájú VLP-t, valamint
fehérje aggregátumokat termel [106,112].
A vakcina hatékonyságnak van még egy fontos paramétere a sokféle antigénen kívül: a
DermaVir-ből származó epitópoknak a kezelt beteg HIV epitópjaival való egyezés
mértéke, vagyis a kereszt-reaktivitás. Hiába generál ugyanis a DermaVir számos HIV
epitópra specifikus CD4+ és CD8+ T sejteket, ha azok nem egyeznek a fertőzött
személy vírusának epitópjaival. Mert nyilvánvalóan csak azok a T sejtek tudják
elpusztítani a HIV-vel fertőzött sejteket, amik felismerik őket. Hogy képet kapjunk az
epitóp egyezések mértékéről, in silico modelleztük a B-szubtípusú HIV fehérjéket
pDNS
12
3
(…)
CD8+ T sejt válasz
Nem reprodukálódik:•Reverz transzkripció gátolt•Replikáció gátolt•Integráció gátolt
a
c
CD4+ T sejt válasz
b
MHCIMHCII
MHCII
T

66
kódoló DermaVir epitópjainak kereszt-reakcióját egy szubtípus-azonos, egy C
szubtípusú, és egy A szubtípusú izolátum esetén. A szubtípus-azonos izolátum esetén a
CD8+ T sejt specifikus epitópok 58%-a és a CD4+ T sejt specifikus epitópok 47%-a
egyezett meg teljesen a DermaVir epitópjaival. (Ez persze a legszigorúbb egyezés
mellett adódó érték, a kereszt-reaktivitás megenged néhány aminosav eltérést, ezért a
valódi keresztvédettséget valamivel erősebbnek várhatjuk.) Ehhez képest viszont a C-,
ill. A szubtípusú fertőzés esetén csak az MHCI epitópok 26%-a ill. 24%-a, míg az
MHCII epitópok 15%-a ill. 14%-a egyezett a DermaVir epitópjaival. A kereszt-
reaktivitást megvizsgáltuk a többi szubtípusra is, valamint az egyes szubtípusok több
izolátumára (az adatokat nem mutatjuk), és hasonló eredményt kaptunk: a nem-B
szubtípusba tartozó izolátumokkal kevesebb keresztvédettséget biztosító egyező epitóp
mutatható ki, mint a szubtípus-azonos izolátumokkal. Ezért indokoltnak láttuk a
szubtípus-optimalizált termékcsalád kifejlesztését, hogy egyszer majd minden beteg
optimális immunválaszt kiváltó DermaVir-t kaphasson.
Az immunválasz szempontú termék optimalizáláshoz egy, a filogenetikai rokonsági
viszonyokat tükröző fával analóg eszközt fejlesztettünk ki: az immunológiai fát, ami a
hasonló epitóp-összetételű szekvenciákat jeleníti meg közös ágon. Információtartalma
minőségileg tér el a filogenetikai fáétól, mert az immunválasz szempontjából legjobban
hasonlító szekvenciákat helyezi egymáshoz legközelebb, ami nem feltétlenül egyezik a
szekvencia alapú távolsággal. Gondoljunk csak az MHCI epitópok képződésének
korábban említett statisztikájára: az antigén feldolgozás során csak minden 7. peptid tud
a TAP molekulán bejutni az endoplazmatikus retikulumba, ahol az MHCI molekulák
csak minden 200. epitópot kötnek meg [43,44]. Két szekvencia közti különbségek tehát
nem jelennek meg az immunológiai fán, ha olyan szakaszon vannak, amikre nem
prediktálható epitóp. Ugyanakkor többszörös súllyal számíthat akár egyetlen bázispár
eltérés, amennyiben az érintett szakaszra több, átfedő epitóp prediktálható.
Az immunológiai fa csoportjai alapján kiválasztottunk négy szubtípust, ill. rekombináns
formát, majd ezekre optimalizált pDNS konstrukciókat terveztünk. Tulajdonképpen
nem is szubtípust/rekombinánst választottunk, hanem “immunológiai
szubtípust/rekombinánst”, de megfelelő elnevezés híján egyelőre a szekvencia-alapú
csoportosítás nevezéktanát használjuk, némi képzavart kockáztatva ezzel. A pDNS-eket
elkészítettük, és az új nanomedicina termékjelölteket in silico és in vitro teszteknek
vetettük alá. Az in silico keresztvédettségi vizsgálataink alapján megállapítottuk, hogy

67
az új termékjelöltek a szubtípusuknak/rekombináns csoportjuknak megfelelő
izolátumok esetében eredményezték a legjobb keresztvédettséget, kivéve egy esetet.
Ebben az immunológiai távolságok eltértek a szubtípus-alapú csoportosítástól, mert a
CRF-BC rekombinánssal való fertőzés modelljében nem a pLWXu-BC termékjelölttel
adódott a legnagyobb keresztvédettség, hanem MHCI esetén a pLWXu-BF
termékjelölttel, MHCII esetén pedig a B szubtípus-specifikus pLWXu1-gyel. Ez a példa
is megerősíti az immunológiai alapú szekvencia csoportosítás létjogosultságát.
A kereszt-reaktivitás vizsgálatát akár személyre szabottan is megtehetjük
módszerünkkel: a páciens HIV szekvenciája és HLA genotípusa ismeretében a
termékjelöltek közül kiválasztható az adott páciensnél legmagasabb keresztvédettséget
biztosító nanomedicina. Ugyanígy prediktálhatjuk egy klinikai kísérletben résztvevő
páciensek HLA genotípusa és vírus szekvenciája ismeretében a várható T sejtes
immunválaszt, segítséget nyújtva az immunogenitási eredmények kiértékeléséhez, ill.
értelmezéséhez.
Az immunológiailag optimalizált szubtípus-specifikus termék-skála tovább bővíthető,
elméletben akár a személyre szabott konstrukciókig. Valószínű azonban, hogy az új
termékcsalád hatékonyságát vizsgáló majdani klinikai mérések alapján nagyobb
immunológiai csoportokat lehet megállapítani, és elég lesz ezekre a csoportokra egy-
egy termékjelöltet készíteni.
A HIV diverzitása miatti kereszt-reaktivitás más vakcinajelöltek esetén is elvárt
tulajdonság, amit többféle megközelítéssel terveznek elérni a fejlesztők, például több
szubtípusra specifikus konszenzus antigének alkalmazásával [113]. A mozaik vakcinák
fejlesztésekor pedig konzervatív, ezáltal kereszt-reaktív epitópok kiválasztására
törekszenek. Tegyük fel, hogy több mozaik fehérjét egyszerre használunk
immunizálásra, pl. a Barouch és munkatársai által in vivo állatmodellben sikeresen
tesztelt Gag, Pol és Env mozaik antigéneket [114]. Egy B szubtípusú fertőzési
modellben (szekvencia génbanki azonosítója DQ487189, a beteg meghatározott HLA
allélje, amit mi is vizsgáltunk: A*0201) 114 MHCI epitóp volt prediktálható a teljes
HIV proteomra. Ha az előbbi mozaik antigéneket használnánk terápiás vakcinaként,
mindössze 5 azonos (kereszt-reaktív) epitóp (4%) adódna. Ehhez képest a DermaVir
(B) esetében 47 egyező epitóp (41%) indukálhatná a beteg HIV vírusát felismerő CD8+
T sejtek termelődését. Egy másik példában valamivel jobb arányt találtunk (szekvencia
génbanki azonosítója DQ487189, a beteg HLA allélje: B*35:01): a beteg 84 epitópjából

