foxo32/2 mice demonstrate reduced numbers of pre-b and
TRANSCRIPT

International Immunology, Vol. 21, No. 7, pp. 831–842doi:10.1093/intimm/dxp049
ª The Japanese Society for Immunology. 2009. All rights reserved.For permissions, please e-mail: [email protected]
Foxo32/2 mice demonstrate reduced numbers ofpre-B and recirculating B cells but normal splenicB cell sub-population distribution
Rochelle M. Hinman1, Whitney A. Nichols1, Tracy M. Diaz1, Teresa D. Gallardo2, Diego H. Castrillon2
and Anne B. Satterthwaite1
1Department of Internal Medicine, Division of Rheumatology, The University of Texas Southwestern Medical Center, Dallas,TX 75390, USA2Department of Pathology and Simmons Comprehensive Cancer Center, The University of Texas Southwestern Medical Center,Dallas, TX 75390, USA
Keywords: B cells, cell activation, knockout mice, signal transduction, transcription factors
Abstract
B cell antigen receptor (BCR) cross-linking promotes proliferation and survival of mature B cells.Phosphoinositide-3-kinase-mediated down-regulation of pro-apoptotic and anti-mitogenic genessuch as the Foxo family of transcription factors is an important component of this process.Previously, we demonstrated that BCR signaling decreases expression of transcripts for Foxo1,Foxo3 and Foxo4. We now show that BCR-induced down-regulation of Foxo3 and Foxo4 mRNAexpression occurs via distinct mechanisms from those established for Foxo1. While Foxo1, Foxo3and Foxo4 bind the same DNA sequence, the differential control of their expression upon B cellactivation suggests that they may have unique functions in the B lineage. To begin to address thisissue, we evaluated B cell development and function in Foxo32/2 mice. No effect of Foxo3 deficiencywas observed with respect to the following parameters in the splenic B cell compartment: sub-population distribution, proliferation, in vitro differentiation and expression of the Foxo target genescyclin G2 and B cell translocation gene 1. However, Foxo32/2 mice demonstrated increased basallevels of IgG2a, IgG3 and IgA. A significant reduction in pre-B cell numbers was also observed inFoxo32/2 bone marrow. Finally, recirculating B cells in the bone marrow and peripheral blood weredecreased in Foxo32/2 mice, perhaps due to lower than normal expression of receptor forsphingosine-1 phosphate, which mediates egress from lymphoid organs. Thus, Foxo3 makesa unique contribution to B cell development, B cell localization and control of Ig levels.
Introduction
Within the immune system, the maintenance of homeostasisrelies on the interplay between survival, proliferation and
death-inducing factors. The number of T and B cells that en-
ter the periphery is only a fraction of the total initially gener-
ated. Programmed cell death eliminates lymphocytes withnon-functional or autoreactive antigen receptors. T and
B cells circulating in the periphery must be kept quiescent,
only to divide and differentiate if/when they encounter theircognate antigen. These activated cells later undergo apo-
ptosis at the termination of the immune response.The Foxo family of transcription factors has been the sub-
ject of much interest with regard to the control of cellular ho-
meostasis. These pro-apoptotic and anti-mitogenic geneswere initially isolated in humans at chromosomal break
points in rhabdomyosarcomas (1) and leukemias (2). Thesetranscription factors play a role in a diverse range of physiologi-cal processes, including cell survival, proliferation, metabo-lism and stress resistance (3–5). The three principal familymembers, Foxo1, Foxo3 and Foxo4, are post-translationallycontrolled via Akt (also known as protein kinase B) in thePhosphoinositide-3-kinase (PI3K) signaling pathway. In numer-ous cell types, including T and B lymphocytes, Akt-mediatedphosphorylation of Foxos in response to cytokines, growthfactors and other stimuli results in their exclusion from thenucleus and subsequent degradation, leading to cell cycle en-try (6–13). In B cells pre-stimulated with LPS, over-expressionof a constitutively active Foxo protein has been shown to resultin cell cycle arrest and apoptosis (13).
Correspondence to: A. B. Satterthwaite; E-mail: [email protected] Received 24 July 2008, accepted 30 April 2009
Transmitting editor: T. Tedder Advance Access publication 5 June 2009
Downloaded from https://academic.oup.com/intimm/article-abstract/21/7/831/820361by gueston 09 April 2018

Foxo1, Foxo3 and Foxo4 share largely overlapping pat-terns of tissue expression and interact preferentially with thesame DNA-binding motif 5#-TTGTTTAC-3# (14). This raisesthe question of functional redundancy. Yet, individual disrup-tion of each gene results in a distinct phenotype (15). Murineknockouts of Foxo1 are embryonic lethal due to defects inangiogenesis (16). Knockouts of both Foxo3 and Foxo4 areviable, however. Foxo3-deficient mice exhibit age-dependentinfertility in females (17, 18) as well as spontaneous T cellactivation and lymphoproliferation over time (19). Mice defi-cient in Foxo4 display decreased migration of vascularsmooth muscle cells (20).
Previous work in our laboratory has established that in ma-ture B cells, there is an additional means of control for Foxosbeyond that established at the post-translational level. Uponengagement of the B cell antigen receptor (BCR) in restingB cells, Foxo mRNA expression is down-regulated via PI3K(21). We now extend our findings to reveal that the mRNAexpression of each of the principal Foxo family members iscontrolled by a unique mechanism downstream of PI3K. Thissuggests that Foxo family members may have unique func-tions during B cell development and activation. Indeed, weshow here that Foxo3�/� mice have reduced frequencies ofpre-B cells in the bone marrow and recirculating B cells inthe blood and bone marrow. They also demonstrate in-creased basal levels of IgG2a, IgG3 and IgA. However,splenic B cell sub-populations are normal and the activationof mature resting B cells in vitro is unimpaired in the ab-sence of Foxo3, suggesting that other Foxo family membersmay play a role in these processes.
Materials and methods
Mice
Animals were housed and studied in accordance with proto-cols approved by the institutional animal care and usecommittee. Wild-type mice used in the inhibitor studieswere of the C57BL/6 genetic background. PhospholipaseC (PLC)c2�/� (22) were of mixed genetic background(C57BL/6 3 129). Littermates were therefore compared di-rectly when possible and experiments repeated with multiplelitters. Foxo3�/� (17) mice were of the FVB background.Experiments with these mice were completed using wild-type littermates as controls.
Flow cytometry
Splenocytes, bone marrow, peritoneal wash and peripheralblood were depleted of RBCs and stained with comb-inations of anti-CD21–FITC, anti-CD43–FITC, anti-IgM–PE,anti-CD23–PE, anti-CD5–PE, anti-B220–PerCP, anti-CD23–biotin, anti-CD93–biotin, anti-CD138–biotin, anti-CD22–biotin(eBioscience, San Diego, CA, USA) and anti-CD127–biotin(eBioscience). Antibodies were purchased from BDPharMingen (San Diego, CA, USA) unless otherwise indi-cated. Biotinylated antibodies were detected with streptavi-din–allophycocyanin (Caltag, Burlingame, CA, USA).Samples were run on a FACS Calibur (Becton Dickinson,San Jose, CA, USA). Data were analyzed with CellQuest(Becton Dickinson) software. Live cells were gated basedon forward and side scatter.
B cell purification
Splenocytes were incubated for 5 min in 0.15 M NH4Cl,1 mM KHCO3 and 0.1 mM Na2EDTA to deplete RBCs.Splenic B cells were then purified by negative selection withanti-CD43 magnetic beads using the MidiMACS system(Miltenyi Biotec, Auburn, CA, USA), according to the manu-facturer’s instructions. As measured by flow cytometry,purified B cells were >90% B220+.
B cell stimulation for quantitative real-time PCR analysis
Purified B cells were harvested at time 0 or stimulated with10 lg ml�1 goat anti-mouse IgM F(ab#)2 (Jackson ImmunoR-esearch Laboratories, West Grove, PA, USA) for 6 h. For in-hibitor studies, cells were pre-treated with either 10 lMLy294002, 50 lM Go6850, 10 lM U0126 or 100 ng ml�1
Cyclosporin A (CsA) for 15 min prior to anti-IgM treatmentor 10 lM 11R-VIVIT for 2 h prior to anti-IgM treatment. Allinhibitors were obtained from Calbiochem (San Diego, CA,USA).
