genesis providing an evolutionary framework for
TRANSCRIPT

Providing anevolutionary frameworkfor biodiversity science
bioGENESIS Science Plan and
Implementation Strategy
bio
GEN
ESIS

DIVERSITAS
Report N°6, bioGENESIS Science Plan andImplementation Strategy
© DIVERSITAS 2009 – ISSN: 1813-7105 ISBN: 2-9522982-7-0
Suggested citation: Michael J. Donoghue, Tetsukazu Yahara, Elena Conti, JoelCracraft, Keith A. Crandall, Daniel P. Faith, Christoph Häuser, Andrew P. Hendry,Carlos Joly, Kazuhiro Kogure, Lúcia G. Lohmann, Susana A. Magallón, Craig Moritz,Simon Tillier, Rafael Zardoya, Anne-Hélène Prieur-Richard, Anne Larigauderie, andBruno A. Walther. 2009. bioGENESIS: Providing an Evolutionary Framework for Biodiversity Science. DIVERSITAS Report N°6. 52 pp.
Cover images credits: J Cracraft, C Körner, B A Walther, D M Hillis, D Zwickl, and R Gutell
Contact address
Michael J. Donoghue, PhD
Department of Ecology and Evolutionary BiologyYale University21 Sachem StreetP.O. Box 208105New Haven, CT 06520-8105, USATel: +1-203-432-2074 Fax: +1-203-432-5176 Email: [email protected]
Tetsukazu Yahara, PhD
Department of BiologyFaculty of SciencesKyushu UniversityHakozaki 6-10-1812-8581 Fukuoka, JapanTel: +81-92-642-2622Fax: +81-92-642-2645Email: [email protected]
www.diversitas-international.org
I C S U
I U B S
S C O P E
UNESCO
© A Hendry

bio
GEN
ESIS
Providing an evolutionary frameworkfor biodiversity science
bioGENESIS Science Plan and Implementation Strategy
Authors: Michael J. Donoghue, Tetsukazu Yahara, Elena Conti, Joel Cracraft,Keith A. Crandall, Daniel P. Faith, Christoph Häuser, Andrew P. Hendry,Carlos Joly, Kazuhiro Kogure, Lúcia G. Lohmann, Susana A. Magallón,Craig Moritz, Simon Tillier, Rafael Zardoya, Anne-Hélène Prieur-Richard,Anne Larigauderie, and Bruno A. Walther.
Approved by the Scientific Committee of DIVERSITAS
© J
Crac
raft


3
The stunning diversity of life is being diminished atan unprecedented rate owing to human impacts onthe environment around the globe. Although muchprogress has been made in understanding biologicaldiversity since Carl Linaeus in the 18th century andCharles Darwin in the 19th, it is clear that we stillknow little of the true diversity of life on Earth. Ourlack of knowledge now severely compromises ourability to recognize and to respond appropriately tothe rapid environmental changes that are occurring.Never before has it been so urgent – for themaintenance of nature and for our own well-being –to discover, to monitor, and to maintain biologicaldiversity.
Although the study of evolution fundamentallyunderlies our understanding of biological diversity,evolutionary biologists – including systematists,paleontologists, biogeographers, and populationgeneticists – have played a rather limited role inresponding to the biodiversity crisis. Yet, it isincreasingly clear that the insights and tools ofevolutionary biology are not just useful, butnecessary in identifying and combating changes inbiodiversity. The successful application ofevolutionary biology in this arena requires increasedintegration, not only among the evolutionarydisciplines, but between evolutionary biologists,ecologists and social scientists, all focusing onproblems of immediate relevance to society.
In recognition of the need to integrate evolutionaryknowledge and perspectives, DIVERSITAS, theinternational programme of biodiversity science, hasinitiated a new Core Project, bioGENESIS. The aimof bioGENESIS is to catalyse the internationalcommunication and integrated research that arenecessary to bring evolutionary approaches to bearon pressing biodiversity issues in a timely fashionand on a global scale. bioGENESIS will inspire a newgeneration of research that will fuel a trulyintegrative, socially relevant biodiversity science.
It is already clear that several areas of evolutionaryinvestigation are of direct significance tounderstanding and managing biodiversity. Weurgently need new strategies and tools to discover
and properly document biodiversity, includinggenetic and phylogenetic diversity. We need tocoordinate interdisciplinary analyses of thedynamics of evolutionary diversification in the past,to make better predictions about responses toglobal change. We also need to understand andincorporate rapid evolutionary change in modellingresponses to anthropogenic drivers. Overall, wemust learn to harness evolutionary knowledge moreeffectively in our efforts to conserve biodiversityand promote human well-being. Corresponding tothese broad needs, three primary foci have beenidentified within bioGENESIS:
FOCUS 1 New strategies and tools for documentingbiodiversity
FOCUS 2 The causes and consequences ofdiversification
FOCUS 3 Evolution, biodiversity, and human well-being
This document, the Science Plan and ImplementationStrategy of bioGENESIS, is the result of internationalmeetings and consultations held over the past threeyears involving scientists from a variety of relevantdisciplines. The Science Plan is by no means an endin itself, but is intended to be a flexible road mapthat will evolve as new data and ideas are generated.We hope that it will engage more and more scientistsworking at the interface of evolutionary biology andbiodiversity science and will generate exciting newavenues of research. Similarly, we hope that theactivities proposed here will enable scientists,resource managers, and policy makers to implementprogrammes and policies that reflect the relevance ofevolution in addressing the biodiversity crisis.
Michael J. Donoghue, Tetsukazu Yahara
Co-chairs, bioGENESIS
Harold A. Mooney
Chair, Scientific Committee DIVERSITAS
Anne Larigauderie
Executive Director, DIVERSITAS
Pre
face
© J Cracraft

4
Michael J. Donoghue
Department of Ecology andEvolutionary BiologyYale UniversityUSA
Tetsukazu Yahara
Department of BiologyKyushu UniversityJapan
bio
GEN
ESIS
Sci
en
tifi
c Com
mit
tee
Co-chairs
Elena Conti
Institute of Systematic BotanyUniversity of ZürichSwitzerland
Joel Cracraft
Department of OrnithologyAmerican Museum of NaturalHistoryUSA
Keith A. Crandall
Brigham Young UniversityUSA
Daniel P. Faith
The Australian MuseumAustralia
Christoph Häuser
Staatliches Museum fürNaturkunde StuttgartGermany
Andrew P. Hendry
Redpath Museum McGill UniversityCanada
Kazuhiro Kogure
Ocean Research InstituteUniversity of TokyoJapan
Lúcia G. Lohmann
Instituto de BiociênciasUniversidade de São PauloBrazil
Susana A. Magallón
Universidad Nacional Autonomade MexicoMexico
Craig Moritz
Museum of Vertebrate ZoologyBerkeleyUSA
Simon Tillier
Département Systématique etEvolutionMuséum National d'HistoireNaturelleFrance
Rafael Zardoya
Department of Biodiversity andEvolutionary BiologyMuseo Nacional de CienciasNaturalesSpain
Scientific Committee Members
© A Hendry

I - DIVERSITAS 6
II - The bioGENESIS Science Plan
FOCUS 1. New strategies and tools for documenting biodiversity 9
TASK 1.1 Discovering the unknown TASK 1.2 Capturing biodiversity information TASK 1.3 Developing phyloinformatics
FOCUS 2. The causes and consequences of diversification 18
TASK 2.1 Evolutionary change in diversity TASK 2.2 The evolutionary history of biotic assembly TASK 2.3 The evolution of functional traits TASK 2.4 Rapid evolution and eco-evolutionary dynamics
FOCUS 3. Evolution, biodiversity, and human well-being 29
TASK 3.1 Evolutionary ecosystem management TASK 3.2 Evolution and climatic change TASK 3.3 Combating diseaseTASK 3.4 Evolutionary conservation
III - Implementation Strategy 43
IV - References 47
V - Acknowledgements 49
VI - List of Acronyms 51
Table
of
Con
ten
ts
© A Hendry

6
DIVERSITAS is an international, non-governmental programme under theauspices of ICSU, IUBS, SCOPE and UNESCO(see side bar), that addresses the complexscientific questions posed by the loss ofand change in global biodiversity. Byconnecting individuals across natural andsocial science disciplines, and acrossnational or regional boundaries it addressesissues of global concern, thereby addingvalue to research projects being undertakenaround the world at the national anddisciplinary levels.
In accordance with the mandate developedby its sponsoring bodies, the mission ofDIVERSITAS is two-fold:
• To promote an integrative biodiversityscience, linking biological, ecological andsocial disciplines in an effort to producesocially relevant new knowledge.
• To provide the scientific basis for theconservation and sustainable use ofbiodiversity.
DIVERSITAS Sponsors
• International Council for Science (ICSU)
• International Union of Biological Sciences(IUBS)
• Scientific Committee on Problems of theEnvironment (SCOPE)
• United Nations Educational, Scientific andCultural Organisation (UNESCO)
Scientific Core Projects
The primary means by which DIVERSITAScarries out its mission is through catalysingresearch aligned with its four Scientific CoreProjects. Collectively, DIVERSITAS CoreProjects comprise a cycle of discovery,analysis and information sharing thatsupports the application of socially relevantknowledge:
• bioGENESIS provides an evolutionaryframework for biodiversity science,focusing on new strategies fordocumenting biodiversity, the causes andconsequences of diversification, andevolution in relation to biodiversityconservation and human well-being.
• bioDISCOVERY focuses on developing ascientific framework to investigate thecurrent extent of biodiversity, monitor itschanges, and predict its future changes.
• ecoSERVICES explores the links betweenbiodiversity and the ecosystem functionsand services that support human well-being, and seeks to determine humanresponses to changes in ecosystemservices.
• bioSUSTAINABILITY concerns itself withthe science-policy interface, looking forways to support the conservation andsustainable use of biological resources.
Cross-cutting Networks
DIVERSITAS also establishes Cross-cuttingNetworks, on specific topics or ecosystems,which embrace issues addressed in all fourCore Projects:
Global Mountain Biodiversity
Assessment (GMBA)
Steep terrain, extreme climates, and severeland-use pressure make mountainecosystems among the most endangered inthe world. Established in 2002, GMBAsynthesises knowledge on ethical,ecological, economic and aesthetic valuesto tackle issues of societal relevance,including land-use management practicessuch as fire, grazing and erosion.
Global Invasive Species Programme (GISP)
Non-native organisms that cause, or havethe potential to cause harm to theenvironment, economies, or human health,invasive alien species (IAS), are one of themost significant drivers of environmentalchange worldwide. GISP aims to conservebiodiversity and sustain human livelihoodsby minimising the spread and impact ofsuch species.
freshwaterBIODIVERSITY
Despite their critical role for basic lifesupport, freshwater ecosystems remainpoorly understood. In addition to identifyingand monitoring freshwater biodiversity andits role in ecosystem functioning, thisnetwork seeks to understand how biologicaland social processes interact.
DIV
ER
SIT
AS
I

7
agroBIODIVERSITY
Examining both agricultural and plantationsystems, this network promotes research onhow contrasting land-use patterns affectbiodiversity, ecological economics, andstandard economic gains.
ecoHEALTH
This network studies the relationshipsbetween plant and animal biodiversity and(re)emergence of infectious diseases andthe consequences for wild biodiversity andhuman societies.
Earth System Science Partnership(ESSP)
Recognising the links between biodiversityand other areas of global concern,DIVERSITAS is a founding member of theEarth System Science Partnership (ESSP;www.essp.org). In addition to DIVERSITAS,this partnership includes three otherprogrammes that focus on global issuessuch as climate change and human impactson the planet:
• International Geosphere-BiosphereProgramme (IGBP)
• International Human DimensionsProgramme on global environmentalchange (IHDP)
• World Climate Research Programme(WCRP)
Established in 2001, ESSP supports theintegrated study of the Earth system: itsstructure and functioning, change occurringwithin the System, and the implications ofchange for global sustainability. ESSPcurrently oversees four Joint Projects.
Global Environmental Change and Food
Systems (GECAFS) develops strategies toaddress food provision concerns while alsoanalysing the environmental andsocioeconomic consequences of adaptationand mitigation.
Global Carbon Project (GCP) investigatescarbon cycles and energy systems todevelop policy relevant knowledge thatencompasses natural and humandimensions, as well as their interactions.
Global Water System Project (GWSP)examines how humans are altering theglobal water cycle, the associatedbiogeochemical cycles, and the biologicalcomponents of the global water system, aswell as human response to these changes.This project is closely aligned tofreshwaterBIODIVERSITY.
Global Environmental Change and
Human Health project (GECHH)investigates how environmental changeworldwide affects human health and well-being, with the aim of developing policiesfor adaptation and mitigation. This projectis developed in conjunction withecoHEALTH.
National Committees and RegionalNetworks
One of DIVERSITAS' primary objectives is tocreate a worldwide network in support ofbiodiversity science that fosters integrationacross disciplines and establishes links atregional and international levels. Two typesof bodies play important roles in theachievement of this objective: NationalCommittees and Regional Networks.
National Committees enlarge DIVERSITAS'scientific and policy networks, thereby
helping to establish crucial links betweennational biodiversity programmes andinternational framework activities. They alsomake it possible to implement and, wherenecessary, to adapt the DIVERSITAS SciencePlan to local and regional concerns.Because many issues related to biodiversitytranscend national boundaries, it is oftenessential for several countries to collaboratein scientific research and policydevelopment.
The knowledge and experience gainedthrough such integrative approaches isinvaluable across the DIVERSITAS network.
Capacity building
The quest to expand knowledge aboutbiological diversity holds inherentchallenges. While most species are locatedin tropical areas, financial resources andtechnical capacity are severely lackingoutside the developed world. Thus, it iscritically important to pursue science whilealso making technological advances morewidely available and building the skillsnecessary to carry out integrative research.As far as possible, all DIVERSITAS activitieswill be designed to support directinvolvement of scientists from all regions ofthe world.

8
Evolutionary biologists – includingsystematists, paleobiologists,biogeographers, and population geneticists– have long endeavoured to document howmany species exist on Earth, to understandthe processes whereby these speciesoriginated and adapted, to chart how theyare distributed, and to infer how they arerelated to one another in the tree of life.These are monumental scientific challengesof direct relevance to understanding andconserving biodiversity. Yet, despite muchprogress, our knowledge of the earth'sdiversity remains far from complete. Infact, in some groups of organisms,including most microbial lineages, the taskof discovering unknown species has onlynow become possible, thanks to thedevelopment of new molecular andphylogenetic tools. Our limited knowledgeseriously compromises efforts to cope withthe rapid erosion of biodiversity around theglobe. We are under-prepared to recognisechanges that are occurring or to formulateappropriate responses.
While much fundamental work remains tobe done, we can formulate broad areas ofevolutionary investigation that are of directsignificance to understanding andmanaging biodiversity. New strategies andtools are urgently needed to discover andproperly document biodiversity. We need tocoordinate interdisciplinary analyses of thedynamics of evolutionary diversification inthe past, so as to provide better predictionsabout responses to global change. We alsoneed to understand and incorporate therapidity of evolutionary change in
modelling biodiversity responses toanthropogenic impacts. More generally, wemust make better use of evolutionarybiology in our efforts to conservebiodiversity and promote human well-being.
Addressing these key issues depends onbringing together, in various newcombinations, our ever-expanding baselinedata on species identities, distributions,and phylogenetic relationships, as well asknowledge of the relevant underlyingevolutionary processes. At present, theability to connect and employ thisinformation is limited and idiosyncratic.While some highly useful tools have beendeveloped within each domain, these are as
yet insufficient, and are poorly integratedand deployed, in part owing to limitedcoordination of the relevant scientificcommunities. DIVERSITAS, through thebioGENESIS Core Project, will play a keyrole in catalysing the internationalcommunication that is needed to tacklethese problems in a timely fashion and ona global scale. In turn, this will promote atruly integrative biodiversity science thatserves the needs of society.
The key issues that require concertedresearch efforts by evolutionary biologistsare identified here as focal areas within thebioGENESIS Core Project of DIVERSITAS.These are highlighted because they are
© A
Hend
ry
Th
e b
ioG
EN
ESIS
Sci
en
ce P
lan
II

9
fundamental to our understanding of theorigins and maintenance of biodiversity, butalso because they will enhance ongoingefforts within the other DIVERSITAS CoreProjects and Cross-cutting Networks. Thislist of focal areas and tasks, however, is notintended to be comprehensive or exclusive.Rather, these are viewed as several amongmany evolutionary issues of direct relevanceto the core mission of DIVERSITAS tounderstand, conserve, and sustainably usebiodiversity. We expect that the scope ofbioGENESIS will be refined and expanded asits plan of work is carried out.
Within each focal area we illustrate a seriesof tasks with boxed examples, whichhighlight concrete studies and findings. Inaddition to the 22 boxes included in thisdocument, in the text below we reference aset of supplemental boxed examples(labelled Box S1, Box S2, etc.), which canbe accessed through the DIVERSITASbioGENESIS web pages(http://www.diversitas-international.org).These supplemental boxes provideadditional case-studies, and boxes will beadded to the web site as the activities ofbioGENESIS develop. Along with each task,we also provide a list of ResearchObjectives and Activities. These are notmeant to be exhaustive, but simply toprovide examples of activities of currentinterest. In all cases, our mission is topromote international networks, scientificparticipation, and access by a wide varietyof user communities.
FOCUS 1. New strategies and tools fordocumenting biodiversity
Tremendous progress has been made indiscovering and documenting biodiversitysince the time of Linnaeus. Over the pastthree centuries approximately 1.7 millionspecies have been described, yet it is clearthat many more species exist – easily morethan ten times the number discovered sofar (Heywood 1995). Likewise, we havediscovered and named only a tiny fractionof the more inclusive branches of the treeof life, which provide us with thefoundation for organizing and navigating allbiological information (Cracraft andDonoghue 2004). At the same time,biodiversity is being lost at anunprecedented rate owing to humanactivities (Pimm et al. 1995, 2006; Hugheset al. 1997). In well-studied groups oforganisms, such as birds and mammals,extinction rates now far exceed thebackground rates characteristic of most ofgeological time. If systematic biologyproceeds along its current trajectory, wehave no hope of discovering the vastmajority of species before many (perhapsmost) of them are driven to extinction.
Dramatic changes are needed to greatlyaccelerate the rate of discovery. Althoughwe recognise the rapidly expanding need fortaxonomic expertise, and fully support theincreased training of specialists onparticular groups of organisms, we alsoappreciate that on the critical time scale– over the next 30 to 50 years – we are notlikely to experience a major increase in thenumber of taxonomists upon whom this
work ultimately depends. Therefore, wemust increase dramatically our efficiency– from the collection of specimens in thefield, to the discovery and description ofnew species and clades, to rendering thesedata readily accessible and interoperablewith other biodiversity information. The aimof Focus 1 is to identify and facilitate theimplementation of new strategies and toolsto greatly speed the pace of thisfundamental research, thereby providing amore effective pipeline from fieldexploration, through laboratory andmuseum studies, to databases accessible torelevant user communities. Here we seeespecially direct connections to theassessment efforts that form one of theprimary foci of the bioDISCOVERY CoreProject.
© M
Don
oghu
e