68
25-öt fedett le a mozaik antigén készlet (30%), míg 40-et a DermaVir antigén készlete
(48%). Ehhez képest egy C szubtípusú fertőzési modellben (szekvencia génbanki
azonosítója AY463231, a beteg HLA allélje: A*68:02) a beteg 95 epitópjával a mozaik
antigének kereszt-reaktivitása 26% volt (25 egyező epitóp), míg a DermaVir antigén
készletével 37% (35 egyező epitóp). Ha a még jobb összehasonlíthatóság érdekében pl.
csak a Gag antigént vizsgáljuk mindkét vakcinából, akkor a B szubtípusú fertőzési
modellekben a vírus Gag fehérjéjére prediktált 15 epitópból csak egy egyezik meg a
mozaik Gag epitópjaival, és 8 a DermaVir-ével, valamint a 16 vírus epitópból 6 a
mozaik Gag epitópjaival és 10 a DermaVir-ével. A C szubtípusú vírus 12 Gag epitópja
közül egyaránt 7-7 egyező epitópot eredményezett a mozaik-Gag és a DermaVir-Gag.
Ez persze messze nem reprezentatív vizsgálat, de ezekben a véletlenszerűen kiválasztott
példákban nem látszott hatékonyabbnak a szubtípustól függetlenül kereszt-reaktívnak
tervezett mozaik antigén, mint a B szubtípusra specifikus DermaVir. A fertőzés
vírusával azonos szubtípusú DermaVir viszont jobbnak adódott, mint a kereszt-reaktív
mozaik antigének, megerősítve a szubtípus-optimalizált termékcsaládtól várt
optimálisabb hatékonyságot.
Az elkészített új nanomedicina termékjelöltek in vitro jellemzését is elvégeztük, és
minőségüket a klinikai DermaVir-éhez hasonlítottuk. Azt találtuk, hogy a
termékjelöltek képesek a kódolt antigén repertoár expressziójára, valamint komplex
VLP+ részecskék expressziójára is, és biológiai aktivitásuk hasonlóan erős, mint a
referencia anyagként használt klinikai tesztelés alatt álló DermaVir-é. Az új
termékjelöltek biztonsági módosításait a szülői pDNS-ként használt pLWXu1-ből
vettük át változtatás nélkül, ezért az új nanomedicina jelölteknél ezen változtatások
eredményét már nem kellett ellenőrizni.
A szubtípus-optimalizált HIV terápiás nanomedicina termékcsalád első tagjai a pDNS
hatóanyagok megfelelő mennyiségben és minőségben történő előállítása után
lényegében készen állnak a klinikai tesztelésre. A termékcsalád hatékonyságára
vonatkozó legfontosabb eredményeket a klinikai mérések szolgáltatják majd.

69
8. Összefoglalás
Kísérletesen és in silico módszerekkel jellemeztük a jelenleg klinikai kísérleti fázisban
lévő HIV-1 B-szubtípus specifikus DermaVir terápiás nanomedicina termékjelölt pDNS
alapú immunogénjét. Továbbá új HIV-1 szubtípus-optimalizált immunterápiás
nanomedicina termékjelölteket terveztünk a vírus diverzitás és a betegek HLA
hátterének figyelembe vételével. A termékjelölteket elkészítettünk és teszteltünk.
Munkánk során bioinformatikai-, molekuláris- és sejtbiológiai módszereket
használtunk, ill. fejlesztettünk.
Kimutattuk, hogy a DermaVir képes a kódolt HIV antigének (tizenhárom teljes és két
részleges antigén) expressziójára humán sejtekben, valamint igazoltuk, hogy ezek az
antigének a vad típusú HIV-vel immunológiai szempontból megegyező komplex vírus-
szerű részecskékké (VLP+) állnak össze. A DermaVir immunogénje emellett
biztonságos, mert nem képes integrálódni, replikálódni, ill. fertőző virionokat termelni.
Az antigén expresszió és a VLP+ termelés kimutatására vizsgálati módszereket
fejlesztettünk. Az európai és amerikai gyógyszerengedélyezési hatóságok
követelményeinek megfelelő validált biológiai aktivitást mérő módszert fejlesztettünk a
termékjelölt antigén termelési potenciáljának mérésére. Eredményeink segítettek
felvázolni a DermaVir hatásmechanizmusának részleteit a DermaVir-ből származó
antigének MHCI és MHCII molekulákon történő antigénbemutatási folyamatában.
A B szubtípusra specifikus DermaVir in silico vizsgálati eredményei, ill. a klinikai
mérési eredmények alapján indokoltnak láttuk szubtípus-optimalizált terápiás vakcina
termékjelöltek kifejlesztését. Prediktáltuk a DermaVir-B T sejt specifikus immunológiai
potenciálját, és modelleztük a keresztvédő képességét nem-B szubtípusú fertőzések
esetén. Azt találtuk, hogy a szubtípusok között csak mérsékelt keresztvédettség
prediktálható a T sejtes immunválaszban, ami a nanomedicina terápiás
hatékonyságának csökkenését vonhatja maga után bizonyos betegpopulációkban. Ezen
eredmények alapján kifejlesztettünk egy in silico immunogén tervezési eszközt. Az
“immunológiai fa” a hasonló epitóp összetételű vírus szekvenciák csoportjaiból épül
fel, amik minőségileg hasonló immunválaszt képesek kiváltani az adott kórokozó(k)
ellen. Ezt az immunológiai szempontú szekvencia csoportosítást használtuk fel négy új
pDNS alapú immunogén tervezésére, amik elsősorban a szubtípus-C, CRF-BC
(rekombináns forma), CRF-BF és CRF-AB-vel fertőzött betegek kezelésére nyújthatnak

