genetic analysis of ten sheep breeds using microsatellite markers
TRANSCRIPT

Genetic analysis of ten sheep breeds using microsatellite markers
A. Farid1,3, E. O’Reilly2, C. Dollard1, and C.R. Kelsey Jr.2
1Department of Animal Science, Nova Scotia Agricultural College, Truro, Nova Scotia, Canada B2N 5E3;2Newfoundland Department of Forest Resources and Agrifoods, Brookfield Road, St. John’s, Newfoundland,
Canada A1B 4J6. Received 9 September 1999, accepted 8 November 1999.
Farid, A., O’Reilly, E., Dollard, C. and Kelsey Jr., C. R. 2000. Genetic analysis of ten sheep breeds using microsatellite mark-ers. Can. J. Anim. Sci. 80: 9–17. The genetic variability of 257 sheep from 10 breeds; North Country Cheviot (NCC), Cheviot(CHE), Dorset (DOR), Suffolk (SUF), Scottish Blackface (SBF), Texel (TEX), Romanov (ROM), Finnish Landrace (FIN),Icelandic (ICE) and Red Masai (MAS) was assessed using 10 microsatellite loci. The average number of alleles per locus was 4.3in ROM, 5.0 in MAS, and spanned a narrow range (5.4 to 6.0) in the other breeds. Estimates of expected heterozygosity (HE) ofthe breeds varied within 0.05 point of each other (0.62 in FIN to 0.67 in CHE), except for ROM (0.53) which was lower (P < 0.05)than those of the other breeds, and in MAS (0.57), which was lower (P < 0.05) than those of NCC, CHE and SBF. Estimates ofobserved heterozygosity (HO) of the breeds were the lowest in MAS, ROM and SUF (0.50 to 0.53) and the highest in NCC andCHE (0.64 and 0.67). The HE was greater (P < 0.01) than HO only in SUF. The results suggest that there have not been drasticlosses of genetic variability in the intensely selected breeds. The low genetic variability of ROM was probably due to a small num-ber of animals imported to North America. The British breeds (NCC, CHE, SUF, DOR, SBF) were genetically close to each other,as were the North European breeds (ROM, ICE, FIN). MAS was remotely related to the British breeds, but it was surprisinglyclose to the North European breeds. TEX was more closely related to the British breeds than to the North European breeds. Morethan 90% of 1000 simulated individuals from each breed were assigned to the correct breed, indicating that this panel of markersis useful for the identification of breed membership of individual animals, and could be used to protect the integrity of registeredbreeds.
Key words: Sheep, genetic variability, genetic distance, microsatellites
Farid, A., O’Reilly, E., Dollard, C. et Kelsey Jr., C. R. 2000. Analyse génétique de dix races ovines au moyen d’un jeu demicrosatelliteb. Can. J. Anim. Sci. 80: 9–17. Nous avons évalué au moyen d’un jeu de dix locus de microsatellites, la variabilitégénétique chez 257 moutons appartenant à dix races : North Country Cheviot (NCC), Cheviot (CHE), Dorset (DOR), Suffolk(SUF), Scottish Blackface (SBF), Texel (TEX), Romanov (ROM), Finnoise (FIN), Islandaise (IS) et Red Masai (MAS). Le nom-bre moyen d’allèles par locus était de 4,3 chez les moutons ROM, 5,0 chez les MAS, et dans les autres races, fluctuait entre 5,4 et6,0. Les valeurs calculées d’hétérozygotie espérée (HE) des races variaient dans un écart de 0,05 point l’une de l’autre, soit de 0,62chez FIN à 0,67 chez CHE, sauf pour ROM où elles étaient plus basses (P < 0,05) que chez toutes les autres races (0,53) et pourMAS chez qui elles étaient plus basses (P < 0,05) que chez NCC, CHE et SBF. Les valeurs d’hétérozygotie observée (HO) les plusbasses appartenaient aux races MAS, ROM et SUF (0,50 à 0,53) et les plus hautes à NCC et CHE (0,64 et 0,67). Ce n’est que dansla race SUF que HE était plus élevé (P < 0,01) que HO . D’après ces observations, on constate qu’il n’y a pas de perte spectacu-laire de la variabilité génétique, même dans les races soumises à une sélection intensive. La faible variabilité génétique chez lesmoutons ROM s’explique probablement par le nombre relativement restreint de sujets importés à l’origine en Amérique du Nord.Les races britanniques : NCC, CHE, SUF, DOR et SBF, étaient génétiquement rapprochées l’une de l’autre, comme l’étouentégalement les races nord-européennes (ROM, FIN, IS). La race MAS était génétiquement éloignée des races britanniques, maisétonnamment proche des races nord-européennes. TEX était plus apparentée aux races britanniques qu’à celles d’Europe du nord.Plus de 90 % des 1 000 individus modélisés de chaque race étaient correctement attribués à leur race d’appartenance, ce qui laisseà penser que le jeu de marqueurs choisis est utile pour l’identification de la race de sujets particuliers et qu’on pourrait s’en servirpour protéger l’intégrité génétique des races enregistrées.
Mots clés: Mouton, variabilité génétique, distance génétique, microsatellites
The extent of genetic variation within breeds of livestockinfluences the rate of genetic progress by selection and thesuccess of genetic resource conservation in the long-term.Several of the sheep breeds in Canada were established by arather small number of individuals (Pickard 1987; Fahmy1996b), which may have resulted in the loss of genetic vari-ability (Nei et al. 1975; Tarr et al. 1998). In addition, selec-tion based on estimated breeding values rather than
phenotypic merits can lead to the extensive use of a smallnumber of elite individuals in purebred livestock popula-tions, and could potentially increase inbreeding (Quinton et
9
Abbreviations: CHE, Cheviot; DOR, Dorset; FIN ,Finnish Landrace;HO, observed heterozygosity; HE,expected heterozygosity; H-W , Hardy-Weinberg; ICE ,Icelandic; MAS, Red Masai; NCC, North Country Cheviot;NJ, Neighbour-joining; PCR, polymerase chain reaction;ROM , Romanov; SBF, Scottish Blackface; SUF, Suffolk;TEX , Texel
3To whom correspondence should be addressed (e-mail:[email protected]).
Can
. J. A
nim
. Sci
. Dow
nloa
ded
from
pub
s.ai
c.ca
by
Uni
vers
ity o
f T
enne
ssee
on
03/0
1/13
For
pers
onal
use
onl
y.

