habitat assessment for a threatened marsupial in temperate
TRANSCRIPT

1
HABITAT ASSESSMENT FOR A THREATENED MARSUPIAL IN TEMPERATE FOREST OF PATAGONIA
BY
MARIANO ALBERTO RODRIGUEZ-CABAL
A THESIS PRESENTED TO THE GRADUATE SCHOOL OF THE UNIVERSITY OF FLORIDA IN PARTIAL FULFILLMENT
OF THE REQUIREMENTS FOR THE DEGREE OF MASTER OF SCIENCE
UNIVERSITY OF FLORIDA
2008

2
© 2008 Mariano Alberto Rodriguez Cabal

3
To Noelia and to all who nurtured my curiosity and love for nature.

4
ACKNOWLEDGMENTS
I thank my supervisory committee (L. C. Branch, K Sieving and E. Bruna) for their
mentoring. I also thank the faculty, staff and grad students at the University of Florida (UF)
Department of Wildlife Ecology and Conservation. I thank Mauricio Failla, Director de Fauna de
la Provincia de Rio Negro, Argentina; Gustavo Iglesias and Claudio Chehebar, Nahuel Huapi
National; Park, and staff of Parque Municipal Llao Llao. Special thanks go to M. Aizen, G.
Amico, N. Barrios, G. Barrios, R. Bunge, J. M. Morales, S. Morgan, A. Martínez, M. Nuñez, E.
Perner, A. Pries, R. Reggio, and N. Tercero-Bucardo for valuable assistance in the field and for
their thoughtful comments and suggestions on previous drafts. Field work was supported by
Wildlife Conservation Society, Scott Neotropical Fund, and the Jennings Scholarship of the UF
Department of Wildlife Ecology and Conservation.

5
TABLE OF CONTENTS page
ACKNOWLEDGMENTS ...............................................................................................................4
LIST OF TABLES...........................................................................................................................6
LIST OF FIGURES .........................................................................................................................7
ABSTRACT.....................................................................................................................................8
CHAPTER
1 HABITAT ASSESSMENT FOR MONITOS DEL MONTE ................................................10
Introduction.............................................................................................................................10 Methods ..................................................................................................................................13
Study Area .......................................................................................................................13 Marsupial Presence - Absence.........................................................................................14 Marsupial Abundance......................................................................................................15 Habitat Variables .............................................................................................................15 Data Analysis...................................................................................................................16
Results.....................................................................................................................................19 Marsupial Presence - Absence.........................................................................................19 Marsupial Abundance......................................................................................................20
Discussion...............................................................................................................................21
2 CONSERVATION IMPLICATIONS....................................................................................25
3 CONCLUSION.......................................................................................................................32
APPENDIX
A CORRELATION ANALYSIS ...............................................................................................33
LIST OF REFERENCES...............................................................................................................35
BIOGRAPHICAL SKETCH .........................................................................................................40

6
LIST OF TABLES
Table page 1-1 Habitat variables for sites where monitos del monte were present or absent.. ..................25
1-2 Logistic regression models at the 28 study sites.. ..............................................................26
1-3 Regression models constructed at 12 study sites.. .............................................................27
A-1 Correlation coefficients between habitat variables in the presence – absence survey ......33
A-2 Correlation coefficients between habitat variables in the abundance survey. ...................34

7
LIST OF FIGURES
Figure page 1-1 Habitat variables at sites where abundances of monito del monte were determined.........28
1-2 Number of monitos del monte vs. the significance predictor variables.............................29

8
Abstract of Thesis Presented to the Graduate School of the University of Florida in Partial Fulfillment of the
Requirements for the Master of Science
HABITAT ASSESSMENT FOR A THREATENED MARSUPIAL IN TEMPERATE FOREST OF PATAGONIA
By
Mariano Alberto Rodriguez-Cabal
May 2008
Chair: Lyn C. Branch Major: Wildlife Ecology and Conservation
Patterns and processes that shape the distribution and abundance of animals depend on the
scale at which they are evaluated. Understanding theses patterns and processes is fundamental
for the study of plant-seed disperser interactions. Most plants that depend on frugivorous
animals for seed dispersal are dispersed by several species of animals and, the majority of animal
dispersers consume the fruit of several plant species. In contrast to the weak mutual dependence
of most plants and their seed dispersers, some mutualisms in the temperate forest of Patagonia
appear to be obligate. The northern portion of the temperate forest harbors a unique plant –
animal interaction in which the seeds of the mistletoe Tristerix corymbosus, a proposed key
species, are dispersed solely by the endemic marsupial Dromiciops gliroides. I developed a set
of habitat models, in which model variables were defined a prior, to test the hypothesis that the
distribution and abundance of monito del monte is associated principally with abundance of
mistletoe in the temperate forest of Patagonia or that, alternatively, habitat structure and/or other
plants determine the distribution and abundance of this marsupial.
Results show that the distribution of monitos del monte at the broad scale was strongly
influenced by bamboo cover. However, at a smaller spatial scale, the abundance of monitos del
monte are explained by the abundance of mistletoe plants and fruits. The lack of dependence

9
among fruiting plants and their seed dispersers often is attributed to the high asymmetry in these
interactions and the important influence of habitat features. Conversely, my results showed that
the mistletoe–monito del monte interaction is more symmetric than many other plant–frugivore
interactions. This interaction is characterized by an obligate partnership of the mistletoe with the
marsupial, and a strong relationship between the abundance of monitos del monte and the
abundance of mistletoe.

10
CHAPTER 1 HABITAT ASSESSMENT FOR MONITOS DEL MONTE
Introduction
Distribution and abundance of species depends on the distribution and abundance of other
species with which they interact and habitat factors a various scales (Wiens,1989).
Understanding the patterns and processes that shape distribution and abundance is fundamental
for the study of plant-seed disperser interactions and biodiversity conservation. Seed dispersal
plays an important role in creating and maintaining diversity in plant communities (Schupp et al.,
2002). At large scales, appropriate habitat may be the most important factor limiting distribution
and abundance of frugivores (Wiens, 1989). Habitat characteristics such as availability of a
variety of food resources, nest sites, and distance to shelter, influence frugivores distribution and
could be play a critical role in regulating the abundance of these species (Fedriani, 2005). At a
small scale, habitat characteristics in which fruiting plants are surrounded may be more
important (Herrera, 1998). Previous studies have demonstrated that frugivores select plants or
good patches based on habitat features rather than on plant phenotypic traits (Herrera, 1998;
Fedriani, 2005). For example, frugivores could minimize predation risk by selecting sites with
high cover of vegetation to avoid predators during foraging (Howe, 1979). Thus, habitat factors
may limit the degree to which frugivorous animals track their food resource, and human impacts
on habitat of interacting species can have cascading effects.
Seed dispersal mutualisms influence patterns of seedling recruitment, plant demography,
and population persistence, and thus potentially have widespread effects on the rest of the
community (Janzen, 1980; Feinsinger, 1987; Bond, 1994; Levey and Benkman,1999; Herrera,
2002; Spiegel and Nathan, 2007; Rodriguez-Cabal et al., 2007). The majority of plants that
depend on frugivorous animals for seed dispersal are dispersed by several species of animals