70
immunológiailag optimalizált megoldást. Ezután elkészítettük a pDNS konstrukciókat
és belőlük a nanomedicina termékjelölteket, majd a szubtípus-B DermaVir vizsgálatára
kifejlesztett módszereinkkel jellemeztük azokat. Minden nanomedicina termékjelölt
antigén termelő képességét (fehérje repertoár, VLP+, biológiai aktivitás) igazoltuk, és a
termékjelöltek minőségét a klinikai vizsgálati DermaVir-hez hasonlónak találtuk.
Emellett in silico modelleztük az új termékjelöltek immunológiai potenciálját és
keresztvédő képességét, ami a megegyező szubtípusú/CRF-ű fertőzések esetében volt a
legjobb. Az új immunológiai fa használatát javasoljuk a legjobb kersztvédő képességű
termékjelölt ill. (személyre szabott) immunterápia kiválasztására is minden beteg
számára.

71
9. Summary
We characterized both in vitro and in silico, the HIV-1 subtype-B specific immunogen
(pDNA) of DermaVir therapeutic nanomedicine presently tested in Phase II clinical
trials in HIV-infected individuals. We have further designed and characterized a HIV-1
subtype-optimized nanomedicine product portfolio taking in consideration of the
genetic sequence variability of the patient’s HLA and HIV. During this work we
employed and developed both bioinformatics, molecular- and cell-biological methods.
We confirmed that DermaVir express fifteen HIV antigens in human cells that assemble
to a complex VLP (VLP+), immunological authentic to the wild-type HIV. However, it
is safe because it cannot integrate or reverse-transcribe new infectious virions. To
characterize the antigen expression and VLP+ release we have developed analytical
methods. To meet the FDA and EMA requirements we have developed a validated
potency assay for DermaVir vaccine that characterized the biological activity of the
product candidate. Our results provided explanation to the mechanism of action for
DermaVir-derived antigen presentation on the MHCI and MHCII molecules.
During the in silico and clinical analysis of DermaVir (subtype-B) we found the
rationale for the development of subtype-optimized vaccine product candidates. We
predicted its T cell specific immunological potential and modeled its cross-protection
capacity in non-B subtype specific infections. We found only modest cross-protection
of DermaVir with other subtype-specific infections of individual patients. This suggests
a decreased immunogenicity of subtype-B specific DermaVir vaccine in non-B subtype
of infections. Based on these results we developed an in silico tool for the rational
design of immunogens. The “immunological tree” clusters sequences containing similar
epitopes that determine the quality of immune responses against the certain antigen(s).
This immunological aspect of sequence grouping were used for the design of four new
pDNA immunogens targeting epidemics caused by subtype-C, CRF-BC (circulating
recombinant form), CRF-BF and CRF-AB. We have cloned these pDNA constructs,
prepared the nanomedicine product and characterized with our molecular- and cell-
biological methods developed for DermaVir subtype B vaccine. Expression of the
antigen repertoire and VLP+ were confirmed for all nanomedicines and quality of the
new product candidates was similar as for the clinically tested DermaVir. Additionally,
we modeled in silico the immunological potency and cross-protection capacity of the
new DermaVir products and showed the highest cross-reactivity with the matching
subtypes. We suggest the application of the novel “immunological tree” as a selection
tool for the best matching product and personalized immunotherapy for each patient.

72
10. Publikációk
Az értekezés alapját képező közlemények
Somogyi E, Xu J, Gudics Á, Tóth J, Kovács AL, Lori F, Lisziewicz J. A plasmid DNA
immunogen expressing fifteen protein antigens and complex virus-like particles (VLP+)
mimicking naturally occurring HIV. Vaccine 2011;29(4):744–753. (IF: 3,572)
Tőke ER, Lőrincz O, Somogyi E, Lisziewicz J. Rational development of a stable liquid
formulation for nanomedicine products. International Journal of Pharmaceutics
2010;392(1-2):261-7. (IF: 3,607)
Referált folyóiratban megjelent konferencia összefoglaló:
Somogyi E, Gudics Á, Forni DD, Molnár L, Lori F, Lisziewicz J. Subtype-optimized
DermaVir for personalized immunotherapy. Abstracts from AIDS Vaccine 2010. AIDS
Research and human retroviruses 2010, 26(10): A-1-A-184. (IF: 2,082)
A témában megjelent egyéb közlemények
Lőrincz O, Tőke ER, Somogyi E, Horkay F, Chandran P, Douglas JF, Szebeni J,
Lisziewicz J. Structure and biological activity of pathogen-like synthetic
nanomedicines. Nanomedicine: NMB 2011. Nyomtatás alatt. (IF: 4,882)
Szabadalom:
Csörgő SBZ, Garaczi E, Gruber L, Hamar P, Kemény L, Kökény G, Lisziewicz J,
Lőrincz O, Molnár M, Mózes M, Ötvös L, Pandur J, Pintér I, Somogyi E, Szabó KÁ,
Szollár L, Tőke ER. Immunogenic nanomedicine composition and preparation and uses
thereof. Pub. No.: WO/2010/125544, International Application No.:
PCT/IB2010/051909, Publication Date: 04.11.2010, International Filing Date:
30.04.2010

