food research international - nestle.com.br
TRANSCRIPT

Probiotic infant cereal improves children’s gut microbiota: Insights using the Simulator of Human Intestinal Microbial Ecosystem (SHIME®)
Mateus Kawata Salgaço a, Natalia Partis Perina b, Thaís Moreno Tome b, Elaine Martins Bento Mosquera b, Tamara Lazarini b, Adilson Sartoratto c, Katia Sivieri a,*
a Department of Food and Nutrition, School of Pharmaceutical Science, Sao Paulo State University-UNESP, Araraquara, Sao Paulo, Brazil b Medical, Scientific and Regulatory Affairs – Nestle Nutrition/Nestle Brazil Ltda, Sao Paulo, Brazil c CPQBA-UNICAMP, Brazil
A R T I C L E I N F O
Keywords: Intestinal metabolites SCFAs Bifidobacterium lactis Microbiota SHIME® model Infant cereal
A B S T R A C T
Infants gut microbiota can be modulated by many factors, including mode of delivery, feeding regime, maternal diet/weight and probiotic and prebiotic consumption. The gut microbiota in dysbiosis has been associated with innumerous diseases. In this sense, early childhood intestinal microbiome modulation can be a strategy for disease prevention. This study had the purpose to evaluate the effect of an infant cereal with probiotic (Bifido-bacterium animalis ssp. lactis BB-12®) on infants intestinal microbiota using SHIME®, which simulates human gastrointestinal conditions. The ascending colon was inoculated with fecal microbiota from three children (2–3 years old). NH+4 , short chain fatty acids (SCFASs) and microbiota composition were determined by selective ion electrode, GC/MS and 16S sequencing, respectively. After treatment, butyric acid production increased (p <0.05) 52% and a decrease in NH+4 production was observed (p < 0.01). The treatment stimulated an increase (p < 0.01) of Lactobacillaceae families, more precisely L. gasseri and L. kefiri. L. gasseri has been associated with the prevention of allergic rhinitis in children and L. kefiri in the prevention of obesity. Thus, infant cereal with BB- 12® is able to stimulate the growth of L. gasseri and L. kefiri in a beneficial way, reducing NH+4 and increasing the production of SCFAs, especially butyric acid, in SHIME®.
1. Introduction
The intestinal microbiota consists of a complex and diverse system of microorganisms that colonize the gastrointestinal tract (GT) (Frick & Autenrieth, 2012). The number of microorganisms that inhabit the GT has been estimated in the literature at over 1014, which is ~10 times the number of human cells and over 100 times the amount of genomic content in the human genome (Thursby & Juge, 2017). The count of microorganisms present in the stomach is usually below 103 CFU/mL due to acidic pH, with a significant increase in the small intestine (104 to 107 CFU/mL) and higher concentrations in the colon region (1012 CFU/ mL) due to favorable conditions such as slow intestinal transit time, nutrient availability and favorable pH (Payne, Zihler, Chassard, & Lacroix, 2012). The intestinal microbiota is mainly composed of strict anaerobic bacteria, which outperform the facultative anaerobic and aerobic bacteria (Walsh, Guinane, O’Toole, & Cotter, 2014). The most abundant species are the members of phyla Firmicutes, Bacteroidetes,
Proteobacteria and Actinobacteria (Gerritsen, Smidt, Rijkers, & De Vos, 2011).
Microbiota at the beginning of life has low microbial diversity and instability, however this same microbiota evolves to stability in the first 24 months of life (Rautava, Luoto, Salminen, & Isolauri, 2012). Bifido-bacteria are generally dominant in the first months of life, especially in breastfed children, due to the bifidogenic effect of breast milk. Meta-genomic analyses show that in adults and children the main components of colon microbiota are Bacteroidetes, followed by various genera belonging to phylum Firmicutes, such as Eubacterium spp., Ruminococcus spp. and Clostridium spp. The composition of intestinal microbiota is influenced by several factors, including the form of delivery (cesarean section or normal delivery), feeding, host genetics, use of antibiotics, and immunological factors (Russell et al., 2011; Scott, Gratz, Sheridan, Flint, & Duncan, 2013). Diet has a significant impact on the intestinal environment, including intestinal transit time, pH and changes in the composition of the microbial community (Scott et al., 2013).
* Corresponding author at: Rodovia Araraquara Jau/Km 1, Brazil. E-mail address: [email protected] (K. Sivieri).
Contents lists available at ScienceDirect
Food Research International
journal homepage: www.elsevier.com/locate/foodres
https://doi.org/10.1016/j.foodres.2021.110292 Received 22 October 2020; Received in revised form 27 February 2021; Accepted 27 February 2021

Intestinal microbiota plays a key role in the host’s health status, since it exerts important functions on the immunological, physiological and metabolic processes of the human body (Gerritsen et al., 2011), espe-cially in the first 1000 days, the period from conception to the 2nd year of life, which is a critical window period with implications for long-term health, programming health and the future risk of illness of an individual (Butel, Waligora-Dupriet, & Wydau-Dematteis, 2018). Microbiota acts on the synthesis of B and K vitamins, on resistance against colonization of pathogenic microorganisms and on short chain fatty acid (SCFAs) synthesis (Davila et al., 2013; Gerritsen et al., 2011; Sekirov, Russell, Antunes, & Finlay, 2010). In addition, it is responsible for modulating the immune system, promoting maturation of immune cells and main-tenance of motor functions of the gastrointestinal tract (Clemente, Ursell, Parfrey, & Knight, 2012; Round & Mazmanian, 2009; Zhuang et al., 2019).
Species and strains representing Lactobacillaceae families and Bifi-dobacteria have been used as probiotics, with the aim of colonizing the intestine of children and modulating the host’s immune response (Enomoto et al., 2014). According to Hill et al. (2014) the term “pro-biotics” says that microorganisms confer health benefits to individuals when administered in appropriate amounts, the true probiotic should preferably be of human origin, safe and free of vectors capable of transferring antibiotic resistance and pathogenicity or toxicity factors. In addition, the probiotic must have a high survival capacity under intes-tinal conditions (acid pH, enzymes, bile salts, etc.). It must also exhibit antagonism against pathogens, stimulate the immune system and, finally, have demonstrable beneficial effects on the host. The continuous activity, viability and growth efficiency of the probiotic must be demonstrated after the technological treatment (Plaza-Díaz, Fontana, & Gil, 2018; Plaza-Diaz, Ruiz-Ojeda, Gil-Campos, & Gil, 2019).
Several studies have demonstrated benefits of treatment with pro-biotics in atopic children and therefore modulation of the baby’s intes-tinal microbiota (Vandenplas, Huys, & Daube, 2015; Rather et al., 2016; D’Elios et al., 2020; Andrade et al., 2020). In addition, evidence from different studies has shown that the occurrence of a disease is often preceded by early changes in microbiota (Zhuang et al., 2019).
Bifidobacteria are considered one of the key genera of the intestinal tract, representing approximately 3% of the microbiota in healthy adult humans
(Solano-Aguilar et al., 2008). BB-12® has been widely used in infant formula, food supplements and fermented dairy products around the world. This strain is technologically suitable, expressing fermentation activity, high aerotolerance, good stability and high tolerance to acids and bile, also as lyophilized products in food supplements. Furthermore, BB-12® has no implications on taste, appearance or mouthfeel of foods and is able to survive in probiotic foods until consumed (Jungersen et al., 2014).
The health benefits of BB-12® includes, for example, bile salt hy-drolase and strong adhesion properties, demonstrated through clinical research (Laparra & Sanz, 2009; Matsumoto, Ohishi, & Benno, 2004; Vernazza, Gibson, & Rastall, 2006; Vinderola & Reinheimer, 2003). Besides the inhibition of pathogens, it is known that BB-12® promotes an improvement in epithelial barrier function and immune interactions, a better intestinal function and consequently a protective effect against diarrhea, thus reducing the side effects of antibiotic treatment (Collado, Grzeskowiak, & Salminen, 2007; Lopez, Gueimonde, Margolles, & Suarez, 2010; Martins et al., 2009). In terms of immune function, clinical studies have shown that BB-12® increases body’s resistance to common respiratory tract infections, reducing the incidence of acute respiratory tract infections (Rizzardini et al., 2012).
