genetic instability and strain degeneration in ... · phenotypically uniform, and most variants...
TRANSCRIPT

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, July 1993, p. 2220-2228 Vol. 59, No. 70099-2240/93/072202-09$02.00/0Copyright ) 1993, American Society for Microbiology
Genetic Instability and Strain Degeneration inStreptomyces rimosus
BIRGIT GRAVIUS,1 TAJANA BEZMALINOVIC,2 DASLAV HRANUELI,3 AND JOHN CULLUM`*LB Genetik, Universitat Kaiserslautern, Postfach 3049, D-67653 Kaiserslautem, Germany, 1 and Faculty of
Food Technology and Biotechnology2 and PLIVA Research Institute,3 Zagreb, Republic of Croatia
Received 11 December 1992/Accepted 15 April 1993
During a strain selection program to improve oxytetracycline production in Streptomyces rimosus R6, isolatesthat showed extreme morphological instability appeared. Propagation via spores gave much higher instabilitythan did propagation via mycelial fragments. Five phenotypic traits were affected: sporulation, pigmentation,colony morphology, oxytetracycline production, and oxytetracycline resistance. The variants were classified onthe basis of oxytetracycline resistance into three classes. Class I variants (99%Yo of variants) showed parentallevels of resistance but were very heterogeneous regarding the other phenotypes. No DNA rearrangements weredetected in primary class I variants. Class II variants (1% of variants, oxytetracycline sensitive) werephenotypically uniform, and most variants carried the same large deletion of ca. 455 kb, including theoxytetracycline resistance gene otrB. Class III variants (0.1% of variants, increased oxytetracycline resistance)were phenotypically uniform and overproduced a brown pigment and oxytetracycline. Most of these variantsalso showed a reproducible large-scale DNA rearrangement, which probably included deletion and a low-levelreiteration (three or four copies) of a DNA fragment. "Revertants" of some class I variants show a similar DNArearrangement to the class III variants, but there is extensive reiteration of sequences of about 200 kb,including the otrB gene. The significance of these results for the problem of strain degeneration and over-production of antibiotics is discussed.
Genetic instability has been observed in many Streptomy-ces strains (8, 21, 23, 35). Most of the strains that have beenstudied at the molecular level were laboratory strains, whichwere not selected for high antibiotic production. In industrialstrains there is often a phenomenon of strain degeneration,which consists of genetic changes deleterious for importantproperties such as antibiotic yield. It has been suggested thatstrain degeneration in Streptomyces species is also a facet ofgenetic instability (25). Among others, Streptomyces aureo-faciens (the producer of chlortetracycline) and S. rimosus(the producer of oxytetracycline) are commercially impor-tant species that are subject to genetic instability (reference8 and references therein). For example, in S. rimosus,oxytetracycline-sensitive variants appear as a result of largedeletions, which may remove all the oxytetracycline synthe-sis genes (6, 31). The only industrial strain in which geneticinstability has been studied in detail at the molecular level isthe tylosin producer S. fradiae. Genomic alterations in S.fradiae were first observed in tylosin-sensitive, tylosin-nonproducing spontaneous mutants obtained after proto-plast regeneration or selection for spectinomycin resistance;these showed high-copy-number tandem repeats of an am-plifiable unit of DNA (10). Further studies revealed that themutants contained deletions involving tylosin productionand resistance genes (7). The cluster of tylosin biosyntheticgenes is interrupted by a structurally unstable segmentcontaining four repeated sequences (3). Although the influ-ence of genetic instability on tylosin production has not beendiscussed, such a structure might be a source of straindegeneration in this industrially important species.
S. rimosus strains have been subjected to intensive selec-tion programs for yield improvement, and genetic maps havebeen established for the two strains, R6 and R7 (ATCC
* Corresponding author.
10970), by using conjugation and protoplast fusion (1, 18,26). During screening programs for yield improvement, itwas noticed that some high-yielding isolates showed highlevels of genetic instability manifested by the production ofmorphological variants. In most Streptomyces strains thathave been studied, genetic instability is associated with largedeletions in the chromosome (4, 6, 7, 11). Pulsed-field gelelectrophoresis showed that deletions may be up to 2 Mb insize (24). In many cases sequences flanking the deletionsundergo high-level DNA amplification, giving rise to severalhundred tandem copies of a chromosomal sequence, and theamplified DNA may account for more than 50% of the totalDNA in the strain (2, 10, 14, 17, 28, 33).
In this paper, we report studies on genetic instability andstrain degeneration in S. rimosus R6. Five phenotypic alter-ations were classified, and pulsed-field gel electrophoresiswas used to detect large-scale DNA rearrangements.