10 CANADIAN JOURNAL OF ANIMAL SCIENCE
al. 1992; Wiener et al. 1994), reduce genetic variance andresponse to selection (Gama and Smith 1993). Although theuse of estimated breeding value for selection has so far beenlimited in sheep, the use of a small number of intenselyselected sires in the future will make census numbers par-ticularly deceiving, since effective population sizes may bevery small despite large census numbers. Measuring theextent of genetic variability in intensely selected livestockpopulations, and those that were established by a small num-ber of founder animals, is thus more informative than demo-graphic parameters. Likewise, knowledge of geneticvariability between breeds is essential for the conservationof genetic resources. It has been suggested that breeds withdiverse genetic backgrounds are needed in the future forgenerating transgressive variation for quantitative trait locimapping and developing new genotypes for particular man-agement systems and market demands (Notter 1999).Preservation of the estimated 3500 to 4000 breeds of mam-malian livestock is neither possible nor necessary (Barker1994; Notter 1999). Genetic distance and phylogeneticanalyses are useful tools in choosing breeds that cover thewidest range of genetic variability (Barker 1994).
Because microsatellite markers are abundant, have highlevels of polymorphism and genotypes of an animal at manyloci can be easily determined, they are the most useful toolsdeveloped to date for the measurement of a wide range of pop-ulation parameters (Bruford and Wayne 1993). Microsatellitemarkers have extensively been used in recent years for esti-
mating population parameters of aquatic and wildlife species,but their use in livestock species in general, and sheep breedsin particular, has so far been limited (Buchanan et al. 1994;Bancroft et al. 1995; Arranz et al. 1998). The objective of thisstudy was to determine levels of genetic variability within andbetween 10 sheep breeds with diverse developmental histo-ries. Although the focus of our study was on the popular sheepbreeds in Canada, a few less-popular and exotic breeds werealso included as reference populations to facilitate interpreta-tion of the results. We also studied the usefulness ofmicrosatellite loci in correctly identifying the breed of an indi-vidual animal. This could be a useful procedure in protectingthe integrity of registered pure breeds.
MATERIALS AND METHODS
AnimalsBlood or semen samples from 24 to 32 sheep each from 10breeds, NCC, CHE, DOR, SUF, SBF, TEX, ROM, FIN, ICEand MAS, were used. Registration information was used toselect animals that were unrelated for at least two genera-tions. The exceptions were SBF and MAS, which are notregistered breeds. Attempts were made to collect samplesfrom the latter breeds as broadly as possible. Most samplesoriginated from various farms across Canada, but because ofgenetic relatedness among sheep on various farms that werewilling to participate in this study, it became necessary tocollect some samples from outside Canada. Information on
Table 1. Origin and number of animals sampled
Breed Country Province/State Number of farms Number of animals Total/breed
North Country Cheviot Canada AL, BC, NS, ON 7 25 25Icelandic Canada ON 2 9 32
Iceland z – 23Romanov Canada AL, BC, ON, NS 4 14 25
CIOQy 8 8USA Nebraska 1 3
Suffolk Canada AL, NS 6 13 25CIOQy 5 6
USA Nebraska 1 6Texel Canada AL 1 14 25
CIOQy 2 2Denmark x 3 6
USA Nebraska 1 3Cheviot Canada BC, QC, NS 6 21 25
UK Scotlandw – 4Dorset Canada QC, NS 5 15 24
CIOQy 12 9Finnsheep Canada QC, NS 2 7 26
CIOQy 1 1USA NY, Nebraska 2 18
Scottish Blackface Canada ON, NS 5 25 25Red Masai Kenya v – 25 25Total 257z DNA samples were kindly provided by Dr. Astridur Palsdottir, Institute of Experimental Pathology, Keldur, Iceland. These samples had been collectedfrom various farms across Iceland. y Centre d’insemination ovine du Quebec. Semen samples obtained from rams that originated from various farms in Canada and the USA.x Samples from Texels that were imported from Denmark in 1994.w Frozen blood samples obtained from the Veterinary Services Laboratory, Scottish Agricultural College. Pedigrees of these animals were not known. v DNA samples were kindly provided by Drs. A. Teale and O. Hanotte, International Livestock Research Institute, Kenya. The source of samples (all ewes)was the experimental flock at the Diani Estate, Mombasa. The experimental flock was established by sampling broadly and representatively from smallholders throughout Kenya.
Can
. J. A
nim
. Sci
. Dow
nloa
ded
from
pub
s.ai
c.ca
by
Uni
vers
ity o
f T
enne
ssee
on
03/0
1/13
For
pers
onal
use
onl
y.

FARID ET AL. — GENETIC ANALYSIS OF SHEEP BREEDS 11
the origin of the samples is presented in Table 1. Blood sam-pling in Canada was performed by experienced techniciansand veterinarians according to the guidelines of theCanadian Council on Animal Care.
Laboratory ProceduresDNA was extracted from blood using the high salt proce-dure of Montgomery and Sise (1990). DNA from semensamples was extracted using a Puregene DNA isolation kit(Vector Biosystems, NC) according to the manufacturer’srecommendations. Ten pairs of unlinked ovine microsatel-lite primers were used to amplify genomic DNA by thePCR. The sequences, chromosomal locations and annealingtemperatures of the microsatellite primers are reported byCrawford et al. (1995). Amplifications were performed in10–15 µL volumes containing 100–200 ng of sample DNA,0.2 mM dNTPs (Boehringer Mannheim, Laval, QC), 0.4 Uof Taq DNA polymease (Boehringer Mannheim), 1X PCRbuffer, 0.01% Tween 20, 1.4 µM unlabelled and 0.6 µMlabelled primer A and 2 µM unlabelled primer B. Primer Awas end labelled with γ-[32P]ATP (ICN pharmaceuticals,Irvine, CA) using T4 polynucleotide kinase (New EnglandBiolabs, Mississauga, ON) according to the manufacturer’sinstruction. PCR cocktails were overlaid by one drop ofmineral oil, and the samples were amplified in a thermalcycler (PTC-100, MJ Research, Inc., Fisher Scientific).Published information on PCR reaction conditions was fol-lowed for each microsatellite (Crawford et al. 1995), withthe exception of OarCP26, for which the annealing temper-ature was increased from 63°C to 64°C to improve band res-olution. After addition of 5 µL stop solution to each PCRtube, mixtures were heated to 95°C for 15 min, 3 µL wereloaded onto 8% polyacrylamide gels and electrophoresedfor 2–3 h. Bands were visualized on Kodak XK-1 films, andscored using M13mp18 sequencing ladder, which wasloaded on both sides of each gel. Samples that producedambiguous results (rare or no alleles, faint or fuzzy bands)were run at least three times to ensure accurate results.
