histological and histochemical studies on the subfornical organ of the squirrel monkey
TRANSCRIPT

Histochemie 13, 331--345 (1968)
Histological and Histochemical Studies on the Subfornical Organ of the Squirrel Monkey
Y. •AKAJIMA*, T. R. SHANTHA**, and G. H. BOURN~
Yerkes Regional Primate Research Center, Emory University, Atlanta, Georgia 30322, U.S.A.
Received March 8, 1968
Summary. Detailed studies have been made on the distribution of several enzymes in the subfornical organ (SFO) of the squirrel monkey. In this species, the nerve cells of the SFO show reactions of varying intensity for enzymes of the glycolytic and aerobic pathways. The nerve cells, glial cells and ependymal cells of the SFO and the choroid plexus are equipped with enzymes of the Embden-Meyerhof (EM) pathway, pentose cycle and tricarboxylic acid (TCA) cycle. Many nerve cells and oligodendroglia in the body of this organ are rich in enzymes of the TCA cycle and the pentose cycle and thus presumably have the capacity of producing adenosine triphosphate (ATP) and reduced nicotinamide adenine dinucleotide phosphate (NADPH2) [reduced triphosphopyridine nucleotide (TPNH)]. In the neurons, ATP is probably used as energy for synaptic transmission, active transport, secretion and various other metabolic processes, whereas NADPH 2 is used for synthetic processes such as the production of fatty acids and some amino acid conversion (e.g., conversion of phenylalanine into tyrosine). The SFO and its stalks contain both cholinergic and adrenergic neurons and fibers. The outermost layer of the perivascular sheath gives a positive reaction for enzymes of the gylcolytic pathways (EM pathway, pentose cycle and TCA cycle), whereas the inner layer of this sheath shows negligible activity for these enzymes. On the other hand, the whole sheath (inner and outer layers) exhibits strong staining for Mg++-activated adenosine triphosphatase (ATPase), and moderate staining for Ca++-activated ATPase. This sheath, rich in ATPase, may carry on active transport and such related functions. Since the outermost layer contains various enzymes of the glycolytic pathways, it is possible that the ATP required for these functions is produced in this layer.
Introduction
The subfornical organ (SFO) is s i tua ted on the an te r ior wall of the th i rd ventr icle . This special s t ruc ture is found in var ious animals (e.g., Amphib ians , Rept i les , Ayes and Mammals ) and i ts funct ion is obscure (AKERT et al., 1961; HOrER, 1965). Not m a n y his tochemical s tudies have been made on this organ. The s tudies made so far on the SFO of the ra t , r a b b i t and guinea pig indica te t h a t anaerobic glycolysis might be a p r e d o m i n a n t fea ture of this organ (SH~MIZU et al., 1957 ; S~IMIZU and MORIKAWA, 1957). Other workers s tudy ing the enzymes of the t r i ca rboxyl ic acid (TCA) cycle, and the Warburg -Diekens p a t h w a y of the SFO in the ra t , c la im t h a t aerobic, not anaerobic glycolysis is p r e d o m i n a n t (NAKA- JI~A, 1964, 1966). Several o ther workers refer to the enzymes of the SFO, bu t mos t of the i r descr ip t ions are shor t and give l i t t le de ta i led in format ion (SrtIMiZU and OKADA, 1957; MATSUNAMI, 1959; AB~ et al., 1963; ATHERTON, 1963). Because of the lack of de ta i l ed s tudies on the higher orders of mammals , especial ly pr imates , and because of the con t roversy as to whe ther an aerobic or an anaerobic t y p e
* Visiting scientist from the Department of Anatomy, Tokyo Medical and Dental Uni- versity, Tokyo, Japan
** T. R. S~ANTH~,VEERAI"I"A in previous publications.

332 Y. NAKAJIMA, T. R. SHANTHA and G. H. BOURN~:
of me tabo l i sm is occurr ing in the SFO, the present s t u d y was unde r t aken . The squirrel m o n k e y (a New Wor ld monkey) was selected because i t has a well deve loped SFO (AKERT et al. , 1961) and is easi ly avai lable for l a b o r a t o r y use.
The term glycolytic pathways is used with different connotations by different workers. Some workers use it to include only the pentose cycle and the Embden-Meyerhof (EM) path- way, whereas others, including the present authors, include the TCA cycle as well.
Materials and Methods Adult male and female Squirrel Monkeys (Saimiri sciureus) were used in this study. The
animals were anesthetized by an appropriate dose of Nembutal. The part of the brain including the SFO was removed, cut into a 5 mm block, placed on aluminum foil and frozen in dry ice. Ten micron thick sagittal sections were cut at -- 20 ~ C in a cryostat. The following histochemi- cal tests were applied to the sections: Acid phosphatase (AC) was tested by BURSTONE'S (1962) method; adenosine triphosphatase (ATPase) by PADYKULA and HERMAN'S (1955), and WACHSTEIN and MEISEL'S (1957) methods; specific (ACHE) and nonspecific (BuChE) cholin- esterase by COUPLA•D and HOL~ES' (1957) method; monoamine oxidase (MAO) by the method of GLE~ICER et al., (1957); amylophosphorylase (AP) by TAKEUCHI and KURIAKI'S (1955) method; glucose-6-phosphate dehydrogenase (G6PD) by the method of HESS et al., (1958); aldolase (AD) by ABE and SHIMIZU'S (1964) method; lactic dehydrogenasc (LDH) by the method of HESS et al., (1958) and BRODY and E~GEL'S (1964) method - - only " tota l" LDH was tested by BRODu and ENGEL'S method through the standard medium; succinic dehydro- genase (SDH) by the method of NAC~LAS etal. , (1957). We also demonstrated G6PD by slightly modifying the above-mentioned method: Approximately 0.1 ml of 0.2 per cent phenazine methosulfate was added to 5.0 ml of incubating solution.
