gap junction protein in rat hippocampus: light microscope immunohistochemical localization
TRANSCRIPT

THE JOURNAL OF COMPARATIVE NEUROLOGY 281269-281(1989)
Gap Junction Protein in Rat Hippocampus: Light lMicroscope Immunohistochemical
Localization
T. YAMAMOTO, S. SHIOSAKA, M.E. WHIlTAKER, E.L. HERTZBERG, AND J.I. NAGY Department of Physiology, Faculty of Medicine, University of Manitoba, Winnipeg, Manitoba,
Canada R3E OW3 (T.Y., S.S., M.E.W., J.I.N.); Department of Neuroscience, Albert Einstein College of Medicine, Bronx, New York 10461 (E.L.H.)
ABSTRACT An affinity-purified antibody against a 27-kD rat liver gap-junctional
protein (GJP) was used to determine the distribution of GJP immunoreactiv- ity in sections of rat hippocampus. Four heterogeneously distributed GJP- immunostaining patterns were observed. The two most common were punc- tate immunoreactive elements ranging in size from 0.3 to 0.7 pm and networks of immunoreactive varicose fibers coursing in a variety of directions within the various hippocampal layers and ranging in length from a few microns up to 200 pm. The density of punctate immunostaining was highest within a portion of the stratum pyramidale, at the border between the stratum pyramidale and stratum oriens, and at the border between the molecular and granule cell layers of the dentate gyrus. Moderate to low densities were observed in other hippocampal areas. Immunoreactive fibers were most concentrated within the border portions of the stratum pyramidale and oriens, moderately distributed in the stratum radiatum and the remaining part of the stratum oriens, and sparse in the alveus. In the dentate gyrus, fiber networks were most evident a t the border between the granule cell and molecular layers and very unevenly distributed in the molecular layer. The two other patterns observed included intense filamentous immunostaining within a small number of neuronal peri- karya located mainly in the stratum pyramidale of areas CA2 and CA3, but rarely in area CAI or the dentate gyrus, and diffuse immunostaining of small cell bodies dispersed throughout the hippocampus but most numerous in the vicinity of the stratum pyramidale and in the alveus. All of these immuno- staining patterns were seen at all rostrocaudal hippocampal levels. These results suggest that if GJP-immunoreactive fibers and neurons observed in the hippocampus have the capacity to form gap junctions, then electrotonic transmission may constitute an important means of information processing within this structure.
Key words: immunohistochemistry, electrotonic transmission, gap-junc- tional coupling
It is now well recognized that electrical transmission via interneuronal gap-junctional contacts occurs to a much greater extent in the mammalian central nervous system (CNS) than previously imagined. Indeed, electrotonic cou- pling between central neurons in mammals may constitute an important mode of neuronal communication and contrib- ute significantly to the integrative properties of particular neural circuits. Reasons to consider the role of electrotonic neuronal interactions in the CNS of mammals have been previously discussed (Bennett, '72; Sotelo and Korn, '78; Sotelo and Triller, '81; Dudek et al., '83; Llinas, '85). Among
0 1989 ALAN R. LISS, INC.
these is the growing body of morphological and electrophy- siological evidence for the existence of electrotonic coupling in an ever-increasing number of CNS structures. For exam-
Accepted October 12,1988. S. Shiosaka's present address is Dept. of Neuroanatomy, Institute of
Higher Nervous Activity, Osaka University Medical School, 4-3-57 Nakano- shima, Kitaku, Osaka, 530 Japan.
Address reprint requests to Dr. J.I. Nagy, Dept. of Physiology, Faculty of Medicine, University of Manitoba, 770 Bannatyne Avenue, Winnipeg, Mani- toba, Canada R3E OW3.

270
ple, the well-known list of brain regions in which electrical transmission has been discovered in the last decade (re- viewed by Bennett and Goodenough, '75; Bennett, '77; Sotelo and Korn, '78; Sotelo and Triller, '81; Dudek et al., '83) has recently been lengthened to include the hypothala- mus (Andrew et al., '81; Cobbett et al., '85), substantia nigra (Grace and Bunney, '83), and the hippocampus (MacVicar and Dudek, '80, '81, '82; Schmalbruch and Jahnsen, '81; Andrew et al., '82; MacVicar et al., '82; Taylor and Dudek, '82; Kosaka and Hama, '85). This evidence together with results presented here and similar data we have compiled regarding other CNS areas (Nagy, unpublished observa- tions) suggests that many more CNS centers harboring neu- ronal gap junctions have yet to be identified.
Investigations of gap-junctional coupling have typically utilized the difficult but direct methods of electrophysiology or electron microscopy. Recently, however, considerable knowledge has been accumulated regarding the biochemis- try of gap junctions in peripheral tissues (Revel et al., '85; Hertzberg and Spray, '85). In particular, the development of methods for the isolation of gap junctions from liver has allowed the characterization of their constituent proteins (Henderson et al., '79; Hertzberg and Gilula, '79; Hertzberg, '80, '84; Hertzberg et al., '85). Homologous 27-kD gap-junc- tion proteins are found widely distributed in peripheral tis- sues in a variety of species (Hertzberg and Skibbens, '84). Antibodies against the 27-kD liver gap-junction protein (GJP) have been successfully used for the immunohisto- chemical visualization of GJP in various tissues (Dermietzel et al., '84; Hertzberg and Skibbens, '84; Hertzberg, '85; Hertzberg and Spray, '85; Paul, '85). Immunohistochemical localization of protein components which endow cells with the potential for gap junctional communication provides a novel approach to studies concerning electrotonic transmis- sion in the CNS. In view of the numerous recent reports that have focussed attention on electrotonic coupling, dye trans- fer, gap-junctional contacts, and electrical interactions be- tween hippocampal neurons (MacVicar and Dudek, '80, '81, '82; Schmalbruch and Jahnson, '81; Andrew et al., '82; Knowles e t al., '82; MacVicar e t al., '82, Taylor and Dudek, '82; Dudek et al., '83; Kosaka, '83a,b; Kosaka and Hama, '85; MacVicar and Jahnsen, '85; Dalkara et al., '86; Rao e t al., '86; Yim et al., '86; Barnes et al., '87), we present here the distribution of GJP immunoreactivity in rat hippocampus as revealed by light microscopy. An accompanying paper contains a correlated light and electron microscope descrip- tion of GJP-immunoreactive (GJP-IR) hippocampal struc- tures. These results have been described in preliminary form elsewhere (Nagy et al., '88).