Probiotic microorganisms are usually consumed as dairy products, such as fermented milk and yogurts (Hekmat, Soltani, & Reid, 2009). However, alternative food matrices, such as infant cereals, could be a good option as vehicles for probiotic consumption, particularly for in-fants and young children, as infant cereal represents one of the first foods introduced at the beginning of complementary feeding period in
many countries (Klerks et al., 2019; Roess et al., 2018). Infant cereals represent an excellent source of energy, providing
substantial amounts of carbohydrates and protein (Agostoni et al., 2008) and also contribute as an important source of vitamins and minerals (Fardet, 2010), specially, iron (Finn et al., 2017; Grimes, Szymlek-Gay, Campbell, & Nicklas, 2015).
In vitro fermentation studies have been developed as tools to study human gut microbiota under highly controlled conditions, thus allowing dynamic sampling over time in reactors that mimic different regions of the colon. The colonic fermentation models have advantages when compared with clinical trials or animal models such as low cost, higher reproducibility, no need of ethical approval and, depending on the experiment, they can be conducted in a shorter time. Colonic models enable the cultivation of human gut microbiota derived from fecal samples under simulated physiological conditions (Pham & Mohajeri, 2018). Thus, these systems allow researchers to study the effects of prebiotics and probiotics on gut microbiota, the fermentation spectra of prebiotics, the survival capacity and function of probiotics throughout the gastrointestinal tract (Bianchi et al., 2018), and the synthesis of polyphenols that modulate the microbial community (Wu et al., 2018). Our research group has been using the Simulator of Human Intestinal Microbial Ecosystem (SHIME®) for several years. Thus, we were able to advance in the studies correlating intestinal microbiota, probiotics, prebiotics and bioactive compounds, contributing to scientific knowl-edge on the following topics: the impact of Enterococcus faecium CRL 183 (Sivieri et al., 2014) and Lactobacillus acidophilus CRL 1014 on the in-testinal microbiota (Sivieri et al., 2013), influence of a fermented vegetable drink with Lacticaseibacillus casei on the intestinal microbiota (Bianchi et al., 2014), as well as the prebiotic effect of fructo- oligosaccharide (Sivieri et al., 2014), “multifunctional” milk-based drink of fermented goat (Freire et al., 2017) and probiotic ice cream (Rodrigues et al., 2020) in the modulation of the intestinal microbiota.
Therefore, the evaluation of the influence of infant cereals on the modulation of the intestinal microbiota is still little explored. As so, the aim of this work was to evaluate the effect of infant cereal with Bifido-bacterium animalis ssp. lactis BB-12® on children’s gut microbiota using the Simulator of Human Intestinal Microbial Ecosystem (SHIME®), which simulates human gastrointestinal conditions, from the stomach portion to colon.
2. Experimental methods
2.1. Simulated digestion in the dynamic colonic model
The Simulator of Human Intestinal Microbial Ecosystem (SHIME®) is a computer controlled simulator and consists of 5 closed compartments representing the stomach, small intestine, ascending colon, transverse colon and descending colon (Molly, Woestyne, Smet, & Verstraete, 1994). In this experiment, SHIME® was adapted for the triplication of the ascending colon, where the transverse and descending colon have been substituted according to Rodrigues et al. (2020).
The volumetric capacity, pH, temperature (37 ◦C) and retention time (24 h) were controlled (Possemiers, Verthe, Uyttendaele, & Verstraete, 2004). Anaerobiosis of the system was achieved by the addition of ni-trogen and the pH value corrected in each vessel using hydrochloric acid or sodium hydroxide accordingly, to be in the range from 5.6 to 5.9 (Molly et al., 1994; Possemiers et al., 2004).
The compartments were colonized with feces from three healthy children volunteers, inclusion criteria were ages 2–3 years, without food allergy or intolerance and exclusion criteria were dietary supplements and medication for gastrointestinal or metabolic disease, probiotics or prebiotics in the last 3 months, and antibiotics in the last 6 months. 24- hour total feces were collected and kept at 4 ◦C until its collection by the researcher. They were homogenized and a portion was stored in a sterile plastic tube (10 g) and held at − 80 ◦C until analysis. According to Possemiers, Marzorati, Verstraete, and Van de Wiele (2010) the 40 g of
M.K. Salgaço et al.

feces were homogenized and diluted in 200 mL of phosphate buffer (pH 6.5), composed of 7.08 g/L of monosodium phosphate (Synth, SP, Brazil), 5.98 g/L of disodium phosphate (Synth, SP, Brazil) and 1 g/L of sodium thioglycollate (Merck, Brazil). This mixture was then homoge-nized in a stomacher and centrifuged at 3000g for 15 min. The super-natant was collected and 10 mL added to each of the last 3 compartments together with 500 mL of sterile feed medium (van den Abbeele et al., 2019), which is a carbohydrate-based medium [4.0 g/L starch (Maizena, Sao Paulo,BR), 4.0 g/L mucin (Sigma, St. Louis, USA), 3.0 g/L yeast extract (Difco,Becton Dickinson Co., Sparks, MD, USA), 1.0 g/L arabinogalactan (Sigma, St. Louis, USA), 1.0 g/L xylan (Sigma, St. Louis, USA), 1.0 g/L peptone (Himedia, Mumbai, India), 0.5 g/L cysteine (Sigma, St. Louis, USA), and 0.4 g/L glucose (Synth, Diadema, BR]. Stomach conditions were simulated in reactor 1 by adding HCl for the regulation of pH, along with the carbohydrate-based medium. The passage through the duodenum was simulated by 60 mL of artificial pancreatic juice (12.5 g/L of NaHCO3, 6 g/L of Oxgall and 1.9 g/L of pancreatin, Sigma-Aldrich, St. Louis, USA) with an important role in environmental adaptation and inoculum growth, with the formation of a stable and representative community (Molly et al., 1994).
2.2. Experimental protocol
The experimental period in the SHIME® reactor lasted 4 weeks. For the microbiota stabilization period, the feed medium (240 mL) was inserted into the system and left to stabilize for 14 days (Bianchi et al., 2018). After these two weeks, for one week, daily, the 210 mL of feed medium and 60 mL of pancreatic juice was inserted in the system (control period). Afterward, the treatment was administered two times a day for 7 days. Treatment: 10.5 g of infant cereal with probiotic (Bifi-dobacterium animalis ssp. lactis- BB12®, 2.7 × 107 cfu.log/g, that cor-responds to 5.4 × 108 cfu.log per portion) (Nestle, Brazil) prepared with 85 mL of whole cow’s milk (Ninho, Nestle, Brazil). The experiment was carried out in biological triplicate.
2.3. Metabolic Activity: Ammonium (NH+4 ) analysis and short chain fatty acids (SCFASs)
After the simulated digestion, the samples from the colon compart-ment (n = 3) were collected and stored at − 20 ◦C. The ammonium ions (NH+4 ) were quantified according to Bianchi et al. (2014) using a specific ion meter (Model 710A, Orion) coupled to a selective ammonia ion electrode (Model 95-12, Orion).
Short chain fatty acids were analyzed according to the protocol adopted by Duque, Monteiro, Adorno, Sakamoto, and Sivieri (2016), with modifications. The samples (n = 3, second week of the colon re-actors) were centrifuged (14,000g, 5 min) followed by the dilution of 100 μL of the supernatant in 1900 μL of ultrapurewater. Next, NaCl (1g) and crotonic acid (100 μL) were added, as well as isobutanol (70 μL) and 2 M H2SO4 (200 μL). Analytical curves were constructed from stock solutions of the acids of interest (acetic, propionic and butyric). The SCFAs analysis was conducted using a HP-5975 model gas chromato-graph (Agilent Technologies, Santa Clara-CA, USA) equipped with a split/splitless injector, a mass selective detector and a HP-7683B auto-mated sampler. Separation of the SCFAs took place through a ZB-WAX column (60 m × 0.25 mm × 0.25 μm) (Phenomenex, Torrance-CA, USA). The GC was performed using the column set at 35 ◦C, 2 ◦C/min, 38 ◦C; 10 ◦C/min, 75 ◦C; 35 ◦C/min, 120 ◦C (1 min); 10 ◦C/min, 170 ◦C (2 min); 40 ◦C/min, 170 ◦C (2 min), and the temperature of the injector and the detector was maintained at 220 ◦C and 250 ◦C, respectively. Helium was used as the carrier gas, the flow rate was set at 1 mL/min (Hoving, Heijink, van Harmelen, van Dijk, & Giera, 2018).