Quantitative real-time PCR
Total RNA was prepared using the RNeasy Kit (Qiagen,Valencia, CA, USA). cDNA was generated with a cDNAArchive Kit (Applied Biosystems, Foster City, CA, USA).Real-time PCR was performed in an Applied Biosystems7300 Real-Time PCR system using TaqMan reagentsspecific for mouse B cell translocation gene 1(Btg-1), cyclinD2, cyclin G2, Foxo1, Foxo3, Foxo4, Edg1 [receptor forsphingosine-1 phosphate (S1P1)], Rag2 and the internalcontrol glyceraldehyde-3-phosphate dehydrogenase(GAPDH) (Applied Biosystems). Data were normalized toGAPDH using the delta comparative threshold cycle (Ct)method.
B cell proliferation, survival and differentiation
Splenic B cells were plated at 106 ml�1 in RPMI + 10% fetalbovine serum (FBS) with media alone, 2 or 20 lg ml�1 anti-IgM F(ab#)2 fragments (Jackson ImmunoResearch Laborato-ries), 0.1, 1 or 10 lg ml�1 LPS (Sigma, St. Louis, MO, USA),10 ng ml�1 phorbol myristate acetate (Sigma) + 1 lg ml�1
ionomycin (Calbiochem) or 10 ng ml�1 IL-4 (R&D Systems,Minneapolis, MN, USA) + 1 lg ml�1 or 10 lg ml�1 anti-CD40 (BD PharMingen) for 48 h. Proliferation was measuredby labeling cells with [3H]thymidine ([3H]TdR) (Amersham,Piscataway, NJ, USA) for the final 8 h. Apoptosis was mea-sured by staining with Annexin V–PE and 7-AAD (BD Phar-Mingen). For differentiation studies, splenic B cells wereplated at 5 3 105 ml�1 in RPMI + 10% FBS with mediaalone, 20 lg ml�1 LPS (Sigma) or 1 lg ml�1 anti-CD40 (BDPharMingen) + 10 ng ml�1 IL-4 (R&D Systems) for 72 h andsubsequently stained with antibodies against B220 andCD138.
Measurement of Ig Levels by ELISA
Mice were bled prior to and 1 and 2 weeks after immuniza-tion with either 10 lg trinitrophenol (TNP)–Ficoll (BiosearchTechnologies, Novato, CA, USA) in PBS or 100 lg NP–CGG(Biosearch Technologies) in Inject Alum (Pierce, Rockford,
832 Foxo3 in B cell development and activation
Downloaded from https://academic.oup.com/intimm/article-abstract/21/7/831/820361by gueston 09 April 2018
IL, USA). Serial dilutions of sera were assayed for total IgM,IgG1, IgG2a, IgG2b, IgG3 and IgA by ELISA using themouse clonotyping system (Southern Biotechnology Associ-ates, Birmingham, AL, USA). Serum Ig concentrations werecalculated based on purified Ig standards (Southern Bio-technology Associates). To assay anti-TNP and anti-NP anti-bodies, ELISAs were performed on serial dilutions of serumusing plates coated with TNP– or NP–BSA (Biosearch Tech-nologies) and secondary antibodies from the mouse clono-typing kit (Southern Biotechnology Associates). ELISAswere developed with an alkaline phosphatase substrate kit(Bio-Rad, Hercules, CA, USA) and OD405 read on a Bio-TekInstruments microplate reader.
IL-7 cultures
Bone marrow was cultured at 106 ml in RPMI + 10% FBS +
varying concentrations of IL-7 (R&D Systems) for 5 days.Proliferation was measured by [3H]TdR (Amersham) for anadditional 24 h. Additional analysis was performed on cellsexpanded for 5 days in 10 ng ml�1 IL-7. B220+ cells wereenumerated by flow cytometry, cDNA was prepared forquantitative real-time PCR (Q-PCR) analysis of Rag2 expres-sion and cell cycle status was assessed by fixing cells in70% ethanol for >24 h and then incubating with propidiumiodide (Roche, Indianapolis, IN, USA) and RNase A (Qiagen)for 20 min prior to analysis by flow cytometry.
Results
Differential regulation of Foxo family members upon B cellactivation
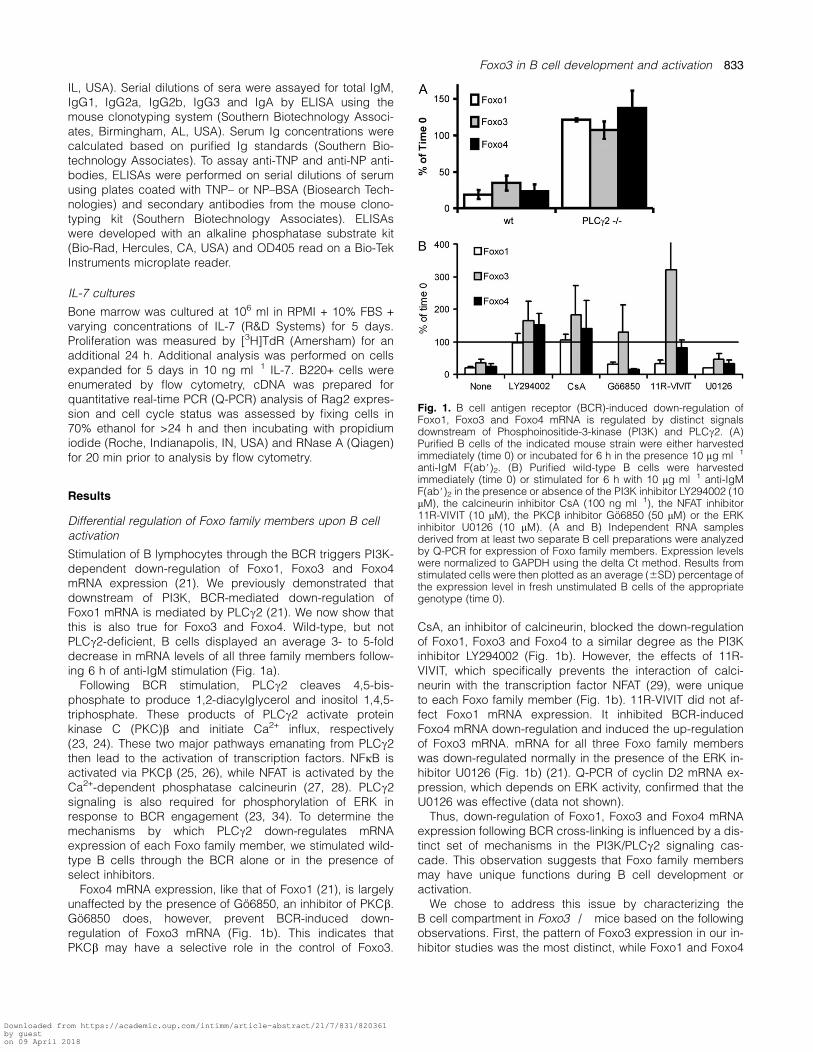
Stimulation of B lymphocytes through the BCR triggers PI3K-dependent down-regulation of Foxo1, Foxo3 and Foxo4mRNA expression (21). We previously demonstrated thatdownstream of PI3K, BCR-mediated down-regulation ofFoxo1 mRNA is mediated by PLCc2 (21). We now show thatthis is also true for Foxo3 and Foxo4. Wild-type, but notPLCc2-deficient, B cells displayed an average 3- to 5-folddecrease in mRNA levels of all three family members follow-ing 6 h of anti-IgM stimulation (Fig. 1a).
Following BCR stimulation, PLCc2 cleaves 4,5-bis-phosphate to produce 1,2-diacylglycerol and inositol 1,4,5-triphosphate. These products of PLCc2 activate proteinkinase C (PKC)b and initiate Ca2+ influx, respectively(23, 24). These two major pathways emanating from PLCc2then lead to the activation of transcription factors. NFjB isactivated via PKCb (25, 26), while NFAT is activated by theCa2+-dependent phosphatase calcineurin (27, 28). PLCc2signaling is also required for phosphorylation of ERK inresponse to BCR engagement (23, 34). To determine themechanisms by which PLCc2 down-regulates mRNAexpression of each Foxo family member, we stimulated wild-type B cells through the BCR alone or in the presence ofselect inhibitors.
Foxo4 mRNA expression, like that of Foxo1 (21), is largelyunaffected by the presence of Go6850, an inhibitor of PKCb.Go6850 does, however, prevent BCR-induced down-regulation of Foxo3 mRNA (Fig. 1b). This indicates thatPKCb may have a selective role in the control of Foxo3.