11
and, most animal dispersers consume the fruit of several plant species (Herrera, 1998; 2002).
Mutual dependence of most plants and seed dispersers is weak, and asymmetry is common.
Asymmetry in plant–animal interactions is represented in terms of the effect of one species on
another species relative to the strength of the reciprocal effect (Vazquez and Aizen, 2004;
Vazquez et al., 2005; Bascompte et al., 2006; Vazquez et al., 2007). Asymmetry is particularly
evident for specialist plants, which tend to interact mostly with generalist animals (Vazquez and
Aizen, 2004; Bascompte et al., 2003; Bascompte et al., 2006). Generalist plants most often
interact with generalist and specialist animals (Vazquez and Aizen, 2004; Bascompte et al.,
2003; Bascompte et al., 2006).
In contrast to the weak mutual dependence of most plants and their seed dispersers, some
mutualisms in the temperate forest of Patagonia appear to be obligate. This system provides an
ideal opportunity to examine whether the distribution and abundance of an animal disperser
depends on the distribution and abundance of a specific fruit. A large proportion of the flora in
this temperate forest depends on mutualistic animals (Armesto et al., 1996; Aizen and Ezcurra,
1998; Aizen et al., 2002). This level of mutualism is one of the highest recorded for a temperate
ecosystem and is comparable to those from tropical forests (Willson et al., 1989; Willson, 1991).
Unlike the mutualism structure in tropical forests, in this temperate forest many species of flora
depend on a few mutualistic animals for their survival (Aizen et al., 2002).
The northern portion of the temperate forest harbors a unique triangle of keystone
mutualists comprised of a hummingbird (Sephanoides sephaniodes), a mistletoe (quintral,
Tristerix corymbosus), and the marsupial (monito del monte, Dromiciops gliroides, Amico and
Aizen, 2000). This hummingbird is responsible for pollinating nearly 20% of the endemic
species of woody flora in this biome (Armesto et al., 1996; Aizen et al., 2002). The mistletoe

12
blooms in winter and is the only source of nectar for the hummingbird during that period (Smith-
Ramírez, 1993; Aizen and Ezcurra, 1998). The monito del monte is the only known disperser of
the mistletoe in this temperate forest (Amico and Aizen, 2000). Moreover, the passage of the
seed through the marsupial’s gut is crucial to triggering germination (Amico and Aizen, 2000).
The isolation from other similar biotas since the disruption of Gondwana allowed the evolution
of monito del monte which is the only extant member from the ancient family Microbiotheriidae.
Because of the long evolutionary history of association of monitos del monte and the
mistletoe and the specific fruiting characteristics and distribution of mistletoes in this temperate
forest, I hypothesized that the distribution and abundance of monitos del monte and mistletoe
might be related closely. Mistletoe produces a large crop size every year and is the most
abundant fleshy fruit in the northern temperate forest of Patagonia (Aizen, 2003, Amico et al., in
press). Once mistletoe fruits are ripe, monitos del monte consume almost exclusively mistletoe
fruits (Rodriguez-Cabal et al., 2007). This mistletoe presents an aggregated distribution and,
therefore, may serve as high density resource patches for the monito del monte. Alternatively,
populations of monitos del monte may not track any single plant species as they are known to eat
and disperse seeds of 80% of the flora with fleshy fruit (Amico et al., in press). Because of the
diversity of fruits available, their populations maybe constrained largely by other habitat factors
such as vegetation cover and structural complexity of the habitat that influence nest site
availability, predation risk, and other aspects of their biology. In this study, I developed a set of
habitat models, in which model variables were defined a prior, to examine the hypothesis that the
distribution and abundance of monito del monte is associated principally with abundance of
mistletoe in the temperate forest of Patagonia or that, alternatively, habitat structure and/or other
plants determine the distribution and abundance of this marsupial. Although experiments are

13
required to fully determine the degree of mutual dependence between monitos del monte and the
mistletoe, landscape scale studies such as this provide a foundation for understanding
environmental constraints on these relationships.
Methods
Study Area
I conducted my research in Nahuel Huapi National Park (705,000 ha) and Llao-Llao
Municipal Reserve (1226 ha) in northwestern Patagonia. Nahuel Huapi National Park
encompasses a marked west-to-east rainfall gradient caused by the Andes, which form an
effective barrier to westerly air masses coming from the southern Pacific Ocean. The most xeric
forest site in my study, situated at the xeric boundary with the Patagonian steppe (Cerro Otto,
41°12' S, 71°19' W), has a mean annual precipitation of 1200 mm of which only 10.5 % falls
during the growing season. The western limit of my study located in Puerto Blest (41º 01' S, 71º
49' W) is an area of Valdivian rainforest, one of the rainiest places in the Patagonia (mean annual
precipitation, 4000 mm). Llao-Llao Municipal Reserve (41º 08' S, 71º 19' W) has intermediate
rainfall (mean annual precipitation, 1800 mm). Mean January temperature is 15ºC, and mean
July temperature is 3ºC.
The native forest vegetation in the study area belongs to the Subantartic biogeographical
region, which is distinctive from the Neotropical forest of the rest of South America (Cabrera,
1976). The most common trees are evergreen southern beech (Nothofagus dombeyi), a lenga
beech (Nothofagus pumilio) and a conifer (Austrocedrus chilensis). The understory is dominated
by the bamboo (Chusquea culeou) and the shrub (Aristotelia chilensis, here after A. chilensis).
That is the main host of the mistletoe in the study area (Mermoz and Martín, 1986). The two
forest layers are well differentiated with tree canopy reaching up to 40-m height and understory
reaching up to 5-m height.

14
I conducted presence–absence surveys of monitos del monte and examined habitat
variables at 16 sites in Nahuel Huapi National Park and 12 sites in the adjacent Llao-Llao
Municipal Reserve. In addition, trapping was conducted at the 12 sites in the Llao-Llao
Municipal Reserve to determine the relationship between abundance of monitos del monte and
abundance of mistletoe. Sites in Nahuel Huapi National Park were chosen randomly to
incorporate the range of habitats found in the study area. Potential sites in the Llao-Llao
Municipal Reserve were stratified by mistletoe density, and four sites were chosen randomly
within each of the following categories: high (>35 reproductive individuals of mistletoe), low
(<20 reproductive individuals), and no mistletoe. Sites were stratified on abundance of mistletoe
because mistletoe has a patchy distribution and a range of all other variables can be found within
each stratum defined by mistletoe density. I conducted most of the field work during the austral
summer (December 2006 –March 2007) because monitos del monte hibernate during winter and
the main fruiting and dispersal season of the mistletoe is summer (Aizen 2003).
Marsupial Presence - Absence
In order to facilitate rapid surveys of presence - absence of monitos del monte, the 28 study
sites were grouped by presence or absence of mistletoe. At sites with mistletoe, I examined all
plants < 15 cm DHB in each site for feces of monitos del monte that contained mistletoe seeds. I
concluded that monitos del monte were present if I found mistletoe seeds dispersed. If no
dispersed seeds were found in the site, I used trapping to determine the presence of monito del
monte. This technique also was used for sites with no mistletoe. Within each site, I placed a 5 x 5
grid (0.25 ha) of Tomahawk-style traps about 10-m apart. Each trap was placed between 1-2 m
above a ground in the shrub closest to the sample point. Traps were baited with apple and
banana. Because the objective was to document presence, once a monito del monte was trapped,
I stopped trapping at this site. Sampling was continued for four nights if no monitos del monte