MATERIALS AND METHODS
Strains, media, and culture conditions. All strains used inthis study are derivatives of the genetically unstable isolateR6-500 of S. rimosus R6 (the Zagreb strain, [21]), obtainedfrom the Faculty of Food Technology and Biotechnology,University of Zagreb (1). S. rimosus R6 is used by thecompany PLIVA for commercial oxytetracycline produc-tion. Strain R6-500 harbors a prophage, RP3 (29). Some ofthe spontaneous variants (see Table 1) were isolated fromstrain R6-501, which was obtained from R6-500 by lysogenywith a second temperate actinophage, RP2 (19). Represen-tative morphological variants isolated in the course of thisstudy are listed in Table 1. S. lividans 66-U7 (9) was usedas a positive control in the course of screening for thepresence of high-level amplifications. Escherichia coliHB101(pUT1954) (30) was used as a source of the otrB genefor DNA hybridization. Methods for propagation and main-
2220
on July 26, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from

GENETIC INSTABILITY IN S. RIMOSUS 2221
TABLE 1. Morphological and biochemical properties of S. rimosus R6-500 variants
Class andClassand ~~~~~~~~~~~~~~~~~RelativeOTC OtCr MICfrequency of Isolate Phenotype' yield'(Otg/ml)occurrence
Parent R6-500 Spo+ Pig+ Kug+ Otc+ Otcr 1.00 290
Class I (99%) MV1 Spo- Pig- Kug- Otc- Otcr <0.01 290MV3 Spo- Pig' Kug+ NDC ND ND NDMV6 Spo- Pig- Kug+ Otc- Otcr <0.01 290MV14 Spo- Pig+ Kug+ Otc+ Otcr 0.70 290MV19 Spo- Pig+ Kug+ Otc+ Otcr 0.75 290MV27 Spo- Pig+ Kug+ Otc+ Otcr 0.68 290MV28 Spo- Pig- Kug- Otc- Otcr <0.01 290MV32 Spo- Pig- Kug+ ND ND ND ND
Class II (1%) MV4 Spot Pig- Kug+ Otc- Otcs <0.01 5MV7 Spot Pig- Kug+ Otc- Otcs <0.01 5MV8 Spo- Pig- Kug- Otc- Otcs <0.01 5MV9 Spot Pig- Kug+ Otc- Otc' <0.01 5MV1o Spo' Pig- Kug+ Otc- Otcs <0.01 5MV11 Spo- Pig- Kug+ Otc- Otc5 <0.01 5MV15 Spot Pig- Kugt Otc- Otcs <0.01 5
Class III (0.1%) MV17d Spo-t Pig++ Kug+ Otc+ Otcrr 1.40 340MV25 Spot- Pig++ Kug+ Otc+ + Otcrr 1.30 340MV34 Spo-t Pig` Kug+ Otc+ + Otcrr 1.28 330MV36 Spot Pig++ Kug+ Otc+ + Otcrr 1.36 310MV37 Spo' Pig` Kug+ Otc+ + OtcrT 1.38 320MV41d Spot Pig` Kug+ Otc" Otcr 1.36 330MV42d Spot Pig` Kug+ Otc++ Otcrr 1.36 350
a Spo, sporulation; Pig, pigmentation; Kug, colony morphology (Kug+, "Kuglof" round regular crater morphology, Kug-, irregular flatter morphology); Otc+,Otc+, Otc-, oxytetracycline synthesis; Otcr, Otcs, oxytetracycline resistance and sensitivity, respectively. +, intermediate phenotype; + +, overproduction; rr,increased resistance.
b Relative to 1 for the parental strain. OTC, oxytetracycline.C ND, not determined because of very high reversion rates.d These variants were derived from R6-501 (see Materials and Methods).
tenance of Streptomyces stocks and the complete agarmedium (19), minimal agar medium (1), and complete pre-cultivation liquid medium (27) used were described previ-ously.
Determination of oxytetracycline resistance and productiv-ity. The MIC was determined on minimal agar medium byusing oxytetracycline.HCl (PLIVA, Zagreb, Republic ofCroatia). For the determination of the average relativeoxytetracycline yield by the parental type and morphologicalvariants, a plate-based method was used (5).DNA manipulation. Total DNA from Streptomyces strains
was isolated by the "Kirby mix" method (16). Plasmid DNAfrom E. coli was purified by cesium chloride-ethidium bro-mide gradient centrifugation (32). The DNA was digestedwith restriction enzymes as recommended by the supplierand subjected to horizontal agarose gel electrophoresis inTris-borate-EDTA (TBE) buffer (32). Southern blotting wascarried out under alkaline conditions with a Hi-Bond-Nmembrane (Amersham). DNA probes were labeled withdigoxigenin by using a random-priming kit (BoehringerMannheim). Hybridization was carried out at 68°C in plasticbags containing 5 x SSC (lx SSC is 0.15 M NaCl plus 0.015M sodium citrate) and 0.1% sodium dodecyl sulfate (SDS). Itwas followed by washes in 2x SSC-0.1% SDS and O.1xSSC-0.1% SDS. For detection of bands we used eithera phosphatase kit or light production kit with 4-methoxy-4- (phosphatephenyl)spiro(1,2 - dioxetane-3,2' - adamantane)(Boehringer Mannheim). Light was detected by using Hy-perfilm-MP (Amersham).
Pulsed-field gel electrophoresis. The method used for
pulsed-field gel electrophoresis was slightly modified fromthat of Hausler et al. (15). Liquid cultures inoculated frommycelium grown on complete agar medium were grown for36 h at 30°C in complete precultivation liquid medium. Thecultures were harvested by centrifugation (1,250 x g for 10min) and washed twice with SucTE buffer (0.3 M sucrose, 25mM Tris [pH 8.0], 25 mM EDTA). The washed cultures werediluted to an optical density at 600 nm of 1.8 in SucTE andmixed carefully with 1.5% low-melting-point agarose (Incertagarose; FMC Biozym) to a final concentration of 1%. Theagarose plugs were incubated for 16 to 24 h at 37°C inlysozyme (2 mg/ml) in SucTE buffer. They were then sub-jected to two 24-h incubations at 50°C with NDSP lysissolution (0.5 M EDTA, 1% N-lauroylsarcosine, 2 mg ofproteinase K [Boehringer], per ml [pH 8.0]). After the plugshad been washed several times in TE buffer (10 mM Tris-HCl, 1 mM EDTA [pH 8.0]), they were incubated twice for4 h each at 4°C with 40 ,ug of phenylmethylsulfonyl fluoride(Sigma) per ml to inactivate the protease. The plugs werethen washed several times in TE buffer.
