hepatic metabolism of n-hydroxy-n-methyl-4...
TRANSCRIPT

[CANCER RESEARCH 36, 2350-2359, July 1976]
Hepatic Metabolism of N-Hydroxy-N-methyl-4-aminoazobenzene and Other N-Hydroxy Arylamines to Reactive Sulfuric Acid Esters
F r e d F. K a d l u b a r , 2 J a m e s A . M i l l e r , a n d E l i z a b e t h C . M i l l e r
McArd/e Laboratory for Cancer Research, University of Wisconsin Medica/ Center, Madison, Wisconsin 53706
S U M M A R Y
Hepatic cytosols catalyzed a 3'-phosphoadenosine 5'- phosphosulfate (PAPS)-dependent O-sulfonation of N-hy- droxy-N-methyl-4-aminoazobenzene (N-HO-MAB) and ot several other N-hydroxy arylamines. The presumed product from N-HO-MAB, N-methyl-4-aminoazobenzene-N-sulfate, reacted with added guanosine to yield N-(guanosin-8-yl)-N- methyl-4-aminoazobenzene, with methionine to form a sul- fonium derivative that decomposed to yield 3-methylmer- capto-N-methyl-4-aminoazobenzene, and with ribosomal RNA to give a bound derivative. N-Methyl-4-aminoazoben-
zene was converted to N-(guanosin-8-yl)-N-methyl-4-ami- noazobenzene in concerted N-oxidation and O-sulfonation reactions conducted aerobically with a fortified 10,000 • g rat liver supernatant. In the absence of an added nucleo- phile, metabolically formed N-methyl-4-aminoazobenzene- N-sulfate (or the nitrenium ion from this unstable ester) was reduced by N-HO-MAB to form N-methyl-4-aminoazoben- zene; the N-HO-MAB was oxidized, probably through a ni- trone intermediate, to yield products that included N-hy- droxy-4-aminoazobenzene and formaldehyde. An analo- gous reaction .was noted between N-benzoyloxy-N-methyl- 4-aminoazobenzene and N-HO-MAB in the absence of cytosol and PAPS.
Hepatic N-HO-MAB sulfotransferase activities were in the order: male rat > female rat, male rabbit, male guinea pig, male mouse > male hamster. Male'rat kidney and small intestine cytosols had low activities; the other tissues stud- ied had little or no activity. Hepatic sulfotransfera'~e activi- ties for N-HO-MAB and N-hydroxy-N-acetyl-2-aminoflu- orene displayed different pH optima and inhibitor and acti- vator responses.
The rates of PAPS-dependent rat liver cytosol-catalyzed esterification of N-hydroxy-N-ethyl-4-aminoazobenzene, N- hydroxy-4-aminoazobenzene, and N-hydroxy-1- and 2- naphthylamine were 20 to 50% of that for N-HO-MAB. Ac- tivities for t r a n s - N - h y d r o x y - 4 - a m i n o s t i l b e n e , N-hydroxy-2- aminofluorene, N-hydroxyanil ine, and N-hydroxy-N-methyl- N-benzylamine were not detected.
No microsomal reduced nicotinamide adenine dinucleo- t ide-dependent reduction or reduced nicotinamide adenine dinucleotide phosphate-dependent oxidation or cytosolic transferase reactions for N-HO-MAB, except the above-de-
1 This research was supported in part by funds from USPHS Grants CA- 07175 and CA-15785.
2 Postdoctoral Fellow of the Foundation for Cancer Research. Chicago. Received January 26, 1976; accepted April 7, 1976.
scribed PAPS-dependent reaction, were detected in rat liver.
I N T R O D U C T I O N
The metabolic N-oxidation of carcinogenic arylam- ines and arylamides appears to be a necessary step in their conversion to ultimate carcinogenic derivatives (48). Two hepatic microsomal enzymes, a cytochrome P-450-depend- ent oxidase (21, 33, 69, 73) and a flavoprotein amine oxi- dase (33, 77, 78), have been implicated as catalysts for these oxidations. The N-hydroxy amines and N-hydroxy amides are then subject to further hepatic metabolism. A NADH-de- pendent microsomal N-hydroxy amine reductase and a NADPH- and oxygen-dependent microsomal N-hydroxy amine oxidase catalyze the reduction and oxidation, re- spectively, of a number of N-hydroxy amines (32, 34, 53). A microsomal deacetylase for N-HO-AAF 3 has been demon- strated in the livers of a number of species (26).
Hepatic cytosols from rats and some other species contain N-hydroxy arylacylamide-dependent N-hydroxy arylamine acyltransferase (6, 7, 36), ATP-dependent seryl- transferase (66, 67), and PAPS-dependent N-hydroxy amide sulfotransferase (16, 37, 64) activities. These enzymes catalyze the conversion of certain N-hydroxy arylamines and N-hydroxy arylamides to their reactive acetic acid, serine, and sulfuric acid esters, respectively. The highly reactive and mutagenic sulfuric acid ester of N-HO-AAF is strongly implicated as an ultimate carcinogenic metabolite of AAF in rat liver (16, 17, 23, 41, 74). Similarly, reactive ester metabolites of other N-hydroxy compounds have been considered as ultimate carcinogens (48). The NADPH- dependent reduction of N-hydroxy-2-naphthylamine (52) and an S-adenosylmethionine-dependent O-methylation of N-HO-AAF (39) by hepatic cytosols have also been reported.
This communication reports the PAPS-dependent esteri- fication of N-HO-MAB, a metabolite of the hepatocarcino- gen MAB (33), and of several other carcinogenic and non- carcinogenic N-hydroxy arylamines. Some enzymatic prop-
3 The abbreviations used are: N-HO-AAF, N-hydroxy-N-acetyl-2-aminoflu- orene; PAPS, 3'-phosphoadenosine 5'-phosphosulfate; AAF, N-acetyl-2-ami- nofluorene; N-HO-MAB, N-hydroxy-N-methyl-4-aminoazobenzene; MAB, N- methyl-4-aminoazobenzene; Bis-Tris, bis(2-hydroxyethyl)-Tris; PAP, 3'-phos- phoadenosine 5'-phosphate; N-HO-AB, N-hydroxy-4-aminoazobenzene; N- HO-EAB, N-hydroxy-N-ethyl-4-aminoazobenzene; N-(guanosin-8-yl)-MAB, N- (guanosin-8-yl)-N-methyl-4-aminoazobenzene; 3-CHsS-MAB, 3-methylmer- capto-N-methyl-4-aminoazobenzene; MAB-N-sulfate, N-methyl-4-aminoazo- benzene-N-sulfate; N-benzoyloxy-MAB, N-benzoyloxy-N-methyl-4-amino- azobenzene.
2350 CANCER RESEARCH VOL. 36
Research. on June 26, 2018. © 1976 American Association for Cancercancerres.aacrjournals.org Downloaded from

erties of the sulfotransferase(s) and the chemical reactivi- ties of the metabolically formed esters were examined.
MATERIALS AND METHODS
Materials. NAD § NADP +, glucose 6-pl~osphate, Bis-Tris, ATP, 3',5'-ADP(PAP), p-nitrophenol, o-iodosobenzoate, S- adenosylmethionine iodide, S-acetyI-CoA, catalase (type C-40), and serum albumin (type V) were purchased from the Sigma Chemical Company (St. Louis, Mo.). NADPH and NADH were obtained from P-L Biochemicals, Inc. (Milwau- kee, Wis.). Glucose-6-phosphate dehydrogenase (Code ZF) was purchased from Worthington Biochemicals (Freehold, N. J.). rRNA and PAPS were prepared from rat liver by the methods of Irving and Veazey (28) and Irving et al. (27), respectively. Radioactive materials were obtained as fol- lows: [8-14C]guanosine, Schwarz/Mann (Orangeburg, N. Y.); [8J4C]adenosine, Amersham/Searle Corp. (Arlington Heights, II1.); [2,6-14C]uridine, Nuclear Dynamics (El Monte, Calif.); and [2,6-~4C]thymidine, New England Nuclear (Bos- ton, Mass.). [G-3H]-N-Hydroxy-2-naphthylamine (110 mCi/ mmole) was prepared from [G-3H]-2-nitronaphthalene (Amersham/Searle custom tritiation) (75). [G-'3H]-N-HO - MAB was synthesized as described previously (33).
The following amine derivatives were synthesized by pub- lished procedures: N-hydroxy-N-methyI-N-benzylamine (20), N-hydroxyaniline (35), N-HO-AB (56), N-HO-MAB and N-HO-EAB (33), N-hydroxy-2-aminofluorene (40), N-HO- AAF (50), trans-N-hydroxy-4-aminostilbene (1), N-hydroxy- 4-aminobiphe~yl (42), N-hydroxy-1- and 2-naphthylamine (75), 2-amino-l-naphthylsulfate (13), 2,2'-azoxynaphthalene (43), MAB (46), 3-methylmercapto-4-aminoazobenzene and its N-methyl derivative (57), and 3-methylmercapto-4-ami- nobiphenyl (16). 3-Methylmercapto-N-ethyl-4-aminoazo- benzene was synthesized by the procedure described for the synthesis of its N-methyl analog (57); after purification by chromatography on alumina and crystallization (m.p. 38~ it eluted as a single peak on gas-liquid chromatogra- phy and had the expected mass spectrum (M + = 271). Analyses for each element were within 0.2% of theory (Huff- man Laboratories, Wheatridge, Colo.). 3-Methylmercapto- 2-aminofluorene and 3-methylmercapto-N-acetyl-2-amino- fluorene were generously supplied by Dr. To L. Fletcher (The Fred Hutchinson Cancer Research Center, Seattle, Wash.).