15
were captured. Based on previous capture data for monitos del monte (Rodriguez-Cabal, 2003),
and following procedures in MacKenzie et al. (2002), probability of capture was 99.6% in four
night if monitos del monte were present. I concluded that monitos del monte were absent after
four nights of trapping if no marsupials were captured.
Marsupial Abundance
Abundance of the monito del monte was estimated using multiple capture-recapture
methods with trapping grids as described above. One trapping grid was placed at each of the 12
sites in Llao-Llao Municipal Reserve. Animals were trapped for four periods of four nights each
per site. Traps were checked daily and all trapped marsupials were marked with an individual
code based on ear perforations and released at the point of capture (for more detail see
Rodriguez-Cabal, 2003; Rodriguez-Cabal et al., 2007). Recaptures were noted in subsequent
trapping periods.
Habitat Variables
At each of the 16 sites in Nahuel Huapi National Park, a 5 x 5 grid was established with
points 10 m apart. I measured habitat variables in 5-m radius circular plots centered on every
second point (n = 13 per site). Similar sampling was conducted at sites trapped in Llao-Llao
Municipal Reserve with plots centered on every second trap. I measured the following variables
to represent the biological and physical structure of the habitat: number of reproductive
individuals and crop size of three understory plants that produce fleshy fruits (mistletoe, A.
chilensis, and Azara microphylla), cover and height of bamboo, complexity of habitat structure,
canopy cover, understory cover and the number of the two most common tree species (N.
dombeyi and Austrocedrus chilensis). Reproductive individuals were plants that presented sexual
structures (i.e., flowers or fruits). In addition for mistletoe, I counted the number of non-
reproductive individuals (2-4 years old), seedlings (those presenting the first two true leaves),

16
and number of seeds dispersed. To determine crop size of understory plants, I counted the
number of fruits on a subset of three branches dispersed in each crown and then estimated total
crop size by multiplying mean number of fruits per branch by total number of branches in the
crown for each plant. As with mistletoe, seeds of A. chilensis and A. microphylla are dispersed
by monitos del monte, and passage of their seeds through the digestive tract of a monito del
monte increases chance of germination (Amico et al., in press). Cover of bamboo was estimated
visually by summing the total area occupied by bamboo stems and leaves in each plot. Also, I
measured the height of the tallest bamboo stem in each plot. As a measure of habitat complexity
within each plot, I counted the number of contacts with a vertical pole (3-m height) for branches
5 – 10-cm in diameter and oriented < 45° relative to the ground. Good connectivity between
plants in the understory stratum could play a key role in the mobility of this animal, and more
seeds dispersed in monito del monte feces are found on branches with these characteristics
(Amico 2000). Percent cover of the canopy and understory were estimated using a densiometer
at the center of the circular plot. Finally, I counted the total number of the most common trees for
all individuals taller than four meters.
Data Analysis
I used multiple logistic regressions to determine which habitat variables predicted monito
del monte presence-absence at the 28 study sites, and I used multiple regressions to assess the
contribution of habitat variables to explaining abundance of monitos del monte at the 12 study
sites in Llao-Llao Municipal Reserve. Data were log-transformed when necessary to achieve
normality and reduce heteroscedasticity (Table 1-1). For both analyses, I only used explanatory
variables that did not correlate significantly after a Bonferroni correction (Appendix A-1 and A-
2; Tables 1-1, 1-2 and 1-3). I did not used bamboo height and understory cover because these
measures were correlated with bamboo cover and habitat complexity, respectively, and bamboo

17
cover and habitat complexity may be more directly related to the biology of monitos del monte,
particular for nest material and mobility. I did not include crop size of A. microphylla in
analyses because this understory plant did not produce fruits during the study.
Seven models were built to identify habitat variables related to presence – absence and
abundance of monitos del monte. Rather than examine all possible models, models were
developed a priori, that focused on mistletoe, other food plants, and habitat structure as key
factors in the distribution and abundance of this marsupial. I examined the following models: (1)
Global -- all habitat variables, (2) Food plants -- number of reproductive individuals of major
food plants that produce fleshy fruits eaten by monitos del monte, (3) Fruit abundance and
bamboo -- crop size of plants with fleshy fruits and cover of bamboo, (4) Mistletoe plants and
bamboo -- number of reproductive mistletoes and bamboo cover, (5) Mistletoe plants, (6)
Bamboo, (7) Habitat structure -- all habitat variables unrelated to food plants (Table 1-2 and 1-
3). Bamboo cover was included as a separate variable because this plant is an important source
of nest material (Jimenez and Rageot 1979), provides connectivity for arboreal movement, and
may inhibit predation by aerial predators. I used an approach that compared the suite of models
using Akaike’s information criteria (AIC), which allows direct comparison of models with
different number of parameters. AIC is calculated for a suite of models and the best fitting one
has the smallest AIC (Burnham and Anderson, 2002; Whittingham et al., 2005). I applied small-
sample version of AIC (AICc), because the ratio of number of observations to number of
parameters was <40. AICc was calculated for each model with the algorithm:
a) AIC = -2 log (L) + 2 K
b) AICc = -2 log (L) + 2 K + (2 K (K + 1))/ (n - K - 1)

18
where log (L) is the log-likelihood function evaluated for the maximum likelihood parameter
estimates, K is the number of parameters in the model and n is the number of observations. ∆AIC
represents the difference between the AICc score for each model and the AICc score of the best
model. The absolute value of AIC is unimportant; instead ∆AIC indicates relative support for the
models. ∆AIC values <2 indicate that models are similar. When ∆AIC was <2, I chose the most
parsimonious model as the best model (i.e., the model that used the least number of parameters).
To evaluate goodness-of-fit for the multiple logistic regressions models, I used Nagelkerke
R2. A true R2, which measures the variability in the dependent variable that is explained by a
linear regression models, cannot be calculated for logistic regression models (Hosmer and
Lemeshow, 1989). However, because deviance can be thought of as a measure of how poorly the
model fits (i.e., lack of fit between observed and predicted values), an analogy can be made to
sum of squares residual in ordinary least squares. This pseudo-R2 is designed to have similar
properties to the true R2. The pseudo-R2 is based on comparison of the likelihood of the current
model to the “null” model (one without predictors). A larger pseudo-R2 indicates that more of the
variation is explained by the model. Here, I reported the Nagelkerke R2 which is a modified
version of Cox and Snell R2 that ranges from 0 to 1 (Hosmer and Lemeshow 1989). I evaluated
the importance of significant independent variables in logistic regression using the standardized
regression coefficient β, which is the predicted change in odds for a unit increase in the
corresponding independent variable (Hosmer and Lemeshow, 1989). Values of β <1 correspond
to decreases in the odds with increases in the dependent variable; values >1 correspond to
increases in the odds with increases in the independent variable; and values close to 1 indicate
that unit changes in the independent variable do not affect the dependent variable.

