gc content-based pan-pox universal pcr assays for poxvirus...
TRANSCRIPT

JOURNAL OF CLINICAL MICROBIOLOGY, Jan. 2010, p. 268–276 Vol. 48, No. 10095-1137/10/$12.00 doi:10.1128/JCM.01697-09Copyright © 2010, American Society for Microbiology. All Rights Reserved.
GC Content-Based Pan-Pox Universal PCR Assays forPoxvirus Detection�
Yu Li,1* Hermann Meyer,2 Hui Zhao,1 and Inger K. Damon1
Centers for Disease Control and Prevention, National Center for Zoonotic, Vector-Borne, and Enteric Diseases, Division ofViral and Rickettsial Diseases, Poxvirus and Rabies Branch, Atlanta, Georgia 30329,1 and Bundeswehr Institute of
Microbiology, Neuherbergstrasse 11, 80937 Munich, Germany2
Received 31 August 2009/Returned for modification 16 October 2009/Accepted 30 October 2009
Chordopoxviruses of the subfamily Chordopoxvirinae, family Poxviridae, infect vertebrates and consist of atleast eight genera with broad host ranges. For most chordopoxviruses, the number of viral genes and theirrelative order are highly conserved in the central region. The GC content of chordopoxvirus genomes, however,evolved into two distinct types: those with genome GC content of more than 60% and those with a content ofless than 40% GC. Two standard PCR assays were developed to identify chordopoxviruses based on whetherthe target virus has a low or high GC content. In design of the assays, the genus Avipoxvirus, which encodesmajor rearrangements of gene clusters, was excluded. These pan-pox assays amplify DNA from more than 150different isolates and strains, including from primary clinical materials, from all seven targeted genera ofchordopoxviruses and four unclassified new poxvirus species. The pan-pox assays represent an importantadvance for the screening and diagnosis of human and animal poxvirus infections, and the technology used isaccessible to many laboratories worldwide.
The poxviruses (family Poxviridae) cause illness character-ized by generalized or localized cutaneous lesions, and mostmember viruses have broad host ranges. The overall broad hostrange of this family is demonstrated by the two subfamilies ofthe Poxviridae. The subfamily Entomopoxvirinae infects insects,and the subfamily Chordopoxvirinae infects vertebrates; thelatter consists of eight genera, and other “unclassified chor-dopoxviruses.” The classified genera are Orthopoxvirus, Parapox-virus, Avipoxvirus, Capripoxvirus, Leporipoxvirus, Suipoxvirus,Molluscipoxvirus, and Yatapoxvirus.
Poxvirus infections of humans, cattle, sheep, goats, compan-ion animals, birds, and zoo animals have been reported world-wide (2) but in general represent an underappreciated cause ofhealth care utilization. Within the United States, until recentlythe international classification of diseases (ICD) codes werenot available for many of these virus infections. Poxvirus in-fections can be clinically confused with other cutaneous dis-ease, and other poxviruses are emerging or reemerging infec-tions in various parts of the world. The majority of humanpoxvirus infections are zoonotic. Poxvirus infections are also asignificant burden to agricultural communities. Capripoxvirusinfections of ruminants cause significant morbidity and mor-tality. Parapoxvirus infections of ruminants and their handlersare endemic in the United States and worldwide; infectedhumans often turn to veterinarians for diagnostic assistance(14). Human monkeypox (an orthopoxvirus) is an emerginginfection with smallpox-like characteristics that was introducedinto the United States in 2003 via global commerce in theexotic pet animal trade. The evolution of monkeypox virus andits two major clades in Africa represents an incompletely un-
derstood emerging infectious risk. Smallpox, caused by variolavirus, an eradicated disease, remains a significant biothreatagent of international concern and the subject of World HealthOrganization-approved research activities. A recent study onhealth care utilization for the other nonzoonotic human pox-virus infections (21) began to define the burden (22/10,000health care visits) of molluscum contagiosum infections in thecommunity. Over the past 5 years, an increasing number ofpotential poxvirus therapies have been studied. Better diag-nostics targeting poxviruses as the cause of cutaneous infec-tions will decrease the inappropriate treatment of these infec-tions. For instance, parapoxvirus infections are oftenmisdiagnosed as cutaneous anthrax, which unnecessarily con-tributes to overuse of antibacterial agents. By using effectivetherapies and prevention measures, after appropriate diagno-sis, the interhuman transmissibility of poxvirus infections willbe reduced, and days lost from work will be reduced.
Poxviruses represent one of largest viruses known and rep-licate in the cytoplasm of the infected cell and encode mostenzymes for their life cycle. The genomes of poxviruses are alinear double-stranded DNA genome in the range of 134 kb(Parapoxvirus) to 330 kb (Avipoxvirus) and encode more than130 genes. The central region, comprised of nearly 100 geneswhich encode viral gene expression, DNA replication, andvirion formation, has a structural arrangement that is con-served in most chordopoxviruses. Between genera of chor-dopoxviruses, host specificity and genome sequence have di-verged. Interestingly, the chordopoxviruses have two distincttypes of genome based on GC content: parapoxviruses, mol-luscipoxviruses, and crocodilepox virus (an unclassified poxvi-rus most similar to molluscipoxvirus) have high GC contents(�60%); the genomes of the other six genera of chordopoxvi-rus have a low GC (30 to 40%) content. The evolutionaryfactors for this divergence are unknown.
The preponderance of data suggests that although the pox-
* Corresponding author. Mailing address: Centers for Disease Con-trol and Prevention, 1600 Clifton Road, MS G43, Atlanta, GA 30333.Phone: (404) 639-2185. Fax: (404) 639-3111. E-mail: [email protected].
� Published ahead of print on 11 November 2009.
268
on June 14, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from
viruses can readily recombine under tissue culture conditions(23), the genomes are quite stable in evolutionary time (15).To develop a PCR assay, or assays, which could easily be usedto screen for the presence of a poxvirus in a clinical sample, wedeveloped two PCR signatures, one that would be expected toamplify nucleic acid from poxviruses with high GC content(high-GC PCR) while the other would be expected to amplifynucleic acid from most poxviruses with low GC content(low-GC PCR).