Data AnalysisGenetic polymorphism for each breed was measured as themean number of alleles per locus. Conformation of geno-
type frequencies to H-W equilibrium was tested using theGENEPOP computer package, version 1.2 (Raymond andRousset 1995). Prior to this test, alleles with frequency ofless than 5% over all breeds were considered as rare, andwere pooled together, or were added to the allele class withthe lowest frequency when there was only one rare allele ata locus. The observed heterozygosity (Ho) was computed foreach breed at each locus by direct count. Expected unbiasedheterozygosity (HE) of each breed at each locus was com-puted according to Nei and Roychoudhury (1974). The Hoand HE of each breed were computed by taking the arith-metic mean of the estimates at each locus weighted for thenumber of individuals genotyped at that locus. Pairwise testsfor the equality of HE of the breeds were performed usingthe likelihood ratio χ2 (SAS Institute, Inc. 1996). The F-sta-tistics (FST, FIS and FIT; Wright 1978) were computed usingthe DIPLOID program of Weir (1990) supplied by theGENEPOP program. The PHYLIP (Phylogeny InferencePackage) computer program, version 3.57 (Felsenstein1995) was used to compute Nei’s genetic distances (Nei1972) and to construct consensus unrooted Neighbour-join-ing (NJ) tree (Saitou and Nei 1987) from 1000 bootstrapreplications of the allele frequency matrix. No out-groupwas specified.
The usefulness of this panel of microsatellite loci in cor-rectly identifying the breed of an individual was determinedby an assignment test using a custom-made Fortran pro-gram. For each breed, 1000 allele pairs at every locus weregenerated by Monte Carlo simulation using the observedallele frequency distributions. For each simulated individ-ual, the likelihood of the genotypes was calculated for eachof the 10 breeds, and the individual was assigned to thatbreed giving the maximum likelihood. To improve the accu-racy of the procedure (Paetkau et al. 1995), alleles of eachsimulated individual were added to the arrays of allelic dis-tributions of every breed, except its own.
RESULTS
Intra-breed Genetic VariabilityThe 10 microsatellite primers detected 93 alleles in the 10breeds (Table 2). The number of alleles at each locus in the
Table 2. Number of alleles at the 10 loci for the 10 breeds
Breedz
Locus NCC ICE ROM SUF TEX CHE DOR FIN SBF MAS Pooled
MAF64 7 10 5 7 9 8 10 9 5 10 16 OarFCB11 8 7 6 7 7 6 6 7 8 5 10 OarFCB128 6 7 3 7 5 5 6 4 6 5 8OarCP26 6 7 5 11 5 5 7 9 5 5 14OarFCB5 3 1 2 3 3 2 3 3 3 2 3OarAE129 5 4 3 3 2 5 3 4 4 3 8OarAE101 3 5 3 5 7 6 5 4 6 2 9OarCP21 5 5 5 5 5 4 6 6 5 4 6TGLA53 8 7 8 7 6 10 9 9 10 10 12MAF35 5 4 3 4 5 4 3 5 6 4 7Average No. 5.6 5.7 4.3 5.9 5.4 5.5 5.8 6.0 5.8 5.0 9.3
z In this and other Tables: NCC = North Country Cheviot, ICE = Icelandic, ROM = Romanov, SUF = Suffolk, TEX = Texel, CHE = Cheviot, DOR = Dorset, FIN = Finnish Landrace, SBF = Scottish Blackface, MAS = Red Masai.
Can
. J. A
nim
. Sci
. Dow
nloa
ded
from
pub
s.ai
c.ca
by
Uni
vers
ity o
f T
enne
ssee
on
03/0
1/13
For
pers
onal
use
onl
y.
12 CANADIAN JOURNAL OF ANIMAL SCIENCE
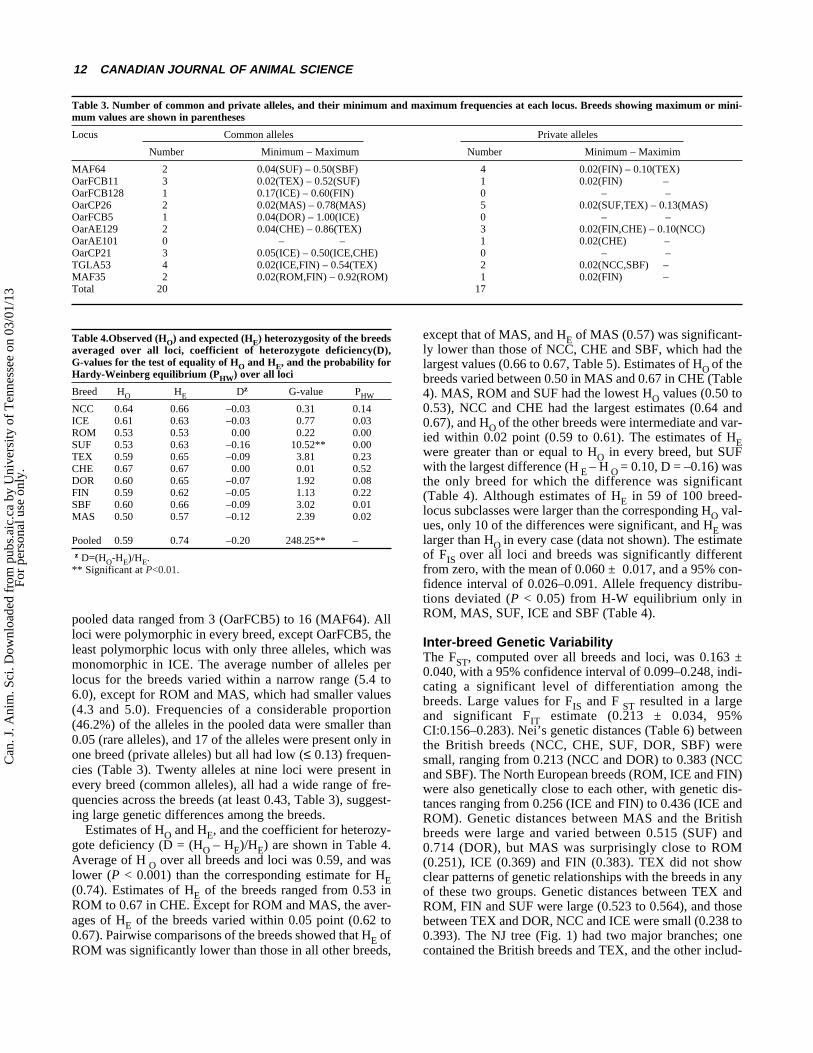
pooled data ranged from 3 (OarFCB5) to 16 (MAF64). Allloci were polymorphic in every breed, except OarFCB5, theleast polymorphic locus with only three alleles, which wasmonomorphic in ICE. The average number of alleles perlocus for the breeds varied within a narrow range (5.4 to6.0), except for ROM and MAS, which had smaller values(4.3 and 5.0). Frequencies of a considerable proportion(46.2%) of the alleles in the pooled data were smaller than0.05 (rare alleles), and 17 of the alleles were present only inone breed (private alleles) but all had low (≤ 0.13) frequen-cies (Table 3). Twenty alleles at nine loci were present inevery breed (common alleles), all had a wide range of fre-quencies across the breeds (at least 0.43, Table 3), suggest-ing large genetic differences among the breeds.