Histological studies on the SFO were made on formalin-fixed, paraffin-embedded sections. These sections were stained with the standard Hematoxylin and Eosin (H&E) method, the Nissl staining method using cresyl violet, the periodic acid SCHIFF (PAS) method of McMANuS (1946), WEIL'S (1945) method for myelin sheaths and BODIA~'S (1936) method for nerve fibers.
Results
The SFO of the squirrel m o n k e y is s i tua ted a t the middle of the an te r ior wall of the th i rd ventr icle , a t the level of the in te rven t r i cu la r foramen (Figs. 1, 2). The pos ter ior border of this organ projects in to this ventr icle . I t measures abou t 0.95 m m dorsovent ra l ly , 0.45 m m ros t rocaudal ly , and 0.55 m m medio la te ra l ly (AKERT et al. , 1961). The choroid plexus is a t t a c h e d to i ts dorsa l aspect . Three f luid sys tems meet a t the S F O : (1) the cercbrospinal f luid of the ventr ic le pene- t r a t es into the dep ths of the SFO th rough the ependyma l canalicul i found on i ts ven t r icu la r surface (ANDRES, 1965) ; (2) the f luid of the subarachnoida l space reaches the SFO th rough the t ransverse fissure (SrRANKEL, 1960) (Fig. 2); (3) the blood supp ly of the SFO enters th rough a branch of the anter ior cerebral a r t e ry and also th rough a ve r tebra l a r t e ry by way of the choroid ar ter ies (SeoER~L 1963). AKERT et al. , (1961) emphas ized the re la t ionship of the SFO with cer ta in nuclei of the septa l region as well as t h a t wi th the fornix sys tem (Figs. 1, 2). This organ can be subd iv ided into three por t ions ; a body, a ven t ra l s ta lk and a dorsal s ta lk (Fig. 2) (AKERT et al. , 1961). The neuropi l of the SFO is loose due to the presence of wide in tercel lu lar spaces. Innumerab le blood vessels arc found in it. There are m a n y sinusoids sur rounded b y P A S posi t ive per ivascular sheaths which remain posi t ive even af te r the dias tase digest ion tes t (Figs. 23, 24). This indica tes t h a t the P A S posi t ive ma te r i a l is not glycogen, and, is p r o b a b l y a complex mucopoly- saccharide. The SFO contains as t rocytes , ol igodendrogl ia and smal l nerve cells.

Studies on the Subfornical Organ 333
" ' - v J
c c
::'.. ~ _ 7..!!."
: , . ' . i" .": .".i.: : . : �9 i.:-. .. � 9
f f f
Fig. 1 Fig. 2
Fig. 1. A coronal section of squirrel monkey brain passing through the SFO, showing the relationship of this structure to the surrounding components of the brain at this level.
AC anterior commissure OCH A P L area praeoptica lateralis P M A P M area praeoptica medialis SCH CC corpus callosum CD caudatc nucleus SF CHO choroid plexus SFO D dorsal stalk of the subfornical organ SG F fornix SH HC hippocampal commissure S T L T lamina terminalis TF M I massa intermedia T R S NCA nucleus commissuralis anterior V N F D B nucleus fasciculus diagonalis Broca I I I
optic chiasm nucleus praeopticus medianus nucleus suprachiasmaticus hypo-
thalami nucleus septalis fimbrialis subfornical organ septal giant cells nucleus septohippocampalis stria terminalis transverse fissure nucleus triangularis septi ventral stalk of the subfornical organ ventriculus tertius
Fig. 2. A saggital section through the squirrel monkey brain showing the SFO and its relation to surrounding structures. (Based on AKERT et al., 196I)
The nerve cells are oval, fusiform and t r iangular in shape, and have fine Nissl granules in their cytoplasm (Figs. 25, 26, 27). The dorsal and vent ra l stalks conta in

334 Y. NAKAJTMA, T. R. SHANTHA and G. H. BOURNE:
Fig. 3. Body of the SFO showing moderate AC positive nerve cells with mild nucleolar staining (arrows), a moderately positive astrocyte (arrow A) and strongly stained oligodendroglia
(arrow 0). Note the positive nerve process (arrow P). • 1,125
Fig. 4. Dorsal stalk of the SFO showing a strongly AC positive nerve cell and nerve fibers (arrows). • 1,125
Fig. 5. Ventral stalk of the SFO. AC test. Note the strong reaction in the apical part of epen- dymal cells. • 1,125
Fig. 6. Body of the SFO. Ca++-activated ATPase test. The nerve cells show moderate nuclear, very strong nucleolar (arrows) and mild to moderate cytoplasmic staining. An astrocyto shows a moderate reaction (arrow A). Oligodendroglia is moderately to strongly positive (arrows O).
• 1,125
Fig. 7. Body of the SFO showing strongly Ca++-activated ATPase positive vascular sheaths. • 300
Fig. 8. Dorsal stalk showing many strongly Ca++-activated ATPase positive fibers and BV's. • 1,125

Studies on the Subfomical Organ 335
a lesser number of blood vessels, glial cells and nerve cells than does the body of the SFO. The dorsal stalk has many nerve fibers tha t are demonstrable by BODIA:N'S and W~IL'S methods (Figs. 28, 29). The ependymal cells covering the body are cuboidal and squamous in character (Fig. 30). Typical (columnar) ependymal cells are found only on the surface of the ventral stalk, below which are evident many subependymal vacuoles (Fig. 31). The dorsal stalk gives at- tachment to the choroid plexus and has no ependymal lining (Fig. 32).
A. Acid Phosphatase
The neuropil of the body gave a moderate to strong reaction. Some oligo- dendroglia and astrocytes showed very strong activity, whereas other glial cells were moderately positive (Fig. 3). Some nerve cells were strongly active and others were moderately stained ; their cytoplasm showed both granular and diffuse types of reactions; the nucleoli of the neurons were mildly positive (Figs. 3, 4). Mildly to moderately positive nerve processes and strongly stained nerve fibers were found in the body (Fig. 3). The endothelial cells of the blood vessels (BV's) were moderately positive. The perivascular sheaths showed a negligible reaction ex- cept for a moderate to strong reaction in the outermost layer. The neuropil of the stalks showed mild activity. Strongly stained nerve fibers were found in the stalks (Fig. 4). The ependymal cells showed strong activity confined to their apical parts (Fig. 5). The choroid plexus exhibited a very strong reaction.