In this study, we describe the design and validation of thetwo new pan-chordopoxvirus standard PCR assays using over150 chordopoxvirus isolates. We also describe the use of thesenew PCR assays to diagnose and discover previously unknownpoxviruses as the cause of infections based on the sequenceinformation from the resultant PCR amplicons.
MATERIALS AND METHODS
Viruses and DNA preparation. The origins of the viruses and methods forpreparing DNA from purified virions or infected cell cultures are describedelsewhere (5–7, 13, 16, 18, 20, 22). DNA from the CDC poxvirus collection wasused for assay development. In the initial validation of the low-GC PCR assay,we used 10 Eurasian or Old World orthopoxviruses (OPV) including a monkey-pox virus (MPXV_US03_39), a cowpox virus (CPXV_BRT), a variola virus(VARV_BSH75), a laboratory vaccinia virus (VACV_WR), a rabbit poxvirus(RPXV), an ectromelia virus (ETCV_MOS), a camelpox virus (CMLV_V78903), ataterapoxvirus (TATV_DAH68), and two recent vaccinia virus human clinical sam-ples (VACV_07065 and VACV_07070) (1); three North American OPV includinga racoonpox virus (RACV_MD61), a volepox virus (VPXV_CA85), and a skunkpoxvirus (SKPV_WA78); two leporipoxviruses, myxoma virus (MYXV_Lau), rabbitfibroma virus (RFV_KAS); two yatapoxviruses, Yaba monkey tumor virus(YMTV_V83) and tanapox virus (TANV_04) (3); three unclassified poxviruses(Cotia virus from South America [8, 17], the clinical isolate NY_014 from New Yorkstate, and a North American deer poxvirus DPXV_V89); and a nucleic acid sampleextracted from a fox squirrel body lesion (POX_08040).
The development of the high-GC PCR assay utilized nucleic acid extracted
from clinical specimens of parapoxviruses including two orf viruses (ORFV)from Missouri (ORFV_06044 and ORFV_08041); four pseudocowpox viruses(PCPV) from Missouri (PCPV_06025), West Virginia (PCPV_08024), and Ban-gladesh (PCPV_07012 and PCPV 07013); two bovine papular stomatitis viruses(BPSV) from Bangladesh (BPSV_07005) and Washington (BPSV_07058); andmolluscum contagiosum virus (MOCV) (MOCV_08031).
Genome sequence alignment and phylogenetic analysis of sequences of PCRamplicons. A multiple sequence alignment program MAFFT (11) was used forthe alignment of poxvirus genome sequences. The conserved sequences frommultiple genome sequence alignments were studied manually for the design ofpan-poxvirus PCR primers. The sequence editing and alignment from high- andlow-GC PCR amplicons used the DNASTAR Lasergene, version 8, softwarepackage (DNASTAR, Inc, Madison, WI) and BioEdit software (http://www.mbio.ncsu.edu/BioEdit/bioedit.html).
To infer the phylogeny of clinical poxvirus specimens, the Bayesian analysissoftware package BEAST, BEAUti, and Tracer (4) were used. The analyses run aMarkov chain Monte Carlo (MCMC) chain length of 5,000,000 to 10,000,000 (untilan EES value of �100) using the HKY (Hasegawa-Kishino-Yano) nucleotide sub-stitution model with gamma (categories 4) distribution and a proportion of invariantsites with a fixed mean substitution rate at 1.0 and sampling of every 1,000 states.
Primer design and PCR amplification. (i) Low-GC PCR primers. Viral ge-nomes from genera with high-GC content were aligned separately from thosewith low-GC content group. The primer sequences were selected based on thegenome sequence alignment results obtained with strains of low-GC contentpoxviruses (Table 1). The conserved sequences were screened and analyzed byconsidering the sequence specificity, annealing temperature, and potential sec-ondary structure formation with the assistance of Primer Express (version 1.5;Applied Biosystems) software. The selected forward primer was 5�-ACA CCAAAA ACT CAT ATA ACT TCT (insulin metalloproteinase-like protein gene),and the reverse primer was CCT ATT TTA CTC CTT AGT AAA TGA T(intracellular mature virion [IMV] membrane protein gene). The amplicon sizefrom the low-GC PCR assay is about 220 bp (bp). The annealing temperature forthe PCR assay is 50°C as the length of conserved sequences is restricted.
(ii) High-GC PCR primers. The primer design for the high-GC PCR assay wasbased on the available genome sequences of poxvirus strains with high GCcontent. The selection criterion used for the primer selection was similar to thatof the low-GC PCR assay. The selected forward primer was CAT CCC CAAGGA GAC CAA CGA G, and the reverse primer was TCC TCG TCG CCGTCG AAG TC (both primers target an RNA polymerase subunit gene). The
TABLE 1. Selected chordopoxvirus strains representing different genera and %GC contenta
GC contenttype Genus Species Abbreviation GenBank no. Size (bases) %GC
Low GC Capripoxvirus Goatpox virus strain G20-LKV AY077836 149,723 25.3Lumpy skin disease virus strain Neethling 2490 LDSV_NW_LW NC_003027 150,773 25.9Sheeppox virus strain A SPPA_A AY077833 150,057 25
Leporipoxvirus Myxoma virus strain Lausanne MYXV_Lau NC_001132 161,773 43.6Rabbit fibroma virus strain Kasza RFV_KAS NC_001266 159,857 39.5
Orthopoxvirus Camelpox virus strain CMS AY009089 202,205 33.2Cowpox virus strain Germany 91-3 CPXV_Ger91-3 DQ437593 228,250 33.5Ectromelia virus strain Moscow NC_004105 209,771 33.2Monkeypox virus strain USA_2003_039 MPXV_US03_39 DQ011157 198,780 33.1Taterapox virus strain Dahomey 1968 NC_008291 198,050 33.3Vaccinia virus strain Western Reserve NC_006998 194,711 33.4Vaccinia virus strain Copenhagen VACV_COP M35027 191,738 33.4Variola major virus strain Bangladesh 1975 VARV_BSH75 L22579 186,103 32.7
Suipoxvirus Swinepox virus strain Nebraska 17077-99 SWPV_Neb NC_003389 146,454 27.4Yatapoxvirus Tanapox virus strain Kenya TANV_KEN NC_009888 144,565 27
Yaba monkey tumor virus strain Amano YMTV_Amano NC_005179 134,721 29.8Unclassified Poxviridae Deerpox virus strain W-1170-84 DPXV_W1170 AY689437 170,560 27Avipoxvirus Canarypox virus strain ATCC VR111 NC_005309 359,853 30.4
Fowlpox virus strain Iowa NC_002188 288,539 30.9
High GC Molluscipoxvirus Molluscum contagiosum virus strain subtype 1 MOCV_T1 NC_001731 190,289 63.4Parapoxvirus Bovine papular stomatitis virus strain BV-AR02 BPSV_AR02 NC_005337 134,431 64.5
Orf virus strain OV-SA00 ORFV_SA00 NC_005336 139,962 64.3Orf virus strain OV-IA82 ORFV_IA82 AY386263 137,241 64.3
Unclassified Poxviridae Crocodilepox virus strain Zimbabwe CROV_Nile NC_008030 190,054 61.9
a The genome sequences and their references are listed at http://www.poxvirus.org/.