Estimates of HO and HE, and the coefficient for heterozy-gote deficiency (D = (HO – HE)/HE) are shown in Table 4.Average of HO over all breeds and loci was 0.59, and waslower (P < 0.001) than the corresponding estimate for HE(0.74). Estimates of HE of the breeds ranged from 0.53 inROM to 0.67 in CHE. Except for ROM and MAS, the aver-ages of HE of the breeds varied within 0.05 point (0.62 to0.67). Pairwise comparisons of the breeds showed that HE ofROM was significantly lower than those in all other breeds,
except that of MAS, and HE of MAS (0.57) was significant-ly lower than those of NCC, CHE and SBF, which had thelargest values (0.66 to 0.67, Table 5). Estimates of HO of thebreeds varied between 0.50 in MAS and 0.67 in CHE (Table4). MAS, ROM and SUF had the lowest HO values (0.50 to0.53), NCC and CHE had the largest estimates (0.64 and0.67), and HO of the other breeds were intermediate and var-ied within 0.02 point (0.59 to 0.61). The estimates of HEwere greater than or equal to HO in every breed, but SUFwith the largest difference (HE – HO = 0.10, D = –0.16) wasthe only breed for which the difference was significant(Table 4). Although estimates of HE in 59 of 100 breed-locus subclasses were larger than the corresponding HO val-ues, only 10 of the differences were significant, and HE waslarger than HO in every case (data not shown). The estimateof FIS over all loci and breeds was significantly differentfrom zero, with the mean of 0.060 ± 0.017, and a 95% con-fidence interval of 0.026–0.091. Allele frequency distribu-tions deviated (P < 0.05) from H-W equilibrium only inROM, MAS, SUF, ICE and SBF (Table 4).
Inter-breed Genetic VariabilityThe FST, computed over all breeds and loci, was 0.163 ±0.040, with a 95% confidence interval of 0.099–0.248, indi-cating a significant level of differentiation among thebreeds. Large values for FIS and FST resulted in a large and significant FIT estimate (0.213 ± 0.034, 95%CI:0.156–0.283). Nei’s genetic distances (Table 6) betweenthe British breeds (NCC, CHE, SUF, DOR, SBF) weresmall, ranging from 0.213 (NCC and DOR) to 0.383 (NCCand SBF). The North European breeds (ROM, ICE and FIN)were also genetically close to each other, with genetic dis-tances ranging from 0.256 (ICE and FIN) to 0.436 (ICE andROM). Genetic distances between MAS and the Britishbreeds were large and varied between 0.515 (SUF) and0.714 (DOR), but MAS was surprisingly close to ROM(0.251), ICE (0.369) and FIN (0.383). TEX did not showclear patterns of genetic relationships with the breeds in anyof these two groups. Genetic distances between TEX andROM, FIN and SUF were large (0.523 to 0.564), and thosebetween TEX and DOR, NCC and ICE were small (0.238 to0.393). The NJ tree (Fig. 1) had two major branches; onecontained the British breeds and TEX, and the other includ-
Table 3. Number of common and private alleles, and their minimum and maximum frequencies at each locus. Breeds showing maximum or mini-mum values are shown in parentheses
Locus Common alleles Private alleles
Number Minimum – Maximum Number Minimum – Maximim
MAF64 2 0.04(SUF) – 0.50(SBF) 4 0.02(FIN) – 0.10(TEX) OarFCB11 3 0.02(TEX) – 0.52(SUF) 1 0.02(FIN) –OarFCB128 1 0.17(ICE) – 0.60(FIN) 0 – –OarCP26 2 0.02(MAS) – 0.78(MAS) 5 0.02(SUF,TEX) – 0.13(MAS)OarFCB5 1 0.04(DOR) – 1.00(ICE) 0 – –OarAE129 2 0.04(CHE) – 0.86(TEX) 3 0.02(FIN,CHE) – 0.10(NCC)OarAE101 0 – – 1 0.02(CHE) –OarCP21 3 0.05(ICE) – 0.50(ICE,CHE) 0 – –TGLA53 4 0.02(ICE,FIN) – 0.54(TEX) 2 0.02(NCC,SBF) –MAF35 2 0.02(ROM,FIN) – 0.92(ROM) 1 0.02(FIN) –Total 20 17
Table 4.Observed (HO) and expected (HE) heterozygosity of the breedsaveraged over all loci, coefficient of heterozygote deficiency(D), G-values for the test of equality of HO and HE, and the probability forHardy-Weinberg equilibrium (P HW) over all loci
Breed HO HE Dz G-value PHW
NCC 0.64 0.66 –0.03 0.31 0.14ICE 0.61 0.63 –0.03 0.77 0.03ROM 0.53 0.53 0.00 0.22 0.00SUF 0.53 0.63 –0.16 10.52** 0.00TEX 0.59 0.65 –0.09 3.81 0.23CHE 0.67 0.67 0.00 0.01 0.52DOR 0.60 0.65 –0.07 1.92 0.08FIN 0.59 0.62 –0.05 1.13 0.22SBF 0.60 0.66 –0.09 3.02 0.01MAS 0.50 0.57 –0.12 2.39 0.02
Pooled 0.59 0.74 –0.20 248.25** –z D=(HO-HE)/HE.** Significant at P<0.01.
Can
. J. A
nim
. Sci
. Dow
nloa
ded
from
pub
s.ai
c.ca
by
Uni
vers
ity o
f T
enne
ssee
on
03/0
1/13
For
pers
onal
use
onl
y.

FARID ET AL. — GENETIC ANALYSIS OF SHEEP BREEDS 13
ed the North European breeds and MAS. The tree suggestthat TEX was genetically closer to the British breeds than tothe North European breeds, and SBF was the closest of theBritish breeds to the North European breeds.
Between 90.2% (CHE) and 99.7% (ROM) of the 1000simulated individuals were correctly assigned (Table 7).The largest number of misassigned animals were fromBritish breeds, most of them were misassigned to otherBritish breeds. Among the British breeds, CHE sheep weremost often misassigned to NCC and vice versa, suggestingthat they were genetically the closest of the British breeds.In agreement with the results of phylogenetic analysis, TEXsheep were misassigned more frequently to the British
breeds than to North European breeds or MAS. A few indi-viduals (1.8%) were incorrectly assigned to MAS, and a fewMAS sheep (0.6%) were incorrectly assigned to the NorthEuropean breeds and none to the British breeds or TEX,confirming the remoteness of the MAS from all other breedsstudied, particularly the British breeds and TEX.
