finlay j.b. ,g.f.esteban & t. fenchel (1998) .protozoan diversity.converging estimates of the...
TRANSCRIPT

Protist, Vol. 149, 29-37, February 1998 © Gustav Fischer Verlag
ORIGINAL PAPER
Protist
Protozoan Diversity: Converging Estimatesof the Global Number of Free-Living Ciliate Species
Bland J. Finlaya,1, Genoveva F. Estebana, and Tom Fenchelb
a Institute of Freshwater Ecology, Windermere laboratory, The Ferry House, Ambleside, Cumbria LA22 alP, UKb Marine Biological laboratory (University of Copenhagen), Strandpromenaden 5, DK - 3000, Helsingm, Denmark
Submitted October 6, 1997; Accepted November 14, 1997Monitoring Editor: Michael Melkonian
Protozoa are the most abundant phagotrophs in the biosphere, but no scientific strategy hasemerged that might allow accurate definition of the dimensions of protozoan diversity on a globalscale. We have begun this task by searching for the common ground between taxonomy and ecology.We have used two methods - taxonomic analysis, and extrapolation from ecological datasets - to estimate the global species richness of free-living ciliated protozoa in the marine interstitial and freshwater benthos. The methods provide estimates that agree within a factor of two, and it is apparentthat the species-area curves for ciliates must be almost flat (the slope z takes the very low value of0.043 in the equation: [number of species] = [constant][area]Z). Insofar as independent ecologicaldatasets can be extrapolated to show similiar, flat, species-area relations, and that these convergewith an independent estimate from taxonomic analysis, we conclude that the great majority of freeliving ciliates are ubiquitous. This strengthens our recent claim that the global species richness offree-living ciliated protozoa is relatively low (-3000).
Introduction
It is rather difficult to provide accurate estimates forthe number of species in any of the larger taxonomicgroups (see May 1988, 1990). Furthermore, thescale of the problem seems to be inversely relatedto the size of the organisms concerned. Estimatingthe global species richness of birds is probably anachievable task (Zink 1996), but more difficult for theinsects (Gaston 1992) and, apparently, extraordinarily difficult for micro-organisms (see UNEP 1995),where in many cases we do not even have a clearidea of what a species is, let alone a sound strategyfor estimating global diversity.
1 Corresponding author;fax 44-15394-46914;e-mail [email protected]
Some of the methods used for estimatingspecies richness of larger organisms are inadequate when it comes to micro-organisms. For example, it may under certain circumstances be possible to extrapolate species numbers in size categories down to about 1 em, but this becomes a verydubious procedure for size classes smaller than this(Fenchel 1993). The main reason may be themarked tendency towards cosmopolitanism in thesmallest organisms. For these, barriers to migrationand dispersal appear to be ineffective: thus rates ofextinction and speciation may be low, and the samespecies will tend to be distributed worldwide. Sothe global species richness of micro-organismscould be relatively low, and the task of accuratelyestimating the number of microbial species (includ-
Protist, Vol. 149, 29-37, February 1998 © Gustav Fischer Verlag
ORIGINAL PAPER
Protist
Protozoan Diversity: Converging Estimatesof the Global Number of Free-Living Ciliate Species
Bland J. Finlaya,1, Genoveva F. Estebana, and Tom Fenchelb
a Institute of Freshwater Ecology, Windermere laboratory, The Ferry House, Ambleside, Cumbria LA22 alP, UKb Marine Biological laboratory (University of Copenhagen), Strandpromenaden 5, DK - 3000, Helsingm, Denmark
Submitted October 6, 1997; Accepted November 14, 1997Monitoring Editor: Michael Melkonian
Protozoa are the most abundant phagotrophs in the biosphere, but no scientific strategy hasemerged that might allow accurate definition of the dimensions of protozoan diversity on a globalscale. We have begun this task by searching for the common ground between taxonomy and ecology.We have used two methods - taxonomic analysis, and extrapolation from ecological datasets - to estimate the global species richness of free-living ciliated protozoa in the marine interstitial and freshwater benthos. The methods provide estimates that agree within a factor of two, and it is apparentthat the species-area curves for ciliates must be almost flat (the slope z takes the very low value of0.043 in the equation: [number of species] = [constant][area]Z). Insofar as independent ecologicaldatasets can be extrapolated to show similiar, flat, species-area relations, and that these convergewith an independent estimate from taxonomic analysis, we conclude that the great majority of freeliving ciliates are ubiquitous. This strengthens our recent claim that the global species richness offree-living ciliated protozoa is relatively low (-3000).
Introduction
It is rather difficult to provide accurate estimates forthe number of species in any of the larger taxonomicgroups (see May 1988, 1990). Furthermore, thescale of the problem seems to be inversely relatedto the size of the organisms concerned. Estimatingthe global species richness of birds is probably anachievable task (Zink 1996), but more difficult for theinsects (Gaston 1992) and, apparently, extraordinarily difficult for micro-organisms (see UNEP 1995),where in many cases we do not even have a clearidea of what a species is, let alone a sound strategyfor estimating global diversity.
1 Corresponding author;fax 44-15394-46914;e-mail [email protected]
Some of the methods used for estimatingspecies richness of larger organisms are inadequate when it comes to micro-organisms. For example, it may under certain circumstances be possible to extrapolate species numbers in size categories down to about 1 em, but this becomes a verydubious procedure for size classes smaller than this(Fenchel 1993). The main reason may be themarked tendency towards cosmopolitanism in thesmallest organisms. For these, barriers to migrationand dispersal appear to be ineffective: thus rates ofextinction and speciation may be low, and the samespecies will tend to be distributed worldwide. Sothe global species richness of micro-organismscould be relatively low, and the task of accuratelyestimating the number of microbial species (includ-

30 B. J. Finlay, G. F. Esteban, and T. Fenchel
ing protozoa) may be simpler than previouslythought.
What evidence is there that protozoan species aretypically cosmopolitan and that the global numberof these species is modest? We have begun to answer this question for one large protozoan group inparticular - the ciliates. There are four main bodiesof evidence. First, and in common with most othergroups of protists, there is no good evidence thatciliates have a biogeography (Bary 1950; Bowersand Pratt 1995; Corliss 1974; Ekebom et al. 1996;Kristiansen 1996; Lackey 1938; Larsen and Patterson 1990; Ogden and Hedley 1980; Patterson andSimpson 1996; Sandon 1927; Stout 1956; Tyler1996). On the contrary, it seems that the same ciliatespecies are found wherever their preferred habitat isfound (e.g. Smith 1978). Second, enrichment cultureexperiments indicate that local species richness ofciliates is a significant proportion of the globalspecies richness (Fenchel et al. 1997; Finlay et al.1996a). Third, the diversity of free-living ciliatespecies described in the international literature is arelatively small number (close to 3000, after revision), and unlikely to increase significantly in the future (Finlay et al. 1996b). Fourth, ciliates and otherprotists have high absolute abundance and, in manycases, effective passive dispersal (e.g. Maguire1963; Parsons et al. 1966; Maguire and Belk 1967;Schlichting and Sides 1969; Kristiansen 1996; Finlay1997). Much evidence indicates that they are continually being distributed everywhere', and newlyformed habitats such as freshwater ponds and volcanic islands are rapidly colonised (e.g. Holmbergand Pejler 1972; Scourfield 1944). Free-living ciliatesmay, in fact, be ubiquitous.
If we take this assumption of ubiquity and use it toextrapolate from ecological datasets for relatively
small areas to produce global estimates of the number of ciliate species, will we obtain estimates thatare similar to the global number of nominal ciliatespecies? We have tested this proposition, usinglarge datasets for free-living ciliates in the marine interstitial and in the freshwater benthos. The category 'free-living ciliates' is here defined as all thoseextant ciliates that do not live exclusively as gutsymbionts (e.g. rumen ciliates), or as parasites orsymphorionts of specific metazoans. A detailed listof taxa considered not to be free-living is given inFinlay et al. (1996b).