VOL. 48, 2010 GC CONTENT-BASED PCR ASSAYS TO IDENTIFY POXVIRUSES 269
on June 14, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

amplicon size from the high-GC PCR assay is about 630 bp. The PCR assay hasa predicted annealing temperature of 65°C.
(iii) PCR conditions. The validation PCR assays were set up as follows. PCRmixtures contained �10 to 100 ng of viral DNA and a 20 �M primer pair in 50�l of a solution of 50 mM Tris-HCl buffer (pH 9.2); 16 mM (NH4)2SO4; 2.25 mMMgCl2; 2% (vol/vol) dimethyl sulfoxide; 0.1% (vol/vol) detergent Tween 20; 350�M (each) dATP, dCTP, dGTP, and dTTP; and 2 units of the DNA polymerasesTaq and Pwo provided in the Expand Long Template PCR Kit (Roche MolecularBiologicals, Indianapolis, IN). Using a mixture of proofreading DNA poly-merases reduces the potential sequence error from PCR amplification. For PCRamplification, we used a Model 9700 thermocycler (Perkin-Elmer Cetus, Boston,MA) programmed as follows: after 2 min at 92°C for DNA denaturing, reactionmixtures were thermocycled 10 times through successive denaturing (92°C for10 s), annealing (50°C or 65°C for 30 s), and elongation (68°C for 1 min) stepsand then through 20 cycles of denaturing, annealing, and elongation in whicheach successive elongation step added 2 s. Four microliters was inspected for theamplicon size by electrophoresis in a 4% E-gel (Invitrogen-Novex, Carlsbad, CA)run for 30 min (preloaded ethidium bromide). PCR products were stored at 4°C.
RFLP analysis. A four-base-cutter restriction endonuclease, TaqI (5�-TCGA-3�; New England Biolabs, Beverly, MA), was initially used to generate restrictionfragment length polymorphism (RFLP) profiles for differentiation: the PCRamplicons were digested with 2 units of TaqI in 10-�l reaction mixtures incu-bated at 65°C for a minimum of 1 h. Other frequent-cutter restriction enzymescould be used to generate RFLP patterns for DNA which has no TaqI restrictionsite. To visualize the restriction fragments, 5 �l of each digest was separated bya 4% E-gel (Invitrogen-Novex, Carlsbad, CA) run for 30 min (preloadedethidium bromide). The Fluor-S system (Bio-Rad, Hercules, CA) was used tocreate gel digital photo images in tagged image file format (TIFF).
DNA sequencing. The low- and high-GC PCR amplicons were sequenced afterPCR product cleanup using ExoSAP-IT for incubation with the PCR product at37°C for 15 min, followed by incubation at 80°C for 15 min to inactivate theenzymes (USB, Cleveland, OH). The purified products (0.2 to 0.5 �g) weresequenced in separate 10-�l reaction mixtures containing 1 �M (each) oligonu-cleotide primer pair and 4 �l of BigDye Terminator, version 3.1, cycle sequenc-ing RR-100 reagent (Applied Biosystems, Foster City, CA). Mixtures were ther-mocycled 35 times through 96°C for 10 s, 50°C for 5 s, and 60°C for 1 min. Thereaction products were clarified using a DyeEx 2.0 Spin kit (QIAGENE, Valen-cia, CA). Then, 20 �l of Hi-Di Formamide (Applied Biosystems, Foster City,CA) was added to 10 �l of purified sequencing reaction product. Samples wereloaded in a capillary electrophoresis system (Model 3130X1 Genetic Analyzer,Applied Biosystems, Foster City, CA).
Nucleotide sequence accession numbers. The new sequences were depositedin the GenBank, National Center for Biotechnology Information (http://www.ncbi.nlm.nih.gov/). The low-GC content sequences were deposited under thefollowing accession numbers: SKPV_WA78, GQ923123; VPXV_CA85,GQ923124; RACV_MD61, GQ923125; DPXV_V89, GQ923126; 2001_960,GQ923127 (Pox_WA01960); POX_OH08040, GQ923128; Cotia virus,GQ923129; NY_014, GQ923130; VACV_07065, GQ923131; VACV_07070,GQ923132. The high-GC content sequences were deposited under the followingaccession numbers: PCPV_06025, GQ902049; PCPV_08024, GQ902050;PCPV_07012, GQ902051; PCPV 07013, GQ902052; BPSV_07058, GQ902053;BPSV_07005, GQ902054; ORFV_06044, GQ902055; ORFV_08041, GQ902056;MOCV_08031, GQ902057.