2.4. Survival of Bifidobacterium animalis ssp. lactis –BB12® under simulated conditions of the stomach and duodenum in the SHIME®
During treatment period with the probiotic infant cereal formula-tion, samples were collected from the reactors corresponding to the stomach and duodenum in order to verify the survival of Bifidobacterium animalis ssp. lactis –BB12®. One mL of samples from each reactor were suspended in 9 mL of sterile peptone water. Serial dilutions were carried and plated in MRS agar supplemented with sodium propionate (3 g.L-1) (Sigma-Aldrich®, USA) and lithium chloride (2 g.L-1) (Merck®, Ger-many). The plates were incubated in anaerobiosis (Probac, Brazil) at 37 ◦C for 72 h (Vinderola & Reinheimer, 1999).
2.5. Microbiological analysis employing 16S rRNA gene sequencing
The microbiological analysis employing 16S rRNA gene sequencing (n = 2) was performed by Next-generation Sequencing (Neoprospecta Microbiome Technologies, Brazil) on what specific primers amplified the V3–V4 region of the 16S rRNA, 341F and 806R of 1 ng of DNA (Caporaso, Lauber, Walters, Berg-Lyons, Lozupone, Turnbaugh, Fierer, & Knight, 2011; Wang & Qian, 2009). The analysis of the sequences and the identification of the taxonomic units were based on the methodology of Giongo et al. (2010) and Hong et al., (2006). The PCR were carried out in triplicate using a Platinum Taq (Invitrogen USA) under the following conditions: initial denaturation at 94 ◦C for 3 min; 20 cycles of denaturation at 94 ◦C for 45 s, annealing at 50 ◦C for 30 s, extension at 65 ◦C for 90 s and final extension at 65 ◦C for 10 min. The PCR products are purified using AMPure beads (Beckman coulter) (Christoff, Fer-nanda, Sereia, Rodrigues, Lucio, & Moraes, 2017) and quantified by fluorometry using Qubit® 2.0 Fluorometer (Invitrogen, USA) (Fagen et al., 2012). The sequencing libraries were prepared according to Neoprospecta Microbiome Technologies, and the sequencing was car-ried out using the MiSeq platform (Illumina).
Bioinformatics analyzes were performed using the QIIME program (Quantitative Insights Into Microbial Ecology) according to procedures described by Caporaso et al. (2010). Taxonomic descriptions were generated based on the database NCBI taxonomy (http://www.ncbi.nl m.nih.gov) for the RDP database entries using the set of scripts listed in the TaxCollector pipeline Giongo et al. (2010). The strings were filtered and classified according to similarity. A minimum of 80% sim-ilarity was considered for domain and phylum identification, 90% for order and class identification, 90% for family, 95% for gender and 99% similarity for species identification (Savy et al., 2018).
2.6. Statistical analysis
Comparison of normally distributed data of the different control and treatment weeks on microbial metabolic markers and microbial com-munity parameters were performed with a Student’s T-test for pairwise comparisons. Differences were significant if p < 0.05, using the statis-tical software Prism 7.0 (Software MacKiev© 1994–2016).
3. Results
3.1. Metabolic activity
Fermentation activity correlated with NH+4 and SCFAs profiles (ac-etate, propionate and butyrate), as shown in Fig. 1. A reduction (p <0.05) in the production of ammonia ions was observed in treatment studied with values below 10 ppm.
Levels of acetic (14.76–12.42 mmol/L) and propionic acid (5.69–2.03 mmol/L) had a decrease during the treatment with probiotic infant cereal when compared to control. Therefore, butyrate levels significantly increased (p < 0.001) (0.89–1.70 mmol/L) in the ascending colon during treatment (see Fig. 2.).
M.K. Salgaço et al.
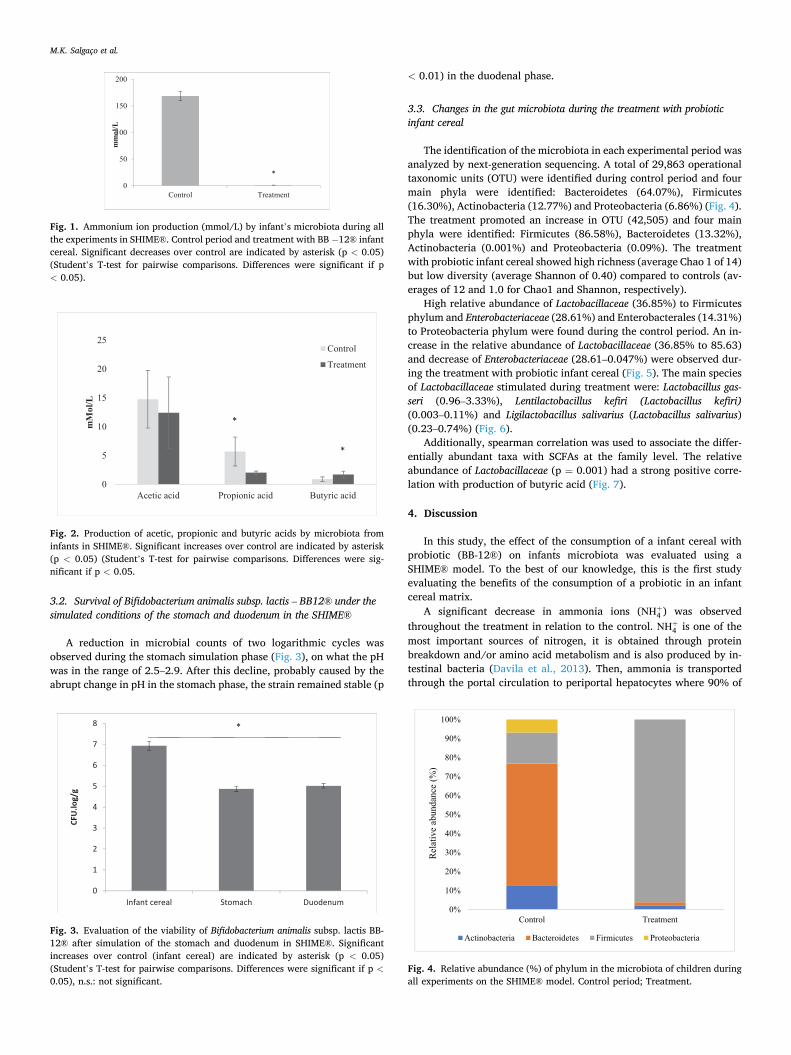
3.2. Survival of Bifidobacterium animalis subsp. lactis – BB12® under the simulated conditions of the stomach and duodenum in the SHIME®
A reduction in microbial counts of two logarithmic cycles was observed during the stomach simulation phase (Fig. 3), on what the pH was in the range of 2.5–2.9. After this decline, probably caused by the abrupt change in pH in the stomach phase, the strain remained stable (p
< 0.01) in the duodenal phase.
3.3. Changes in the gut microbiota during the treatment with probiotic infant cereal
The identification of the microbiota in each experimental period was analyzed by next-generation sequencing. A total of 29,863 operational taxonomic units (OTU) were identified during control period and four main phyla were identified: Bacteroidetes (64.07%), Firmicutes (16.30%), Actinobacteria (12.77%) and Proteobacteria (6.86%) (Fig. 4). The treatment promoted an increase in OTU (42,505) and four main phyla were identified: Firmicutes (86.58%), Bacteroidetes (13.32%), Actinobacteria (0.001%) and Proteobacteria (0.09%). The treatment with probiotic infant cereal showed high richness (average Chao 1 of 14) but low diversity (average Shannon of 0.40) compared to controls (av-erages of 12 and 1.0 for Chao1 and Shannon, respectively).
High relative abundance of Lactobacillaceae (36.85%) to Firmicutes phylum and Enterobacteriaceae (28.61%) and Enterobacterales (14.31%) to Proteobacteria phylum were found during the control period. An in-crease in the relative abundance of Lactobacillaceae (36.85% to 85.63) and decrease of Enterobacteriaceae (28.61–0.047%) were observed dur-ing the treatment with probiotic infant cereal (Fig. 5). The main species of Lactobacillaceae stimulated during treatment were: Lactobacillus gas-seri (0.96–3.33%), Lentilactobacillus kefiri (Lactobacillus kefiri) (0.003–0.11%) and Ligilactobacillus salivarius (Lactobacillus salivarius) (0.23–0.74%) (Fig. 6).
Additionally, spearman correlation was used to associate the differ-entially abundant taxa with SCFAs at the family level. The relative abundance of Lactobacillaceae (p = 0.001) had a strong positive corre-lation with production of butyric acid (Fig. 7).