B. Adenosine Triphosphatase
PADYKULA and HERMAN'S Method. The ncuropil of the body of the SFO was moderately positive for Ca++-activated ATPase. The astrocytes of the body reacted moderately. The oligodendroglia showed moderate to strong activity (Fig. 6). The cytoplasm of the nerve cells revealed mild to moderate activity, while their nuclei were stained either moderately or strongly. The nucleoli of the neurons were very strongly positive (Fig. 6). The nerve processes and nerve fibers of the body showed a moderate reaction. The endothelial cells and perivascular sheaths reacted strongly (Fig. 7). In the stalks, the neuropil showed a mild to
Fig. 9. Body of the SFO. Mg++-activated ATPase test. The nerve cells reveal strong nuclear, very strong nucleolar (arrows) and moderate cytoplasmic staining. Astrocytes (arrows A)
are mildly or moderately positive and oligodendroglia (arrow O) moderately positive. • 1,125
Fig. 10. Body of the SFO showing AChE positive nerve cells. Note strongly (arrow S), moderately (arrows m) and mildly positive neurons (arrows). • 480
Fig. 11. Ventral stalk showing moderately BuChE positive ependymal cells. • Fig. 12. Body of the SFO. MAO test. Nerve cells are moderate to strongly positive with moderate nucleolar staining (arrows). Note the strongly stained nerve fibers (arrows n).
• 1,125 Fig. 13. Body showing a moderately AP positive nerve cell with nucleolar staining (arrow), moderately positive astrocytes (arrows A) and strongly positive oligodendroglia (arrows O).
• 1,125 :Fig. 14. Ventral stalk of the SFO. A moderate AP reaction is found in the apical part of
ependymal cells (arrows). • 1,125
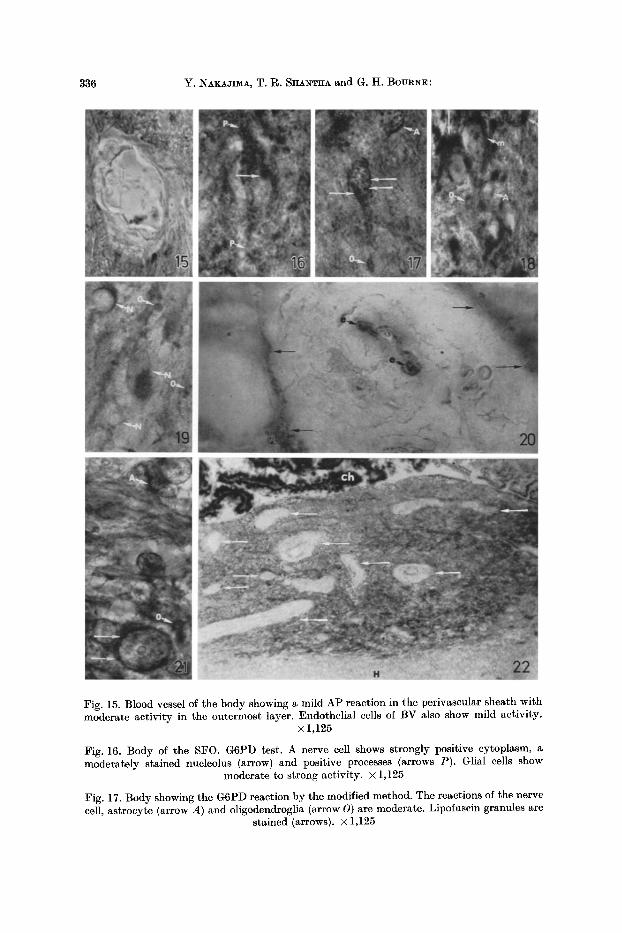
336 Y. NAKAJIMA, T. R. SHA~T~A and G. H. BOURNE:
Fig. 15. Blood vessel of the body showing a mild AP reaction in the perivascular sheath with moderate activity in the outermost layer. Endothelial cells of BV also show mild activity.
• 1,125
Fig. 16. Body of the SFO. G6PD test. A nerve cell shows strongly positive cytoplasm, a moderately stained nucleolus (arrow) and positive processes (arrows P). Glial cells show
moderate to strong activity. • 1,125
Fig. 17. Body showing the G6PD reaction by the modified method. The reactions of the nerve cell, astrocyte (arrow A) and oligodendroglia (arrow O) are moderate. Lipofnscin granules are
stained (arrows). • 1,125

Studies on the Subfornical Organ 337
modera te reac t ion and the glial cells were mode ra t e ly act ive. The reac t ion of the nerve cells of the s ta lks was the same as t h a t of the body. Very s t rongly s ta ined nerve fibers were found in the s ta lks ; especial ly the dorsa l s ta lk which conta ined m a n y posi t ive fibers (Fig. 8). The ependyma l cells were mode ra t e ly act ive. The cells of the choroid plexus showed a modera t e to s t rong reac t ion in the cytoplasm.
WACHSTEIN and MEISEL'S Method. The neuropi l showed a mild react ion. The ol igodendrogl ia reac ted modera te ly . The as t rocy tes were mi ld ly or mode ra t e ly posi t ive (Fig. 9). The nerve cells had mode ra t e ly ac t ive cytoplasm, a mode ra t e ly or s t rongly s ta ined nucleus and a ve ry s t rongly s ta ined nncleolus (Fig. 9). Some- t imes, posi t ive nerve processes were observed ex tend ing f rom the pe r ika ryon of the nerve cells. S t rong ly s ta ined nerve fibers were found in the body and s ta lks ; m a n y such fibers were seen in the dorsa l s talk. The endothe l ia l cells of the BV's showed a s t rong react ion. The pcr ivascular shea th was mode ra t e ly posi t ive. The e p e n d y m a l cells and choroid plexus reac ted s t rongly.