RESULTS AND DISCUSSION
Genome sequence alignment of chordopoxviruses. A newmultiple sequence alignment program, MAFFT, was used toalign poxvirus genome sequences (10). MAFFT constructedinitial alignments by the progressive method and finishedalignments using iterative refinement methods, which im-proved the speed of the alignment of large, complex genomesequences of multiple genera of poxviruses. The genomesequence alignment of each genus used is depicted in Fig. 1.The entomopoxvirus genome sequences were not includedin the analysis as they diverged significantly from chor-dopoxviruses, with considerable gene order rearrangement.Within the chordopoxvirus subfamily, avipoxviruses havethe largest genome and are more diverged from other gen-
era of chordopoxviruses. In addition, avipoxvirus genomescontain multiple rearrangements in comparison to otherchordopoxviruses, which introduced numerous gaps in thegenome sequence alignments, reducing the quality of thealignment results. Thus, for the low-GC PCR assay design,we did not include the avipoxvirus genome.
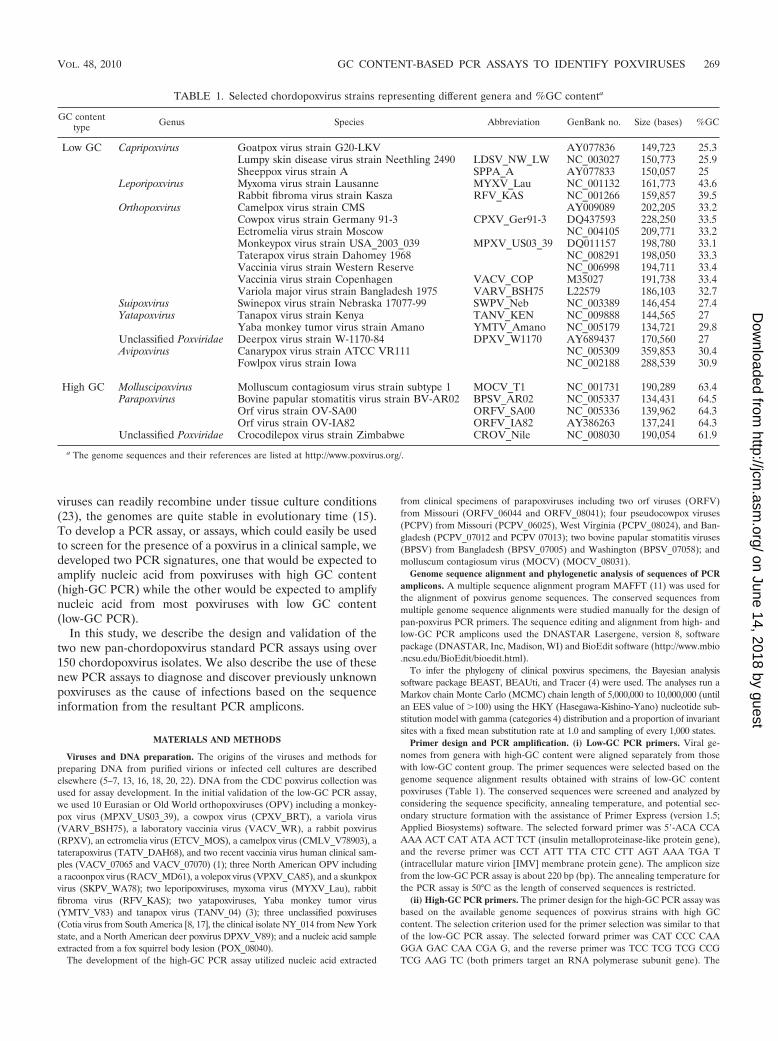
The genome sequence alignments were manually studied toidentify suitable primer targets. The low-GC PCR assay targetselected spans a portion of insulin metalloproteinase-like pro-tein (forward primer, G1L ortholog) to the IMV membraneprotein gene (reverse primer, G3L ortholog) and generates anamplicon about 230 bp in size (Fig. 1A). The primer annealingsite sequences are conserved except for a single nucleotidevariation in the SWPV_NEB and RFV_KAS forward primerannealing sites. Neither single nucleotide polymorphism (SNP)is expected to affect the PCR amplification of these DNA. TheBLAST analysis of low-GC primer sequences with Avipoxvirusgenomes suggests that the low-GC PCR could not amplifyavipoxvirus DNA as the primer annealing target sequenceshave limited homology, and the target size changed signifi-cantly. Genome sequences with high-GC content are limited;most sequences are from the genus of parapoxvirus, but onegenome is from the genus of Molluscipoxvirus and an unclas-sified poxvirus, crocodilepox virus. The high-GC PCR genometarget selected was one of the RNA polymerase subunit genes(VAC-COP J6R ortholog); the primer sequences are con-served, and the primers are predicted to generate an ampliconabout 630 bp in size (Fig. 1B).
Low-GC PCR assay validation. The initial validation usedpurified poxvirus DNAs. As expected, the primers do not am-plify the high-GC poxvirus DNAs. The low-GC PCR assayamplified an appropriately sized amplicon from all the poxvi-rus isolates or clinical materials with known low-GC contenttested (Fig. 2A). This included 10 Eurasian OPV, three NorthAmerican OPV, two leporipoxviruses, two yatapoxviruses,three unclassified poxviruses, and a new poxvirus isolate froma fox squirrel lesion. Concordant with the predicted ampliconsequence alignments, the amplicon sizes are similar among theisolates in different genera (Fig. 2A). The RFLP analysis of theamplicons with TaqI restriction endonuclease digestion pro-vided an additional, convenient way to differentiate some gen-era or species. All Eurasian OPV yield an identical double-banded (about 127 and 107 nucleotides in size) RFLP pattern;North American OPV also yield double bands, but the sizes ofthese bands (about 170 and 64 nucleotides in size) are differentfrom those of the Eurasian OPV. Of note, the newly discov-ered poxvirus NY_014 (CDC personal communication) and adeerpox virus DPXV_V89 from North America have an RFLPpattern similar to that of North American OPV. The twoyatapoxviruses and the unclassified Cotia virus are not cleavedby TaqI. TaqI cleavage of the amplicon produced from the twoleporipoxvirus DNAs produced multiple small fragments; TaqIcleavage of the amplicon generated from the fox squirrel pox-virus lesion also produced multiple bands, but the bandingpattern sizes were not identical to those derived from the twoleporipoxviruses, myxoma virus and rabbit fibroma virus.