4. Discussion
In this study, the effect of the consumption of a infant cereal with probiotic (BB-12®) on infants microbiota was evaluated using a SHIME® model. To the best of our knowledge, this is the first study evaluating the benefits of the consumption of a probiotic in an infant cereal matrix.
A significant decrease in ammonia ions (NH+4 ) was observed throughout the treatment in relation to the control. NH+4 is one of the most important sources of nitrogen, it is obtained through protein breakdown and/or amino acid metabolism and is also produced by in-testinal bacteria (Davila et al., 2013). Then, ammonia is transported through the portal circulation to periportal hepatocytes where 90% of
Fig. 1. Ammonium ion production (mmol/L) by infant’s microbiota during all the experiments in SHIME®. Control period and treatment with BB − 12® infant cereal. Significant decreases over control are indicated by asterisk (p < 0.05) (Student’s T-test for pairwise comparisons. Differences were significant if p < 0.05).
Fig. 2. Production of acetic, propionic and butyric acids by microbiota from infants in SHIME®. Significant increases over control are indicated by asterisk (p < 0.05) (Student’s T-test for pairwise comparisons. Differences were sig-nificant if p < 0.05.
Fig. 3. Evaluation of the viability of Bifidobacterium animalis subsp. lactis BB- 12® after simulation of the stomach and duodenum in SHIME®. Significant increases over control (infant cereal) are indicated by asterisk (p < 0.05) (Student’s T-test for pairwise comparisons. Differences were significant if p <0.05), n.s.: not significant.
Fig. 4. Relative abundance (%) of phylum in the microbiota of children during all experiments on the SHIME® model. Control period; Treatment.
M.K. Salgaço et al.

the ammonia enters the urea cycle and is converted into urea. Yet, ammonia is not excreted, due to its low water solubility, the remaining 10% is transported to perennial hepatocytes where the ammonia is condensed with glutamate to glutamine through glutamine synthetase (GS) (Savy et al., 2018). According to Scott et al. (2013), the release of ammonia ions may be associated with increased metabolic activity of some species of Bifidobacterium, among others, that participate in the deamination processes. The reduction of ammonia in the colon is considered beneficial, because these ions in high quantity can alter the morphology, beyond the metabolism of the intestinal cells, increasing the synthesis of DNA and promoting the tumoral development (Davila et al., 2013). Hughes, Kurth, McGilligan, McGlynn, and Rowland (2008) demonstrated that ammonia can increase the cellular permeability in the colonocytes, causing several diseases in the host. The presence of
ammonia can reduce the absorption and use of short chain fatty acids (SCFAs) by the colonocytes (He et al., 2019). Furthermore, the blood level of ammonia should remain very low because even slightly elevated concentrations (hyperammonemia) are toxic to the central nervous system. The peak ammonia level and duration of hyperammonemia are the main risk factors for neurological deficits related to hyper-ammonemia and death. In children, hyperammonemia is mainly caused by severe liver failure and congenital metabolism errors (Mohiuddin & Khattar, 2020).
According to Cinquin, Le Blay, Fliss, and Lacroix (2006) the SCFAs proportion (acetic:propionic:butyric) observed in infants feces (ratio close to 75:19:6) is different than those observed previously in adults feces (ratio close to 60:20:20) (Bridgman et al., 2017). Interestingly, the SCFAs proportion observed in the colon vessels in control period was
Fig. 5. Relative abundance of bacterial family (%) in the microbiota of children during all experiments on the SHIME® model. Control period; Treatment.
M.K. Salgaço et al.

very similar to adult profile (69:26:4), however, after infant cereal treatment, it changed to 76:12:25. Therefore, a significant increase of butyric acid and decrease of propionic was observed during treatment with infant cereal with probiotic.
The main products of bacterial fermentation in the intestine are SCFAs, that are among the most important components of host micro-organism signaling, are capable of modulating gene expression in host cells, brain function and behavior, as well as modulate host energy metabolism and immune functions (Kim, Qie, Park, & Kim, 2016; Russo et al., 2016). The amount and relative proportion of metabolites pro-duced in the colon of adults and infants have been closely linked to some factors such as diet, as well as the composition, diversity and activity of microbes (Bridgman et al., 2017).
Butyrate has beneficial effects on human intestinal homeostasis (Canani et al., 2011; Macia et al., 2015), host immunological functions (Kim et al., 2016) and is considered a resource compound with prom-ising effects on various neurological and neuropsychiatric disorders, besides being the main source of energy for colonocytes and be involved in maintaining colonic health (Bourassa, Alim, Bultman, & Ratan, 2016; Dinan & Cryan, 2017; Stilling et al., 2016). A study in animal model suggested a dose dependence on the effects of butyrate on brain devel-opment and function (Gagliano, Delgado-Morales, Sanz-Garcia, & Armario, 2014; Kratsman, Getselter, & Elliott, 2016; Takuma et al., 2014). In addition, according to Roduit et al. (2019) children with the highest levels of butyrate were less likely to have a reported diagnosis of food allergy or allergic rhinitis and were less likely to have asthma be-tween 3 and 6 years.
Although propionate decreased during treatment, the concentration
was very similar to that found by Cinquin et al. (2006), in infants feces. An excess of propionate and an inability for it to convert to methyl-malonyl CoA through propionyl-CoA causes propionic acidemia (Hos-seini, Grootaert, Verstraete, & Van de Wiele, 2011). However, beneficial effects were attributed to propionate, such as; decreased serum choles-terol levels (Illman et al., 1988), upregulation of GLP-1 (Zhou et al., 2008), greater feeling of fullness, less hunger and reduced desire to eat (Ruijschop, Boelrijk, & te Giffel, 2008).
In the stomach reactor, there was a decline in the viability of B. animalis - BB12®. However, this viability remained constant in the duodenum reactor. The presence of foods and food ingredients such as dietary fiber have a protective effect on the viability of microorganisms during their passage through the stomach and duodenum (Sendra, Sayas-Barbera, Fernandez-Lopez, & Perez-Alvarez, 2016). The infant cereal product reached the colon with 105 CFU.log/g. According to Hill et al. (2014), the recommended use of the term probiotic in products is when living microorganisms provide a minimum viable and sufficient count of well-defined strains and evaluated with a reasonable expecta-tion of providing benefits for the well-being of the host (ANVISA, Brazil, 2018). Survival capacity in aggressive GI conditions, overcoming the presence of a mixed solution with bile salts and other digestive factors, is crucial for host colonization by probiotics (Koskenniemi et al., 2011). Therefore, the infant cereal with probiotic proved to be a good matrix for carrying B. animalis - BB12®. In addition, bifidobacterial strains are frequently used as probiotic microorganisms and thus incorporated in several fermented products (Meira et al., 2015; Taverniti, Scabiosi, Arioli, Mora, & Guglielmetti, 2014; Verruck, Prudencio, Vieira, Amante, & de Mello Castanho Amboni, 2015).
The administration of an infant cereal with probiotic caused a posi-tive change in the pattern of relative abundance percentage of microbial family during the different experimental periods. Comparing with con-trol period, a high prevalence of Lactobacillaceae was observed after administration of the infant cereal with probiotic. It is important to highlight that the treatment and ascending colon conditions can favor the growth of saccharolytic bacteria such as Lactobacillaceae (Liu et al., 2018). Lactobacillaceae family showed a positive strong correlation with butyric acid production. Increasingly, whole grain is becoming one of the choices as a carrier for probiotics. This is mainly because the formulation of probiotics with cereals products offers consumers bene-fits from both probiotics and cereal. Thus, the use of cereals in formu-lations increases the dietary value of the product, as well as being used as fermentable substrates for intestinal bacteria (Lamsal & Faubion, 2009; Pereira et al., 2019).
Interestingly, the probiotic infant cereal treatment stimulated the increase of L. gasseri, which is involved in the prevention and treatment of children with asthma and allergic rhinitis (Chen et al., 2010). The
Fig. 6. Relative abundance of Lactobacillaceae family in the microbiota of children during all experiments on the SHIME® model. Control period; Treatment.
Fig. 7. Correlation between Short Chain Fat Acids (SCFAs) production (butyric and acetic acids), and bacterial family. Significant correlations are indicated by one asterisk (p < 0.05), two asterisks (p < 0.01), three asterisks (p < 0.001) (Spearman correlation).
M.K. Salgaço et al.