C. Speci/ic Cholinesterase
The neuropi l of the body showed a negligible react ion. The glial cells were negat ive . The nerve cells of the body revealed var ious t ypes of reac t ions : some cells were s t rong ly posi t ive, some mode ra t e ly posi t ive, some mi ld ly posi t ive and others negat ive (Fig. 10). The nucleoli of the neurons showed a neghgible react ion. Some nerve processes were negat ive and others showed a mild or modera te posi t ive reac t ion ex tend ing for some dis tance into the nerve process. Mildly act ive nerve fibers were found in the body. The endothe l ia l cells of the BV's showed a negligible react ion. The per ivascu la r shea th was negat ive . I n the dorsa l and ven t ra l s ta lks some nerve cells were negat ive and others were mode ra t e ly act ive. Modera te ly posi t ive nerve fibers were found in the stalks. The e p e n d y m a l cells and the choroid
plexus were negat ive .
Fig. 18. Body of the SFO. AD test. Note a moderately positive nerve cell (arrow m) and a strongly positive one with formazan granules thick in a part of the cytoplasm (arrow). An astrocyte (arrow A) shows moderate activity and the oligodendroglia (arrows O) reveal
strong activity. • 1,125
Fig. 19. Body of the SFO showing the LDH reaction (by the method of HESS et al.). Nerve cells (arrows N) are strongly and moderately positive. Oligodendroglia (arrows O) are moderately
positive. • 1,I25
Fig. 20. Blood vessel of the body showing LDH activity. The endothelial cells (arrows e) are strongly stained. The vascular sheath shows a negligible reaction except for a moderately
strong reaction in the outermost layer (arrows). • 1,125
Fig. 21. Body of the SFO. BRODY and ENGEL'S total LDH test. A nerve cell, oligodendroglia (arrows O) and astrocyte (arrow A) show moderate to strong activity. Lipofuscin granules in
the neuron are very strongly stained (arrows). • 1,125
Fig. 22. Body of the SFO (in saggital section). SDH test. The choroid plexus is very strongly positive. The neuropil shows moderate activity. The BV's (arrows) show positive activity only in the intima and the outermost layer of the perivascular sheath. Note the presence of in- numerable BV's in the SFO. The hippocampal commissure (fornix) is slightly stained. • 150
23 a Histochemie, Bd. 13

338 Y. •AKAJIMA, T. R. SItANTItA and G. H. BOURNE:
D. Nonspeci/ic Cholinesterase
A negligible reaction was found in the neuropil, glial cells, nerve processes and nerve fibers of the body. Only some nerve cells had moderately positive cyto- plasm, whereas the rest of them showed a negligible reaction. The BV's were negative. In the dorsal and ventral stalks, the nerve cells and nerve fibers revealed a negligible reaction. The ependymal cells were moderately positive (Fig. l l) . The choroid plexus reacted mildly.
E. Monoamine Oxidase
The medial part of the body, which was close to the third ventricle, was more strongly stained than the rest of the SFO. In the body, the neuropil reacted moderately, and the glial cells and nerve processes were mildly positive. Some nerve cells showed negligible activity, while others had moderate to strong cyto- plasmic activity. The positive reaction in neuronal cytoplasm was both granular and diffuse. The nucleoli of the neurons were moderately positive (Fig. 12). The body contained moderately as well as strongly stained nerve fibers (Fig. 12). Only the outermost layer of the perivascular sheath of the BV's showed a positive reaction. The stalks contained many positive nerve fibers. A mild reaction was found in the neuropil, glial cells and nerve cells of the stalks. The ependymal cells reacted mildly to moderately. The choroid plexus revealed moderately strong staining.
F. A m ylophosphor ylase
The neuropil was moderately positive. In the body, the oligodendroglia showed a strong reaction, and astrocytes showed moderate activity (Fig. 13). Some nerve cells of the body had moderately positive cytoplasm, and others were mildly active. Many neurons showed a moderate reaction on their cell membranes (Fig. 13). Nerve processes were mildly stained. The glial cells and nerve cells of the stalks were mildly active. A mild reaction was also observed in the intima of the BV's and also in the perivascular sheath, the outermost layer of which was moderately positive (Fig. 15). The ependymal cells and the choroid plexus lining cells showed moderate activity; the reaction was mostly confined to the apical parts of the lining cells in the ependyma (Fig. 14).
G. Glucose-6-phosphate Dehydrogenase
In the body of the SFO, the neuropil reacted moderately, and the glial cells showed a moderate to strong reaction. Some nerve cells of the body were moder- ately positive, and others were strongly positive. The nucleoli of the neurons were moderately stained. The nerve processes (Fig. 16) and nerve fibers of the body showed moderate to strong activity. The endothelial cells of the BV's were moderately stained. The perivascular sheath was negative except tha t a strong beaded reaction was found in the outermost layer of the sheath. In the stalks, the neuropil showed a mild reaction, and the glial cells, nerve cells and nerve fibers revealed moderate activity. The ependymal cells showed a moderate re- action which was confined to their apical parts. The choroid plexus was very strongly stained.

Studies on the 8ubfornical Organ 339
When phenazine methosulfate was added to the incubating medium, the lipofuscin granules of the neurons (Fig. 17), and the myelin sheaths of the nerve fibers were stained; a strong reaction was found in the ependymal cells.