DISCUSSION
Intra-breed Genetic VariabilityThe similarities among breeds for the average number ofalleles per locus and HE, except for ROM and MAS, werestriking in view of the diverse origin of the breeds and their
Table 5. Pairwise comparisons of expected heterozygosity of the breeds (probability of likelihood ratio χ2)
Breed ICE ROM SUF TEX CHE DOR FIN SBF MAS
NCC 0.41 0.00 0.47 0.78 0.84 0.78 0.34 0.94 0.04ICE 0.02 0.94 0.59 0.30 0.60 0.86 0.46 0.16ROM 0.02 0.00 0.00 0.00 0.04 0.00 0.38SUF 0.66 0.36 0.67 0.81 0.53 0.17TEX 0.63 0.99 0.49 0.85 0.07CHE 0.63 0.25 0.78 0.02DOR 0.51 0.84 0.07FIN 0.38 0.25SBF 0.04
Table 6. Nei genetic distances among the 10 breeds
Breed ICE ROM SUF TEX CHE DOR FIN SBF MAS
NCC 0.560 0.792 0.365 0.393 0.222 0.213 0.475 0.383 0.694ICE 0.436 0.769 0.384 0.501 0.647 0.256 0.464 0.369ROM 0.447 0.525 0.387 0.472 0.405 0.262 0.251SUF 0.564 0.222 0.305 0.484 0.338 0.515TEX 0.445 0.238 0.523 0.465 0.637CHE 0.265 0.297 0.343 0.531DOR 0.488 0.278 0.714FIN 0.493 0.383SBF 0.455
Fig. 1. Unrooted neighbour-joining tree relating the tenbreeds using the Nei’s genticdistance matrix. Bootstrap val-ues at the tree nodes were com-puted over 1000 replications,and indicate the number of treeswhere breeds beyond the node(left) occurred together.
Can
. J. A
nim
. Sci
. Dow
nloa
ded
from
pub
s.ai
c.ca
by
Uni
vers
ity o
f T
enne
ssee
on
03/0
1/13
For
pers
onal
use
onl
y.

14 CANADIAN JOURNAL OF ANIMAL SCIENCE
different developmental histories. The small average num-ber of alleles per locus in ROM may be the result of a smallnumber of founder animals, which were imported to NorthAmerica (5 rams and 14 ewes from 1981 importation and 10pregnant ewes imported in 1985; Fahmy 1996b). Althoughmore ROM sheep have been imported to North America inrecent years, it is possible that representatives of thesenewer lines were not included in the sample used in thisstudy. The FIN sheep population in North America was alsoestablished with a small number of animals (Fahmy 1996b),but this breed had the largest number of alleles per locusamong the breeds (6.0) and a significantly larger HO thanthat in ROM. This may be due to backcrossing of variousbreeds to FIN rams, making them eligible for registration asa purebred when they reached 15/16 FIN ancestry (Fahmy1996b).
The sample of MAS breed was obtained from a flockoriginating from across Kenya, yet it had the second small-est mean number of alleles per locus (5.0). The Awassibreed was reported to have a lower microsatellite geneticvariation than five Spanish sheep breeds, including theMerino (Arranz et al. 1998). Our results on MAS and that ofArranz et al. on Awassi sheep may suggest that either theEuropean breeds descended from a mixture of several typesof ancestral sheep, or unimproved Asian and African breeds,such as MAS, have been kept in isolation for an extendedperiod of time. It is also possible that the latter breedsencountered population crashes, possibly because ofdrought, which resulted in the loss of some low frequencyalleles (Luikart et al. 1998a,b). Low HE values in ROM andMAS were the reflection of the small number of alleles perlocus in these breeds.
The low estimate of HO in SUF, its significantly greaterHE than HO, and a rather high level of genetic variability(average number of alleles per locus of 5.9), suggest thatwhile SUF had a wide genetic base, individual animals weremore inbred than those in most other breeds. The rather highlevel of genetic variability in SUF is not surprising in viewof the fact that this is the most numerous sheep breed inNorth America. The high level of homozygosity may be dueto non-detectable alleles (Callen et al. 1993; Pemberton etal. 1995) or the Wahlund effect, caused by population sub-division into lines or families (Hartl and Clark 1989). Thisphenomenon results in a portion of total genetic variability
to reside among lines, and while the population as a wholepossesses a considerable level of genetic variability, indi-vidual animals are inbred (Nei 1965). There might havebeen a large genetic drift in SUF resulting from high selec-tion pressures that have been applied within each family fordecades. In addition, the physical characteristics and pro-duction performance of SUF is so different from those ofmost other breeds in North America that the likelihood ofany intentional crossbreeding to improve its productivity isnegligible. On the contrary, crossing in other pure registeredbreeds might have occurred in the past which expanded theirgenetic variability. The ROM and MAS breeds, which alsohad low levels of genetic variability, were not possibly sub-divided into lines, and consequently, estimates of HE andHO were not significantly different. The results suggest thatthere have not been drastic losses of genetic variability inthe intensely selected breeds, such as SUF, DOR and TEX,and that the effect of small numbers of founder animals onthe depletion of genetic variability has been more pro-nounced than the intense phenotypic selection pressure. Theresults also imply that the use of estimated breeding valuesin the future, which will result in the extensive use of a smallnumber of elite animals (Quinton et al. 1992; Gama andSmith 1993), may accelerate the rate of reduction of thegenetic variability.
Significant departures of allele frequency distributionsfrom H-W proportions in SUF, ICE, SBF and MAS breedswere caused by excess of homozygosity (larger HE thanHO). Although ROM was inbred, and its allele frequencydistributions significantly deviated from H-W proportions,the estimates of HE and HO were the same. This was theresult of large differences among loci for HE and HO inROM, which were often in opposite directions, a phenome-non that has been observed in populations that have experi-enced bottlenecks (Tarr et al. 1998).
Inter-breed Genetic Variability Gene flow among most of the breeds used in this study hasbeen restricted by geographical and(or) physical (herd-books and pedigree registration) isolation. The limited geneflow has resulted in considerable degrees of genetic differ-entiation among breeds indicated by differences across thebreeds for the frequency of common alleles. Sample sizes inthis study (25) provided a high probability (92.3%) ofdetecting an allele with the frequency of 0.05.