CsA, an inhibitor of calcineurin, blocked the down-regulationof Foxo1, Foxo3 and Foxo4 to a similar degree as the PI3Kinhibitor LY294002 (Fig. 1b). However, the effects of 11R-VIVIT, which specifically prevents the interaction of calci-neurin with the transcription factor NFAT (29), were uniqueto each Foxo family member (Fig. 1b). 11R-VIVIT did not af-fect Foxo1 mRNA expression. It inhibited BCR-inducedFoxo4 mRNA down-regulation and induced the up-regulationof Foxo3 mRNA. mRNA for all three Foxo family memberswas down-regulated normally in the presence of the ERK in-hibitor U0126 (Fig. 1b) (21). Q-PCR of cyclin D2 mRNA ex-pression, which depends on ERK activity, confirmed that theU0126 was effective (data not shown).
Thus, down-regulation of Foxo1, Foxo3 and Foxo4 mRNAexpression following BCR cross-linking is influenced by a dis-tinct set of mechanisms in the PI3K/PLCc2 signaling cas-cade. This observation suggests that Foxo family membersmay have unique functions during B cell development oractivation.
We chose to address this issue by characterizing theB cell compartment in Foxo3�/� mice based on the followingobservations. First, the pattern of Foxo3 expression in our in-hibitor studies was the most distinct, while Foxo1 and Foxo4
Fig. 1. B cell antigen receptor (BCR)-induced down-regulation ofFoxo1, Foxo3 and Foxo4 mRNA is regulated by distinct signalsdownstream of Phosphoinositide-3-kinase (PI3K) and PLCc2. (A)Purified B cells of the indicated mouse strain were either harvestedimmediately (time 0) or incubated for 6 h in the presence 10 lg ml�1
anti-IgM F(ab#)2. (B) Purified wild-type B cells were harvestedimmediately (time 0) or stimulated for 6 h with 10 lg ml�1 anti-IgMF(ab#)2 in the presence or absence of the PI3K inhibitor LY294002 (10lM), the calcineurin inhibitor CsA (100 ng ml�1), the NFAT inhibitor11R-VIVIT (10 lM), the PKCb inhibitor Go6850 (50 lM) or the ERKinhibitor U0126 (10 lM). (A and B) Independent RNA samplesderived from at least two separate B cell preparations were analyzedby Q-PCR for expression of Foxo family members. Expression levelswere normalized to GAPDH using the delta Ct method. Results fromstimulated cells were then plotted as an average (6SD) percentage ofthe expression level in fresh unstimulated B cells of the appropriategenotype (time 0).
Foxo3 in B cell development and activation 833
Downloaded from https://academic.oup.com/intimm/article-abstract/21/7/831/820361by gueston 09 April 2018
differed only with respect to dependence on NFAT (Fig. 1).Second, Foxo3 has been shown to have an effect on T cellactivation, suggesting that it may also play role in other lym-
phocytes (19). Finally, Foxo1�/� mice are embryonic lethal(16), while Foxo3-deficient mice are viable and readily exam-ined for B cell phenotypes (17–19).
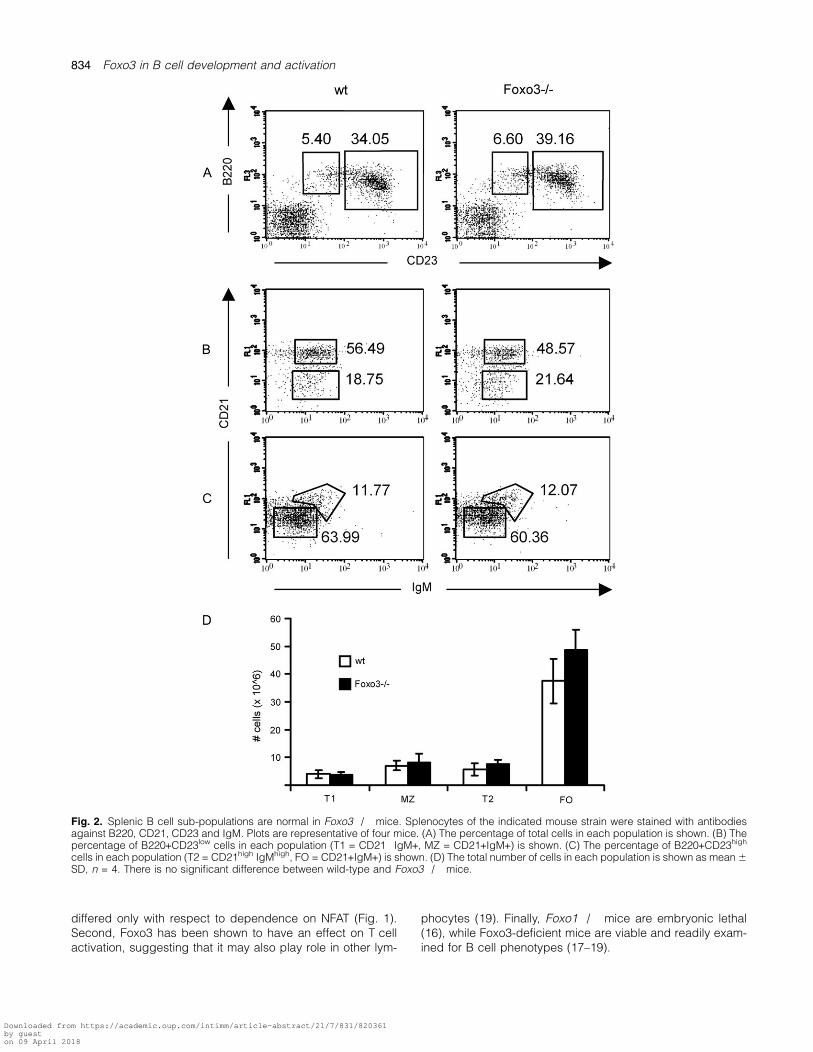
Fig. 2. Splenic B cell sub-populations are normal in Foxo3�/� mice. Splenocytes of the indicated mouse strain were stained with antibodiesagainst B220, CD21, CD23 and IgM. Plots are representative of four mice. (A) The percentage of total cells in each population is shown. (B) Thepercentage of B220+CD23low cells in each population (T1 = CD21�IgM+, MZ = CD21+IgM+) is shown. (C) The percentage of B220+CD23high
cells in each population (T2 = CD21high IgMhigh, FO = CD21+IgM+) is shown. (D) The total number of cells in each population is shown as mean 6SD, n = 4. There is no significant difference between wild-type and Foxo3�/� mice.
834 Foxo3 in B cell development and activation
Downloaded from https://academic.oup.com/intimm/article-abstract/21/7/831/820361by gueston 09 April 2018

Foxo3 is dispensable for splenic B cell development andresponse to BCR engagement
Similar frequencies and total numbers of follicular, marginalzone and transitional B cells were found in the spleens ofwild-type and Foxo3�/� mice (Fig. 2). Despite the anti-mito-genic and pro-apoptotic functions of Foxo3, splenic B cellslacking Foxo3 did not demonstrate altered proliferation(Fig. 3a) or survival (Fig. 3b) either basally or in response toanti-IgM. The lack of an effect of Foxo3 deficiency was notdue to compensatory changes in the expression levels ofFoxo1 or Foxo4 (Fig. 3c). mRNA expression of cyclin G2and Btg-1, cell cycle inhibitory genes which are known tar-gets of Foxo3 (30–34), was comparable in wild-type andFoxo3�/� B cells both basally and upon BCR engagement(Fig. 3d).