T. YAMAMOTO ET AL.
MATERIALS AND METHODS Male Sprague-Dawley rats weighing 250-300 g were
deeply anesthetized with chloral hydrate and perfused transcardially with 60 ml of an ice-cold solution containing 0.9% sodium chloride, 0.1 M sodium phosphate, pH 7.4 (PBS), 0.1% sodium nitrite, and 6G units of heparin. This was followed by 400 ml of an ice-cold solution containing 4% paraformaldehyde, 0.2% picric acid, and 0.1 M sodium phosphate, pH 7.4. Brains were removed, cut into 5-10 mm transverse blocks, and postfixed in fresh perfusion fixative for 12-18 hours at 4°C. The blocks were then transferred to an ice-cold solution containing 25 % sucrose, 10% glycerol, and 0.1 M sodium phosphate, pH 7.4, and maintained in this for 24-48 hours. Brains were quickly frozen on dry ice and
sectioned on a sliding microtome. Sections, cut at a thick- ness of 20 gm, were collected in cold PBS and washed for several hours with three or four changes of this solution. The sections were then allowed to soak overnight a t 4°C in PBS containing 0.3 % Triton X-100 (PBS-Triton). For immuno- histochemical processing, PBS-Triton was used for dilu- tions of all immunoreagents and for all washes between vari- ous incubation steps. Sections treated with PBS-Triton were incubated with affinity-purified sheep anti-GJP anti- body (protein concentration, 0.84 mg/ml; diluted 1:1,000) for 36-48 hours at 4°C. The sections were washed at room temperature for 1 hour, incubated with biotin-conjugated donkey antisheep antibody (diluted 1: lOO) (Amersham) for 1 hour at room temperature, and then incubated with horse- radish peroxidase-conjugated streptavidin (diluted 1:150) (Amersham) for 1 hour. After a 1-hour wash at room tem- perature, the sections were rinsed in 50 mM Tris-HC1, pH 7.4 (Tris buffer) for 15 minutes and placed in Tris buffer containing 0.02 % 3,3'-diaminobenzidine (DAB) and 0.005 % hydrogen peroxide for 3-8 minutes. Following several washes in Tris buffer the sections were mounted from gela- tin-ethanol solution, dehydrated in ethanol, and coverslip- ped with Permount. A series of immunostained sections col- lected at 100-pm intervals through the rostrocaudal extent of the hippocampus was taken for Nissl counterstaining in order to aid visualization of anatomical boundaries. Control sections incubated with preimmune serum or preadsorbed antibody were processed exactly as described above. For antibody absorption, affinity-purified antibody (1 mg/ml) was added to purified rat liver gap junctions (1.5 mg/ml). The mixture was incubated with continuous inversion at room temperature for 1 hour and then at 4°C overnight. The supernatant fraction was then centrifuged at 13,000 rpm for 10 minutes in a Beckman model 11 microfuge, and residual gap junctions were removed by centrifugation in a Beckman Airfuge a t 30 psi for 1 hour. The supernatant fraction from the latter centrifugation was then readsorbed by the same procedure. Preimmune serum was used a t a protein concen- tration similar to that of primary antibody. Sections of liver were included with those of brain and processed with GJP antibody in order to verify the expected pattern of GJP immunostaining in this peripheral tissue. Tissues were ex- amined and photographed by brightfield, darkfield, and dif- ferential-interference-contrast microscopy with a Leitz Dia- lux-20 microscope.
The production, affinity purification, and biochemical and immunohistochemical characterization of the anti-27- kD GJP antibody have been previously described (Hertz- berg, '84, '85; Hertzberg and Skibbens, '84; Hertzberg and Spray, '85; Nagy et al., '88). The above immunohistochemi- cal procedure was found to produce reliable immunostain- ing for GJP in CNS tissues of rats as well as in those of several other species (Nagy, unpublished observations). However, numerous procedures were tested in order to opti- mize immunostaining since GJP is an integral membrane protein and may therefore behave atypically in standard protocols used for immunohistochemical staining of cyto- plasmic antigens. In addition, since several different GJP-immunostaining patterns were evident, it was neces- sary to establish the consistency of these under a variety of conditions. We have tested immunostaining for GJP with 1) unfixed cryostat sections; 2) over five dozen prewash and fixative perfusion conditions; 3) a variety of postfixation times; 4) a wide range of primary antibody concentrations and incubation times; and 5) biotin-avidin-peroxidase, per-

GAP JUNCTION PROTEIN IN HIPPOCAMPUS 271
Fig. 1. Low-magnification photomicrographs of transverse sections show- ing the density of immunostaining for gap junction protein (GJP) in various regions of the rostral (A) and caudal (B) portions of the hippocampus. The levels depicted in A and B correspond to bregma -4.52 and -6.04, respec- tively, according to the atlas of Paxinos and Watson (‘86). Note the higher density of staining associated with the stratum pyramidale in all subregions
of the hippocampus proper and with the granule cell layer in the dentate gyrus. Also note the slightly lower staining density in area CA1 as compared with areas CA2 and CA3. Abbreviations: AH, amygdalohippocampal area; BLP, posterior part of the basolateral amygdaloid nucleus; DG, dentate gyrus; PO, polymorph layer of the dentate gyrus; S, subiculum. x25.

272 T. YAMAMOTO ET AL. With respect to immunoreactive fibers, no differences in
their distribution densities were seen in any subregions or layers rostrocaudally. However, as in the case of punctate staining, area CA1 consistently contained slightly fewer fibers than areas CA2 or CA3. Greater distinctions were seen among the various hippocampal layers. Immunostained fi- bers were rarely found in the alveus or in the outer half of the stratum lacunosum moleculare, moderately concen- trated in the stratum oriens, the stratum radiatum, and the inner half of the stratum lacunosum moleculare, and most abundant in an area surrounding the stratum pyramidale. This distribution closely paralleled that of punctate immu- nostaining. A dense plexus of fibers accumulated in, for example, the inner half of the stratum pyramidale and at the border zone between this layer and the stratum oriens (Figs. 2A,B, 3A). Some differences among layers were also seen in the orientation and average lengths of their immunoreactive fiber constituents (Fig. 3A-C). These were documented in some detail to aid future studies of fiber organization and chemistry. In the alveus the few short fibers (mean length, 10pm; range, 7-12 pm) found were oriented parallel with the layer and largely restricted to a zone near the stratum oriens, which itself contained some fibers penetrating into the alveus. In the stratum oriens, fibers oriented parallel (mean length, 52pm; range 12-150 pm) or oblique (mean length, 50 pm; range, 25-100 pm) to the layer tended to be slightly longer than those traversing it perpendicularly (mean length, 33 pm; range, 15-45 pm). In the stratum pyramidale, the longest fibers (mean length, 32 pm; range, 7.5-60 pm) were oriented parallel with the layer and the shortest (mean length, 7.5 pm; range, 5-10 pm) perpendicu- lar to it. Long fibers were most numerous in the stratum radiatum (Fig. 3B) and the longest of these were oriented either parallel or oblique to this layer (mean lengths, 56 and 78 pm, respectively; range, 15-200 pm). Some fibers within the stratum radiatum were seen penetrating into the stratum lacunosum moleculare where most fibers (mean length, 25 pm; range, 7.5-50 pm) generally ran parallel with it in area CA1 (Fig. 3C) while those in areas CA2 and CA3 were somewhat more disorganized. In the dentate gyrus, fibers in the granule cell layer tended to be short (mean length, 14 pm; range, 7.5-30 pm) and showed no preferred orientations. In the molecular layer, fibers running parallel with the layer were generally longer (mean length, 28 pm; range, 15-60 pm) than those oriented perpendicular to it (mean length, 10 bm; range, 5-25 pm). Fibers in the poly- morph layer were among the shortest (mean length, 10 pm; range, 5-17 pm) and most convoluted. Unless otherwise stated, fibers in most layers showed no predominant orien- tation. I t should be noted that these fiber lengths and orien-
-
oxidase-antiperoxidase, and immunofluorescence-based methods. The results of these tests will be described together with the distribution of GJP immunoreactivity in rat brain (Nagy, in preparation).