73
11. Köszönetnyilvánítás
Köszönöm témavezetőmnek, Dr. Lisziewicz Juliannának, a Genetic Immunity Kft.
vezérigazgatójának, hogy doktori munkám az ő irányítása alatt végezhettem. A
szaktudáson kívül céltudatosságot, kitartást és problémamegoldó gondolkodást tanultam
tőle.
Hálás vagyok kollégáimnak a közös-, és előremutató munkáért. Köszönöm a
bioinformatikus csoportnak: Molnár Leventének és Tóth Józsefnek a “rám szabott”
szoftvereket, amik nélkül sok in silico eredményem csak hipotézis lehetne. A vegyész
laborral mindig öröm volt a sok munka is, hálás vagyok nekik: Dr. Tőke Enikő, Lőrincz
Orsolya, Lakatos Mónika. Köszönöm Szűcsné Pulinka Ildikónak és Földiné Horváth
Erika technikusoknak a precíz és lelkiismeretes munkát, Gudics Ágnesnek a
felejthetetlen MBT éveket, és a csapatmunkát. Hálával tartozom Dr. Davide de
Forninak a klónozási feladatokban való részvételéért és szakmai segítségért, valamint
Dr. Franco Lorinak a mindig hasznos szakmai kritikáiért. Köszönöm Dr. Kovács Attila
Lajosnak az értékes elektronmikroszkópos felvételeket, Dr. Lisziewicz Zsoltnak pedig a
jó hangulatú brainstorming-eket.
A doktori munka elvégzéséhez az NKTH HIKC05 és DVCLIN01 pályázatok, valamint
a Research Institute for Genetic and Human Therapy (RIGHT) nyújtott anyagi
támogatást.
Hálával gondolok családom minden tagjára, mert szerető támogatásuk nélkül most nem
kerülhetne pont a disszertációm végére.

74
12. Irodalomjegyzék
[1]UNAIDS World AIDS Day Report 2011, www.unaids.org
[2]Webster TJ. Nanomedicine: what’s in a definition? International Journal of Nanomedicine 2006;1(2) 115–116.
[3]Hemelaar J, Gouws E, Ghys PD, Osmanov S. Global and regional distribution of HIV-1 genetic subtypes and recombinants in 2004. AIDS. 2006 Oct 24;20(16):W13-23.
[4]World Health Organization. Antiretroviral therapy of HIV infection in infants and children. Recommendations for a public health approach. 2006, Geneva.
[5]Pantaleo G, Demarest JF, Schacker T, Vaccarezza M, Cohen OJ, Daucher M, Graziosi C, Schnittman SS, Quinn TC, Shaw GM, Perrin L, Tambussi G, Lazzarin A, Sekaly RP, Soudeyns H, Corey L, Fauci AS. The qualitative nature of the primary immune response to HIV infection is a prognosticator of disease progression independent of the initial level of plasma viremia. Proceedings of the National Academy of Sciences of the United States of America 1997;94(1):254-8.
[6]Wyand MS, Manson K, Montefiori DC, Lifson JD, Johnson RP, Desrosiers RC. Protection by live, attenuated simian immunodeficiency virus against heterologous challenge. Journal of Virology 1999;73(10):8356-63.
[7]Gundlach BR, Lewis MG, Sopper S, Schnell T, Sodroski J, Stahl-Hennig C, Uberla K. Evidence for recombination of live, attenuated immunodeficiency virus vaccine with challenge virus to a more virulent strain. Journal of Virology 2000;74(8):3537-42.
[8]Paovonen J, Naud P, Salmeron J, Wheeler CM, Chow SN, Apter D, Kitchener H, Teixeira JC, Skinner SR, Jaisamrarn U, Limson G, Romanowski B, Aoki FY, Schwarz TF, Poppe WA, Bosch FX, Harper DM, Huh W, Hardt K, Zahaf T, Descamps D, Struyf F, Dubin G, Lehtinen M; HPV PATRICIA Study Group. Efficacy of human papillomavirus (HPV)-16/18 AS04-adjuvanted vaccine against cervical infection and precancer caused by oncogenic HPV types (PATRICIA): final analysis of a double-blind, randomised study in young women. Lancet 2009;374(9686):301-14.
[9]Villa LL, Perez G, Kjaer SK, Paavonen J, Lehtinen M, Munoz N, et al. Quadrivalent vaccine against human papillomavirus to prevent high-grade cervical lesions. New England Journal of Medicine 2007;356(19):1915-27.
[10]Hildesheim A, Herrero R, Wacholder S, Rodriguez AC, Solomon D, Bratti MC, Schiller JT, Gonzalez P, Dubin G, Porras C, Jimenez SE, Lowy DR; Costa Rican HPV Vaccine Trial Group. Effect of human papillomavirus 16/18 L1 viruslike particle vaccine among young women with preexisting infection - A randomized trial. Jama-Journal of the American Medical Association 2007;298(7):743-53.
[11]Broliden K, Hinkula J, Devito C, Kiama P, Kimani J, Trabbatoni D, Bwayo JJ, Clerici M, Plummer F, Kaul R.Functional HIV-1 specific IgA antibodies in HIV-1 exposed, persistently IgG seronegative female sex workers. Immunology Letters 2001;79(1-2):29-36.