Increased basal IgG2a, IgG3 and IgA levels in Foxo3�/�mice
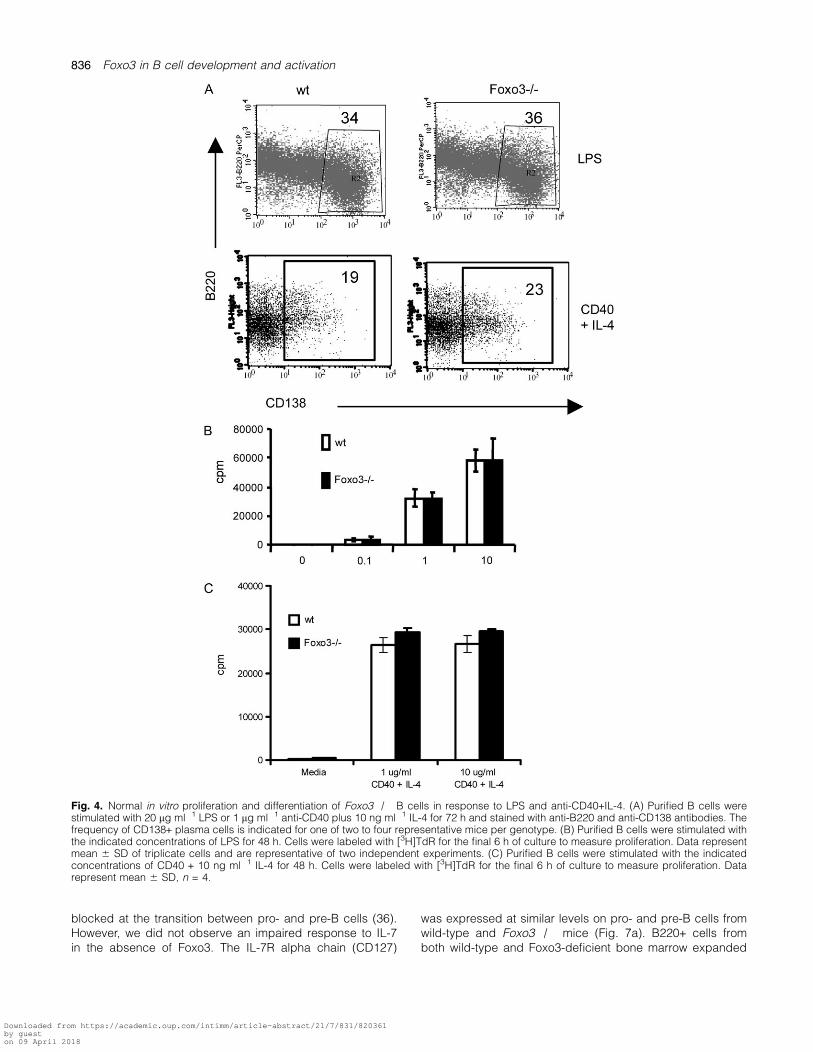
Forced expression of a constitutively active form of Foxo1 inLPS-stimulated B cells inhibits the ability of these cells to dif-ferentiate into plasma cells (35). This suggested that plasmacell differentiation may be increased in the absence ofFoxo3. However, up-regulation of the plasma cell markerCD138 was normal in cultures of Foxo3�/� B cells incu-bated with either LPS or CD40 plus IL-4 (Fig. 4a). Wild-typeand Foxo3�/� B cells also had a similar proliferative re-sponse to these stimuli (Fig. 4b and c).
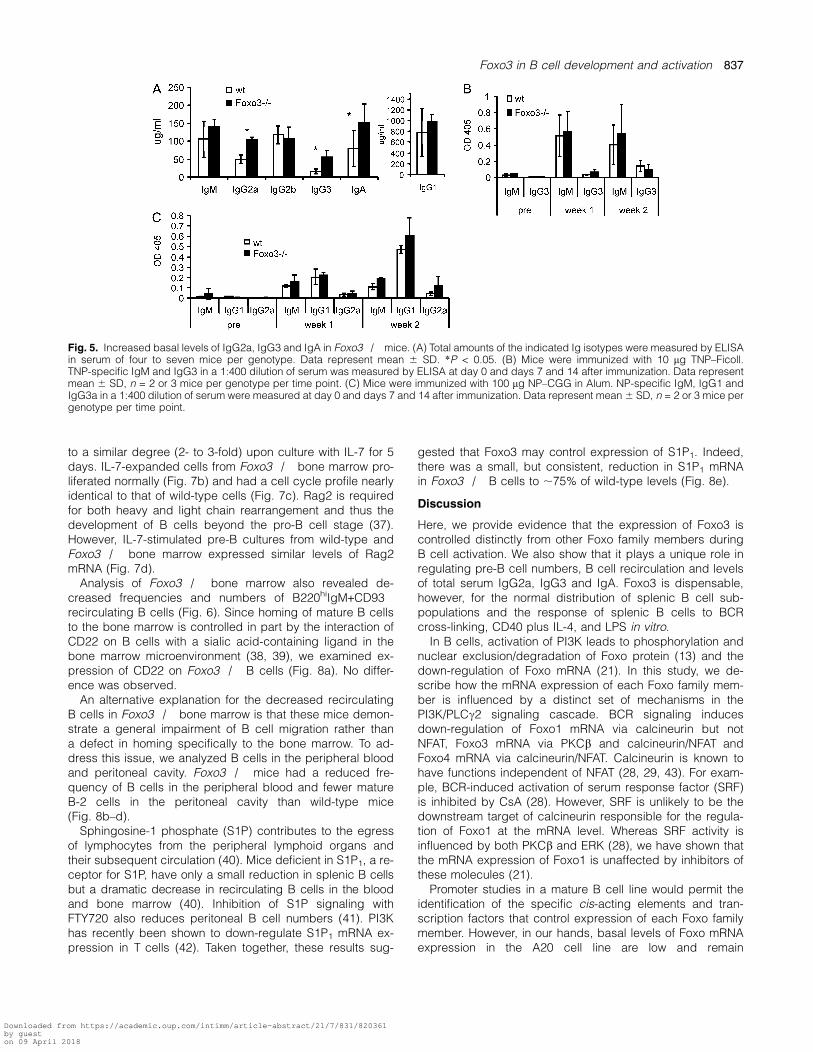
Although there was no general enhancement of plasmacell differentiation in vitro in the absence of Foxo3, it is pos-sible that Foxo3 regulates B cell differentiation and antibodyproduction in vivo. We therefore measured serum Ig levels,both basally and in response to immunization. UnimmunizedFoxo3�/� mice had significantly increased amounts of totalIgG2a, IgG3 and IgA relative to wild-type controls, althoughlevels of total IgM, IgG1 and IgG2b were unchanged(Fig. 5a).
The ability of Foxo3�/� mice to mount a humoral im-mune response to specific antigens was also assessed.Responses to the T-independent antigen TNP–Ficoll(Fig. 5b) and the T-dependent antigen NP–CGG (Fig. 5c)were not significantly different between wild-type andFoxo3�/� mice, although there was a trend towarda slightly increased response to NP–CGG in the absenceof Foxo3 2 weeks after immunization.