19
To evaluate the goodness-of-fit for the multiple regression models for abundance of
monitos del monte, I calculated the R2 of the models. In multiple regressions, I evaluated the
importance of significant independent variables by calculating the standardized regression
coefficient β (Ganas and Robbins, 2005). The standardized regression coefficient β is used to
compare the magnitude of effect of different variables by describing the change in the dependent
variable (in SD units) caused by an increase in one unit (SD) of the independent variable, while
all the other independent variables are held constant.
In addition, I compared habitat variables between sites with and without monitos del monte
and at sites where I measured marsupial abundances using an ANOVA. I performed a Chi-square
test to compare the relationship between the presence of monitos del monte and mistletoe
presence at the 28 sites. For the 12 sites where I measured abundance of monitos del monte, I
report the minimum number alive for all sites to allow comparison among sites (Wilson et al.,
1996). Because of the low number of captures in sites without mistletoe, I could not use other
methods. Finally, I performed a regression analysis to examine the relationship between
abundance of monitos del monte and habitat variables that had significant explanatory power in
the multiple regression models. These analyses were conducted using SAS 9.13 and JMP IN for
Windows Ver. 5.1 (SAS Institute, Inc., Cary, NC).
Results
Marsupial Presence-Absence
Three of the seven multiple logistic regression models received substantial support as the
best predictive models of presence-absence of monitos del monte (Table 1-2). The Global model
perfectly predicted the presence-absence of monitos del monte (i.e., Nagelkerke R2 = 1). The
most parsimonious model (Fruit abundance and bamboo) and the Habitat structure model, which
was not ranked in the best models, also explained considerable variation in presence-absence of

20
monitos del monte (Table 1-2). Bamboo cover was the only significant predictor variable in the
top models (Table 1-2). The seventeen sites where I found marsupials had more bamboo cover
than sites without monito del monte (Table 1-1). Monito del monte was recorded 91% of the
sites that had more than 10% bamboo cover and in only 35% of the sites with less than 10%
cover. The presence-absence of mistletoe and monito del monte were not related at the 28 sites
(Chi-square = 2.06, df = 1, P = 0.15). Five of the 17 sites where I recorded presence of monito
del monte did not have mistletoe. Five of the eleven sites where monitos del monte were absent
had reproductive mistletoe plants, but I did not find mistletoe seeds dispersed, seedlings or non-
reproductive mistletoe (2-4 years old). The lack of seeds, seedlings and non-reproductive
individuals indicates that there has not been recruitment at least in the last four years. The other
12 sites had mistletoe seeds dispersed in monito del monte feces and seedlings.
Marsupial Abundance
The estimated minimum number alive of marsupials on study plots ranged between 0 and
45 individuals. At eleven of the 12 sites where I measured abundances, I captured monitos del
monte in the first four nights of trapping, in spite of the high variability in local abundance of
this species. Two regression models were candidates for the best predictive model for abundance
of monitos del monte, although several additional models also explained high amounts of
variation in abundance of these marsupials (Table 1-3). The number of reproductive mistletoe or
crop size of mistletoe were significant variables in five of the seven models. Habitat complexity,
bamboo cover, and crop size of A. chilensis appear as significant variables in models that were
not supported as the best models, but P-values for these variables were lower (Table 1-3).
Marsupial abundance increased when the number of reproductive mistletoe, crop size of
mistletoe and A. chilensis, or habitat complexity increased (see regression coefficients β, Table
1-3). In contrast, the relationship between bamboo cover and monito del monte abundance was

21
negative. At these 12 sites, bamboo cover was lowest where the abundance of mistletoe was
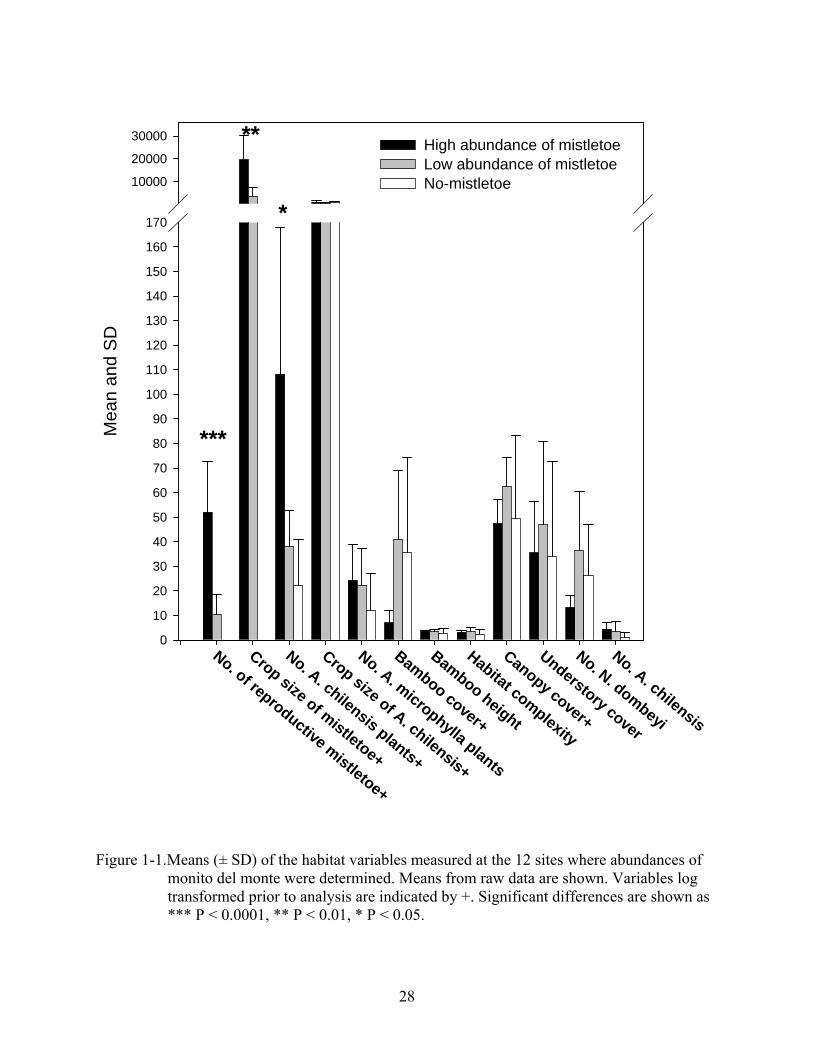
highest, but the over all correlation of these variables is not significant (Figure 1-1, Appendix A-
2). Linear regression analysis of the abundance of monitos del monte and the five significant
variables indicates that abundance of monitos del monte is better explained by number of
reproductive mistletoes and crop size of mistletoe than crop size of A. chilensis or structural
variables of the habitat (Figure 1-2). Abundance of monito del monte also correlated with
abundance of non-reproductive-mistletoe plants (R2 = 0.83; P < 0.0001) and with mistletoe
seedlings (R2 = 0.70; P < 0.001).
Discussion
High asymmetry between fleshy fruit plants and seed dispersers has been emphasized as a
mechanisms limiting coevolution of these plants and seed dispersers (Herrera, 1985; 1993;
Bascompte et al., 2003; Vazquez and Aizen, 2004). Other recent work also suggests that habitat
features constrain these species interactions (Fedriani, 2005). In this study at the scale of the
study area, habitat features played an important role in determining presence of monitos del
monte whereas presence or absence of specific food plants and mistletoe did not have an
appreciable effect. Bamboo cover was the most important habitat feature for the presence of
marsupial populations. Leaves of bamboo have been described as a principal resource for nest
building for this marsupial and because monitos del monte are arboreal, bamboo can increase the
mobility of this marsupial in forest understory where fruits and nests are located (Jimenez and
Rageot, 1979; Mann, 1955). However, presence data alone can overestimate or underestimate the
importance of habitat characteristics, because a species can be present but have very different
abundances in different sites (Royle and Nichols, 2003; Bailey et al., 2004). Although the
distribution of monitos del monte at the broad scale was influenced strongly by bamboo cover,
surveys conducted at Llao-Llao Municipal Reserve did not show a significant association