Table 7. Results of assigning 1000 simulated individuals from each of the 10 breeds (row headings) to every breed (column headings)
Breeds DOR NCC SBF CHE SUF FIN ICE ROM TEX MAS
Dorset 929 12 14 6 5 1 0 0 33 0 NC Cheviot 10 927 11 39 8 1 0 0 2 2S. Blackface 11 13 925 17 6 5 4 13 2 4 Cheviot 4 36 13 902 12 15 1 11 3 3Suffolk 5 4 10 18 961 1 0 0 1 0 Finnsheep 2 0 1 19 2 965 4 1 2 4 Icelandic 0 0 7 2 0 5 973 5 5 3 Romanov 0 0 0 2 0 0 0 997 0 1Texel 34 3 3 4 2 2 7 0 944 1 Red Masai 0 0 0 0 0 1 1 4 0 994
Can
. J. A
nim
. Sci
. Dow
nloa
ded
from
pub
s.ai
c.ca
by
Uni
vers
ity o
f T
enne
ssee
on
03/0
1/13
For
pers
onal
use
onl
y.

FARID ET AL. — GENETIC ANALYSIS OF SHEEP BREEDS 15
The pattern of differentiation revealed by the matrix ofNei’s genetic distances, the tree topology and the assign-ment test, reflected the evolutionary history, geographicaldistributions and the gene flow among the breeds.Differentiation between the British and North Europeanbreeds was anticipated in view of the fact that there has notbeen gene flow between them in the recent history. ROM,ICE and FIN, which showed close genetic relationships,descended from the North European Short-tailed type ofsheep (Ryder 1983). FIN and ICE had the smallest geneticdistance among the North-European Short-tailed breeds,which may be the result of divergence of ICE from the OldNorwegian sheep (Ponting 1980), whose habitat was closeto that of FIN. It has been suggested that ancestors of FINoriginated from the eastern side of the Ural Mountains inRussia (Maijala 1996), which is geographically close to thearea where ROM originated. Similarities between FIN andROM for allele frequencies in several biochemical poly-morphic systems have led to the conclusion that either ROMand FIN had common ancestry (Maijala 1996; Fahmy1996a) or, since both breeds are prolific, those proteinscould be markers of fertility (Maijala 1996). Our results sup-port the former hypothesis, because the majority ofmicrosatellite markers are selectively neutral, and ICE is nota prolific breed.
The observation that MAS was genetically closer to ROMthan were any of the North European breeds to each other,suggests that ancestors of both North European Short-tailedbreeds and sheep that were introduced to east Africa perhapsdiverged from the same type of ancestral sheep. Althoughthe time of introduction of MAS to Africa is not known, theclose genetic relationship between ROM and MAS maysuggest that sheep were spread from their centre of domes-tication in south-west Asia (Ryder 1983), and the same typethat reached the Ural Mountains also spread into northAfrica. The fat-tail of MAS might have evolved after diver-gence from ROM, either in south-west Asia or in north-eastAfrica, through human selection or as an adaptive mecha-nism to withstand dry climates.
Most of the British breeds were established in the mid1700s to the early 1800s from three types of sheep; Black-Faced Horned, White-Faced Hornless and Soay (Ryder1983). SUF, SBF and Southdown are descendants of Black-faced Horned sheep, DOR and CHE are descendants ofSoay, while White-faced Hornless sheep contributed tomany breeds, including SUF, DOR, CHE, Southdown,Leicester and Lincoln (Ryder 1983). Southdown seems tohave contributed to SUF, DOR and CHE, and Leicester con-tributed to NCC, DOR and CHE (Briggs 1958; Ryder 1983).Close genetic relationships between these breeds were,therefore, anticipated. The exact time and the extent of geneflow among the British breeds are subject to speculation.This makes it difficult to correlate the estimated time ofestablishment of the breeds and their lines of ancestry withthe results obtained in this study. Furthermore, the possibil-ity of backcrossing in the ancestry of some registered breedsin North America adds another element of uncertainty to thehistorical evidence on the time of divergence of these breeds.
The closest genetic relationship among the British breeds,
based on the assignment test, was between NCC and CHE,which was expected in view of the fact that NCC derivedfrom CHE (Briggs 1958). The genetic distance betweenNCC and CHE was, however, close to that between NCCand DOR, and the latter breeds were clustered together inthe NJ tree. Although sampling error cannot be ruled out,genetic similarity between NCC and DOR may be the resultof gene flow from common sources, namely Leicester, toboth breeds (Briggs 1958). NCC, CHE and DOR had a com-mon ancestral type of sheep [Tan or White-Faced Hornedsheep of the British Isles (Ryder 1983)], and Southdown andMerino have possibly contributed to DOR and CHE. It ispossible that Leicester might have contributed to all thesethree breeds (Briggs 1958). The close genetic relationshipsbetween DOR and other British breeds may be due to thecontribution of all the three ancestral types of sheep of theBritish Isles to this breed (Briggs 1958; Ryder 1983). Thesmall genetic distance between SUF and CHE, and theobservation that SUF sheep were more frequently misas-signed to CHE than to any other breed, could be the resultof gene flow from Southdown to both breeds, as well as pos-sible contribution of Blackface Highland sheep to CHE(Briggs 1958). Little is known about the history of the SBFbreed, except that it descended from the Black-FacedHorned type of sheep (Ryder 1983; Ponting 1980). Theresults of this study did not clarify the evolutionary historyof SBF either, as this breed could not be unequivocallyassigned to either of the British or North-European branch-es. The results may suggest that SBF is the closest of theBritish breeds to the North-European breeds.
The TEX breed originated from the isle of Texel in theNorth Sea and was expected to be genetically close to theNorth European Short-tailed breeds. TEX lambs are occa-sionally born with short bare tails, suggesting the geneticrelatedness of TEX with the North European Short-tailedbreeds. The finding that TEX was genetically closer to theBritish breeds than to the North European Short-tailedbreeds is possibly the result of crossing the Old Texel breedof sheep with the British breeds, namely Leicester andLincoln (Mason 1988). The contribution of Leicester toDOR and NCC (Briggs 1958; Mason 1988) may explain theclose genetic relationships between TEX, DOR and NCC,which resulted in these breeds being clustered together inthe phylogenetic tree.
Genetic structure of a breed at any time is the result of abalance between genetic drift (founder effect and selection)and gene flow. There have been complex patterns of geneflow among most British breeds during the early years oftheir establishment (Briggs 1958; Ryder 1983), while MASand ICE, and perhaps ROM and FIN originated from onlyone type of ancestral sheep (Fahmy 1996a; Maijala 1996).Additionally, these breeds may have been kept in isolationlonger than the British breeds. Since the accuracy of phylo-genetic reconstruction increases as the time of divergenceincreases (Takazaki and Nei 1996), the tree topology of theBritish breeds are less accurate than those for the North European breeds. The estimates of FST were 0.105 ±0.025 (95% CI: 0.065–0.156) and 0.162 ± 0.038 (95%CI:0.102–0.245) for the five British breeds and the Short-tailed
Can
. J. A
nim
. Sci
. Dow
nloa
ded
from
pub
s.ai
c.ca
by
Uni
vers
ity o
f T
enne
ssee
on
03/0
1/13
For
pers
onal
use
onl
y.