RESULTS In the following description of our observations, hippo-
campus proper is defined as the hippocampus excluding the dentate gyrus. Directions within various layers viewed in transverse sections are defined as outer toward the hippo- campal fissure and inner toward the alveus or hilus. Direc- tions in the longitudinal septa1 and temporal axis are referred to as rostra1 or caudal, respectively, and orienta- tions within the various layers refer to stratification within the transverse plane. Some results are shown by both con- ventional interference-contrast microscopy, which better conveys detail, and by darkfield microscopy, which more readily conveys differences in immunostaining density. The overall distribution and density of immunostaining in the hippocampus and, for comparison, the posterior basolateral amygdaloid nucleus (BLP) are shown at two transverse lev- els in Figure 1. The two most prominent GJP-immunostain- ing patterns observed were immunoreactive punctate ele- ments and fibers. These were seen in all subregions of the hippocampus proper including areas CA1, CA2, and CA3, in all areas of the dentate gyrus, and in all layers of these regions. However, there were considerable regional varia- tions in their densities and in the organization of fiber net- works.
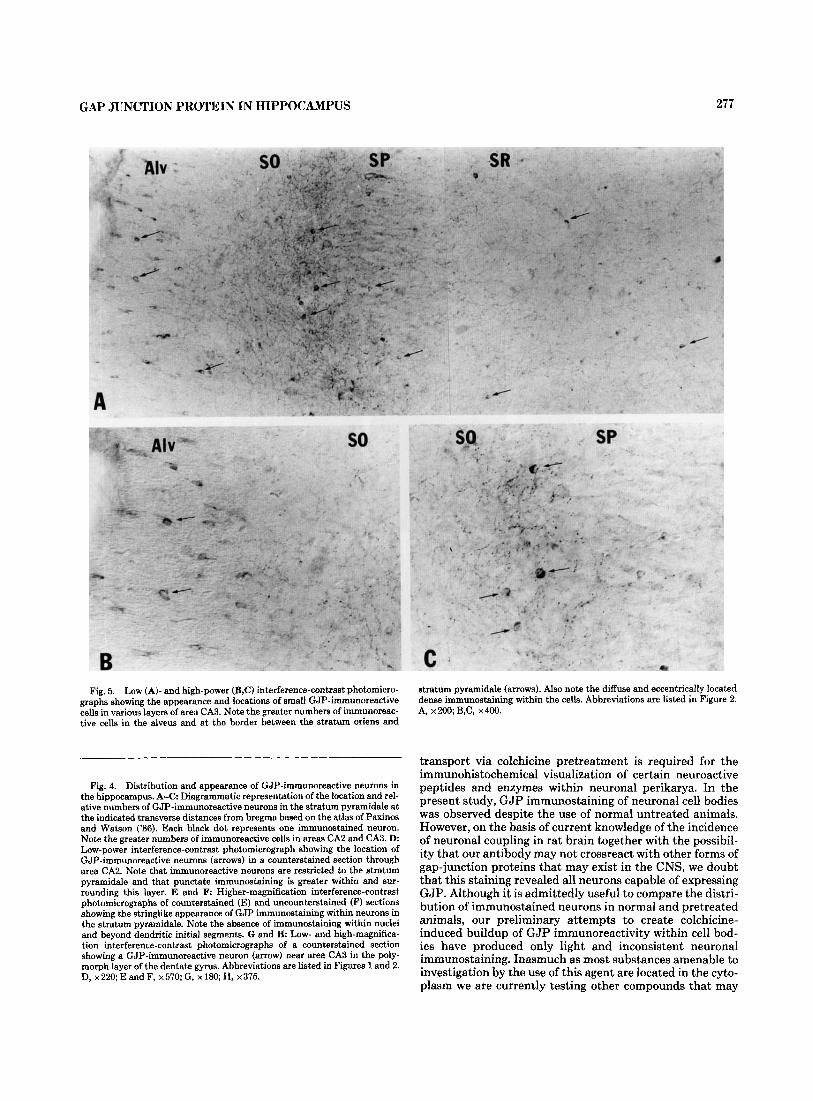
With respect to the punctate pattern, immunostaining was somewhat more widely and uniformly distributed than that of fibers. In the hippocampus proper, the density was lowest in the alveus and outer two-thirds of the stratum lacunosum moleculare, moderate in the stratum radiatum and oriens, and consistently highest in the outer two-thirds of the stratum pyramidale and at the border between this layer and the stratum oriens. The inner part of the stratum pyramidale contained distinctly less immunoreactivity (Fig. 2A,B). No rostrocaudal differences in staining density were evident within any subregions or layers of the hippocampus. Among subregions, however, punctate immunostaining den- sity in all layers of areas CA2 and CA3 was moderately greater than that in area CA1 (Fig. 1). Within the dentate gyrus, staining density was lowest in the inner quarter of the molecular layer, moderate in the rest of the molecular layer and in the polymorph layer, and highest in the granule cell layer and particularly in the outer third of this layer. This distribution produced a striking contrast in density at the border between the granule cell and molecular layer (Fig. 2C,D). No deviation from this regional distribution was seen in the rostrocaudal axis of the dentate gyrus; at anterior lev- els, however, staining density did appear to be slightly greater in the dorsal than in the ventral blade of the granule cell layer. The morphology of the GJP-irnmunoreactive punctate structures was relatively uniform in all areas of the hippocampus proper and dentate gyrus. These ranged from 0.3 to 0.7 pm in size and usually had a round or slightly oval appearance. Although this type of staining was seen hetero- geneously distributed in several other regions of the brain, it is noteworthy that the density in the hippocampus was con- siderably less than seen in some of these regions. An exam- ple of this is shown by the appearance of immunostaining in the posterior basolateral amygdaloid nucleus (Fig. 2E,F) where the density of punctate staining appears to outline its nuclear boundaries.
Fig. 2. A and B Darkfield (A) and interference-contrast (B) photomicro- graphs of the same area showing the density of fibrous and punctate GJP immunostaining in a section through area CA2 of the hippocampus. Note the higher density of staining within and surrounding the stratum pyramidale. C and D Darkfield (C) and interference-contrast (D) photomicrographs of the same area showing the distribution of GJP immunostaining in the rostrd- medial part of the dentate gyrus. Note the greater density of staining at the outer border of the granule cell layer (GL) (arrowheads) and the lower density in the inner zone of the molecular layer. E and F Darkfield (E) and interfer- ence-contrast (F) photomicrographs of the same area showing the appearance of GJP immunoreactivity within the BLP. Punctate immunostaining density is greater and generally of a finer grain (shown at higher magnification, inset in F) than seen in hippocampal areas. Abbreviations: GL, granule cell layer of the dentate gyms; ML, molecular layer of the dentate gyrus; Alv, alveus; SO, stratum oriens; SP, stratum pyramidale; SR, stratum radiatum. A-D, x 190; E,F, x 95; inset in F, x 340.