H. A ldolase
The neuropil in the body of the SFO showed a moderate reaction. The astro- cytes were moderately positive. The oligodendroglia showed strong activity (Fig. 18). Some nerve cells of the body were moderately positive and others were strongly positive with formazan granules distributed evenly throughout the cyto- plasm. In some neurons, the formazan granules were found aggregated at one pole of the cell cytoplasm (Fig. 18). The nucleoli of the neurons revealed moderate reactions at their peripheral zones forming thin rims around them. The nerve processes and nerve fibers of the body were moderately to strongly positive. The endothelial cells of the BV's reacted mildly. The moderately positive and beaded reaction was often found in the outermost layer of the perivascular sheath, but the other part of the sheath showed a negligible reaction. The nerve cells and their processes in the stalks reacted moderately. The ependymal cells were moderately active and most reaction products were found in their apical parts in the form of fine granules. The choroid plexus was very strongly stained.
I. Lactic Dehydrogenase
The Method o/ HEss et al. In the body, the neuropil and the oligodendroglia reacted moderately (Fig. 19) and the astrocytes were mildly active. Some nerve cells of the body showed strong activity, some were only moderate (Fig. 19), and others had mild activity. The nucleoli of the neurons and the nerve fibers were moderately stained. The endothelial cells of the BV's were strongly stained. The perivascular sheath showed a negligible reaction except for a moderately strong reaction in the outermost layer of this sheath {Fig. 20). In the stalks, the neuropil showed mild activity and the glial cells, nerve cells and nerve fibers revealed a moderate reaction. The ependymal cells were moderately active and the choroid plexus was very strongly stained.
BLOODY and ENGEL'S Method. The glial cells of the body showed moderate to strong activity for total LDH (Fig. 21). Some nerve cells of the body were moderately active and others revealed a strong reaction. The lipofuscin of the neurons seemed to be stained very strongly (Fig. 21). In the stalks, the glial cells reacted moderately, the nerve cells showed a mild or moderate reaction and the nerve fibers were strongly positive. As to the other elements, the reactions were the same as those detected by the method of HESS et al.
J. Succinic Dehydrogenase
The neuropil of the body was moderately active (Fig. 22). ' Ihe oligodendroglia were strongly positive and the astrocytes moderately positive. Some nerve cells of the body showed strong positive activity and the rest were moderate. The nucleoli of the neurons were mildly positive at their peripheral parts giving the appearance of thin rims. The neuronal processes and nerve fibers revealed moderate to streng staining. The reaction of the BV's was similar to that of AD. In the
23*
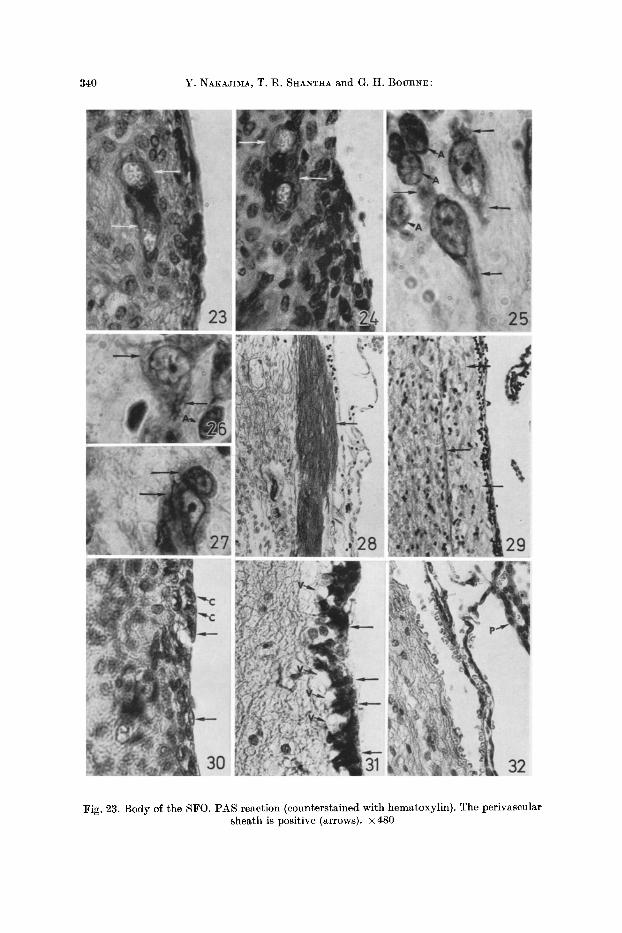
340 Y. NAKAJIMA, T. R. SHANTHA and G. H. BOURNE:
Fig. 23. Body of the SFO. PAS reaction (counterstained with hematoxylin). The perivascular sheath is positive (arrows). • 480

Studies on the Subfornical Organ 341
stalks, the glial cells and nerve cells were moderately positive. The nerve fibers of the stalks showed a moderate to strong reaction. The positive activity in them gave the appearance of beads. The ependymal cells showed moderate activity, which was restricted to their apical parts. The choroid plexus was very strongly stained (Fig. 22).
D i s c u s s i o n
The nature of the "parenchymal cells" in the SFO has been a subject of argument for a long time. AsIDA (1943) thought that these cells of the SFO were undifferentiated and that "genetically" they were to be regarded as nerve cells or at least that they arose from cells very similar to neuroblasts. SPRA:NKEL (1957) reported that the parenchymal cells of the SFO were related to the " typical" nerve cells by transitional forms of cells. On the other hand, WISLOCXI and LEDUC (1952) thought that they were derived from ependymal cells or medullary epithelium. These cells, though small to medium in size, show the presence of Nissl substance in their cytoplasm, suggesting a neuronal nature. The electron microscopical studies of ANDRES (1965) clearly show the neuronal nature of these cells with the typical Nissl substance which is similar to that found in other neurons.
AK]~RT et al. (1961) described the presence of nerve fibers of unknown origin and destination in the SFO. In our study, the dorsal stalk shows more myelinated fibers than the body and ventral stalk. The body and stalks of the SFO contain a number of unmyelinated fibers. As suggested by AK]~RT et al. (1961), it is pos- sible that some of these fibers in the dorsal stalk are derived from some adjoining septal nuclei. Our studies in the preparations stained with W~IL'S and BODIAN'S methods indicate that a number of fibers originate in the body of the SFO. The termination of these fibers is not known and further work is planned to study this problem.