16 CANADIAN JOURNAL OF ANIMAL SCIENCE
breeds (FIN, ROM, ICE, MAS), respectively. This confirmsthe high degree of differentiation among the latter breedscompared with the British breeds. Although sample sizeswere adequate in providing accurate tree topologies, thenumber of loci examined was not large enough to result inan efficient phylogenetic reconstruction (Takazaki and Nei1996).
The assignment test has been proven to be a powerfulmethod for identifying the population membership of indi-viduals in livestock (Buchanan et al. 1994; Belliveau et al.1999), fish (Estoup et al. 1998) and wildlife (Paetkau et al.1995; Reed et al. 1997; Simonsen et al. 1998). As expected,the number of animals from each breed that was misas-signed to other breeds was inversely related to the geneticdistances between the breeds. Correct classification of ahigh proportion of animals from each sample, even for thebreeds such as CHE and NCC that were genetically close toeach other, makes the assignment test with this panel ofmicrosatellites a useful procedure for protecting the integri-ty of pedigreed breeds. This is an issue of considerableimportance to the purebred sheep industry in Canada. Oneproblem with parentage identification, which is currentlyused by the Canadian sheep industry, is mis-mothering.Adoption of a lamb of another ewe occasionally happenswhen several ewes give birth at the same time, and parent-age will not be verified while the lamb is a purebred. If sam-pling is not supervised, parentage testing will notcompletely prevent fraud either. The correct parentage of acrossbred animal will be verified, as long as the DNA sam-ples of the actual sire and dam are tested. The use of theassignment test, in addition to parentage testing, will pro-vide a more accurate method of protecting the integrity ofpurebred animals. One of the limitations of the assignmenttest is that the test individual must be a purebred from abreed with known allele frequency distribution. Crossbredanimals may be assigned to a pure breed to which it has norelationship, but there are methods to reduce the likelihoodof such bias (Farid et al. 1998).
IMPLICATIONSComparison between the breeds which have been subjectedto various degrees of selection pressure suggest that geneticvariability, which is essential for the persistence of geneticimprovement, has not been exhausted in intensely selectedsheep breeds. The use of a small number of founder animalsseems to have a much larger effect on the depletion of genet-ic variability than intense phenotypic selection. The trendtowards using estimated breeding values in the future mayresult in the extensive use of small numbers of elite individ-uals, and could potentially accelerate the rate of reduction ingenetic variability. There seems to be a need for increasingthe genetic variability of the Romanov breed. Although indi-vidual Suffolk sheep seem to be more inbred than animals ofother improved breeds in North America, a considerableamount of genetic variability is still available within thisbreed. Apart from the information on the extent of geneticvariability of the breeds studied, the results indicated thatthe assignment test could be used to improve the accuracy of
the method of protecting the integrity of pure breeds inCanada. This requires establishment of a national databasefor all the pure breeds in Canada using a large number ofmicrosatellite markers.
ACKNOWLEDGEMENTSWe are grateful to Ms. Marg Zillig, from the Department ofAnimal Science, NSAC, Mr. Roy MacKenzie, SheepSpecialist at the Nova Scotia Department of Agriculture andMarketing, Mr. Mike Wallace, Sheep Operation Manager,Roman L. Hruska, US Meat Animal Research Center, ClayCenter, Nebraska, and Mr. Brian Magee, CornellUniversity, NY, USA, for their help in collecting blood sam-ples. DNA samples from the Red Masai sheep were kindlyprovided by Dr. Alan Teale and Dr. Olivier Hanotte, theInternational Livestock Research Institute, Kenya. DNAsamples of some of the Icelandic sheep were kindly provid-ed by Dr. Adsridur Palsdottir, Institute of ExperimentalPathology, Keldur, Iceland. Dr. James Mylne, EdinburghGenetics, Scotland, provided blood samples from someCheviot sheep. We are also obliged to a large number ofsheep breeders across Canada who responded to our requestfor blood sampling and providing pedigree information ontheir sheep. We appreciate the staff of CIOQ, La Pocatiere,Quebec, for providing semen samples. We wish to acknowl-edge Dr. J. Felsenstein for providing the PHYLIP computerprogram, and Drs. M. Raymond and F. Rousset for theGENEPOP program. Financial assistance for this projectwas provided by the Canada/Newfoundland Green Plan andthe Canada/Newfoundland Agricultural Safety Net program.
Arranz, J. J., Bayon, Y. And San Primitivo, F. 1998. Geneticrelationships among Spanish sheep using microsatellites. Anim.Genet. 29: 435–440.Bancroft, D. R., Pemberton, J. M. and King, P. 1995.Extensiveprotein and microsatellite variability in an isolated, cyclic ungulatepopulation. Heredity 74: 121–129.Barker, J. S. F. 1994.A global protocol for determining geneticdistances among domestic livestock breeds. Proc. 5th World Cong.Genet. Appl. Livest. Prod. University of Guelph, Guelph, ON 21:501–508.Belliveau, A. M., Farid, A., O’Connell, M. and Wright, J. M.1999. Assessment of genetic variability in captive and wildAmerican mink (Mustela vison) using microsatellite markers. Can.J. Anim. Sci. 79: 7–16. Briggs, H. M. 1958.Modern breeds of livestock. The MacMillanCo., New York, NY. p. 754.Bruford, M. W. and Wayne, R. K. 1993.Microsatellite and theirapplication to population genetic studies. Curr. Opin. Genet. Dev.3: 939–943.Buchanan, F. C., Adams, L. J., Littlejohn, R. P., Addox, J. F.and Crawford, A. M. 1994. Determination of evolutionary rela-tionship among sheep breeds using microsatellites. Genomics 22:397–403. Callen, D. F., Thompson, A. D., Shen, Y., Phillip, H. A.,Richards, R. I., Mulley, J. C. and Sutherland, G. R. 1993.Incidence and origin of “null” alleles in the (AC)n microsatellitemarkers. Am. J. Hum. Genet. 52: 922–927. Crawford, A. M, Dodds, K. G., Ede, A. J., Pierson, C. A.,
Can
. J. A
nim
. Sci
. Dow
nloa
ded
from
pub
s.ai
c.ca
by
Uni
vers
ity o
f T
enne
ssee
on
03/0
1/13
For
pers
onal
use
onl
y.