Results
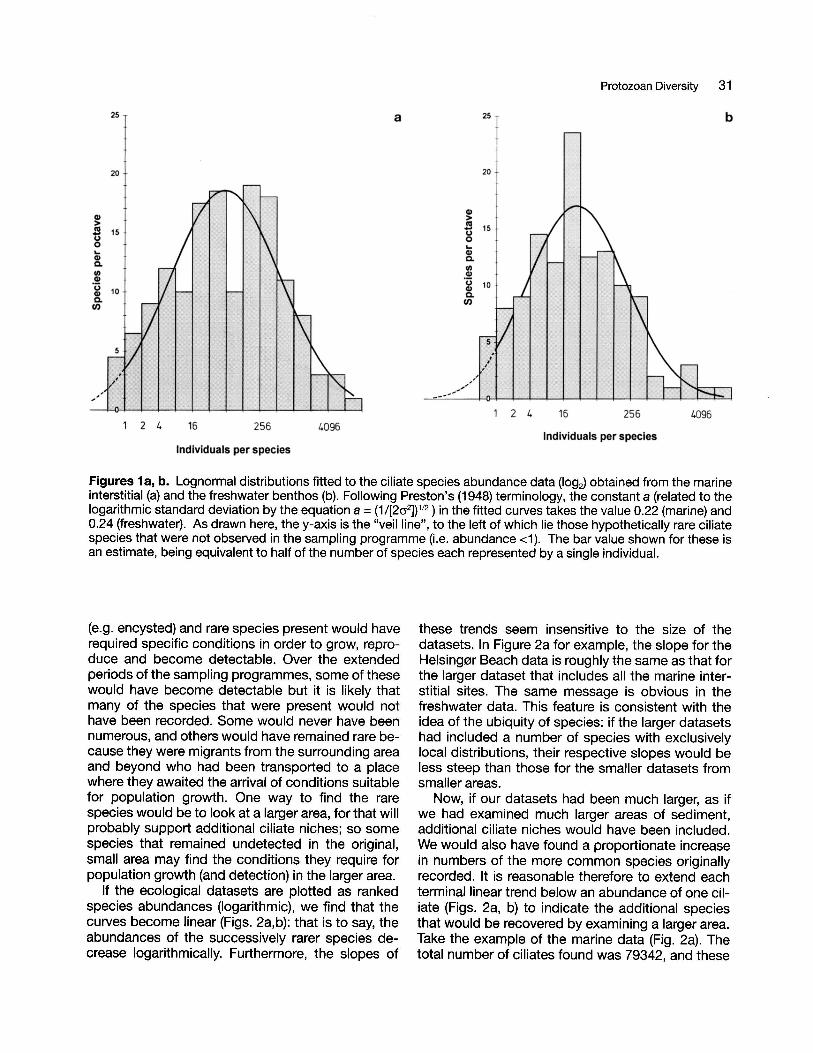
The ecological data for numbers and abundance ofciliate species in marine and freshwater sites aresummarised in Table 1. As we also have data for thenumbers of individuals of each species, the speciesabundance data can be illustrated as frequencycurves. Using the original method of Preston (1948),with log2 octaves on the abscissa (Le. each successive octave represents a doubling in abundance), wecreate species curves indicating that most speciesare of intermediate abundance, a few species arevery abundant, and some are very rare. A normalcurve superimposed upon the logarithmic abundance data (a 'lognormal' distribution) provides anadequate fit (Figs. 1a, b). In this case, as is often sowith ecological data, the distribution is truncated, although nearly 'unveiled' (Le. indicating that mostspecies have been recorded). But this message ismisleading, for it fails to take account of the potential ciliate diversity - the true number of ciliatespecies present, but not detected during the timespan of the sampling programme. Many of the cryptic
Table 1. Summary information from ecological datasets with independent estimates for the global number ofciliate species in the marine interstitial and in the freshwater benthos
Ecological Datasets Global Estimate Based on:
Number of Number of Extrapolation Taxonomicciliates ciliate species from ecological analysis*recorded recorded datasets
Marine Helsing0r Beach 48186 85 597 793Interstitial All marine sites 79342 151
Freshwater Esthwaite Water 20486 104 732 1370Benthos All freshwater sites 35837 125
*Current estimated numbers for all nominal marine and non-marine free-living ciliate species are 1592 and 2152 respectively. These numbers include, for example, planktonic species, many suctorians, and those ciliates found sofar only in soil.
30 B. J. Finlay, G. F. Esteban, and T. Fenchel
ing protozoa) may be simpler than previouslythought.
What evidence is there that protozoan species aretypically cosmopolitan and that the global numberof these species is modest? We have begun to answer this question for one large protozoan group inparticular - the ciliates. There are four main bodiesof evidence. First, and in common with most othergroups of protists, there is no good evidence thatciliates have a biogeography (Bary 1950; Bowersand Pratt 1995; Corliss 1974; Ekebom et al. 1996;Kristiansen 1996; Lackey 1938; Larsen and Patterson 1990; Ogden and Hedley 1980; Patterson andSimpson 1996; Sandon 1927; Stout 1956; Tyler1996). On the contrary, it seems that the same ciliatespecies are found wherever their preferred habitat isfound (e.g. Smith 1978). Second, enrichment cultureexperiments indicate that local species richness ofciliates is a significant proportion of the globalspecies richness (Fenchel et al. 1997; Finlay et al.1996a). Third, the diversity of free-living ciliatespecies described in the international literature is arelatively small number (close to 3000, after revision), and unlikely to increase significantly in the future (Finlay et al. 1996b). Fourth, ciliates and otherprotists have high absolute abundance and, in manycases, effective passive dispersal (e.g. Maguire1963; Parsons et al. 1966; Maguire and Belk 1967;Schlichting and Sides 1969; Kristiansen 1996; Finlay1997). Much evidence indicates that they are continually being distributed everywhere', and newlyformed habitats such as freshwater ponds and volcanic islands are rapidly colonised (e.g. Holmbergand Pejler 1972; Scourfield 1944). Free-living ciliatesmay, in fact, be ubiquitous.
If we take this assumption of ubiquity and use it toextrapolate from ecological datasets for relatively
small areas to produce global estimates of the number of ciliate species, will we obtain estimates thatare similar to the global number of nominal ciliatespecies? We have tested this proposition, usinglarge datasets for free-living ciliates in the marine interstitial and in the freshwater benthos. The category 'free-living ciliates' is here defined as all thoseextant ciliates that do not live exclusively as gutsymbionts (e.g. rumen ciliates), or as parasites orsymphorionts of specific metazoans. A detailed listof taxa considered not to be free-living is given inFinlay et al. (1996b).
Results
The ecological data for numbers and abundance ofciliate species in marine and freshwater sites aresummarised in Table 1. As we also have data for thenumbers of individuals of each species, the speciesabundance data can be illustrated as frequencycurves. Using the original method of Preston (1948),with log2 octaves on the abscissa (Le. each successive octave represents a doubling in abundance), wecreate species curves indicating that most speciesare of intermediate abundance, a few species arevery abundant, and some are very rare. A normalcurve superimposed upon the logarithmic abundance data (a 'lognormal' distribution) provides anadequate fit (Figs. 1a, b). In this case, as is often sowith ecological data, the distribution is truncated, although nearly 'unveiled' (Le. indicating that mostspecies have been recorded). But this message ismisleading, for it fails to take account of the potential ciliate diversity - the true number of ciliatespecies present, but not detected during the timespan of the sampling programme. Many of the cryptic
Table 1. Summary information from ecological datasets with independent estimates for the global number ofciliate species in the marine interstitial and in the freshwater benthos
Ecological Datasets Global Estimate Based on:
Number of Number of Extrapolation Taxonomicciliates ciliate species from ecological analysis*recorded recorded datasets
Marine Helsing0r Beach 48186 85 597 793Interstitial All marine sites 79342 151
Freshwater Esthwaite Water 20486 104 732 1370Benthos All freshwater sites 35837 125
*Current estimated numbers for all nominal marine and non-marine free-living ciliate species are 1592 and 2152 respectively. These numbers include, for example, planktonic species, many suctorians, and those ciliates found sofar only in soil.
25
20
a 25
20
Protozoan Diversity 31
b
~:l 15Uo~
~CoIII~
'2 10CoV)
~>~ 15o~
~CoIII~
'2 10CoV)
5
1 2 I. 16 256 40961 2 I. 16 256 4096
Individuals per speciesIndividuals per species
Figures 1a, b. Lognormal distributions fitted to the ciliate species abundance data (log2) obtained from the marineinterstitial (a) and the freshwater benthos (b). Following Preston's (1948) terminology, the constant a (related to thelogarithmic standard deviation by the equation a =(1/[202])1/2) in the fitted curves takes the value 0.22 (marine) and0.24 (freshwater). As drawn here, the y-axis is the "veil line", to the left of which lie those hypothetically rare ciliatespecies that were not observed in the sampling programme (Le. abundance <1). The bar value shown for these isan estimate, being equivalent to half of the number of species each represented by a single individual.
(e.g. encysted) and rare species present would haverequired specific conditions in order to grow, reproduce and become detectable. Over the extendedperiods of the sampling programmes, some of thesewould have become detectable but it is likely thatmany of the species that were present would nothave been recorded. Some would never have beennumerous, and others would have remained rare because they were migrants from the surrounding areaand beyond who had been transported to a placewhere they awaited the arrival of conditions suitablefor population growth. One way to find the rarespecies would be to look at a larger area, for that willprobably support additional ciliate niches; so somespecies that remained undetected in the original,small area may find the conditions they require forpopulation growth (and detection) in the larger area.