study conducted by Zubiría et al. (2017), showed that the administration of L. gasseri SBT2055 to mice fed a 10% fat diet resulted in reduced expression of pro-inflammatory genes such as CCL2 and CCR2 in fat tissue, preventing body weight gain and fat accumulation. An increase in L. kefiri and L. salivarius were also observed after the probiotic infant cereal treatment. Recently, L. kefiri was associated with obesity pre-vention and cholesterol control, and the mechanisms involved are the direct reduction of excessive lumen cholesterol to decrease the influx of fat and the positive regulation of genes coding for PPAR-α, CPT1 and FABP4 in fat tissue to increase the oxidation of fatty acids, moreover, L. kefiri is also associated with anti-obesity and anti-inflammatory effects (Kim et al., 2017). Supplementation with L. salivarius was associated with improvement of the quality of life of children affected by atopic dermatitis (Niccoli et al., 2014). It is important to highlight that atopic dermatitis is a chronic inflammatory skin disease with multifactorial etiopathology, which is most prevalent in childhood and appears in the first 5 years of life, with about 60% of cases appearing between 0 and 6 months (Monti et al., 2011).
However, some limitations were also observed, such as the duration of the study (7 days), the in vitro study and the lack of previous studies demonstrating the biological effects between the infant cereal and microbiota. Despite the limitations, we could conclude that the dynamic view of microbiota and microbial metabolites demonstrated by treat-ment with infant cereals with Bifidobacterium animalis ssp. lactis – BB12® opened new perspectives on the interaction of probiotics with micro-biota and metabolites. The results indicate that the infant cereal with probiotic has a positive impact on the microbiota with a decrease in the production of ammonia ions, increase in butyric acid, and stimulates the growth of L. kefiri, L. gasseri and L. salivarius with potential benefits to children and promoting a healthy microbiota. Finally, a clinical trial is essential to confirm the results found in the dynamic colonic reactor (SHIME®).
CRediT authorship contribution statement
Mateus Kawata Salgaço: Writing - original draft. Natalia Partis Perina: Methodology, Investigation. Thaís Moreno Tome: Methodol-ogy, Investigation. Elaine Martins Bento Mosquera: Methodology, Investigation. Tamara Lazarini: Methodology, Investigation. Adilson Sartoratto: Investigation. Katia Sivieri: Conceptualization, Methodol-ogy, Investigation, Writing - original draft.
Declaration of Competing Interest
The authors declare the following financial interests/personal re-lationships which may be considered as potential competing interests: ‘Natalia Partis Perina, Thaís Moreno Tome, Elaine Mosquera and Tam-ara Lazarini are employees of Nestle Brazil, that financially supported the study and manufactures the infant cereal used in this study’.
Acknowledgments
This work was supported by Nestle Brazil.
References
ANVISA, 2018. National Sanitary Vigilance Agency: Resolution – No. 241, 26 July 2018. Accessed: http://portal.anvisa.gov.br/documents/10181/3898888/RDC_241_2018_. pdf/941cda52-0657-46dd-af4b-47b4ee4335b7.
Andrade, P. D. M. S. A., Silva, J. M., Vanessa, C., Sacramento, L., Roberti, L., Aragon, D., & Junior, P. R. (2020). Probiotics improve atopic dermatitis in children and adolescents: A double blind, placebo-controlled study. Authorea, 10(1), 1–9. https:// doi.org/10.22541/au.158930908.86661145.
Agostoni, C., Decsi, T., Fewtrell, M., Goulet, O., Kolacek, S., Koletzko, B., … van Goudoever, J. (2008). Complementary feeding: A commentary by the ESPGHAN committee on nutrition. Journal of Pediatric Gastroenterology and Nutrition, 46(1), 99–110. https://doi.org/10.1097/01.mpg.0000304464.60788.bd.
Bianchi, F., Larsen, N., de Mello Tieghi, T., Adorno, M. A. T., Kot, W., Saad, S. M. I., … Sivieri, K. (2018). Modulation of gut microbiota from obese individuals by in vitro
fermentation of citrus pectin in combination with Bifidobacterium longum BB-46. Applied Microbiology and Biotechnology, 102(20), 8827–8840. https://doi.org/ 10.1007/s00253-018-9234-8.
Bianchi, F., Rossi, E. A., Sakamoto, I. K., Adorno, M. A. T., Van de Wiele, T., & Sivieri, K. (2014). Beneficial effects of fermented vegetal beverages on human gastrointestinal microbial ecosystem in a simulator. Food Research International, 64, 43–52. https:// doi.org/10.1016/j.foodres.2014.05.072.
Bourassa, M. W., Alim, I., Bultman, S. J., & Ratan, R. R. (2016). Butyrate, neuroepigenetics and the gut microbiome: Can a high fiber diet improve brain health? Neuroscience Letters, 625(3), 56–63. https://doi.org/10.1016/j. neulet.2016.02.009.
Bridgman, S. L., Azad, M. B., Field, C. J., Haqq, A. M., Becker, A. B., Mandhane, P. J., … Kozyrskyj, A. L. (2017). Fecal short-chain fatty acid variations by breastfeeding status in infants at 4 months: Differences in relative versus absolute concentrations. Frontiers Nutrition, 4(April). https://doi.org/10.3389/fnut.2017.00011.
Butel, M. J., Waligora-Dupriet, A. J., & Wydau-Dematteis, S. (2018). The developing gut microbiota and its consequences for health. Journal of Developmental Origins of Health and Disease, 9(6), 590–597. https://doi.org/10.1017/S2040174418000119.
Canani, R. B., Costanzo, M. Di, Leone, L., Pedata, M., Meli, R., & Calignano, A. (2011). Potential beneficial effects of butyrate in intestinal and extraintestinal diseases. World Journal of Gastroenterology, 17(12), 1519–1528. https://doi.org/10.3748/wjg. v17.i12.1519.
Caporaso, J. G., Kuczynski, J., Stombaugh, J., Bittinger, K., Bushman, F. D., … Knight, R. (2010). QIIME allows analysis of high-throughput community sequencing data. Nature Methods, 7(5), 335–336. https://doi.org/10.1038/nmeth.f.303.
Caporaso, J. G., Lauber, C. L., Walters, W. A., Berg-Lyons, D., Lozupone, C. A., Turnbaugh, P. J., … Knight, R. (2011). Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proceedings of the National Academy of Sciences, 108(Supplement_1), 4516–4522. https://doi.org/10.1073/ pnas.1000080107.
Chen, X., Tian, F., Liu, X., Zhao, J., Zhang, H. P., Zhang, H., & Chen, W. (2010). In vitro screening of lactobacilli with antagonistic activity against Helicobacter pylori from traditionally fermented foods. Journal of Dairy Science, 93(12), 5627–5634. https:// doi.org/10.3168/jds.2010-3449.
Christoff, A. P., Fernanda, A., Sereia, R., Rodrigues, D., Lucio, R., Moraes, … Oliveira, V. De. (2017). Bacterial identification through accurate library preparation and high- throughput sequencing. Neoprospecta Microbiome Technologies, (May 2017), 1–5.
Cinquin, C., Le Blay, G., Fliss, I., & Lacroix, C. (2006). New three-stage in vitro model for infant colonic fermentation with immobilized fecal microbiota. FEMS Microbiology Ecology, 57(2), 324–336. https://doi.org/10.1111/j.1574-6941.2006.00117.x.
Clemente, J. C., Ursell, L. K., Parfrey, L. W., & Knight, R. (2012). The impact of the gut microbiota on human health: An integrative view. Cell, 148(6), 1258–1270. https:// doi.org/10.1016/j.cell.2012.01.035.
Collado, M. C., Grzeskowiak, Ł., & Salminen, S. (2007). Probiotic strains and their combination inhibit in vitro adhesion of pathogens to pig intestinal mucosa. Current Microbiology, 55(3), 260–265. https://doi.org/10.1007/s00284-007-0144-8.
D’Elios, S., Trambusti, I., Verduci, E., Ferrante, G., Rosati, S., Marseglia, G. L., … Peroni, D. G. (2020). Probiotics in the prevention and treatment of atopic dermatitis. Pediatric Allergy and Immunology, 31(S26), 43–45. https://doi.org/10.1111/ pai.13364.
Davila, A. M., Blachier, F., Gotteland, M., Andriamihaja, M., Benetti, P. H., Sanz, Y., & Tome, D. (2013). Intestinal luminal nitrogen metabolism: Role of the gut microbiota and consequences for the host. Pharmacological Research, 68(1), 95–107. https://doi. org/10.1016/j.phrs.2012.11.005.