The cells of the SFO have been shown to contain slight or negative SDH activity in various rodents (Lv, Duc and WISLOC~, 1952 ; SHImzu and MORIXAWA, 1957). On the contrary, NAKAJ1MA (1966) found mild AD, moderate SDH and
Fig. 24. Body of the SFO showing the PAS positive perivascular sheath after digestion by diastase (arrows). x480
Figs. 25, 26 and 27. Body of the SF0. The Nlssl substance is stained with cresy] violet. Note the fine Nissl granules in the cytoplasm of the nerve cells (arrows). The nuclei of the astrocytes
are also stained (arrows .4). • 1,125
Fig. 28. Dorsal stalk of the SFO. BODIA~'S method. Many nerve fibers are stained (arrow). • 150
Fig. 29. Dorsal stalk of the SFO showing a number of myelinated nerve fibers stained with W~IL'S method (arrows). X 150
Fig. 30. Body of the SF0. H&E staining. The ependymal cells covering the body are both squamous (arrows) and cuboidal (arrows C) in nature. • 480
Fig. 31. Ventral stalk of the SFO. H&E staining. Note the columnar ependymal cells (arrows) and the subependymal vacuoles below these lining cells (arrows V). • 480
Fig. 32. Dorsal stalk of the SFO. H&E staining. The ehoroid plexus (arrow P) attaches to the dorsal stalk, at which there is no ependymal lining. • 300
23b Histochemie, Bd. 13

342 Y. NXKAJ~, T. R. S H A ~ and G. H. BouR~:
cytochrome oxidase and strong G6PD and LDH activities in the rat SFO. In the present study, the nerve cells of the SFO, in addition to showing active enzymes, reveal variability in the enzyme reaction from cell to cell for any given enzyme. This clearly shows that the amount of enzyme varies from cell to cell and time to time depending on its functional state. I t is probable that the active cells con- rain more enzyme, whereas the resting cells contain less. This study indicates that at any given time some cells are functionally more active and others are in a state of rest. Contrary to the previous belief (L~,DUC and WISLOCKI, 1952; SHIMIZU and MORIKAWA, 1957) that the SFO derives its energy through anaerobic meta- bolism, the present work as well as NAKAJL~A'S (1966) study shows that it derives its energy by aerobic metabolism through the TCA cycle as do other neurons of the nervous system. I t is possible tha t these cells produce considerable amounts of ATP by the TCA cycle and NADPH 2 by the pcntose cycle which are subsequently used for the various activities of the cells.
The oligodcndroglia like the oligodendroglia in other parts of the nervous system and glial cells of the eye (SHANTHA, VEERAPPA et a~., 1966; SHA~THA et al., 1967 ; IIJ~MA et al., 1967a, b, c ; MA~OCHA et al., 1967) show a large amount of AP. I t is possible that these cells act as energy reservoirs and that the AP in the cells is involved in supplying glucose-l-phosphate to neurons by the breakdown of glycogen. A considerable amount of SDH in these oligodendroglla indicate tha t they are capable of aerobic metabolism.
I t is interesting to note that LV.DUC and WISLOCKI (1952) found AC located exclusively in the cytoplasm of parenchymal cells. In the present study, it was found to be widely distributed in several elements of the SFO. The ependymal cells and the choroid plexus show very high activity for enzymes of the glycolytie pathways (i.e., pentosc cycle, EI~I pathway and TCA cycle) as previously observed (MANoCHA et al., 1967 ; S ~ A ~ A and MA_~OCHA, 1968).
Only the outermost layer of the pcrivascular sheath shows enzymes of the glycolytic pathways (i.e., enzymes of the TCA cycle, EM pathway, pentose cycle), whereas the inner layers of this sheath are almost flee of these enzymes. Similar findings have been reported on the enzymes of the glycolytic pathways in the perivascular sheath of the area postrema which also belongs to the paraventricular structures (IIJrMA et al., 1967). We suggest that this outer element of the peri- vascular sheath is also rich in ATPase and may mediate the function of active transport between BV's and neurons as well as between BV's and glial cells. The amylophosphorylasc (AP) found in the perivascular sheath may be correlated with the breakdown of glycogen into glucose-l-phosphate which is then fed onto the EM pathway with the consequent production of ATP.
ATHERTOI~T (1963) reported the absence of cholinesterase in the SFO of the chick, but we found positive activity for AChE in the nerve cells of the SFO of the squirrel monkey and also found that the nerve fibers in the stalks were moderately positive for this enzyme. In addition, they show a variable degree of (mild to strong) MAO activity, and the stalks contain many positive nerve fibers. A~DRES' (1965) electron microscopic studies on the SFO of the dog showed that it contained neurons with axodendritic and axosomatic synapses, so AChE is presumably involved in the synaptic transmission of impulses at these sites. An inverse relationship between AChE and MAO in the central nervous system (CNS)

Studies on the Subfornieal Organ 343
has been suggested (SHANTHAVEERAPPA and BOURNE, 1965; MANOCHA eta/., 1967; SHANTHA et a[., 1967). The present study indicates that the SFO of the squirrel monkey contains both cholinergic and adrenergic neurons, as well as both types of fibers. Non-specific cholinesterase (BuChE) was found to be present in some nerve cells of the CNS as well as in the BV's and glial cells (Ko•LLV., 1954). This enzyme has been described as a "neurohumoral scavenger" (KOELLE, 1950; SHANTHAVEERAPPA and BOURNE, 1965) and does play any important or direct role in the process of the transmission of impulses (KoELLE, 1954). I t is surprising that the BV's of the SFO do not show any BuChE activity, instead, the neurons show the presence of this enzyme. In neurons, BuChE may hydrolyze the acetyl- choline which has escaped hydrolysis by ACHE. The ependymal cells and the choroid plexus show BuChE activity. I t is possible that this enzyme in these structures is concerned with the permeability as well as the destruction of acetyl- choline which escapes from the CNS into the cerebrospinal fluid and blood without hydrolysis by ACHE.