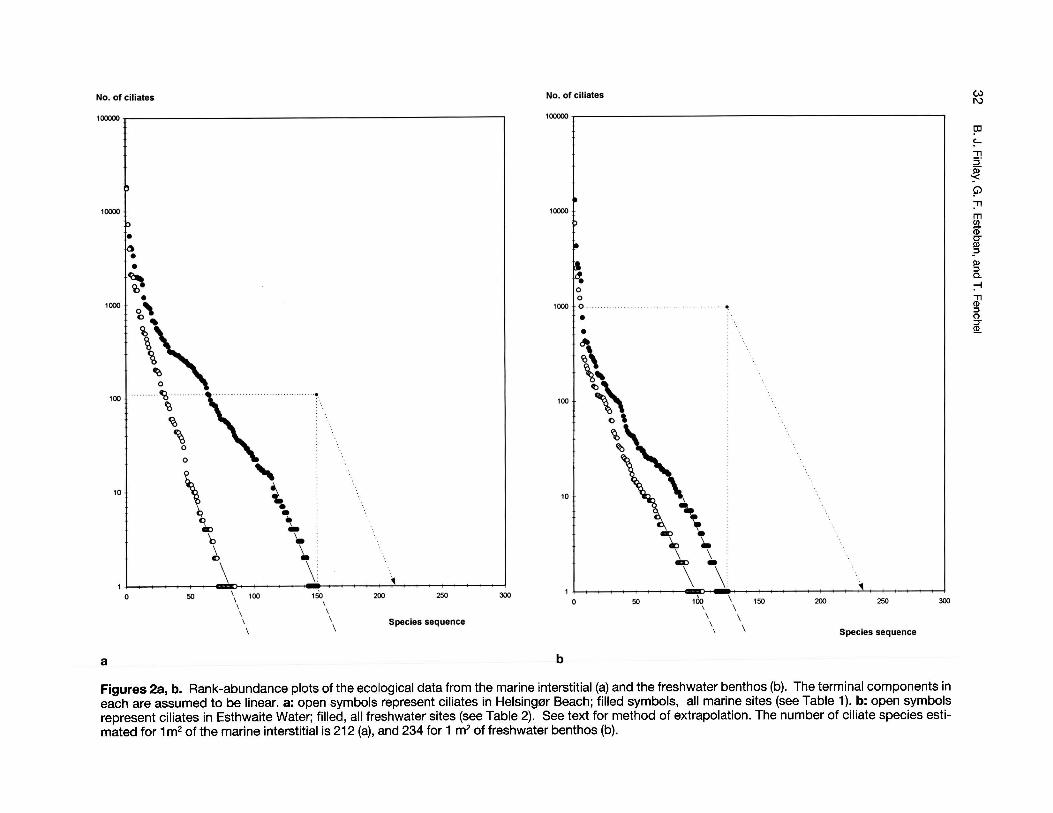
If the ecological datasets are plotted as rankedspecies abundances (logarithmic), we find that thecurves become linear (Figs. 2a,b): that is to say, theabundances of the successively rarer species decrease logarithmically. Furthermore, the slopes of
these trends seem insensitive to the size of thedatasets. In Figure 2a for example, the slope for theHelsing0r Beach data is roughly the same as that forthe larger dataset that includes all the marine interstitial sites. The same message is obvious in thefreshwater data. This feature is consistent with theidea of the ubiquity of species: if the larger datasetshad included a number of species with exclusivelylocal distributions, their respective slopes would beless steep than those for the smaller datasets fromsmaller areas.
Now, if our datasets had been much larger, as ifwe had examined much larger areas of sediment,additional ciliate niches would have been included.We would also have found a proportionate increasein numbers of the more common species originallyrecorded. It is reasonable therefore to extend eachterminal linear trend below an abundance of one ciliate (Figs. 2a, b) to indicate the additional speciesthat would be recovered by examining a larger area.Take the example of the marine data (Fig. 2a). Thetotal number of ciliates found was 79342, and these
25
20
a 25
20
Protozoan Diversity 31
b
~:l 15Uo~
~CoIII~
'2 10CoV)
~>~ 15o~
~CoIII~
'2 10CoV)
5
1 2 I. 16 256 40961 2 I. 16 256 4096
Individuals per speciesIndividuals per species
Figures 1a, b. Lognormal distributions fitted to the ciliate species abundance data (log2) obtained from the marineinterstitial (a) and the freshwater benthos (b). Following Preston's (1948) terminology, the constant a (related to thelogarithmic standard deviation by the equation a =(1/[202])1/2) in the fitted curves takes the value 0.22 (marine) and0.24 (freshwater). As drawn here, the y-axis is the "veil line", to the left of which lie those hypothetically rare ciliatespecies that were not observed in the sampling programme (Le. abundance <1). The bar value shown for these isan estimate, being equivalent to half of the number of species each represented by a single individual.
(e.g. encysted) and rare species present would haverequired specific conditions in order to grow, reproduce and become detectable. Over the extendedperiods of the sampling programmes, some of thesewould have become detectable but it is likely thatmany of the species that were present would nothave been recorded. Some would never have beennumerous, and others would have remained rare because they were migrants from the surrounding areaand beyond who had been transported to a placewhere they awaited the arrival of conditions suitablefor population growth. One way to find the rarespecies would be to look at a larger area, for that willprobably support additional ciliate niches; so somespecies that remained undetected in the original,small area may find the conditions they require forpopulation growth (and detection) in the larger area.
If the ecological datasets are plotted as rankedspecies abundances (logarithmic), we find that thecurves become linear (Figs. 2a,b): that is to say, theabundances of the successively rarer species decrease logarithmically. Furthermore, the slopes of
these trends seem insensitive to the size of thedatasets. In Figure 2a for example, the slope for theHelsing0r Beach data is roughly the same as that forthe larger dataset that includes all the marine interstitial sites. The same message is obvious in thefreshwater data. This feature is consistent with theidea of the ubiquity of species: if the larger datasetshad included a number of species with exclusivelylocal distributions, their respective slopes would beless steep than those for the smaller datasets fromsmaller areas.
Now, if our datasets had been much larger, as ifwe had examined much larger areas of sediment,additional ciliate niches would have been included.We would also have found a proportionate increasein numbers of the more common species originallyrecorded. It is reasonable therefore to extend eachterminal linear trend below an abundance of one ciliate (Figs. 2a, b) to indicate the additional speciesthat would be recovered by examining a larger area.Take the example of the marine data (Fig. 2a). Thetotal number of ciliates found was 79342, and these
No. of ciliates No. of ciliates c.vN
10000
100000 I I
~
~•
~c....'TIS'
~P:nm!eoCD0$I)
.~
$I)~
Co
:-f'TICD~
o:::r~•
~\~,
100+ ~() ,~
10
\C>
~~\ \~ -
1 I \ \ ~~ -0 50 100 \ 150 200 250 300
\ \\ \
\ \\ Species sequence
~•oo
1000 to. .•
10000 t
100000 I I
300
Species sequence
200 250150\\\\
\ 100\\\\
\
50
10
•~
•~,,
\~.... . .100 V······· ~
%'%
o
\ '\t.J '-
~~. I
~\ \ ~~~~;:-------;-1 Io
1000
a b
Figures 2a, b. Rank-abundance plots of the ecological data from the marine interstitial (a) and the freshwater benthos (b). The terminal components ineach are assumed to be linear. a: open symbols represent ciliates in Helsing0r Beach; filled symbols, all marine sites (see Table 1). b: open symbolsrepresent ciliates in Esthwaite Water; filled, all freshwater sites (see Table 2). See text for method of extrapolation. The number of ciliate species estimated for 1m2 of the marine interstitial is 212 (a), and 234 for 1 m2 of freshwater benthos (b).
No. of ciliates No. of ciliates
100000 .,-----------------------------, 100000.,----------------------------,
10000
p•~
300250
Species sequence
200150\\
\\
~\\;,~~
"~ ~
\ \
\ -\.1 +---+-~...--.___+_....-.-+-__~__-+-..........__..........._~,.>..1· ..........._~---+---I
o 50 100\
\\
\
10
100
I-
~•oo
1000 0 .
••
10000
300
Species sequence
•~
•0'
~\"\ '\
&l .-b '-~ ~
\ \-1 -I-_--+-~.......... .,......... ........___.........._..,,1 +_ ....j
o 50 \ 100 150 200 250\ \\ \\ \\ \
10
100
1000
a b
Figures 2a, b. Rank-abundance plots of the ecological data from the marine interstitial (a) and the freshwater benthos (b). The terminal components ineach are assumed to be linear. a: open symbols represent ciliates in HelsingeJr Beach; filled symbols, all marine sites (see Table 1). b: open symbolsrepresent ciliates in Esthwaite Water; filled, all freshwater sites (see Table 2). See text for method of extrapolation. The number of ciliate species estimated for 1m2 of the marine interstitial is 212 (a), and 234 for 1 m2 of freshwater benthos (b).
Protozoan Diversity 33
6
Insects
10
Ciliates
5o
Log Area (km2)
-5
2
5
4
O+------f-------+------+---------l-10
3
IIIIIIQ)c:.c:(.)
~InQ)
'0Q)a.