75
[12]Rowland-Jones S, Sutton J, Ariyoshi K, Dong T, Gotch F, McAdam S, et al. HIV-specific cytoptoxic T-cells in HIV-exposed but uninfected Gambian women. Nature Medicine 1995;1(1):59-64.
[13]Rosenberg ES, LaRosa L, Flynn T, Robbins G, Walker BD. Characterization of HIV-1-specific T-helper cells in acute and chronic infection. Immunology Letters 1999;66(1-3):89-93.
[14]Lisziewicz J, Rosenberg E, Lieberman J, Jessen H, Lopalco L, Siliciano R, Walker B, Lori F. Control of HIV despite the discontinuation of antiretroviral therapy. New England Journal of Medicine 1999;340(21):1683-4.
[15]Johnson RP, Glickman RL, Yang JQ, Kaur A, Dion JT, Mulligan MJ, Desrosiers RC. Induction of vigorous cytotoxic T-lymphocyte responses by live attenuated simian immunodeficiency virus. Journal of Virology 1997;71(10):7711-8.
[16]Busch M, Abel K, Li J, Piatak M, Lifson JD, Miller CJ. Efficacy of a SHIV 89.6 proviral DNA vaccine against mucosal SIVmac239 challenge. Vaccine 2005;23(31):4036-47.
[17]McElrath MJ, De Rosa SC, Moodie Z, Dubey S, Kierstead L, Janes H, Defawe OD, Carter DK, Hural J, Akondy R, Buchbinder SP, Robertson MN, Mehrotra DV, Self SG, Corey L, Shiver JW, Casimiro DR; Step Study Protocol Team. HIV-1 vaccine-induced immunity in the test-of-concept Step Study: a case-cohort analysis. Lancet 2008;372(9653):1894-905.
[18]Weboldal: http://idshowcase.lshtm.ac.uk/
[19]Gelderblom HR, Ozel M, Pauli G. Morphogenesis and morphology of HIV. Structure-function relations. Arch Virol 1989;106:1-13.
[20]Sagar M. J Infect Dis. 2010 Oct 15;202 Suppl 2:S289-96.
[21]Fanales-Belasio E, Raimondo M, Suligoi B, Buttò S. Ann Ist Super Sanita. 2010;46(1):5-14.
[22]Bera S, Pandey KK, Vora AC, Grandgenett DP. Molecular interactions between HIV-1 integrase and the two viral DNA ends within the synaptic complex that mediates concerted integration. Journal of Molecular Biology 2009;389(1):183–98.
[23]McDougal JS, Mawle A, Cort SP, Nicholson JK, Cross GD, Scheppler-Campbell JA, Hicks D, Sligh J. Cellular tropism of the human retrovirus HTLV-III/LAV. I. Role of T cell activation and expression of the T4 antigen. J Immunol. 1985 Nov;135(5):3151-62.
[24]Siliciano RF, Greene WC. HIV Latency. Cold Spring Harb Perspect Med. 2011 Sep;1(1):a007096.
[25]Karn, J. Tackling Tat. J. Mol. Biol. 1999;293, 235-254.
[26]Feinberg MB, Baltimore D, Frankel AD.The role of Tat in the human immunodeficiency virus life cycle indicates a primary effect on transcriptional elongation. Proc Natl Acad Sci USA 1991;88: 4045-4049.
[27]Hope TJ. The ins and outs of HIV Rev. Arch Biochem Biophys. 1999;15;365(2):186-91.
[28]Ganser-Pornillos BK, Yeager M, Sundquist WI. The structural biology of HIV assembly. Curr Opin Struct Biol. 2008 Apr;18(2):203-17.
[29]Weboldal: http://www.uptodate.com/contents/immunology-of-hiv-1-infection

76
[30]Moore JP. Coreceptors: implications for HIV pathogenesis and therapy. Science. 1997;276(5309):51.
[31]Grouard G, Clark EA. Role of dendritic and follicular dendritic cells in HIV infection and pathogenesis. Curr Opin Immunol. 1997;9(4):563.
[32]Geijtenbeek TB, Kwon DS, Torensma R, van Vliet SJ, van Duijnhoven GC, Middel J, Cornelissen IL, Nottet HS, KewalRamani VN, Littman DR, Figdor CG, van Kooyk Y.DC-SIGN, a dendritic cell-specific HIV-1-binding protein that enhances trans-infection of T cells. Cell. 2000 Mar 3;100(5):587-97.
[33]Kahn JO, Walker BD. Acute human immunodeficiency virus type 1 infection. N Engl J Med. 1998;339(1):33.
[34]Pantaleo G, Fauci AS.Immunopathogenesis of HIV infection. Annu Rev Microbiol. 1996;50:825-54.
[35]Brenchley JM, Price DA, Schacker TW, Asher TE, Silvestri G, Rao S, Kazzaz Z, Bornstein E, Lambotte O, Altmann D, Blazar BR, Rodriguez B, Teixeira-Johnson L, Landay A, Martin JN, Hecht FM, Picker LJ, Lederman MM, Deeks SG, Douek DC. Microbial translocation is a cause of systemic immune activation in chronic HIV infection. Nat Med. 2006;12(12):1365.
[36]Hunt PW. Th17, gut, and HIV: therapeutic implications. Curr Opin HIV AIDS. 2010;5(2):189.
[37]Cooper MA, Fehniger TA, Fuchs A, Colonna M, Caligiuri MA. NK cell and DC interactions. Trends Immunol. 2004 Jan;25(1):47-52.
[38]Villadangos JA, Schnorrer P. Intrinsic and cooperative antigen-presenting functions of dendritic-cell subsets in vivo. Nat Rev Immunol. 2007 Jul;7(7):543-55.
[39]Bachmann MF, Lutz MB, Layton GT, Harris SJ, Fehr T, Rescigno M, Ricciardi-Castagnoli P. Dendritic cells process exogenous viral proteins and virus-like particles for class I presentation to CD8+ cytotoxic T lymphocytes. Eur J Immunol. 1996;26(11):2595.
[40]Sabado RL, Babcock E, Kavanagh DG, Tjomsland V, Walker BD, Lifson JD, Bhardwaj N, Larsson M. Pathways utilized by dendritic cells for binding, uptake, processing and presentation of antigens derived from HIV-1. Eur J Immunol. 2007 Jul;37(7):1752-63.
[41]Schwartz O, Marechal V, LeGall S, Lemonnier F, Heard JM. Endocytosis of major histocompatibility complex class I molecules is induced by the HIV-1 Nef protein. Nature Medicine 1996;2(3):338–42.
[42]Stumptner-Cuvelette P, Morchoisne S, Dugast M, Le Gall S, Raposo G, Schwartz O, Benaroch P. HIV-1 Nef impairs MHCclass II antigen presentation and surface expression. Proceedings of the National Academy of Sciences of the United States of America 2001;98(21):12144–9.
[43]Uebel S, Kraas W, Kienle S, Wiesmuller KH, Jung G, Tampe R. Recognition principle of the TAP transporter disclosed by combinatorial peptide libraries. Proc Natl Acad Sci U S A 1997 94:8976–81.
[44]Yewdell JW, Bennink JR. Immunodominance in major histocompatibility complex class I-restricted T lymphocyte responses. Annu Rev Immunol. 1999;17:51–88.