10
TASK 1.1 Discovering the unknown
DIVERSITAS, through bioGENESIS, will playa key role in establishing and maintaininginternational expertise, protocols,infrastructure, and data sharing mechanismsrelevant to the discovery and description ofbiodiversity. Through bioGENESIS workinggroups and the formation of strategicpartnerships with other relevantorganizations (GBIF, TDWG, IBOL, GTI, GEO,CoML, EDIT, etc.), we envision thedevelopment of international standards forbiodiversity repositories and “speciesdiscovery” centres, including best-practicesfor field work, for the collection andcuration of voucher specimens, for imagingand DNA sequencing technologies, for thedigitization of legacy data, for tissue – andDNA – banking and microbial culturefacilities, and for the development,maintenance, and connectivity of relevantdatabases.
We will especially promote efforts todocument the biodiversity of bacteria (Box 1),fungi (Box 2), and other microbes. Thesegroups are expected to contain the majorityof unknown taxa (Box S1), some of whichwill prove to be of enormous value tohumans (e.g., in the development of newdrugs and bio fuels; e.g., Smith et al. 2008;Box 2). As biological communities aregenerally composed of few dominantspecies, and many more that are rare,special attention must be given toimproving the efficiency of discovering anddescribing low-abundance organisms. Formicrobes, the sequencing of environmentalsamples (metagenomics) has created thepossibility of advancing our understandingof microbial diversity by orders ofmagnitude. Likewise, for small plants andanimals, new “industrial-strength” samplingprotocols have greatly accelerated thediscovery of unknowns (e.g., Bouchet et al.2002; Box S1).
bioGENESIS working groups will also addressissues surrounding (1) access to materialsand collection, which currently presentimpediments to progress in the discovery
and description of biodiversity; (2) thedevelopment of local taxonomic capacity(including the work-force of in-countrypara-taxonomists; Box S2), focusing onproviding training in low- and high-techfield, museum, and laboratory methods; and(3) access to, and utilisation of, spatialenvironmental information and new toolsfor spatial analysis, including GIS-basedniche-modelling and prediction techniques,to increase the efficiency of field-samplingand discovery.
Research Objectives and Activities
• Develop improved strategies and best-practices for global inventory and thediscovery and description of biodiversity,increasing the efficiency of fieldsampling and the quality andaccessibility of collections. Thisobjective integrates with activities beingcarried out under the GTI and in GBIF,and links directly with the bioDISCOVERYCore Project and its ongoinginvolvements in the GEOSS effort (GEOBON).
• Promote the application of nextgeneration DNA sequencing technologiesto reveal the biodiversity and functions ofmicrobes. This initiative relates directly tothe assessment of global genetic diversitydescribed below under Task 3.4.
• Facilitate the formulation of newpolicies regarding the collection ofbiological specimens for scientificresearch. This relates directly to Accessand Benefit Sharing (ABS) under theCBD. bioGENESIS activities have alreadystimulated discussions that areinforming the development of aninternational ABS regime.
© M
Don
oghu
e

11
BOX 1. Molecular approaches to discovering microbial diversity in the oceans
From the very beginning of life on Earth, microorganisms have been responsible for thecycling of materials through the biosphere. Although knowledge of the diversity andfunctioning of microbes is indispensable for understanding biological processes in theocean, relatively little is known to date. Most microbes are not yet cultured and thefunction of most of their genes is still unknown. It is highly likely that undiscoveredmicrobial diversity will reveal entirely new biological functions and chemical compounds,some of which will provide the raw material for human innovation into the future.
The application of molecular techniques, especially metagenomic approaches, has revealedan enormous diversity of undescribed, mostly uncultivable microbes (National ResearchCouncil 2007). Recent studies conducted in the Sargasso Sea using whole genome shotgunsequencing revealed the presence of 148 previously unknown bacterial phylotypes (Venteret al. 2004; Fig. 1.1). In addition, more than 1.2 million previously unknown genes wereidentified. Progress in sequencing technology has made it possible to analyse hugenumbers of genes very rapidly. Sogin et al. (2006), for example, developed the parallel tagsequence method and used this in combination with pyrosequencing technology. Theiranalyses revealed the presence of an immense number of low-abundance populations,which accounted for most of the observed phylogenetic diversity in deep-sea samples fromthe Atlantic (Fig. 1.2).
References
National Research Council. 2007. The new science of metagenomics: revealing the secrets of our microbial planet.Committee on Metagenomics: Challenges and Functional Applications, National Research Council. National AcademiesPress.
Sogin ML, Morrison HG, Huber JA, Welch DM, Huse SM, Neal PR, Arrieta JM and Herndl GJ. 2006. Microbial diversityin the deep sea and the underexplored “rare biosphere.” Proceedings of the National Academy of Sciences USA103:12115-12120
Venter JC, Remington K, Heidelberg JF,Halpern AL, Rusch D, Eisen JA, Wu D,Paulsen I, Nelson KE, Nelson W, Fouts DE,Levy S, Knap AH, Lomas MW, Nealson K,White O, Peterson J, Hoffman J,Parsons R, Baden-Tillson H, Pfannkoch C,Rogers YH and Smith HO. 2004.Environmental genome shotgunsequencing of the Sargasso Sea. Science304:66-74
FIG. 1.2. Rarefaction of microbes (after Sogin et al. 2006), ©K Kogure.OUT, Operational Taxonomic Units.
Number of tags sampled
OT
Us
FIG. 1.1. Collection of seawater for metagenomicanalyses, ©K Kogure.

12
BOX 2. “Myco-diesel” from an endophytic fungus
One argument for conserving biodiversity is that undiscovered species might yieldproducts of importance to humans. However, the link between undiscoveredbiodiversity and potential usefulness has largely been conjectural. Recent studies ofendophytic fungi living inside of tropical plants have provided direct evidence of thepotential usefulness of undescribed species (Strobel and Daisy 2003). Combiningfield collecting and culture studies with bioinformatic analyses and assays forbioactive compounds, Smith et al. (2008) showed that manypreviously unknown microbes produce bioactive compounds. Severalof the most genetically dissimilar and phylogenetically isolatedendophytes were among those that showed high levels ofbioactivity.
Studies of one such endophytic fungus, Gliocladium roseum, havedocumented the production of a series of volatile hydrocarbons– including octane and other compounds normally associated withdiesel fuel – when grown on a cellulose-based medium and extractsof the host flowering plant, Eucryphia cordifolia (Strobel et al.2008). This discovery of “myco-diesel” has clear implications for theproduction of biofuels, which migh potentially decrease reliance onfossil fuels.
References
Smith SA, Tank DC, Boulanger L-A, Bascom-Slack CA, Eisenman K, Kingery D, Babbs B,Fenn K, Greene JS, Hann BD, Keehner J, Kelley-Swift EG, Kembaiyan V, Lee SJ, Li P,Light DY, Lin EH, Ma C, Moore E, Schorn MA, Vekhter D, Nunez PV, Strobel GA,Donoghue MJ and Strobel SA. 2008. Bioactive endophytes warrant intensifiedexploration and conservation. Public Library of Science One 3(8):e3052
Strobel GA and Daisy B. 2003. Bioprospecting for microbial endophytes and theirnatural products. Microbiology and Molecular Biology Reviews 67(4):491-502
Strobel GA, Knighton B, Kluck K, Ren Y, Livinghouse T, Griffen M, Spakowicz D andSears J. 2008. The production of myco-diesel hydrocarbons and their derivatives by theendophytic fungus Gliocladium roseum. Microbiology 154:3319-3328
FIG. 2.1. Eucryphiacordifolia (Cunoniaceae),the host plant fromwhich Gliocladium roseumwas isolated, ©M Donoghue.
FIG. 2.2. Gliocladium sp.,showing hyphae and spores,©M Donoghue.

13
TASK 1.2 Capturing biodiversityinformation
The goal of this task is to improve methodsof capturing biodiversity information. ThebioDISCOVERY Core Project focuses uponadvancing biodiversity assessment, trackingbiodiversity changes, and developingscenarios of future biodiversity. ThebioGENESIS and bioDISCOVERY Core Projectstherefore share a common goal to improvethe methods of biodiversity assessment,across both spatial and temporal scales, atthe level of genes, species, and ecosystems,and in terms of the various functions andprocesses manifested by biodiversity.bioGENESIS will focus specifically onadvancing biodiversity assessment based onevolutionary biology, including systematicbiology, phylogenetics, and evolutionarygenetics. Both bioGENESIS andbioDISCOVERY will contribute to GEO BON, aglobal network of biodiversity observation(Box 3). Here too, the aim of bioGENESIS isspecifically to enable the deployment ofevolutionary methods.
The accurate identification of species isfundamental to biodiversity assessment. Itwould revolutionise our ability to inventoryand monitor biodiversity to develop rapid-capture technologies for identifying knownspecies and discovering new ones. Ofspecial interest is the development of acost-effective, hand-held, automatedspecies-identifier. The idea is to analyse atiny sample of an organism; quickly extract,amplify, and sequence a set of target DNAmarkers; and then to compare these toknown sequences to situate the unknown
within the tree of life. The design of such atool is already being pursued for medicaldiagnostic purposes, but a bioGENESISworking group could help to guide itsdevelopment to insure maximum benefit tothe biodiversity community. Perhaps mostimportantly, we must help to design theinterface between international “bar-coding” efforts (IBOL) and the rapidlyexpanding “tree of life” databases beinggenerated through individual andcoordinated efforts (e.g., the US NSF's ATOLprogramme).
A key bioGENESIS initiative will be tocontinue to help in connecting the bar-code and tree of life efforts. The first suchmeeting, convened at NESCent (Durham,North Carolina, USA) in 2007, focusedattention on what new insights might begained by linking these activities. A crucialrole will be to help to scale-up bar-codingstandards to multi-locus assays, and in thisway increase the connectivity to tree of lifedatabases. Additionally, such workshops willserve to promote the sharing of biologicalmaterials and the development ofcommunity-wide standards for DNAisolation, vouchering, and storage. They willalso address emerging issues surroundingthe environmental sequencing of microbes(metagenomics), including the use of thesesequences in isolating currentlyuncultivable organisms and identifying theirphenotypes.
New technologies also are needed to makeoptimal use of digital images. This includesthe utilisation of satellite technology forreal-time analysis of field images, the use
of remote-controlled internet cameras andmicroscopes, and the development of imagerecognition identification tools madeavailable as web-services. It is also crucialto develop digital tools to extract data thatare currently locked in the world's naturalhistory museums. There may be as many asthree billion specimens in these collections,each one accompanied by at least someinformation on where and when it wascollected. We need automated methods tocapture these data, requiring little humanintervention (Donoghue and Smith 2005).An example is the HERBIS project(www.herbis.org; Box S3), which isattempting to produce a system of web-services to automatically capture andupload the label data associated with aherbarium specimen with a single click of adigital camera.
Another focus will be on streamlining thedescription of newly discovered species andclades, and developing better ways totranslate new discoveries into theknowledge-bases used by decision-makers,including the emerging “Encyclopedia ofLife” (EOL). Our bioGENESIS effort, whichwill be closely aligned with those of EDIT(Box 4) and related developments withinnatural history museums, will consider thedigitisation of existing taxonomic literatureand specimens and the development of newelectronic publishing mechanisms. We alsoneed to explore how nomenclatural codesand associated databases might best bedesigned to facilitate rapid publication andthe dissemination of new taxonomicdiscoveries.

14
Finally, bioGENESIS will coordinate withbioDISCOVERY on the development of bettermechanisms to utilise specimen data andrelated information to facilitate the work ofenvironmental policy makers and thestewards of natural resources on local,regional, and national levels (Box S4). Inparticular, bioGENESIS can help tocoordinate regional efforts (e.g., ERIN inAustralia, CONABIO in Mexico, andBIOTA/FAPESP in Brazil), and promote thewidespread use of successful strategies.
BOX 3. bioGENESIS and GEO BON
In recognition of the rapid erosion of biodiversity at the level of ecosystems, species,and genes, the international community established targets to significantly reducethe rate of biodiversity loss by 2010. To achieve this and future such targets, GEO,DIVERSITAS, and NASA announced the formation of a new global partnership, GEOBON (http://www.earthobservations.org/cop_bi_geobon.shtml). The newlyestablished Biodiversity Observation Network seeks to develop open resourcedatabases of various observations at the ecosystem, species and gene levels,coordinate and develop the monitoring of biodiversity changes in structure,composition, and function, and provide scientifically rigorous and up-to-dateknowledge about on-going biodiversity changes.
In parallel with goals for species and ecosystems, GEO BON will facilitate the globalmonitoring of genetic diversity, including species phylogenetic diversity, using acombination of remote sensing and in situ approaches (Scholes et al. 2008). Threebroad strategies for observation and analysis of genetic diversity will be employed byGEO BON:
(1) Repeated observation of specific genetic components in selected target species.
(2) Repeated observation of other components (e.g., geographic range) integratedwith models that create links to genetic diversity.
(3) Repeated observation of changes inland/water condition (e.g., using remotesensing) integrated with models that act as a“lens” for inferences about correspondinggenetic changes.
GEO BON will promote these strategies as a wayto enable a range of monitoring approaches,extending from detailed observations for keyspecies to model-based inferences of regionaland global genetic diversity loss. Incollaboration with GEO BON, and withbioDISCOVERY, bioGENESIS will play a leadingrole in designing and promoting global geneticdiversity assessments to monitor how rapidlygenetic diversity is being lost.
References
Scholes RJ, Mace GM, Turner W, Geller GN, Jürgens N,Larigauderie A, Muchoney D, Walther BA and Mooney HA.2008. Toward a global biodiversity observing system. Science321(5892):1044-1045
biodiversityindicators
maps ofpriority areas
conversationplans
ecosystemservices
biodiversity-based
ressource trends
researchoutputs
User-definedneeds
tabulations
models
GIS
attributemaps
Observationproducts
GEO
BO
N p
orta
l
remotelysensedimages
interactions • type• intensity• from• to
ecosystem • name• polygon• date• attributes
abiotic • climate• substrate• topography• disturbance
community • location• date• species 1• ...• species n
uses • product• offtake rate• value• users
Observation
specimen • species• location• date• source
population • species• location• date• abundance• func type
gene • species• location• date• gene• alleles
species • scientific name• authority• synonyms• folk name
gazetteer • place name• polygon or latitude & longitude
taxonomy • species• genus• family• class etc
Supportinginformation
Core of the biodiversity observation network
feedbackfeedback
FIG. 3.1. A schematic representation of the core data types, observation products and end uses of anintegrated biodiversity observation system. Most of the elements already exist, but are incomplete ordispersed among a wide range of partners. The proposed implementation strategy involves linking themusing data-sharing protocols, followed by incremental, needs-led and opportunistic growth (fromScholes et al. 2008).

15
BOX 4. EDIT Scratchpads: Unifying revisionary taxonomy on the Web
Using traditional methods and technologies our knowledge about species isaccumulating too slowly to provide even basic information about most living speciesin less than several centuries. This limits the contribution taxonomy can make tobiodiversity analysis. Modern IT technologies can increase both the rate of creationand the accessibility of knowledge on species, simply by individual researcherssharing a common on-line storage facility.
The idea behind the EDIT Scratchpads is simple: make web sites easy to get, easy touse, and easy to read by computers and humans alike. It starts from a vision: being ataxonomist, what would you like to see up on the web for your taxonomic group?Scratchpads are simply template web sites equipped with a number of tools andservices that make it easy to add taxonomically relevant material. Initially, this maybe no more than a personal statement about research on a group of organisms, but itmay grow quickly as it is shared with others having similar interests.
Scratchpads are equipped with a number of tools and services that make it easy toadd taxonomically relevant material. As an illustration, one of the first EDITScratchpads was devoted to the family Sciaroidea (fungus gnats). While the pageswere being built in the UK, taxonomists from Norway found the site through a searchengine, and with the permission of the site manager, started adding data of theirown. This epitomises the vision that the Scratchpads will nucleate new communitiesof taxonomic experts.
EDIT Scratchpads illustrate how taxonomy can be transformed through thedevelopment and use of newtools that go beyond a change inmethod and imply a deepcultural change in how scientificknowledge is produced anddisseminated. In the DIVERSITASframework, they contribute tostreamlining research to improveour understanding andsustainable management ofbiodiversity.
References
http://www.editwebrevisions.info
FIG. 4.1. Examples of Scratchpads servingdifferent community functions, ©D Robert.
Research Objectives and Activities
• Develop mechanisms to promoteelectronic access to specimen-basedinformation, such as best-practices fordigitization and georeferencing projects.
• Promote research on innovativeidentification tools and the developmentof electronic field guides. This connectsdirectly to elements within bioDISCOVERY.
• Collaborate with the Global MountainBiodiversity Assessment (GMBA) on theintegration of elevational data ingeoreferencing, distribution mapping,and niche-modelling.