(J)
Clo...J
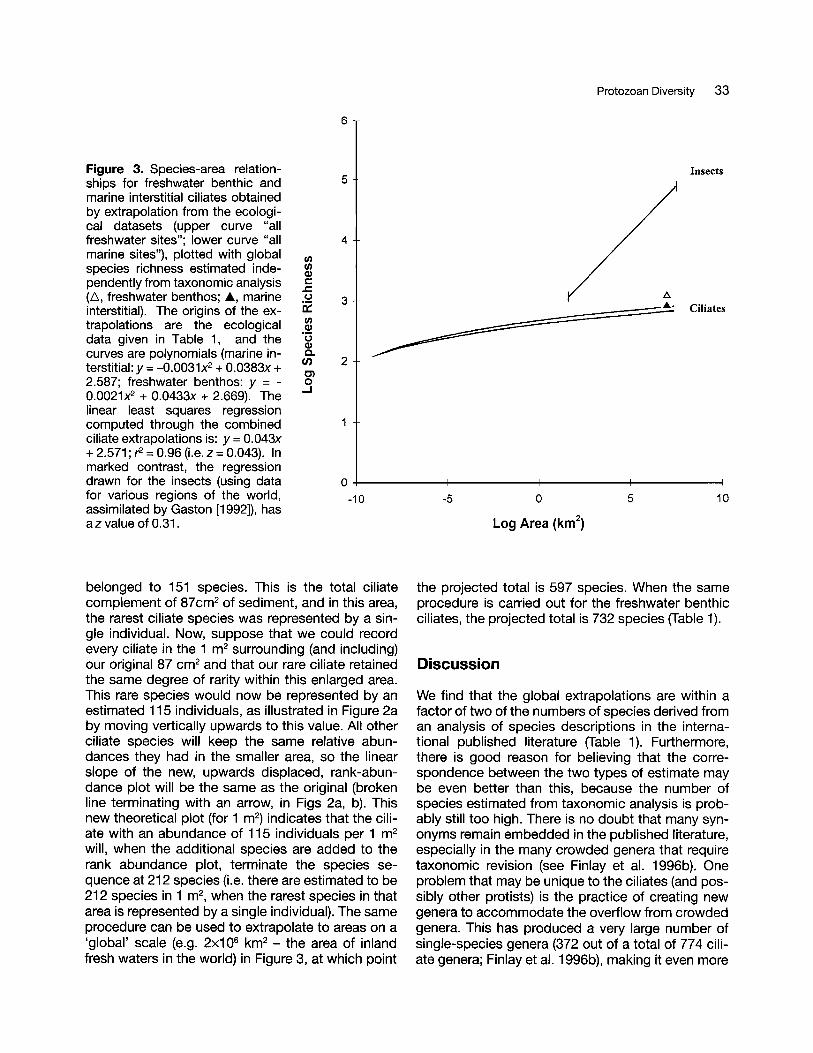
Figure 3. Species-area relationships for freshwater benthic andmarine interstitial ciliates obtainedby extrapolation from the ecological datasets (upper curve "allfreshwater sites"; lower curve "allmarine sites"), plotted with globalspecies richness estimated independently from taxonomic analysis(1'" freshwater benthos; A, marineinterstitial). The origins of the extrapolations are the ecologicaldata given in Table 1, and thecurves are polynomials (marine interstitial: y =-0.0031x2 + 0.0383x +2.587; freshwater benthos: y = 0.0021x2 + 0.0433x + 2.669). Thelinear least squares regressioncomputed through the combinedciliate extrapolations is: y =0.043x+ 2.571 ; r2 =0.96 (Le. z =0.043). Inmarked contrast, the regressiondrawn for the insects (using datafor various regions of the world,assimilated by Gaston [1992]), hasaz value of 0.31.
belonged to 151 species. This is the total ciliatecomplement of 87cm2 of sediment, and in this area,the rarest ciliate species was represented by a single individual. Now, suppose that we could recordevery ciliate in the 1 m2 surrounding (and including)our original 87 cm2 and that our rare ciliate retainedthe same degree of rarity within this enlarged area.This rare species would now be represented by anestimated 115 individuals, as illustrated in Figure 2aby moving vertically upwards to this value. All otherciliate species will keep the same relative abundances they had in the smaller area, so the linearslope of the new, upwards displaced, rank-abundance plot will be the same as the original (brokenline terminating with an arrow, in Figs 2a, b). Thisnew theoretical plot (for 1 m2) indicates that the ciliate with an abundance of 115 individuals per 1 m2
will, when the additional species are added to therank abundance plot, terminate the species sequence at 212 species (Le. there are estimated to be212 species in 1 m2
, when the rarest species in thatarea is represented by a single individual). The sameprocedure can be used to extrapolate to areas on a'global' scale (e.g. 2x106 km2 - the area of inlandfresh waters in the world) in Figure 3, at which point
the projected total is 597 species. When the sameprocedure is carried out for the freshwater benthicciliates, the projected total is 732 species (Table 1).
Discussion
We find that the global extrapolations are within afactor of two of the numbers of species derived froman analysis of species descriptions in the international published literature (Table 1). Furthermore,there is good reason for believing that the correspondence between the two types of estimate maybe even better than this, because the number ofspecies estimated from taxonomic analysis is probably still too high. There is no doubt that many synonyms remain embedded in the published literature,especially in the many crowded genera that requiretaxonomic revision (see Finlay et al. 1996b). Oneproblem that may be unique to the ciliates (and possibly other protists) is the practice of creating newgenera to accommodate the overflow from crowdedgenera. This has produced a very large number ofsingle-species genera (372 out of a total of 774 ciliate genera; Finlay et al. 1996b), making it even more
Protozoan Diversity 33
6
Insects
10
Ciliates
5o
Log Area (km2)
-5
2
5
4
O+------f-------+------+---------l-10
3
IIIIIIQ)c:.c:(.)
~InQ)
'0Q)a.
(J)
Clo...J
Figure 3. Species-area relationships for freshwater benthic andmarine interstitial ciliates obtainedby extrapolation from the ecological datasets (upper curve "allfreshwater sites"; lower curve "allmarine sites"), plotted with globalspecies richness estimated independently from taxonomic analysis(1'" freshwater benthos; A, marineinterstitial). The origins of the extrapolations are the ecologicaldata given in Table 1, and thecurves are polynomials (marine interstitial: y =-0.0031x2 + 0.0383x +2.587; freshwater benthos: y = 0.0021x2 + 0.0433x + 2.669). Thelinear least squares regressioncomputed through the combinedciliate extrapolations is: y =0.043x+ 2.571 ; r2 =0.96 (Le. z =0.043). Inmarked contrast, the regressiondrawn for the insects (using datafor various regions of the world,assimilated by Gaston [1992]), hasaz value of 0.31.
belonged to 151 species. This is the total ciliatecomplement of 87cm2 of sediment, and in this area,the rarest ciliate species was represented by a single individual. Now, suppose that we could recordevery ciliate in the 1 m2 surrounding (and including)our original 87 cm2 and that our rare ciliate retainedthe same degree of rarity within this enlarged area.This rare species would now be represented by anestimated 115 individuals, as illustrated in Figure 2aby moving vertically upwards to this value. All otherciliate species will keep the same relative abundances they had in the smaller area, so the linearslope of the new, upwards displaced, rank-abundance plot will be the same as the original (brokenline terminating with an arrow, in Figs 2a, b). Thisnew theoretical plot (for 1 m2) indicates that the ciliate with an abundance of 115 individuals per 1 m2
will, when the additional species are added to therank abundance plot, terminate the species sequence at 212 species (Le. there are estimated to be212 species in 1 m2
, when the rarest species in thatarea is represented by a single individual). The sameprocedure can be used to extrapolate to areas on a'global' scale (e.g. 2x106 km2 - the area of inlandfresh waters in the world) in Figure 3, at which point
the projected total is 597 species. When the sameprocedure is carried out for the freshwater benthicciliates, the projected total is 732 species (Table 1).
Discussion
We find that the global extrapolations are within afactor of two of the numbers of species derived froman analysis of species descriptions in the international published literature (Table 1). Furthermore,there is good reason for believing that the correspondence between the two types of estimate maybe even better than this, because the number ofspecies estimated from taxonomic analysis is probably still too high. There is no doubt that many synonyms remain embedded in the published literature,especially in the many crowded genera that requiretaxonomic revision (see Finlay et al. 1996b). Oneproblem that may be unique to the ciliates (and possibly other protists) is the practice of creating newgenera to accommodate the overflow from crowdedgenera. This has produced a very large number ofsingle-species genera (372 out of a total of 774 ciliate genera; Finlay et al. 1996b), making it even more

34 B. J. Finlay, G. F. Esteban, and T. Fenchel
laborious to discover synonyms. Another problemthat is certainly not unique to ciliates is that toomany 'new' species are still being described on thebasis of trivial differences, or in ignorance of speciesdescriptions that have already been published. Onelikely consequence of the resolution of these problems is that our estimates of global species richnessmay, in the course of time, show even better convergence than they do at present. In addition, we mayalso have some tentative evidence that the extrapolations provide realistic estimates on a local scale.We are engaged in an intensive long-term study ofthe ciliate fauna in a natural, freshwater pond in England (e.g. Finlay et al. 1988; 1996a,c). The number ofciliate species that are predicted to find suitablehabitats in one hectare (the area of the pond) is 375(from the polynomial in Fig. 3). Our list for the pondcurrently contains 244 ciliate species (only one ofwhich may be new to science); and although thenumber continues to grow with continued samplingeffort, it is becoming noticeably more difficult torecord additional species for this water body.