Microsomal and cytosolic cell fractions were obtained by differential centrifugation according to the principles out- lined by Hogeboom et al. (25). Livers were perfused in situ with 0.9% NaCI solution prior to homogenization. Adult animals were obtained as follows: CD random-bred rats and CD-t mice, Charles River Breeding Laboratories (Wilming- ton, Mass); rabbits, Sand Valley Farms (Spring Green, Wis.); guinea pigs, O'Brien Co. (Madison, Wis.); and ham- sters (Con Olson Co. (Madison, Wis.). The rats, mice, and hamsters were fed Wayne Breeder Blox pellets (Allied Mills, Inc., Chicago, II1.); the rabbits and guinea pigs were given Teklad Guinea Pig Diet (Teklad Mills, Winfield, Iowa).
Enzyme Assays. The enzyme-catalyzed reactions of N- HO-AAF, N-HO-MAB, and other N-hydroxy arylamines were measured by monitoring the loss of these substrates from the assay media. Each N-hydroxy compound was dissolved
N-Hydroxy Arylamine Sulfotransferase
or diluted in absolute ethanol to a concentration of 25 mM and was added to the enzyme incubation mixture at a final concentration of 0.5 mM. The oxidative decomposition of the N-hydroxy arylamines in the reaction media was mini- mized by the inclusion of 0.5 mM EDTA which prevented metal-catalyzed autoxidation (31) and/or lipid peroxidation (29). This low level of EDTA stabilized the N-hydroxy deriva- tives even in assay media that required high concentrations of Mg 2+. Assays that did not require oxygen were carried out in an argon atmosphere to minimize further nonenzy- matic oxidation of these compounds. Unless otherwise indi- cated, hepatic cell fractions from adult male rats (age, 4 to 6 months) were used as enzyme sources.
Microsomal N-hydroxy amine oxidase and reductase assays were carried out as previously reported (32). Unless otherwise indicated, the assay media for measuring cyto- solic S-adenosylmethionine-dependent O-methylase (39), N-hydroxy arylacetamide-dependent acetyltransferase (7), ATP-dependent seryltransferase (66), and NADPH-depend- ent N-hydroxy arylamine reductase (52) were used as described in the references.
Attempts to detect acetyI-CoA-dependent metabolism in N-HO-MAB were carried out in assay media that contained 100 mM Tris-HCI buffer (pH 7.3 at 37~ 0.5 mM EDTA, 1 mM acetyI-CoA, 20 to 100 mM methionine, 3 mg cytosol protein per ml, and 0.5 mM N-HO-MAB.
The assay medium that was found to be optimal for hepa- tic N-hydroxy arylamine sulfotransferase activity contained 100 mM Bis-Tris-HCI buffer (pH 6.2 at 37~ 5 mM MgCI.,, 0.5 mM EDTA, 1 mM PAPS, 0.5 to 5.0 mg cytosol protein per ml, and the specified N-hydroxy amine (0.5 mM). The assay medium that was optimal for N-HO-AAF sulfotransferase contained 100 mM Bis-Tris-HCI buffer (pH 6.6 at 37~ 5 mM MgCI2, 0.5 mM EDTA, 0.65 mM PAPS, 0.2 to 1.0 mg cytosol protein per ml, and N-HO-AAF (0.5 mM). When compared directly with the assay medium described by DeBaun et al. (16), the above medium yielded 10 to 20% higher activi- ties. L-Methionine (100 mM for N-hydroxy amine assays; 20 mM for N-HO-AAF assays), [8-14C]guanosine (1 mM, 4 mCi/ mmole), or rRNA (6.7 mg/ml) was routinely added to the sulfotransferase assays to trap the enzymatically generated sulfuric acid esters.
Each of the above assays was carried out in the presence or absence of the coenzymes and with intact or heat- denatured (85 ~ for 3 min) cell fractions so that coenzyme- and enzyme-dependent metabolism could be ascertained.
Chemical Assays. The concentrations of the N-hydroxy amines in aliquots of the incubation mixtures were deter- mined colorimetrically as n-amyl acetate-extractable Fe 3+- reducing equivalents. Aliquots (0.5 ml) were agitated on a vortex mixer with 3 ml of water-saturated n-amyl acetate, and the phases were separated by centrifugation. The con- centrations of the N-hydroxy secondary amines (N-HO- MAB, N-HO-EAB, N-hydroxy-N-methyI-N-benzylamine)in the amyl acetate extracts were determined by a method described for the estimation of "lipid-soluble" N-hydroxy amines (32), except that, in order to stabilize the chromo- phore, 0.05 ml of 1.7 M acetic acid was added after the specified addition of EDTA.
The concentrations of N-hydroxy primary arylamines and
JULY 1976 2351
Research. on June 26, 2018. © 1976 American Association for Cancercancerres.aacrjournals.org Downloaded from

F. F. Kad lubar et al.
phenolic amines in the amyl acetate extracts were mea- sured by a modif icat ion of a method developed for the estimation of tocopherols as Fe3§ equivalents (72), in which 0.2 ml of the amyl acetate extract was mixed with a solution that consisted of 0.1 ml of 0.4 M sodium acetate:0.6 M acetic acid, 0.6 ml of 95% ethanol, 0.2 ml of 4,7-diphenyl- l ,10-phenanthrol ine [10 mg/ml in anhydrous amyl acetate:absolute ethanol (15:1)], and 0.04 ml of 0.01 M Fe(NO~),~ :0 .1 M acetic acid; after 1 min the reaction was terminated by addit ion of 0.04 ml of 0.02 M H:~PO4. The absorbance at 535 nm was 39,200 (2 equivalents of Fe 3+ reduced per equivalent of N- or o-hydroxy amine) for each of the fol lowing compounds: N-HO-AB, N-hydroxyani l ine, N-hydroxy-1- and 2-naphthylamine, N-hydroxy-4-aminobi- phenyl, N-hydroxy-2-aminof luorene, N-hydroxy-4-amino- sti lbene, 2-amino- l -naphthol , and o- and p-aminophenol . If reducing compounds are formed from a N-hydroxy amine substrate dur ing an enzyme-catalyzed reaction, this assay can provide only a min imum estimate of the amount of N- hydroxy amine metabolized.
N-HO-AAF was determined spectrophotometr ical ly as its ferric chelate as described by Booth and Boyland (9). Pro- tein was determined by the biuret reaction (22).
Reactivity Studies and Product Identification. When me- th ionine was used to trap reactive metabolites, the methi- onyl derivatives of the aminoazo dyes were decomposed and analyzed as 3-methylmercapto derivatives as described by Poirier et ah (51), except that the yield of the methylmer- capto compounds was increased 30 to 50% by decomposi- tion of the sul fonium adduct in alkali at 90 ~ for 45 min. The identif ication of 3-CH:3S-MAB (M + = 257) from N-HO-MAB sulfotransferase reaction mixtures was conf irmed by gas chromatography-mass spectrometry. The sul fonium deriva- tive of 4-aminobiphenyl was decomposed and analyzed as 3-methylmercapto-4-aminobiphenyl by gas-l iquid chroma- tography as previously described (7).
The format ion of [8-1"C]-N-(guanosin-8-yl)-MAB in N-HO- MAB sulfotransferase assays that contained [8-1"C]guan - osine was measured by thin-layer chromatography (Ref. 38, Solvents A to D) and l iquid scint i l lat ion spectrophotom- etry. Simi lar incubation mixtures that contained other [14C]nucleosides were chromatographed by the procedures of Poirier et al. (51). When the N-hydroxy amine sulfo- transferase assays contained rRNA, the recovery and esti- mation of RNA-bound substrate were carried out according to the method of Wislocki et ah (76).
For structural identi f icat ion the products formed dur ing the PAPS-dependent esterif ication of N-HO-MAB were iso- lated by extraction 3 times with 2 volumes of ethyl acetate, concentrat ion of the extract under reduced pressure, and thin-layer chromatography of the concentrate in Solvents A, B, and F (33). After elution the products were analyzed by mass spectrometry. Quantitat ion of these products was achieved with the use of [3H]-N-HO-MAB (503 mCi /mmole) as fol lows. A 0.5-ml al iquot of the incubation medium was mixed with an equal volume of 95% ethanol that contained carrier dyes (0.01 M MAB, N-HO-MAB, and N-HO-AB); after centr i fugat ion 2 /~1 were analyzed as previously described (33) by thin- layer chromatography in Solvents A, B, and F and l iquid scint i l lat ion spectrophotometry. Formaldehyde
was measured by the method of Nash (49), except that prior extraction with chloroform was required to remove dye derivatives; semicarbazide (1 mM) was included in the reac- tion mixture to obtain quantitative recovery of the formalde- hyde.
Products from the [G-3H]-N-hydroxy-2-naphthylamine (110 mCi/mmole) sulfotransferase reaction were isolated similarly by thin-layer chromatography on si l ica gel (East- man 6060 sheets) with 1-butanol: l -propanol:0.1 M NH:~ (2:1:1) and 1-butanol:acetic acid:H.,O (10:1:1) [Solvents (a) and (m) of Boyland and Manson (11)] and wi th Solvents A and F (33).
Instrumentation. Mass spectra were obtained with a Var- ian CH-7 mass spectrometer (Varian Associates, Palo Alto, Calif.) equipped with a mass marker and a magnet-driven direct insertion probe (Variset Corp., Madison, Wis.). Elec- tronic spectra were measured with a Zeiss PMQ II or Beck- man DB spectrophotometer. 3H was determined in Scintisol (Isolab, Inc., Akron, Ohio) with a Packard Tri-Carb scintil la- tion spectrometer. Gas chromatography was performed with a Barber-Colman Model 10 chromatograph equipped with a 0.6- x 120-cm column packed with 3% OV-1 on Chromsorb W/HP (Pierce Chemical Co., Rockford, III.).