16
TASK 1.3 Developing phyloinformatics
One of the most profound contributions ofthe systematics community to ourunderstanding of biodiversity is knowledgeof how species are related to one anotherin the tree of life (Cracraft and Donoghue2004). This phylogenetic knowledgeprovides the basis for organising (inpredictive classification systems) andnavigating biological information of allkinds. Yet, at the present time, our abilityto harness this rapidly expanding base ofphylogenetic information is greatlyhindered by the lack of an appropriatecyber-infrastructure allowing phylogeneticnavigation – through clade-based queries –of other biodiversity information. Forexample, we currently have no way toquickly and accurately chart the geographicdistribution of any particular clade ofinterest, such as the clade that includes theseveral major lineages of C4 grasses(Edwards et al. 2007). This is problematicas phylogenetic knowledge is accumulatingrapidly, and older classifications are quicklybecoming misleading.
We envision a bioGENESIS working groupforming strategic alliances with a variety ofrelevant partners to shape the developmentof a comprehensive phyloinformaticsinfrastructure. Of special relevance is theongoing effort under CIPRES to re-developand greatly extend the capabilities ofTreeBASE (www.treebase.org; Box 5). Anumber of efforts are also underway tocode phylogenetic trees in KML (thelanguage for drawing objects on GoogleEarth; http://earth.google.com/), which, inturn, will allow us to visualise patterns ofbiological diversification across the face ofthe planet. This technique is useful notonly in analysing the geographicalevolution of plants and animals (Box 5),
but also in determining the site of originand spread of infectious diseases (Box 6).
Likewise, the databasing efforts underlyingthe Tree of Life web project(www.tolweb.org) are crucial to thisdevelopment, as are the activities of theEncyclopedia of Life (EOL) and the NSF's
iPlant phylogeny project that are focusedon visualising large phylogenetic trees.There are many other such projects andconnections to be made, such as with themajor domain-specific data repositories,including GenBank and GBIF. Here we seespecial opportunities to developinternational standards and protocols forthe rapid, efficient connection of diversedata types. Among other things, this willentail the establishment of “ontologies” (orcontrolled vocabularies) for morphologicaland behavioural traits, and for connectingphenotypes to physiological and ecologicalfunctions.
Underlying all such efforts is the continueddevelopment of confident phylogeneticknowledge. bioGENESIS will play animportant role in helping to stimulate thisactivity internationally, and in coordinatingindividual and national efforts along thelines of the US NSF's ATOL project. Forexample, bioGENESIS has already beensuccessful in promoting tree of life researchfunding and collaborations within China(Hong et al. 2008). Likewise, bioGENESIScan help guide the development ofstandards for the naming of clades and thedatabasing of clade names.
Research Objectives and Activities
• Promote and coordinate internationalefforts to assemble the tree of life,including extension of the US NationalScience Foundation's “Assembling theTree of Life” (ATOL) programme (seehttp://atol.sdsc.edu/) to Europe, Asia,and other regions.
• Develop mechanisms to integratephylogenetic knowledge with geographic,morphological, paleontological, andecological data to promoteinterdisciplinary studies of biodiversitydynamics.
• Coordinate with ongoing efforts withinCIPRES, EOL, and iPlant on the assemblyand visualisation of large phylogenetictrees, especially to insure broadinternational participation and access.
© M
Don
oghu
e

17
BOX 5. Visualising Evolution on a Global Scale
Evolution, as represented by phylogeny, is a central, unifying, expressionof the history of life on Earth, and as such it can serve as a nexus for allsorts of biodiversity patterns: the interconnected causes of gene evolutionand adaptation; of ecological diversification and morphological change;and of vicariance and speciation. Phylogenetic data are being published inthe scientific literature in ever-increasing abundance (Fig. 5.1; Piel et al.2002), and we can estimate that at least 60,000 phylogenies have beenpublished since inexpensive computers and efficient DNA sequencingmethods became available. It is only natural to find ways to trace andvisualise spatial patterns of diversification. Google Earth,™ a freelyavailable computer programme for 3D rendering of the earth andgeoreferenced data, is a perfect choice for visualising the interplaybetween phylogeny and geography (e.g. Fig. 5.1).
A number of efforts are underway to code phylogenetic trees in KML– the format for rendering objects and features in Google Earth. Theseinclude the TreeBASE Google Earth tree browser(http://www.treebase.org/getrees/); SupraMap (Janies et al. 2007);Mesquite Cartographer(http://mesquiteproject.org/packages/cartographer/); andGeoPhyloBuilder (https://www.nescent.org/wg_EvoViz/GeoPhyloBuilder).Additionally, GeoPhyloBuilder is built on the ArcGIS information system(ESRI), which potentially takes the user beyond visualization, allowingcomputational analyses at the interface of phylogeny and geography.
References
Janies D, Hill AW, Guralnick R, Habib F, Waltari E and Wheeler WC. 2007. Genomic analysis andgeographic visualization of the spread of avian influenza (H5N1). Systematic Biology 56:321-329
Piel WH, Donoghue MJ andSanderson MJ. 2002. TreeBASE: adatabase of phylogeneticknowledge. In: Shimura J, WilsonK and Gordon D (Eds). Theinteroperable “Catalog of Life.”Research Report, NationalInstitute for EnvironmentalStudies No. 171
FIG. 5.1. Evolution of the TúngaraFrog across the Isthmus of Panama.Phylogenetic relationships show astrong congruence with geography,in which the earliest branchingevents in the tree reflect an ancientloss of a land bridge between thecontinents. The TreeBASE Google Earth tree browser (http://www.treebase.org/getrees/) generated thecoding of the tree for display in Google Earth, ©W Piel.
BOX 6. Geographic evolution and thespread of infectious diseases
One of the great services of evolutionarybiology arises through the examination ofevolutionary patterns in a geographic contextto gain insights into the historicalbiogeography of groups of species. The sametechniques developed to analyse natural historydata can be applied directly to the question ofthe origin, spread, and transmission ofinfectious diseases. One such studyinvestigated the spread of the “severe acuterespiratory syndrome” (SARS) virus, whichbecame a novel human infectious disease in2002. Using genomic data to estimatephylogenetic relationships, the resulting treeshows the rapid and global spread of the SARSvirus (Fig. 6.1). The tree also shows host shiftsinto carnivore lineages, contradicting theoriginal hypothesis of transmission fromcarnivores to humans. Similar approaches haveshed light on the origin and spread of West Nilevirus, Hanta virus, and other such emergingdiseases.
References
Janies D, Habib F, Alexandrov B, Hill A and Pol D. 2008.Evolution of genomes, host shifts and the geographic spreadof SARS-CoV and related coronaviruses. Cladistics 24:111-130
FIG. 6.1. Phylogeny ofSARS-CoV (yellowlineages) and relatedcoronoviruses frompotential sourcepopulations (othercoloured lineage).Superimposingphylogeny ongeography (usingGoogle Earth) allows usto visualise the originof the virus in Asia andits rapid evolution
(represented by long branch lengths) and spread through human populations,©D Janies.

18
FOCUS 2. The causes and consequences ofdiversification
Our understanding and management ofextant biodiversity can benefit greatly fromknowledge of changes in diversity throughtime. This provides both a historicalperspective within which to interpretmodern patterns and a framework forgenerating predictions about futurechanges. The idea behind Focus 2 is tounderstand how various drivers (climatechange, episodes of invasion, etc.) haveinfluenced biodiversity in the past, so as toenable better predictions of future change.
Great progress has been made inunderstanding the causes and consequencesof diversification through time using acombination of paleontological andphylogenetic approaches. However, thereare still wide gaps both in knowledge andin communication among the practitionersof these disciplines, as well as with thosestudying the genetics of speciation andadaptive radiation. Furthermore, andperhaps most importantly from thestandpoint of DIVERSITAS, connectionsbetween evolutionary research, includingthe possibility of rapid evolutionary change,and ecology, including the study of present-day ecosystem function and the design ofpredictive models of biodiversity change,have been very limited. Through thebioGENESIS Core Project, DIVERSITAS willimprove integration among these disparatecommunities.
TASK 2.1 Evolutionary change indiversity
It is clear that past environmental changeshave impacted diversity on a variety ofspatial and temporal scales. Such problemshave been approached from variousdirections, with results of varying accuracyand precision. For example, Egan andCrandall (2008) successfully documentedhow diversification rates changed with pastglobal climate changes resulting fromPleistocene glacial cycles (Box S5).However, it remains difficult in many cases– using paleontological data directly, orusing phylogenies – to distinguish among avariety of possible causal factors (e.g.,climate change vs. entry of a clade into anew geographic region vs. the origin ofinnovations in a clade) and downstreamconsequences (e.g., on rate of speciationvs. rate of extinction; Moore and Donoghue2007). One common concern relates to theprecision with which we can infer thetiming of lineage-splitting events. Anotheris the reliability of the models used tolocate shifts in diversification in the fossilrecord or in phylogenetic trees.
A bioGENESIS working group will focusdirectly on these issues. The aim of thisgroup will be to enhance internationalcommunication across the relevantdisciplines, and also to refine the key toolsand models needed to make comparisonsfrom which relevant biodiversity predictionscan be drawn. For example, it may bepossible to refine measures of the lineage-specific or functional selectivity of massextinctions in the past, or the resilience orrebound dynamics following mass extinctionevents, again measured in bothphylogenetic and functional terms. More
generally, this activity would encourage theintegration of tree-based andpaleobiological approaches, and wouldexpand modelling efforts to encompass thewide variety of outcomes now apparent inphylogenetic comparisons (e.g., theexistence of ancient clades now containingjust one or a few long-surviving species).
Critical to our understanding of the genesisof biodiversity is the relationship betweenspeciation, the evolution of adaptive traits,and patterns of geographical distribution.Aspects of the problem have been studiedin relative isolation by phylogeneticbiologists, students of speciation,evolutionary developmental biologists,
Research Objectives and Activities
• Promote the integrated analysis(involving phylogeneticists,paleobiologists, population geneticists,and ecologists) of rates of speciationand extinction in relation toenvironmental changes, morphologicaland physiological innovations, and themovement of lineages into newgeographic regions.
• Develop better methods toextrapolate/predict overall biodiversitypatterns from studies of selectedlineages, e.g., based on responses tocommon environmental gradients andhistorical processes.
• Apply these new approaches in thecontext of a chosen system; e.g.,working with the Global MountainBiodiversity Assessment (GMBA) ondiversification at high elevations.

19
paleobiologists, and biogeographers.Integration across these disciplines wouldprovide a much clearer understanding of therelationship between the origin of noveltieswithin populations and the evolution ofspecies differences, and between modes ofspeciation (sympatric, allopatric, etc.) andpatterns of character evolution and/or theresulting distribution of geographic ranges.In-depth studies on groups showingexceptionally high diversity, such as cichlidfishes (Box 7), and those involving hybridspeciation, such as arctic primroses (Box 8),are accelerating our understanding of theserelationships. A bioGENESIS working groupfocused on these issues will draw togetherthese disciplines to develop a coherentgeneral model of diversification, whichcould then be applied, for instance, tounderstanding the dynamics underlying theexistence of biodiversity hotspots.
BOX 7. Rapid radiation of cichlid fishes in East African lakes
One of the most intriguing questions in evolutionary biology is how biologicaldiversity has been generated, especially when this appears to have taken place veryrapidly. The Great East African Lakes (Victoria, Malawi, and Tanganyika) havewitnessed the evolution of nearly 2,000 species of cichlids in the last 1-10 millionyears. In Lake Victoria, most of the species appear to have arisen in only15,000 years, before which the lake bed was almost entirely dry. This spectacularburst of recently diverged but phenotypically diverse species offers a specialopportunity to studying the underlying generative mechanisms.
Diversification in this system is thought to be the result of both natural and sexualselection. Natural selection has acted through adaptation to habitat (e.g., rocks,sand) and diet (e.g., snails, fish, zooplankton). Sexual selection has acted throughdivergence in male colouration and behaviour, coupled with female preferences forthose colours and behaviours. These drivers of speciation may be acting in aparticular sequence, starting with divergence in habitat use, then feedingmorphology, and then colour (Kocher 2004).
Diversification in African cichlids is now being impacted by human activities. Inaddition to extinction caused by introduced predatory fishes, such as the Nile perch,eutrophication has reduced water quality and thereby degraded the sexual signalsthat influence species recognition (Seehausen et al. 1997). Jointly these processeshave greatly reduced the diversity of cichlid species in Lake Victoria. These findingscall for more attention to the nature of species differences and how these willrespond to human influences.
References
Kocher TD. 2004. Adaptive evolution and explosive speciation: the cichlid fish model. Nature Review Genetics.5:288-298
Seehausen O, van Alphen JJM and Witte F. 1997. Cichlid fish diversitythreatened by eutrophication that curbs sexual selection. Science277:1808-1811
FIG. 7.1. Diversity of cichlids: radiation of trophic morphologiesand male colour patterns (Kocher, 2004), ©L Ruber.

20
BOX 8. Climate change, habitat fragmentation, and hybrid speciation inprimroses
A relatively large proportion of plant species, especially in the arctic, are polyploids,i.e., their chromosome numbers are multiples of the number in related diploidspecies. One proposed explanation for this observed distribution of ploidy levels isthe so-called secondary contact model (Stebbins 1985). According to this model,climatic and geological changes that cause the fragmentation and subsequentsecondary contact between differentiated populations favour the origin of newspecies through hybridization, followed by chromosome doubling. Successive cyclesof climate change, habitat fragmentation, and secondary contact can drive theevolution of species to progressively higher ploidy levels (Fig. 8.1).
In a group of arctic primroses (Primula sect. Aleuritia), polyploid, self-fertilisingspecies tend to occur at higher latitudes than their diploid, outcrossing relatives.Evidence from phylogenetic trees generated from maternally – and biparentally –inherited genes clarified the relationships between diploid progenitors and polyploidderivatives (Figure 8.1), suggesting that speciation via secondary contact provides aplausible scenario for correlated changes in ploidy level and reproductive strategies.During the Pleistocene, glacial advancesfragmented the ranges of the diploidspecies, while glacial retreats allowedranges to come into contact again,promoting the origin of polyploid taxathrough hybridization. The scarcity ofsuitable pollinators at higher latitudesmight explain the higher re-colonisationsuccess of the polyploid, selfing speciesin comparison with their diploid,outcrossing relatives.
References
Guggisberg A, Mansion G, Kelso S and Conti E.2006. Evolution of biogeographic patterns, ploidylevels, and breeding systems in a diploid-polyploidspecies complex of Primula. New Phytologist171(3):617-632
Stebbins GL. 1985. Polyploidy, hybridization, andthe invasion of new habitats. Annals of the MissouriBotanical Garden 72:824-832
P. nutans P. mistassinicaP. egaliksensis
2n = 2x = 18 2n = 4x = 40 2n = 2x = 22
P. egaliksensis
P. mistassinicaP. nutans
P. egaliksensis
chloroplast DNAphylogeny
nuclear DNAphylogeny
FIG. 8.1. Phylogenetic trees show that thediploid, outcrossing Primula mistassinica andP. nutans gave rise to the tetraploid, selfingP. egaliksensis. The geographic distribution ofP. egaliksensis overlaps with the ranges of thetwo diploid parents, ©E Conti.