Dinan, T. G., & Cryan, J. F. (2017). The microbiome-gut-brain axis in health and disease. Gastroenterology Clinics of North America, 46(1), 77–89. https://doi.org/10.1016/j. gtc.2016.09.007.
Duque, A. L. R. F., Monteiro, M., Adorno, M. A. T., Sakamoto, I. K., & Sivieri, K. (2016). An exploratory study on the influence of orange juice on gut microbiota using a dynamic colonic model. Food Research International, 84, 160–169. https://doi.org/ 10.1016/j.foodres.2016.03.028.
Enomoto, T., Sowa, M., Nishimori, K., Shimazu, S., Yoshida, A., Yamada, K., … Xiao, J. Z. (2014). Effects of bifidobacterial Bifidobacterial supplementation to pregnant women and infants in the prevention of allergy development in infants and on fecal microbiota. Allergology International, 63(4), 575–585. https://doi.org/10.2332/ allergolint.13-OA-0683.
Fagen, J. R., Giongo, A., Brown, C. T., Davis-Richardson, A. G., Gano, A. K., & Triplett, E. W. (2012). Characterization of the relative abundance of the citrus pathogen Ca. Liberibacter asiaticus in the microbiome of its insect vector, Diaphorina citri, using high throughput 16s rRNA sequencing. Open Microbiology Journal, 6(29–33), 2012. https://doi.org/10.2174/1874285801206010029.
Fardet, A. (2010). New hypotheses for the health-protective mechanisms of whole-grain cereals: What is beyond fibre? Nutrition Research Reviews, 23(1), 65–134. https://doi. org/10.1017/S0954422410000041.
Finn, K., Callen, C., Bhatia, J., Reidy, K., Bechard, L. J., & Carvalho, R. (2017). Importance of dietary sources of iron in infants and toddlers: Lessons from the FITS Study. Nutrients, 9(7), 16–19. https://doi.org/10.3390/nu9070733.
Freire, F. C., Adorno, M. A. T., Sakamoto, I. K., Antoniassi, R., Chaves, A. C. S. D., dos Santos, K. M. O., & Sivieri, K. (2017). Impact of multi-functional fermented goat milk beverage on gut microbiota in a dynamic colon model. Food Research International, 99(May), 315–327. https://doi.org/10.1016/j.foodres.2017.05.028.
Frick, J. -S., & Autenrieth, I. B. (2012). The gut microflora and its variety of roles in health and disease. In Assessment & evaluation in higher education (Vol. 37, pp. 273–289). https://doi.org/10.1007/82_2012_217.
Gagliano, H., Delgado-Morales, R., Sanz-Garcia, A., & Armario, A. (2014). High doses of the histone deacetylase inhibitor sodium butyrate trigger a stress-like response.
M.K. Salgaço et al.

Neuropharmacology, 79, 75–82. https://doi.org/10.1016/j. neuropharm.2013.10.031.
Gerritsen, J., Smidt, H., Rijkers, G. T., & De Vos, W. M. (2011). Intestinal microbiota in human health and disease: The impact of probiotics. Genes and Nutrition, 6(3), 209–240. https://doi.org/10.1007/s12263-011-0229-7.
Giongo, A., Davis-Richardsom, A., Crabb, D. B., & Triplett. (2010). TaxCollector: Modifying current 16S rRNA databases for the rapid classification at six taxonomic levels. Diversity, 2(7), 1015–1025. https://doi.org/10.3390/d2071015.
Grimes, C. A., Szymlek-Gay, E. A., Campbell, K. J., & Nicklas, T. A. (2015). Food sources of total energy and nutrients among U.S. infants and toddlers: National Health and Nutrition Examination Survey 2005–2012. Nutrients, 7(8), 6797–6836. https://doi. org/10.3390/nu7085310.
He, X., Parenti, M., Grip, T., Lonnerdal, B., Timby, N., Domellof, M., … Slupsky, C. M. (2019). Fecal microbiome and metabolome of infants fed bovine MFGM supplemented formula or standard formula with breast-fed infants as reference: A randomized controlled trial. Scientific Reports, 9(1), 1–14. https://doi.org/10.1038/ s41598-019-47953-4.
Hekmat, S., Soltani, H., & Reid, G. (2009). Growth and survival of Lactobacillus reuteri RC-14 and Lactobacillus rhamnosus GR-1 in yogurt for use as a functional food. Innovative Food Science and Emerging Technologies, 10(2), 293–296. https://doi.org/ 10.1016/j.ifset.2008.10.007.
Hill, C., Guarner, F., Reid, G., Gibson, G. R., Merenstein, D. J., Pot, B., … Sanders, M. E. (2014). Expert consensus document: The international scientific association for probiotics and prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nature Reviews Gastroenterology and Hepatology, 11(8), 506–514. https://doi.org/10.1038/nrgastro.2014.66.
Hong, S. H., Bunge, J., Jeon, S. O., & Epstein, S. S. (2006). Predicting microbial species richness. Proceedings of the National Academy of Sciences of the United States of America, 103(1), 117–122. https://doi.org/10.1073/pnas.0507245102.
Hosseini, E., Grootaert, C., Verstraete, W., & Van de Wiele, T. (2011). Propionate as a health-promoting microbial metabolite in the human gut. Nutrition Reviews, 69(5), 245–258. https://doi.org/10.1111/j.1753-4887.2011.00388.x.
Hoving, L. R., Heijink, M., van Harmelen, V., van Dijk, K. W., & Giera, M. (2018). GC-MS analysis of short-chain fatty acids in feces, cecum content, and blood samples. Clinical Metabolomics, 1730, 247–256. https://doi.org/10.1007/978-1-4939-7592-1_ 17.
Hughes, R., Kurth, M. J., McGilligan, V., McGlynn, H., & Rowland, I. (2008). Effect of colonic bacterial metabolites on caco-2 cell paracellular permeability in vitro. Nutrition and Cancer, 60(2), 259–266. https://doi.org/10.1080/ 01635580701649644.
Illman, R. J., Topping, D. L., Mclntosh, G. H., Trimble, R. P., Storer, G. B., Taylor, M. N., & Cheng, B.-Q. (1988). Hypocholesterolaemic effects of dietary propionate: Studies in whole animals and perfused rat liver. Annals of Nutrition and Metabolism, 32(2), 97–107. https://doi.org/10.1159/000177414.
Jungersen, M., Wind, A., Johansen, E., Christensen, J., Stuer-Lauridsen, B., & Eskesen, D. (2014). The science behind the probiotic strain Bifidobacterium animalis subsp. lactis BB-12®. Microorganisms, 2(2), 92–110. https://doi.org/10.3390/ microorganisms2020092.
Kim, D.-H., Jeong, D., Kang, I.-B., Kim, H., Song, K.-Y., & Seo, K.-H. (2017). Dual function of Lactobacillus kefiri DH5 in preventing high-fat-diet-induced obesity: Direct reduction of cholesterol and upregulation of PPAR-α in adipose tissue. Molecular Nutrition & Food Research, 61(11), 1700252. https://doi.org/10.1002/ mnfr.201700252.
Kim, M., Qie, Y., Park, J., & Kim, C. H. (2016). Gut microbial metabolites fuel host antibody responses. Cell Host & Microbe, 20(2), 202–214. https://doi.org/10.1016/j. chom.2016.07.001.
Klerks, M., Bernal, M. J., Roman, S., Bodenstab, S., Gil, A., & Sanchez-Siles, L. M. (2019). Infant cereals: Current status, challenges, and future opportunities for whole grains. Nutrients, 11(2), 1–25. https://doi.org/10.3390/nu11020473.
Koskenniemi, K., Laakso, K., Koponen, J., Kankainen, M., Greco, D., Auvinen, P., … Varmanen, P. (2011). Proteomics and transcriptomics characterization of bile stress response in probiotic Lactobacillus rhamnosus GG. Molecular and Cellular Proteomics, 10(2). https://doi.org/10.1074/mcp.M110.002741.
Kratsman, N., Getselter, D., & Elliott, E. (2016). Sodium butyrate attenuates social behavior deficits and modifies the transcription of inhibitory/excitatory genes in the frontal cortex of an autism model. Neuropharmacology, 102, 136–145. https://doi. org/10.1016/j.neuropharm.2015.11.003.