We have assumed throughout that ciliates areubiquitous. Absolute abundance is so large, andpassive dispersal so effective, that every specieshas some probability of being transported, at somepoint in time, anywhere in the biosphere. Eachspecies will grow and reproduce where it finds asuitable habitat, and if the habitats are found inmany different parts of the world, that species willbe considered to have a cosmopolitan distribution.But perhaps we are mistaken - perhaps we are unable to detect subtle but important differences separating ciliates, so we identify ciliates from differentplaces as the same species only because we are unable to tell them apart. These separate speciescould have different spatial distributions, and in anextreme case, a ciliate referred to as a cosmopolitanmorphospecies could consist of many similarspecies, each with its own geographical distribution.There is, however, one good piece of evidence indicating that this is not usually the case. In some ciliate genera (e.g. Paramecium and Tetrahymena), abiological species concept does apply. The differentsyngens are in most cases morphologically indistinguishable, and yet they are readily identifiable usinglaboratory tests of their mutual reproductive isolation. We would expect this reproductive isolation tobe correlated with geographic isolation, but the evidence is to the contrary. Most syngens in the Paramecium aurelia complex have cosmopolitan distributions (Nyberg 1988), many in the Tetrahymenapyriformis complex have been found on two or morecontinents (Nanney and McCoy 1976), and the apparent absence of species from other regions in the
world might easily be contradicted with additionalsampling effort (Corliss and Daggett 1983). It appears as if those ciliates holding the most promise ofrevealing a species biogeography within a commonmorphotype, fail to do just that.
A second argument against cosmopolitanism inciliates is fuelled by the so-called endemics. Newspecies do continue to be discovered. Invariablythese come from unusual or previously unexploredhabitats, such as solution lakes (Esteban et al.1993), wetlands in tropical Africa (Dragesco andDragesco-Kerneis 1986 ) or Antarctic sea-ice (Petzet al. 1995). The relevant point is that these speciesare found in these places because of the habitatsthat the places provide. This is graphically illustratedby the species of sea-ice ciliates that appear to beidentical in the Arctic and in the Antarctic (Agatha etal. 1993; Petz et al. 1995). There is also an undeniable tendency for 'endemics' to acquire a broadergeographical distribution in response to additionalsampling effort, and the true number is probably low(Fenchel 1993; Foissner 1997). Wilbert and Kahan(1981) described a very large and unusual ciliate(Condylostoma reichi) from Solar Lake in Eilat. It wassubsequently found in tropical Africa (Dragesco andDragesco-Kerneis 1986), and a ciliate most closelyresembling C. reichi has recently been found in theAntarctic (Petz et al. 1995). An 'endemic' of theHawaiian archipelago (Foissner 1994) was recentlyfound in wet moss by a river in central Spain (Olmoand Tellez 1996).
The assumption of cosmopolitanism in free-livingciliates is, in general, justified, and it may even betrue for the ciliates living in habitats that are relatively rare. These ciliates will have lower absoluteglobal abundances and (for purely statistical reasons) rates of dispersal that are much lower thanthose of ciliates living in common habitats. Ciliatesliving and growing on unusual 'islands' that are separated by large distances (e.g. the sea-ice of theArctic and the Antarctic) may rarely if ever be detected in intermediate regions; but the evidencedoes indicate the reality of global dispersal of these'island' species, even if the magnitude of this dispersal is small compared to that of the many common ciliate morphospecies co-occurring in common habitats in, for example, the Antarctic, Nigeriaand Scotland.
We have recently (July 1997) obtained some supporting evidence from a real 'island' - a small lakelying in the crater of an extinct volcano in Victoria,Australia. We are still adding species to the recordfor this water body, but preliminary results indicatethat of the 80 ciliate species recorded, 79 are already known from Northern Europe. The only other
34 B. J. Finlay, G. F. Esteban, and T. Fenchel
laborious to discover synonyms. Another problemthat is certainly not unique to ciliates is that toomany 'new' species are still being described on thebasis of trivial differences, or in ignorance of speciesdescriptions that have already been published. Onelikely consequence of the resolution of these problems is that our estimates of global species richnessmay, in the course of time, show even better convergence than they do at present. In addition, we mayalso have some tentative evidence that the extrapolations provide realistic estimates on a local scale.We are engaged in an intensive long-term study ofthe ciliate fauna in a natural, freshwater pond in England (e.g. Finlay et al. 1988; 1996a,c). The number ofciliate species that are predicted to find suitablehabitats in one hectare (the area of the pond) is 375(from the polynomial in Fig. 3). Our list for the pondcurrently contains 244 ciliate species (only one ofwhich may be new to science); and although thenumber continues to grow with continued samplingeffort, it is becoming noticeably more difficult torecord additional species for this water body.
We have assumed throughout that ciliates areubiquitous. Absolute abundance is so large, andpassive dispersal so effective, that every specieshas some probability of being transported, at somepoint in time, anywhere in the biosphere. Eachspecies will grow and reproduce where it finds asuitable habitat, and if the habitats are found inmany different parts of the world, that species willbe considered to have a cosmopolitan distribution.But perhaps we are mistaken - perhaps we are unable to detect subtle but important differences separating ciliates, so we identify ciliates from differentplaces as the same species only because we are unable to tell them apart. These separate speciescould have different spatial distributions, and in anextreme case, a ciliate referred to as a cosmopolitanmorphospecies could consist of many similarspecies, each with its own geographical distribution.There is, however, one good piece of evidence indicating that this is not usually the case. In some ciliate genera (e.g. Paramecium and Tetrahymena), abiological species concept does apply. The differentsyngens are in most cases morphologically indistinguishable, and yet they are readily identifiable usinglaboratory tests of their mutual reproductive isolation. We would expect this reproductive isolation tobe correlated with geographic isolation, but the evidence is to the contrary. Most syngens in the Paramecium aurelia complex have cosmopolitan distributions (Nyberg 1988), many in the Tetrahymenapyriformis complex have been found on two or morecontinents (Nanney and McCoy 1976), and the apparent absence of species from other regions in the
world might easily be contradicted with additionalsampling effort (Corliss and Daggett 1983). It appears as if those ciliates holding the most promise ofrevealing a species biogeography within a commonmorphotype, fail to do just that.
A second argument against cosmopolitanism inciliates is fuelled by the so-called endemics. Newspecies do continue to be discovered. Invariablythese come from unusual or previously unexploredhabitats, such as solution lakes (Esteban et al.1993), wetlands in tropical Africa (Dragesco andDragesco-Kerneis 1986 ) or Antarctic sea-ice (Petzet al. 1995). The relevant point is that these speciesare found in these places because of the habitatsthat the places provide. This is graphically illustratedby the species of sea-ice ciliates that appear to beidentical in the Arctic and in the Antarctic (Agatha etal. 1993; Petz et al. 1995). There is also an undeniable tendency for 'endemics' to acquire a broadergeographical distribution in response to additionalsampling effort, and the true number is probably low(Fenchel 1993; Foissner 1997). Wilbert and Kahan(1981) described a very large and unusual ciliate(Condylostoma reichi) from Solar Lake in Eilat. It wassubsequently found in tropical Africa (Dragesco andDragesco-Kerneis 1986), and a ciliate most closelyresembling C. reichi has recently been found in theAntarctic (Petz et al. 1995). An 'endemic' of theHawaiian archipelago (Foissner 1994) was recentlyfound in wet moss by a river in central Spain (Olmoand Tellez 1996).
The assumption of cosmopolitanism in free-livingciliates is, in general, justified, and it may even betrue for the ciliates living in habitats that are relatively rare. These ciliates will have lower absoluteglobal abundances and (for purely statistical reasons) rates of dispersal that are much lower thanthose of ciliates living in common habitats. Ciliatesliving and growing on unusual 'islands' that are separated by large distances (e.g. the sea-ice of theArctic and the Antarctic) may rarely if ever be detected in intermediate regions; but the evidencedoes indicate the reality of global dispersal of these'island' species, even if the magnitude of this dispersal is small compared to that of the many common ciliate morphospecies co-occurring in common habitats in, for example, the Antarctic, Nigeriaand Scotland.
We have recently (July 1997) obtained some supporting evidence from a real 'island' - a small lakelying in the crater of an extinct volcano in Victoria,Australia. We are still adding species to the recordfor this water body, but preliminary results indicatethat of the 80 ciliate species recorded, 79 are already known from Northern Europe. The only other

record for the remaining species (Oxytricha salmastraY is from tropical Africa (Dragesco and DragescoKernt3is, 1986). This directed search for 'endemics'in a place where we might have had a realisticchance of finding some, has so far failed to revealany.
Species-area relations, and extinctions
Two important features of the species-richness offree-living ciliates become clear, and both are a consequence of their small size. First, in comparisonwith macroscopic animals and plants, individualsand species are both very densely packed in nature.A square metre of freshwater benthos will typicallycontain 2x107 ciliates represented by 234 species.The equivalent figures for the marine interstitial are107 ciliates and 212 species. How do so manyspecies manage to live in a small area? The answeris easily explained in terms of 'fractal geometry':that the world is equally complex at all scales, so forexample 1cm2 of sediment could be as complex ahabitat for protozoa as a beech tree in mid-summeris for insects. The reality of this phenomenon will ofcourse be obvious to anyone who has used a microscope to examine a small sediment aggregate andfound attached peritrich ciliates, Chilodonella andAspidisca crawling and browsing over the surface,Spirostomum and various small scuticociliates filterfeeding in the pore volume, and the neck of an embedded Lacrymaria periodically emerging to grabalgae and other microbial food items.