Decreased pre-B and recirculating B cells in Foxo3�/� mice
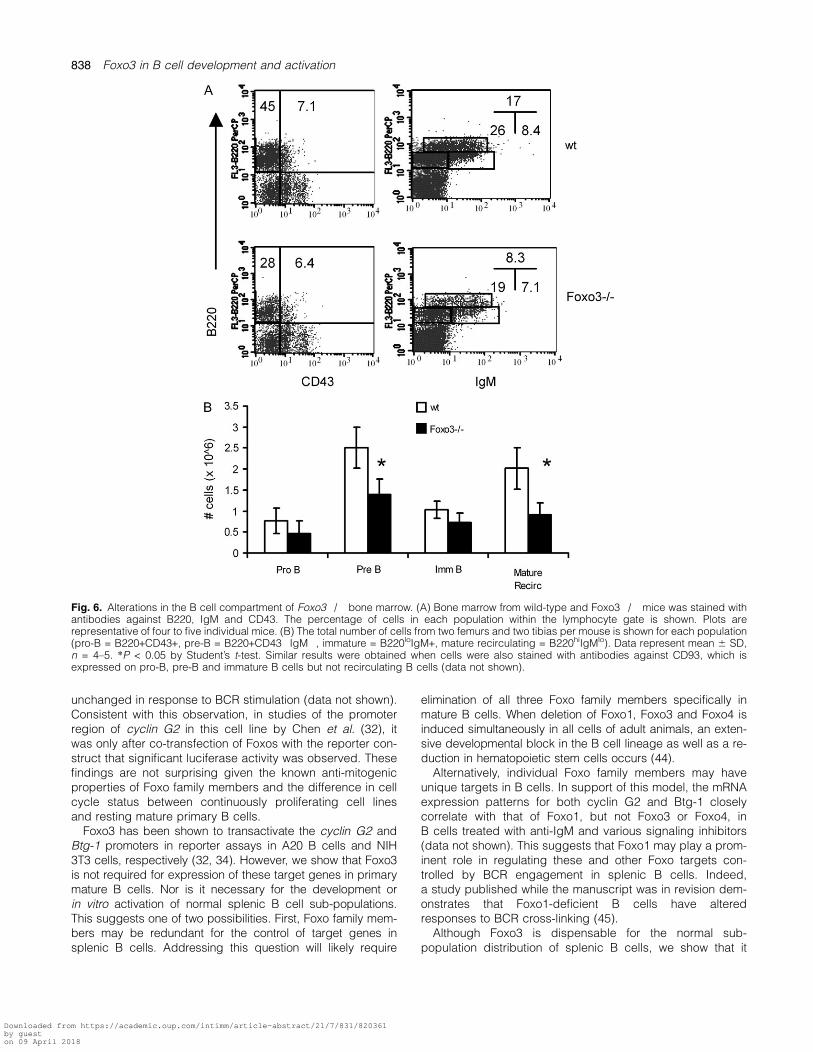
To determine whether Foxo3 uniquely contributes to otherstages of B cell development, we examined the bone mar-row of Foxo3�/� mice. Intriguingly, while both B220+CD43+pro-B cells and B220+CD93+IgM+ immature B cells werepresent at normal numbers, there was a significant reductionin both the percentage and total number of B220+CD43�CD93+IgM� pre-B cells in the absence of Foxo3 (Fig. 6).IL-7 is an important growth and differentiation factor forB cell progenitors. In its absence, B cell development is
Fig. 3. Normal response to BCR cross-linking in the absence of Foxo3. (A) Splenic B cells were stimulated for 48 h with media alone, 2 or 20 lgml�1 anti-IgM F(ab’)2 fragments or 10 ng ml�1 phorbol myristate acetate + 1 lg ml�1 ionomycin and proliferation measured by [3H]TdRincorporation. Data represent mean 6 SD, n = 3. (B) Purified B cells were cultured in media alone or 10 lg ml�1 anti-IgM F(ab#)2 for 48 h andstained with Annexin V and 7-AAD. The frequency of live (Annexin V� and 7-AAD�), early apoptotic (Annexin V+ and 7-AAD�) and dead(Annexin V+ and 7-AAD+) cells is shown for one of three representative mice per genotype. (C and D) Purified B cells of the indicated mousestrain were either harvested immediately (time 0) or incubated for 6 h in the presence 10 lg ml�1 anti-IgM F(ab#)2. Independent RNA samplesderived from three separate B cell preparations were analyzed by Q-PCR for expression of (C) Foxo1 and Foxo4 as well as (D) cyclin G2 and Btg-1. Expression levels for each gene were normalized to GAPDH using the delta Ct method and plotted as an average (6SD) percentage of theexpression level of fresh unstimulated wild-type B cells (time 0).
Foxo3 in B cell development and activation 835
Downloaded from https://academic.oup.com/intimm/article-abstract/21/7/831/820361by gueston 09 April 2018
blocked at the transition between pro- and pre-B cells (36).However, we did not observe an impaired response to IL-7in the absence of Foxo3. The IL-7R alpha chain (CD127)
was expressed at similar levels on pro- and pre-B cells fromwild-type and Foxo3�/� mice (Fig. 7a). B220+ cells fromboth wild-type and Foxo3-deficient bone marrow expanded
Fig. 4. Normal in vitro proliferation and differentiation of Foxo3�/� B cells in response to LPS and anti-CD40+IL-4. (A) Purified B cells werestimulated with 20 lg ml�1 LPS or 1 lg ml�1 anti-CD40 plus 10 ng ml�1 IL-4 for 72 h and stained with anti-B220 and anti-CD138 antibodies. Thefrequency of CD138+ plasma cells is indicated for one of two to four representative mice per genotype. (B) Purified B cells were stimulated withthe indicated concentrations of LPS for 48 h. Cells were labeled with [3H]TdR for the final 6 h of culture to measure proliferation. Data representmean 6 SD of triplicate cells and are representative of two independent experiments. (C) Purified B cells were stimulated with the indicatedconcentrations of CD40 + 10 ng ml�1 IL-4 for 48 h. Cells were labeled with [3H]TdR for the final 6 h of culture to measure proliferation. Datarepresent mean 6 SD, n = 4.
836 Foxo3 in B cell development and activation
Downloaded from https://academic.oup.com/intimm/article-abstract/21/7/831/820361by gueston 09 April 2018
to a similar degree (2- to 3-fold) upon culture with IL-7 for 5days. IL-7-expanded cells from Foxo3�/� bone marrow pro-liferated normally (Fig. 7b) and had a cell cycle profile nearlyidentical to that of wild-type cells (Fig. 7c). Rag2 is requiredfor both heavy and light chain rearrangement and thus thedevelopment of B cells beyond the pro-B cell stage (37).However, IL-7-stimulated pre-B cultures from wild-type andFoxo3�/� bone marrow expressed similar levels of Rag2mRNA (Fig. 7d).
Analysis of Foxo3�/� bone marrow also revealed de-creased frequencies and numbers of B220hiIgM+CD93�recirculating B cells (Fig. 6). Since homing of mature B cellsto the bone marrow is controlled in part by the interaction ofCD22 on B cells with a sialic acid-containing ligand in thebone marrow microenvironment (38, 39), we examined ex-pression of CD22 on Foxo3�/� B cells (Fig. 8a). No differ-ence was observed.
An alternative explanation for the decreased recirculatingB cells in Foxo3�/� bone marrow is that these mice demon-strate a general impairment of B cell migration rather thana defect in homing specifically to the bone marrow. To ad-dress this issue, we analyzed B cells in the peripheral bloodand peritoneal cavity. Foxo3�/� mice had a reduced fre-quency of B cells in the peripheral blood and fewer matureB-2 cells in the peritoneal cavity than wild-type mice(Fig. 8b–d).
Sphingosine-1 phosphate (S1P) contributes to the egressof lymphocytes from the peripheral lymphoid organs andtheir subsequent circulation (40). Mice deficient in S1P1, a re-ceptor for S1P, have only a small reduction in splenic B cellsbut a dramatic decrease in recirculating B cells in the bloodand bone marrow (40). Inhibition of S1P signaling withFTY720 also reduces peritoneal B cell numbers (41). PI3Khas recently been shown to down-regulate S1P1 mRNA ex-pression in T cells (42). Taken together, these results sug-
gested that Foxo3 may control expression of S1P1. Indeed,there was a small, but consistent, reduction in S1P1 mRNAin Foxo3�/� B cells to ;75% of wild-type levels (Fig. 8e).
Discussion
Here, we provide evidence that the expression of Foxo3 iscontrolled distinctly from other Foxo family members duringB cell activation. We also show that it plays a unique role inregulating pre-B cell numbers, B cell recirculation and levelsof total serum IgG2a, IgG3 and IgA. Foxo3 is dispensable,however, for the normal distribution of splenic B cell sub-populations and the response of splenic B cells to BCRcross-linking, CD40 plus IL-4, and LPS in vitro.
In B cells, activation of PI3K leads to phosphorylation andnuclear exclusion/degradation of Foxo protein (13) and thedown-regulation of Foxo mRNA (21). In this study, we de-scribe how the mRNA expression of each Foxo family mem-ber is influenced by a distinct set of mechanisms in thePI3K/PLCc2 signaling cascade. BCR signaling inducesdown-regulation of Foxo1 mRNA via calcineurin but notNFAT, Foxo3 mRNA via PKCb and calcineurin/NFAT andFoxo4 mRNA via calcineurin/NFAT. Calcineurin is known tohave functions independent of NFAT (28, 29, 43). For exam-ple, BCR-induced activation of serum response factor (SRF)is inhibited by CsA (28). However, SRF is unlikely to be thedownstream target of calcineurin responsible for the regula-tion of Foxo1 at the mRNA level. Whereas SRF activity isinfluenced by both PKCb and ERK (28), we have shown thatthe mRNA expression of Foxo1 is unaffected by inhibitors ofthese molecules (21).
Promoter studies in a mature B cell line would permit theidentification of the specific cis-acting elements and tran-scription factors that control expression of each Foxo familymember. However, in our hands, basal levels of Foxo mRNAexpression in the A20 cell line are low and remain
Fig. 5. Increased basal levels of IgG2a, IgG3 and IgA in Foxo3�/� mice. (A) Total amounts of the indicated Ig isotypes were measured by ELISAin serum of four to seven mice per genotype. Data represent mean 6 SD. *P < 0.05. (B) Mice were immunized with 10 lg TNP–Ficoll.TNP-specific IgM and IgG3 in a 1:400 dilution of serum was measured by ELISA at day 0 and days 7 and 14 after immunization. Data representmean 6 SD, n = 2 or 3 mice per genotype per time point. (C) Mice were immunized with 100 lg NP–CGG in Alum. NP-specific IgM, IgG1 andIgG3a in a 1:400 dilution of serum were measured at day 0 and days 7 and 14 after immunization. Data represent mean 6 SD, n = 2 or 3 mice pergenotype per time point.