High-GC PCR assay validation. The high-GC PCR assaywas designed to amplify the known high-GC content poxvi-ruses: parapoxvirus, molluscum contagiosum virus, and croco-dilepox virus strain Zimbabwe. Neither the parapoxviruses nor
270 LI ET AL. J. CLIN. MICROBIOL.
on June 14, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from
molluscum contagiosum virus is easily propagated in immor-talized cell culture systems. For the validation of the high-GCPCR assay, we directly used nucleic acid from parapoxvirusand molluscum contagiosum virus clinical material for thePCR amplification and sequencing confirmation. Two ORFV,four PCPV, two BPSV, and one molluscum contagiosum virusDNA were amplified (Fig. 2B). The corresponding TaqI RFLPpatterns are unique for each species and agree with the pre-dicted RFLP patterns predicted from sequences (unpublishedresults). The RFLP patterns of BPSV isolates are similar butdifferent from the pattern of MOCV_08031, as predicted bysequences. In comparison, the predicted TaqI RFLP pattern ofMOCV_08031 shows variation from that of MOCV_T1. The
sequences show that MOCV_08031 is about 97% identical toMOCV type I.
Evaluation of the pan-pox assays with a large collection ofpoxvirus isolates. The low- and high-GC PCR assays werefurther tested with 146 poxviruses including nucleic acids di-rectly extracted from one swinepox virus, one crocodilepoxvirus, and clinical materials from one molluscum contagiosumvirus (Table 2). The low-GC PCR assay was able to amplifyorthopoxviruses (including 8 camelpox, 78 cowpox, 17 monkey-pox, 8 mousepox, 14 vaccinia, and 1 raccoonpox virus isolates),capripoxviruses (including 1 lumpy skin disease virus and 1 sheep-pox virus), suipoxvirus (including 1 swinepox virus), yatapoxvi-ruses (including 1 tanapox virus), and 1 Yaba-like disease virus.
FIG. 1. Sequence alignment of the low- and high-GC PCR amplicons. (A) Alignment of amplicon sequences of low-GC PCR assay: 12predicted amplicon sequences represent 12 species within five chordopoxvirus genera and one unclassified poxvirus. The names for the virusabbreviations used on the figure are given in Table 1. (B) Alignment of amplicon sequences of high-GC PCR assay: five predicted ampliconsequences represent four species of three chordopoxvirus genera and one unclassified poxvirus. The primer positions are indicated with arrow bars.
VOL. 48, 2010 GC CONTENT-BASED PCR ASSAYS TO IDENTIFY POXVIRUSES 271
on June 14, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

The high-GC PCR assay was able to amplify DNA from 13parapoxvirus isolates including, 9 orf viruses, 2 bovine papularstomatitis viruses, 1 pseudocowpox virus, and 1 camel parapox-virus species and DNA from 1 molluscum contagiosum virusand 1 crocodilepox virus. Both low-GC and high-GC PCRassays worked well with poxvirus DNA from cell cultures andclinical samples. For the high-GC PCR assay, the DNA wasextracted from clinical samples as high-GC poxviruses are dif-ficult to grow in cell culture systems.
Diagnostics for suspected poxvirus clinical samples or un-known rash specimens. The low- and high-GC PCR assayshave been used successfully to aid in the identification of sus-pected poxviruses in a variety of animal-derived rash speci-mens. The newly discovered poxvirus NY_014, which caused aprogressive panniculitis in an immune-suppressed patient, wasamplified by the low-GC PCR. A DNA sample (POX_08040)extracted from a fox squirrel (Sciurus niger) with multiple nod-ular lesions was tested to rule out the possible infection with
FIG. 2. Low- and high-GC PCR and TaqI RFLP assays. (A) Low-GC PCR amplicons and TaqI RFLP patterns. The top panel shows low-GCPCR amplicons from 20 poxvirus DNA samples. The bottom panel shows corresponding TaqI restriction endonuclease RFLP patterns.(B) High-GC PCR amplicons and TaqI RFLP patterns. The left panel shows high-GC PCR amplicons from nine clinical poxvirus DNA samples.The right panel shows the corresponding TaqI restriction endonuclease RFLP patterns.
272 LI ET AL. J. CLIN. MICROBIOL.
on June 14, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from
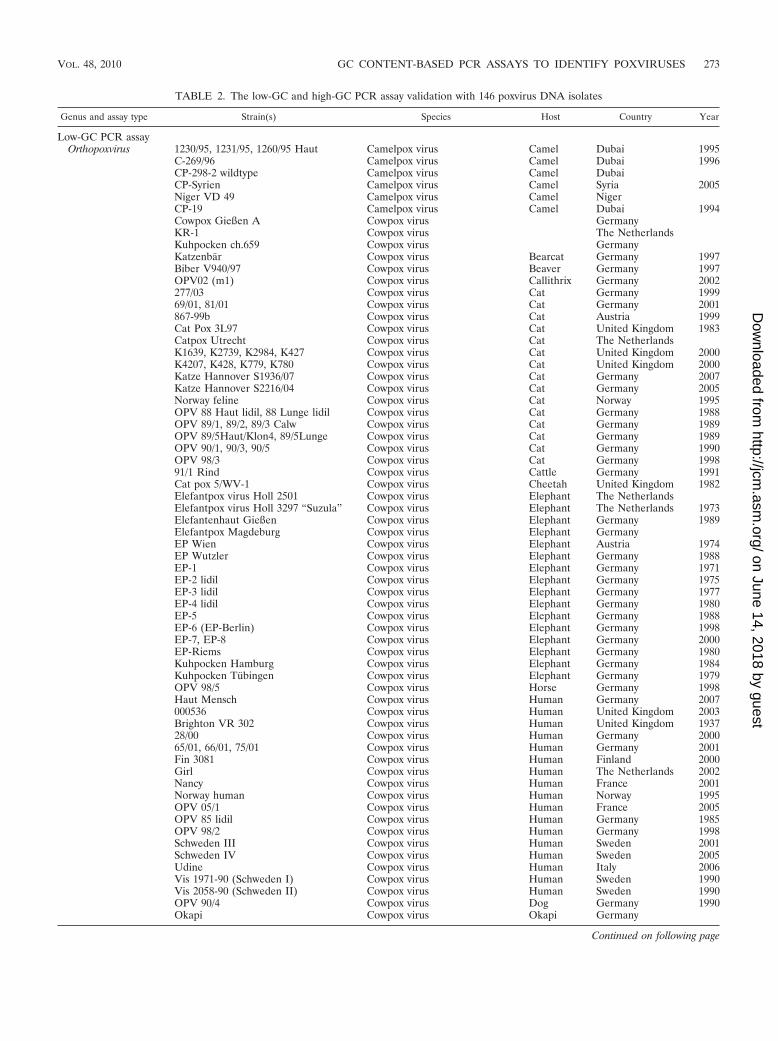
TABLE 2. The low-GC and high-GC PCR assay validation with 146 poxvirus DNA isolates
Genus and assay type Strain(s) Species Host Country Year