The second important feature is the low rate ofspecies addition for increasing area. The bestknown general equation for the relation is S = CN,where S is number of species, A is area, and C and zare constants that vary from one group of organismsto another (MacArthur and Wilson 1967). In moststudies where the relationship has been fitted (Le.the macroscopic flora and fauna of islands), z takesa value in the range 0.2 to 0.35. The value is usuallysmaller (0.12-0.17) when the areas are locatedwithin continents (reflecting enhanced migration between areas); and birds (easily dispersed) havelower values than land snails. The average slope ofthe extrapolation for ciliates in Figure 3 (z = 0.043)falls well below either of these ranges and is consistent with the high rates of dispersal assumed for ciliates. In this connection it is interesting that Dodson(1991, 1992) also reports a low value (z = 0.05) foranother group of relatively small and easily dispersed animals - the crustacean zooplankton of European and North American lakes. He too, ascribesthe low value to high rates of immigration fromneighbouring areas.
Protozoan Diversity 35
One important implication of the low slope for theciliate species-area relationship is that the global diversity of ciliate species is largely unaffected by lossof habitat. One can readily calculate (e.g. Wilson,1992) that in the larger metazoans, with a typical zvalue of 0.3, a reduction in habitat area (e.g. of rainforest) to one tenth of its original size, will eventuallylead to loss of one half of the original number ofspecies. For ciliates (z = 0.043), the same reductionin area would lead to a 10% reduction in speciesnumber. But even this level of threatened extinctionis probably unrealistically severe, as a specific ciliatehabitat is unlikely to be lost simultaneously from allplaces in the biosphere where it exists (and thesame ciliate species probably live in 'rainforests'that are geographically isolated from each other).Moreover, at a local level, habitat destruction is perhaps only rarely so thorough that it reduces theabundance of any ciliate species-population to suchan extent that stochastic extinction becomes likely.
If our extrapolation was based on only onedataset, the closeness of the global estimate to thatobtained by taxonomic analysis might be considered fortuitous. The fact that the two independentdata sets can each be extrapolated to show thesame general species-area relation and that theseextrapolations converge with the additional independent estimate of taxonomic analysis indicatesthat our conclusions are firm: free-living ciliatespecies are ubiquitous, many have cosmopolitandistributions and their global species richness is relatively low.
Methods
Marine data: The localities, sampling techniquesand enumeration methods are described in Fenchel(1969). Sampling was concentrated within threemain areas, and at water depths down to 22 m: inthe 0resund, the Isefjord area, and in the Balticsouth of Stockholm. The largest data set is fromHelsing0r Beach (Denmark).
Freshwater data: The largest dataset we usedwas derived from a study of the benthos of a lake(Esthwaite Water) in the UK. These data were obtained from monthly sampling in the period October1978 to November 1981. A total of 228 Jenkin sediment cores were collected. The data from AirthreyLoch in Scotland (UK) were obtained from examination of 92 sediment cores taken with approximatelymonthly sampling at three sites in the period January 1975 to December 1976. The remainder of thedata were obtained from 18 excursions to samplefive shallow freshwater streams on the Jos Plateau
record for the remaining species (Oxytricha salmastraY is from tropical Africa (Dragesco and DragescoKernt3is, 1986). This directed search for 'endemics'in a place where we might have had a realisticchance of finding some, has so far failed to revealany.
Species-area relations, and extinctions
Two important features of the species-richness offree-living ciliates become clear, and both are a consequence of their small size. First, in comparisonwith macroscopic animals and plants, individualsand species are both very densely packed in nature.A square metre of freshwater benthos will typicallycontain 2x107 ciliates represented by 234 species.The equivalent figures for the marine interstitial are107 ciliates and 212 species. How do so manyspecies manage to live in a small area? The answeris easily explained in terms of 'fractal geometry':that the world is equally complex at all scales, so forexample 1cm2 of sediment could be as complex ahabitat for protozoa as a beech tree in mid-summeris for insects. The reality of this phenomenon will ofcourse be obvious to anyone who has used a microscope to examine a small sediment aggregate andfound attached peritrich ciliates, Chilodonella andAspidisca crawling and browsing over the surface,Spirostomum and various small scuticociliates filterfeeding in the pore volume, and the neck of an embedded Lacrymaria periodically emerging to grabalgae and other microbial food items.
The second important feature is the low rate ofspecies addition for increasing area. The bestknown general equation for the relation is S = CN,where S is number of species, A is area, and C and zare constants that vary from one group of organismsto another (MacArthur and Wilson 1967). In moststudies where the relationship has been fitted (Le.the macroscopic flora and fauna of islands), z takesa value in the range 0.2 to 0.35. The value is usuallysmaller (0.12-0.17) when the areas are locatedwithin continents (reflecting enhanced migration between areas); and birds (easily dispersed) havelower values than land snails. The average slope ofthe extrapolation for ciliates in Figure 3 (z = 0.043)falls well below either of these ranges and is consistent with the high rates of dispersal assumed for ciliates. In this connection it is interesting that Dodson(1991, 1992) also reports a low value (z = 0.05) foranother group of relatively small and easily dispersed animals - the crustacean zooplankton of European and North American lakes. He too, ascribesthe low value to high rates of immigration fromneighbouring areas.
Protozoan Diversity 35
One important implication of the low slope for theciliate species-area relationship is that the global diversity of ciliate species is largely unaffected by lossof habitat. One can readily calculate (e.g. Wilson,1992) that in the larger metazoans, with a typical zvalue of 0.3, a reduction in habitat area (e.g. of rainforest) to one tenth of its original size, will eventuallylead to loss of one half of the original number ofspecies. For ciliates (z = 0.043), the same reductionin area would lead to a 10% reduction in speciesnumber. But even this level of threatened extinctionis probably unrealistically severe, as a specific ciliatehabitat is unlikely to be lost simultaneously from allplaces in the biosphere where it exists (and thesame ciliate species probably live in 'rainforests'that are geographically isolated from each other).Moreover, at a local level, habitat destruction is perhaps only rarely so thorough that it reduces theabundance of any ciliate species-population to suchan extent that stochastic extinction becomes likely.
If our extrapolation was based on only onedataset, the closeness of the global estimate to thatobtained by taxonomic analysis might be considered fortuitous. The fact that the two independentdata sets can each be extrapolated to show thesame general species-area relation and that theseextrapolations converge with the additional independent estimate of taxonomic analysis indicatesthat our conclusions are firm: free-living ciliatespecies are ubiquitous, many have cosmopolitandistributions and their global species richness is relatively low.
Methods
Marine data: The localities, sampling techniquesand enumeration methods are described in Fenchel(1969). Sampling was concentrated within threemain areas, and at water depths down to 22 m: inthe 0resund, the Isefjord area, and in the Balticsouth of Stockholm. The largest data set is fromHelsing0r Beach (Denmark).
Freshwater data: The largest dataset we usedwas derived from a study of the benthos of a lake(Esthwaite Water) in the UK. These data were obtained from monthly sampling in the period October1978 to November 1981. A total of 228 Jenkin sediment cores were collected. The data from AirthreyLoch in Scotland (UK) were obtained from examination of 92 sediment cores taken with approximatelymonthly sampling at three sites in the period January 1975 to December 1976. The remainder of thedata were obtained from 18 excursions to samplefive shallow freshwater streams on the Jos Plateau

36 B. J. Finlay, G. F. Esteban, and T. Fenchel
in Nigeria, in the period December 1977 to May1978. The enumeration methods used are describedin Finlay and Guhl (1992). Further information relating to the Airthrey and Esthwaite datasets appearsin Finlay (1980, 1982 respectively).
Taxonomic analysis: The method used inanalysing all ciliate species descriptions publishedin the period 1758 to 1996, was published in Finlayet al. (1996b). The dataset established therein hasbeen further analysed, with allocation of all free-living ciliate species to one of two categories: marine,and non-marine. Some ciliates (e.g. Cyclidiumglaucoma) live in the sea and in fresh water, sonumbers of these have been divided equally between the marine and non-marine categories. Themarine category was then further divided - intothose species that are typically interstitial, andthose that are not (e.g. tintinnids and other planktonic ciliates). This procedure is not easily appliedto the freshwater ciliates, of which few can be classified as truly interstitial species. The interstitialhabitat is much rarer in the typically finely-grainedfreshwater sediments, and many freshwater ciliatesperform seasonal benthic-planktonic migrations inresponse to the development of deep-water anoxia·(Finlay 1981). But many freshwater ciliates are apparently well-adapted for a permanently planktoniclifestyle (e.g. many oligotrichs), and we finally allocated species to the freshwater benthos category ifthey were not obviously planktonic, and if theweight of evidence (our personal experience combined with that in the published literature) indicatedthat the benthos provided their 'preferred' habitat.It may be noted that some (perhaps many) 'soil ciliates' also make a living in fresh waters (Foissner1987); especially in riverine and littoral sediments.These species are included in our 'freshwater benthos' total in Table 1.
The species concept used throughout is the concept of 'morphospecies' as described in Finlay et al.(1996b).
Data handling: All data handling and statisticalprocesses were performed using Microsoft EXCEL(5.0).
Acknowledgements
This work was supported financially by the Centrefor Ecology and Hydrology (NERC, United Kingdom), The British Council (newIMAGES), and theDanish Natural Science Research Council. The interim results from Australia are the product of an ongoing collaboration with Prof. PA Tyler, Deakin University, Australia.