RESULTS
Microsomal N-Hydroxy Arylamine Metabolism
In view of the metabolic formation of N-hydroxy amines by the endoplasmic reticulum of liver cells, the ability of male rat liver microsomes to metabolize certain N-hydroxy arylamines was examined. Under the assay condi t ions used (cf. "Materials and Methods"), the NADH-dependent reduc- tion of N-hydroxy-N-methyI -N-benzy lamine and N-hydroxy- anil ine occurred at rates of 8.6 -+ 0.8 and 2.1 +-- 0.5 nmoles/ min/mg microsomal protein (n = 3). On the basis of analy- ses for extractable reducing equivalents, no NADH-depend- ent loss of N-HO-MAB or N-HO-EAB was observed, and these compounds were recovered quantitat ively from the assay medium with incubation periods of up to 6 min. Incubation for longer times resulted in signif icant losses, but similar losses occurred with heat-denatured micro- somes. Similarly, 93 to 98% of the N-hydroxy derivatives of 4-aminoazobenzene, 4-aminobiphenyl, 2-aminofluorene, 4- aminosti lbene, and 1- and 2-naphthylamine were recovered as extractable reducing equivalents after 6-min incubations with NADH-fortif ied microsomes. With fort i f ied heat-dena- tured microsomes only 20 to 50% of each N-hydroxy pri- mary arylamine was recovered under the same conditions. The much higher recoveries of these compounds from incu- bations with fresh microsomes may be due to a reduced pyridine nucleot ide-dependent enzymatic reduction of aerobic oxidation products (nitroso derivatives?) of the N-hydroxy amines (54).
Under the standard condit ions (pH 7.6 in air, cf. "Mate- rials and Methods") for measuring microsomal NADPH-de- pendent N-hydroxy amine oxidase activity, the N-hydroxy aminoazo dyes oxidized rapidly (20 to 50 nmoles /min /ml of reaction mixture) even in the absence of NADPH and/or
2352 CANCER RESEARCH VOL. 36
Research. on June 26, 2018. © 1976 American Association for Cancercancerres.aacrjournals.org Downloaded from
microsomes. Thus, if an NADPH-dependent enzymatic oxi- dation occurred, it could not be detected under these con- ditions. The rapid air oxidation of N-hydroxy amines under aerobic condit ions (pH > 7) has been studied (31,33).
Cytosolic N-Hydroxy Arylamine Metabolism
In preliminary experiments with liver cytosol from male rats no coenzyme-dependent loss of N-HO-MAB or N-HO- EAB was detected (<0.1 nmole/min/mg cytosol protein) using assay media described (cf. "Materials and Methods") for N-hydroxyaryl acetamide- or acetyI-CoA-dependent O- acetylation, ATP-dependent O-serylation, S-adenosylme- thionine-dependent O-methylation, or NADPH-dependent re- duction of certain N-hydroxy amines or N-hydroxy amides. Furthermore, the formation of a methionyl-dye adduct was not detected (<0.01 nmole/min/mg protein) in methionine- supplemented O-acetylation or O-serylation incubation mix- tures. Attempts to detect these latter activities at other pH's (6.2, 6.6, 7.0), with substrate concentrations up to 0.5 mM, or with protein concentrations of 1 to 5 mg/ml were unsuc- cessful. However, PAPS-dependent losses of N-HO-MAB of greater than 10% occurred under the assay conditions used to demonstrate the formation of the sulfuric acid ester of N- Q HO-AAF (16), and 3-CH:}S-MAB was recovered when methio- nine was included in the incubation medium. Similar results were obtained with several other N-hydroxy arylamines. These experiments suggested that rat hepatic cytosol con- tains enzyme(s) that catalyze the PAPS-dependent esterifi- cation of N-hydroxy amines, and the properties of this enzy- matic activity are described in the fol lowing section.
N-Hydroxy Amine Sulfotransferase Activity
Assay Requirements and Comparisons with N-HO-AAF Sul- fotransferase Activity
The loss of N-HO-MAB and formation of 3-CH.~S-MAB from methionine-supplemented incubation mixtures were dependent on the presence of PAPS and cytosol (Table 1).
Table 1 Rat hepatic N-HO-MAB sulfotransferase: assay requirements
Relative rates"
Formation Loss of N- of 3-CH:~S-
Incubation mixture HO-MAB MAB
Complete system 100 100 With heat-denatured cytosol 5 <1 - PAPS 8 <1 - PAPS + PAP (1 mM) 10 <1 - PAPS + ATP (1 mM) 12 <1 - Methionine 101 <1 + Serum albumin (5 mg/ml) 101 59
" Assays were carried out as described in "Materials and Meth- ods" with 5 to 10 different preparations of hepatic cytosol from male rats; the rates are expressed relative to that of the complete system. The mean N-HO-MAB sulfotransferase activity of the com- plete system was 4.7 + 1.6 nmoles N-HO-MAB metabolized per min per mg protein (n = 10). The formation of 3-CH:~S-MAB by the complete system averaged 0.61 - 0.05 nmole/min/mg protein (n = 10).
N-Hydroxy Arylamine Sulfotransferase
Neither ATP nor PAP could substitute for PAPS, and heat- denatured cytosol did not support the reaction. A low PAPS- independent loss of N-HO-MAB was achieved only with cytosols from 0.9% NaCI solution-perfused livers. The loss of N-HO-MAB was 5 to 10 times greater in cytosols from nonperfused livers, and variable amounts of heme or Fe 2§ in these cytosols may have promoted the rapid decomposit ion of the N-HO-MAB (31). Omission of methionine from the assay medium did not affect the PAPS-dependent loss of N- HO-MAB, but addition of methionine was essential to the formation of a sulfonium-dye conjugate. Protein (added serum albumin) apparently competed with methionine for the reactive ester.
Experiments with N-HO-MAB and other N-hydroxy arylam- ines indicated that the optimal condit ions for N-hydroxy amine sulfotransferase activity differed from those for sulfo- transferase activity for N-HO-AAF or 3-hydroxyxanthine (16, 64); the major differences were the optimal pH and the protein and PAPS concentrations. The optimal assay condi- tions for sulfotransferase activity for N-HO-MAB and N-HO- AAF (cf. "Materials and Methods") were defined after test- ing different methods of rat hepatic cytosol preparation (with or without liver perfusion, various homogenization media, gel fi ltration of the cytosol), incubation times, pH, buffer composit ions, and the concentrations of Mg 2+, EDTA, PAPS, substrate, substrate solvent (ethanol, ace- tone, or dimethyl sulfoxide), and cytosol protein. Under the optimal assay condit ions the PAPS-dependent loss of N-HO- MAB and the formation of 3-(methion-S-yl)-N-methyl-4- aminoazobenzene (determined as 3-CH:,S-MAB) were 1st order with respect to protein concentration up to 5 mg/ml and linear with time for 10 to 15 min. The loss of N-HO-AAF and the formation of 1- and 3-(methion-S-yl)-N-acetyl-2-ami- nofluorene (as o-methylmercapto-N-acetyl-2-aminoflu- orene) were 1st order with protein concentration up to 1 mg/ml and linear with time for 30 to 60 min. Inhibit ion by PAPS occurred at concentrat ions above 1.0 and 0.65 mM, respectively, for N-HO-MAB and N-HO-AAF sulfotransferase activities.
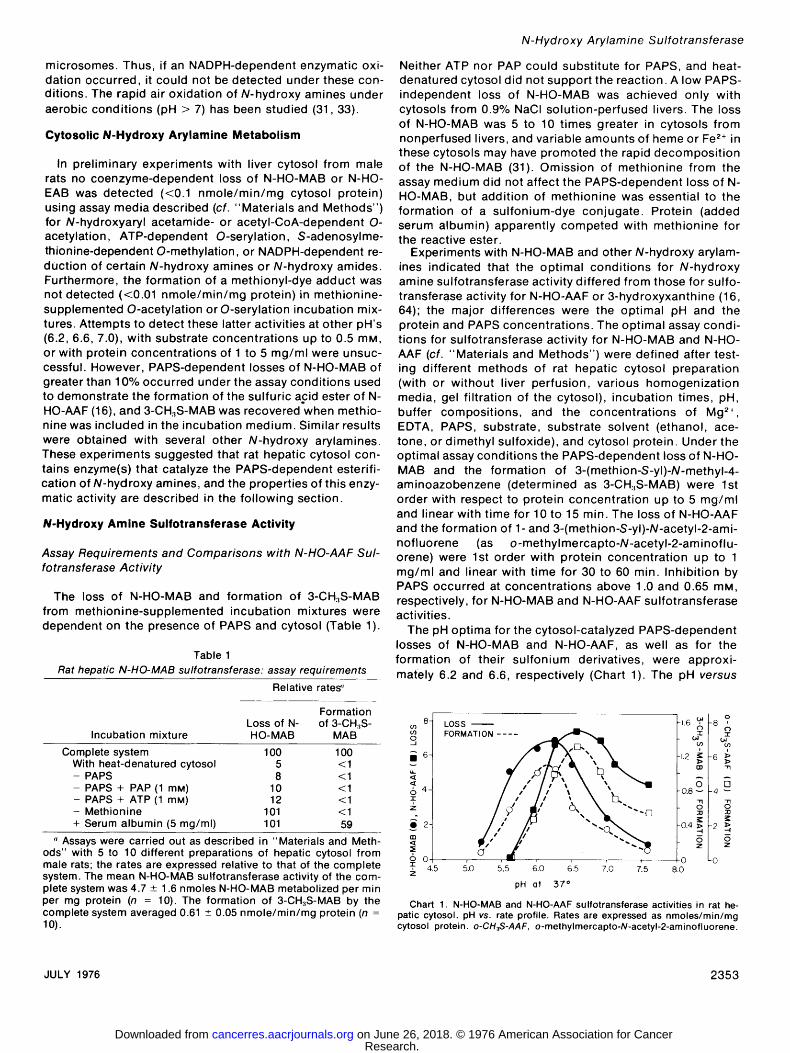
The pH optima for the cytosol-catalyzed PAPS-dependent losses of N-HO-MAB and N-HO-AAF, as well as for the formation of their sulfonium derivatives, were approxi- mately 6.2 and 6.6, respectively (Chart 1). The pH versus
LOSS ? FORMATION .... 0 9 %
0 | 0 LO , 4.5 ~.o ~.5- 6'.o 1.5 7'.o 7'.~ 8 .o z
pH 01 37 ~
Chart 1. N-HO-MAB and N-HO-AAF sul fotransferase act iv i t ies in rat he- patic cytosol, pH vs. rate profile. Rates are expressed as nmoles/min/mg cytosol protein, o-CH3S-AAF, o-methylmercapto-N-acetyl-2-aminofluorene.