Foxo3 in B cell development and activation 837
Downloaded from https://academic.oup.com/intimm/article-abstract/21/7/831/820361by gueston 09 April 2018
unchanged in response to BCR stimulation (data not shown).Consistent with this observation, in studies of the promoterregion of cyclin G2 in this cell line by Chen et al. (32), itwas only after co-transfection of Foxos with the reporter con-struct that significant luciferase activity was observed. Thesefindings are not surprising given the known anti-mitogenicproperties of Foxo family members and the difference in cellcycle status between continuously proliferating cell linesand resting mature primary B cells.
Foxo3 has been shown to transactivate the cyclin G2 andBtg-1 promoters in reporter assays in A20 B cells and NIH3T3 cells, respectively (32, 34). However, we show that Foxo3is not required for expression of these target genes in primarymature B cells. Nor is it necessary for the development orin vitro activation of normal splenic B cell sub-populations.This suggests one of two possibilities. First, Foxo family mem-bers may be redundant for the control of target genes insplenic B cells. Addressing this question will likely require
elimination of all three Foxo family members specifically inmature B cells. When deletion of Foxo1, Foxo3 and Foxo4 isinduced simultaneously in all cells of adult animals, an exten-sive developmental block in the B cell lineage as well as a re-duction in hematopoietic stem cells occurs (44).
Alternatively, individual Foxo family members may haveunique targets in B cells. In support of this model, the mRNAexpression patterns for both cyclin G2 and Btg-1 closelycorrelate with that of Foxo1, but not Foxo3 or Foxo4, inB cells treated with anti-IgM and various signaling inhibitors(data not shown). This suggests that Foxo1 may play a prom-inent role in regulating these and other Foxo targets con-trolled by BCR engagement in splenic B cells. Indeed,a study published while the manuscript was in revision dem-onstrates that Foxo1-deficient B cells have alteredresponses to BCR cross-linking (45).
Although Foxo3 is dispensable for the normal sub-population distribution of splenic B cells, we show that it
Fig. 6. Alterations in the B cell compartment of Foxo3�/� bone marrow. (A) Bone marrow from wild-type and Foxo3�/� mice was stained withantibodies against B220, IgM and CD43. The percentage of cells in each population within the lymphocyte gate is shown. Plots arerepresentative of four to five individual mice. (B) The total number of cells from two femurs and two tibias per mouse is shown for each population(pro-B = B220+CD43+, pre-B = B220+CD43�IgM�, immature = B220loIgM+, mature recirculating = B220hiIgMlo). Data represent mean 6 SD,n = 4–5. *P < 0.05 by Student’s t-test. Similar results were obtained when cells were also stained with antibodies against CD93, which isexpressed on pro-B, pre-B and immature B cells but not recirculating B cells (data not shown).
838 Foxo3 in B cell development and activation
Downloaded from https://academic.oup.com/intimm/article-abstract/21/7/831/820361by gueston 09 April 2018

has a unique role in regulating pre-B cell numbers. Despitethis observation, no difference in IL-7R expression or expan-sion of B220+ cells in IL-7 cultures was observed between
wild-type and Foxo3�/� bone marrow. Rag2 expressionwas also normal in IL-7-expanded pre-B cells from Foxo3�/�mice. Impairment of earlier stages of B lymphopoiesis may
Fig. 7. Foxo3�/� bone marrow cells respond normally to IL-7. (A) Bone marrow was stained with antibodies against IgM, B220, CD43 andIL-7Ra (CD127). The level of IL-7Ra on pro- and pre-B cells (B220+IgM�) is shown for wild-type (shaded histogram) and Foxo3�/� (openhistogram, dark line) mice. (B) Bone marrow cells were plated in media alone or varying concentrations of IL-7 at 106 ml�1 for 5 days. Proliferationwas measured by [3H]TdR incorporation for an additional 24 h. Data represent mean 6 SD, n = 3. (C) Bone marrow cells were plated in 10 ng ml�1
IL-7 at 106 ml�1 for 5 days and stained with propidium iodide (PI) to assess cell cycle status. The frequency of apoptotic (sub 2n) and S/G2/Mcells (>2n) is indicated. Plots are representative of n = 3–4. (E) cDNA was prepared from day 5 IL-7 bone marrow cultures and subjected to Q-PCR for Rag2. Expression levels were normalized to GAPDH using the delta Ct method and plotted as a percentage of the average expressionlevel in wild-type B cells. Data represent mean 6 SD, n = 2.
Fig. 8. Reduced recirculating B cells in Foxo3�/� mice. (A) Splenocytes were stained with antibodies against CD22, B220, CD21 and CD23.The level of CD22 expression on follicular B cells (B220+CD23+CD21+) from wild-type (shaded histogram) and Foxo3�/� (open histogram, darkline) mice is shown. Data are representative of three mice. (B) B220 expression is shown for cells in the lymphocyte gate of peripheral blood fromwild-type (shaded histogram) and Foxo3�/� (open histogram, dark line) mice. Data are representative of three mice. (C) Peritoneal cells werestained with antibodies against B220 and CD5. The frequency of B-2 (B220+CD5�) and B-1 (B220lowCD5+) cells in the lymphocyte gate isindicated. Data are representative of three mice. (D) The mean 6 SD of three mice analyzed as in (B and C) is shown. *P < 0.05 by pairedStudent’s t-test. (E) Purified B cells were analyzed by Q-PCR for expression of S1P1 (Edg1). Expression levels were normalized to GAPDH usingthe delta Ct method and plotted as a percentage of the average expression level in wild-type B cells. Each symbol represents an independentexperiment with one mouse of each genotype. *P < 0.05 by paired Student’s t-test.
Foxo3 in B cell development and activation 839
Downloaded from https://academic.oup.com/intimm/article-abstract/21/7/831/820361by gueston 09 April 2018

play a role, however. While the frequency and numberof pro-B cells was not significantly different between wild-type and Foxo3�/� mice, there was a trend toward a slightreduction in this population in the absence of Foxo3. In addi-tion, fewer cells fell into the lymphoid gate in Foxo3�/� bonemarrow. Thus, some of the effect of loss of all three Foxofamily members on hematopoietic stem cells and pro-B cells(44) may be due specifically to Foxo3. Foxo3 may also act inthe bone marrow microenvironment to support the develop-ment or survival of pre-B cells. Intriguingly, mice lackingFoxo1 specifically in the B lineage were recently shown toalso have a block in development at the pre-B stage (45).However, unlike the case in Foxo3�/� mice, the absence ofFoxo1 results in decreased expression of both Rag2 andIL-7R in early B lineage cells (45, 46). Thus, Foxo3 andFoxo1 maintain pre-B cell numbers via distinct mechanisms.
Foxo3 deficiency also results in a decreased frequency ofrecirculating B cells in the bone marrow and blood as wellas a reduction in B-2 cells in the peritoneum. This functionof Foxo3 is B cell intrinsic, as a similar result was observedin a recent study examining mice lacking Foxo3 specificallyin B cells (47). B cell expression of CD22, which mediates Bcell homing to the bone marrow (38, 39), was normal in theabsence of Foxo3. However, mRNA encoding S1P1, whichmediates S1P-induced egress of B and T cells from lymphoidorgans (40), was reduced in B cells from Foxo3�/� mice.This is consistent with recent observations indicating thatPI3K signaling down-regulates S1P1 expression via Foxo tran-scription factors in T cells (42, 48). B cells from S1P1+/� micedemonstrate impaired exit from peripheral lymphoid organs,suggesting that degree of change in S1P1 expression seen inFoxo3�/� B cells is physiologically relevant (49). S1P has re-cently been shown to promote egress by counteracting otherchemokines that retain lymphocytes in the spleen and lymphnodes (50). Foxo3 deficiency may disrupt the balance ofthese signals in favor of retention by altering responses to ei-ther S1P or other chemokines. It is possible that Foxo3 targetsin addition to S1P1 also promote B cell egress, migration orhoming. It should be noted that the block in B cell recircula-tion is incomplete in Foxo3�/� mice, indicating that otherFoxo family members, or Foxo-independent mechanisms,likely contribute this process as well.