22
between abundance of this marsupial and cover of bamboo, except in one non-competitive model
where bamboo cover was related negatively related to abundance of monitos del monte. This
negative relationship may have occurred because the lowest amount of bamboo was in areas with
the highest amount of mistletoe (Figure 1-1). The general lack of importance of bamboo for
explaining abundance of monitos del monte in Llao-Llao Municipal Reserve could be explained
by the fact that when the minimum requirement for bamboo cover is met, which based on my
results may be about 10%, mistletoe plays a more important role than bamboo in regulating the
number of marsupials the habitat can support.
Numerous studies have addressed the link between fruit abundance and frugivore
abundance but clear examples of a single plant regulating the abundance of seed dispersers are
scarce (Levey, 1988; Herrera, 1998; 2002; Loiselle and Blake, 1991; Jordano, 1994; 2000;
Malizia, 2001; but see Moegenburg and Levey, 2003). My data suggest that the relationship
between the abundance of the monito del monte and mistletoe may be stronger and more
symmetric than in many other plant frugivory systems (Herrera, 1998; Moegenburg and Levey,
2003). Even though the monito del monte consumes and disperses 80% of the flora with fleshy
fruits, a significant correlation occurred at the Llao-Llao Municipal Reserve between the
abundance of monito del monte and the abundance of mistletoe plants and fruits. This
relationship was much stronger than with other fruiting species such as A. chilensis. Sites with
high abundance of mistletoe had from 2 to 5 times more monitos del monte that sites without
mistletoe. The strong correlation between mistletoe abundance and abundance of monito del
monte could occur because mistletoe directly affects the abundance of monitos del monte or
because both species are responding to another habitat variable not measured in this study.
Experiments are needed to resolve this issue. However, other data also support the conclusions of

23
the strong link between monitos del monte and mistletoe. Once mistletoe fruits are ripe, monitos
del monte change their diet from generalist to consume mistletoe fruits almost exclusively
(Rodriguez-Cabal et al., 2007).
The distribution and abundance of monitos del monte found in this study may be linked to
different habitat variables operating at different scales. This arboreal marsupial lives mainly in
the understory. At a large scale, the distribution of this marsupial may be limited by bamboo
cover, which increases nest sites and/or nest building material and connectivity to where food is
more abundant above ground. At a smaller spatial scale, the abundance of monitos del monte
tracked abundance of mistletoe. This spatial pattern may result from progressive gathering of
fine-grain foraging information by individuals in those forest sectors with high abundance of
reproductive mistletoe.
One of the most common explanations for lack of coevolution in plant-seed dispersal
interactions is the high level of asymmetry found (Bascompte et al., 2003). Specialized species
will have limited chance for coevolution because they are asymmetrically influenced by their
generalized species (Vazquez et al., 2007). Although the monito del monte could exert strong
selection on reproductive traits of this mistletoe as its only seed disperser in the temperate forest,
there is no evidence to support the coevolution of traits in this interaction. Furthermore, the
evidence points in the other direction, in which most of the relevant features of the mistletoe in
the temperate forest of Patagonia like the green color of the mature fruits, a color associated with
mammalian dispersers (van der Pijl, 1982), is driven only by environmental factors, such as
period of day light and temperatures (Amico, 2007; Amico et al., 2007). In the Chilean shrubland
this mistletoe species has fruits that are yellow when ripe, and birds disperse the seeds (Amico,
2007). As in the Australian mistletoe (Loranthaceae and Viscaceae) and mistletoebird (Dicaeum

24
hirundinaceum) mutualisms (Reid, 1987), the mistletoe-monito del monte interaction is an
ecological association rather than a coevolved interaction.

25
Table 1-1. Means (± SD) of habitat variables and summary ANOVA for comparison of means for sites where monitos del monte were present or absent. Means from raw data are shown. Variables that were log-transformed prior to analysis are indicated by †. Significant P-values are indicated by *.
Mean ± SD
Habitat variables Present (n = 17)
Absent (n = 11) F df P-value
No. reproductive mistletoe† 1.68 ± 1.88 1.09 ± 1.56 0.958 1 0.34Crop size of mistletoe† 610.17 ± 725.25 468.68 ± 927.70 2.263 1 0.15No. of A. chilensis plants† 4.06 ± 4.05 1.35.63 ± 1.60 5.322 1 0.03*Crop size of A. chilensis † 62.11 ± 57.30 47.90 ± 146.57 2.263 1 0.15No. of A. microphylla plants 0.73 ± 1.01 0.69 ± 2.29 0.013 1 0.91Bamboo cover (%)† 24.97 ± 27.50 3.57 ± 8.23 11.625 1 0.002*Bamboo height (m) 3.13 ± 1.57 0.98 ± 1.51 11.234 1 0.003*Habitat complexity 3.44 ± 0.93 2.48 ± 1.36 4.87 1 0.04*Understory cover (%) 55.10 ± 23.39 54.24 ± 36.50 0.006 1 0.94Canopy cover (%)† 54.70 ± 20.60 46.28 ± 35.79 2.87 1 0.10Average number of trees 2.94 ± 2.21 1.12 ± 1.54 5.66 1 0.03*

26
Table 1-2. Summary of the seven logistic regression models constructed to determine which habitat variables predicted presence-absence of monitos del monte at the 28 study sites. Competitive models (∆AIC <2) and the significant variables in each model are shown in bold.
Predicted correct (%)
Models Variables AIC AICc ∆AIC df Nagelkerke
R2 β Absent Present Global 20.124 30.124 0.641 9 1 100 100
All variables Food plants 39.129 40.129 10.646 3 0.28 72.7 82.4
No. of reproductive mistletoe No. of A. chilensis plants 45.37 No. of A. microphylla plants
Fruit abundance and 28.483 29.483 0 3 0.62 81.8 88.2 bamboo Crop size of mistletoe
Crop size of A. chilensis Bamboo cover 13.37
Mistletoe plants and 30.817 31.297 1.814 0.49 72.7 82.4 bamboo No. of reproductive mistletoe
Bamboo cover 11.29 Mistletoe plants 40.502 40.656 11.173 1 0.05 0 100
No. of reproductive mistletoe Bamboo 31.746 31.900 2.417 1 0.40 72.7 82.4
Bamboo cover 8.27 Habitat structure 30.517 32.256 2.773 4 0.62 81.8 88.2
Bamboo cover 10.99 Habitat complexity Canopy cover Average number of trees