References
Agatha 5, Spindler M, Wilbert N (1993) Ciliated protozoa (Ciliophora) from Arctic sea ice. Acta Protozool 32:261-268
Bary BM (1950) Studies on the freshwater ciliates ofNew Zealand. Part II. An annotated list of species fromthe neighbourhood of Wellington. Proc Roy Soc N.Z.78:311-323
Bowers NJ, Pratt JR (1995) Estimation of genetic variation among soil ciliates of Colpoda inflata (Stokes)(Protozoa: Ciliophora) using the polymerase chain reaction and restriction fragment length polymorphism analysis. Arch Protistenkd 145: 29-36
Corliss JO (1974) Time for evolutionary biologists totake more interest in phylogenetics? Taxon 23: 497-522
Corliss JO, Daggett P-M (1983) 'Paramecium aurelia'and 'Tetrahymena pyriformis'; current status of the taxonomy and nomenclature of these popularly known andwidely used ciliates. Protistologica 19: 307-322
Dodson 5 (1991) Species richness of crustacean zooplankton in European lakes of different sizes. Verh Internat Verein theor ang Limnol 24: 1223-1229
Dodson 5 (1992) Predicting crustacean zooplanktonspecies richness. Limnol Oceanogr 37: 848-856
Dragesco J, Dragesco-Kerneis A (1986) Cilies Libresde l'Afrique Intertropicale. [Collection Faune Tropicaleno. 26], ORSTOM, Paris
Ekebom J, Patterson OJ, Vors N (1996) Heterotrophicflagellates from coral reef sediments (Great BarrierReef, Australia). Arch Protistenkd 146: 251-272
Esteban G, Finlay BJ, Embley TM (1993) New speciesdouble the diversity of anaerobic ciliates in a Spanishlake. FEMS Microbiol Lett 109: 93-100
Fenchel T (1969) The ecology of marine microbenthosIV. Structure and function of the benthic ecosystem, itschemical and physical factors and the microfauna communities with special reference to the ciliated protozoa.Ophelia 6: 1-182
Fenchel T (1993) There are more small than largespecies? Oikos 68: 375-378
Fenchel T, Esteban GF, Finlay BJ (1997) Local versusglobal diversity of microorganisms: cryptic diversity ofciliated protozoa. Oikos 80: 220-225
Finlay BJ (1980) Temporal and vertical distribution ofciliophoran communities in the benthos of a small eutrophic loch with particular reference to the redox profile. Freshwat Bioi 10: 15-34
Finlay BJ (1981) Oxygen availability and seasonal migrations of ciliated protozoa in a freshwater lake. J GenMicrobiol123: 173-178
Finlay BJ (1982) Effects of seasonal anoxia on the community of benthic ciliated protozoa in a productive lake.Arch Protistenkd 125: 215-222
36 B. J. Finlay, G. F. Esteban, and T. Fenchel
in Nigeria, in the period December 1977 to May1978. The enumeration methods used are describedin Finlay and Guhl (1992). Further information relating to the Airthrey and Esthwaite datasets appearsin Finlay (1980, 1982 respectively).
Taxonomic analysis: The method used inanalysing all ciliate species descriptions publishedin the period 1758 to 1996, was published in Finlayet al. (1996b). The dataset established therein hasbeen further analysed, with allocation of all free-living ciliate species to one of two categories: marine,and non-marine. Some ciliates (e.g. Cyclidiumglaucoma) live in the sea and in fresh water, sonumbers of these have been divided equally between the marine and non-marine categories. Themarine category was then further divided - intothose species that are typically interstitial, andthose that are not (e.g. tintinnids and other planktonic ciliates). This procedure is not easily appliedto the freshwater ciliates, of which few can be classified as truly interstitial species. The interstitialhabitat is much rarer in the typically finely-grainedfreshwater sediments, and many freshwater ciliatesperform seasonal benthic-planktonic migrations inresponse to the development of deep-water anoxia·(Finlay 1981). But many freshwater ciliates are apparently well-adapted for a permanently planktoniclifestyle (e.g. many oligotrichs), and we finally allocated species to the freshwater benthos category ifthey were not obviously planktonic, and if theweight of evidence (our personal experience combined with that in the published literature) indicatedthat the benthos provided their 'preferred' habitat.It may be noted that some (perhaps many) 'soil ciliates' also make a living in fresh waters (Foissner1987); especially in riverine and littoral sediments.These species are included in our 'freshwater benthos' total in Table 1.
The species concept used throughout is the concept of 'morphospecies' as described in Finlay et al.(1996b).
Data handling: All data handling and statisticalprocesses were performed using Microsoft EXCEL(5.0).
Acknowledgements
This work was supported financially by the Centrefor Ecology and Hydrology (NERC, United Kingdom), The British Council (newIMAGES), and theDanish Natural Science Research Council. The interim results from Australia are the product of an ongoing collaboration with Prof. PA Tyler, Deakin University, Australia.
References
Agatha 5, Spindler M, Wilbert N (1993) Ciliated protozoa (Ciliophora) from Arctic sea ice. Acta Protozool 32:261-268
Bary BM (1950) Studies on the freshwater ciliates ofNew Zealand. Part II. An annotated list of species fromthe neighbourhood of Wellington. Proc Roy Soc N.Z.78:311-323
Bowers NJ, Pratt JR (1995) Estimation of genetic variation among soil ciliates of Colpoda inflata (Stokes)(Protozoa: Ciliophora) using the polymerase chain reaction and restriction fragment length polymorphism analysis. Arch Protistenkd 145: 29-36
Corliss JO (1974) Time for evolutionary biologists totake more interest in phylogenetics? Taxon 23: 497-522
Corliss JO, Daggett P-M (1983) 'Paramecium aurelia'and 'Tetrahymena pyriformis'; current status of the taxonomy and nomenclature of these popularly known andwidely used ciliates. Protistologica 19: 307-322
Dodson 5 (1991) Species richness of crustacean zooplankton in European lakes of different sizes. Verh Internat Verein theor ang Limnol 24: 1223-1229
Dodson 5 (1992) Predicting crustacean zooplanktonspecies richness. Limnol Oceanogr 37: 848-856
Dragesco J, Dragesco-Kerneis A (1986) Cilies Libresde l'Afrique Intertropicale. [Collection Faune Tropicaleno. 26], ORSTOM, Paris
Ekebom J, Patterson OJ, Vors N (1996) Heterotrophicflagellates from coral reef sediments (Great BarrierReef, Australia). Arch Protistenkd 146: 251-272
Esteban G, Finlay BJ, Embley TM (1993) New speciesdouble the diversity of anaerobic ciliates in a Spanishlake. FEMS Microbiol Lett 109: 93-100
Fenchel T (1969) The ecology of marine microbenthosIV. Structure and function of the benthic ecosystem, itschemical and physical factors and the microfauna communities with special reference to the ciliated protozoa.Ophelia 6: 1-182
Fenchel T (1993) There are more small than largespecies? Oikos 68: 375-378
Fenchel T, Esteban GF, Finlay BJ (1997) Local versusglobal diversity of microorganisms: cryptic diversity ofciliated protozoa. Oikos 80: 220-225
Finlay BJ (1980) Temporal and vertical distribution ofciliophoran communities in the benthos of a small eutrophic loch with particular reference to the redox profile. Freshwat Bioi 10: 15-34
Finlay BJ (1981) Oxygen availability and seasonal migrations of ciliated protozoa in a freshwater lake. J GenMicrobiol123: 173-178
Finlay BJ (1982) Effects of seasonal anoxia on the community of benthic ciliated protozoa in a productive lake.Arch Protistenkd 125: 215-222

Finlay BJ (1997). The diversity and ecological role ofprotozoa in fresh waters. In Sutcliffe DW, Jones JG(eds) The Microbiological Quality of Water. FBAIISWA,pp 108-125
Finlay BJ, Berninger U-G, Clarke KJ, Cowling AJ,Hindle RM, Rogerson A (1988) On the abundance anddistribution of protozoa and their food in a productivefreshwater pond. Europ J Protistol23: 205-217
Finlay BJ, Esteban GF, Fenchel T (1996a) Global diversity and body size. Nature 383:132-133
Finlay BJ, Corliss JO, Esteban GF, Fenchel T (1996b)Biodiversity at the microbial level: the number of freeliving ciliates in the biosphere. Quart Rev Bioi 71:221-237
Finlay BJ, Maberly SC, Esteban GF (1996c) Spectacular abundance of ciliates in anoxic pond water: contribution of symbiont photosynthesis to host respiratoryoxygen requirements. FEMS Microbiol Ecol 20:229-235
Finlay BJ, Guhl BE (1992) Benthic sampling - freshwater. Pages In Lee JJ, Soldo AT (eds) Protocols in Protozology. Society of Protozoologists, Lawrence,Kansas, pp B-2.1- 5
Foissner W (1987) Soil protozoa: fundamental problems, ecological significance, adaptations in ciliatesand testaceans, bioindicators, and guide to the literature. Prog Protistol 2: 69-212.