JULY 1976 2353
Research. on June 26, 2018. © 1976 American Association for Cancercancerres.aacrjournals.org Downloaded from

F. F. Kadlubar et al.
rate profile for N-HO-AAF sulfotransferase activity is similar to that described by DeBaun et al. (16).
Previous studies on the sulfonation of phenols, amines, and steroids have demonstrated that several cytosolic pro- teins catalyze PAPS-dependent sulfonation reactions (19). These sulfotransferases have mult iple and overlapping sub- strate affinities and differ in their pH optima and sensitivity to inhibitors and activators. A "phenol" sulfotransferase that reportedly esterifies only phenols is unaffected by added Mg 2§ has optimal activity at pH 5.8, and, depending on the thiol content of the enzyme preparation, is activated or inhibited by o-iodosobenzoate (3, 4, 15, 45). Another sulfotransferase that acts on androgens, cholesterol, 2- naphthylamine, p-nitrophenol, and L-tyrosine methyl ester is stimulated by Mg 2+, has optimal activity at pH 7.0 to 7.5, and is inhibited by o-iodosobenzoate (3, 5, 15). A 3rd sulfo- transferase that acts on estrogens, 2-naphthylamine, and p-nitrophenol is stimulated by Mg2+; it has optima! activity at pH 6.2 for estrogens and 2-naphthylamine and at pH 7.0 for p-ni t rophenol (3, 19). Sulfonation of 2-naphthylamine is stimulated by addition of steroids while steroid sulfonation is inhibited by added 2-naphthylamine (3, 55). Another steroid sulfotransferase is thought to catalyze the esterifi- cation of deoxycort icosterone, but no further characteriza- tion has been made (3). An o-iodosobenzoate-sensitive sulfotransferase with activity toward L-tyrosine methyl ester, tyramine, and p-ni trophenol has also been described (5, 44).
As an aid in evaluating the roles of the various sulfotrans- ferases in the esterif ication of N-HO-AAF and N-HO-MAB, the effects of several inhibitors and activators on these reactions were investigated. The assays were carried out either under optimal condit ions for N-HO-MAB (1 mM PAPS, pH 6.2) and for N-HO-AAF (0.65 mM PAPS, pH 6.6) or under intermediate condit ions (0.65 mM PAPS, pH 6.4), so that both activities could be measured on a comparable basis (Table 2). At pH6.4, omission of Mg 2+ decreased the activity toward both N-HO-MAB and N-HO-AAF by 10 to 20%. At pH 7.0, the effect on N-HO-AAF activity was more pronounced; the rate in the absence of added Mg 2+ was only 50 to 60% of the control level. These differences may reflect the catalytic participation of several sulfotransferases and/or the inhibi- tory effect of Mg 2+ on PAPS-sulfohydrolase (18). o-lodoso- benzoate inhibited both N-HO-MAB and N-HO-AAF sulfo- transferase activity, but it was a much more effective inhibi- tor of the latter activity. Both 2-naphthylamine and p-nitro- phenol inhibited the esterif ication of N-HO-MAB and N-HO- AAF. Dehydroepiandrosterone, estrone, and deoxycorticos- terone increased sulfotransferase activity for N-HO-AAF by 20 to 40%, while only estrone and deoxycort icosterone ap- peared to increase sulfotransferase activity for N-HO-MAB. L-Tyrosine methyl ester did not signif icantly alter either N- HO-MAB. or N-HO-AAF sulfotransferase activity. The addi- tion of N-HO-AAF to assays for N-HO-MAB sulfotransferase activity caused a 40% stimulation, while N-HO-MAB was strongly inhibitory to N-HO-AAF sulfotransferase activity. Although several sulfotransferases may catalyze the esterifi- cation of both N-HO-MAB and N-HO-AAF, the activity for N- HO-MAB appears to be similar to that described for
Table 2 Effects of various agents on rat hepatic N-HO-MAB and N-HO-AAF
sulfotransferase activities
Relative rates of forma- tion ~
o-Methyl- mercapto- N-acetyl-2-
3-CH3S- aminoflu- Reaction conditions" MAB orene
Complete system 100 100 - M g =+ 78 86 + o-lodosobenzoate (0.25 mM) 42 4 + 2-Naphthylamine (2.0 mM) 23 33 + p-Nitrophenol (1.0 mM) 1 2 + Dehydroepiandrosterone (0.1 mM) 98 128 + Estrone (0.1 mM) 113 120 + Deoxycorticosterone (0.1 mM) 132 138 + L-Tyr0sine methyl ester (3 mM) 104 108 + N-HO-AAF (0.5 mM. ) 139 + N-HO-MAB (0.5 mM) 13
" The incubation medium was a compromise between the opti- mal systems for N-HO-MAB and N-HO-AAF and contained 100 mM Bis-Tris-HCI buffer (pH 6.4 at 37~ 100 mM methionine, 5 mM MgCI._,, 0.5 mM EDTA, 0.65 mM PAPS, rat liver cytosol protein (1 mg/ml), and 0.5 mM N-HO-MAB or N-HO-AAF. Incubation time was 15 min.
"Assays were carried out as described in "Materials and Meth- ods" using 4 to 6 different cytosol preparations; the rates are ex- pressed relative to the control. The mean rates of methylmercapto- arylamine formation in the complete system were 0.31 +- 0.03 and 6.1 - 0.5 nmoles/min/mg (n = 6) for N-HO-MAB and N-HO-AAF, re- spectively. Similar relative rates were obtained under the various reaction conditions by measuring the PAPS-dependent loss of N-HO-MAB or N-HO-AAF.
"phenol" sulfotransferase, while the activity for N-HO-AAF exhibits properties similar to those described for both phenol and steroid sulfotransferases.
Reactivity of MAB-N-sul fate
A relatively high concentration of methionine (100 mM) was required to trap appreciable amounts of the sulfuric acid ester of N-HO-MAB, and under these condit ions the formation of a sulfonium dye-conjugate accounted for only 5 to 15% of the PAPS-dependent loss of N-HO-MAB (Chart 2). With N-HO-AAF as substrate 20 mM methionine trapped 65 to 85% of the reactive ester as a methionyl derivative.
Addition of [8-1"C]guanosine (optimal concentration, 0.8 to 1.2 mM) to the sulfotransferase incubation medium yielded a [8-14C]guanosinyl adduct that accounted for 6 to 12% of the N-HO-MAB metabolized. The adduct was identi- fied as [8-14C]-N-(guanosin-8-yl)-MAB on the basis of its chromatographic properties in 4 thin-layer systems (38). Similar sulfotransferase incubation mixtures that contained 1 mM [~4C]adenosine, [14C]cytidine, [~4C]uridine, or [~4C]- thymidine yielded no significant amounts of nucleoside-dye adducts (<0.5% of the N-HO-MAB metabolized). This result was consistent with the lack of detectable reactivity of these nucleosides with the synthetic ester N-benzoyloxy-MAB (51). The sulfuric acid ester of N-HO-MAB also reacted with added rRNA (6.7 mg/ml), but only 1 to 3% of the N-hydroxy- amine metabolized was recovered as an rRNA adduct.
2354 CANCER RESEARCH VOL. 36
Research. on June 26, 2018. © 1976 American Association for Cancercancerres.aacrjournals.org Downloaded from

Tissue, Sex, and Species Distribution
Of the ex t rahepa t i c t issues s tud ied on ly the cy toso l frac- t ions of rat k idney and small in test ine con ta ined detec tab le su l fo t ransferase act iv i ty for N-HO-MAB (Table 3). These ac- t iv i t ies are no more than 20% of that found in hepat ic cytosol , but a comparab le level of su l fo t rans ferase act iv i ty for N-HO-AAF was not detected in rat k idney or smal l intes- t ine (16). No act iv i ty for N-HO-MAB was detec ted in rat muscle or lung cytosols .
The h ighest hepat ic su l fo t ransferase act iv i ty for N-HO- MAB was found in male rats. Female rat l iver and the l ivers from males of the o ther species s tud ied, except the ham- ster, had at least 25% of the act iv i ty of male rat l iver (Table 4) . N-HO-AAF su l fo t ransferase act iv i ty has been detec ted only in rat and rabbi t l iver cytosols (16).