The nucleoli of the neurons of the SFO show mild reactions for AC, AP and SDH, moderate reactions for G6PD, LDH, AD and MA0, and a very strong reaction for ATPase. Siml]ar findings have been reported on the neurons of the SFO of the rat (NAKAJ~A, 1966), and on some other neurons of the CNS (KuMA- GAI, 1927; IIJIMA and NAKAJIMA, 1964; N~gA.UMA, 1965; SHANTHA et a~., 1967; IIJIMA et al., 1967). As the nucleolus is actively involved in the synthesis of proteins, it is possible that some of these enzymes probably take part in protein synthesis and the subsequent liberation of protein into the cytoplasm.
The moderate or strong staining of the nucleus of the neuron for ATPase tests may be an artifact resulting from the deposition of reaction products (i.e., metallic and nonmetallic in character) on the nucleus. Also the staining of llpofuscin granules and myelin sheaths, which is observed when phenazine methosulfate is used as a flavoprotein substitute for the demonstration of G6PD and LDH, may be an artifact caused by the deposition of formazan onlipid (BRoDY and ENG~L, 1964).
Our present study throws no light on the problem of the function of the SF0. Several hypothetical functions, such as chemoreceptor for cerebrospinal fluid, neurosecretion, the regulation of quantity and quality of cerebrospinal fluid, etc., have been given (YAMADA and HASUNUMA, 1955; HASU~UMA, 1956; DIm~CKX, 1963 ; PACttOMOV, 1963 ; SPOE~P~, 1963 ; ANDRv.S, 1965). The present histochemical studies indicate that the SFO is very active enzymatically and appears to be capable of performing any one or all of the above suggested functions. Further experimental studies are needed to pinpoint the exact function performed by the SFO.
Aclcrwwledgment. The present study was supported by Grant No. FR-00165 from the U.S. Public Health Service, Anlmal Resources Branch and Grant No. MGR-11-001-016 from NASA.
References ABE, T., and N. SH~ZU: Histochemieal method for demonstrating aldolase. Histochemie 4,
209---212 (1964). - - y . yAMAUA, P. H. I:~ASHIMOTO, and N. Snv~zu: Histochemical study of glucose-6-phos-
phate dehydrogenase in the brain of normal adult rat. Med. J. Osaka Univ. 14, 67--98 (1963).

344 Y. NAWA.rmA, T. R. S~ANT~A and G. H. BOURNE:
AKERT, K., H. D. POTTER, and J. W. ANDERSON: The subfornical organ in mammals. I. Com- parative and topographical anatomy. J. comp. Neurol. 116, 1--14 (1961).
A~DRES, K. H. : Der Feinbau des Subfornikalorganes vom Hund. Z. Zellforseh. 68, 445---473 (1965).
ASIDA, K. : Beitr~ge zur Kenntnis der Morphologie und Entwicklungsgeschichte des sub- fornikalen Organs. Keijo J. IVied. 12, 45--94 (1943).
ATHERTON, G.W.: An investigation of the specificity of cholinesterase in the developing brain of the chick. Histochemie 3, 214---221 (1963).
BODIAN, D. : A new method of staining nerve fibers and nerve endings in mounted paraffin sections. Anat. Rec. 65, 89--97 (1936).
BLOODY, I.A., and W. K. ENGEL: Isozyme histochemistry: The display of selective lactate dehydrogenase isozymes in sections of skeletal muscle. J. Histochem. Cytochem. 12, 6 8 7 4 9 5 (1964).
BURSTO~E, M. S. : Enzyme histochemistry and its application in the study of neoplasm. New York and London: Academic Press 1962.
COUrLAND, R. E., and R. L. HOI~ES: The use of cholinesterase techniques for the demon- stration of peripheral nervous structures. Quart. J. micr. Sci. 98, 327--330 (1957).
DIERICKX, K. : The subfornical organ, a specialized osmorcceptor. Naturwissenschaften 50, 163--164 (1963).
GLENNER, G. G., H. J. BURT~ER, and G. W. BROWN: The histochemical demonstration of monoamine oxidase activity by tetrazolinm salts. J. Histochem. Cytochem. 5, 591--600 (1957).
HXSUNUM_A, S.: Comparative anatomical studies on the subfornical organ (intereolumnar tubercle) of the mammals and man. Bull. Tokyo reed. dent. Univ. 8, 159--170 (1956).
HEss, R., D. G. SCARPF_Z~I, and A. G. E. P~,ARSE : The cytochemical localization of oxidative enzymes. II. Pyridine nucleotide-linked dehydrogenases. J. biophys, biochem. Cytol. 4, 753--760 (1958).
HOFER, H. : Die circumventrikul~iren 0rgane des Zwischenhirns. Primatologia, Bd. II/2, S. 13. Basel and New York: S. Karger 1965.
IIJn~A, K., G. H. BOURNE, and T. R. S~AN~A: Histoehemical studies on the distribution of enzymes of glycolytic pathways in the area postrema of the Squirrel Monkey. Acta histochem. (Jena) 27, 42---54 (1967a).
- - , and Y. NAKAJ--~A: Histechemical studies on the oxidative enzymes of the human cerebel- lum. Bull. Tokyo reed. dent. Univ. 11, 103--135 (1964).
- - T. R. S~ANTHA, and G. H. BOUR~E: Enzyme-histochemical studies on the hypothalamns with special reference to the supraoptic and paraventricular nuclei of Squirrel Monkey (Saimiri sciureus). Z. Zellforsch. 79, 76--91 (1967b).
- - - - - - Histochemical studies on the distribution of some enzymes of the glycolytie path- ways in the olfactory bulb of the Squirrel Monkey (Saimiri sciureus). Histochemie 10, 224--229 (1967 c).