GAP JUNCTION PROTEIN IN HIPPOCAMPUS 273
Figure 2

274 T. YAMAMOTO ET AL.
Figure 3

GAP JUNCTION PROTEIN IN HIPPOCAMPUS 275
tations refer to those seen in transverse planes of section; different results may emerge in other planes, which we did not examine.
Some detailed features of GJP-immunoreactive fibers are shown in Figure 3D-F. On rare occasions fibers were seen giving off collaterals (Fig. 3D). Such points of collateraliza- tion, however, were difficult to identify with confidence because of light immunostaining of intervaricose segments. In addition, attempts to quantify these were confounded by the high level of punctate immunostaining. Similar difficul- ties prevented thorough documentation of possible sites of fiber contacts. Estimation of the frequency with which two immunoreactive fibers made close encounters of the parallel kind as compared with their random crossings may be sug- gestive of such contacts. Only a few examples of two fibers exhibiting this configuration were found after excluding those located in slightly different planes of focus (Fig. 3E). An interesting feature of immunoreactive fibers in both the stratum pyramidale (Fig. 3F) and granule cell layer was their frequent localization around neuronal somata. Such fibers may be axons making somatic contacts (unpublished observations). Throughout the hippocampus and dentate gyrus, varicosity sizes ranged from 0.5 to 1.7 pm (mean, 1.0 pm) and intervaricose distances from 0.3 to 2.8 pm (mean, 1.8 pm). Distinct classes of immunostained fibers could not be recognized since only a single broad continuum of param- eters such as staining intensity, fiber caliber, varicosity size, intervaricosity distance, and fiber lengths were observed. Nevertheless, the wide variation in these parameters sug- gests that the fibers may originate from more than one and probably several different cellular sources.
In marked contrast to the high density of GJP-immunore- active fibers and punctate elements seen in the hippocam- pus, GJP-immunoreactive neuronal cell bodies were rela- tively rare. Despite their small numbers, these were readily recognizable by virtue of their characteristic pattern and intensity of immunostaining. Their distribution in the hip- pocampus is shown schematically in Figure 4A-C. Two con- sistent features were evident regarding their localization in the hippocampus proper. First, areas CA2 and CA3 con- tained the most GJP-immunoreactive neurons, about zero to 12 per 20-pm section, and area CAI contained zero or two per section. Second, the neurons were located exclusively either within the stratum pyramidale or occasionally at the border between the stratum pyramidale and stratum oriens (Fig. 4D-F). These neurons usually had oval to fusiform shapes with major axes ranging from 18 to 27 pm (mean, 23 pm) and minor axes ranging from 8 to 17 pm (mean 15 pm). No spherical GJP-immunoreactive neurons were observed. Within the cells, GJP immunoreactivity was restricted to slender tortuous strands of various lengths (Fig. 4E,F).
Fig. 3. A-C: Interference-contrast photomicrographs showing the relative density and configurations of GJP-immunoreactive fibers in the stratum oriens just above (with respect to orientation of the micrograph) the stratum pyramidale (A), the stratum radiatum (B), and stratum lacunosum moleculare (C) of area CA1. D-F High-magnification photo- micrographs of the appearance of GJP-immunoreactive fibers in various hippocampal layers. D A fiber is shown giving off a collateral (arrow- head) in the stratum radiatum. E: Varicose portions of two fibers are shown in close proximity to each other in the stratum radiatum (arrow- head). Note varicose appearance of fibers and immunostaining of inter- varicose segments (arrows). F: A varicose GJP-immunoreactive fiber is shown circumventing a neuronal cell body in the stratum pyramidale (arrows; counterstained section). Abbreviations: SR, stratum radiatum; SLM, stratum lacunosum moleculare. A-C, x580; D-F, x 1,430.
These strands were oriented mainly parallel to the major axis of the neurons and extended to the proximal processes. The major axis of the cells appeared to be oriented perpen- dicular, or very nearly so, to the stratum pyramidale. Within the dentate gyrus, the presence of GJP-immunoreactive neurons having the appearance of those in the hippocampus proper was more variable and inconsistent among animals, thus precluding any firm conclusions regarding their rela- tive numbers and distribution. In some animals, a few such neurons were seen in the polymorph layer near the loosely packed cells of area CA3 (Fig. 4G,H) and in the granule cell layer.
Small GJP-immunoreactive cells were heterogeneously distributed throughout the hippocampus proper (Fig. 5A) and the dentate gyrus (not shown). These were generally oval or spherical and had diameters ranging from 5 to 7 pm. Oval cells were most numerous in the alveus (Fig. 5B) and stratum lacunosum moleculare. Spherical cells were most abundant in the inner portion of the stratum pyramidale and at the border between this layer and the stratum oriens (Fig. 5C). Fewer cells of either shape were seen in the outer part of the stratum pyramidale and in the stratum radia- tum, and they were least evident in the stratum oriens. On the basis of their size and sparse cytoplasmic areas, these cells were likely astrocytes and oligodendrocytes. Processes extending from these cells could not be visualized in the present material.
The results of the preabsorbed antibody and preimmune serum controls are shown in Figure 6B and C by brightfield and in 6E-F by darkfield microscopy, respectively. Preab- sorption of antibody substantially reduced but did not totally eliminate immunostaining in both brain (Fig. 6B,E) and liver. The patterns of the staining which remained in both tissues were qualitatively similar to that obtained with unabsorbed antibody. We attribute the persistence of some staining to incomplete blockade of primary antibody, and to the sensitivity of immunohistochemical detection by immu- noperoxidase methods. No punctate, fibrous, or neuronal cell body staining was seen with preimmune serum (Fig. 6C,F). The typical pattern of GJP immunoreactivity ob- tained in liver is shown in Figure 6G. Consistent with previ- ous reports using immunofluorescence, punctate reaction product is seen surrounding the borders of cells. No signifi- cant intracellular staining is observed.
DISCUSSION The morphological patterns of GJP immunoreactivity ob-
served in the hippocampus appear deceptively straightfor- ward; immunostaining is restricted to a particular class of neuronal perikarya, to a network of fibers, to a large number of dispersed punctate elements, and to some small cells. A high level of complexity becomes evident, however, upon consideration of some interpretational and technical limita- tions imposed by the nature of the antigen under investiga- tion. In addition, conclusions concerning structural or func- tional relationships are made difficult by our current lack of information regarding 1) the anatomical origin of GJP- immunoreactive hippocampal fibers and the extent to which these represent axons or dendrites, 2) the axonal projections or arborizations of GJP-immunoreactive cell bodies, and 3) the identity and cellular origin of GJP-immunoreactive punctate elements.