Restriction digests were carried out with AseI (Biolabs)and XbaI (Pharmacia) and the buffer recommended by theenzyme supplier. The plugs were equilibrated with the buffer(twice for 4 h at 4°C). Digests were then carried out for 24 hwith 20 U per plug. Appropriate slices from the plugs wereembedded in agarose gels (1% in 0.5 x TBE) and used forpulsed-field gel electrophoresis (Bio-Rad CHEF DRII appa-ratus). The running conditions were selected to give optimalresolution in the molecular weight regions of interest. Sac-charomyces cerevisiae chromosomes or lambda ladders
VOL. 59, 1993
on July 26, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from

2222 GRAVIUS ET AL.
were used routinely as markers. However, the sizes forfragments under 500 kb were recalculated with a correctionincluded for the high G+C content (13).
RESULTS
Appearance of morphological variants. During a strainselection program to improve oxytetracycline production inS. nmosus R6, isolates that showed extreme morphologicalinstability appeared. This phenomenon was studied furtherby using strain R6-500, which is a high producer of oxytet-racycline and also shows high resistance levels to the anti-biotic. When mycelial fragments were plated onto completeagar medium, 1 to 3% of colonies were morphologicalvariants (Fig. 1A). However, when spores were plated,about 80% of colonies were variants (Fig. 1B). A possibleexplanation for this difference in frequency would be that themycelial fragments are multinucleate and complementationproduces a parental colony morphology. It had been ob-served (20) that during growth of S. nmosus R6 in liquidmedium, the morphology altered from small pellets throughmore dispersed growth to extremely dispersed small myce-lial fragments. If the differences in frequency of variantsbetween spore and mycelial fragments were due to thedifferences in numbers of nuclei, we would expect that thesmall mycelial fragments from later growth phases wouldalso show an increased frequency of variants compared withthe larger pellets seen earlier. Samples were plated fromcultures after 6, 24, and 48 h of growth and scored for thefrequency of variants. In two experiments there were nosignificant changes with culture age (the two experimentsgave 1.2, 1.6, and 2.4% and 3.2, 2.4, and 1.9% morphologicalvariants at the three sampling times).The genetic instability affects one or more of five phenotypic
traits (9). Variants that were altered in sporulation, pigmenta-tion, colony morphology, oxytetracycline production, andoxytetracycline resistance arose. (i) For sporulation variants,the parent strain sporulated well, whereas most variants hadreduced or no sporulation. (ii) For pigmentation variants, theparent strain produced a brown pigment and variants oftendiffered in amount of pigmentation. The nature of the pigmentis unknown, but its production is usually correlated withoxytetracycline production. (iii) Colony morphology variantsoften show loss of the parental round regular crater "Kuglof'morphology. (iv) Oxytetracycline production variants oftenshow reduced or no production; rare variants show increasedproduction. (v) Oxytetracycline resistance variants may showsensitivity or increased resistance.
It proved convenient to classify variants on the basis ofoxytetracycline resistance. Most variants (about 99%) re-tained normal resistance levels, often grew faster than strainR6-500 (data not shown), and formed the very heterogeneousclass I. Some typical phenotypic combinations are listed inTable 1. Figure 1C and D shows two members of class I thatrepresent the extremes of phenotypic variation. MV14 (Fig.1C) does not sporulate and has normal (Kug+) colonymorphology but has reduced pigmentation and oxytetracy-cline production. On the other hand, MV1 (Fig. 1D) showsabnormal colony morphology and has lost sporulation, pig-ment production, and oxytetracycline production. The classI variants span a whole spectrum in the degree to which eachphenotypic characteristic is affected (Table 1), and theredoes not seem to be any correlation among the differentphenotypes. Variants often show continued instability, andphenotypic characteristics may be lost or regained. In somecases, it is clear that phenotypic characteristics "revert."
For example, MV1 and MV28 give rise to pigmented vari-ants in about 1% of colonies (Fig. 2). In other cases (e.g.,MV3 and MV32), the variants revert to a parental phenotypeat such high frequencies (more than 80% of colonies) thatscoring of production phenotypes is no longer possible. Theheterogeneity in class I might reflect the operation of severalmechanisms. We chose six variants representing a range ofphenotypic combinations and relatively stable strains forfurther study (all except MV3 and MV32 [Table 1]).