Young adul t an imals were used in these compara t i ve studies. In young rats hepat ic N-HO-MAB su l fo t ransferase act ivi ty was cons is ten t l y lower than in o lder rats (cf. Tables
I.O- -5.0
o z o o.8- ~4.0 I-- -r ,~ oJ
o 0.6- -3.0 la.. -3'i A
0 [] m 0.4- -2.0 -,1 < O
m
" 0 !
o 2'0 40 eo ,oo
METHIONINE (raM) Chart 2. Effect of methionine on methylmercaptoarene formation from N-
HO-MAB and N-HO-AAF. Rates are expressed as nmoles methylmercaptoar- erie isolated per rain per mg cytosol protein, o-CH~-S-AAF, o-methylmer- capto-N-acetyl-2-aminoftuorene.
Table 3 N-HO-MAB sulfotransferase activity in various tissues from male
rats
Tissue
PAPS-dependent loss of N- HO-MAB (nmoles/min/mg
protein)"
Liver 2.7 _+ 0.5 ~,' Kidney 0.5 - 0.3 b Small intestine mucosa 0.5 _ 0.3 ~ Muscle <0.2" Lung <0.2'
" Assays were carried out with the cytosol fraction of each tissue homogenate, and the values reported represent determinations on tissues from 3 animals.
b For each tissue at least 2 reactions were also carried out jn the presence of methionine. In each case the amount of 3-CH3S-MAB that could be isolated at the end of the incubation period was equivalent to 5 to 10% of the PAPS-dependent loss of N-HO-MAB.
" Mean _ S.D. a These values were judged to be the lower limit of detection.
N-Hydroxy Arylamine Sulfotransferase
1 and 5). Thus, the lower N-HO-MAB act iv i t ies shown in Tables 3 and 4 ref lect th is age var iat ion.
Substrate Specif icity
N-HO-EAB, N-HO-AB, N - h y d r o x y - 1 - a n d 2-naphthy la - mine, and N -hyd roxy -4 -am inob ipheny l also appeared to be conver ted to react ive esters by rat hepat ic cy toso l and PAPS (Table 5). The PAPS-dependen t metabo l i sm of each of these N-hyd roxy amines was opt imal unde r the assay cond i t ions . fo r N-HO-MAB su l fo t rans ferase rather than those for N-HO-AAF ester i f i ca t ion . In the absence of PAPS, 80 to
Table 4 N-HO-MAB sulfotransferase activity in hepatic cytosols from
various species
PAPS-dependent loss of N-HO-MAB
No. of ani- (nmoles/min/mg Species Sex mals protein)"
Rat M 10 3.6 - 1.8 ~' Rat F 5 i.1 _+ 0.5 Rabbit M 3 1.9 -+ 0.5 Guinea pig M 3 1.3 + 0.3 Mouse M 3 1.1 _ 0.1 Hamster M 3 0.2 - 0.03
" See "Materials and Methods" for assay procedure. For the livers from each species at least 2 reactions were carried out in the presence of methionine. In each case the amount of 3-CH:~S-MAB that could be isolated at the end of the incubation period was equivalent to 5 to 10% of the PAPS-dependent loss of N-HO-MAB.
t, Mean _+ S.D.
Table 5 Substrate specificity of rat hepatic N-hydroxy amine
sulfotransferase(s)
Substrate
PAPS-de- pendent loss % isolable of substrate as methyl-
(nmoles/min/ mercapto- mg protein)" arylamine b
N-HO-MAB 4.8 _+ 1.8" 5-20 N-HO-EAB 2.2 -+ 0.6 8-12 N-HO-AB 1.9 _+ 0.6 1-3 N-Hydroxy-l-naphthylamine 2.4 _ 1.0 - ' N-Hydroxy-2-naphthylamine 1.4 __+ 0.3 - " N-Hydroxy-4-aminobiphenyl 1.1 + 0.4 25-40 trans-N-Hyd roxy-4-am i n osti I be n e, <0.1 - '
N-hyd roxy-2-aminofluorene, N- hydroxyaniline, and N-hydroxy- N-methyI-N-benzylamine
" Assays were carried out as described in "Materials and Meth- ods."
b The trapping efficiencies differed between liver cytosols, possi- bly because of variations in the amounts of endogenous nucleo- philes.
e Mean - S.D. for analyses of 5 to 10 different cytosol prepara- tions. The wide standard deviations reflect the variations in activity from one liver preparation to another.
' Not studied. The methylmercaptoarylamine was too labile to air oxidation to
permit its isolation and estimation by chromatography. Therefore, rRNA was used as a trapping agent; 0.2 to 0.3% of the PAPS- and cytosol-dependent loss of [:~H]-N-hydroxy-2-naphthylamine was ac- counted for as a rRNA-bound adduct. By comparison, 1 to 3% of the N-HO-MAB metabolized was recovered as rRNA-bound dye.
JULY 1976 2355
Research. on June 26, 2018. © 1976 American Association for Cancercancerres.aacrjournals.org Downloaded from

F. F. Kadlubar et al.
100% of each N-hydroxy amine added could be recovered after the 10-min incubation period; the low PAPS-independ- ent losses were presumed to be nonenzymatic. When me- thionine or rRNA was added to the complete assay media, methionyl or rRNA adducts were formed.
Products Formed in the Sulfotransferase Incubat ion Me- d ium
From N-HO-MAB. Incubation of N-HO-MAB in the sulfo- transferase assay in the absence of added guanosine, methionine, or RNA yielded the fol lowing products (ex- pressed as the percentage of the N-HO-MAB metabolized): MAB (50 to 60%), N-HO-AB (25 to 35%), formaldehyde (20 to 30%), and 2 unidentified products (15 to 25%). MAB and N-HO-AB were identified by thin-layer chro- matography, their UV absorption maxima in 95% ethanol (402 and 375 nm, respectively), and their mass spectra (M + = 211 and 213, respectively). Formaldehyde was character- ized spectrally as the 2,4-pentanedione-ammonia adduct (~ ...... = 412 nm). The unidentified products exhibited ab- sorption maxima at 390 to 400 nm in 95% ethanol and at 520 to 530 nm in 40% ethanol:0.5 M HCI; these products had low volatilities and did not yield molecular ions on electron impact mass spectrometry. These properties are consistent with those expected for a dinitrone or a nitrone-N-hydroxy amine addition product. All of the above reaction products were also obtained in assays supplemented with methionine or guanosine, but the yields were proportionately de- creased.
A reaction mechanism that is consistent with the above results is presented in Chart 3. In view of the great lability of the N-sulfate of N-HO-AAF and some other esters of N- hydroxy metabolites (7, 41,48, 51, 60), the apparent initial product from N-HO-MAB, MAB-N-sulfate, is presumed to be rapidly converted to a reactive nitrenium ion. This ion could then bind to cellular nucleophiles (cf. Ref. 59). However, unlike N-hydroxy amides (e.g., N-HO-AAF), N-hydroxy
- L ~ ~OS%H j \
~ : ' H ~ A B TM ~ TLR EN~II U M ~ON
r % N "CH2 ] A ~,-N "CH3 | ~ ~o I ~ ",4 ~ - ~ "H ] N-(p-PHENYLAZOPHENYL)[ . . . . . . . . . . . . L NITRONE j . . . . . . . . - .....
HCHO ,~ H20 "%'"" (20-30 % )~ ~'~ OTHER DYE PRODUCTS
H (15-25 %) N~OH
N-HO-AB (25- 35%)
%= YIELD FROM N-HOMAB
Chart 3. Reaction scheme for the formation of the products obtained from the PAPS-dependent cytosol-catalyzed esterification of N-HO-MAB in the absence of added nucleophiles.
amines such as N-HO-MAB are strong reducing agents. The latter property may account for the apparent reduction of the aminoazo nitrenium ion to MAB and the concomitant formation of oxidation products (N-HO-AB and nitrone-de- rived adducts) of N-HO-MAB. Reduction of acylaryl nitren- ium ions by several reducing agents has been demonstrated (61, 65). This reaction sequence may account for the low yields of arylamine-nucleophile adducts formed from N- hydroxy amines, as compared to the high yields of such adducts obtained from esters of N-HO-AAF.
Further support for this mechanism was obtained by incu- bation of 0.5/.Lmole of N-benzoyloxy-MAB with 0.5/~mole of N-HO-MAB under the sulfotransferase assay condit ions, ex- cept for the omission of PAPS and cytosol. Both of the added dyes were consumed within 10 min, and 0.16/~mole of MAB, 0.28 /~mole of N-HO-AB, 0.14 /~mole of formalde- hyde, and 5 to 6 other unidentified dye-containing products were obtained. Incubation of N-HO-MAB in the absence of N-benzoyloxy-MAB under these conditions yielded only the starting material, while incubation of N-benzoyloxy-MAB alone yielded several of the above unidentif ied dye prod- ucts.
From N-Hydroxy-2-naphthylamine. A similar reaction se- quence appeared to occur when [3H]-N-hydroxy-2-naphthyl- amine was incubated with PAPS and hepatic cytosol (Chart 4), but the identification of the naphthalene derivatives was hampered by the relatively low rate of esterification of this N-hydroxy amine. The tentative identif ications of the prod- ucts are based on their chromatographic behaviors in 2 thin-layer systems and the characteristic f luorescence of each product under 254 nm light (10). The formation of each product was dependent on the presence of PAPS in the incubation mixture. 2-Amino-l-naphthyl sulfate accounted for 30 to 35% of the substrate metabolized. 2-Naphthyla-
OH ~NH 2 * H(~+ HS04 e
2-AMINO-I- NAPHTHOL
~
'0'" I , so:. ~N:oH J 2-AMINO-I-NAPHTHYL-
SULFATE P"PSTcY'~176 T N ~H N'. \OH cytosol ) .-\'OSO3
/ 2 " ~ M I N E : ~ "~
y> ; >, + HSO~
Chart 4. Reaction scheme for the formation of the prod ucts obtained from the PAPS-dependent cytosol-catalyzed esterification of N-hydroxy-2-naph- thylamine in the absence of added nucleophiles.