Low-GC PCR assayOrthopoxvirus 1230/95, 1231/95, 1260/95 Haut Camelpox virus Camel Dubai 1995
C-269/96 Camelpox virus Camel Dubai 1996CP-298-2 wildtype Camelpox virus Camel DubaiCP-Syrien Camelpox virus Camel Syria 2005Niger VD 49 Camelpox virus Camel NigerCP-19 Camelpox virus Camel Dubai 1994Cowpox Gießen A Cowpox virus GermanyKR-1 Cowpox virus The NetherlandsKuhpocken ch.659 Cowpox virus GermanyKatzenbar Cowpox virus Bearcat Germany 1997Biber V940/97 Cowpox virus Beaver Germany 1997OPV02 (m1) Cowpox virus Callithrix Germany 2002277/03 Cowpox virus Cat Germany 199969/01, 81/01 Cowpox virus Cat Germany 2001867-99b Cowpox virus Cat Austria 1999Cat Pox 3L97 Cowpox virus Cat United Kingdom 1983Catpox Utrecht Cowpox virus Cat The NetherlandsK1639, K2739, K2984, K427 Cowpox virus Cat United Kingdom 2000K4207, K428, K779, K780 Cowpox virus Cat United Kingdom 2000Katze Hannover S1936/07 Cowpox virus Cat Germany 2007Katze Hannover S2216/04 Cowpox virus Cat Germany 2005Norway feline Cowpox virus Cat Norway 1995OPV 88 Haut lidil, 88 Lunge lidil Cowpox virus Cat Germany 1988OPV 89/1, 89/2, 89/3 Calw Cowpox virus Cat Germany 1989OPV 89/5Haut/Klon4, 89/5Lunge Cowpox virus Cat Germany 1989OPV 90/1, 90/3, 90/5 Cowpox virus Cat Germany 1990OPV 98/3 Cowpox virus Cat Germany 199891/1 Rind Cowpox virus Cattle Germany 1991Cat pox 5/WV-1 Cowpox virus Cheetah United Kingdom 1982Elefantpox virus Holl 2501 Cowpox virus Elephant The NetherlandsElefantpox virus Holl 3297 “Suzula” Cowpox virus Elephant The Netherlands 1973Elefantenhaut Gießen Cowpox virus Elephant Germany 1989Elefantpox Magdeburg Cowpox virus Elephant GermanyEP Wien Cowpox virus Elephant Austria 1974EP Wutzler Cowpox virus Elephant Germany 1988EP-1 Cowpox virus Elephant Germany 1971EP-2 lidil Cowpox virus Elephant Germany 1975EP-3 lidil Cowpox virus Elephant Germany 1977EP-4 lidil Cowpox virus Elephant Germany 1980EP-5 Cowpox virus Elephant Germany 1988EP-6 (EP-Berlin) Cowpox virus Elephant Germany 1998EP-7, EP-8 Cowpox virus Elephant Germany 2000EP-Riems Cowpox virus Elephant Germany 1980Kuhpocken Hamburg Cowpox virus Elephant Germany 1984Kuhpocken Tubingen Cowpox virus Elephant Germany 1979OPV 98/5 Cowpox virus Horse Germany 1998Haut Mensch Cowpox virus Human Germany 2007000536 Cowpox virus Human United Kingdom 2003Brighton VR 302 Cowpox virus Human United Kingdom 193728/00 Cowpox virus Human Germany 200065/01, 66/01, 75/01 Cowpox virus Human Germany 2001Fin 3081 Cowpox virus Human Finland 2000Girl Cowpox virus Human The Netherlands 2002Nancy Cowpox virus Human France 2001Norway human Cowpox virus Human Norway 1995OPV 05/1 Cowpox virus Human France 2005OPV 85 lidil Cowpox virus Human Germany 1985OPV 98/2 Cowpox virus Human Germany 1998Schweden III Cowpox virus Human Sweden 2001Schweden IV Cowpox virus Human Sweden 2005Udine Cowpox virus Human Italy 2006Vis 1971-90 (Schweden I) Cowpox virus Human Sweden 1990Vis 2058-90 (Schweden II) Cowpox virus Human Sweden 1990OPV 90/4 Cowpox virus Dog Germany 1990Okapi Cowpox virus Okapi Germany
Continued on following page
VOL. 48, 2010 GC CONTENT-BASED PCR ASSAYS TO IDENTIFY POXVIRUSES 273
on June 14, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

the oral rabies vaccine (a live vaccinia-rabies glycoprotein re-combinant virus) (9). The CDC standard OPV generic andvaccinia-specific real-time PCR assays did not amplify DNAderived from the sample (16); that the low-GC PCR amplifiedthe DNA and sequence from the amplicon shows that theinfection was caused by a leporipoxvirus-like poxvirus (Fig.3A), perhaps related to the squirrel fibroma virus which wasfirst reported by King et al. over 35 years ago (12).The low-GCPCR also identified a new poxvirus (POX_01960) from nucleicacid extracted from formalin-fixed tissue of a rat skin lesion;the rat was a contact of a child with a febrile rash illness in thenorthwestern United States (Fig. 3A) (CDC personal commu-
nication). The amplicon-derived sequence did not have signif-icant identity to any known poxvirus nucleotide sequences in aGenBank search (http://blast.ncbi.nlm.nih.gov). This may sup-port the phylogram, which suggests that sample POX_01960contains a virus which is a member of a new genus of chor-dopoxvirus as it forms a distinct clade.