Several points regarding technical uncertainties are as follows. First, it is well known that inhibition of axonal
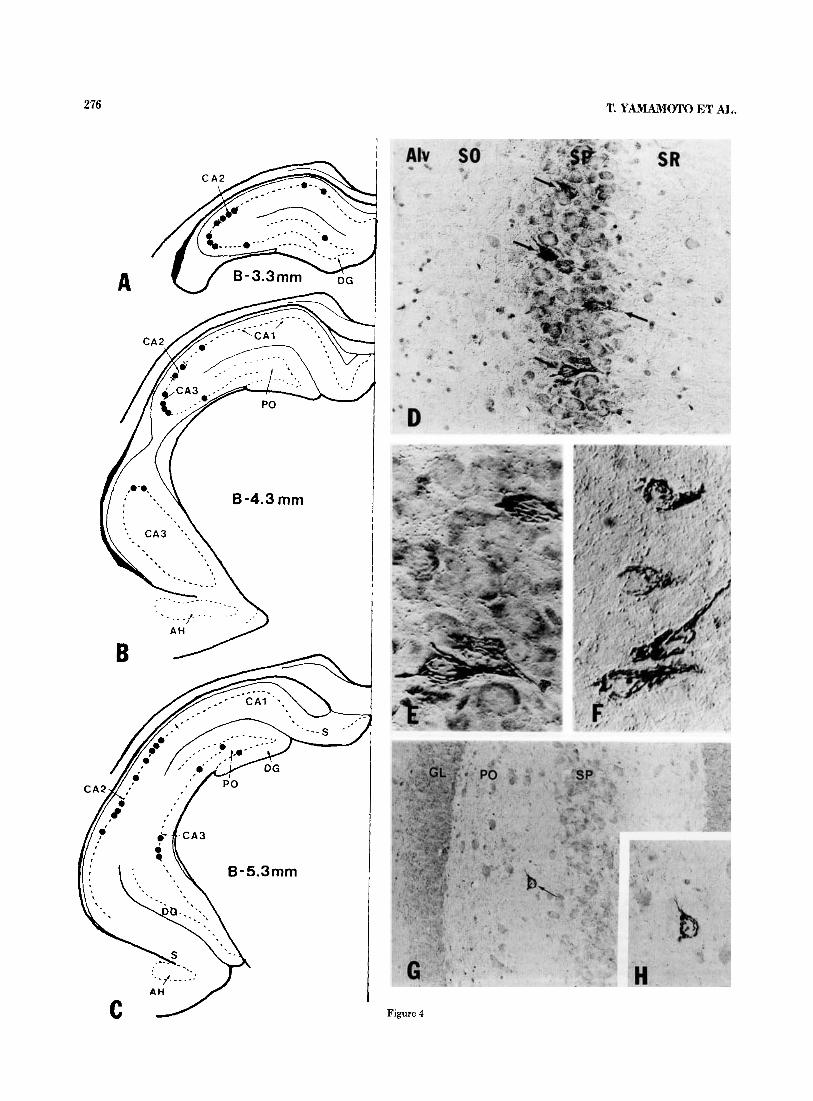
276 T. YAMAMOTO ET AL,.
Figure 4
GAP JUNCTION PROTEIN IN HIPPOCAMPUS 277
Fig. 5. Low (A)- and high-power (B,C) interference-contrast photomicro- graphs showing the appearance and locations of small GJP-immunoreactive cells in various layers of area CA3. Note the greater numbers of immunoreac- tive cells in the alveus and at the border between the stratum oriens and
Fig. 4. Distribution and appearance of GJP-immunoreactive neurons in the hippocampus. A-C Diagrammatic representation of the location and rel- ative numbers of GJP-immunoreactive neurons in the stratum pyramidale at the indicated transverse distances from bregma based on the atlas of Paxinos and Watson ('86). Each black dot represents one immunostained neuron. Note the greater numbers of immunoreactive cells in areas CA2 and CA3. D. Low-power interference-contrast photomicrograph showing the location of GJP-immunoreactive neurons (arrows) in a counterstained section through area CA2. Note that immunoreactive neurons are restricted to the stratum pyramidale and that punctate immunostaining is greater within and sur- rounding this layer. E and F: Higher-magnification interference-contrast photomicrographs of counterstained (E) and uncounterstained (F) sections showing the stringlike appearance of GJP immunostaining within neurons in the stratum pyramidale. Note the absence of immunostaining within nuclei and beyond dendritic initial segments. G and H: Low- and high-magnifica- tion interference-contrast photomicrographs of a counterstained section showing a GJP-immunoreactive neuron (arrow) near area CA3 in the poly- morph layer of the dentate gyrus. Abbreviations are listed in Figures 1 and 2. D,x220;EandF,x570;G,x180;H, x375.
stratum pyramidale (arrows). Also note the diffuse and eccentrically located dense immunostaining within the cells. Abbreviations are listed in Figure 2. A, x200;B,C, x400.
transport via colchicine pretreatment is required for the immunohistochemical visualization of certain neuroactive peptides and enzymes within neuronal perikarya. In the present study, GJP immunostaining of neuronal cell bodies was observed despite the use of normal untreated animals. However, on the basis of current knowledge of the incidence of neuronal coupling in rat brain together with the possibil- ity that our antibody may not crossreact with other forms of gap-junction proteins that may exist in the CNS, we doubt that this staining revealed all neurons capable of expressing GJP. Although it is admittedly useful to compare the distri- bution of immunostained neurons in normal and pretreated animals, our preliminary attempts to create colchicine- induced buildup of GJP immunoreactivity within cell bod- ies have produced only light and inconsistent neuronal imrnunostaining. Inasmuch as most substances amenable to investigation by the use of this agent are located in the cyto- plasm we are currently testing other compounds that may