Oxytetracycline-sensitive variants (class II) accounted forabout 1% of variants. They all showed a similar phenotype(Table 1; Fig. 1E): reduced sporulation, parental-type colonymorphology, and no detectable pigment or oxytetracyclineproduction. All seven class II variants were tested forreversion to oxytetracycline resistance. No revertants wereseen in 5 x 107 to 10 x 107 mycelial fragments tested. It wasinteresting that no variants intermediate in resistance be-tween the class II mutants and the parent strain were seen.The class III variants have a typical dark-brown appear-
ance (Fig. 1F) due to increased pigment production. Theyaccount for only about 0.1% of variants and have uniformphenotypical characteristics (Table 1): reduced sporulation,normal colony morphology, and increased production andresistance to oxytetracycline. They revert to a parentalphenotype and also produce other variants. This instabilitymakes them unsuitable for antibiotic production.DNA rearrangements in variants. In many Streptomyces
strains, genetic instability is associated with the generationof high-level DNA amplifications (2, 10, 14, 17, 28, 33). Wetherefore screened DNA prepared from primary variantstrains for the presence of amplifications. Total DNA wasprepared from four representatives of each class; digestedwith the restriction enzymes BamHI, EcoRI, HindIII, PstI,and SalI; and subjected to agarose gel electrophoresis.Typical results are shown in Fig. 3. No high-level DNAamplifications could be observed in any of the variantstested, but amplified DNA could be seen clearly in thecontrol strain S. lividans 66-U7 (9) (Fig. 3, lane 10).
Pulsed-field gel electrophoresis is a useful technique todetect large-scale DNA rearrangements in Streptomycesspecies (24). Preliminary experiments showed that S. rimo-sus R6-500 carries a giant linear plasmid and that theenzymesAseI andXbaI generate convenient sizes of restric-tion fragments for genomic analysis (9). Six class I variants(MV1, MV6, MV14, MV19, MV27 and MV28 [Table 1])were examined by pulsed-field gel electrophoresis (data notshown). All carried a giant linear plasmid of identical size tothat in the parent strain, and none of the variants showed anychanges in the AseI or XbaI digestion patterns. Southerntransfers of the gels were hybridized with a probe containingthe otrB oxytetracycline resistance gene (plasmid pUT1954[30]) that lies near one end of the oxytetracycline synthesiscluster (6). None of the variants showed alterations in thehybridization pattern.
Southern hybridization experiments of genomic DNAtransferred from horizontal agarose gels indicated that theclass II variants had deleted the otrB gene (data not shown).DNA from seven class II variants was used for pulsed-fieldgel electrophoresis. No changes were observed in the giantlinear plasmid (data not shown). When AseI digests wereexamined, six of the seven independently isolated variants(MV4, MV7, MV8, MV10, MV11, and MV15 [Table 1])showed a new AseI band of 340 kb; the exception (MV9[Table 1]) showed a band of 240 kb (Fig. 4A). Hybridizationof a Southern blot with an otrB probe (Fig. 4B) showed thatthe 795-kb band which hybridizes in the parent strain is
APPL. ENvIRON. MICROBIOL.
on July 26, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from
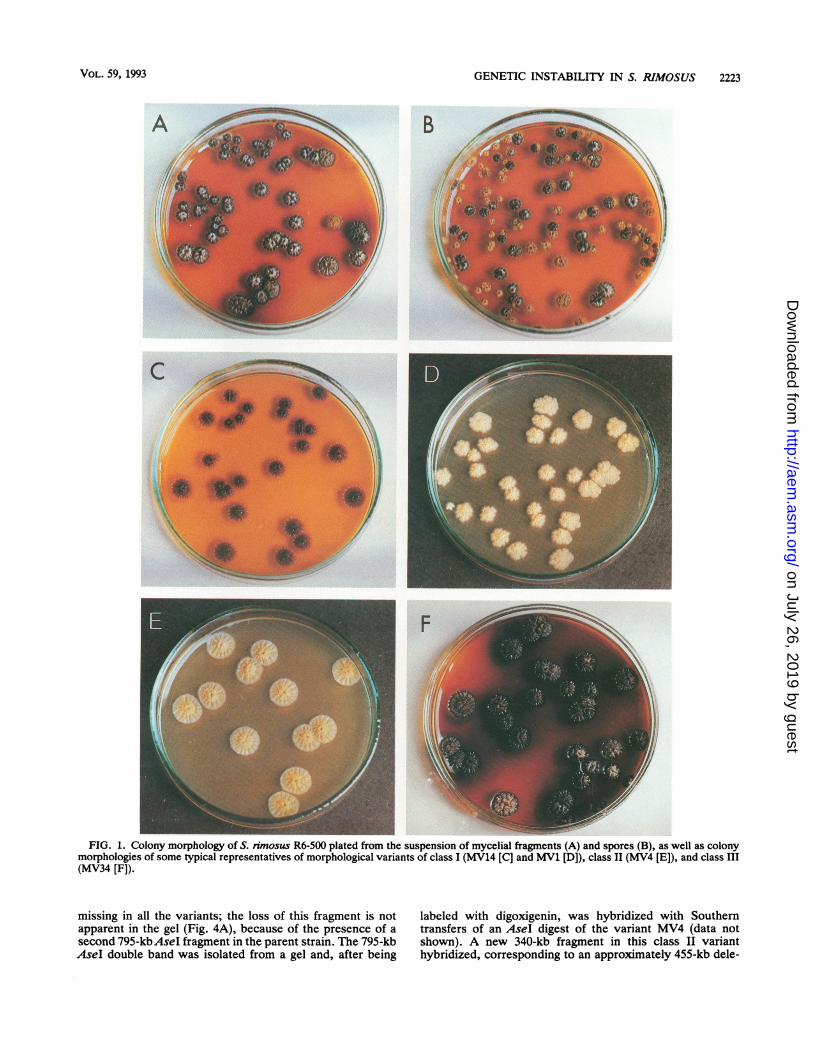
GENETIC INSTABILITY IN S. RIMOSUS 2223
B
- w_FIG. 1. Colony morphology of S. rimosus R6-500 plated from the suspension of mycelial fragments (A) and spores (B), as well as colony
morphologies of some typical representatives of morphological variants of class I (MV14 [C] and MV1 [D]), class II (MV4 [E]), and class III(MV34 [F]).