21
TASK 2.2 The evolutionary historyof biotic assembly
Generations of biologists have focused ondescribing and understanding thecomposition and function of bioticcommunities. Ecological assemblages, atwhatever spatial scale, have histories – theywere pieced together through time, by themovement of species into and out of thesystem, or by in situ speciation andextinction events. Interactions amongspecies and ecological functions havechanged throughout this assembly process.The species in a local assemblage may beclustered phylogenetically (i.e., closelyrelated) if habitat filtering was thedominant process shaping communitymemberships, or phylogenetically widelydistributed or overdispersed (i.e., lessclosely related) if competitive exclusion ofecologically similar close relatives was thepredominant factor (Webb et al. 2002;Box S6). Likewise, spatial patterns ofbiodiversity have changed over time(Box 9), and our understanding of thesepatterns would benefit from historicalanalysis. At present we possess adequatespatial data for relatively few areas andgroups of organisms. Much more detailedinformation is necessary to identify hotspots of diversity and to explain howspecies came together to form theecosystems we now know. The aim ofTask 2.2 is to integrate the historical andspatial analysis of biotic assemblages acrossa variety of scales, from local assemblagesto ecosystems and regional species pools.
The species now present in any assemblagecame to occupy the same area – and tointeract with one another – through aparticular sequence of historical events.Some of the species in the system have
been interacting (and potentiallycoevolving) for longer, and some for muchshorter, periods of time. At times in thehistory of life there have been episodesduring which species have come newly, andquite rapidly, into contact (e.g., Vermeij1991). Examples include the great bioticinterchange between North and SouthAmerica (Marshall et al. 1982), and thewholesale effects on species ranges ofglacial cycles during the Pleistocene (Roy etal. 1996; see also Box 8). Human activitieshave recently brought species together atan unprecedented rate through theintroduction of exotic species, some ofwhich having become aggressive invasives.
Some progress has been made in inferringsequences of community assembly andpatterns of species movements through theintegration of paleontological,phylogenetic, and biogeographic methods(Box 9). These same approaches can revealhow environmental changes have shapedcurrent genetic diversity. However, muchremains to be done to comprehend theassembly of particular biotas and to extractany general assembly rules. This requiresthe development of a conceptual andmethodological framework for bringingtogether knowledge of the fossil record,phylogeny, biogeography, and lineagedivergence times, along with geographicand climatological information related toorganismal function (e.g., physiology anddispersal ability).
Evolutionary analyses may also help toexplain spatial patterns of diversity, forexample, the widely cited latitudinaldiversity gradient (Box 10). Criticalelements in elucidating the establishmentof such gradients include the inference ofancestral and descendant areas of
distribution, the timing of lineagediversification and changes in rates ofspeciation and extinction, the extent towhich lineages track their ancestralecological niches through time and space(“niche conservatism”), and the effects ofQuaternary climate change on localextinction and colonization. Bringing theseelements together will result in a far richerunderstanding of the origin of biodiversitygradients.
A bioGENESIS working group will beorganised to foster collaboration amongdifferent scientific communities, includingpaleobiologists, phylogeneticists,ecologists, climatologists, and geologists.These connections will enableinvestigations of such questions as: In anygiven ecosystem, how many of the speciesoriginated in situ, versus having originatedelsewhere and then filtered into the systemat a later date? For those that moved intothe system, are there any patternsinvolving source regions and times ofimmigration? Are there particular functionaltraits that originated within the system, orpreferentially filtered into the systemthrough species movement? Can we identifycharacter shifts that were brought about bythe entry of species into the system andsubsequent interactions in a new setting?How does the phylogenetic relatedness ofspecies within an ecosystem relate to thefunctioning of the system? Focus 2.2 willserve to explore and promote theintegration of historical and ecologicalknowledge relevant to such questions aboutbiotic assembly.
Greater knowledge of the effects of pastclimatic cycles on species origins can alsoinform the prediction of future evolutionarydynamics. How will climate change affect

22
habitat fragmentation and therefore,hybridisation between species in the future(cf. Box 8)? Can we predict howtemperature increases will change bioticdistributions and opportunities forsecondary contact among differentiatedpopulations? By fostering the integrationof phylogenetic tools with ecological nichemodelling, bioGENESIS activities willimprove our ability to make informedchoices for the conservation of areas wheredifferent species and biotic communitiescome in contact, under current and futureclimate projections (see also Focus 3).
Research Objectives and Activities
• Promote the integration of analyses ofbiome and community-level assemblyprocesses, bringing togetherphylogenetic, biogeographic, andecological approaches.
• Apply new analytical approaches in thecontext of selected study systems; e.g.,tropical forests of Southeast Asia inconjunction with CTFS and DIWPA;tropical montane diversity inconjunction with NESCent workinggroups; and Mediterranean ecosystems.
BOX 9. The assembly of regional biotas
Understanding how diversity evolves within local, regional, and continental biotas,and thus contributes to diversity within assemblages at these different scales,requires analyses of speciation among closely related groups, insights into how Earthhistory may have driven large-scale shifts in species distributions, and informationon the effects of climate change on the expansion, contraction, and shuffling ofspecies distributions over time. Examples of highly diverse biotas that might bestudied from these perspectives include the rainforests of the Amazon Basin ornortheastern Australia, the deserts of Australia, the savannah/grasslands biotas ofeastern and southern Africa, and the biotas of high montane areas (e.g., Penningtonet al. 2006).
It has been proposed, for example, that lowland forest populations of SouthAmerican birds and other organisms were passively transported to high altitudes bythe rapid uplift of the Andes, thus contributing to the montane ecosystems we seethere today. Many of these montane species assemblages were later subjected torepeated bouts of allopatricand sympatric speciation asaltitudinal vegetation zoneswere raised and lowered as afunction of cyclical climatechange during Plio-Pleistocenetimes (Ribas et al. 2007).
References
Pennington RT, Richardson JE andLavin M. 2006. Insights into thehistorical construction of species-richbiomes from dated plant phylogenies,neutral ecological theory andphylogenetic community structure. NewPhytologist 172:605-616
Ribas CC, Moyle RG, Miyaki CY andCracraft J. 2007. The assembly ofmontane biotas: linking Andeantectonics and climatic oscillations toindependent regimes of diversificationin Pionus parrots. Proceedings of theRoyal Society of London B 274:2399-2408
mindo
ensis
12
00-3
000m
cora
llinu
s
12
00-3
000m
satu
ratu
s
20
0-24
00m
pons
i
10
0-22
00m
sord
idus
10
0-18
00m?
ante
lius
10
0-18
00m?
lace
rus
0-
1000
m
siy
0-10
00m
melan
obleph
arus
0-
1000
m
maximiliani
fuscus
0-
1000
m
mindoensis
corallinus
saturatusponsi
sordidusantelius
lacerus
siy
melanoblepharus
fuscus
max
imili
ani
0-
1000
m
fuscus
sordidus species-group maximiliani species-group
FIG. 9.1. Distributional and altitudinal ranges of one lineage of South American parrots in thegenus Pionus (from Ribas et al. 2007). One lineage, the maximiliani species-group, is lowland,whereas its sister-lineage, the sordidus species-group, is high montane. Species in the Andes breedat high elevations but extend down to lower elevations during the winter in search of food. Thesister-lineage of all of these species (not shown) also has lowland-high montane sister-clades. Moredistant relatives are lowland forest taxa, thus showing that montane lineages have beenindependently derived from lowland ancestors.

23
BOX 10. Evolutionary factors shaping species diversity gradients
The differential distribution of species richness across geographical regions andbiomes has been documented for many different types of organisms, both on landand in the marine realm. Recent studies have integrated evolutionary and ecologicalprocesses to explain the high concentration of species in tropical regions and thecorresponding latitudinal diversity gradient.
The “tropical niche conservatism” hypothesis (reviewed in Wiens and Donoghue 2004)explains higher tropical diversity as a function of the longer occupation of tropicalareas by many lineages, with expansion into temperate areas limited mostly byfreezing temperatures. In support of this hypothesis, a study of hylid frogs in theAmericas documented a significant correlation between species richness and time ofcolonisation of an area, and identified temperature seasonality as the most importantfactor limiting the spread of four hylid lineages (Wiens et al. 2006).
Similarly, the “out of the tropics” model of Jablonski et al. (2006) postulates higherrates of taxonomic origination in tropical waters, and subsequent expansion intoextratropical regions. The spatial patterns of first occurrences and changes inlatitudinal distribution of marine bivalves, documented from the Neogene fossilrecord, support this model by finding higher speciationand lower extinction rates in the tropics, and greaterimmigration into extratropical regions.
It is likely that many different factors play a role inestablishing such diversity gradients, including therelative ease or difficulty of adapting to new climaticregimes and the availability of corridors for movement(Donoghue 2008).
References
Donoghue MJ. 2008. A phylogenetic perspective on the distribution ofplant diversity. Proceedings of the National Academy of Sciences USA 105Supplement 1:11549-11555
Jablonski D, Roy K and Valentine JW. 2006. Out of the tropics:evolutionary dynamics of the latitudinal diversity gradient. Science314:102-106
Wiens JJ and Donoghue MJ. 2004. Historical biogeography, ecology, andspecies richness. Trends in Ecology and Evolution 19:639-644
Wiens JJ, Graham CH, Moen DS, Smith SA and Reeder TW. 2006.Evolutionary and ecological causes of the latitudinal diversity gradient inhylid frogs: Treefrog trees unearth the roots of high tropical diversity.American Naturalist 168(5):579-596
3
7
11
14
16
10
7
4
10
7
4
16
14
11
7
3
FIG. 10.1. Left: Exerodontamelanomma (Hylidae), a treefrogfrom Oaxaca, Mexico, ©U García;right: Agropecten circularis(Pectinidae), a marine bivalvefrom the Pliocene InfiernoFormation, Baja California,Mexico, ©F Sour; bottom: origination and initialdiversification in the tropics(shown on the right) as anexplanation for the latitudinalspecies diversity gradient(represented on the left) (fromWiens and Donoghue 2004).

24
TASK 2.3 The evolution of functionaltraits
Phylogenetic analyses frequently reveal therecurrence of form in evolution, includingparallelisms, convergence, and theindependent loss of traits. These cases canbe examined for historical correlations withother parameters. This may, for example,help us to understand whether a particulartrait evolved as an adaptation followingexposure to certain environmentalcircumstances, or, rather, the evolution ofthe trait under different circumstancesallowed the subsequent occupation of anew environment (Box 11, S7). Suchstudies provide powerful insights into theecological functions of traits as well as intothe forces that shape community assembly(see Task 2.2).
It is clear that many important ecologicalfunctions have evolved independently inseparate lineages, sometimes apparentlytightly clustered in phylogeny, andsometimes arising very distantly in the treeof life. This raises an important, thoughstill largely unaddressed question, namely,the extent to which independently evolvedtraits really do functionsimilarly enough thatthey can be treated assingle categories inmodelling ecosystem-level processes (Box 12).
That is, to what extent are independentlyevolved traits really functionally redundant?Alternatively, to what extent shouldindependently evolved traits be modelled aspossibly responding quite differently torelevant environmental changes?
There are already some importantindications that independently evolvedfunctions might perform differently enoughto be misleading in some circumstances. Forexample, species from several grass lineagesthat appear to have evolved C4photosynthesis independently have shownsignificantly different growth responses toelevated CO2 levels. Likewise, levels of theenzyme carbonic anhydrase have beenshown to differ significantly among C4grasses representing independently evolvedlineages. Levels of carbonic anhydrase arecritical in interpreting oxygen isotopefractionation and, in turn, in calculatingglobal primary productivity and identifyingpotential carbon sinks (Box 12).
A bioGENESIS working group focused onthese issues will draw phylogeneticists andfunctional biologists together to provide acritical assessment of the validity of using
BOX 11. Drivers of morphologicalevolution in Bignoniaceae
Analyses of the relative timing of evolutionarychanges in organismal traits can test specifichypotheses on the drivers of morphologicalevolution and shed light on the ecologicalfunction of traits. For example, ecologicalchanges that occur after the occupation of anew habitat suggest adaptation to the novelconditions (Larson and Losos 1996).Alternatively, morphological changes that occurbefore the occupation of a particular habitatsuggest that the new morphology may haveserved as a “key innovation” allowingoccupation of the novel conditions.
In the plant clade Bignonieae (Bignoniaceae)the evolution of several distinctivemorphological characters is directly linked tocertain habitat transitions. Specifically, repeatedtransitions from the liana growth form to theshrub habit are strongly correlated with thecolonization of dry open habitats, together withthe loss of tendrils, and the evolution ofrounded seed wings. In general, these featuresappear to have evolved after the inferredhabitat transitions, suggesting that the derivedmorphological traits are indeed adaptations tothe new habitat (Lohmann et al. in prep.).
References
Larson A and Losos JB. 1996.Phylogenetic systematics ofadaptation. In: Rose MR andLauder GV (Eds). Adaptation,Academic Press
Lohmann LG, Bell C andWinkworth RC. Environmentalchange as a key driver ofmorphological diversificationin Bignonieae (Bignoniaceae).In preparation.
FIG. 11.1. Sample of the morphological diversity in Bignonieae, ©L G Lohmann.

25
BOX 12. Taking trait evolution into account in modelling global climatechange
Global ecological models, such as those that enable predictions of climate change,have accommodated variation in the traits of organisms mainly by reference to majorfunctional classes, such as “trees” or “C4 grasses.” These models can be refined bytaking into account phylogeny and evolutionary changes in the traits of interest.
For example, grasses in which C4 photosynthesis originated independently mayrespond differently to elevated levels of CO2 (Kellogg et al. 1999). Likewise, plantsfrom separate C4 grass lineages show different levels of the key enzyme carbonicanhydrase, and estimates of primary productivity that assume uniformly low levels inC4 grasses (e.g., Gillon and Yakir 2001) may be biased. Furthermore, somewidespread and locally dominant C3 grasses, such as Phragmites australis, producethe low levels of carbonic anhydrase found in their C4 relatives (Edwards et al.2007). Knowledge of carbonic anhydrase levels is critical in interpreting oxygenisotope fractionation and, inturn, in inferring globalprimary productivity. A moreaccurate estimate of theglobal uptake of atmosphericCO2 could be obtained bytaking into account thephylogenetic distribution ofthis key enzyme.
References
Edwards EJ, Still CJ and DonoghueMJ. 2007. The relevance of phylogenyto studies of global change. Trends inEcology and Evolution 22(5):243-249
Gillon J and Yakir D. 2001. Influenceof carbonic anhydrase activity interrestrial vegetation on the O-18content of atmospheric CO2. Science291:2584-2487
Kellogg EA, Farnsworth EJ, Russo ETand Bazzaz FA. 1999. Growthresponses of C4 grasses ofcontrasting origin to elevated CO2.Annals of Botany 84:279-288
Conifers
Angiosperms
Juniperus excelsaMetasequola glyptostroboides
Sequoia sempervirensPodocarpus falcatusAraucaria excelsia
Monocots Arum sp.
Crocosmia sp.Ruscus sp.
Asparagus sp.Commelina communis
Typha sp.Cyperus sp.2Cyperus sp.1
Poaceae Eleusine indica
Cynodon dactylonPhragmites australisDigitaria sanguinalis
Setaria vulgarisEchinochloa crus-galli
Sorghum bicolorSorghum halepense
Zea maysBambusa sp.Stipa capensis
Bromus sp.Lolium rigidumPolypogon sp.
Apera sp.
Eudicots Platanus orientalisAmaranthus sp.1Amaranthus sp.2Amaranthus sp.3Malus orientalis
Prunus divaricataCeltis tournefortii
Glycine soyaQuercus robur
Quercus pedunculifloraQuercus bossieriiMercurialis sp.
Pistacia atlanticaPistacia lentiscusDals cotinifolia
Hibiscus sp.Eucalypus woodwardii
Liquidambar styracifluaSonchus sp.
Nerium oleanderNicotiana tabacumLycium barbarum
Paulownia tomentosa
PACCAD grass clade
BEP grass clade
Caryophyllales
Rosids
Asterids
(a) (b)
H eq
0.01 - 0.1090.109 - 0.2080.208 - 0.3070.307 - 0.4060.406 - 0.5050.505 - 0.6040.604 - 0.7030.703 - 0.8020.802 - 0.9010.901 - 1.01.0
ConiferTree/shrubC3 herbC4 herbC3 monocotC3 grassC4 monocot
Plantfunctional type
TRENDS in Ecology & Evolution
FIG. 12.1. Evolutionary shifts in grasses between C3 and C4 functional groups (shown on the right) arenot significantly correlated with evolutionary shifts (shown on the left) in the level of the enzymecarbonic anhydrase (measured by eq) (from Edwards et al. 2007). Note that Echinocloa crus-galli,which represents a separate origin of C4 photosynthesis, has higher values than other C4 grasses. Also,the C3 grass Phragmites australis has low values similar to its C4 relatives.
H
independently evolved functional groups inmodelling responses to environmentalchange or in comparing similar ecosystemsassembled independently in differentregions. Here we see immediateopportunities for collaborations involvingthe ecoSERVICES and bioSUSTAINABILITYCore Projects within DIVERSITAS. A cross-project working group has already beenformed to address these issues through theCentre for Population Biology at SilwoodPark (Imperial College, UK). This effort isfocused on how ecosystem functions relateto the evolution of underlying organismaltraits, and how the phylogeneticdistribution of the key functions themselvesmight bear on the assessment of risks toecosystem services in the face ofenvironmental changes. The focus here on“feature diversity” also bears directly onthe development of appropriate measures ofbiodiversity within bioSUSTAINABILITY CoreProject (see also Focus 3).
Research Objectives and Activities
• Develop analyses to assess the possibleimpacts of the convergent evolution oftraits on their ecological function; e.g.,using global plant trait databases.
• Stimulate analyses of the evolution ofkey enzymes and metabolic pathwaysrelated to ecosystem function, especiallybridging genomics and ecology inmicroorganisms.
• Develop new approaches to assessingrisks to ecosystem services that directlyincorporate the phylogenetic distributionof ecological functions.