2356 CANCER RESEARCH VOL. 36
Research. on June 26, 2018. © 1976 American Association for Cancercancerres.aacrjournals.org Downloaded from

mine and 2,2'-azoxynaphthalene, which were only partially resolved by chromatography, accounted for another 35 to 40%. While the 5 to 10% yield of 2-amino-l-naphthol was dependent on PAPS, 2-amino-l-naphthol was converted to 2-amino-l-naphthyl sulfate in the sulfotransferase reaction medium at 5 to 10 times the rate of conversion of N-hydroxy- 2-naphthylamine to this product. The reaction scheme, while only tentative, allows for both sulfamate and ester formation and is consistent with the reported chemical properties of N- and O-sulfonates of N-hydroxy arylamines (12-14).
Conversion of MAB to N-(Guanosin-8-yl)-MAB by the 10,000 x g Supernatant of Rat Liver
By appropriate modifications of the MAB N-oxidase (33) and N-HO-MAB sulfotransferase assay procedures, it was possible to demonstrate the conversion of MAB to a nucleo- side adduct on incubation with the 10,000 x g supernatant fraction of rat liver. For this purpose [3H]MAB and guano- sine were used as substrate and trapping agent, respec- tively. Demonstration of this overall reaction required the selection of an intermediate pH and elimination of potas- sium phosphate buffer, which inhibited sulfotransferase ac- tivity,, and Tris-HCI buffer, which inhibited N-oxidation. Me- thionine (100 raM) could not be used as the trapping agent, since it completely inhibited the N-oxidase. PAPS was also somewhat inhibitory (40%) to the N-oxidase. The incubation medium selected for demonstration of the reaction con- tained 100 mM Bis-Tris-HCI buffer (pH 7.0 at 37~ 1 mM NAN3, 5 mM MgCI=, 0.5 mM EDTA, 1 mM PAPS, an NADPH- and NADH-generating system (33), 1 mM guanosine, 5 mg protein from 10,000 x g supernatant per ml, and 0.5 mM [3H]MAB (1.5 Ci/mmole). The isolation, separation, and estimation of [3H]-N-(guanosin-8-yl)-MAB were carried out as described above for the 1"C-labeled derivative. Under these conditions the formation of [3H]-N-(guanosin-8-yl)- MAB from [3H]MAB was demonstrated and shown to be enhanced by the addition of both PAPS and a NADPH, NADH-generating system (Chart 5).
MAB ~ PAPS ; N-HO-MAB l, MAB-N-SULFATE NADPH
3.0- GUANOSINL~ N" (GUANOSIN- 8 -YL ) MAB
Y COMPLETE S Y S T E M ~
2.0- ? d,
i z
1.0- - PAPS /'k
-NADPH -NADH G
O O 5 IO 115
MIN
Chart 5. N-Oxidat ion of MAB and O-esteri f icat ion of N-HO-MAB by the 10,000 x g supernatant of rat liver.
N-Hydroxy Arylamine Sulfotransferase
DISCUSSION
The hepatocarcinogen MAB is N-hydroxylated by a mixed-function amine oxidase in the hepatic endoplasmic reticulum (33). As shown in this report the product of this oxidation, N-HO-MAB, is sulfonated by 1 or more sulfo- transferases in the hepatic cytosol in a PAPS-dependent reaction to form a reactive electrophile, presumably the ester MAB-N-sulfate. This electrophile forms products of known structure with nucleophiles such as methionine and guanosine that are identical to products derived from the protein- and nucleic acid-bound dyes formed in vivo in the livers of rats fed MAB (38, 58). MAB-N-sulfate is thus a possible ultimate carcinogenic metabolite of MAB. Evi- dence supporting this concept has come from the recent observations of Blunck and Crowther (8) that the hepatocar- cinogenicity of the closely related dye, 3'-methyI-N,N-di- methyl-4-aminoazobenzene, is greatly enhanced in the rat by dietary administration of a high level of sodium sulfate. These data on the metabolism and carcinogenicity of MAB thus closely parallel data previously obtained for the metab- olism and carcinogenicity of AAF in the rat liver (16, 17, 23, 48, 69, 74), although the N-oxidase and sulfotransferase activities for MAB and N-HO-MAB, respectively, differ from those for AAF and N-HO-AAF. The microsomal N-oxidase for MAB does not depend on cytochrome P-450 and pre- sumably is a flavoprotein amine oxidase similar to that described by Ziegler et al. (33, 78). The microsomal oxidase for AAF involves cytochrome P-450 (69). Similarly, as shown in the present report (cf. "Results"), the hepatic sulfotrans- ferase activities for N-HO-AAF and N-HO-MAB appear to be different. The former activity appears to have properties noted for both phenol and steroid sulfotransferases, while the latter activity appears to be more similar to that of the phenol sulfotransferase.
The inability to detect either O-acetylation or O-serylation of N-HO-MAB, as has been demonstrated for other carcino- genic N-hydroxy arylamines (6, 7, 36, 66, 67), emphasizes the probable importance of O-sulfonation in the metabolism of this N-hydroxy dye. The failure of hepatic cytosols (NADPH-dependent) and of hepatic microsomes (NADH-de- pendent) to catalyze the reduction of certain N-hydroxy arylamines further conserves these substrates for this met- abolic pathway.
The tissue, sex, and species distributions of microsomal N-oxidase activity for MAB (33) and cytosolic sulfotransfer- ase activity for N-HO-MAB (Tables 3 and 4) are consistent with the observation that MAB and related aminoazo dyes are strongly carcinogenic only in the liver of the male rat (47). Likewise, the low hepatocarcinogenicit ies of EAB and AB (2, 47, 68), 1-naphthylamine (Ref. 24, pp. 100-101; Ref. 30, p. 168; Ref. 62, p. 183; Ref. 63, p. 126; Ref. 70, p. 559; Ref. 71, pp. 271-272), 2-naphthylamine (Ref. 24, pp. 101- 103; Ref. 30, pp. 169-170; Ref. 62, pp. 183-185; Ref. 63, pp. 127-128; Ref. 70, pp. 559-564; Ref. 71, p. 272), and 4- aminobiphenyl (Ref. 30, p. 104; Ref. 62, p. 102; Ref. 63, p. 71; Ref. 70, pp. 375-376; Ref. 71, p. 200) may result in part from the lower rates of O-sulfonation of the N-hydroxy metabolites of these amines in the rat liver as compared to those noted for N-HO-MAB (Table 5).
JULY 1976 2357
Research. on June 26, 2018. © 1976 American Association for Cancercancerres.aacrjournals.org Downloaded from

F. F. K a d l u b a r e t a l .
N - ( p - P h e n y l a z o ) p h e n y l n i t r o n e a lso d e s e r v e s c o n s i d e r a t i o n as a p o s s i b l e u l t i m a t e c a r c i n o g e n o f N - H O - M A B . As n o t e d
p r e v i o u s l y (33), N - H O - M A B is eas i l y o x i d i z e d a e r o b i c a l l y to a
r e a c t i v e d e r i v a t i v e , p r e s u m a b l y a n i t r o n e , t h a t can f o r m
a d d u c t s in a n h y d r o u s m e d i a w i t h c a r b o n - c a r b o n and car -
b o n - n i t r o g e n d o u b l e b o n d s in a v a r i e t y o f t i s s u e c o n s t i t u -
en t s . F rom d a t a p r e s e n t e d in t h i s r e p o r t , t h i s n i t r o n e a lso a p p e a r s to be f o r m e d d u r i n g t he o x i d a t i o n o f N - H O - M A B by
m e t a b o l i c a l l y f o r m e d M A B - N - s u l f a t e to y ie ld p r o d u c t s in-
c l u d i n g M A B , N - H O - A B , and f o r m a l d e h y d e . T h e la t te r 2
p r o d u c t s m a y ar ise by h y d r o l y s i s o f N - ( p - p h e n y l -
a z o ) p h e n y l n i t r o n e , an in i t ia l p r o d u c t in t he o x i d a t i o n - r e -
d u c t i o n r e a c t i o n . A r e a c t i o n b e t w e e n N - b e n z o y l o x y - M A B
and N - H O - M A B o c c u r r e d in the a b s e n c e o f c y t o s o l and
P A P S and y i e l d e d s i m i l a r p r o d u c t s . In a p r e v i o u s r e p o r t (33) it w a s n o t e d t h a t N - H O - E A B d id no t f o r m a r e a c t i v e n i t r o n e
u p o n a e r o b i c o x i d a t i o n . T h u s , t h e s e o b s e r v a t i o n s are c o n -
s i s t e n t w i t h t he ea r l i e r f i n d i n g s t h a t an N - m e t h y l g r o u p w a s
r e q u i r e d fo r t he f o r m a t i o n o f h i g h leve ls o f p r o t e i n - b o u n d dye i n v i v o and fo r s t r o n g h e p a t o c a r c i n o g e n i c i t y by ami -
n o a z o d y e s in t he rat (2, 47, 68).
A C K N O W L E D G M E N T S
The authors wish to express their gratitude to Nancy Korda for her expert technical assistance during the course of these studies.
R E F E R E N C E S
1. Andersen, R. A., Enomoto, M., Miller, E. C., and Miller, J. A. Carcino- genesis and Inhibition of the Walker 256 Tumor in the Rat by trans-4- Acetylaminostilbene, Its N-Hydroxy Metabolite, and Related Com- pounds. Cancer Res., 24:128-143, 1964.