missing in all the variants; the loss of this fragment is notapparent in the gel (Fig. 4A), because of the presence of asecond 795-kbAsel fragment in the parent strain. The 795-kbAseI double band was isolated from a gel and, after being
labeled with digoxigenin, was hybridized with Southerntransfers of an AseI digest of the variant MV4 (data notshown). A new 340-kb fragment in this class II varianthybridized, corresponding to an approximately 455-kb dele-
VOL. 59, 1993
on July 26, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from

2224 GRAVIUS ET AL.
FIG. 2. Colonies of morphological variant MV1 showing one Pig' Otc+ "revertant."
tion in one of the 795-kb fragments. In XbaI digests of classII variants (Fig. 5), a band of about 415 kb that carries theotrB gene disappeared, but we have been unable to locate anew junction band.DNA from seven independently derived class III variants
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
FIG. 3. Electrophoretogram of total DNA of S. rimosus R6-500and some representatives of morphological variants of classes I, II,and III, digested with BamHI and Sall after conventional agarosegel electrophoresis. Lanes: 1 and 2, R6-500; 3 and 4, MV14; 5 and 6,MY1; 7 and 8; MV4; 9, lambda DNA digested with HindIll; 10, S.lividans TK64-U7 DNA digested with PstI; 11 and 12, MV7; 13 and14, MV25; 15 and 16, MV34.
(MV17, MV25, MV34, MV36, MV37, MV41, and MV42[Table 1]) was also examined by pulsed-field gel electro-phoresis and Southern hybridization. It was observed that inXbaI digests of all variants except MV17, the 415-kb bandcarrying the otrB gene had been replaced by a smaller bandof 200 kb (data not shown). In five of these six variants, the795-kb AseI band was replaced by a band of 920 kb, and inMV34 a new 1,050-kb band was found (data not shown).Therefore, there are reproducible large-scale DNA rear-rangements associated with most of the class III variants.Two of the variants, MV17 and MV25, showed alterationsinvolving the giant linear plasmid, which will be discussedelsewhere (12), but their significance for the instabilitybehavior is unclear.The nonpigmented class I variants MV1 and MV28 give
rise to further variants that have regained the ability toproduce brown pigment. Three such "revertants" wereisolated from each strain and analyzed by pulsed-field gelelectrophoresis. In Fig. 6, it can be seen that all six "rever-tants" had lost the 415-kb XbaI otrB-containing band andthat otrB was carried on a fragment of 200 kb, which is ofidentical size to that seen in the class III variants. It is clearthat this 200-kb fragment is present at a substantially in-creased copy number and that two small XbaI fragments (29and 31 kb) are also amplified (Fig. 6). In AseI digests, theotrB-containing fragment has increased in size from 795 tomore than 1,600 kb (Fig. 7) and is probably larger than theupper resolution limit under the electrophoresis conditionsused.
DISCUSSION
S. nmosus R6-500 shows a high frequency of productionof morphological variants, especially when propagated via
APPL. ENvIRON. MICROBIOL.
on July 26, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from

GENETIC INSTABILITY IN S. RIMOSUS 2225
1 2 3 4 5 6 7 8 9 10
4-795kb -
*-340kbi-*240kbi
A BFIG. 4. (A) Pulsed-field gel electrophoresis ofAseI digests of chromosomal DNA of class II (Otcs) mutants. Lanes: 1 and 10, chromosomes
of Saccharomyces cerevisiae; 2, parent strain R6-500; 3 to 9, Otcs mutants MV4, MV7, MV8, MV10, MV11, MV15, and MV9, respectively.The electrophoresis parameters were 200 V; 14 h: 40 to 50-s pulse time slope; 16 h: 60- to 130-s slope. (B) Southern transfer of gel in panelA hybridized with a plasmid carrying the otrB resistance gene (pUT1954).
spores. The difference between spores and mycelial frag-ments cannot be accounted for solely by the number ofnuclei involved, because the frequency of variants remainsconstant during growth of a culture, when the size ofmycelial fragments varies widely. This behavior might beexplained if many chromosomes during mycelial growthwere multimers and segregation into spores required theformation of haploid products. That the existence of rela-tively stable structures carrying more than one chromosomeis possible is indicated by the presence of heterogenotesafter protoplast fusion (18). On the other hand, it has beensuggested that an increase in mutability could be a conse-quence of changes in the stability of the replisome-DNAcomplex caused by growth conditions that modify the repli-cation rhythm (such as spore germination), resulting inerrors in DNA replication (23).