27
Table 1-3.Summary of the seven regression models constructed to determine which habitat variables predicted abundances of monito del monte at 12 study sites. Competitive models (AIC <2) and the significant variables in each model are shown in bold. Model variables are the same as in Table 1-2 except for habitat structure. For this analysis, number of N. dombeyi and Austrocedrus chilensis were used instead of average number of trees.
Models Significant variables AIC AICc ∆AIC df R2 β P-value Global 44.623 134.623 86.154 9 0.87 No. of reproductive mistletoe 1.50 <0.0001 Habitat complexity 2.02 0.05Food plants 48.070 51.070 2.600 3 0.80 No. of reproductive mistletoe 0.72 <0.0001Fruit abundance and 48.292 51.292 2.823 3 0.83 bamboo Crop size of mistletoe 0.75 0.0005 Crop size of A. chilensis 0.45 0.04Mistletoe plants and 48.065 49.398 0.929 2 0.83 bamboo No. of reproductive mistletoe 0.95 <0.0001Mistletoe 48.069 48.469 0 1 0.80 No. of reproductive mistletoe 0.89 <0.0001Bamboo 65.311 65.711 17.242 1 0.01 Habitat structure 54.068 64.068 15.599 5 0.67 Bamboo cover -2.02 0.01
28
No. of reproductive mistletoe+
Crop size of mistletoe+
No. A. chilensis plants+
Crop size of A. chilensis+
No. A. microphylla plants
Bamboo cover+
Bamboo height
Habitat complexity
Canopy cover+
Understory cover
No. N. dombeyi
No. A. chilensis
Mea
n an
d S
D
0
10
20
30
40
50
60
70
80
90
100
110
120
130
140
150
160
170
10000
20000
30000 High abundance of mistletoeLow abundance of mistletoeNo-mistletoe
***
**
*
Figure 1-1.Means (± SD) of the habitat variables measured at the 12 sites where abundances of
monito del monte were determined. Means from raw data are shown. Variables log transformed prior to analysis are indicated by +. Significant differences are shown as *** P < 0.0001, ** P < 0.01, * P < 0.05.

29
Bamboo cover (log)
0 1 2 3 40
10
20
30
40
50R2 = 0.01, P = 0.80
No. of reproductive mistletoe (log)
0.0 0.5 1.0 1.5 2.0
No.
mon
itos
del m
onte
0
10
20
30
40
50R2 = 0.80, P < 0.001
Crop size of A. chilensis (log)
0 1 2 3 4 50
10
20
30
40
50R2 = 0.22, P = 0.13
Crop size of mistletoe (log)
0 1 2 3 4 5 6 7 80
10
20
30
40
50R2 = 0.72, P < 0.001
Habitat complexity
0 1 2 3 4 50
10
20
30
40
50R2 = 0.26, P = 0.09
Figure 1-2. Simpler linear regression analysis of the number of monitos del monte vs. the significance predictor variables of the multiple regression models. The coefficient of determination (R2) and P-value are shown.

30
CHAPTER 2 CONSERVATION IMPLICATIONS
The interaction between this mistletoe and its marsupial disperser may have consequences
at the community and ecosystem levels and habitat changes that impact either of these species
could have indirect cascading effects throughout different interaction webs. This mistletoe is
considered a keystone species due to the production of sugar-rich nectar and protein-rich fruits
during periods of food scarcity. This mistletoe also increases mortality of host trees and could
play an important role in plant community diversity. As the only known disperser for the
mistletoe, the monito del monte is important in maintaining these processes. Habitat
fragmentation disrupts the interaction between the mistletoe and monitos del monte (Rodriguez-
Cabal et al., 2007). Fragmentation negatively affects marsupial abundance, fruit removal, seed
dispersal, and seedling recruitment. The local extinction of monitos del monte has been
associated with the complete disruption of mistletoe seed dispersal (Rodriguez-Cabal et al.,
2007). The reason for the local extinction of monitos del monte in fragmented habitats is
unknown. One possible mechanism is reduction of bamboo in fragments results in loss of nest
sites and connectivity. Grazing by cattle and other introduced ungulates significantly alters
habitat structure and composition of the forest, including reduction of bamboo (Veblen et al.,
1992). Effects of grazing are particularly severe in forest fragments where introduced ungulates
have easy access to entire patch.
In this temperate forest, the bamboo blooms and dies at approximately 40-60 years
intervals. Although this is a natural process, the die-off of the bamboo also likely has important
consequences for populations of the monito del monte by reducing nest sites and connectivity.
Loss of bamboo within habitat fragments may have particularly severe consequences because

31
these patches are isolated, reducing the probability of demographic rescue and recolonization
following regeneration of bamboo within the patch.

32
CHAPTER 3 CONCLUSION
In the temperate forest of southern South America, the northern portion (to 41oS) has the
highest biodiversity and number of endemic species, the lowest proportion of protected areas,
and the highest rate of disturbance in this forest (Armesto et al., 1998, Rodriguez-Cabal et al., in
press). The biodiversity of this temperate forest currently is threatened by the highest rates of
destruction, fragmentation, and degradation ever experienced by this system, with more than
120,000 ha of native forest destroyed or degraded annually (Lara et al., 1996).
The alteration of natural habitats through human activities is the principal cause of the
decline and extinction of natural populations (Gaston and Spicer, 2004). Long-term plans to
conserve biodiversity must not only focus in maintaining the elements that form biodiversity but
also on the interactions among these elements (Vazquez and Simberloff, 2003). Understanding
the species and processes that shape ecosystems is fundamental for effective conservation
(Wayne et al., 2006). No previous study has investigated explicitly factors that determine this
austral marsupial’s distribution and abundance or the causes of its present decline. This study
constitutes the first detailed ecological research focused on the identification of the habitat
variables that play the most important role in maintaining the marsupial’s populations. My work
demonstrates that both habitat factors (bamboo) and interacting species (mistletoe) are important
in determining the distribution and abundance of this species.

33
APPENDIX CORRELATION ANALYSIS
Table A-1. Correlation coefficients (r) between habitat variables in the presence-absence survey (n = 28). Significant correlations
after the Bonferroni correction are indicated by * (P < 0.0009).
Crop size of mistletoe
No. of A. chilensis plants
Crop size of A. chilensis
No. of A. microphylla plants
Bamboo cover (%)
No. of reproductive mistletoe 0.89* 0.56 0.15 0.17 -0.19 Crop size of mistletoe 0.50 0.10 0.18 -0.17 No. of A. chilensis plants 0.66* 0.42 0.29 Crop size of A. chilensis 0.10 0.39 No. of A. microphylla plants 0.33 Bamboo cover (%) Bamboo height (m) Habitat complexity Understory cover (%) Canopy cover (%)
Table A-1.
Bamboo height (m)
Habitat complexity
Understory cover (%) Canopy cover (%)
Average no. trees
No. of reproductive mistletoe 0.11 0.29 -0.004 - 0.38 -0.24 Crop size of mistletoe 0.02 0.01 -0.01 - 0.44 -0.28 No. of A. chilensis plants 0.47 0.02 -0.23 -0.07 0.13 Crop size of A. chilensis 0.40 0.19 -0.004 -0.06 0.16 No. of A. microphylla plants 0.36 -0.06 - 0.39 -0.05 0.004 Bamboo cover (%) 0.88* 0.35 -0.11 0.37 0.43 Bamboo height (m) 0.20 -0.29 0.29 0.40 Habitat complexity 0.73* 0.43 0.44 Understory cover (%) 0.23 0.15 Canopy cover (%) 0.88*