2. Arcos, J. C., and Simon, J. Effect of 4'-Substituents on the Carcinogenic Activity of 4-Aminoazobenzene Derivatives. ArzneimitteI-Forsch., 12: 270-275, 1962.
3. Banerjee, R. K., and Roy, A. B. The Sulfotransferases of Guinea Pig Liver. Mol. Pharmacol., 2: 56-66, 1966.
4. Banerjee, R. K., and Roy, A. B. Kinetic Studies of the Phenol Sulfotrans- ferase Reaction. Biochim. Biophys. Acta, 151: 573-586, 1968.
5. Barford, D. J., and Jones, J. G. ThioI-Dependent Changes in the Proper- ties of Rat Liver Sulfotransferases. Biochem. J., 123: 427-434, 1971.
6. Bartsch, H., Dworkin, C., Miller, E. C., and Miller, J. A. Formation of Electrophilic N-Acetoxyarylamines in Cytosols from Rat Mammary Gland and Other Tissues By Transacetylation from the Carcinogen N-Hydroxy- 4-acetylaminobiphenyl. Biochim. Biophys. Acta, 304: 42-55, 1973.
7. Bartsch, H., Dworkin, M., Miller, J. A., and Miller, E. C. Electrophilic N- Acetoxyaminoarenes Derived from Carcinogenic N-Hydroxy-N-acetyl- aminoarenes by Enzymatic Deacetylation and Transacetylation in Liver. Biochim. Biophys. Acta, 286: 272-298, 1972.
8. Blunck, J. M., and Crowther, C. E. Enhancement of Azo Dye Carcinogen- esis by Dietary Sodium Sulfate. European J. Cancer, 11: 23-32, 1975.
9. Booth, J., and Boyland, E. The Biochemistry of Aromatic Amines. 10. Enzymic N-Hydroxylation of Arylamines and Conversion of Arylhydroxyl- amines into o-Aminophenols. Biochem. J., 91: 362-369, 1964.
10. Booth, J., Boyland, E., and Manson, D. Metabolism of Polycyclic Com- pounds. 9. Metabolism of 2-Naphthylamine in Rat Tissue Slices. Bio- chem. J., 60: 62-71, 1955.
11. Boyland, E., and Manson, D. The Biochemistry of Aromatic Amines. 2- Formamido-l-naphthyl Hydrogen Sulfate, a Metabolite of 2-Naphthyla- mine. Biochem. J., 99: 189-199, 1966.
12. Boyland, E., and Manson, D. The Biochemistry of Aromatic Amines. The Metabolism of 2-Naphthylamine and 2-Naphthylhydroxylamine Deriva- tives. Biochem. J., 101: 84-102, 1966.
13. Boyland, E., Manson, D., and Sims, P. The Preparation of o-Amino- phenyl Sulfates. J. Chem. Soc., 3623-3628, 1953.
14. Boyland, E., and Nery, R. Arylhydroxylamines. Part II. Phenylhydroxyl- amine N- and O-Sulphonic Acids. J. Chem. Soc., 5217-5222, 1962.
15. Carroll, J. and McEvoy, F. A. The Activation of Liver Phenol Sulfotrans- ferase. Biochem. J., 119: 27p, 1970.
16. DeBaun, J. R., Miller, E. C., and Miller, J. A. N-Hydroxy-2-acetylamino- fluorene Sulfotransferase: Its Probable Role in Carcinogenesis and in Protein-(methion-S-yl) Binding in Rat Liver. Cancer Res., 30: 577-595, 1970.
17. DeBaun, J. R., Smith, J. Y. R., Miller, E. C., and Miller, J. A. Reactivity in Vivo of the Carcinogen N-Hydroxy-2-acetylaminofluorene: Increase by Sulfate Ion. Science, 167: 184-186, 1970.
18. Denner, W. H. B., Stokes, A. M., Rose, F. A., and Dodgson, K. S. Separation and Properties of the Soluble 3'-Phosphoadenosine-5'-Phos- phosulfate-Degrading Enzymes of Bovine Liver. Biochim. Biophys. Acta, 315: 394-401, 1973.
19. Dodgson, K. S., and Rose, F. A. Sulfoconjugation and Sulfohydrolysis. In: W. H. Fishman (ed.), Metabolic Conjugation and Metabolic Hydroly- sis, Vol. 1, pp. 239-325. New York: Academic Press, Inc., 1970.
20. Exner, O. Derivatives of Oximes. I1. Reduction of O- and N-Alkyl Oximes with Lithium Aluminum Hydride. Collection Czech. Chem. Commun., 20: 202-209, 1955.
21. Gillette, J. R., Davis, D. C., and Sasame, H. A. Cytochrome P-450 and Its Role in Drug Metabolism. Ann. Rev. Pharmacol., 12: 57-84, 1972.
22. Gornall, A. G., Bardawill, C. J., and David, M. M. Determination of Serum Proteins by Means of the Biuret Reaction. J. Biol. Chem., t77: 751-766, 1949.
23. Gutmann, H. R., Malejka-Giganti, D., Barry, E. J., and Rydell, R. E. On the Correlation between the Hepatocarcinogenicity of the Carcinogen, N-2-Fluorenylacetamide, and Its Metabolic Activation by the Rat. Cancer Res., 32: 1554-1561, 1972.
24. Hartwell, J. L. Survey of Compounds Which Have Been Tested for Carci- nogenic Activity. U. S. Public Health Service Publication No. 149. Wash- ington, D. C.: U. S. Government Printing Office, 1951.
25. Hogeboom, G. H., Schneider, W. C., and Palade, G. E. Cytochemical Studies of Mammalian Tissues. I. Isolation of Intact Mitochondria From Rat Liver: Some Biochemical Properties of Mitochondria and Submi- croscopic Particulate Material. J. Biol. Chem., 172: 619-635, 1948.
26. Irving, C. C. Enzymatic Deacetylation of N-Hydroxy-2-Acetylamino-flu- orene by Liver Microsomes. Cancer Res., 26 (Part 1): 1390-1396, 1966.
27. Irving, C. C., Janss, D. H., and Russell, L. T. Lack of N-Hydroxy-2- acetylaminofluorene Sulfotransferase Activity in the Mammary Gland and Zymbal's Gland of the Rat. Cancer Res., 31: 387-391, 1971.
28. Irving, C. C., and Veazey, R. A. Isolation of Deoxyribonucleic Acid and Ribosomal Ribonucleic Acid from Rat Liver. Biochim. Biophys. Acta, 166: 246-248, 1968.
29. Jacobson, M., Levin, W., Lu, A. Y. H., Conney, A. H., and Kuntzman, R. The Rate of Pentobarbital and Acetanilide Metabolism by Liver Micro- somes: A Function of Lipid Peroxidation and Degradation of Cytochrome P-450 Heine. Drug. Metab. Disposition, I : 766-774, 1973.
30. John I. Thompson and Co. Survey of Compounds Which Have Been Tested for Carcinogenic Activity, Vol. 1968-1969, U. S. Public Health Service Publication No. 149. Washington, D. C.: U. S. Government Printing Office, 1973.
31. Johnson, D. H., Rogers, M. A. T., and Trappe, G. Aliphatic Hydroxyla- mines. Part I1. Autoxidation. J. Chem. Soc., 1093-1103, 1956.
32. Kadlubar, F. F., McKee, E. M., and Ziegler, D. M. Reduced Pyridine Nucleotide-Dependent H-Hydroxy Amine Oxidase and Reductase Activi- ties of Hepa tic Microsomes. Arch. Biochem. Biophys., 156: 46-57, 1973.
33. Kadlubar, F. F., Miller, J. A., and Miller, E. C. Microsomal N-Oxidation of the Hepatocarcinogen N-Methyl-4-aminoazobenzene and the Reactivity of N-Hydroxy-N-methyl-4-aminoazobenzene. Cancer Res., 36: 1196- 1206, 1976.
34. Kadlubar, F. F., and Ziegler, D. M. Properties of a NADH-Dependent N- Hydroxy Amine Reductase Isolated from Pig Liver Microsomes. Arch. Biochem. Biophys., 162: 83-92, 1974.
35. Kamm, O. /3-Phenylhydroxylamine. In: H. Gilman (ed.), Organic Syntheses, Coll. Vol. 1, pp. 445-447. New York: John Wiley and Sons, Inc., 1941.
36. King, C. M. Mechanism of Reaction, Tissue Distribution, and Inhibition of Arylhydroxamic Acid Acyltransferase. Cancer Res., 34: 1503-1515, 1974.
37. King, C. M., and Phillips, B. N-Hydroxy-2-fluorenylacetamide. Reaction of the Carcinogen with Guanosine, Ribonucleic Acid, Deoxyribonucleic Acid, and Protein following Enzymatic Deacetylation or Esterification. J. Biol. Chem., 244: 6209-6216, 1969.
38. Lin, J. K., Miller, J. A., and Miller, E. C. Structures of Hepatic Nucleic Acid-Bound Dyes in Rats Given the Carcinogen N-Methyl-4-aminoazo- benzene. Cancer Res., 35: 844-850, 1975.
39. Lotlikar, P. D. Enzymatic N-O-methylation of Hydroxamic Acids. Biochim. Biophys. Acta, 170: 468-471, 1968.
40. Lotlikar, P. D., Miller, E. C., Miller, J. A., and Margreth, A. The Enzymatic Reduction of the N-Hydroxy Derivatives of 2-Acetylaminofluorene and Related Carcinogens by Tissue Preparations. Cancer Res., 25: 1743- 1752, 1965.