77
[45]Janssen EM, Lemmens EE, Wolfe T, Christen U, von Herrath MG, Schoenberger SP. CD4(+) T cells are required for secondary expansion and memory in CD8(+) T lymphocytes. Nature 2003;421(6925):852-6.
[46]Letvin NL, Walker BD. Immunopathogenesis and immunotherapy in AIDS virus infections. Nat Med. 2003 Jul;9(7):861-6.
[47]Wherry EJ, Ahmed R. Memory CD8 T-cell differentiation during viral infection. J Virol. 2004 Jun;78(11):5535-45.
[48]Poignard P, Sabbe R, Picchio GR, Wang M, Gulizia RJ, Katinger H, Parren PW, Mosier DE, Burton DR. Neutralizing antibodies have limited effects on the control of established HIV-1 infection in vivo. Immunity. 1999 Apr;10(4):431-8.
[49]Richman DD, Wrin T, Little SJ, Petropoulos CJ. Rapid evolution of the neutralizing antibody response to HIV type 1 infection. Proc Natl Acad Sci U S A. 2003 Apr 1;100(7):4144-9.
[50]Pelak K, Goldstein DB, Walley NM, Fellay J, Ge D, Shianna KV, Gumbs C, Gao X, Maia JM, Cronin KD, Hussain SK, Carrington M, Michael NL, Weintrob AC, Infectious Disease Clinical Research Program HIV Working Group, National Institute of Allergy and Infectious Diseases Center for HIV/AIDS Vaccine Immunology (CHAVI). Host determinants of HIV-1 control in African Americans. J Infect Dis. 2010;201(8):1141.
[51]Scorza Smeraldi R, Fabio G, Lazzarin A, Eisera N, Uberti Foppa C, Moroni M, Zanussi C. HLA-associated susceptibility to AIDS: HLA B35 is a major risk factor for Italian HIV-infected intravenous drug addicts. Hum Immunol. 1988;22(2):73
[52]De Leys R, Vanderborght B, Van den Haesevelde M, Heyndrickx L, Van Geel A, Wauters C, Bernaerts R, Saman E, Nijs P, Willems B. Isolation and partial characterization of an unusual human immunodeficiency retrovirus from two persons of west-central African origin. J Virol 1990;64(3):1207-16.
[53]Keele BF, Van Heuverswyn F, Li Y, Bailes E, Takehisa J, Santiago ML, Bibollet-Ruche F, Chen Y, Wain LV, Liegeois F, Loul S, Ngole EM, Bienvenue Y, Delaporte E, Brookfield JF, Sharp PM, Shaw GM, Peeters M, Hahn BH. Chimpanzee reservoirs of pandemic and nonpandemic HIV-1. Science. 2006 Jul 28;313(5786):523-6.
[54]Taylor BS, Sobieszczyk ME, McCutchan FE, Hammer SM. The Challenge of HIV-1 Subtype Diversity. N Engl J Med 2008; 358:1590-1602.
[55]Korber B, Muldoon M, Theiler J, Gao F, Gupta R, Lapedes A, Hahn BH, Wolinsky S, Bhattacharya, T. Timing the Ancestor of the HIV-1 Pandemic Strains. Science. 2000 Jun 9;288(5472):1789-96.
[56]Van Heuverswyn F, Li Y, Neel C, Bailes E, Keele BF, Liu W, Loul S, Butel C, Liegeois F, Bienvenue Y, Ngolle EM, Sharp PM, Shaw GM, Delaporte E, Hahn BH,Peeters M.. Human immunodeficiency viruses: SIV infection in wild gorillas. Nature 2006;444:164.
[57]Rambaut A, Posada D, Crandall KA, Holmes EC. The causes and consequences of HIV evolution. Nat Rev Genet. 2004 Jan;5(1):52-61.
[58]Perelson AS, Neumann AU, Markowitz M, Leonard JM, Ho DD. HIV-1 dynamics in vivo: virion clearance rate, infected cell life-span, and viral generation time. Science 1996;271:1582-6.

78
[59]Robertson DL, Anderson JP, Bradac JA, Carr JK, Foley B, Funkhouser RK, Gao F, Hahn BH, Kalish ML, Kuiken C, Learn GH, Leitner T, McCutchan F, Osmanov S, Peeters M,Pieniazek D, Salminen M, Sharp PM, Wolinsky S, Korber B. HIV-1 nomenclature proposal. Science. 2000 Apr 7;288(5463):55-6.
[60]Natz E, Lisziewicz J. Rational design of formulated DNA vaccines: the DermaVir approach. Gene Vaccines 2012, 127-143
[61]Lori F, Lewis MG, Xu J, Varga G, Zinn DE Jr, Crabbs C, Wagner W, Greenhouse J, Silvera P, Yalley-Ogunro J, Tinelli C, Lisziewicz J (2000) Control of SIV rebound through structured treatment interruptions during early infection. Science 290(5496):1591–1593.
[62]Walker BD. Elite control of HIV Infection: implications for vaccines and treatment. Top HIV Med. 2007 Aug-Sep;15(4):134-6.
[63]Toke ER, Lorincz O, Somogyi E, Lisziewicz J. Rational development of a stable liquid formulation for nanomedicine products. Int J Pharm 2010; 392(1-2):261-267.
[64]Lisziewicz J, Trocio J, Whitman L, Varga G, Xu J, Bakare N, Erbacher P, Fox C, Woodward R, Markham P, Arya S, Behr JP, Lori F. DermaVir: a novel topical vaccine for HIV/AIDS. J Invest Dermatol 2005;124(1):160–169.
[65]Collins KL, Chen BK, Kalams SA, Walker BD, Baltimore D. HIV-1 Nef protein protects infected primary cells against killing by cytotoxic T lymphocytes. Nature. 1998 Jan 22;391(6665):397-401.
[66]Lőrincz O, Tőke ER, Somogyi E, Horkay F, Chandran P, Douglas JF, Szebeni J, Lisziewicz J. Structure and biological activity of pathogen-like synthetic nanomedicines. Nanomedicine: NMB 2011. In press.
[67]Lisziewicz J, Gabrilovich DI, Varga G, Xu JQ, Greenberg PD, Arya SK, Bosch M, Behr JP, Lori F. Induction of potent human immunodeficiency virus type 1-specific T-cell-restricted immunity by genetically modified dendritic cells. Journal of Virology 2001;75(16):7621-8.
[68]Boussif O, Lezoualc’h F, Zanta MA, Mergny MD, Scherman D, Demeneix B, Behr JP. A versatile vector for gene and oligonucleotide transfer into cells in culture and in vivo: polyethylenimine. Proc Natl Acad Sci USA 1995;92(16):7297–7301.
[69]Matzinger P. The danger model: a renewed sense of self. Science 2002; 296(5566): 301-305.
[70]Nicolas JF and Guy B Intradermal, epidermal and transcutaneous vaccination: from immunology to clinical practice. Expert. Rev. Vaccines 2008; 7(8): 1201-1214.
[71]Bauer J, Bahmer FA, Worl J, Neuhuber W, Schuler G et al A strikingly constant ratio exists between Langerhans cells and other epidermal cells in human skin. A stereologic study using the optical disector method and the confocal laser scanning microscope. J. Invest. Dermatol. 2001;116(2):313-318.
[72]Lisziewicz J, Trocio J, Xu J, Whitman L, Ryder A, Bakare N, Lewis MG, Wagner W, Pistorio A, Arya S, Lori F. Control of viral rebound through therapeutic immunization with DermaVir. AIDS 2005;19(1):35–43.