34
Table A-2. Correlation coefficients (r) between habitat variables in the abundance survey (n = 12). Significant correlations after the Bonferroni correction are indicated by * (P < 0.0006).
Crop size of
mistletoe No. of A.
chilensis plants Crop size of A.
chilensis No. of A.
microphylla plants Bamboo cover
(%) No. of reproductive mistletoe 0.94* 0.78 0.30 0.09 -0.28 Crop size of mistletoe 0.70 0.16 0.12 -0.13 No. of A. chilensis plants 0.41 0.33 -0.32 Crop size of A. chilensis 0.05 0.46 No. of A. microphylla plants 0.10 Bamboo cover (%) Bamboo height (m) Habitat complexity Understory cover (%) Canopy cover (%) N. dombeyi Table A-2.
Bamboo
height (m) Habitat
complexity Understory cover (%)
Canopy cover (%) N. dombeyi
A. chilensis
No. of reproductive mistletoe 0.33 0.28 0.13 0.21 -0.35 0.46 Crop size of mistletoe 0.39 0.34 0.18 0.28 0.20 0.55 No. of A. chilensis plants 0.63 0.39 0.11 0.56 -0.02 0.53 Crop size of A. chilensis 0.38 0.70 0.67 0.36 -0.14 -0.42 No. of A. microphylla plants 0.32 0.00 -0.23 0.10 0.18 0.09 Bamboo cover (%) 0.65 0.70 0.71 0.68 0.46 -0.26 Bamboo height (m) 0.67 0.50 0.96* 0.33 0.30 Habitat complexity 0.89* 0.58 -0.16 -0.08 Understory cover (%) 0.40 -0.16 -0.35 Canopy cover (%) 0.51 0.27 N. dombeyi 0.17

35
LIST OF REFERENCES
Aizen, M. A., 2003. The relative influence of animal pollination and seed dispersal on flowering time in a winter flowering mistletoe. Ecology 84, 2613-2627.
Aizen, M. A., Ezcurra C., 1998. High incidence of plant-animal mutualisms in the woody flora
of the temperate forest of southern South America: biogeographical origin and present ecological significance. Ecología Austral 8, 217-236.
Aizen, M. A., Vázquez, D. P., Smith-Ramírez, C., 2002. Historia natural de los mutualismos
planta-animal del Bosque Templado de Sudamérica Austral. Revista Chilena de Historia Natural 75, 7 9-97.
Amico, G. C., 2000. Dispersión del quintral (Tristerix corymbosus) por el Monito del Monte
(Dromiciops australis). Master Theses. Universidad Nacional del Comahue, Bariloche, RN Argentina.
Amico, G. C., 2007. Variación geográfica en la coloración de los frutos del muérdago Tristerix
corymbosus (Loranthaceae): efecto del ambiente, de los dispersores de semillas y de los hospedadores. Universidad Nacional del Comahue, Bariloche, RN, Argentina.
Amico, G. C., Aizen, M. A., 2000. Marsupial dispersal of mistletoe seeds in temperate South
America. Nature 408, 929-930. Amico, G. C., Vidal-Russell, R., Nickrent, D., 2007. Phylogenetic relationships and ecological
speciation in the mistletoe Tristerix (Loranthaceae): the influence of pollinators, dispersers, and hosts. American Journal of Botany 95, 558-567.
Amico, G. C., Rodríguez-Cabal, M. A., Aizen, M. A., 2008. Diet, frugivory and the potential
seed-dispersing role of the marsupial, Dromiciops gliroides (Microbiotheriidae). Mammalian Biology In press.
Armesto, J. J., Smith-Ramírez, C., Sabag, C., 1996. The importance of plant-bird mutualisms in
the temperate rainforest of southern South America. In: Lawford R.G., Alaback P.B. and Fuentes E. (Eds.), High latitude rainforests and associated ecosystems of the west coast of the Americas. Springer, New York pp. 248-265.
Armesto, J.J., R. Rozzi, C. Smith-Ramirez, Arroyo, M.T.K., 1998. Conservation targets in South
American temperate forests. Science 282, 1271-1272. Bailey, L. L., Simons, T. R., Pollock, K. H., 2004. Estimating site occupancy and species
detection probability parameters for terrestrial salamanders. Ecology 14, 692-702. Bascompte, J., Jordano, P., Melian, C. J., Olesen, J. M., 2003. The nested assembly of plant-
animal mutualistic networks. Proceedings of Natural Academic of Science USA 100, 9383-9387.

36
Bascompte, J., Jordano, Olesen, J. M., 2006. Asymmetric coevolutionary networks facilitate biodiversity maintenance. Science 312, 431-433.
Bond, W. J., 1994. Do mutualisms matter? Assessing the impact of pollinator and disruption
plant extinction. Philosophical transaction of the Royal Society of London 344, 83-90. Burnham, K. P., Anderson, D. R., 2005. Model selection and multimodel inference. Springer,
New York, NY. Cabrera, L. A., 1976. Regiones fitogeográficas argentinas. Editorial ACME, Buenos Aires. Fedriani, J. M., 2005. Do frugivorous mice choose where or what to feed on? Journal of
Mammalogy 86, 576-586. Feinsinger, P., 1987. Approaches to nectarivore-plant interactions in the New World. Revista
Chilena de Historia Natural 60, 285-319. Gaston, K. J., Spicer, J. I., 2004. Biodiversity: An Introduction. Blackwell Publising, Melbourne. Herrera, C. M., 1985. Determinants of plant-animal coevolution: the case of mutualistic dispersal
of seeds by vertebrates. Oikos 44, 132-141. Herrera, C. M., 1993. Selection on floral morphology and environmental determinants of
fecundity in a hawk moth-pollinated violet. Ecological Monograph 63, 251-275. Herrera, C. M., 1998. Long-term dynamics of Mediterranean frugivorous birds and fleshy fruits:
a 12-yr study. Ecological Monograph 68, 511-538. Herrera C. M., 2002. Seed dispersal by vertebrates. In: Herrera C.M., Pellmyr O. (Eds.), Plant-
animal interactions: an evolutionary approach. Blackwell, Oxford, UK pp. 185–208. Hosmer, D. W., Lemeshow, S.,1989. Applied Logistic Regression. John Wiley & Sons, Inc, New
York, USA. Howe, H. F., 1979. Fear and frugivory. American Naturalist 114, 925-931. Janzen, D. H., 1980. When is it coevolution? Evolution 34, 611-612. Jimenez, J., Rageot, R., 1979. Notas sobre la biología del monito del monte (Dromiciops
australis Philippi 1893). Anales del Museo de Historia Natural 12, 83-88. Jordano, P., 1994 . Spatial and temporal variation in the avian-frugivore assemblage ofn Prunus
mahaleb: patterns and consequences. Oikos 71, 479-491. Jordano, P., 2000. Fruits and frugivory In: Fenner, M. (Ed.), Seeds: the ecology of regeneration
in plant community. CAB International Publishing, Wallingford, UK pp 125-165.