It proved to be convenient to classify variants according totheir oxytetracycline resistance. Most variants had parentallevels of resistance. They showed a variety of phenotypes
1 2 3 4 5 6 7 8 9 10
affected to differing degrees in sporulation, colony morphol-ogy, pigmentation, and oxytetracycline production. Variantswere often unstable, giving rise to further variants and toreversion to some of the parental characteristics. Screeningfor high-level DNA amplification by gel electrophoresis orother large-scale DNA rearrangements by using pulsed-fieldgel electrophoresis showed no changes in primary class Ivariants compared with the parent strain. Therefore, anyDNA changes in this heterogeneous common class of vari-ants were below the limits of resolution of the methods used.It is possible that small changes in the copy number ofcontrol genes, such as suggested for the RES1 elementaffecting level of chloramphenicol resistance in S. lividans 66(34), was not detected in our experiments.
In contrast to class I variants, class II variants, which aresensitive to oxytetracycline, show very uniform phenotypicproperties. No variants with intermediate levels of resis-tance were observed. All variants had deleted the otrBresistance gene, and the changes seen in pulsed-field gel
1 2 3 4 5 6 7 8 9 10
A BFIG. 5. (A) Pulsed-field gel electrophoresis of XbaI digests of chromosomal DNA of class II mutants. Lanes: 1 and 10, chromosomes of
Saccharomyces cerevisiae; 2, parent strain R6-500; 3 to 9, Otcs mutants MV4, MV7, MV8, MV10, MV11, MV15, and MV9, respectively. Theelectrophoresis parameters were 200 V; 28 h: 10- to 100-s pulse time slope. (B) Southern transfer of gel in panel A hybridized with an otrBprobe (pUT1954).
VOL. 59, 1993
on July 26, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from

2226 GRAVIUS ET AL.
3 45tSo67t008 9 10
FIG. 6. (A) Pulsed-field gel electrophoresis of Xbal digests of DNA from class I "revertants." Lanes: 1, Saccharomyces cerevisiaechromosomes; 2, parent strain R6-500; 3, MV1; 4 to 6, "revertants" MV1rA, MV1rB, and MV1rC, respectively; 7, MV28; 8 to 10,"revertants" MV28rA, MV28rB, and MV28rC, respectively. This gel was run under the following conditions: 200 V, 28 h: 4- to 40-s pulsetime slope. (B) Southern transfer of gel in panel A hybridized with an otrB probe (pUlT1954).
electrophoresis suggest that large deletions are present,which may remove all the oxytetracycline synthesis andresistance genes as reported for the strain of S. nmosusstudied at Pfizer (6, 31). It was striking that most (six ofseven) of the independently isolated variants showed anidentical change in restriction pattern; this suggests thatthere are specific deletion end points. This is in contrast tothe deletions observed in most Streptomyces species, inwhich a variety of end points in a region are used (4, 24).The class III variants, which were the rarest observed
(about 0.1% of variants), also exhibit a uniform phenotype.They show 30 to 40% overproduction of oxytetracycline in aplate test and also have increased resistance. Most (six ofseven) of the independently isolated variants tested showedthe same specific alteration in pulsed-field gel electrophore-sis that affected the XbaI fragment carrying the otrB gene.The only exception, MV17, had acquired extra copies of the
1 2 3 4 5 6 7 8 9 10
otrB gene (and probably the biosynthesis genes) carried bythe giant linear plasmid which is present in S. rimosus R6(12). In this case, the increases in production and resistancemight be caused by the increased copy number of the genesinvolved. In most of the class III strains, the otrB gene ispresent on a smallerXbaI fragment (200 kb instead of 415 kb)and there is probably also a low-level reiteration (three orfour copies) of this fragment. One of the class III strains(MV25) had integrated parts of the giant linear plasmid intothe chromosome (12), but it is not clear whether the integra-tion plays a role in the phenotypic changes, since MV25 doesnot differ phenotypically from the other class III variantswith autonomous plasmid.The "revertants" of the class I strains MV1 and MV28
show the same change in size of the otrB-containing XbaIfragment, but the fragment is also amplified together withtwo small XbaI fragments (Fig. 6); preliminary copy number
1 2 3 4 5 6 7 8 9 10
j. ,.r.f,"Iiif
i _-P* t4w:-wf
.
"O.~~~~~~~~~~~~~~~~~~
*--l.6Mb
| -795kb
A BFIG. 7. (A) Pulsed-field gel electrophoresis of AseI digests of DNA from class I "revertants." Lanes: 1, Saccharomyces cerevisiae
chromosomes; 2, parent strain R6-500; 3, MV1; 4 to 6, "revertants" MV1rA, MV1rB, and MV1rC, respectively; 7, MV28; 8 to 10,"revertants" MV28rA, MV28rB, and MV28rC, respectively. This gel was run under the same conditions as those used for the gel in Fig. 4.(B) Southern transfer of the gel in panel A hybridized with an otrB probe (pUT1954).
I5
APPL. ENvIRON. MICROBIOL.
on July 26, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from

GENETIC INSTABILITY IN S. RIMOSUS 2227
estimates from the intensity of the gel bands suggest copynumbers of 20 to 50. It is likely that this amplificationinvolves tandem repeats of the 200-kb unit in the chromo-some; the presence of very large fragments in AseI digests(Fig. 7) would support this idea. There may well be deletionof sequences to produce the reduced molecular weight of theXbaI-amplified fragment. One explanation for these resultsis that the class I variants MV1 and MV28 carry a mutationin a control gene that results in poor expression of pigmentand oxytetracycline production genes. This mutation couldbe suppressed by amplification of the structural genes asreported for RES1-dependent mutations in S. lividans (34).