26
TASK 2.4 Rapid evolution and eco-evolutionary dynamics
Charles Darwin famously believed thatevolution required “the long lapse of ages.”This perspective continued to prevail formore than a century, at which timeevidence began to emerge that evolutioncan actually occur rapidly (Palumbi 2002).Darwin's own Galapagos finches illustratethis point: the beak size of the mediumground finch, Geospiza fortis, evolvedrecently in response to changing seedavailability (Grant and Grant 2006; Box S8).This is a good example of the action of themain driver of evolution, natural selection –individuals with genes that improve survival(or fecundity) produce more offspring andthereby increase the frequency of thosegenes into the future. Rapid evolutionarychange can also be driven by sexualselection, which increases the frequency ofgenes that influence mating success. Anexample comes from male Trinidadianguppies (Poecilia reticulata) that evolvehigh colour in low-predation environments– females prefer to mate with colourfulmales and predation no longer selectsagainst high colour (Endler 1980; Magurran2005). Finally, evolutionary change can bedriven by random effects, for example,genetic drift. If population sizes are smallenough, these changes can be rapid, suchas when relatively few individuals withparticular traits colonise a new location.
Speciation – the evolutionary divergence ofone species into two – can also be rapid,sometimes driven by the mechanismsdescribed above. For example, Africancichlid fishes in Lake Victoria havediversified through natural selection (diet)and sexual selection (mate choice) from afew species into hundreds in less than
15,000 years (Box 7). On even shorter timescales, many species of insects have, overless than 100 years, formed new “hostraces” adapted to introduced plants andhave become reproductively isolated fromtheir ancestors on native plants (seeTask 3.1; Box S9). Other geneticmechanisms, such as polyploidyzation, cancause very rapid speciation (see Task 2.1;Box 8).
It has now been firmly established thatsome species show appreciable evolutionarychange on very short time scales. Thesechanges presumably influence the successof those organisms – but this has beenlittle studied except in parasites andpathogens. HIV population size, forexample, decreases after anti-viral drugs aregiven to a patient, but then increases againafter the evolution of resistance to thosedrugs (see Task 3.3; Box 19). Similardynamics have been observed in rabbits(Oryctolagus cuniculus) introduced intoAustralia and the myxoma virussubsequently introduced to control them(Dwyer et al. 1990). These results suggestthe need to add evolution into models ofpopulation dynamics, so as to better predictpathogen outbreaks, species invasion, andpopulation declines (see Task 3.3).
Rapid evolution of one species also mayhave consequences for other organisms, forcommunity structure, and perhaps even forecosystem function. One major researcheffort in this area falls under the umbrellaof “community genetics.” Recent findingsindicate, for example, that plant genesaccount for a significant proportion ofvariation in water cycles, nutrient cycles,below-ground carbon storage, and theabove-ground diversity of herbivores
(Whitham et al. 2008; Box 13). Theseresults clearly demonstrate that standinggenetic variation matters to ecologicalprocess. More recent work is concentratingon whether rapid evolutionary changes inthis genetic variation have significantecological consequences. A US NSF-FIBRproject is examining whether the rapidevolution of guppy populations in Trinidad,referred to above, is influencing aquaticcommunities and nutrient cycling(Palkovacs et al. in press).
It is critical to note that many rapidevolutionary changes are associated withhuman impacts. Fish, mammal, and plantpopulations subjected to human harvestinghave rapidly evolved smaller sizes andearlier reproduction (Box 14). Additionalexamples are presented below under Focus3 of rapid evolution (and even speciation)in response to climate change andintroduced species. Collectively, these casesillustrate the critical role that rapidevolution can play in the response ofpopulations to human-causedenvironmental change, and the need totake evolution into account in modellingresponses to such changes. A bioGENESISworking group focused on these issues willpromote the integration of evolutionaryapproaches in understanding ecologicaldynamics, and provide clear links to relatedDIVERSITAS efforts.
Research Objectives and Activities
• Stimulate theoretical predictions andempirical testing of rapid evolution andits ecological consequences.
• Promote applications of evolutionaryapproaches to elucidate the dynamicrelationship between genetic variationand aspects of ecosystem function.

27
BOX 13. Genetic variation matters to ecological processes
The genetic properties of organisms have long been known to influence their survivaland reproductive success. More recently, it has been recognised that these propertiescan also influence community structure and ecosystem processes.
Recent work in the area of “community genetics” (Whitham et al. 2006, 2008) hasshown that cottonwood trees (Populus spp.) that are less genetically similar to oneanother house less similar arthropod communities, and that these communities are,to some extent, passed on from parent to offspring trees. Some community-levelconsequences of genetic variation are therefore heritable. Perhaps even more striking,selection on genetic variation has been shown to influence ecosystem function.Specifically, the amount of foliar condensed tannins in cottonwood trees has agenetic basis. This genetic variation affects nitrogen cycling in forests; increasedtannins in the leaf litter increases nitrogen mineralization and, therefore, treegrowth. In addition, beavers (Castor canadensis) avoid high-tannin trees, whichincreases the relative frequency of such trees in areas with beavers. In turn, thisleads to increased tannins in the surrounding water and soil, which changes theaquatic invertebrate community, influences decomposition rates, and feeds back ontree growth.
References
Bangert RK, Turek RJ, Rehill B, Allan GJ,Wimp GM, Schweitzer JA, Bailey JK,Martinsen GD, Keim P, Lindroth RL, andWhitham TG. 2006. A genetic similarity ruledetermines arthropod community structure.Molecular Ecology 15:1379-1391
Whitham TG, Bailey JK, Schweitzer JA,Shuster SM, Bangert RK, LeRoy CJ,Lonsdorf EV, Allan GJ, DiFazio SP, Potts BM,Fischer DG, Gehring CA, Lindroth RL,Marks JC, Hart SC, Wimp GM and Wooley SC.2006. A framework for community andecosystem genetics: from genes toecosystems. Nature Review Genetics 7:510-523
Whitham TG, DiFazio SP, Schweitzer JA,Shuster SM, Allan GA, Bailey JK andWoolbright SA. 2008. Extending genomics tonatural communities and ecosystems.Science 320:492-495
FIG. 13.1. Cottonwood trees,which are more geneticallydissimilar to one another housemore dissimilar communities ofarthropods (from Bangert et al.2006). This pattern was observedin trees planted in a “commongarden” and in wild populations.Beavers favor trees with particulargenotypes, which may influencecommunity and ecosystemproperties.
© M
B E
dlun
d

28
BOX 14. Harvesting can cause undesirable rapidevolutionary change
Humans often harvest animal and plant populations for commercialor recreational benefit. When harvest rates are high or select forparticular phenotypes, populations may crash or otherwise undergochange in ways that make them less useful to humans. Some ofthese unfortunate changes are the result of evolution.
Bighorn sheep (Ovis canadensis) on Ram Mountain, Alberta, providean example. These sheep are hunted as trophies, and hunters areonly allowed to take individuals with the largest horns. Thisgenerates strong natural selection against individuals whose hornsgrow particularly fast. A consequence is the rapid evolution ofsmaller horn size, which then reduces the valuefor hunters (Coltman et al. 2004). Anotherexample of undesirable evolutionary changecaused by harvesting is the evolutionarydecrease in size at maturity in Atlantic cod(Gadus morhua), which may have contributedto the collapse of this fishery (Olsen et al.2004).
References
Coltman DW, O'Donoghue P, Jorgenson JT, Hogg JT, Strobeck Cand Festa-Bianchet M. 2003. Undesirable evolutionaryconsequences of trophy hunting. Nature 426:655-658
Olsen EM, Heino M, Lilly GR, Morgan MJ, Brattey J, Ernande Band Dieckmann U. 2004. Maturation trends indicative of rapidevolution preceded the collapse of northern cod. Nature428:932-935
80
70
60
50
40
30
Mea
n ho
rn le
ngth
(cm
)
1970
1975
1980
1985
1990
1995
2000
2005
Year
FIG. 14.1. The horn size ofbighorn sheep rams has decreasedduring the last 30 years ofselective harvesting. The plotshows phenotypic change, butadditional work has shown thatthis has a genetic basis (fromColtman et al. 2003).
© A
Hend
ry

29
FOCUS 3. Evolution, biodiversity, andhuman well-being
The United Nations World Summit onSustainable Development (2002) identifieda set of key objectives: biodiversity andecosystem services must be maintained (orrestored), food security must be improved,and global population health must beenhanced. Moreover, the development ofrenewable energy is seen as crucial in viewof the adverse consequences of globalwarming on society. Further, security frombiological threats – from the spread ofdebilitating diseases to the possible use ofdeadly bioagents – is an increasingconcern. The bioGENESIS Core Project willbring an evolutionary perspective directlyto bear on these pressing issues, to identifynew uses for evolutionary knowledge inconserving biodiversity and promotinghuman well-being.
In general, research in evolutionary biologyis playing an increasingly important role inaddressing these key societal objectives.Advances across a number of evolutionarydisciplines (e.g., comparative genomics)provide a necessary foundation foraddressing numerous challenges. Forexample, the rapid identification ofemergent diseases now depends directly onmolecular phylogenetic methods, as doesthe identification of their bio geographicorigins (Box 6), the prediction of spread,and the design of appropriate responses(including vaccines). The same is true ofintroduced species more generally, whereeffective management of invasive organismsoften depends critically on knowledge ofevolutionary relationships and processes.
As noted above (Task 2.4), recent work oneco-evolutionary dynamics is providingprofound insights into how organisms willrespond to anthropogenic environmentalchanges. In fact, we now recognise thathuman impacts, including pollution, land-use change, climate change,agriculture/aquaculture, and introducedspecies, may be among the most importantdrivers of modern evolutionary change(Palumbi 2002). In each of these casesthere are a range of evolutionaryconsequences, from the erosion of geneticdiversity, to the evolution and spread ofdiseases, and even human-inducedspeciation. Analyses of such evolutionaryresponses are critical to the development ofa truly predictive biodiversity science thatcan provide a sound basis for ecosystemmanagement.
The emerging field of metagenomics – theuse of genomic tools to probeenvironmental samples (from a gram of soilor a drop of sea water) for microbialbiodiversity – relies directly on evolutionaryanalysis for the identification andcharacterization of what now appears to bea vast reservoir of uncharted biodiversity(Box 1). Predictions based on thephylogenetic relationships of theseunknown organisms provide the first step inunderstanding their possible ecosystem-level functions and global impacts.Phylogenetic studies also provide the basisfor sound “bioprospecting”, focusingattention on the close relatives oforganisms that produce medicinally usefulchemicals or that may be important in thedevelopment of sustainable energy. Recentstudies of endophytic fungi living inside oftropical plants, for example, have provideda glimpse of the enormous biological and
chemical diversity that remains to bediscovered and potentially harnessed forhuman purposes. Some of these fungiproduce entirely novel bioactivecompounds, and a few have even beenfound to produce octane and may one dayprovide the basis for new biofuels (Box 2).
While Focus 1 concerns the discovery anddocumentation of biodiversity, and Focus 2deals with the causes and consequences ofdiversification, Focus 3 addressesevolutionary management and conservation.DIVERSITAS, through bioGENESIS, can playa significant role in promoting these areasof investigation, by providing a focal pointfor identifying and tackling evolutionaryquestions of immediate relevance tomanaging biodiversity. bioGENESIS can helpto coordinate the development of anintegrated system for evolutionaryecosystem management aimed at mitigatingthe effects of climate change, combatingemerging diseases, and conservingthreatened species and their evolutionarypotential. Through its working groups andactivities, bioGENESIS will promote theapplication of new tools and theexploration of creative new ways to harnessevolutionary biology in the service ofbiodiversity and sustainability.
© J
Crac
raft

30
TASK 3.1 Evolutionary ecosystemmanagement
To prevent biodiversity changes undesirablefor our own existence, we need increasinglyto manage the protection of threatenedspecies, the eradication of invasive species,and the restoration of native ecosystems.Current practices in what has becomeknown as “ecosystem management”generally undervalue evolutionaryconsiderations. A bioGENESIS working groupin this area will promote new approaches tothe science of ecosystem managementbased on evolutionary biology.Retrospective approaches can help us tounderstand how our activities haveimpacted ecosystems during humanevolution, migration, and populationexpansion. Complementary prospectiveapproaches enable us to project howhumans are likely to affect ecosystemfunctions and how we might mitigateagainst deterioration.
Homo sapiens has continuously expandedits population since moving out of Africaaround 80,000 year ago (Hawks et al.2007). During the human journey, ourspecies has had profound impacts, locallyand globally, through the use of fire, the
development of agriculture, the processingof metals, the construction of cities, theexpansion of trade, and the development ofscience and technology. These activitieshave driven extraordinary biodiversitychange, including the evolution of weeds,pests, and pathogens, and the massextinction of wild species. Historicalanalyses of how biodiversity and ecosystemservices have been changed by humanactivities provide unique insights into theeffects of such changes on human well-being. The first aim of Task 3.1 is tofacilitate the integration of variousretrospective approaches to reconstructingthe history of biodiversity change in orderto develop better models to predict long-term trends.
The second aim of Task 3.1 is to provide ananalysis of the on-going evolutionaryresponses of both native and alien speciesto recent environmental perturbations. Oneimportant class of outcomes entails therapid evolution of invasive species whichresults in their even more rapid spread(e.g., the cane toad in Australia, Phillips etal. 2006; Box S10). In other cases,hybridization can result between native andintroduced species, possibly promoting
rapid hybrid speciation and cycles of furtherinvasion and ecosystem disturbance. Suchgene flow between natives and non-nativesmay be quite common in some groups oforganisms, and could have devastatingconsequences (e.g., Kim et al. 2008;Box S11). Also of concern are cases of therapid spread of aggressive genotypesthrough invasive populations, as appears tohave happened in the reed grass,Phragmites, in North America (Box 15). Insome cases, hybridization or selectivesweeps appear to be associated with majorchanges in breeding system, such as shiftsto self fertilization or asexual reproduction.Likewise, there may be significant impactson predator-prey, host-parasite, or host-pathogen dynamics and evolution with theintroduction of new species into a system(e.g., Schwarz et al. 2007; Box S9).
To achieve the aims of Task 3.1, bioGENESISwill bring together relevant evolutionarybiologists around these issues, especiallywhere clear connections can be made toother DIVERSITAS Core Projects and Cross-cutting Networks. For example, there areobvious links to the maintenance ofagricultural biodiversity and food security,and we envision an activity focused on theevolutionary impacts of genetically modifiedorganisms (GMOs) on non-target species.
Research Objectives and Activities
• Facilitate studies on the history ofbiodiversity change as a function ofhuman evolution, migration, andpopulation expansion.
• Coordinate a synthetic analysis of theimpacts of introduced species on rapidevolution, hybridisation, and speciationinvolving native species.
• Promote an analysis of the potentialevolutionary impacts of GMOs on non-target species.
© J
Crac
raft

31
BOX 15. Invasion of the reed grass, Phragmitesaustralis: genetic analysis of a “sleeper weed”
Invasive species pose a major threat to the maintenance ofnative biodiversity, and in some cases have enormous ecologicaland economic consequences. Therefore, it is important tounderstand why and how they spread. Evolutionary analyses areincreasingly being applied to this problem.
The rapid spread of the common reed grass, Phragmitesaustralis, in northeastern North America (and beyond), providesan example of the application of molecular genetic techniquesin tracing the history of invasion. Until the beginning of the1900's a native form of this species remained limited indistribution in North America. Since then, P. australis has spreadrapidly, and expanded into many freshwater habitats, chokingout the native vegetation. Analyses of several chloroplastgenetic markers, both from living plants andfrom herbarium specimens going back over100 years, demonstrated that a non-nativestrain of Phragmites moved into NorthAmerica from Eurasia and has rapidlyreplaced the native populations in manyareas and greatly expanded the range of thespecies (Saltonstall 2002). The spread of theexotic genotype also has been documented inQuebec, Canada, where its movement inlandappears to have been promoted by thedevelopment of the highway systembeginning in the 1960's (Lelong et al. 2007).The introduction of a new, more aggressivegenotype appears to explain the suddenawakening of this “sleeper weed.”
References
Lelong BL, C., Jodoin Y and Belzile F. 2007. Expansionpathways of the exotic common reed (Phragmitesaustralis): A historical and genetic analysis. Diversity andDistributions 13:430-437
Saltonstall K. 2002. Cryptic invasion of a non-nativegenotype of the common reed, Phragmites australis, intoNorth America. Proceedings of the National Academy ofScience USA 99:2445-2449
Before 1900 1901 - 1920
1921 - 1940 1941 - 1960
After 1960
FIG. 15.1. Changes in the geographicdistribution of Phragmites australis genotypesover 20 year time intervals in Connecticut,Massachusetts, and Rhode Island, USA (fromSaltonstall 2002). Green and red trianglesrepresent the native genotype and invasivehaplotype, respectively.
© M
Don
oghu
e

32
BOX 16. Rapid evolution of native species in response to invaders
The invasion of a species into a new geographicallocation should impose strong natural selection on theinvader and on the native species with which itinteracts. This might then result in rapid evolutionarychange. An example of this phenomenon comes fromsoapberry bugs – insects that use a “beak” to piercethe fruits of certain plants and then feed on the seedsinside. As larger fruits require longer beaks, and viceversa, evolutionary changes in beak length have takenopposite trajectories in different parts of the world,depending on the relative sizes of the fruits of thenative and introduced plants (Carroll et al. 2005).
In Florida, the native soapberry bug (Jaderahaematoloma) feeds on a native plant (balloon vine,Cardiospermum corindum) with a large fruit, but itevolved a shorter beak when it switched to anintroduced plant with smaller fruits. In Australia, thereverse occurred. There, the native soapberry bug(Leptocoris tagalicus) feeds on native plants (Alectryonspp.) with small fruits but evolved a longer beak whenit switched to a balloon vine (Cardiospermumgrandiflorum) introduced from the neotropics. Theballoon vine is a major pest in Australia and soapberrybugs are the only native species to have colonised it.It has been suggested that the evolution of nativesoapberry bugs in response to this plant may decreaseits spread, averting the need for biocontrol throughfurther species introductions.
References
Carroll SP, Loye JE, Dingle H, Mathieson M, Famula TR and Zalucki MP.2005. And the beak shall inherit – evolution in response to invasion.Ecology Letters 8:944-951
FIG. 16.2. The left-hand part of the plot (circles) shows the smallerbeak length of museum specimens before 1925-1964 vs. after 1964-2004 when balloon vine became common. The right-hand part of thefigure also shows that soapberry bugs in Australia are diverging intotwo forms – those adapted to the native plant (diamonds) and thoseadapting to the introduced balloon vine (triangles) (from Carroll et al.2005).
Beak
leng
th (
mm
)
(mm
)
1925 -34 35 -44 45 -54 55 - 64 65 - 74 75 -84 85 - 94 95- 2004
Decade
6
6.5
7
Males
6.5
8
7.5
7
6
Females
FIG. 16.1. The nativeAustralian soapberry bugon its native host (left)and on the invasiveballoon vine (right). Thelarger fruits of theballoon vine have causedthe evolution of a longerbeak (seen being insertedinto the fruit capsule) inthese bugs, ©S P Carroll.