Finally, Foxo3 has a unique role in regulating basal levelsof serum IgG2a, IgG3 and IgA. This could result from eithera heightened ability of B cells to respond to factors that in-duce switching to these isotypes (i.e. IFNc and transforminggrowth factor b) or an increased production of these factorsby other cell types. The latter is a likely possibility given thatgeneralized CD4+ T cell hyperactivity has been reported inan independently generated line of Foxo3�/� mice (19).Mice were able to respond and class switch upon immuniza-tion with TNP–Ficoll and NP–CGG in the absence of Foxo3.This is in contrast to mice lacking Foxo1 in mature B cells,which fail to class switch in response to either T-independentor T-dependent antigens (45). Taken together, these resultsindicate that Foxo1 and Foxo3 regulate Ig production viadistinct mechanisms.
Foxos and their targets play important roles in diseasesassociated with altered proliferation and/or activation of Bcells. Resistance to cell death in B-chronic lymphocytic leu-
kemia has been linked to inactivation of Foxo3 via proteinphosphorylation (51). Similarly, BCR–ABL, which causesboth chronic myelogenous leukemia and acute lymphocyticleukemia, acts in part by down-regulating Foxo3 (52, 53). Ithas also been proposed that increased PKCb signaling,which we show here down-regulates Foxo3 mRNA levels innormal B cells stimulated through the BCR, could contributeto lymphoma (26). These findings are interesting in that theysupport the theory that both transcriptional and post-transla-tional control of Foxo family members could have conse-quences for B cell malignancy. Btg-1, like the Foxotranscription factors themselves (2), has been associatedwith chromosomal translocations in leukemia (54). mRNA ex-pression of this target gene is also up-regulated in EBV-trans-formed B cells from X-linked agammaglobulinemia patients(55), suggesting that aberrant expression of Btg-1 has conse-quences not only in cancer but immunodeficiency as well.Foxo family members may also play a role in immune toler-ance. Foxo-induced Rag expression in immature B cells islikely to promote elimination of autoreactive B cells via recep-tor editing (46, 47). Preliminary studies have also shown al-tered Foxo1 transcipt levels in the PBMC of both systemiclupus erythematosus and rheumatoid arthritis patients (56).Thus, teasing out the roles for each Foxo family member andits associated target genes in the maintenance of B cell ho-meostasis has implications for prevention and/or treatment ofcancer, immunodeficiency and autoimmunity.
Funding
National Institutes of Health Grants (AI049248 to A.B.S.,1K26RR024196 to D.H.C., T32 AI 005284-28 to R.M.H.); Uni-versity of Texas Southwestern Summer Undergraduate Re-search Fellowship Program to W.A.N.; Southwestern MedicalFoundation Scholarship in Biomedical Research to A.B.S.
Acknowledgements
We thank Toni Gutierrez and Sandirai Musuka for both technicalassistance and mouse genotyping. We also thank Laurie Davis, ToniGutierrez and Sandirai Musuka for helpful suggestions and criticalreading of the manuscript. James Ihle provided the PLCc2�/� mice.
Abbreviations
BCR B cell antigen receptorBtg-1 B cell translocation gene 1CsA cyclosporin ACt comparative threshold cycleFBS fetal bovine serumGAPDH glyceraldehyde-3-phosphate dehydrogenase[3H]TdR [3H]thymidinePI3K phosphoinositide-3-kinasePKC protein kinase CPLC phospholipase CQ-PCR quantitative real-time PCRS1P sphingosine-1 phosphateS1P1 receptor for sphingosine-1 phosphateSRF serum response factorTNP trinitrophenol
References
1 Barr, F. G. 2001. Gene fusions involving PAX and FOX family
members in alveolar rhabdomyosarcoma. Oncogene 20:5736.
840 Foxo3 in B cell development and activation
Downloaded from https://academic.oup.com/intimm/article-abstract/21/7/831/820361by gueston 09 April 2018

2 Borkhardt, A., Repp, R., Haas, O. et al. 1997. Cloning andcharacterization of AFX, the gene that fuses to MLL in acuteleukemias with a t(X;11)(q13;q23). Oncogene 14:195.
3 Burgering, B. M. T. and Kops, G. J. P. L. 2002. Cell cycle anddeath control: long live forkheads. Trends Biochem. Sci. 27:352.
4 Birkenkamp, K. U. and Coffer, P. J. 2003. FOXO transcriptionfactors as regulators of immune homeostasis: molecules to diefor? J. Immunol. 171:1623.
5 Tran, H., Brunet, A., Griffith, E. C. and Greenberg, M. E. 2003. Themany forks in FOXO’s road. Sci. STKE 2003:re5.
6 Biggs, W. H. III, Meisenhelder, J., Hunter, T., Cavenee, W. K. andArden, K. C. 1999. Protein kinase B/Akt-mediated phosphorylationpromotes nuclear exclusion of the winged helix transcription factorFKHR1. Proc. Natl Acad. Sci. USA 196:7421.
7 Brunet, A., Bonni, A., Zigmond, M. J. et al. 1999. Akt promotes cellsurvival by phosphorylating and inhibiting a forkhead transcriptionfactor. Cell 96:857.
8 Tang, E. D., Nunez, G., Barr, F. G. and Guan, K.-L. 1999. Negativeregulation of the forkhead transcription factor FKHR by Akt. J. Biol.Chem. 274:16741.
9 Kops, G. J. P. L., de Ruiter, N. D., De Vries-Smits, A. M. M., Powell,D. R., Bos, J. L. and Burgering, B. M. T. 1999. Direct control of theforkhead transcription factor AFX by protein kinase B. Nature398:630.
10 Rena, G., Guo, S., Cichy, S. C., Unterman, T. G. and Cohen, P.1999. Phosphorylation of the transcription factor forkheadfamily member FKHR by protein kinase B. J. Biol. Chem.274:17179.
11 Takaishi, H., Konishi, H., Matsuzaki, H. et al. 1999. Regulation ofnuclear translocation of forkhead transcription factor AFX byprotein kinase B. Proc. Natl Acad. Sci. USA 96:11836.
12 Fabre, S., Lang, V., Harriague, J. et al. 2005. Stable activation ofphosphatidylinositol 3-kinase in the T cell immunological synapsestimulates Akt signaling to FoxO1 nuclear exclusion and cellgrowth control. J. Immunol. 174:4161.
13 Yusuf, I., Zhu, X., Kharas, M. G., Chen, J. and Fruman, D. A. 2004.Optimal B-cell proliferation requires phosphoinositide 3-kinase-dependent inactivation of FOXO transcription factors. Blood104:784.
14 Furuyama, T., Nakazawa, T., Nakano, I. and Mori, N. 2000.Identification of the differential distribution patterns of mRNAs andconsensus binding sequences for mouse DAF-16 homologues.Biochem. J. 349:629.
15 Hosaka, T., Biggs, W. H. III, Tieu, D., Boyer, A. D., Varki, N. M.,Cavenee, W. K. and Arden, K. C. 2004. Disruption of forkheadtranscription factor (FOXO) family members in mice reveals theirfunctional diversification. Proc. Natl Acad. Sci. USA 101:2975.
16 Furuyama, T., Kitayama, K., Shimoda, Y. et al. 2004. Abnormalangiogenesis in Foxo1 (Fkhr)-deficient mice. J. Biol. Chem.279:34741.
17 Castrillon, D. H., Miao, L., Kollipara, R., Horner, J. W. and DePinho,R. A. 2003. Suppression of ovarian follicle activation in mice by thetranscription factor Foxo3a. Science 301:215.
18 John, G. B., Shirley, L. J., Gallardo, T. D. and Castrillon, D. H.2007. Specificity of the requirement for Foxo3 in primordial follicleactivation. Reproduction 133:855.
19 Lin, L., Hron, J. and Peng, S. 2004. Regulation of NF-jB, Thactivation, and autoinflammation by the forkhead transcriptionfactor Foxo3. Immunity 21:203.
20 Hao, L., Laing, J., Castrillon, D. H., DePinho, R. A., Olson, E. N.and Liu, Z. 2007. FoxO4 regulates tumor necrosis factor alpha-directed smooth muscle cell migration by activating matrixmetalloproteinase 9 gene transcription. Mol. Cell. Biol. 27:2676.
21 Hinman, R. M., Bushanam, J. N., Nichols, W. A. and Satterthwaite,A. B. 2007. B cell receptor signaling down-regulates forkhead boxtranscription factor class O 1 mRNA expression via phosphatidy-linositol 3-kinase and Bruton’s tyrosine kinase. J. Immunol.178:740.