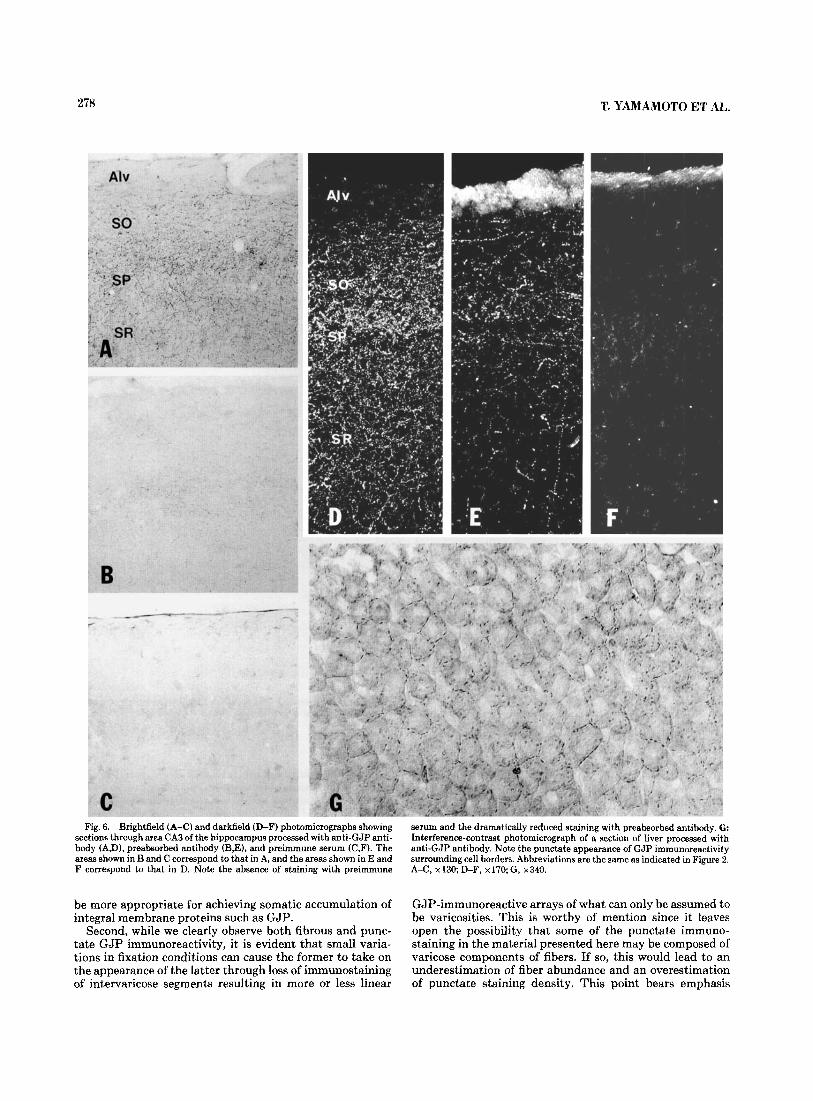
278 T. YAMAMOTO E:T AL.
Fig. 6. Brightfield (A-C) and darkfield (D-F) photomicrographs showing section8 through area CA3 of the hippocampus processed with anti-GJP anti- body (A,D), preabsorbed antibody (B,E), and preimmune serum (C,F). The areas shown in B and C correspond to that in A, and the areas shown in E and F correspond to that in D. Note the absence of staining with preimmune
serum and the dramatically reduced staining with preabsorbed antibody. G Interference-contrast photomicrograph of a section of liver processed with anti-GJP antibody. Note the punctate appearance of GJP immunoreactivity surrounding cell borders. Abbreviations are the same as indicated in Figure 2. A-C, x130; D-F, x170;G, x340.
be more appropriate for achieving somatic accumulation of integral membrane proteins such as GJP.
Second, while we clearly observe both fibrous and punc- tate GJP immunoreactivity, it is evident that small varia- tions in fixation conditions can cause the former to take on the appearance of the latter through loss of immunostaining of intervaricose segments resulting in more or less linear
GJP-immunoreactive arrays of what can only be assumed to be varicosities. This is worthy of mention since it leaves open the possibility that some of the punctate immuno- staining in the material presented here may be composed of varicose components of fibers. If so, this would lead to an underestimation of fiber abundance and an overestimation of punctate staining density. This point bears emphasis

GAP JUNCTION PROTEIN IN HIPPOCAMPUS
since we cannot discern the cellular or subcellular elements represented by purely punctate staining at the light micro- scope level. On the basis of results in the accompanying elec- tron microscope study (Shiosaka et al., '88), punctate stain- ing may be localized to gap junctions between neuronal and/ or between glial elements, GJP-immunoreactive terminal boutons, or portions of glial processes. In contrast, GJP- immunoreactive fibers seen at the light microscope level can be categorized eventually, if not in the present study, as dendritic or axonal. Since the sizes of the largest gap junc- tions we observe approach those of the smallest varicosities, at least in the hippocampus, though perhaps not in all brain areas, it is desirable to minimize confusion between these two different morphological entities by choosing fixation conditions most suitable for revealing continuous immuno- staining of fibers.
A third point concerns the validity of any assumption that the presence of GJP immunoreactivity within a cell neces- sarily endows it with the capability to form gap junctions. Intracellular immunostaining for GJP is often seen in junc- tionally coupled neuronal elements and glial processes (Shiosaka et al., '88). Nevertheless, this may be an unsafe assu nption a t our present stage of knowledge regarding GJP in at least CNS tissues; the functional significance of, for example, GJP-immunoreactive vesicles within axons and atypical gap junctional structures which have been observed in the hippocampus remains to be determined (Shiosaka et al., '88). Thus, although present evidence would strongly suggest that the antibody employed here detects a 27-kD GJP in rat brain (Nagy et al., '88) and that the physiological actions mediated by gap junctions are functionally dependent on and structurally supported by GJPs (Peracchia, '80; Hertzberg et al., '81, '85; Loewenstein, '81; Warner et al., '84; Spray et al., '86; Dahl e t al., '87; Ding-E Young et al., '87; Dudek et al., '88), the converse may not necessarily hold; i t is conceivable that GJP or a very similar immunologically crossreacting protein may have functions associated with, but not confined to, gap junc- tions.
Our observations of GJP-immunoreactive neuronal cell bodies in the stratum pyramidale are consistent with reports by Kosaka ('83a) and Kosaka and Hama ('85), who found gap junctions between dendrites originating from neuronal perikarya located within or straddling this cell layer. Unfortunately, the dendritic morphology and arbori- zations of these cells could not be deduced since GJP immu- nostaining did not extend very far into their initial dendritic segments. However, as discussed in the accompanying paper (Shiosaka et al., '88), we suspect that they are nonpyramidal cells. This would be in keeping with the varicose nature of nonpyramidal cell dendrites which form gap junctions (Ko- saka and Hama, '85) and the possibility that some GJP- immunoreactive varicose fibers identified here represent dendritic processes. The presence of a few GJP-immunore- active neuronal somas in the polymorph layer of the dentate gyrus is also in agreement with observations of gap-junc- tionally connected neurons in this region (Kosaka, '83b). Although the appearance of immunostaining within hippo- campal neuronal cell bodies is highly unusual, this staining is consistent with the localization of GJP in the Golgi appa- ratus of these cells (Shiosaka et al., '88). To date we have found GJP-immmunoreactive neuronal perikarya having this appearance in only a few other brain areas, a finding that makes their hippocampal localization all the more dis- tinctive. Since none of these other areas includes sites
279
known to have projections to the hippocampus, it seems rea- sonable to consider that the GJP-immunoreactive fibers within this structure originate from local GJP-positive neu- rons. However, since a fair proportion of these immunoreac- tive fibers are axons (Shiosaka et al., '88), we cannot exclude the possibility that some or all of the axonal contingent of GJP-positive hippocampal fibers originate from extrinsic neurons which fail to stain for GJP. The presence of GJP- immunoreactive fiber networks in area CA1, despite the very few GJP-immunoreactive neurons observed in this region, also suggests that these networks may not arise exclusively from local GJP-immunoreactive neurons.
Although speculative, the disproportionate distribution of GJP-immunoreactive neurons among hippocampal sub- regions may reflect the relative density of a t least one type of neuronal gap junction in these regions. Based on the fewer GJP-immunoreactive neurons and generally lower im- munostaining densities in area CA1 than either area CA2 or CA3, it might be predicted that many more dendrodendritic gap junctions of the kind described by Kosaka and Hama ('85) may be found in the latter two regions. This may have some bearing on the relative contribution of electrotonic coupling to functional interactions between neurons in the various hippocampal subdivisions. A related point concerns the observation that the densities of the four immunostain- ing patterns described parallel each other in some of the layers of the hippocampus proper. This was most striking in the region straddling the stratum pyramidale and oriens where GJP-immunoreactive punctate, fibrous, neuronal, and small cell body immunostaining were all most abun- dant. If both glial and neuronal junctions are heavily con- centrated within this region, then this restricted area may represent an example of not only a heterogeneous distri- bution of glial junctions but also of a distribution of these paralleling that of neuronal junctions. The functional signif- icance of this potential codistribution is open to specula- tion.