FARID ET AL. — GENETIC ANALYSIS OF SHEEP BREEDS 17
Montgomery, G. W., Garmonsway, H. G., Geattie, A. E.,Davies, K., Maddox, J. F. Kappes, S. W., Stone, R. T., Nguyen,T. C., Penty, J. M., Lord, E. A., Broom, J. E., Buitkamp, J.,Schwaiger, W., Epplen, J. T., Matthew, P., Matthews, M. E.,Hulme, D. J., Beh, K. J., McGraw, R. A. and Beattie, C. W.1995.An autosomal genetic linkage map of the sheep genome.Genetics 140: 703–724.Estoup, A., Rousset, F., Michalakis, Y., Cornuet, J.-M,Adriamanga, M. and Guyomard, R. 1998.Comparative analysisof microsatellite and allozyme markers: a case study investigatingmicrogeographic differentiation in brown trout (Salmo trutta).Mol. Ecol. 7: 339–353.Fahmy, M. H. 1996a. The Romanov. Pages 47–72 in M. H.Fahmy, ed., Prolific sheep. CAB International, University Press,Cambridge, UK. Fahmy, M. H. 1996b.The spread of prolific sheep breeds in vari-ous countries. The Americas. Pages 228–233 in M.H. Fahmy, ed.,Prolific sheep. CAB International, University Press, Cambridge,UK.Farid, A., Almudevar, A., Dollard, C. and O’Reilly, E. 1998.Classification of crossbred animals using microsatellites. 6thWorld Cong. Genet. Appl. Livest. Prod., Armidale, NSW,Australia 24: 208–211.Felsenstein, J. 1995. PHYLIP- Phylogeny Inference Package.,Version 3.75c. University of Washington, Seattle, WA.Gama, L. T. and Smith, C. 1993.The role of inbreeding depres-sion in livestock production systems. Livest. Prod. Sci. 36:203–211.Hartl, D. L. and A. G. Clark. 1989. Principles of populationgenetics. Sinauer Associates, Sunderland, MA. p. 682.Luikart, G., Allendorf, F. W., Cornuet, J.-M and Sherwin, W.B. 1998a.Distortion of allele frequency distributions provides atest for recent population bottlenecks. J. Hered. 89: 238–247.Luikart, G., Sherwin, W. B. Steele, B. M. and Allendorf, F. W.1998b.Usefulness of molecular markers for detecting populationbottlenecks via monitoring genetic change. Mol. Ecol. 7: 963–974.Maijala, K.1996. The Finnsheep. Pages 10–46 in M. H. Fahmy,ed. Prolific sheep. CAB International, University Press,Cambridge, UK.Mason, I. L. 1988.Dictionary of livestock breeds. 3rd ed. CABInternational, Wallingford, UK. p. 348.Montgomery, G. W. and Sise, J. A. 1990.Extraction of DNAfrom sheep blood cells. N.Z. J. Agric. Res. 33: 437–441.Nei, M. 1965.Variation and covariation of gene frequencies insubdivided populations. Evolution 19: 256–258.Nei, M. 1972.Genetic distance between populations. Am. Nat.106: 283–292.Nei, M. and Roychoudhury, A. K. 1974.Sampling variance ofheterozygosity and genetic distance. Genet. 76: 379–390.
Nei, M., Maruyama, T. and Chakraborty, R. 1975.The bottle-neck effect and genetic variability in populations. Evolution 29: 1–10. Notter, D. R. 1999.The importance of genetic diversity in live-stock populations of the future. J. Anim. Sci. 77: 61–69.Paetkau, D., Calvert, W. Stirling, I. and Strobeck, C. 1995.Microsatellite analysis of population structure in Canadian polarbears. Mol. Ecol. 4: 347–354.Pemberton, J. M., Slate, J., Bancroft, D. R. and Barrett, J. A.1995.Non-amplifying alleles at microsatellite loci: a caution forparentage and fingerprinting studies. Mol. Ecol. 4: 519–520.Pickard, L. 1987.Early sheep importations. Sheep Canada maga-zine 12:24.Ponting, K. 1980.Sheep of the world. Blandford Press Ltd.,Blandford, UK. p. 145.Quinton, M., Smith, C. and Goddard, M. E. 1992.Comparisonof selection methods at the same level of inbreeding. J. Anim. Sci.70: 1060–1067. Raymond, M. and Rousset, F. 1995.GENEPOP (Version 1.2):Population genetics software for exact tests and ecumenicism. J.Hered. 86: 248–249.Reed, J. Z.,Tollit, D. J., Thompson, P. M. and Amos, W. 1997.Molecular scatology: the use of molecular genetic analysis toassign species, sex and individual identity to seal faeces. Mol.Ecol. 6: 225–234. Ryder, M. L. 1983. Sheep and man. Duckworth & Co. Ltd.,London, UK. p. 846.Saitou, N. and Nei, M. 1987.The Neighbor-joining method: Anew method for reconstructing phylogenic trees. Mol. Biol. Evol.4: 406–425. SAS Institute, Inc. 1996.Statistical analysis system, SAS user’sguide. SAS Institute Inc., Cary, NC.Simonsen , B. T., Siegismund H. R. and Arctander, P. 1998.Population structure of African buffalo inferred from mtDNAsequences and microsatellite loci: high variation but low differen-tiation. Mol. Ecol. 7: 225–237.Takezaki, N. and Nei, M. 1996.Genetic distances and recon-struction of phylogenetic trees from microsatellie DNA. Genet.144: 389–399.Tarr, C. L., Conant, S. and Fleischer, R. C. 1998.Founderevents and variation at microsatellite loci in an insular passerinebird, the Laysan finch (Telespiza cantans). Mol. Ecol. 7: 719–731.Weir, B. S. 1990.Genetic data analysis. Methods for discrete pop-ulation genetic data. Sinauer Asso. Inc. Pub., Sunderland, MA.Wiener, G., Lee, G. J. and Wooliams, J. A. 1994.Consequencesof inbreeding for financial returns from sheep. Anim. Prod. 59:245–249. Wright, S. 1978.Evolution and the genetics of populations. Vol.4, Variability within and among natural populations. University ofChicago Press, Chicago, IL. 580 pp.
Can
. J. A
nim
. Sci
. Dow
nloa
ded
from
pub
s.ai
c.ca
by
Uni
vers
ity o
f T
enne
ssee
on
03/0
1/13
For
pers
onal
use
onl
y.

Can
. J. A
nim
. Sci
. Dow
nloa
ded
from
pub
s.ai
c.ca
by
Uni
vers
ity o
f T
enne
ssee
on
03/0
1/13
For
pers
onal
use
onl
y.