It seems clear that the variants responsible for classicalstrain degeneration in S. rimosus R6-500 are variants belong-ing to class I, in which no DNA rearrangements weredetected. These variants, unlike the oxytetracycline-sensi-tive class II variants, would survive under production con-ditions, especially since they usually grow faster than theparental type, allowing accumulation of a nonproducingsubpopulation. More work is needed to detect the alterationsat the DNA level in class I variants and to understand themechanisms involved. In addition, better characterization ofthe class III and class I "revertant" rearrangements mightallow the introduction of a selectable marker gene to stabi-lize the overproducing variants by using an approach similarto that of Kaiser et al. (22).
ACKNOWLEDGMENTS
This work was supported by EC International Scientific Cooper-ation grant CI1*/0527-C(MB) (to J.C. and D.H.) and by grant1-08-021 (to D.H.) from the Ministry of Science, Technology andInformatics, Republic of Croatia. We thank the state of Rheinland-Pfalz for a graduate stipendium (to B.G.) and the Ministry ofScience, Technology and Informatics, Republic of Croatia, forfinancial support for a graduate student (T.B.).We thank Goran Biukovic for providing strain MV17. We grate-
fully acknowledge the receipt of plasmid pUT1954 from G. Tiraby.We are also indebted to Helena Kobelscak and Visnja Horvat fortheir skilled technical assistance. We thank Sonja Schwarz and AnjaEichner for their contributions during research practicals.
REFERENCES1. Ala&evic, M., M. Stragek-Vesligaj, and G. Sermonti. 1973. The
circular linkage map of Streptomyces nmosus. J. Gen. Micro-biol. 77:173-185.
2. Altenbuchner, J., and J. Cullum. 1984. DNA amplification andan unstable arginine gene in Streptomyces lividans 66. Mol.Gen. Genet. 195:134-138.
3. Beckmann, R. J., K. Cox, and E. T. Seno. 1989. A cluster oftylosin biosynthetic genes is interrupted by a structurally unsta-ble segment containing four repeated sequences, p. 176-186. InC. L. Hershberger, S. W. Queener, and G. Hegeman (ed.),Genetics and molecular biology of industrial microorganisms.Proceedings of the 4th ASM Conference on the Genetics andMolecular Biology of Industrial Microorganisms. AmericanSociety for Microbiology, Washington, D.C.
4. Birch, A., A. Hausler, M. Vogtli, W. Krek, and R. Hutter. 1989.Extremely large chromosomal deletions are intimately involvedin genetic instability and genomic rearrangements in Streptomy-ces glaucescens. Mol. Gen. Genet. 217:447-458.
5. Bofnjak, M., J. Pigac, R. Valinger, M. Vampola, and M.Velligaj. 1990. Kinetics of product formation in microbial colo-nies, p. 345-358. In A. Blazej and A. Ottova (ed.), Mathematicalmodelling in biotechnology. Proceedings of the InternationalSymposium on Biotechnology. Elsevier Biomedical Press, Am-sterdam.
6. Butler, M. J., E. J. Friend, I. S. Hunter, F. S. Kaczmarek, D. A.Sugden, and M. Warren. 1989. Molecular cloning of resistancegene and architecture of a linked gene cluster involved in the
biosynthesis of tetracycline by Streptomyces rimosus. Mol.Gen. Genet. 215:231-238.
7. Cox, K. L., S. E. Fishman, J. L. Larson, R. Stanzak, P. A.Reynolds, W.-K. Yeh, R. M. Van Frank, V. A. Birmingham,C. L. Hershberger, and E. T. Seno. 1987. Cloning and charac-terization of genes involved in tylosin biosynthesis, p. 337-346.In M. Alacevic, D. Hranueli, and Z. Toman (ed.), Genetics ofindustrial microorganisms. Proceedings of the 5th InternationalSymposium on the Genetics of Industrial Microorganisms.PLIVA, Zagreb, Yugoslavia.
8. Cullum, J., J. Altenbuchner, F. Flett, and W. Piendi. 1986. DNAamplification and genetic instability in Streptomyces. Biotech-nol. Genet. Eng. Rev. 4:59-78.
9. Cullum, J., F. Flett, B. Gravius, D. Hranueli, K. Miyashita, J.Pigac, U. Rauland, and M. Redenbach. 1991. Analysis of ampli-fications and deletions in Streptomyces species, p. 265-272. InS. Baumberg, H. Kriigel, and D. Noack (ed.), Genetics andproduct formation in Streptomyces. Proceedings of the Interna-tional Symposium on Genetics and Product Formation in Strep-tomyces. Plenum Press, New York.
10. Fishman, S. E., and C. L. Hershberger. 1983. Amplified DNA inStreptomyces fradiae. J. Bacteriol. 155:459-466.
11. Flett, F., and J. Cullum. 1987. DNA deletions in spontaneouschloramphenicol-sensitive mutants of Streptomyces coelicolorA3(2) and Streptomyces lividans 66. Mol. Gen. Genet. 207:499-502.
12. Gravius, B., K. Pandia, D. Glocker, J. Pigac, D. Hranueli, andJ. Cullum. Unpublished data.
13. Gravius, B., J. Cullum, and D. Hranueli. Submitted for publi-cation.
14. Hasegawa, M., G. Hintermann, J.-M. Simonet, R. Crameri, J.Piret, and R. Hutter. 1985. Certain chromosomal regions inStreptomyces glaucescens tend to carry amplifications anddeletions. Mol. Gen. Genet. 200:375-384.