33
TASK 3.2 Evolution and climaticchange
A rapidly expanding area of inquiry isfocused on predicting the impacts ofclimate change on natural ecosystems, agroecosystems, and water supplies. Thedramatic increase in temperature over thepast few decades has been having seriousimpacts on biodiversity, including localextinction, range shifts, and adaptation torising temperatures. Phenological shifts(leading to earlier reproduction)corresponding to increasing springtemperatures have now been documented(Box 17), although the genetic bases forsuch shifts remain uncertain in most cases.In some cases, shifts in timing haveresulted in mismatches with pollinators orwith the abundance of food supplies, as inthe decline of the pied flycatcher in Europe(Both et al. 2006).
It is expected that phylogenetic constraintsand evolutionary potential will influenceresponses of various organisms to climaticchanges (Reusch and Wood 2007). Forexample, a recent analysis of the florasurrounding Henry David Thoreau's WaldenPond in Concord, Massachusetts, USA,demonstrated that members of specificangiosperm lineages that had failed to shifttheir flowering times in response totemperature change are now greatlydecreasing in abundance (Box 18).
Here again, insights can be gained fromboth retrospective and prospectiveanalyses. Retrospective approaches,integrating geological data on climaticchange during the Pleistocene and thePliocene, and even earlier in the Tertiary,with knowledge of phylogeny, help us tounderstand the relative importance of
migration/niche conservatism and adaptiveevolution to climate change, and to testthe validity of climate envelop models inpredicting future changes. New insights arenow being provided by modellingenvironmental factors associated withpresent geographical distributions usingdata obtained from specimens in naturalhistory collections and reconstructing pastdistributions and the evolution of nicheparameters using phylogenetic trees. Suchstudies permit broader comparisons amongclades and can be scaled-up to makepredictions about ecosystem compositionand function in the face of climate change.Finally, the phylogenetic distribution ofspecies at risk of extinction due to climatechange can be used to predict themaintenance of diversity (Yesson andCulham 2006; Box S12).
At the same time, prospective approachesbased on the principles and methods ofpopulation biology are critical in predictingthe demographic and genetic effects ofclimate change and how populations are
likely to respond. Such approaches areparticularly important in predicting how theboundaries between different vegetationtypes are likely to shift under climaticchange. At present, however, most evidenceof adaptive response to climatic change isindirect and it remains uncertain how largethe genetic variance is in wild populationsfor adaptation to climatic changes. The aimof Task 3.2 is to stimulate productivecollaborations between evolutionarybiologists and archaeologists, geologists,and ecologists focused on climate change.Here we envision close ties with elementswithin the bioDISCOVERY Core Project,especially regarding the analysis ofgeographical ranges and climate nichemodelling.
Research Objectives and Activities
• Promote the integration of phylogenyand abiotic niche reconstructions intoclimatic change research to test andimprove climate envelope models.
• Facilitate analyses of short-termevolutionary responses to climatechange.
• Coordinate collaborative research onvegetation boundary shifts incorporatingthe evolutionary responses ofpopulations.
© A Hendry

34
BOX 17. Evolutionary responses to climate change
One of the major global changes over the past few decades has been the dramaticincrease in temperature, and this change is expected to accelerate in the comingdecades. Since organisms are often adapted to local temperatures, populations willneed to either move to new locations or evolve rapidly to remain adapted. Suchadaptations are expected to be particularly likely in relation to the timing ofreproduction.
Climate warming is expected to be especially acute at high latitudes. In the Yukon,for example, the average April-June temperature has increased by 2º C over the last26 years, and the production of spruce cones on which the red squirrels (Tamiasciurushudsonicus) depend has come correspondingly earlier. The squirrels have, in response,advanced the date at which they give birth to young (parturition) by almost 26 days.This advance has been accomplished by acombination of phenotypic plasticity and geneticchange (Réale et al. 2003). Many other examplesof shifts in timing are known, but the relativecontributions of plastic versus genetic responsesare less certain. Another case where the responseis known to be genetic is the shift in photoperiodin pitcher plant mosquitoes, Wyeomyia smithii(Bradshaw and Holzafel 2001).
References
Bradshaw W and Holzapfel C. 2001. Genetic shift inphotoperiodic response correlated with global warming.Proceedings of the National Academy of Sciences USA 98:14509-14511
Réale D, McAdam AG, Boutin S and Berteaux D. 2003. Geneticand plastic responses of a northern mammal to climate change.Proceedings of the Royal Society of London B 270(1515):591-596
135
130
125
120
115
110
Mea
n lif
etim
e pa
rtur
itio
n da
te
1989
1990
1991
1992
1993
1994
1995
1996
Year of birth
1997
1998
FIG. 17.1. Red squirrels in theYukon that were born in 1998reproduced (parturition) almost 20days earlier (averaged over theirentire life) than did those born in1989. This plot shows phenotypicchange, but additional work hasestablished that this phenotypicchange has a genetic basis (fromRéale et al. 2003).
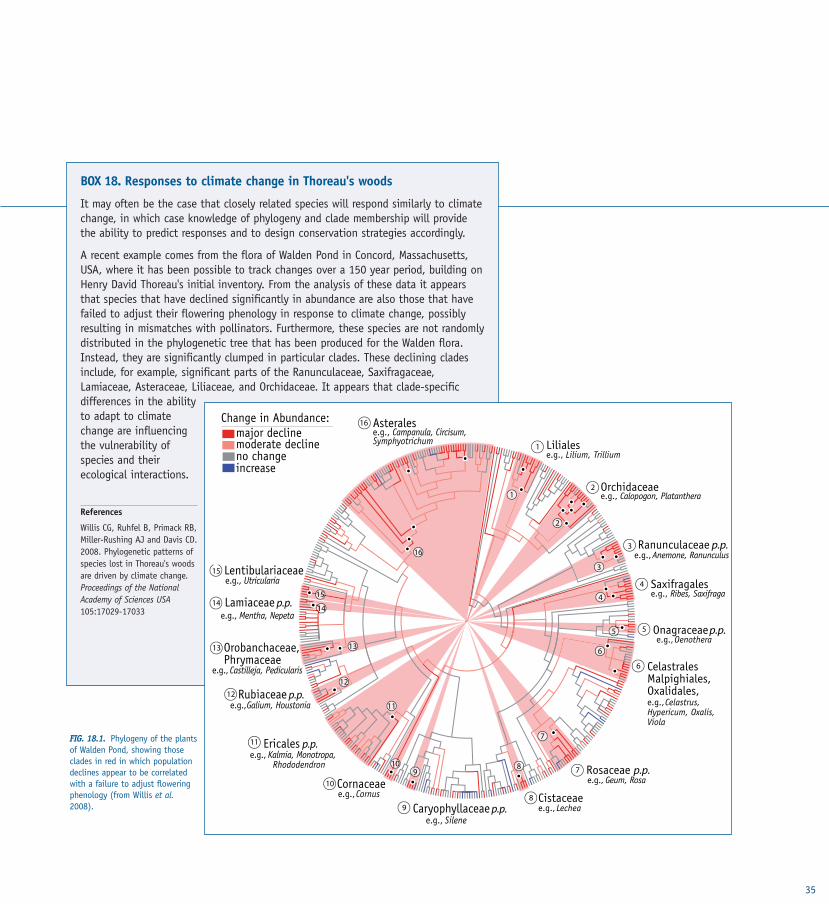
35
BOX 18. Responses to climate change in Thoreau's woods
It may often be the case that closely related species will respond similarly to climatechange, in which case knowledge of phylogeny and clade membership will providethe ability to predict responses and to design conservation strategies accordingly.
A recent example comes from the flora of Walden Pond in Concord, Massachusetts,USA, where it has been possible to track changes over a 150 year period, building onHenry David Thoreau's initial inventory. From the analysis of these data it appearsthat species that have declined significantly in abundance are also those that havefailed to adjust their flowering phenology in response to climate change, possiblyresulting in mismatches with pollinators. Furthermore, these species are not randomlydistributed in the phylogenetic tree that has been produced for the Walden flora.Instead, they are significantly clumped in particular clades. These declining cladesinclude, for example, significant parts of the Ranunculaceae, Saxifragaceae,Lamiaceae, Asteraceae, Liliaceae, and Orchidaceae. It appears that clade-specificdifferences in the abilityto adapt to climatechange are influencingthe vulnerability ofspecies and theirecological interactions.
References
Willis CG, Ruhfel B, Primack RB,Miller-Rushing AJ and Davis CD.2008. Phylogenetic patterns ofspecies lost in Thoreau's woodsare driven by climate change.Proceedings of the NationalAcademy of Sciences USA105:17029-17033
Asteralese.g., Campanula, Circisum, Symphyotrichum
Lentibulariaceaee.g., Utricularia
Lamiaceae p.p.e.g., Mentha, Nepeta
Orobanchaceae,Phrymaceae
e.g., Castilleja, Pedicularis
Rubiaceae p.p.e.g., Galium, Houstonia
Ericales p.p.e.g., Kalmia, Monotropa,
Rhododendron
Cornaceaee.g., Cornus
Caryophyllaceae p.p.e.g., Silene
Cistaceaee.g., Lechea
Rosaceae p.p.e.g., Geum, Rosa
CelastralesMalpighiales,Oxalidales,e.g., Celastrus, Hypericum, Oxalis, Viola
Saxifragalese.g., Ribes, Saxifraga
Ranunculaceae p.p.e.g., Anemone, Ranunculus
Orchidaceaee.g., Calopogon, Platanthera
Lilialese.g., Lilium, Trillium
Onagraceae p.p.e.g., Oenothera
increaseno change
major decline moderate decline
Change in Abundance:
1
16
16
2
3
4
6
7
89
10
11
12
13
14
15
1
2
3
4
6
7
89
10
11
12
13
14
15
5 5
FIG. 18.1. Phylogeny of the plantsof Walden Pond, showing thoseclades in red in which populationdeclines appear to be correlatedwith a failure to adjust floweringphenology (from Willis et al.2008).

36
TASK 3.3 Combating disease
Of all biodiversity changes, the migrationand evolution of zoonotic pathogensperhaps most directly threatens human life.Because pathogens generally have veryshort generation times and much highermutation rates than their hosts, they canquickly adapt to environmental changes,including the antibodies in our immunesystems (Box 19). Evolutionary approachesare therefore absolutely necessary for theidentification, surveillance, and control ofemerging diseases (Box S13, S14, S15).
Many factors have contributed to theemergence of infectious diseases such asWest Nile fever and the avian flu, includingglobal human movements, expanded tradein various goods including wild animals,and anthropogenic environmental changes.However, it remains unclear when, where,and how novel pathogens such as HIV andSARS moved from their natural hosts intohuman populations. Furthermore, forpersistent human infections such asinfluenza, evolutionary approaches providenot only insights into the origins anddiversification of the pathogens, but alsoallow us to make predictions about futureoutbreaks and anticipated strains (Bush etal. 1999). bioGENESIS, in cooperation withecoHEALTH Cross-cutting Network, willpromote integrative studies of such issues,bringing evolutionary biologists togetherwith ecologists, health scientists, andresearchers in related disciplines.
The spread of disease in relation to globalchanges affects not only humans, but otheranimals and the plants upon which wedepend. The migration and evolution ofpathogens through wild species often drivesdramatic ecosystem changes. For example,migrations of a chestnut fungus from
eastern Asia to eastern North America, andof a pine nematode from eastern NorthAmerica to eastern Asia, caused precipitousdeclines of chestnut and pine forests,respectively (Box 20). The ongoing loss ofhemlock trees in eastern North America dueto the introduction of the woolly adelgidfrom Japan provides another example ofthis devastating phenomenon. bioGENESIS,again with ecoHEALTH, will facilitateinternational, interdisciplinary projectsaimed at better understanding themigration and evolution of pathogensinfecting wild organisms.
Another issue directly relevant to Task 3.3is the evolution of pathogens resistant topesticides, herbicides, fungicides, andantibiotics. Uses of these chemicals inagriculture and human medicine have drivenrapid and repeated evolution of resistantpathogens. Evolutionary studies of theacquisition of resistance have oftenrevealed unexpected mechanisms such asrecombination and the horizontal transferof resistance genes. Whereas the molecularevolution of resistance has beendocumented in a few systems (especiallyviral and bacterial systems), our knowledgeof the evolution of resistance remainslimited in many pests and weeds. AbioGENESIS working group will play animportant role in promoting comparativestudies of the evolution of resistance inpathogens brought on by human activities.
Population dynamics of pathogens havebeen of continued interest in medicalscience and also in ecology. While classicmodels describing dynamics withoutevolution made significant contributions toour understanding of epidemiology, thedynamics of hosts and pathogens is a morecomplicated process owing to the evolution
of resistance in hosts, virulence inpathogens, and drug resistance inpathogens. Thus, evolutionary approachesare critical in developing more predictivemodels of host-pathogen dynamics. Ifhistorical samples are available, pastpopulation sizes can be estimated frompopulation genetic theory (Box S14), andinferred population dynamics could, in turn,be used to test evolutionary models. AbioGENESIS working group will coordinatetheoretical and empirical studies of host-pathogen dynamics in human and non-human systems that specifically incorporateevolutionary approaches.
Research Objectives and Activities
• Promote integrative studies on theorigin, migration, and diversification ofpathogens.
• Facilitate analyses of the mechanismsunderlying the evolution of resistanceand its consequences for theconservation of biodiversity
• Coordinate evolutionary studies of host-pathogen dynamics in human and non-human diseases in relation tohuman-induced environmental change.
© A
Hend
ry

37
BOX 19. Drug resistance in HIV
The emergence of drug resistance in HIV has perhaps been the single largest setbackin the treatment of AIDS. Once touted as a cure for HIV infection, early drugtherapies failed to take into account the rapid evolution of the virus resulting indrug resistant strains. No doubt, the development of highly active antiretroviraltherapy (HAART) has greatly extended life expectancy and quality of life for thosesuffering from AIDS who can access these expensive drugs. The failure as a cure hasled to many important lessons concerning our understanding of HIV biology. Usingthe same phylogenetic approaches developed to investigate evolutionaryrelationships among species andwithin populations, we can estimaterelationships among viruses isolatedfrom a single patient (Fig. 19.1)using viral sequence data collectedover time (both before the start ofdrug therapy and after theemergence of drug resistant strains).Using the inferred evolutionaryhistory, we can quickly identify themutations associated with drugresistance that have resulted inwholesale changes in the HIVpopulation through selective sweeps.
References
Crandall KA, Vasco D, Posada D and ImamichiH. 1999. Advances in understanding theevolution of HIV. AIDS 13:S39-S47
Rambaut A, Posada D, Crandall KA and HolmesEC. 2004. The causes and consequences of HIVevolution. Nature Review Genetics 5:52-61
54 a.a. position
82 a.a. position
160 A G
245 T C
ATC (Ile)
GTC (Val)
GTC (Val)
GCC (Ala)
1 Nucleotide difference
N° ofclones
10 (7)*
8 (4)*
11 (4)*
9 (4)*
12 (4)*
9 (3)*
11 (5)*
Heterogeneity (%)
2.13±1.17
0.25±0.22
0.18±0.22
0.36±0.29
0.25±0.22
0.34±0.24
0.29±0.22
HIV-RNA(copies/ml)
119,000
<500
<500
510
869
42,730
63,890
Day 0
Wk. 6
Wk. 100
Wk. 104
Wk. 108
Wk. 113
Wk. 116
* : Number of unique haplotypes within the pod.
FIG. 19.1. Evolutionary network of HIVsequences from the protease gene collected atdifferent time points (from Crandall et al.1999). Note the evolution of amino acidreplacements conferring drug resistance(highlighted in the boxes) and the associatedwholesale replacement of the HIV population bydrug resistant forms. Circles represent the HIVsequence types; diameters represent thefrequency of each type in the population.

38
BOX 20. Origin and evolution of introduced pests in forest ecosystems
Some of the most dramatic changes in forest ecosystems have been driven byintroduced pests. Phylogenetic approaches have been successfully utilised todetermine the origin of those pests. In the case of a pinewood nematode,Bursaphelenchus xylophilus, that caused epidemics of pine wilt disease and rapiddecline of pine forests in Japan (Togashi et al. 2004), molecular phylogeneticevidence showed that it is a lineage native to the USA (Mamiya 1988). This Americanpinewood nematode is not harmful to pines in its native range. Likewise, the nativeJapanese pinewood nematode, B. mucronatus, is avirulent on Japanese pines. It isprobable that the nematodes have weakened their virulence to their native hostsduring their co-evolution.
To predict the fate of invasive pests, monitoring the evolution of virulence iscritically important. It is theoretically expected that a-virulent strains evolve withdecreased host density and this is supported by observation in the mixoma virusintroduced to Australia in order to control naturalized rabbits. A recent study inB. xylophilus showed that its virulence is a heritable trait, with high virulence beingclosely related to a high rate of reproduction and within-tree dispersal, and lessvirulent strains of the pine nematode being selected under decreased host density(Togashi and Jikumaru 2006). It is predicted that the evolution of less virulentstrains may stabilise the coexistence of Japanese pines and the introducednematode. Even if this prediction were supported, large area of pine forest havealready been lost in Japan; the annual loss of pines reached a maximum value of2,430,000 m3 in 1979, and has held steady at about 1,000,000 m3 since the early1990's. Eradication in the early stage ofcolonisation is of primary importance incontrolling invasive pests, andphylogenetic knowledge of B. xylophilusis now being utilised in surveillance inEast Asian countries and in Europe.
References
Mamiya Y. 1988. History of pine wilt disease inJapan. Journal of Nematology 20:219-226
Togashi K, Chung Y-I and Shibata E. 2004. Spreadof an introduced tree pest organism - the pinewoodnematode. In: Hong S-K, Lee JA, Ihm B-S, FarinaA, Son Y, Eun-Shik K and Choe JC (Eds). Ecologicalissues in a changing world: Status, response andstrategy. Kluwer Academic Publishers
Togashi K and Jikumaru S. 2006. Evolutionary change in a pine wilt system following the invasion of Japanby the pinewood nematode, Bursaphelenchus xylophilus. Ecological Research 22:862-868
FIG. 20.1. Left: A pine forest devastated by an epidemic of pine wiltdisease. Right: The pinewood nematode, Bursaphelenchus xylophilus,causes pine wilt disease by damaging vessels in the wood andpreventing water use, ©K Togashi.