Lamsal, B. P., & Faubion, J. M. (2009). The beneficial use of cereal and cereal components in probiotic foods. Food Reviews International, 25(2), 103–114. https:// doi.org/10.1080/87559120802682573.
Laparra, J. M., & Sanz, Y. (2009). Comparison of in vitro models to study bacterial adhesion to the intestinal epithelium. Letters in Applied Microbiology, 49(6), 695–701. https://doi.org/10.1111/j.1472-765X.2009.02729.x.
Liu, L. S., Firrman, J., Tanes, C., Bittinger, K., Thomas-Gahring, A., Wu, G. D., … Tomasula, P. M. (2018). Establishing a mucosal gut microbial community in vitro using an artificial simulator. PLoS ONE, 13(7), 1–20. https://doi.org/10.1371/ journal.pone.0197692.
Lopez, P., Gueimonde, M., Margolles, A., & Suarez, A. (2010). Distinct Bifidobacterium strains drive different immune responses in vitro. International Journal of Food Microbiology, 138(1–2), 157–165. https://doi.org/10.1016/j. ijfoodmicro.2009.12.023.
Macia, L., Tan, J., Vieira, A. T., Leach, K., Stanley, D., Luong, S., … Mackay, C. R. (2015). Metabolite-sensing receptors GPR43 and GPR109A facilitate dietary fibre-induced gut homeostasis through regulation of the inflammasome. Nature Communications, 6. https://doi.org/10.1038/ncomms7734.
Martins, F. S., Silva, A. A., Vieira, A. T., Barbosa, F. H. F., Arantes, R. M. E., Teixeira, M. M., & Nicoli, J. R. (2009). Comparative study of Bifidobacterium animalis, Escherichia coli, Lactobacillus casei and Saccharomyces boulardii probiotic properties. Archives of Microbiology, 191(8), 623–630. https://doi.org/10.1007/ s00203-009-0491-x.
Matsumoto, M., Ohishi, H., & Benno, Y. (2004). H+-ATPase activity in Bifidobacterium with special reference to acid tolerance. International Journal of Food Microbiology, 93 (1), 109–113. https://doi.org/10.1016/j.ijfoodmicro.2003.10.009.
Meira, Q. G. S., Magnani, M., de Medeiros Júnior, F. C., Queiroga, R. de C. R. do E., Madruga, M. S., Gullon, B., … de Souza, E. L. (2015). Effects of added Lactobacillus acidophilus and Bifidobacterium lactis probiotics on the quality characteristics of goat ricotta and their survival under simulated gastrointestinal conditions. Food Research International, 76, 828–838. https://doi.org/10.1016/j.foodres.2015.08.00 2.
Molly, K., Woestyne, M. Vande, Smet, I. De, & Verstraete, W. (1994). Validation of the simulator of the human intestinal microbial ecosystem (SHIME) reactor using microorganism-associated activities. Microbial Ecology in Health and Disease, 7(4), 191–200. https://doi.org/10.3109/08910609409141354.
Monti, F., Agostini, F., Gobbi, F., Neri, E., Schianchi, S., & Arcangeli, F. (2011). Quality of life measures in Italian children with atopic dermatitis and their families. Italian Journal of Pediatrics, 37(1), 59. https://doi.org/10.1186/1824-7288-37-59.
Mohiuddin, S. S., Khattar, D. (2020) Biochemistry, ammonia. In: StatPearls. Treasure Island (FL): StatPearls Publishing, Biochemistry. PMID: 31082083.
Niccoli, A. A. N., Artesi, A. L., Candio, F., Ceccarelli, S., Cozzali, R., Ferraro, L., … Drago, L. (2014). Preliminary results on clinical effects of probiotic Lactobacillus salivarius LS01 in children affected by atopic dermatitis. Journal of Clinical Gastroenterology, 48, S34–S36. https://doi.org/10.1097/mcg.0000000000000233.
Payne, A. N., Zihler, A., Chassard, C., & Lacroix, C. (2012). Advances and perspectives in in vitro human gut fermentation modeling. Trends in Biotechnology, 30(1), 17–25. https://doi.org/10.1016/j.tibtech.2011.06.011.
Pereira, J. O., Soares, J., Monteiro, M. J. P., Amaro, A., Gomes, A., & Pintado, M. (2019). Cereal bars functionalized through: Bifidobacterium animalis subsp. Lactis BB-12 and inulin incorporated in edible coatings of whey protein isolate or alginate. Food and Function, 10(10), 6892–6902. https://doi.org/10.1039/c9fo00370c.
Pham, V. T., & Mohajeri, M. H. (2018). The application of in vitro human intestinal models on the screening and development of pre-And probiotics. Beneficial Microbes, 9(5), 725–742. https://doi.org/10.3920/BM2017.0164.
Plaza-Díaz, J., Fontana, L., & Gil, A. (2018). Human milk oligosaccharides and immune system development. Nutrients, 10(8). https://doi.org/10.3390/nu10081038.
Plaza-Diaz, J., Ruiz-Ojeda, F. J., Gil-Campos, M., & Gil, A. (2019). Mechanisms of action of probiotics. Advances in Nutrition, 10, S49–S66. https://doi.org/10.1093/ advances/nmy063.
Possemiers, S., Marzorati, M., Verstraete, W., & Van de Wiele, T. (2010). Bacteria and chocolate: A successful combination for probiotic delivery. International Journal of Food Microbiology, 141(1–2), 97–103. https://doi.org/10.1016/j. ijfoodmicro.2010.03.008.
Possemiers, Sam, Verthe, K., Uyttendaele, S., & Verstraete, W. (2004). PCR-DGGE-based quantification of stability of the microbial community in a simulator of the human intestinal microbial ecosystem. FEMS Microbiology Ecology, 49(3), 495–507. https:// doi.org/10.1016/j.femsec.2004.05.002.
Rather, I. A., Bajpai, V. K., Kumar, S., Lim, J., Paek, W. K., & Park, Y.-H. (2016). Probiotics and atopic dermatitis: An overview. Frontiers in Microbiology, 7. https:// doi.org/10.3389/fmicb.2016.00507.
Rautava, S., Luoto, R., Salminen, S., & Isolauri, E. (2012). Microbial contact during pregnancy, intestinal colonization and human disease. Nature Reviews Gastroenterology and Hepatology, 9(10), 565–576. https://doi.org/10.1038/ nrgastro.2012.144.
Rizzardini, G., Eskesen, D., Calder, P. C., Capetti, A., Jespersen, L., & Clerici, M. (2012). Evaluation of the immune benefits of two probiotic strains Bifidobacterium animalis ssp. lactis, BB-12® and Lactobacillus paracasei ssp. paracasei, L. casei 431® in an influenza vaccination model: A randomised, double-blind, placebo-controlled study. British Journal of Nutrition, 107(6), 876–884. https://doi.org/10.1017/ S000711451100420X.
Rodrigues, V. C. D. C., Rocha Faria Duque, A. L., De Carvalho Fino, L., Simabuco, F. M., Sartoratto, A., Cabral, L. L., … Antunes, A. E. C. (2020). Modulation of the intestinal microbiota and the metabolites produced by the administration of ice cream and a dietary supplement containing the same probiotics. British Journal of Nutrition, 124 (1), 57–68. https://doi.org/10.1017/S0007114520000896.
Roduit, C., Frei, R., Ferstl, R., Loeliger, S., Westermann, P., Rhyner, C., … Doekes, G. (2019). High levels of butyrate and propionate in early life are associated with protection against atopy. Allergy: European Journal of Allergy and Clinical Immunology, 74(4), 799–809. https://doi.org/10.1111/all.13660.
Roess, A. A., Jacquier, E. F., Catellier, D. J., Carvalho, R., Lutes, A. C., Anater, A. S., & Dietz, W. H. (2018). Food consumption patterns of infants and toddlers: Findings from the feeding infants and toddlers study (FITS) 2016. Journal of Nutrition, 148(9), 1525S–1535S. https://doi.org/10.1093/jn/nxy171.
Round, J. L., & Mazmanian, S. K. (2009). The gut microbiota shapes intestinal immune responses during health and disease. Nature Reviews Immunology, 9(5), 313–323. https://doi.org/10.1038/nri2515.
Ruijschop, R. M. A. J., Boelrijk, A. E. M., & te Giffel, M. C. (2008). Satiety effects of a dairy beverage fermented with propionic acid bacteria. International Dairy Journal, 18(9), 945–950. https://doi.org/10.1016/j.idairyj.2008.01.004.