79
[73]Lisziewicz J, Calarota SA, Banhegyi D, Lisziewicz Z, Ujhelyi E, Lori F. Single DermaVir patch treatment of HIV+ individuals induces long-lasting, high-magnitude, and broad HIV-specific T cell responses. Poster presentation. 15th conference on retroviruses and opportunistic infections, Boston, MA, 3–6 Feb 2008.
[74]Lisziewicz J, Bakare N, Calarota SA, Bánhegyi D, Szlávik J, Újhelyi E, Tőke ER, Molnár L, Lisziewicz Z, Autran B, Lori F. Single DermaVir Immunization: Dose-dependent Expansion of Precursor/Memory T Cells Against All HIV Antigens in HIV-1 Infected Individuals. Plos One 2012. In press.
[75]Calarota SA, Foli A, Maserati R, Baldanti F, Paolucci S Young MA, Tsoukas CM, Lisziewicz J, Lori F. HIV-1-Specific T Cell Precursors with High Proliferative Capacity Correlate with Low Viremia and High CD4 Counts in Untreated Individuals. J. Immunol. 2008; 180(9): 5907-5915.
[76]Rodriguez B, Asmuth D, Matining R, Spritzler J, Li X, Jacobson J, Read S, Lisziewicz J, Lori F, Pollard R, Team AS. Repeated-dose transdermal administration of DermaVir, a candidate plasmid DNA-based therapeutic HIV vaccine, is safe and well-tolerated: a 61-week analysis of ACTG Study 5176. Presented at the XVIIIth International AIDS Conference, 2010, Vienna, Austria
[77]van Lunzen J, Pollard R, Stellbrink HJ, Plettenberg A, Natz E, Lisziewicz Z, Freese R, Molnar L, Calarota SA, Lori F, Lisziewicz J. DermaVir for initial treatment of HIV-infected subjects demonstrates preliminary safety, immunogenicity and HIV-RNA reduction versus placebo immunization. Presented at the XVIII international AIDS conference, 2010, Vienna, Austria
[78]ICH Q2(R1) guideline: Validation of analytical procedures: Text and Methodology
[79]Weboldal: http://tools.immuneepitope.org/
[80]Peters B, Bui HH, Frankild S, Nielsen M, Lundegaard C, Kostem E, Basch D, Lamberth K, Harndahl M, Fleri W, Wilson SS, Sidney J, Lund O, Buus S, Sette A. A community resource benchmarking predictions of peptide binding to MHC-I molecules. PLoS Computational Biology 2006;2(6):574–84.
[81]Wang P, Sidney J, Dow C, Mothe B, Sette A, Peters B. A systematic assessment of MHC class II peptide binding predictions and evaluation of a consensus approach. PLoS Computational Biology 2008;4(4).
[82]Nielsen M, Lundegaard C, Worning P, Lauemoller SL, Lamberth K, Buus S, Brunak S, Lund O. Reliable prediction of T-cell epitopes using neural networks with novel sequence representations. Protein Science 2003;12(5):1007–17.
[83]Korber TMB, Brander C, Haynes BF, Koup R, Moore JP, Walker BD, et al., editors. HIV Molecular Immunology 2008. Los Alamos, New Mexico: Los Alamos National Laboratory, Theoretical Biology and Biophysics, 2008.
[84]Saitou, N., Nei, M. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987; 4;406-625.
[85]Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980;16;111-120.
[86]Felsenstein, J. Confidence limits on phylogenies: an approach using the bootstrap. Evolution 1985;39;783-791.

80
[87]Lori F, Veronese FD, Devico AL, Lusso P, Reitz MS, Gallo RC. Viral-DNA carried by human-immunodeficiency-virus type-1 virions. Journal of Virology 1992;66(8):5067-74.
[88]Malykh A, Reitz MS, Louie A, Hall L, Lori F. Multiple determinants for growth of human-immunodeficiency-virus type-1 in monocyte-macrophages. Virology 1995;206(1):646-50.
[89]Piccinini G, Foli A, Comolli G, Lisziewicz J, Lori F. Complementary antiviral efficacy of hydroxyurea and protease inhibitors in human immunodeficiency virus-infected dendritic cells and lymphocytes. Journal of Virology 2002;76(5):2274-8.
[90]Cara A, Guarnaccia F, Reitz MS, Gallo RC, Lori F. Self-limiting, cell typedependent replication of an integrase defective human-immunodeficiencyvirus type-1 in human primary macrophages but not T-lymphocytes. Virology 1995;208(1):242–8.
[91]Gruters RA, van Baalen CA, Osterhaus A. The advantage of early recognition of HIV-infected cells by cytotoxic T-lymphocytes. Vaccine 2002;20(15):2011–5.
[92]Somogyi E, Xu J, Gudics Á, Tóth J, Kovács AL, Lori F, Lisziewicz J. A plasmid DNA immunogen expressing fifteen protein antigens and complex virus-like particles (VLP+) mimicking naturally occurring HIV. Vaccine 2011;29(4):744–753.
[93]Quan FS, Steinhauer D, Huang CZ, Ross TM, Compans RW, Kang SM. A bivalent influenza VLP vaccine confers complete inhibition of virus replication in lungs. Vaccine 2008;26(26):3352-61.
[94]Altfeld M, Allen TM, Yu XG, Johnston MN, Agrawal D, Korber BT, Montefiori DC, O'Connor DH, Davis BT, Lee PK, Maier EL, Harlow J, Goulder PJ, Brander C,Rosenberg ES, Walker BD. HIV-1 superinfection despite broad CD8+ T-cell responses containing replication of the primary virus. Nature. 2002 Nov 28;420(6914):434-9.
[95]Streeck H, Li B, Poon AF, Schneidewind A, Gladden AD, Power KA, Daskalakis D, Bazner S, Zuniga R, Brander C, Rosenberg ES, Frost SD, Altfeld M, Allen TM. Immune-driven recombination and loss of control after HIV superinfection. J Exp Med. 2008 Aug 4;205(8):1789-96.
[96]Somogyi E, Gudics Á, Forni DD, Molnár L, Lori F, Lisziewicz J. Subtype optimized DermaVir for personalized immunotherapy. Abstracts from AIDS Vaccine 2010. AIDS Research and human retroviruses 2010, 26(10): A-1-A-184.
[97]Weboldal: http://www.hiv.lanl.gov/
[98]Pantaleo G, Koup RA. Correlates of immune protection in HIV-1 infection: what we know, what we don't know, what we should know. Nat Med. 2004 Aug;10(8):806-10.
[99]Mascola JR, Montefiori DC. The Role of Antibodies in HIV Vaccines. Annu. Rev. Immunol. 2010. 28:413–44.
[100]Koup RA, Graham BS, Douek DC. The quest for a T cell-based immune correlate of protection against HIV: a story of trials and errors. Nat Rev Immunol. 2011 Jan;11(1):65-70.