39
TASK 3.4 Evolutionary conservation
The knowledge provided by various globalbiodiversity observations now beingintegrated by GEO BON (see Task 1.2; Box3) will need to be integrated withconservation efforts. Evolutionarydisciplines, including systematics,phylogenetics, and population genetics, canand must make significant contributions tothis effort. A fundamental aim here is toidentify strategies for protecting theprocesses that generate diversity andmaintain adaptation; this is crucial tomaintain diversity in the face of rapidglobal change (Stockwell et al. 2003;Pressey et al. 2007). A further goal is toimprove the prediction of spatial patternsof genetic and phylogenetic diversity lossusing methods from evolutionarybiogeography.
At the ecosystem level, phylogenetic treesof component species enable us to measurethe diversity of the system in terms ofevolutionary lineages, and to compare thisphylogenetic diversity (Faith 1992) amongecosystems, regions, and local communities(Box 21). This information adds animportant evolutionary dimension inassessing conservation priorities. Currentmethods typically focus on measuring andcomparing endemism in representativegroups, such as birds and plants.Phylogenetic methods are necessary toestablish whether endemic species haveevolved recently (neoendemics) or representmuch older lineages (paleoendemics), andthus provide a more nuanced evaluation ofthe risks and choices for conservation.
The flip-side of seeking to maximisephylogenetic diversity in protected areas
(Box S16, S17) is to identify “evolutionaryhotspots” that are geographic areas andlandscapes in which species haveundergone recent diversification. Afascinating example of an evolutionaryhotspot is Lake Victoria where hundreds ofcichlid species with extremely diversemorphologies have evolved in less than15,000 years (Box 7). While phylogeneticdiversity is very low in the Lake Victoriacichlids, the evident morphological diversityreflects the divergence of functional genescontrolling body colour patterns, spectralsensitivity, jaw morphology and others(Kocher 2004). Additional examples ofevolutionary hotspots are found in the CapeFloristic Province of South Africa, whichharbours an extremely diverse flora, and incoastal and montane California withconcentrations of recently evolved mammals(Box 21).
At the species level, the recognition ofdistinct taxa is obviously a prerequisite toany conservation effort. Molecularphylogenetic techniques are increasinglyused in discovering and properly diagnosingcryptic taxa, and thus in establishing thedegree to which evolutionary lineages arethreatened. For example, mtDNA sequencedata revealed an ancient phylogenetic splitwithin the common carp between the LakeBiwa lineage endemic to Japan and theEurasian wild lineage (Mabuchi et al. 2005).The Lake Biwa lineage is now properlyrecognised as a threatened cryptic speciesurgently in need of conservation.Comparative phylogeographic data also serveto identify geographic foci of distinctlineages and their interactions, which can beincorporated into strategies for protectingboth pattern and process (Box 21, 22).
© A
Hend
ry

40
At the level of within-population geneticdiversity, it is widely appreciated thatdecreases in population size can have avariety of deleterious effects and increaseextinction risk. Loss of genetic variance in apopulation restricts evolutionary potentialunder environmental change. For example,some lowland-rainforest species ofDrosophila have lost the capacity torespond to selection for desiccationresistance (Kellermann et al. 2006),perhaps reflecting severely reducedpopulation size during the last glacialmaximum. In plants, mating success isreduced if allelic diversity is lost at self-incompatibility loci. Increased inbreeding isassociated with a decrease of populationmean fitness due to increased homozygosityof deleterious alleles. The evolutionaryanalysis of these effects is crucial indevising appropriate conservation measures.
While preserving within-population geneticdiversity is important for maintaining viable
wild populations, greater genetic diversityis maintained between species, and thefunctions and processes of ecosystems arelargely dependent on this between-speciesgenetic diversity. In view of the rapid lossof ecosystems such as tropical forests andcorals, we urgently need to quantify howmany genes and alleles are being lost atlocal, regional, and global scales.
Rapid advances in the efficiency of DNAsequencing have enabled the examinationof far larger samples of organisms andgenes, which greatly expands our capacityto utilise such data in monitoringbiodiversity change and in devisingconservation strategies. In somecircumstances it may be possible to identifyand monitor variation in the genes thatdetermine response to environmental stress(Hoffmann and Willi 2008). A bioGENESISworking group in this area will foster theapplication of advanced genomictechnologies and evolutionary approaches
to conservation biology, forging alliancesbetween evolutionary and conservationbiologists focused on particular systemsand promoting global efforts to monitorbiodiversity change by integrating gene-level observations with species diversityand ecosystem function.
Research Objectives and Activities
• Promote the concept of protectingevolutionary processes, and the use ofmeasures of phylogenetic diversity inconservation decision making.
• Accelerate recognition and description ofcryptic lineages in need of conservation.
• Develop approaches to predicting spatialpatterns of genetic and phylogeneticdiversity loss.
• Promote global genetic diversityobservation by coordinating theapplication of advanced genomicapproaches to biodiversity monitoringand conservation practice.
© B
A W
alth
er

41
FIG. 21.1. For each quarter degree square within the Cape ofSouth Africa, Forest et al. (2007) calculated (b) genus diversity;(c) phylogenetic diversity (PD); (d) residuals from a regression ofPD on genus richness; and (e) unusual PD values, where blueindicates significantly lower PD, © F Forest.
a: © istockphoto.com/M Dabell
BOX 21. Phylogenetic diversity, evolutionary mechanisms, andconservation priorities
Biodiversity assessments typically focus on the number of species as the quantity ofprimary interest, and it is tempting to base priorities for conservation on the numberof species that might be protected in different areas. Increasingly, however, it hasbeen recognised that the number of species in an area is not necessarily directlyrelated to the amount of phylogenetic diversity that is represented there. Two areasmay have the same number of species, but one might contain species that are veryclose relatives that originated in situ only recently, whereas the other might includedistantly related species that represent much deeper splits in the tree of life. Thelater area encompasses more evolutionary history. Choosing conservation areas so asto maximise phylogenetic diversity (PD) may preserve the greatest “featurediversity”, the greatest evolutionary potential, and the greatest number of optionsfor the future use of biodiversity.
One early application of the phylogenetic approach examined the amount ofphylogenetic diversity uniquely represented by the mammals endemic to the world'shotspots (Sechrest et al. 2002). The phylogenetic clumpingof the endemic taxa within hotspots implies that loss ofthese species would mean that not only terminal branchesbut also deeper branches would be lost. Thus, loss of theglobal hotspots would imply a large loss of evolutionaryhistory.
A recent phylogenetic diversity application focused on theextraordinarily diverse flora of the Cape region of SouthAfrica (Forest et al. 2007). The western Cape harbours morespecies but less phylogenetic diversity, in part owing torecent evolutionary radiations in several major lineageswithin that area. In contrast, the Eastern Cape harboursfewer species, but relatively more phylogenetic diversity. Thisknowledge of phylogenetic relationships highlights theexistence of different, potentially decoupled, objectives forconservation efforts, and will generally promote thedevelopment of wiser strategies for biodiversity management.
References
Forest F, Grenyer R, Rouget M, Davies TJ, Cowling RM, Faith DP, Balmford A,Manning JC, Proches S, van der Bank M, Reeves G, Hedderson TAJ andSavolainen V. 2007. Preserving the evolutionary potential of floras inbiodiversity hotspots. Nature 445(7129):757-760
Sechrest W, Brooks TM, da Fonseca GAB, Konstant WR, Mittermeier RA, PurvisA, Rylands AB and Gittleman JL. 2002. Hotspots and the conservation ofevolutionary history. Proceedings of the National Academy of Sciences USA99:2067-2071
b c
d e
a
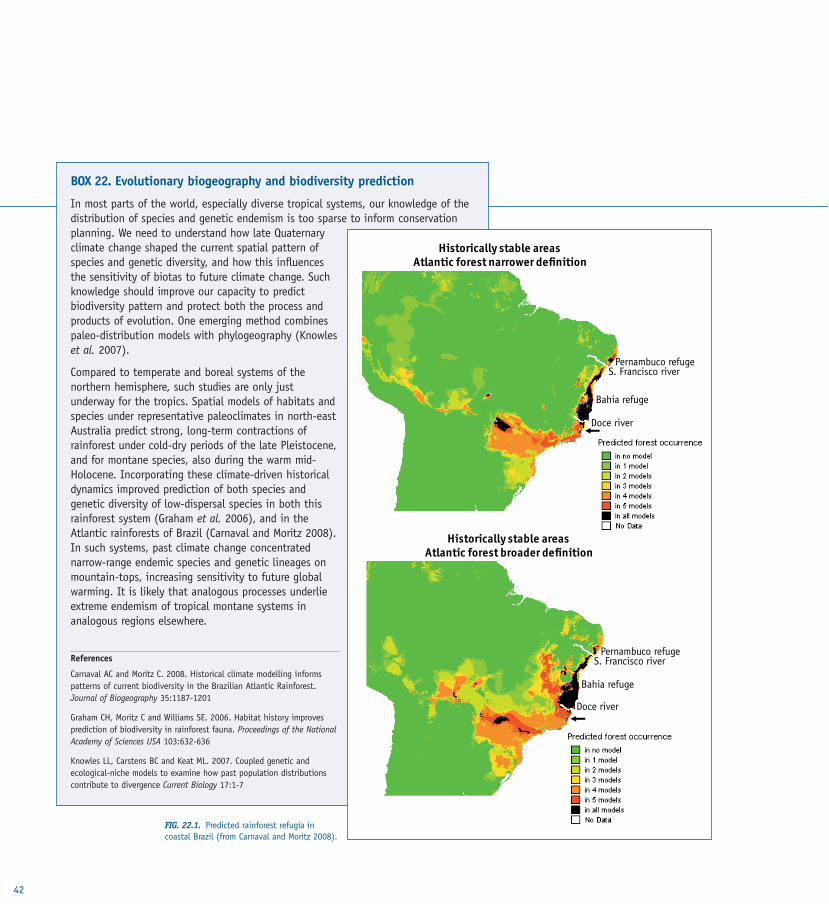
42
BOX 22. Evolutionary biogeography and biodiversity prediction
In most parts of the world, especially diverse tropical systems, our knowledge of thedistribution of species and genetic endemism is too sparse to inform conservationplanning. We need to understand how late Quaternaryclimate change shaped the current spatial pattern ofspecies and genetic diversity, and how this influencesthe sensitivity of biotas to future climate change. Suchknowledge should improve our capacity to predictbiodiversity pattern and protect both the process andproducts of evolution. One emerging method combinespaleo-distribution models with phylogeography (Knowleset al. 2007).
Compared to temperate and boreal systems of thenorthern hemisphere, such studies are only justunderway for the tropics. Spatial models of habitats andspecies under representative paleoclimates in north-eastAustralia predict strong, long-term contractions ofrainforest under cold-dry periods of the late Pleistocene,and for montane species, also during the warm mid-Holocene. Incorporating these climate-driven historicaldynamics improved prediction of both species andgenetic diversity of low-dispersal species in both thisrainforest system (Graham et al. 2006), and in theAtlantic rainforests of Brazil (Carnaval and Moritz 2008).In such systems, past climate change concentratednarrow-range endemic species and genetic lineages onmountain-tops, increasing sensitivity to future globalwarming. It is likely that analogous processes underlieextreme endemism of tropical montane systems inanalogous regions elsewhere.
References
Carnaval AC and Moritz C. 2008. Historical climate modelling informspatterns of current biodiversity in the Brazilian Atlantic Rainforest.Journal of Biogeography 35:1187-1201
Graham CH, Moritz C and Williams SE. 2006. Habitat history improvesprediction of biodiversity in rainforest fauna. Proceedings of the NationalAcademy of Sciences USA 103:632-636
Knowles LL, Carstens BC and Keat ML. 2007. Coupled genetic andecological-niche models to examine how past population distributionscontribute to divergence Current Biology 17:1-7
Historically stable areas
Atlantic forest narrower definition
Historically stable areas
Atlantic forest broader definition
Bahia refuge
Pernambuco refugeS. Francisco river
Doce river
Bahia refuge
Pernambuco refugeS. Francisco river
Doce river
FIG. 22.1. Predicted rainforest refugia incoastal Brazil (from Carnaval and Moritz 2008).

43
Management Structure
The activities of the bioGENESIS CoreProject are overseen by a ScientificCommittee (SC) with expertise in theevolutionary aspects of biodiversity science.The SC-bioGENESIS is appointed by theDIVERSITAS Scientific Committee. Duties ofthe SC-bioGENESIS include:
• Providing scientific guidance in thedevelopment and implementation ofbioGENESIS, especially in stimulating thedevelopment of research networks, andidentifying funding for, and monitoringthe progress of, working groups, andeducational activities centred around thefocal areas and tasks outlined in thisScience Plan ;
• Linking bioGENESIS activities to the otherCore Projects and Cross-cutting Networksof DIVERSITAS and to other relevantnational and international programmes ;
• Encouraging national governments andregional funding agencies to supportbioGENESIS-related research at national,regional, and international levels, andproviding expert advice relevant to theformulation of biodiversity policy andconservation management.
International Project Office
The activities of bioGENESIS are supportedthrough the bioGENESIS InternationalProject Office (IPO), currently hosted by theDIVERSITAS Secretariat in Paris, France.Along with the SC-bioGENESIS members, theIPO is responsible for helping to obtainfunding for the proposed activities, toorganise these activities, and to facilitatelinks across DIVERSITAS and the widercommunity, including with relevant nationaland international programmes. The resultsof bioGENESIS activities are communicatedthrough reports, publications, and theDIVERSITAS website (www.diversitas-international.org).
bioGENESIS Activities
In its start-up phase (2007-2008),bioGENESIS co-sponsored a set of relevantmeetings/symposia as a mechanism topromote its scientific agenda and publicisethe existence of the programme. In 2007,these included meetings to (1) helpcoordinate international “tree of life” and“barcoding” activities (USA); (2) foster thedevelopment of phylogenetic research andapplications in China (Beijing);(3) promote the development and use ofphylogenetic diversity (PD) measures inconservation efforts (New Zealand);(4) explore the origin and evolution of theMediterranean biota (Switzerland); and(5) focus attention on the uses ofphylogenies in tropical ecology (Mexico).These meetings yielded a variety ofproducts, including publications ofsymposium proceedings in the Journal ofBiogeography and in Evolution andSystematics. SC-bioGENESIS members alsoparticipated in pre-meetings of the CBD'sConference of the Parties in Curitiba, Brazil(COP8).
Imple
men
tati
on
Str
ate
gy
III
© M
Don
oghu
e

44
In 2008, bioGENESIS co-sponsored meetingson (1) future trends in taxonomy(Portugal); (2) biome boundary shifts(Japan); (3) phylogenetic communityecology (Japan); (4) the GBIF 2010Campaign (UK); (5) evolutionaryapplications in conservation (Canada); (6)
dating phylogenetic divergences (Spain);(7) speciation and biodiversity inPatagonia (USA); and (8) the establishmentof an international regime on access andbenefit sharing (Germany). SC-bioGENESISmembers also participated in pre-conference events for the CBD COP9 inBonn, Germany, and in the Global TaxonomyInitiative (GTI) meeting, in Bonn, Germany.
In its current phase, SC-bioGENESISmembers are committed to leading a varietyof activities associated to the focal areasand specific tasks described in the SciencePlan. Specific areas of responsibility areprovided on the bioGENESIS web site. Theseactivities include the formation of workinggroups and the organization of meetings tostimulate relevant science and outreachactivities. The activities of each workinggroup will take place over a specified timeperiod, as appropriate, with new groupsbeing formed as others complete theirmissions. The SC-bioGENESIS will monitorthe progress of the working groups and willentertain requests for support for specificworking group activities. As appropriate,these activities will be supported, in part,directly through DIVERSITAS. However, ingeneral, outside funding and partnershipswill be sought from national and regionalsources, foundations, and non-governmentalorganizations.
Special efforts will be made to linkbioGENESIS with the other DIVERSITAS CoreProjects and Cross-cutting Networks, in allcases providing an evolutionary perspectiveon the problem at hand. Recently,bioGENESIS has engaged with the GlobalMountain Biodiversity Assessment (GMBA)to help integrate georeferencing and nichemodelling in assessing the past, present,and future of mountain biodiversity.bioGENESIS is also now involved in aworking group on the evolution offunctional traits in relation to ecosystemfunction. This project, funded through theCentre for Population Biology (SilwoodPark, Imperial College, UK), forms aproductive link between bioGENESIS andecoSERVICES. Similarly, we are engagingwith colleagues in bioDISCOVERY to explorethe connection between rapid evolution,climate change, and ecosystem function,and an Imperial College, UK, working groupfocused on “eco-evolutionary dynamics” is
working toward the publication of itsfindings. With bioDISCOVERY we also areinvolved in the ongoing development ofGEO BON, focusing special attention onmechanisms to assess and monitor globalgenetic/phylogenetic diversity. Theseconnections will be fostered through ouractive participation in the DIVERSITASannual Scientific Committee meetings andits Open Science Conferences.
We will continue to encourage and co-sponsor relevant workshops and symposia,and will especially promote activitiesfocused on education and capacity building.For example, we are sponsoring a workshopfor Latin American students onphylogenetic methods in relation tobiodiversity, organised through theUniversity of Mexico.
bioGENESIS will also provide concrete linksbetween DIVERSITAS and biodiversityorganizations such as the Global
© B
A W
alth
er