22 Wang, D., Feng, J., Wen, R. et al. 2000. Phospholipase Cc2 isessential in the functions of B cell and several Fc receptors.Immunity 13:25.
23 Kurosaki, T. 1999. Genetic analysis of B cell antigen receptorsignaling. Annu. Rev. Immunol. 17:555.
24 Kurosaki, T., Maeda, A., Ishiai, M., Hashimoto, A., Inabe, K. andTakata, M. 2000. Regulation of the phospholipase C-gamma2pathway in B cells. Immunol. Rev. 176:19.
25 Saijo, K., Mecklenbrauker, I., Santana, A., Leitges, M., Schmedt,C. and Tarakhovsky, A. 2002. Protein kinase C beta controlsnuclear factor kappaB activation in B cells through selectiveregulation of the IkappaB kinase alpha. J. Exp. Med. 195:1647.
26 Su, T. T., Guo, B., Kawakami, Y. et al. 2002. PKC-b controls IjBkinase lipid raft recruitment and activation in response to BCRsignaling. Nat. Immunol. 3:780.
27 Rao, A., Luo, C. and Hogan, P. G. 1997. Transcription factors ofthe NFAT family: regulation and function. Annu. Rev. Immunol.15:707.
28 Hao, S., Kurosaki, T. and August, A. 2003. Differential regulation ofNFAT and SRF by the B cell receptor via a PLCc-Ca2+-dependentpathway. EMBO J. 22:4166.
29 Aramburu, J., Yaffe, M. B., Lopez-Rodriguez, C., Cantley, L. C.,Hogan, P. G. and Rao, A. 1999. Affinity-driven peptide selection ofan NFAT inhibitor more selective than cyclosporin A. Science285:2129.
30 Rouault, J. P., Rimokh, R., Tessa, C. et al. 1992. BTG1, a memberof a new family of antiproliferative genes. EMBO J. 11:1663.
31 Bennin, D. A., Don, A. S. A., Brake, T. et al. 2002. Cyclin G2associates with protein phosphatase 2A catalytic and regulatoryB’ subunits in active complexes and induces nuclear aberrationsand a G1/S phase cell cycle arrest. J. Biol. Chem. 277:27449.
32 Chen, J., Yusuf, I., Andersen, H. and Fruman, D. A. 2006. FOXOtranscription factors cooperate with dEF1 to activate growthsuppressive genes in B lymphocytes. J. Immunol. 176:2711.
33 Bakker, W. J., Blazquez-Domingo, M., Kolbus, A. et al. 2004. Foxo3aregulates erythroid differentiation and induces BTG1, an activatorof protein arginine methyl transferase 1. J. Cell. Biol. 164:175.
34 Bakker, W. J., Van Dijk, T. B., Parren-Van Amelsvoort, M. et al.2007. Differential regulation of Foxo3a target genes in erythropoi-esis. Mol. Cell. Biol. 27:3839.
35 Omori, S. A., Cato, M. H., Anzelon-Mills, A. et al. 2006. Regulationof class-switch recombination and plasma cell differentiation byphosphatidylinositol 3-kinase signaling. Immunity 25:545.
36 von Freeden-Jeffry, U., Vieira, P., Lucian, L. A., McNeil, T.,Burdach, S. E. and Murray, R. 1995. Lymphopenia in interleukin(IL)-7 gene-deleted mice identifies IL-7 as a nonredundantcytokine. J. Exp. Med. 181:1519.
37 Shinkai, Y., Rathbun, G., Lam, K. P. et al. 1992. RAG-2-deficientmice lack mature lymphocytes owing to inability to initiate V(D)Jrearrangement. Cell 68:855.
38 Nitschke, L., Floyd, H., Ferguson, D. J. P. and Crocker, P. R. 1999.Identification of CD22 ligands on bone marrow sinusoidalendothelium implicated in CD22-dependent homing of recirculat-ing B cells. J. Exp. Med. 189:1513.
39 Ghosh, S., Bandulet, C. and Nitschke, L. 2006. Regulation of Bcell development and B cell signaling by CD22 and its ligandsa2,6-linked sialic acids. Int. Immunol. 18:603.
40 Matloubian, M., Lo, C. G., Cinamon, G. et al. 2004. Lymphocyteegress from thymus and peripheral lymphoid organs is dependenton S1P receptor 1. Nature 427:355.
41 Kunisawa, J., Kurashima, Y., Gohda, M. et al. 2007. Sphingosine1-phosphate regulates peritoneal B cell trafficking for subsequentintestinal IgA production. Blood 109:3746.
42 Sinclair, L. V., Finlay, D., Feijoo, C. et al. 2008. Phosphatidylino-sitol-3-OH kinase and nutrient sensing mTOR pathways controlT lymphocyte trafficking. Nat. Immunol. 9:513.
43 Friday, B. B., Horsley, V. and Pavlath, G. K. 2000. Calcineurinactivity is required for the initiation of skeletal muscle differenti-ation. J. Cell Biol. 149:657.
44 Tothova, Z., Kollipara, R., Huntly, B. J. et al. 2007. FoxOs arecritical mediators of hematopoietic stem cell resistance tophysiologic oxidative stress. Cell 128:325.
45 Dengler, H. S., Baracho, G. V., Omori, S. A. et al. 2008. Distinctfunctions for the transcription factor Foxo1 at various stages of Bcell differentiation. Nat. Immunol. 9:1388.
46 Amin, R. H. and Schlissel, M. S. 2008. Foxo1 directly regulates thetranscription of recombination-activating genes during B celldevelopment. Nat. Immunol. 9:613.
Foxo3 in B cell development and activation 841
Downloaded from https://academic.oup.com/intimm/article-abstract/21/7/831/820361by gueston 09 April 2018

47 Herzog, S., Hug, E., Meixlsperger, S. et al. 2008. SLP-65 regulatesimmunoglobulin light chain gene recombination through thePI(3)K-PKB-Foxo pathway. Nat. Immunol. 9:623.
48 Fabre, S., Carrette, F., Chen, J. et al. 2008. FOXO1 regulates L-Selectin and a network of human T cell homing moleculesdownstream of phosphatidylinositol 3-kinase. J. Immunol.181:2980.
49 Lo, C. G., Xu, Y., Proia, R. L. and Cyster, J. G. 2005. Cyclicalmodulation of sphingosine-1-phosphate receptor 1 surfaceexpression during lymphocyte recirculation and relationship tolymphoid organ transit. J. Exp. Med. 201:291.
50 Pham, T. H., Okada, T., Matloubian, M., Lo, G. C. and Cyster, J. G.2008. S1P1 receptor signaling overrides retention mediated byG alpha i-coupled receptors to promote T cell egress. Immunity28:122.
51 Ticchioni, M., Essafi, M., Jeandel, P. Y. et al. 2007. Homeostaticchemokines increase survival of B-chronic lymphocytic leukemiacells through inactivation of transcription factor Foxo3a. Onco-gene 26:7081.
52 Fernandez de Mattos, S., Essafi, A., Soeiro, I. et al. 2004. Foxo3aand BCR-ABL regulate cyclin D2 transcription through a STAT5/BCL6-dependent mechanism. Mol. Cell. Biol. 24:10058.
53 Essafi, A., Fernandez de Mattos, S., Hassen, Y. A. M. et al. 2005.Direct transcriptional regulation of Bim by Foxo3a mediatesSTI571-induced apoptosis in Bcr-Abl-expressing cells. Oncogene24:2317.
54 Rimokh, R., Rouault, J. P., Wahbi, K. et al. 2005. A chromosome 12coding region is juxtaposed to the MYC protooncogene locus ina t(8;12)(q24;q22) translocation in a case of B-cell chroniclymphocytic leukemia. Genes Chromosomes Cancer 3:24.
55 Islam, T. C., Lindvall, J., Wennborg, A., Branden, L. J., Rabbani, H.and Smith, C. I. E. 2002. Expression profiling in transformedhuman B cell: influence of Btk mutations and comparison to B celllymphomas using filter and oligonucleotide arrays. Eur.J. Immunol. 32:982.
56 Kuo, C. and Lin, S. 2007. Altered FOXO1 transcript levels inperipheral blood mononuclear cells of systemic lupus erythema-tosus and rheumatoid arthritis patients. Mol. Med. 13:561.
842 Foxo3 in B cell development and activation
Downloaded from https://academic.oup.com/intimm/article-abstract/21/7/831/820361by gueston 09 April 2018