Russell, W. R., Gratz, S. W., Duncan, S. H., Holtrop, G., Ince, J., Scobbie, L., … Flint, H. J. (2011). High-protein, reduced-carbohydrate weight-loss diets promote metabolite profiles likely to be detrimental to colonic health. American Journal of Clinical
M.K. Salgaço et al.

Nutrition, 93(5), 1062–1072. https://doi.org/10.3945/ajcn.110.002188. INTRODUCTION.
Russo, R., De Caro, C., Avagliano, C., Cristiano, C., La Rana, G., Mattace Raso, G., … Calignano, A. (2016). Sodium butyrate and its synthetic amide derivative modulate nociceptive behaviors in mice. Pharmacological Research, 103, 279–291. https://doi. org/10.1016/j.phrs.2015.11.026.
Savy, N., Brossier, D., Brunel-Guitton, C., Ducharme-Crevier, L., Du Pont-Thibodeau, G., & Jouvet, P. (2018). Acute pediatric hyperammonemia: Current diagnosis and management strategies. Hepatic Medicine: Evidence and Research, 10, 105–115. https://doi.org/10.2147/hmer.s140711.
Scott, K. P., Gratz, S. W., Sheridan, P. O., Flint, H. J., & Duncan, S. H. (2013). The influence of diet on the gut microbiota. Pharmacological Research, 69(1), 52–60. https://doi.org/10.1016/j.phrs.2012.10.020.
Sekirov, I., Russell, S. L., Caetano M Antunes, L., & Finlay, B. B. (2010). Gut microbiota in health and disease. Physiological Reviews, 90(3), 859–904. https://doi.org/10.1152/ physrev.00045.2009.
Sendra, E., Sayas-Barbera, M. E., Fernandez-Lopez, J., & Perez-Alvarez, J. A. (2016). Effect of food composition on probiotic bacteria viability. Probiotics, prebiotics, and synbiotics: Bioactive foods in health promotion. Elsevier Inc.. https://doi.org/10.1016/ B978-0-12-802189-7.00017-4.
Sivieri, K., Morales, M. L. V., Adorno, M. A. T., Sakamoto, I. K., Saad, S. M. I., & Rossi, E. A. (2013). Lactobacillus acidophilus CRL 1014 improved “gut health” in the SHIME® reactor. BMC Gastroenterology, 13(1). https://doi.org/10.1186/1471-230X- 13-100.
Sivieri, K., Morales, M. L. V., Saad, S. M. I., Adorno, M. A. T., Sakamoto, I. K., & Rossi, E. A. (2014). Prebiotic effect of fructooligosaccharide in the Simulator of the Human Intestinal Microbial Ecosystem (SHIME® Model). Journal of Medicinal Food, 17(8), 894–901. https://doi.org/10.1089/jmf.2013.0092.
Solano-Aguilar, G., Dawson, H., Restrepo, M., Andrews, K., Vinyard, B., & Urban, J. F. (2008). Detection of Bifidobacterium animalis subsp. lactis (Bb12) in the intestine after feeding of sows and their piglets. Applied and Environmental Microbiology, 74 (20), 6338–6347. https://doi.org/10.1128/AEM.00309-08.
Stilling, R. M., van de Wouw, M., Clarke, G., Stanton, C., Dinan, T. G., & Cryan, J. F. (2016). The neuropharmacology of butyrate: The bread and butter of the microbiota- gut-brain axis? Neurochemistry International, 99, 110–132. https://doi.org/10.1016/ j.neuint.2016.06.011.
Takuma, K., Hara, Y., Kataoka, S., Kawanai, T., Maeda, Y., Watanabe, R., … Matsuda, T. (2014). Chronic treatment with valproic acid or sodium butyrate attenuates novel object recognition deficits and hippocampal dendritic spine loss in a mouse model of autism. Pharmacology Biochemistry and Behavior, 126, 43–49. https://doi.org/ 10.1016/j.pbb.2014.08.013.
Taverniti, V., Scabiosi, C., Arioli, S., Mora, D., & Guglielmetti, S. (2014). Short-term daily intake of 6 billion live probiotic cells can be insufficient in healthy adults to modulate the intestinal bifidobacteria and lactobacilli. Journal of Functional Foods, 6 (1), 482–491. https://doi.org/10.1016/j.jff.2013.11.014.
Thursby, E., & Juge, N. (2017). Introduction to the human gut microbiota. Biochemical Journal, 474(11), 1823–1836. https://doi.org/10.1042/BCJ20160510.
Van den Abbeele, P., Duysburgh, C., Vazquez, E., Chow, J., Buck, R., & Marzorati, M. (2019). 2′-Fucosyllactose alters the composition and activity of gut microbiota from formula-fed infants receiving complementary feeding in a validated intestinal model. Journal of Functional Foods, 61(July). https://doi.org/10.1016/j.jff.2019.103484.
Vandenplas, Y., Huys, G., & Daube, G. (2015). Probiotics: An update. Jornal de Pediatria, 91(1), 6–21. https://doi.org/10.1016/j.jped.2014.08.005.
Vernazza, C. L., Gibson, G. R., & Rastall, R. A. (2006). Carbohydrate preference, acid tolerance and bile tolerance in five strains of Bifidobacterium. Journal of Applied Microbiology, 100(4), 846–853. https://doi.org/10.1111/j.1365-2672.2006.02832.x.
Verruck, S., Prudencio, E. S., Vieira, C. R. W., Amante, E. R., & de Mello Castanho Amboni, R. D. (2015). The buffalo Minas Frescal cheese as a protective matrix of Bifidobacterium BB-12 under invitro simulated gastrointestinal conditions. LWT – Food Science and Technology, 63(2), 1179–1183. https://doi.org/10.1016/j. lwt.2015.04.014.
Vinderola, C. G., & Reinheimer, J. A. (1999). Culture media for the enumeration of Bifidobacterium bifidum and Lactobacillus acidophilus in the presence of yoghurt bacteria. International Dairy Journal, 9(8), 497–505. https://doi.org/10.1016/S0958- 6946(99)00120-X.
Vinderola, C. G., & Reinheimer, J. A. (2003). Lactic acid starter and probiotic bacteria: A comparative “in vitro” study of probiotic characteristics and biological barrier resistance. Food Research International, 36(9–10), 895–904. https://doi.org/ 10.1016/S0963-9969(03)00098-X.
Walsh, C. J., Guinane, C. M., O’Toole, P. W., & Cotter, P. D. (2014). Beneficial modulation of the gut microbiota. FEBS Letters, 588(22), 4120–4130. https://doi. org/10.1016/j.febslet.2014.03.035.
Wang, Y., & Qian, P. Y. (2009). Conservative fragments in bacterial 16S rRNA genes and primer design for 16S ribosomal DNA amplicons in metagenomic studies. PLoS ONE, 4(10). https://doi.org/10.1371/journal.pone.0007401.
Wu, T., Grootaert, C., Pitart, J., Vidovic, N. K., Kamiloglu, S., Possemiers, S., … Van Camp, J. (2018). Aronia (Aronia melanocarpa) polyphenols modulate microbial community in Simulator of the Human Intestinal Microbial Ecosystem (SHIME®) and decrease secretion of pro-inflammatory markers in a Caco-2/endothelial cell co- culture model. Molecular Nutrition & Food Research, 1800607. https://doi.org/ 10.1002/mnfr.201800607.
Zhou, J., Martin, R. J., Tulley, R. T., Raggio, A. M., McCutcheon, K. L., Shen, L., … Keenan, M. J. (2008). Dietary resistant starch upregulates total GLP-1 and PYY in a sustained day-long manner through fermentation in rodents. American Journal of Physiology – Endocrinology and Metabolism, 295(5), 1160–1167. https://doi.org/ 10.1152/ajpendo.90637.2008.
Zhuang, L., Chen, H., Zhang, S., Zhuang, J., Li, Q., & Feng, Z. (2019). Intestinal microbiota in early life and its implications on childhood health. Genomics, Proteomics and Bioinformatics, 17(1), 13–25. https://doi.org/10.1016/j. gpb.2018.10.002.
Zubiría, M. G., Gambaro, S. E., Rey, M. A., Carasi, P., Serradell, M. de los A., & Giovambattista, A. (2017). Deleterious metabolic effects of high fructose intake: The preventive effect of Lactobacillus kefiri administration. Nutrients, 9(5). https://doi. org/10.3390/nu9050470.
M.K. Salgaço et al.