MOCV_08031 was obtained from a specimen initially sus-pected to be a herpes virus infection. The high-GC PCR assayamplified the clinical sample, and the sequences of the ampli-fied amplicon identified it as a molluscum contagiosum-likevirus (unpublished results). The MOCV has at least threesubtypes (19), and the sequence of the amplicon derived from
TABLE 2—Continued
Genus and assay type Strain(s) Species Host Country Year
Rat Cowpox virus Rat The Netherlands 2002Nashornpocken ch. 15 Cowpox virus Rhino275/03 Cowpox virus Tapir Germany 1999AP-5, WRAIR Monkeypox virus Cynomolgus DRCa 197104MPX095c Monkeypox virus Human DRC 200404MPX099v Monkeypox virus Human DRC 200406MPX 575v, 609v, 699c Monkeypox virus Human DRC 200606MPX 727c, 739c, 743c Monkeypox virus Human DRC 200606MPX 785c, 802v, 850v, 854c Monkeypox virus Human DRC 2006MSF 10, 2, 6 Monkeypox virus Human DRC 2001AP-4, 65-31 Monkeypox virus Monkey DRC 1965Blaufuchs Mousepox virus Fox Czech Republic 1993LTK Moskau Mousepox virus Mouse RussiaMP C99-505 Mousepox virus Mouse United States 1999MP-1 Mousepox virus Mouse Germany 1983MP-3 Mousepox virus Mouse GermanyUS 33221, 4908, Organ9 Mousepox virus Mouse United States 1995ATCC VR 838 Raccoon poxvirus Raccoon United StatesElstree “Wyeth” NYCBHL Vaccinia virusCopenhagen host range Vaccinia virusCopenhagen wild type Vaccinia virusCVA Plaque 7, Hagen lidil, WR Vaccinia virusHolland vaccine, IHD, Levaditi Vaccinia virusBuffelpocken ch.3 Vaccinia virus BuffaloBuffelpox BP-1 lidil Vaccinia virus Buffalo144/89/K. Vaccinia virus Human Germany 1989170/05 Buffelpox Vaccinia virus Human Pakistan 200535/99 Vaccinia virus Human Germany 1999
Capripoxvirus Lumpy skin Lumpy skin disease virusSheeppox vaccine Sheeppox virus
Suipoxvirus Swinepox Swinepox virus SwineYatapoxvirus Tanapox Tanapox virus
Yaba-like Davis strain Yaba-like disease virus Monkey
High-GC PCRassayParapoxvirus MM 7.KOP, B014 Orf virus Human Germany 1996
ORF Orf virus SheepOrf 11 Orf virus Sheep Scotland 1996B032 Namibia Orf virus Sheep Namibia 2003B047 6. BEL Orf virus Goat Germany 1987NZ-2 Orf virus Sheep New Zealand 1982S1 Japan Orf virus Sheep Japan 1985B020 Orf virus Goat Germany 2004V590/75 Bovine papular stomatitis Cattle Germany 1979Kamerun 21 Bovine papular stomatitis Cattle Cameroon 2002B035 Pseudocowpox virus Cattle Germany 2004CE 48 Camel parapoxvirus Camel Sudan 2006
Molluscipoxvirus Molluscum contagiosum Molluscum contagiosum virus Human Germany 2008Unclassified Crocopox Crocodile
a Democratic Republic of the Congo.
274 LI ET AL. J. CLIN. MICROBIOL.
on June 14, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

FIG. 3. Phylogenetic analysis of a chordopoxvirus clinical specimen. (A) The phylogram of low-GC PCR amplicon sequences from which newpoxvirus isolates were inferred (labeled in blue). POX_08040 forms a cluster with the genus Leporipoxvirus, and an unknown poxvirus(POX_01960) forms a distinct clade to all known chordopoxviruses. (B) The phylogram of high-GC PCR amplicon sequences confirms the clinicaldiagnostics of the PCPV and BPSV specimens, which form distinct clades relative to ORFV. MOCV08031 forms a relatively large divergence withmolluscum contagiosum virus type I (MOCV_T1).
VOL. 48, 2010 GC CONTENT-BASED PCR ASSAYS TO IDENTIFY POXVIRUSES 275
on June 14, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

the clinical sample MOCV 08031 is 3% divergent based onnucleic acid content relative to MOCV subtype I (Fig. 3B); thismay indicate that it is another subtype. The high-GC PCRassay amplified three PCPV isolates and two BPSV isolateswhich were initially diagnosed by PCPV and BPSV real-timePCR assays, respectively (unpublished results). Comparativesequence analysis of the amplicons generated by the high-GCPCR assay was concordant with the prior real-time PCR assayresults and additionally indicated that PCPV and BPSV con-tain subclades, as does ORFV (Fig. 3B).
The designs of the low-GC and high-GC pan-poxvirus assayswere based on the available chordopoxvirus genome se-quences. Avipoxviruses have greatly divergent genome se-quences and need a separate diagnostic PCR assay. Thehigh-GC PCR assay was generated with limited but widelydiverged genome sequences and could be further modifiedwhen more high-GC genome sequences become available. Ourinitial test showed that the high-GC PCR did not amplify asealpox virus DNA.
The new low-GC and high-GC PCR assays significantly in-creased our ability to diagnose unknown rash illnesses as pox-virus in origin and gave us preliminary information about ad-ditional, novel poxviruses. Broader use of these assays inscreening for unknown vesiculo-pustular illness may increaseour awareness of poxvirus infections in humans and nonhumananimals worldwide. The assays can be used to monitor zooanimals, domestic animals, or wild animals for mild rash ill-nesses. We expect many new poxviruses can be discovered bythe screening of rash-derived specimens using these pan-poxassays.
ACKNOWLEDGMENTS
We thank M. Buettner (Oberschleissheim) for providing parapoxvirus DNA and W. Eichhorn (Munich) for providing clinical materialof swinepox virus. The skilled assistance of L. Dobrzykowski is greatlyacknowledged.