A well-known property of the hippocampus is its propen- sity for epileptiform activity. In addition, certain popula- tions of hippocampal neurons are known to be predisposed to abnormal synchronous electrical discharge (Kandel and Spencer, '61; Dudek et al., '83; Prince, '83; Schwartzkroin, '83). A major focus of investigations for many years has been elucidation of the anatomical connections and cellular mechanisms that are responsible for the manifestation of these properties. Although the possibility has been consid- ered that electrotonic coupling between hippocampal neu- rons may contribute to the synchronization of their dis- charge (Dudek et al., '83; Schwartzkroin, '83; Traub et al., '85a,b) in a manner analogous to the synchronous activity found in other coupled neuronal networks (Bennett, '74), the relative importance of this as compared with other synchronizing mechanisms remains uncertain. While the present study provides little clarification of this issue, it does suggest that there is ample opportunity for electrotonic interaction between GJP-immunoreactive hippocampal neuronal elements.
ACKNOWLEDGMENTS The authors thank Lyn Polson for typing the manuscript
and Lyn Polson and Barbaro Tinner for excellent technical assistance. This work was supported by grants from the Manitoba Health Research Council (MHRC), the Univer- sity of Manitoba Medical Faculty Fund, the Canadian Med-

280
ical Research Council (MRC), and the National Scienee and Engineering Research Council (NSERC) to J.I. Nagy and from the National Institute of Health (NIH) (GM 30667) to E.L. Hertzberg. Fellowship support to S. Shiosaka and T. Yamamoto was provided by the MHRC and the Faculty Fund. M.E. Whittaker was supported by the University of Manitoba BSc./M.D. program. J.I. Nagy is a Scholar of the MRC and E.L. Hertzberg is a recipient of a Research Career Development Award (HD 00713) from the NIH.
LITERATURE CITED Andrew, R.D., B.A. MacVicar, F.E. Dudek, and G.I. Hatton (1981) Dye trans-
fer through gap junctions between neuroendocrine cells of rat hypothala- mus. Science 211:1187-1189.
Andrew, R.D., C.P. Taylor, R.W. Snow, and F.E. Dudek (1982) Coupling in rat hippocampal slices: Dye transfer between CA1 pyramidal cells. Brain Res. Bull. 8211-222.
Barnes, C.A., G. Rao, and B.L. McNaughton (1987) Increased electrotonic coupling in aged rat hippocampus: A possible mechanism for cellular excitability changes. J. Comp. Neurol. 259:549-558.
Bennett, M.V.L. (1972) A comparison of electrically and chemically mediated transmission. In G.D. Pappas and D.P. Purpura (eds): Structure and Function of Synapses. New York Raven Press, pp. 221-256.
Bennett, M.V.L. (1974) Flexibility and rigidity in electrotonically coupled systems. In M.V.L. Bennett (ed): Synoptic Transmission and Neuronal Interaction. New York Raven Press, pp. 153-178.
Bennett, M.V.L., and D.A. Goodenough (1975) Gap junctions, electrotonic coupling and intercellular communication. Neurosci. Res. Prog. Bull. 16:371-486c.
Bennett, M.V.L. (1977) Electrical transmission: A functional analysis and comparison with chemical transmission. In E.R. Kandel (ed): Handbook of Physiology, Section I: The Nervous System, Vol. 1, Cellular Biology of Neurons, Part 1. Bethesda, Maryland American Physiological Society, pp. 357-416.
Cobbett, P., K.G. Smithson, and G.I. Hatton (1985) Dye-coupled magnocellu- lar peptidergic neurons of the rat paraventricular nucleus show homo- typic immunoreactivity. Neuroscience 16.S85-895.
Dahl, G., T. Miller, D. Paul, R. Voellmy, and R.L. Werner (1987) Expression of functional cell-cell channels from cloned rat liver gap junction comple- mentary DNA. Science 236r129LL-1293.
Dalkara, T., K. Krnjevic, N. Ropert, and C.Y. Yim (1986) Chemical modula- tion of ephaptic activation of CA3 hippocampal pyramids. Neuroscience 17:361-370.
Dermietzel, R., A. Leibstein, U. Frixen, U. Janssen-Timmen, 0. Traub, and K. Willecke (1984) Gap junctions in several tissues share antigenic deter- minants with liver gap junctions. EMBO J. 3:2261-2270.
Ding-E Young, J., Z.A. Cohn, and N.B. Gilula (1987) Functional assembly of gap junction conductance in lipid bilayers: Demonstration that the major 27 kD protein forms the junctional channel. Cell 48:733-743.
Dudek, F.E., R.D. Andrew, B.A. MacVicar, R.W. Snow, and C.P. Taylor (1983) Recent evidence for and possible significance of gap junctions and electrotonic synapses in the mammalian brain. In H.H. Jasper and N.M. van Gelder (eds): Basic Mechanisms of Neuronal Hyperexcitability. New York: Alan R. Liss, Inc., pp. 31-73.
Dudek, F.E., V.K. Gribkoff, J.E. Olson, and E.L. Hertzberg (1988) Reduction of dye coupling in glial cultures by microinjection of antibodies against the liver gap junction polypeptide. Brain Res. 439:275-280.
Grace, A.A., and B.S. Bunney (1983) Intracellular and extracellular electro- physiology of nigral dopaminergic neurons-3. Evidence for electrotonic coupling. Neuroscience 10:333-348.
Henderson, D., H. Eibl, and K. Weber (1979) Structure and biochemistry of mouse hepatic gap junctions. J. Mol. Biol. 132193-218.
Hertzberg, E.L., and N.B. Gilula (1979) Isolation and characterization of gap junctions from rat liver. J. Biol. Chem. 254:213&2147.
Hertzberg, E.L. (1980) Biochemical and immunological approaches to the study of gap junctional communication. In Vitro 16t1057-1067.
Hertzberg, E.L., T.S. Lawrence, and N.B. Gilula (1981) Gap junctional com- munication. Annu. Rev. Physiol. 43:479-491.
Hertzberg, E.L. (1984) A detergent-independent procedure for the isolation of gap junctions from rat liver. J. Biol. Chem. 259:99369943.
Hertzberg, E.L., and R.V. Skibbens (1984) A protein homologous to the 27,000 dalton liver gap junction protein is present in a wide variety of spe- cies and tissues. Cell 39.649.
T. YAMAMOTO ET AL.
Hertzberg, E.L. (1985) Antibody probes in the study of gap junctional com- munication. Annu. Rev. Physiol. 47:305-318.