81
[101]Mouquet H, Scheid JF, Zoller MJ, Krogsgaard M, Ott RG, Shukair S, Artyomov MN, Pietzsch J, Connors M, Pereyra F, Walker BD, Ho DD, Wilson PC, Seaman MS, Eisen HN, Chakraborty AK, Hope TJ, Ravetch JV, Wardemann H, Nussenzweig MC. Polyreactivity increases the apparent affinity of anti-HIV antibodies by heteroligation. Nature 2010;467(7315):591-595.
[102]Mouquet H, Warncke M, Scheid JF, Seaman MS, Nussenzweig MC. Proc Natl Acad Sci U S A. Enhanced HIV-1 neutralization by antibody heteroligation. 2012 Jan 17;109(3):875-80.
[103]Betts MR, Casazza JP, Koup RA. Monitoring HIV specific CD8+ T cell responses by intracellular cytokine production. Immunol Lett. 2001 Nov 1;79(1-2):117-25.
[104]Abbott A. Therapeutic HIV vaccines show promise. Nature 2010;466,539, published online. http://www.nature.com/news/2010/100727/full/466539a.html
[105]Sandstrom E, Nilsson C, Hejdeman B, Brave A, Bratt G, Robb M, Cox J, Vancott T, Marovich M, Stout R, Aboud S, Bakari M, Pallangyo K, Ljungberg K, Moss B,Earl P, Michael N, Birx D, Mhalu F, Wahren B, Biberfeld G; HIV Immunogenicity Study 01/02 Team. Broad Immunogenicity of a Multigene, Multiclade HIV-1 DNA Vaccine Boosted with Heterologous HIV-1 Recombinant Modified Vaccinia Virus Ankara. Journal of Infectious Diseases 2008;198(10):1482-90.
[106]Ellenberger D, Wyatt L, Li B, Buge S, Lanier N, Rodriguez IV, Sariol CA, Martinez M, Monsour M, Vogt J, Smith J, Otten R, Montefiori D, Kraiselburd E, Moss B,Robinson H, McNicholl J, Butera S. Comparative immunogenicity in rhesus monkeys of multi-protein HIV-1 (CRF02_AG) DNA/MVA vaccines expressing mature and immxature VLPs. Virology 2005;340(1):21-32.
[107]Masemola A, Mashishi T, Khoury G, Mohube P, Mokgotho P, Vardas E, Colvin M, Zijenah L, Katzenstein D, Musonda R, Allen S, Kumwenda N, Taha T, Gray G,McIntyre J, Karim SA, Sheppard HW, Gray CM; HIVNET 028 Study Team. Hierarchical Targeting of Subtype C Human Immunodeficiency Virus Type 1 Proteins by CD8+ T Cells: Correlation with Viral Load. Journal of Virology 2004; 78(7):3233–3243.
[108]Kiepiela P, Ngumbela K, Thobakgale C, Ramduth D, Honeyborne I, Moodley E, Reddy S, de Pierres C, Mncube Z, Mkhwanazi N, Bishop K, van der Stok M, Nair K, Khan N, Crawford H, Payne R, Leslie A, Prado J, Prendergast A, Frater J, Brander C, Learn GH, Nickle D, Rousseau C, Coovadia H, Mullins JI, Heckerman D, Walker BD, Goulder P. CD8+ Tcell responses to different HIV proteins have discordant associations with viral load. Nat Med. 2007 Jan;13(1):46-53.
[109]Vider-Shalit T, Fishbain V, Raffaeli S, Louzoun Y. Phase-dependent immune evasion of herpesviruses. Journal of Virology 2007;81(17):9536-45.
[110]Fischer W, Perkins S, Theiler J, Bhattacharya T, Yusim K, Funkhouser R, Kuiken C, Haynes B, Letvin NL, Walker BD, Hahn BH, Korber BT. Polyvalent vaccines for optimal coverage of potential T-cell epitopes in global HIV-1 variants. Nat Med 2007;13(1)100-6.
[111]Blauvelt A, Asada H, Saville MW, KlausKovtun V, Altman DJ, Yarchoan R, Katz SI. Productive infection of dendritic cells by HIV-1 and their ability to capture virus are mediated through separate pathways. Journal of Clinical Investigation 1997;100(8):2043-53.

82
[112]Ellenberger D, Li B, Smith J, Yi H, Folks T, Robinson H, Butera S. Optimization of a multi-gene HIV-1 recombinant subtype CRF02_AG DNA vaccine for expression of multiple immunogenic forms. Virology 2004;319(1):118-30.
[113]Weaver EA, Lu Z, Camacho ZT, Moukdar F, Liao HX, Ma BJ, Muldoon M, Theiler J, Nabel GJ, Letvin NL, Korber BT, Hahn BH, Haynes BF, Gao F. Cross-subtype T-cell immune responses induced by a human immunodeficiency virus type 1 group M consensus env immunogen. J Virol 2006;80:6745-56.
[114]Barouch DH, O'Brien KL, Simmons NL, King SL, Abbink P, Maxfield LF, Sun YH, La Porte A, Riggs AM, Lynch DM, Clark SL, Backus K, Perry JR, Seaman MS,Carville A, Mansfield KG, Szinger JJ, Fischer W, Muldoon M, Korber B. Mosaic HIV1 vaccines expand the breadth and depth of cellular immune responses in rhesusmonkeys. Nat Med. 2010 Mar;16(3):319-23.