Foissner W (1994) Bryometopus hawaiiensis sp. n., anew colpodid ciliate from a terrestrial biotope of theHawaiian Archipelago. Annals Naturhist Mus Wien 96B:19-27
Foissner W (1997) Global soil ciliate (Protozoa, Ciliophora) diversity: a probability-based approach usinglarge sample collectives from Africa, Australia, andAntarctica. Biodiv Conserv (In press)
Gaston K J (1992) Regional numbers of insect andplant species. Funct Ecol6: 243-247
Holmberg 0, Pejler B (1972) On the terrestrial microfauna of Surtsey during the summer 1970. Surtsey ResProgr Rep 6: 69-72
Kristiansen J (1996) Dispersal of freshwater algae - areview. Hydrobiologia 336: 151-157
Lackey JB (1938) A study of some ecologic factors affecting the distribution of protozoa. Ecol Monogr 8:503-527
Larsen J, Patterson OJ (1990) Some flagellates (Protista) from tropical marine sediments. J Nat Hist 24:801-937
MacArthur RH, Wilson EO (1967) The Theory of IslandBiogeography. Princeton University Press, Princeton
Maguire B (1963) The passive dispersal of smallaquatic organisms and their colonization of isolatedbodies of water. Ecol Monogr33: 161-185
Maguire B, Belk 0 (1967) Paramecium transport byland snails. J Protozool 14: 445-447
Protozoan Diversity 37
May RM (1988) How many species are there on Earth?Science 241: 1441-1449
May RM (1990) How many species? Phil Trans RSLond 330: 293-304
Nanney OL, McCoy JW (1976) Characterization of thespecies of the Tetrahymena pyriformis complex. TransAmer Microsc Soc 95: 664-682
Nyberg 0 (1988) The species concept and breedingsystems. In G6rtz HD (ed) Paramecium. Springer-Verlag, Berlin, pp 41-58
Ogden CG, Hedley RH (1980) An Atlas of FreshwaterTestate Amoebae. British Museum/Oxford UniversityPress, Oxford
Olmo JL, Tellez C (1996). An European population ofBryometopus hawaiiensis Foissner, 1994 (Protozoa: Ciliophora). Acta Protozool 35: 317-320
Parsons WM, Schlichting HE, Stewart KW (1966) Inflight transport of algae and protozoa by selectedOdonata. Trans Amer Microsc Soc 85: 520-527
Patterson OJ, Simpson AGB (1996) Heterotrophicflagellates from coastal marine and hypersaline sediments in Western Australia. Europ J Protistol 32:423-448
Petz W, Song W, Wilbert N (1995) Taxonomy and ecology of the ciliate fauna (Protozoa Ciliophora) in the endopagial and pelagial of the Weddell Sea Antarctica.Stapfia 40: 223pp.
Preston FW (1948) The commonness and rarity ofspecies. Ecology 29: 254-283
Sandon H (1927) The Composition and Distribution ofthe Protozoan Fauna of the Soil. Oliver and Boyd, Edinburgh.
Schlichting HE, Sides SL (1969) The passive transportof aquatic microorganisms by selected Hemiptera. JEcoI57:759-764
Scourfield OJ (1944) The nannoplankton of bombcrater pools in Epping Forest. Essex Natural 27:231-241
Smith HG (1978) The distribution and ecology of terrestrial protozoa of sub-Antarctic and maritime Antarcticislands. Scient Reps Brit Antarc Surv No 95,BAS/NERC, Cambridge
Stout JO (1956) Reaction of ciliates to environmentalfactors. Ecology 37: 178-191
Tyler PA (1996) Endemism in freshwater algae. Hydrobiologia 336: 127-135
UNEP (1995) Global Biodiversity Assessment. Heywood VH (exec ed) Cambridge University Press, Cambridge
Wilbert N, Kahan 0 (1981) Ciliates of Solar Lake on theRed Sea shore. Arch Protistenkd 124: 70-95
Wilson EO (1992) The diversity of life. Penguin, London
Zink RM (1996) Bird species diversity. Nature 381: 566
Finlay BJ (1997). The diversity and ecological role ofprotozoa in fresh waters. In Sutcliffe DW, Jones JG(eds) The Microbiological Quality of Water. FBAIISWA,pp 108-125
Finlay BJ, Berninger U-G, Clarke KJ, Cowling AJ,Hindle RM, Rogerson A (1988) On the abundance anddistribution of protozoa and their food in a productivefreshwater pond. Europ J Protistol23: 205-217
Finlay BJ, Esteban GF, Fenchel T (1996a) Global diversity and body size. Nature 383:132-133
Finlay BJ, Corliss JO, Esteban GF, Fenchel T (1996b)Biodiversity at the microbial level: the number of freeliving ciliates in the biosphere. Quart Rev Bioi 71:221-237
Finlay BJ, Maberly SC, Esteban GF (1996c) Spectacular abundance of ciliates in anoxic pond water: contribution of symbiont photosynthesis to host respiratoryoxygen requirements. FEMS Microbiol Ecol 20:229-235
Finlay BJ, Guhl BE (1992) Benthic sampling - freshwater. Pages In Lee JJ, Soldo AT (eds) Protocols in Protozology. Society of Protozoologists, Lawrence,Kansas, pp B-2.1- 5
Foissner W (1987) Soil protozoa: fundamental problems, ecological significance, adaptations in ciliatesand testaceans, bioindicators, and guide to the literature. Prog Protistol 2: 69-212.
Foissner W (1994) Bryometopus hawaiiensis sp. n., anew colpodid ciliate from a terrestrial biotope of theHawaiian Archipelago. Annals Naturhist Mus Wien 96B:19-27
Foissner W (1997) Global soil ciliate (Protozoa, Ciliophora) diversity: a probability-based approach usinglarge sample collectives from Africa, Australia, andAntarctica. Biodiv Conserv (In press)
Gaston K J (1992) Regional numbers of insect andplant species. Funct Ecol6: 243-247
Holmberg 0, Pejler B (1972) On the terrestrial microfauna of Surtsey during the summer 1970. Surtsey ResProgr Rep 6: 69-72
Kristiansen J (1996) Dispersal of freshwater algae - areview. Hydrobiologia 336: 151-157
Lackey JB (1938) A study of some ecologic factors affecting the distribution of protozoa. Ecol Monogr 8:503-527
Larsen J, Patterson OJ (1990) Some flagellates (Protista) from tropical marine sediments. J Nat Hist 24:801-937
MacArthur RH, Wilson EO (1967) The Theory of IslandBiogeography. Princeton University Press, Princeton
Maguire B (1963) The passive dispersal of smallaquatic organisms and their colonization of isolatedbodies of water. Ecol Monogr33: 161-185
Maguire B, Belk 0 (1967) Paramecium transport byland snails. J Protozool 14: 445-447
Protozoan Diversity 37
May RM (1988) How many species are there on Earth?Science 241: 1441-1449
May RM (1990) How many species? Phil Trans RSLond 330: 293-304
Nanney OL, McCoy JW (1976) Characterization of thespecies of the Tetrahymena pyriformis complex. TransAmer Microsc Soc 95: 664-682
Nyberg 0 (1988) The species concept and breedingsystems. In G6rtz HD (ed) Paramecium. Springer-Verlag, Berlin, pp 41-58
Ogden CG, Hedley RH (1980) An Atlas of FreshwaterTestate Amoebae. British Museum/Oxford UniversityPress, Oxford
Olmo JL, Tellez C (1996). An European population ofBryometopus hawaiiensis Foissner, 1994 (Protozoa: Ciliophora). Acta Protozool 35: 317-320
Parsons WM, Schlichting HE, Stewart KW (1966) Inflight transport of algae and protozoa by selectedOdonata. Trans Amer Microsc Soc 85: 520-527
Patterson OJ, Simpson AGB (1996) Heterotrophicflagellates from coastal marine and hypersaline sediments in Western Australia. Europ J Protistol 32:423-448
Petz W, Song W, Wilbert N (1995) Taxonomy and ecology of the ciliate fauna (Protozoa Ciliophora) in the endopagial and pelagial of the Weddell Sea Antarctica.Stapfia 40: 223pp.
Preston FW (1948) The commonness and rarity ofspecies. Ecology 29: 254-283
Sandon H (1927) The Composition and Distribution ofthe Protozoan Fauna of the Soil. Oliver and Boyd, Edinburgh.
Schlichting HE, Sides SL (1969) The passive transportof aquatic microorganisms by selected Hemiptera. JEcoI57:759-764
Scourfield OJ (1944) The nannoplankton of bombcrater pools in Epping Forest. Essex Natural 27:231-241
Smith HG (1978) The distribution and ecology of terrestrial protozoa of sub-Antarctic and maritime Antarcticislands. Scient Reps Brit Antarc Surv No 95,BAS/NERC, Cambridge
Stout JO (1956) Reaction of ciliates to environmentalfactors. Ecology 37: 178-191
Tyler PA (1996) Endemism in freshwater algae. Hydrobiologia 336: 127-135
UNEP (1995) Global Biodiversity Assessment. Heywood VH (exec ed) Cambridge University Press, Cambridge
Wilbert N, Kahan 0 (1981) Ciliates of Solar Lake on theRed Sea shore. Arch Protistenkd 124: 70-95
Wilson EO (1992) The diversity of life. Penguin, London
Zink RM (1996) Bird species diversity. Nature 381: 566