Hertzberg, E.L., and D.C. Spray (1985) Studies of gap junctions: Biochemical analysis and use of antibody probes. In M.V.L. Bennett and D.C. Spray (eds): Gap Junctions. New York: Cold Spring Harbor, pp. 57-65.
Hertzberg, E.L., D.C. Spray, and M.V.L. Bennett (1985) Reduction of gap junctional conductance by microinjection of antibodies against the 27- !&a liver gap junction polypeptide. Proc. Natl. Acad. Sci. USA 822412- 2416.
Kandel, E.R., and W.A. Spencer (1961) Excitation and inhibition of single pyramidal cells during hippocampal seizure. Exp. Neurol. 4:162-179.
Knowles, W.D., P.G. Funch, and P.A. Schwartzkroin (1982) Electrotonic and dye coupling in hippocampal CA1 pyramidal cells in vitro. Neuroscience 7t1713-1722.
Kosaka, T. (1983a) Gap junctions between non-pyramidal cell dendrites in the rat hippocampus (CA1 and CA3 regions). Brain Res. 271:157-161.
Kosaka, T. (1983b) Neuronal gap junctions in the polymorph layer of the rat dentate gyrus. Brain Res. 277:347-351.
Kosaka, T., and K. Hams (1985) Gap junctions between non-pyramidal cell dendrites in the rat hippocampus (CA1 and CA3 regions): A combined Golgi-electron microscopy study. J. Comp. Neurol. 231:150-161.
Llinh, R.R. (1985) Electrotonic transmission in the mammalian central ner- vous system. In M.V.L. Bennett and D.C. Spray (eds): Gap Junctions. New York Cold Spring Harbor, pp. 337-353.
Loewenstein, W.R. (1981) Junctional intercellular communication: The cell- to-cell membrane channel. Physiol. Rev. 61t829-913.
MacVicar, B.A., and F.E. Dudek (1980) Dye-coupling between CA3 pyrami- dal cells in slices of rat hippocampus. Brain Res. 196t494-497.
MacVicar, B.A., and F.E. Dudek (1981) Electrotonic coupling between pyramidal cells: A direct demonstration in rat hippocampal slices. Science 213:782-785.
MacVicar, B.A., and F.E. Dudek (1982) Electrotonic coupling between gran- ule cells of rat dentate gyrus: Physiological and anatomical evidence. J. Neurophysiol. 47:579-592.
MacVicar, B.A., N. Ropert, and K. Krnjevic (1982) Dye-coupling between pyramidal cells of rat hippocampus in vivo. Brain Res. 238.239-244,
MacVicar, B.A., and H. Jahnsen (1985) Uncoupling of CA3 pyramidal neu- rons by proprionate. Brain Res. 33Ot141-145.
Nagy, J.I., T. Yamamoto, S. Shiosaka, K.M. Dewar, M.E. Whittaker, and E.L. Hertzberg (1988) Immunohistochemical localization of gap junction pro- tein in rat CNS: A preliminary account. In E.L. Hertzberg and R. Johnson (eds): Gap Junctions. New York Alan R. Liss, Inc., pp. 375-389.
Paul, D.L. (1985) Antibody against liver gap junction 27-KD protein is tissue specific and cross-reacts with a 54-KD protein. In M.V.L. Bennett and D.C. Spray (eds): Gap Junctions. New York Cold Spring Harbor, pp. 107-122.
Paxinos, G., and C. Watson(1986) The Rat Brain in Stereotaxic Coordinates. New York Academic Press.
Peracchia, C. (1980) Structural correlates of gap junction permeation. Int. Rev. Cytol. 66:81-146.
Prince, D.A. (1983) Mechanisms of epileptogenesis in brain-slice model sys- tems. In A.A. Ward, Jr., J.K. Penry, and D. Purpura (eds): Epilepsy. New York Raven Press, pp. 29-52.
Rao, G., C.A. Barnes, and B.L. McNaughton (1986) Intracellular fluorescent staining with carboxyfluorescein: A rapid and reliable method for quan- tifying dye-coupling in mammalian central nervous system. J. Neurosci. Methods 16.251-263.
Revel, J.-P., B.J. Nicholson, and S.B. Yancey (1985) Chemistry of gap junc- tions. Annu. Rev. Physiol. 47263-279.
Schmalbruch, H., and H. Jahnsen (1981) Gap junctions on CA3 pyramidal cells of guinea pig hippocampus shown by freeze-fracture. Brain Res. 217:175-178.
Schwartzkroin, P.A. (1983) Local circuit considerations and intrinsic nen- ronal properties involved in hyperexcitability and cell synchronization. In H.H. Jasper and N.M. van Gelder (eds): Basic Mechanisms of Neuronal Hyperexcitability. New York Alan R. Liss, Inc., pp. 75-108.
Senha, E., P.E. Daddona, T. Watanabe, J.-Y. Wu, and J.I. Nagy (1985) Coex- istence of adenosine deaminase, histidine decarboxylase, and glutamate decarboxylase in hypothalamic neurons of the rat. J. Neurosci. 5:3393- 3402.
Shiosaka, S., T. Yamamoto, E.L. Hertzberg, and J.1. Nagy (1988) Gap junc- tion protein in rat hippocampus: Correlative light and electron micro- scope immunohistochemical localization. J. Comp. Neurol. 280.W-000.
Sotelo, C., and H. Korn (1978) Morphological correlates of electrical and

GAP JUNCTION PROTEIN IN HIPPOCAMPUS 281
Traub, R.D., F.E. Dudek, C.P. Taylor, and W.D. Knowles (1985a) Simulation of hippocampal after discharges synchronized by electrical interactions. Neuroscience 14:1033-1038.
Traub, R.D., F.E. Dudek, R.W. Snow, and W.D. Knowles (1985b) Computer simulations indicate that electrical field effects contribute to the shape of the epileptiform field potential. Neuroscience 15:947-958.
Warner, A.E., S.C. Guthrie, and N.B. Gilula (1984) Antibodies to gap-junc- tional protein selectively disrupt junctional communication in the early amphibian embryo. Nature 311t127-131.
Yim, C.C., K. Krnjevic, and T. Dalkara (1986) Ephaptically generated poten- tials in CA1 neurons of rat’s hippocampus in situ. J. Neurophysiol. 5699- 122.
other interactions through low-resistance pathways between neurons of the vertebrate central nervous system. Int. Rev. Cytol. 55:67-107.
Sotelo, C., and A. Triller (1981) Morphological correlates of electrical, chemi- cal and dual modes of transmission. In L. Stjame, P. Hedquist, H. Lager- crantz, and A. Wennmaln (eds): Chemical Neurotransmission 75 Years. London: Academic Press, pp. 13-28.
Spray, D.C., J.C. Saez, D. Brosius, M.V.L. Bennett, and E.L. Hertzberg (1986) Isolated liver gap junctions: Gating of transjunctional currents is similar to that in intact pairs of rat hepatocytes. Proc. Natl. Acad. Sci. USA 83:5494-5497.
Taylor, C.P., and F.E. Dudek (1982) A physiological test for electrotonic cou- pling between CA1 pyramidal cells in rat hippocampal slices. Brain Res. 235:351-357.