KOELL~, G. B. : The histochemical differentiation of types of cholinesterases and their localiza- tion in tissues of the cat. J. Pharmacol. exp. Ther. 109, 158--179 (1950).
- - The histochemical localization of cholinesterases in the central nervous system of the rat. J. comp. Neurol. 10O, 211--235 (1954).
KUMAGAI, K. : Beitr~ge zur Kenntnis der Indophenoloxidase-Reaktion des Nervensystems. Okayama Igakkai Zasshi 447, 446---454 (1927).
L]~DUC, E.H., and G. B. WISLOCKI: The histochemical localization of acid and alkaline phosphatases, non-specific esterase and succinic dehydrogenase in the structures comprising the hematoencephalic barrier of the rat. J. comp. Neurol. 97, 241--280 (1952).
1VIANoCHA, S.L., T. R. S~ANTm~, and G. H. BOURNE: Histochemical studies on the spinal cord of the Squirrel Monkey (Saimiri sciureus). Exp. Brain Res. 3, 25~39 (1967).
MATSUNA~, T. : Histochemical study of lactic dehydrogenase in the brain (Japanese). Osaka Daigaku Igaku Zasshi l l , 3617--3631 (1959).
McMAm~s, J. F. A. : Histological demonstration of mucin after periodic acid. Nature (Lond.) 158, 202 (1946).

Studies on the Subfornical Organ 345
NACItLAS, M. ~[., K. C. Tsou, E. DE SOUZA, C. S. CHENG, and A. M. SEUGMAN: Cytochemical demonstration of succinic dehydrogenase by the use of a new p-nitrophenyl substituted ditetrazole. J . Histoehem. Cytochem. 5, 420---436 (1957).
NAKAJIMA, Y. : Histochemieal studies on the distribution of amylophosphorylase in the sub- fornieal organ of the rat. Bull. Tokyo med. dent. Univ. 11, 391--402 (1964).
- - Histochemical studies on carbohydrate metabolism of the mesencephalie nucleus of the trigeminal nerve in the rat. Bull. Tokyo med. dent. Univ. 12, 265--282 (1965).
- - Histochemical studies of the carbohydrate metabolism of the subfornical organ in the rat. Bull. Tokyo med. dent. Univ. 13, 125--146 (1966).
PACHOMOV, N. : Morphologische Untersuchung zur Frage der Funktion des subfornikalen Organs der Ratte. Dtsch. Z. Nervenheilk. 185, 13--19 (1963).
PADYKULA, H. A., and E. HERMAN: Factors affecting the activity of adenosine triphosphatase and other phosphatases measured by histochemieal techniques. J . Histoehem. Cytochem. 3, 161--169 (1955).
- - - - The specificity of the histoehemical method for adenosine triphosphatase. J . Histochem. Cytochem. 8, 170--195 (1955).
S~ANTHA, T. R., K. IIJ~Lr and G. H. BOURNE: Histochemieal studies on the cerebellum of Squirrel Monkey (Saimiri sciureus). Acta histochem. (Jena) 27, 129--162 (1967).
- - S. L. MANOCHA: Enzyme histochemistry of the choroid plexus in rat and squirrel monkey. Histochemie, in Press (1968).
SHANTHAVEER~ePA, T. R., and G. H. BOURNV,: Histoehemical studies on the distribution of dephosphorylating and oxidative enzymes and esterases in the olfactory bulb of the squirrel monkey. J . nat. Cancer Inst. 35, 153--165 (1965).
- - M. B. WArrZ~AN, and G. H. BOURNn: Studies on the distribution of phosphorylase in eyes of the rabbit and the squirrel monkey. Histoehemie 7, 80--95 {1966).
SHIMIZlY, N., and N. MOR~WA: Histochemical studies of sueeinic dehydrogenase of the brains of mice, rats, guinea pigs and rabbits. J . Histochem. Cytoehem. 5, 334---345 (1957).
, and Y. ISHI: Histoehemical studies of suecinic dehydrogenase and cytochrome oxidase of the rabbit brain, with special reference to the results in the paraventricular structures. J . eomp. Neurol. 108, 1---21 (1957).
- - , and M. OKADA : Histochemical distribution of phosphorylase in rodent brain from newborn to adults. J . Histochem. Cytochem. 6, 459--471 (1957).
SPOERRI, O. : 12~ber die Gef~versorgung des Subfornikalorgans der Ratte. Acta anat. (Basel) 54, 333--348 (1963).
SPRANKEL, H. : Zur Zytologie des subfornikalen Organes bei Affen. Verb. dtsch, zool. Ges., Graz 4, 44 51 (1957).
- - Uber die Beziehung des Plexus des dritten Ventrikels zum subfornikalen Organ bei den Primaten. Naturwissenschaften 47, 383--384 (1960).
TAKE~C~, T., and H. KmCTA~T: Histochemieal detection of phosphorylase in animal tissues. J . Histochem. Cytochem. 3, 153--160 (1955).
WACHST]~IN, M., and E. M~m~L: Histochemistry of hepatic phosphatases at a physiologic pH with special reference to the demonstration of bile canaliculi. Amer. J . clin. Path. 27, 13--23 (1957).
WEre, A.: Textbook of neuropathology. New York: Grune & Stratton 1945. WISv.OCKI, G. B., and E. H. L~DUC: Vital staining of the hematoencephalic barrier by silver
nitrate and trypan blue, and cytological comparison of the neurohypophysis, pineal body, area postrema, intercolumnar tubercle and supraoptic crest. J . comp. Neurol. 96, 3 7 1 ~ 1 3 (1952).
YAMADA, H., and S. HASUN~w~: Finer structure of the subfornical organ (intereolumnar tubercle) of the dog. Bull. Tokyo med. dent. Univ. 2, 67--76 (1955).
Dr. T. R. S ~ A Emory University Yerkes Regional Primate Research Center Atlanta, Georgia 30322 U.S.A.