41. Maher, V. M., Miller, E. C., Miller, J. A., and Szybalski, W. Mutations and Decreases in Density of Transforming DNA Produced by Derivatives of
2 3 5 8 CANCER RESEARCH VOL. 36
Research. on June 26, 2018. © 1976 American Association for Cancercancerres.aacrjournals.org Downloaded from

the Carcinogens 2-Acetylaminofluorene and N-Methyl-4-aminoazoben- zene. Mol. Pharmacol., 4:411-426, 1968.
42. Maher, V. M., Miller, J. A., Miller, E. C., and Summers, W. C. Mutations and Loss of Transforming Activity of Bacillus subtillis DNA after Reaction with Esters of Carcinogenic N-Hydroxy Aromatic Amides. Cancer Res., 30: 1473-1480, 1970.
43. Manson, D. Oxidation of N-Naphthylhydroxylamines to Nitrosonaphthols by Air. J. Chem. Soc. Perkin Trans. I, 192-194, 1974.
44. Mattock, P., and Jones, J. G. Partial Purification and Properties of an Enzyme from Rat Liver That Catalyzes the Sulfation of L-Tyrosyl Deriva- tives. Biochem. J., 116: 797-803, 1970.
45. McEvoy, F. A., and Carroll, J. The Isolation of Rat Liver Phenol Sulfo- transferase. Biochem. J., 119: 26p-27p, 1970.
46. Miller, J. A., and Baumann, C. A. The Determination ofp-Dimethylamino- azobenzene, p-Monomethylaminoazobenzene and p-Aminoazobenzene in Tissue. Cancer Res., 5: 157-161, 1945.
47. Miller, J. A., and Miller, E. C. The Carcinogenic Aminoazo Dyes. Advan. Cancer Res., 1: 339-396, 1953.
48. Miller, J. A., and Miller, E. C. The Metabolic Activation of Aromatic Amines and Amides. Progr. Exptl. Tumor Res., 11: 273-301, 1969.
49. Nash, T. The Colorimetric Estimation of Formaldehyde by Means of the Hantzsch Reaction. Biochem. J., 55: 416-421, 1953.
50. Poirier, L. A., Miller, J. A., and Miller, E. C. The N- and Ring-Hydroxyl- ation of 2-Acetylaminofluorene and the Failure to Detect N-Acetylation of 2-Aminofluorene in the Dog. Cancer Res., 23: 790-800, 1963.
51. Poirier, L. A., Miller, J. A., Miller, E. C., and Sato, K. N-Benzoyloxy-N- methyl-4-aminoazobenzene: Its Carcinogenic Activity in the Rat and Its Reaction with Proteins and Nucleic Acids and Their Constituents in Vitro. Cancer Res., 27: 1600-1613, 1967.
52. Poirier, L. A., and Weisburger, J. H. Enzymic Reduction of Carcinogenic Aromatic Nitro Compounds by Rat and Mouse Liver Fractions. Biochem. Pharmacol., 23: 661-669, 1974
53. Poulsen, L. L., Kadlubar, F. F., and Ziegler, D. M. Role of the Microsomal Mixed-Function Amine Oxidase in the Oxidation of N,N-Disubstituted Hydroxylamines. Arch. Biochem. Biophys., 164: 774-775, 1974.
54. Poulsen, L. L., Masters, B. S. S., and Ziegler, D. M. Mechanism of 2- Naphthyiamine Oxidation Catalyzed by Pig Liver Microsomes. Xenobi- otica, in press.
55. Roy, A. B. The Enzymic Synthesis of Aryl Sulphamates. Biochem. J., 74: 49-56, 1960.
56. Sato, K., Poirier, L. A., Miller, J. A., and Miller, E. C. Studies on the N- Hydroxylation and Carcinogenicity of 4-Aminoazobenzene and Related Compounds. Cancer Res., 26: 1678-1687, 1966.
57. Scribner, J. D., and Miller, J. A. Intramolecular Hydrogen Bonding in o- Methylmercapto Derivatives of N-Methylaniline and N-Methyl-4-aminoa- zobenzene. J. Org. Chem., 32: 2348-2349,-1967.
58. Scribner, J. D., Miller, J. A., and Miller, E. C. 3-Methylmercapto-N- Methyl-4-aminoazobenzene: An Alkaline-Degradation Product of a La- bile Protein-Bound Dye in the Livers of Rats Fed N,N-Dimethyl-4-aminoa- zobenzene. Biochem. Biophys. Res. Commun., 20: 560-565, 1965.
59. Scribner, J. D., Miller, J. A., and Miller, E. C. Nucleophilic Substitution on Carcinogenic N-Acetoxy-N-arylacetamides. Cancer Res., 30: 1570- 1579, 1970.
60. Scribner, J. D., and Naimy, N. K. Reactions of Esters of N-Hydroxy-2- acetamidophenanthrene with Cellular Nucleophiles and the Formation
N - H y d r o x y A r y l a m i n e S u l f o t r a n s f e r a s e
of Free Radicals upon Decomposition of N-Acetoxy-N-arylacetamides. Cancer Res., 33: 1159-1164, 1973.
61. Scribner, J. D., and Naimy, N. K. Destruction of Triplet Nitrenium Ion by Ascorbic Acid. Experientia, 31: 470-471, 1975.
62. Shubik, P., and Hartwell, J. L. Survey of Compounds Which Have Been Tested for Carcinogenic Activity, Suppl. 2, U. S. Public Health Service Publication No. 149. Washington, D. C.: U. S. Government Printing Office, 1969.
63. Shubik, P., and Hartwell, J. L. Survey of Compounds Which Have Been Tested for Carcinogenic Activity, Suppl. 1. U. S. Public Health Service Publication No. 149. Washington D. C.: U. S. Government Printing Office, 1957.
64. StOhrer, G., Corbin, E., and Brown, G. B. Enzymatic Activation of the Oncogen 3-Hydroxyxanthine. Cancer Res., 32: 637-642, 1972.
65. StOhrer, G., and Salemnick, G. Oxidizing Action of Purine N-Oxide Esters. Cancer Res., 35: 122-131, 1975.
66. Tada, M., and Tada, M. Enzymatic Activation of the Carcinogen 4- Hydroxyaminoquinoline 1-Oxide and Its Interaction with Cellular Macro- molecules. Biochem. Biophys. Res. Commun., 46: 1025-1032, 1972.
67. Tada, M., and Tada, M. Seryl-tRNA Synthetase and Activation of the Carcinogen 4-Nitroquinoline 1-Oxide. Nature, 255: 510-512, 1975.
68. Terayama, H. Aminoazo Carcinogenesis-Methods and Biochemical Problems. In: H. Busch (ed.), Methods in Cancer Research, Vol. 1, pp. 399-449. New York: Academic Press, Inc., 1967.
69. Thorgeirsson, S. S., Jollow, D. J., Sasame, H. A., Green, I., and Mitchell, J. R. The Role of Cytochrome P-450 in N-Hydroxylation of 2-Acetylami- nofluorene. Mol. Pharmacol., 9: 398-404, 1973.
70. Tracor/Jitco. Survey of Compounds Which Have Been Tested for Carci- nogenic Activity, Vol. 1961-1967. U. S. Public Health Service Publication No. 149. Washington, D. C.: U. S. Government Printing Office, 1972.
71. Tracor/Jitco. Survey of Compounds Which Have Been Tested for Carcinogenic Activity, Vol. 1970-1971. U. S. Public Health Service Publi- cation No. 149. Washington, D. C.: U. S. Government Printing Office, 1974.
72. Tsen, C. C. An Improved Spectrophotometric Method for the Determina- tion of Tocopherols Using 4,7-Diphenyl-l,10-phenanthroline. Anal. Chem., 33: 849-851, 1961.
73. Uehleke, H. The Role of Cytochrome P-450 in the N-Oxidation of Individ- ual Amines. Drug. Metab. Disposition, I : 299-313, 1973.
74. Weisburger, J. H., Yamamoto, R. S., Williams, G. M. Grantham, P. H., Matsushima, T., and Weisburger, Eo K. On the Sulfate Ester of N-Hy- droxy-N-2-fluorenylacetamide as a Key Ultimate Hepatocarcinogen in the Rat. Cancer Res., 32: 491-500, 1972.
75. Willstatter, R., and Kubli, H. Uber die Reduktion von Nitroverbindungen nach der Methode von Zinin. Chem. Ber., 41: 1936-1940, 1908.
76. Wislocki, P. G., Borchert, P., Miller, J. A., and Miller, E. C. The Metabolic Activation of the Carcinogen l'-Hydroxysafrole in Vivo and in Vitro and the Electrophilic Reactivities of Possible Ultimate Carcinogens. Cancer Res., 36: 1686-1695, 1976.
77. Ziegler, D. M., McKee, E. M., and Poulsen, L. L. Microsomal Flavopro- tein-Catalyzed N-Oxidation of Arylamines. Drug Metab. Disposition, 1: 314-321, 1973.
78. Ziegler, D. M., and Mitchell, C. H. Microsomal Oxidase: IV. Properties of a Mixed-Function Amine Oxidase Isolated from Pig Liver Microsomes. Arch. Biochem. Biophys., 150: 116-125, 1972.
JULY 1976 2 3 5 9
Research. on June 26, 2018. © 1976 American Association for Cancercancerres.aacrjournals.org Downloaded from

1976;36:2350-2359. Cancer Res Fred F. Kadlubar, James A. Miller and Elizabeth C. Miller to Reactive Sulfuric Acid Esters
-Hydroxy ArylaminesN-methyl-4-aminoazobenzene and Other N-Hydroxy-NHepatic Metabolism of
Updated version
http://cancerres.aacrjournals.org/content/36/7_Part_1/2350
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/36/7_Part_1/2350To request permission to re-use all or part of this article, use this link
Research. on June 26, 2018. © 1976 American Association for Cancercancerres.aacrjournals.org Downloaded from