37
Lara, A., C. Donoso, Aravena, J. C., 1996. La conservación del bosque nativo en Chile: problemas y desafíos. In: J.J. Armesto, C. Villagrán and M.T.K. Arroyo, (Eds.), Ecología de los bosques nativos de Chile. Editorial Universitaria, Santiago de Chile pp. 335-362.
Levey, D. J., 1988. Spatial and temporal variation in Costa Rican fruit and fruit-eating bird
abundance. Ecological Monograph 58, 251-269. Levey, D. J., Benkman, C. W., 1999. Fruit-seed disperser interactions: timely insights from a
long-term perspective. Trends in Ecology and Evolution 14, 41-43. Loiselle, B. A., Blake, J. G., 1991. Potential consequences of local frugivore extinction for plant
population in neotropical lowland wet forest. In: Levey D. J., Silva W. R. and Galetti M. (Eds.), Seed dispersal and frugivory: ecology, evolution and conservation. CAB International Press, Oxfordshire, UK pp. 397-406.
MacKenzie, D. I., Nichols, J. D., Hines, J. E., Knutson, M. G., Franklin, A. B., 2002. Estimating
site occupancy, colonization, and local extinction when a species is detected imperfectly. Ecology 84, 2200-2207.
Malizia, L. R., 2001. Seasonal fluctuations of birds, fruits, and flowers in a subtropical forest of
Argentina. Condor 103, 45-61. Mann, G. F., 1955. Monito del monte Dromiciops australis Philippi. Investigación Zoológica
Chilenas 2, 159-166. Mermoz, M., Martín, C., 1986. Mapa de vegetación del Parque y la Reserva Nacional Nahuel
Huapi. Secretaría de Ciencia y Técnica de la Nación, Delegación Regional Patagonia, S. C. de Bariloche, Argentina.
Moegenburg, S. M., Levey, D. J. ,2003. Do frugivores respond to fruit harvest? An experimental
study of short-term responses. Ecology 84,2600-2612. Reid, N., 1987. The mistletoebird and Australian mistletoe: co-evolution or coincidence? Emu
87, 130-131. Rodríguez-Cabal, M. A., 2003. Habitat fragmentation’s effect on a mutualism of dispersal in the
temperate forest of South America. Universidad Nacional del Comahue, Bariloche, RN, Argentina.
Rodríguez-Cabal, M. A., Aizen, M. A., Novaro, A. J., 2007. Habitat fragmentation disrupts a
plant-disperser mutualism in the temperate forest of South America. Biological Conservation 139, 195-202.
Royle, J. A., Nichols, J. D., 2003. Estimating abundance from repeated presence-absence data or
point counts. Ecology 84, 777-790.

38
Schupp, E.W., Milleron, T., Russo, S.E., 2002. Dissemination limitation and the origin of species-rich tropical forests. In: Levey D.J., Silva W.R. and Galetti M. (eds.) Seed Dispersal and Frugivory: Ecology, Evolution, and Conservation. CAB International, Wallingford, Oxon, UK, pp. 19-43.
Smith-Ramirez, C., 1993. Los picaflores y su recurso floral en el bosque templado de la isla de
Chiloé, Chile. Revista Chilena de Historia Natural 66, 65-73. Spiegel, O., Nathan, R., 2007. Incorpating dispersal distance into the disperser effectiveness
framework: frugivorous birds provide complementary dispersal to plants in a patchy environment. Ecology Letters 10, 718-728.
van der Pijl, L., 1982 Principles of dispersal in higher plants. Springer, Berlin, Germany. Vazquez, D. P., Simberloff, D,. 2003. Changes in interaction biodiversity induced by an
introduced ungulate. Ecology Letters 6, 1077-1083. Vazquez, D. P., Aizen, M. A., 2004. Degree distribution in plant-animal mutualistic networks:
forbidden links or random interactions. Oikos 108, 421-426. Vazquez, D. P., Morris, W. F., Jordano, P., 2005. Interaction frequency as surrogate for the total
effect of animal mutualists on plants. Ecology Letters 8, 1088-1094. Vazquez, D. P., Melian, C. J., Williams, N. M., Bluthgen, N., Krasnov, R., Poulin, R., 2007.
Species abundance and asymmetric interaction strength in ecological networks. Oikos 116, 1120-1127.
Veblen, T V., Mermoz, M., Martin, C., Kitzberger, T., 1992. Ecological impacts of introduced
animals in Nahuel Huapi National Park, Argentina. Conservation Biology 6, 71-83. Wayne, A. F., Cowling, A., Lindenmayer, D. B., Ward, C. G., Vellios, C. V., Donnelly, C. F.,
Calver, M. C., 2006. The abundance of a threatened arboreal marsupial in relation to anthropogenic disturbance at local and landscape scales in Mediterranean-type forest in south-western Australia. Biological Conservation 127, 463-476.
Wiens, J. A., 1989. Spatial scaling in ecology. Functional Ecology 3, 385-397. Whittingham, M. J., Swetnam, R. D., Wilson, J.D., Chamberlain, D. E., Freckleton, R. P., 2005.
Habitat selection by yellowhammers Emberiza citrinella on lowland farmland at two spatial scales: implications for conservation management. Journal of Applied Ecology 42, 270-280.
Wilson, D. E., Cole, F. R., Nichols, J. D., Rudran, R., Foster, M., 1996. Measuring and
monitoring biological diversity. Standard methods for mammals. Smithsonian Institution Press, Washington, D. C, USA.

39
Willson, M. F., Irvine, A. K., Walsh, N.G., 1989. Vertebrate dispersal syndromes in some Australian and New Zealand plant communities, with geographic comparisons. Biotropica 21, 133-147.
Willson, M. F., 1991. Dispersal of seeds by frugivorous animals in temperate forests. Revista
Chilena de Historia Natural 64, 537-554.

40
BIOGRAPHICAL SKETCH
Mariano Alberto Rodriguez Cabal was born in Buenos Aires, Argentina in 1976. His
interest in biology began when he very young. Nature has always exhibited a powerful attraction
for him. He studied in a religious high school and obtained a degree in economics. When he
finished his secondary studies, he moved to San Carlos de Bariloche in the foothills of the
Andes, in western Argentina. The magnificent environment around this city, which is within a
national park, led him unequivocally to pursue university studies in biology.
In 1997, he began studying Biology at Universidad Nacional del Comahue. In July 2003,
he graduated as a “Licenciado en Ciencias Biológicas.” Under the guidance of Dr. Marcelo
Aizen and Dr. Andrés Novaro, he conducted his undergraduate thesis research on the effects of
habitat fragmentation on a dispersal mutualism in the temperate forest of southwestern
Argentina. After two years of field work, he completed and defended his thesis, obtaining the
highest grade.
In 2004, he received a Fulbright Scholarship to study abroad. In the next year, he moved
to Florida, USA, where he started his master’s studies at the University of Florida in the
Department of Wildlife Ecology and Conservation. During those years worked under the
supervision of Professor Lyn C. Branch. This work was recognized with three grants. Twice he
earned the Outstanding Academic Achievement Award at UF. In December 2007, he defended
his master’s thesis. He made a firm decision to devote himself to scientific research. He hopes to
continue his doctoral education in the United States of America. On completing his studies, he
will return to his country to contribute to the welfare of his people and the conservation of
Argentina’s biodiversity through teaching and research.