The findings and conclusions in this report are those of the author(s)and do not necessarily represent the official position of the Centers forDisease Control and Prevention/the Agency for Toxic Substances andDisease Registry.
REFERENCES
1. Centers for Disease Control and Prevention. 2008. Laboratory-acquiredvaccinia exposures and infections—United States, 2005–2007. MMWRMorb. Mortal. Wkly. Rep. 57:401–404.
2. Damon, I. 2007. Poxviruses, p. 2947–2976. In D. M. Knipe, P. M. Howley,D. E. Griffin, R. A. Lamb, M. A. Martin, B. Roizman, and S. E. Straus (ed.),Fields virology, 5th ed. Lippincott Williams & Wilkins, Philadelphia, PA.
3. Dhar, A. D., A. E. Werchniak, Y. Li, J. B. Brennick, C. S. Goldsmith, R.Kline, I. Damon, and S. N. Klaus. 2004. Tanapox infection in a collegestudent. N. Engl. J. Med. 350:361–366.
4. Drummond, A. J., and A. Rambaut. 2007. BEAST: Bayesian evolutionaryanalysis by sampling trees. BMC Evol. Biol. 7:214.
5. Esposito, J. J., R. Condit, and J. Obijeski. 1981. The preparation of or-thopoxvirus DNA. J. Virol. Methods 2:175–179.
6. Esposito, J. J., and J. C. Knight. 1985. Orthopoxvirus DNA: a comparison ofrestriction profiles and maps. Virology 143:230–251.
7. Esposito, J. J., J. F. Obijeski, and J. H. Nakano. 1978. Orthopoxvirus DNA:strain differentiation by electrophoresis of restriction endonuclease frag-mented virion DNA. Virology 89:53–66.
8. Esposito, J. J., E. L. Palmer, E. C. Borden, A. K. Harrison, J. F. Obijeski,and F. A. Murphy. 1980. Studies on the poxvirus Cotia. J. Gen. Virol.47:37–46.
9. Hanlon, C. A., M. Niezgoda, A. N. Hamir, C. Schumacher, H. Koprowski,and C. E. Rupprecht. 1998. First North American field release of a vaccinia-rabies glycoprotein recombinant virus. J. Wildl. Dis. 34:228–239.
10. Katoh, K., K. Kuma, H. Toh, and T. Miyata. 2005. MAFFT version 5:improvement in accuracy of multiple sequence alignment. Nucleic AcidsRes. 33:511–518.
11. Katoh, K., K. Misawa, K. Kuma, and T. Miyata. 2002. MAFFT: a novelmethod for rapid multiple sequence alignment based on fast Fourier trans-form. Nucleic Acids Res. 30:3059–3066.
12. King, J. M., A. Woolf, and J. N. Shively. 1972. Naturally occurring squirrelfibroma with involvement of internal organs. J. Wildl. Dis. 8:321–324.
13. Knight, J. C., C. S. Goldsmith, A. Tamin, R. L. Regnery, D. C. Regnery, andJ. J. Esposito. 1992. Further analyses of the orthopoxviruses volepox virusand raccoon poxvirus. Virology 190:423–433.
14. Lederman, E. R., G. M. Green, H. E. DeGroot, P. Dahl, E. Goldman, P. W.Greer, Y. Li, H. Zhao, C. D. Paddock, and I. K. Damon. 2007. ProgressiveORF virus infection in a patient with lymphoma: successful treatment usingimiquimod. Clin. Infect. Dis. 44:e100–e103.
15. Li, Y., D. S. Carroll, S. N. Gardner, M. C. Walsh, E. A. Vitalis, and I. K.Damon. 2007. On the origin of smallpox: correlating variola phylogenics withhistorical smallpox records. Proc. Natl. Acad. Sci. U. S. A. 104:15787–15792.
16. Li, Y., V. A. Olson, T. Laue, M. T. Laker, and I. K. Damon. 2006. Detectionof monkeypox virus with real-time PCR assays. J. Clin. Virol. 36:194–203.
17. Lopesode, J., P. Lacerde, I. E. Fonseca, D. P. Castro, O. P. Forattini, andE. X. Rabello. 1965. Cotia virus: a new agent isolated from sentinel mice inSao Paulo, Brazil. Am. J. Trop. Med. Hyg. 14:156–157.
18. Massung, R. F., L. I. Liu, J. Qi, J. C. Knight, T. E. Yuran, A. R. Kerlavage,J. M. Parsons, J. C. Venter, and J. J. Esposito. 1994. Analysis of the com-plete genome of smallpox variola major virus strain Bangladesh-1975. Vi-rology 201:215–240.
19. Porter, C. D., and L. C. Archard. 1992. Characterisation by restriction map-ping of three subtypes of Molluscum contagiosum virus. J. Med. Virol.38:1–6.
20. Reed, K. D., J. W. Melski, M. B. Graham, R. L. Regnery, M. J. Sotir, M. V.Wegner, J. J. Kazmierczak, E. J. Stratman, Y. Li, J. A. Fairley, G. R. Swain,V. A. Olson, E. K. Sargent, S. C. Kehl, M. A. Frace, R. Kline, S. L. Foldy, J. P.Davis, and I. K. Damon. 2004. The detection of monkeypox in humans in theWestern Hemisphere. N. Engl. J. Med. 350:342–350.
21. Reynolds, M. G., R. C. Holman, K. L. Yorita Christensen, J. E. Cheek, andI. K. Damon. 2009. The incidence of molluscum contagiosum among Amer-ican Indians and Alaska Natives. PLoS One 4:e5255.
22. Ropp, S. L., Q. Jin, J. C. Knight, R. F. Massung, and J. J. Esposito. 1995.PCR strategy for identification and differentiation of smallpox and otherorthopoxviruses. J. Clin. Microbiol. 33:2069–2076.
23. Yao, X. D., and D. H. Evans. 2003. Characterization of the recombinantjoints formed by single-strand annealing reactions in vaccinia virus-infectedcells. Virology 308:147–156.
276 LI ET AL. J. CLIN. MICROBIOL.
on June 14, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from