15. Hiusler, A., A. Birch, W. Krek, J. Piret, and R. Hutter. 1989.Heterogeneous genomic amplification in Streptomycesglaucescens: structure, location and junction sequence analysis.Mol. Gen. Genet. 217:437-446.
16. Hopwood, D. A., M. J. Bibb, K. F. Chater, T. Kieser, C. J.Bruton, H. M. Kieser, D. J. Lydiate, C. P. Smith, J. M. Ward,and H. Schrempf. 1986. Genetic manipulation in Streptomyces:a laboratory manual. The John Innes Foundation, Norwich,England.
17. Hornemann, U., C. J. Otto, G. G. Hoffman, and A. C. Bertuni-son. 1987. Spectinomycin resistance and associated DNA am-plification in Streptomyces achromogenes subsp. rubradiris. J.Bacteriol. 169:2360-2366.
18. Hranueli, D., J. Pigac, T. Smokvina, and M. Alacevic. 1983.Genetic interactions in Streptomyces rimosus mediated by con-jugation and by protoplast fusion. J. Gen. Microbiol. 129:1415-1422.
19. Hranueli, D., J. Pigac, and M. Vesligaj. 1979. Characterizationand persistence of actinophage RP2 isolated from Streptomycesrimosus ATCC 10970. J. Gen. Microbiol. 114:295-303.
20. Hranueli, D., T. Smokvina, and M. Alacevic. 1986. A compara-tive study of protoplast preparation and regeneration in Strep-tomyces rimosus and Streptomyces lividans. Microbiologica9:387-392.
21. Hutter, R., and T. Eckhardt. 1988. Genetic manipulation, p.89-184. In M. Goodfellow, S. T. Williams, and M. Mordarski(ed.), Actinomycetes in biotechnology. Academic Press, Inc.,San Diego, Calif.
22. Kaiser, P., F. Flett, and J. Cullum. 1992. Stabilization ofStreptomyces lividans by homologous recombinational inser-tion. Bio/Technology 10:570-573.
23. Leblond, P., P. Demuyter, J.-M. Simonet, and B. Decaris. 1990.Genetic instability and hypervariability in Streptomyces ambo-faciens: towards an understanding of a mechanism of genomeplasticity. Mol. Microbiol. 4:707-714.
24. Leblond, P., P. Demuyter, J.-M. Simonet, and B. Decaris. 1991.Genetic instability and associated genome plasticity in Strepto-myces ambofaciens: pulsed-field gel electrophoresis evidencefor large DNA alterations in a limited genomic region. J.
VOL. 59, 1993
on July 26, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from

APPL. ENVIRON. MICROBIOL.
Bacteriol. 173:4229-4233.25. Miyoshi, T., M. Iseki, T. Konomi, and H. Imanaka. 1980.
Biosynthesis of bicyclomycin. I. Appearance of aerial mycelianegative strains (am-). J. Antibiot. 33:480-487.
26. Pigac, J., and M. Ala&vic. 1979. Mapping of oxytetracyclinegenes in Streptomyces rimosus. Period. Biol. 81:575-582.
27. Pigac, J., D. Hranueli, T. Smokvina, and M. Alacevic. 1982.Optimal cultural and physiological conditions for handlingStreptomyces rimosus protoplasts. Appl. Environ. Microbiol.44:1178-1186.
28. Potekhin, Y., and V. N. Danilenko. 1985. The determinant ofkanamycin resistance of Streptomyces nmosus: amplification inthe chromosome and reversed genetic instability. Mol. Biol.(Moscow) 19:805-817.
29. Rausch, H., M. Vesligaj, D. Pocta, G. Biukovic, J. Pigac, J.Cullum, H. Schmieger, and D. Hranueli. The temperate phagesRP2 and RP3 of Streptomyces nmosus. J. Gen. Microbiol., inpress.
30. Reynes, J. P., T. Calmels, D. Drocourt, and G. Tiraby. 1988.Cloning, expression in Eschenchia coli and nucleotide sequence
of a tetracycline-resistance gene from Streptomyces nimosus. J.Gen. Microbiol. 134:585-598.
31. Rhodes, P. M., I. S. Hunter, E. J. Friend, and M. Warren. 1984.Recombinant DNA methods for the oxytetracycline producerStreptomyces rimosus Biochem. Soc. Trans. 12:586-587.
32. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecularcloning: a laboratory manual, 2nd ed. Cold Spring HarborLaboratory, Cold Spring Harbor, N.Y.
33. Schrempf, H. 1983. Deletion and amplification of DNA se-quences in melanin-negative variants of Streptomyces reticuli.Mol. Gen. Genet. 189:501-505.
34. Starodubtseva, L. I., A. S. Taisova, and V. N. Danilenko. 1985.Study on amplification of kanamycin resistance determinant(KanR) in constructed hybrid plasmids of Streptomyces lividans.Antibiotiki 30:565-572.
35. Young, M., and D. Hranueli. 1988. Chromosomal gene amplifi-cation in gram-positive bacteria, p. 157-200. In J. A. Thomson(ed.), Recombinant DNA and bacterial fermentation. CRCPress, Inc., Boca Raton, Fla.
2228 GRAVIUS ET AL.
on July 26, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from