45
Biodiversity Information Facility (GBIF), theTaxonomic Database Working Group (TDWG),the International Barcode of Life (IBOL),the Encyclopedia of Life (EOL), and theEuropean Distributed Institute of Taxonomy(EDIT). bioGENESIS members have beenactive in connection with the Conventionon Biological Diversity (CBD), especiallythrough SBSTTA activities and the GlobalTaxonomy Initiative “CoordinationMechanism” (GTI CM). In general, we willactively engage in activities that connectbiodiversity science to policy, and willparticipate in relevant national, regional,and global biodiversity conservation efforts.
Getting involved
There are many ways to participate in DIVERSITAS and to support the activities of the bioGENESISCore Project, as an individual scientist, through the establishment or participation in a NationalCommittee, or as a funder.
The activities highlighted above are meant only to provide examples of projects that might becarried out in connection with bioGENESIS. We encourage scientists to propose additional activitiesthat support the goals outlined in the bioGENESIS Science Plan including:
• Proposals for collaborative research or educational initiatives • Meetings, symposia, and workshops• Synthetic activities and products, including databases and web resources
bioGENESIS also welcomes requests for the endorsement of activities that embrace its goals. Suchproposals should be submitted in the early planning stages of the event or initiative. We welcomeyour involvement in fulfilling the mission of bioGENESIS!
© B
A W
alth
er

46

47
Both C, Bouwhuis S, Lessells CM and Visser ME.2006. Climate change and population declinesin a long-distance migratory bird. Nature441(7089):81-83
Bouchet P, Lozouet P, Maestrati P and Heros V.2002. Assessing the magnitude of speciesrichness in tropical marine environments:exceptionally high numbers of molluscs at aNew Caledonia site. Biological Journal of theLinnean Society 75:421-436
Bush RM, Bender CA, Subbarao K, Cox NJ andFitch WM. 1999. Predicting the evolution ofhuman influenza A. Science 286:1921-1925
Cracraft J and Donoghue MJ. 2004. Assemblingthe Tree of Life. Oxford University Press
Donoghue MJ. 2008. A phylogenetic perspectiveon the distribution of plant diversity.Proceedings of the National Academy of ScienceUSA 105 Supplement 1:11549-11555
Donoghue MJ and Smith M. 2005. Biodiversityinventory: Reflections on preparedness andefficiency. In: Le Duc J-P (Eds). Biodiversity:Science and governance.
Dwyer G, Levin SA and Buttel L. 1990. Asimulation model of the population dynamicsand evolution of myxomatosis. EcologicalMonographs 60:423-447
Edwards EJ, Still CJ and Donoghue MJ. 2007. Therelevance of phylogeny to studies of globalchange. Trends in Ecology and Evolution22(5):243-249
Egan AN and Crandall KA. 2008. Divergence anddiversification in North American Psoraleae(Fabaceae) due to climate change. BMC Biology.In press
Endler JA. 1980. Natural selection on colourpatterns in Poecilia reticulata. Evolution 34:76-91
Faith DP. 2002. Conservation evaluation andphylogenetic diversity. Biological Conservation61:1-10
Grant PR and Grant BR. 2006. Evolution ofcharacter displacement in Darwin's finches.Science 313:224-226
Hawks J, Wang ET, Cochran GM, Harpending HCand Moyzis RK. 2007. Recent acceleration ofhuman adaptive evolution. Proceedings of theNational Academy of Sciences USA 104:20753-20758
Heywood VH. 1995. Global biodiversityassessment. Cambridge University Press
Hoffmann AA and Willi Y. 2008. Detecting geneticresponses to environmental change. NatureReview Genetics 9:421-432
Hong DY, Chen ZD, Qui YL and Donoghue MJ.2008. Tracing patterns of evolution through theTree of Life: Introduction. Journal ofSystematics and Evolution 2008 46(3): 237-238
Hughes JB, Daily GC and Ehrlich PR. 1997.Population diversity: its extent and extinction.Science 278:689-692
Kellermann VM, Heerwaarden BV, Hoffmann AAand Sgro CM. 2007. Very low additive geneticvariance and evolutionary potential in multiplepopulations of two rainforest Drosophila species.Evolution 60:1104-1108
Kim ST, Sultan SE and Donoghue MJ. 2008.Allopolyploid speciation in Persicaria(Polygonaceae): Insights from a low-copynuclear region. Proceedings of the NationalAcademy of Sciences USA 105(34):12370-12375
Kocher TD. 2004. Adaptive evolution andexplosive speciation: the cichlid fish model.Nature Review Genetics 5:288-298
Mabuchi K, Miya M, Senou H, Suzuki T andNishida M. 2006. Complete mitochondrial DNAsequence of the Lake Biwa wild strain ofcommon carp (Cyprinus carpio L.). Aquaculture257:68-77
Magurran AE. 2005. Evolutionary ecology: TheTrinidadian guppy. Oxford University Press
Refe
ren
ces IV
© J Cracraft

48
Marshall LG, Webb SD, Sepkoski JJ and Raup DM.1982. Mammalian evolution and the greatAmerican interchange. Science 215:1351-1357
Moore BR and Donoghue MJ. 2007. Correlates ofdiversification in the plant clade Dipsacales:Geographic movement and evolutionaryinnovations. American Naturalist 170:S28-S55
Palkovacs EP, Marshall MC, Lamphere BA, LynchBR, Weese DJ, Fraser DF, Reznick DN, Pringle CMand Kinnison MT. 2009. Experimental evaluationof evolution and coevolution as agents ofecosystem change in tropical streams.Philosophical Transactions of the Royal Societyof London B. In press
Palumbi SR. 2001. The evolution explosion: Howhumans cause rapid evolutionary change.Norton
Phillips BL, Brown GP, Webb JK and Shine R.2006. Invasion and the evolution of speed intoads. Nature 439(7078):803
Pimm SL, Russell GJ, Gittleman JL and Brooks TM.1995. The future of biodiversity. Science269:347-350
Pimm S, Raven P, Peterson A, Sekercioglu CH andEhrlich PR. 2006. Human impacts on the ratesof recent, present, and future bird extinctions.Proceedings of the National Academy of SciencesUSA 103:10941-10946
Pressey RL, Cabeza M, Watts ME, Cowling RM andWilson KA. 2007. Conservation planning in achanging world. Trends in Ecology and Evolution22:583-592
Reusch TBH and Wood TE. 2007. Molecularecology of global change. Molecular Ecology16(19):3973-3992
Roy K, Valentine JW, Jablonski D and Kidwell SM.1996. Scales of climatic variability and timeaveraging in Pleistocene biotas: implications forecology and evolution. Trends in Ecology andEvolution 11:458-463
Schwarz D, Shoemaker KD, Botteri NL andMcPheron BA. 2007. A novel preference for aninvasive plant as a mechanism for animal hybridspeciation. Evolution 61:245-256
Smith SA, Tank DC, Boulanger LA, Bascom-SlackCA, Eisenman K, Kingery D, Babbs B, Fenn K,Greene JS, Hann BD, Keehner J, Kelley-SwiftEG, Kembaiyan V, Lee SJ, Li P, Light DY, Lin EH,Ma C, Moore E, Schorn MA, Vekhter D, NunezPV, Strobel GA, Donoghue MJ and Strobel SA.2008. Bioactive endophytes warrant intensifiedexploration and conservation. Public Library ofScience One 3(8):e3052
Stockwell CA, Hendry AP and Kinnison MT. 2003.Contemporary evolution meets conservationbiology. Trends in Ecology and Evolution 18:94-101
Vermeij GJ. 1991. When biotas meet:understanding biotic interchange. Science253:1099-1104
Webb CO, Ackerly DD, McPeek MA and DonoghueMJ. 2002. Phylogenies and community ecology.Annual Review of Ecology and Systematics33:475-505
Whitham TG, DiFazio SP, Schweitzer JA, ShusterSM, Allan GA, Bailey JK and Woolbright SA.2008. Extending genomics to naturalcommunities and ecosystems. Science 320:492-495
Wiens JJ and Donoghue MJ. 2004. Historicalbiogeography, ecology, and species richness.Trends in Ecology and Evolution 19:639-644
Yesson C and Culham A. 2006. A phyloclimaticstudy of Cyclamen. BMC Evolutionary Biology 6:Art.No. 72

49
This bioGENESIS Science Plan and ImplementationStrategy is the product of a major co-operativeeffort.
DIVERSITAS would like to thank the members of thescoping meeting who met 27-30 April 2006, atICSU, in Paris, France:
Sandra Baldauf, University of York, UK • Brigitte
Bremer, Bergius Botanic Garden, Sweden • Elena
Conti, University of Zürich, Switzerland • Woody
Cotterill, University of Stellenbosch, South Africa• Joel Cracraft, American Museum of NaturalHistory, USA • Jorge Crisci, National Academy ofSciences, Argentina • Michael J. Donoghue, YaleUniversity, USA • James Edwards, GBIF • Daniel P.
Faith, The Australian Museum, Australia • Catherine
Graham, SUNY Stony Brook, USA • Christoph L.
Häuser, Staatliches Museum für NaturkundeStuttgart, Germany • David Hillis, University ofTexas, USA • Kent Holsinger, University ofConnecticut, USA • Anne Larigauderie, DIVERSITAS• Susana Magallon, Universidad Nacional Autonomade México, México • Michel C. Milinkovitch,Université libre de Bruxelles, Belgique • Craig
Moritz, Museum of Vertebrate Zoology, USA• Leandro de Oliveira Salles, Universidade Federaldo Rio de Janeiro, Brazil • Anne-Hélène Prieur-
Richard, DIVERSITAS • Susanne S. Renner,University of Missouri, USA • Isabel Sanmartin
Bastida, Uppsala University, Sweden • Vincent
Savolainen, Kew Royal Botanic Gardens, UK• Simon Tillier, Muséum National d'HistoireNaturelle, France • Rafael Zardoya, Museo Nacionalde Ciencias Naturales, Spain.
DIVERSITAS would also like to thank members of theScientific Committee of bioGENESIS (2006-08), whobuilt upon the work of the scoping team, finalised
the bioGENESIS Science Plan and ImplementationStrategy, and began its implementation:
Elena Conti, University of Zürich, Switzerland• Joel Cracraft, American Museum of NaturalHistory, USA • Keith A. Crandall, Brigham YoungUniversity, USA • Michael J. Donoghue, YaleUniversity, USA, co-Chair • Daniel P. Faith, TheAustralian Museum, Australia • Christoph Häuser,Staatliches Museum für Naturkunde Stuttgart,Germany • Andrew P. Hendry, McGill University,Canada • Carlos Joly, State University of Campinas,Sao Paulo, Brazil • Kazuhiro Kogure, The Universityof Tokyo, Japan • Lúcia G. Lohmann, Universidadede São Paulo, Brazil • Susana A. Magallón,Universidad Nacional Autónoma de México, México• Craig Moritz, Museum of Vertebrate Zoology, USA• Simon Tillier, Muséum National d'HistoireNaturelle, France • Rafael Zardoya, Museo Nacionalde Ciencias Naturales, Spain • Tetsukazu Yahara,Kyushu University, Japan, co-Chair.
DIVERSITAS expresses its gratitude to the followinginstitutions or projects which offered funding for thedevelopment of this Core Project:
• European Distributed Institute of Taxonomy (EDIT,MNHN; Simon Tillier)
• Japan Society for the Promotion of Science, and
• Kyushu University 80th anniversary memorial fund,Japan (Tetsukazu Yahara)
• BIOTA/FAPESP, Brazil (Carlos Joly)
• Universidade de São Paulo, Brazil (Lúcia G. Lohmann)
• Centre National de la Recherche Scientifique(CNRS; poste rouge to Bruno A. Walther), France
• Yale University (Michael J. Donoghue)
Ack
now
ledgem
en
tsV
© A Hendry

50
Finally, DIVERSITAS thanks its members whoprovided core funding:
• Academia Sinica, China-Taipei
• Federal Ministry of Education, Science andCulture, Austria
• French Ministry of Foreign Affairs (MAE)
• French Ministry of Research
• German Federal Ministry of Education andResearch (BMBF)
• German Research Foundation (DFG)
• National Council on Science and Technology(CONACYT), Mexico
• National Council for Scientific and TechnologicalResearch (CONICET), Argentina
• National Research Foundation (NRF), South Africa
• National Science Foundation (NSF), USA
• Natural Environment Research Council (NERC), UK
• The Netherlands Organisation for ScientificResearch (NWO)
• Politique Scientifique Fédérale de Belgique,Belgium
• The Research Council, Norway
• Royal Netherlands Society of Arts and Sciences(KNAW)
• Slovak Academy of Sciences
• Spanish Ministry of Science and Technology(MYCT)
• Swedish Natural Science Research Council (NFR)
• Swiss National Science Foundation (SNF)
• US National Academy of Sciences (NAS).

51
Lis
t of
acr
onym
sVI
ABS
ATOL
BIOTA
CBD
CIPRES
CoML
CONABIO
CTFS
DIWPA
EDIT
EOL
ERIN
ESSP
FAPESP
FIBR
GBIF
GCP
GECAFS
GECHH
GenBank
GEO
GEO BON
GEOSS
GISP
GMBA
GTI
GTI CM
GWSP
HERBIS
IBOL
ICSU
IGBP
IHDP
Access and Benefit Sharing
Assembling the Tree of Life
Programa de Pesquisas em Caracterização, Conservação e Uso Sustentável da Biodiversidade do Estadode São Paulo
UN Convention on Biological Diversity
Cyberinfrastructure for Phylogenetic Research
Census of Marine Life
Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (Mexico)
Center for Tropical Forest Science
DIVERSITAS in Western Pacific and Asia
European Distributed Institute of Taxonomy
Encyclopedia of Life
Environmental Resources Information Network (Australia)
Earth System Science Partnership (DIVERSITAS, IGBP, IHDP, WCRP)
Fundaçao de Amparo à Pesquisa do Estado de Sao Paolo
Frontiers in Integrative Biological Research (US NSF project)
Global Biodiversity Information Facility
Global Carbon Project (ESSP)
Global Environmental Change and Food Systems (ESSP)
Global Environmental Change and Human Health project (ESSP)
NIH (US National Institutes of Health) genetic sequence database
Group on Earth Observations
Group on Earth Observations Biodiversity Observation Network
Global Earth Observation System of Systems
Global Invasive Species Programme
Global Mountain Biodiversity Assessment
Global Taxonomy Initiative
Global Taxonomy Initiative “Coordination Mechanism“
Global Water System Project (ESSP)
Erudite Recorded Botanical Information Synthesiser
International Barcode of Life
International Council for Science
International Geosphere-Biosphere Programme
International Human Dimensions Programme on Global Environmental Change
© J Cracraft

52
IUBS
NASA
NESCent
NSF
SBSTTA
SCOPE
TDWG
TOL
TreeBASE
UNESCO
WCRP
International Union of Biological Sciences
National Aeronautics and Space Administration
National Evolutionary Synthesis Center, Durham, NC, USA
National Science Foundation, USA
Subsidiary Body for Scientific, Technical and Technological Advice (UN CBD)
Scientific Committee on Problems of the Environment
Biodiversity Information Standards, formerly Taxonomic Database Working Group
Tree of Life
Relational database of phylogenetic information hosted by Yale Peabody Museum
United Nations Educational, Scientific and Cultural Organization
World Climate Research Programme

Gett
ing i
nvolv
ed…
The success of DIVERSITAS is directly related to thevoluntary involvement of scientists andorganisations from around the world. The followingparagraphs briefly describe the primary means ofcontributing to this dynamic network of integratedbiodiversity science. More detailed information isavailable in the Getting involved section of our web site: www.diversitas-international.org
as a Scientist
DIVERSITAS invites individual scientists to make theSecretariat aware of their ongoing research and tosuggest ways to integrate local and internationalinitiatives. The DIVERSITAS Secretariat, as well asthe Core Project and Cross-cutting Network offices,welcome proposals for collaborative activities(research projects, workshops, syntheses, etc.) thatsupport the implementation of the DIVERSITASScience Plan.
as a National Committee
DIVERSITAS encourages the establishment ofNational Committees as a means of building a trulyinternational network to support integratedbiodiversity science. These Committees play animportant role in linking national and internationalprogrammes, as well as interacting with policymakers and other stakeholders in their homecountries.
as a Funder
Funding DIVERSITAS initiatives provides an excellentopportunity for individuals and organisations todemonstrate a strong commitment to conservationand sustainable use of biodiversity – issues thatoften have strong appeal for their own stakeholdersand publics. DIVERSITAS welcomes the opportunityto collaborate with private industry, non-governmental/inter-governmental organizations,foundations and associations.
© M Donoghue

I C S U
I U B S
S C O P E
UNESCO
DIVERSITAS SecretariatMuséum National d'Histoire Naturelle (MNHN)Maison Buffon / 57, rue Cuvier – CP 41 / 75231 Paris, Cedex 05, FranceTel: +33 (0)1 40 79 80 40 / Fax: +33 (0)1 40 79 80 [email protected] / www.diversitas-international.org
bioDISCOVERYDeveloping a scientific framework toinvestigate the current extent of biodiversity,monitor its changes and predict biodiversityfutures.
ecoSERVICESExploring the link between biodiversity andecosystem functions and services thatsupport human well-being; seeking todetermine human responses to changes inecosystem services.
DIVERSITASis an international, non-governmental programmewith a dual mission:
• To promote an integrativebiodiversity science, linkingbiological, ecological andsocial disciplines in aneffort to produce sociallyrelevant new knowledge
• To provide the scientificbasis for the conservationand sustainable use ofbiodiversity.
Our
Mis
sion
Collectively, DIVERSITAS Core Projects
comprise a cycle of discovery, analysis
and information sharing that supports the
application of socially relevant knowledge.
The bioGENESIS Science Plan
complements efforts in related areas of:
Desi
gner
Mar
o Ha
as -
Jan
uary
200
9 –
This
doc
umen
t is
prin
ted
on 1
00%
rec
ycle
d pa
per,
cert
ified
by
the
labe
ls B
laue
Eng
el, No
rdic
Eco
labe
l and
Eco
labe
l. eu
ropé
en.
bioSUSTAINABILITYLooking at the science-policy interface forways to support the conservation andsustainable use of biological resources.