fifty-thousand-year vegetation and climate history of noel ... · fifty-thousand-year vegetation...
TRANSCRIPT

www.elsevier.com/locate/yqres
Quaternary Research 61 (2004) 215–230
Fifty-thousand-year vegetation and climate history of Noel Kempff
Mercado National Park, Bolivian Amazon
Rachel E. Burbridge,a Francis E. Mayle,a,* and Timothy J. Killeenb,c
aDepartment of Geography, University of Leicester, Leicester LE1 7RH, UKbMuseo de Historia Natural ‘‘Noel Kempff Mercado,’’ Avenida Irala 565, Casilla 2489, Santa Cruz de la Sierra, Santa Cruz, BoliviacCenter for Applied Biodiversity Science, Conservation International, 2501 M Street, NW, Suite 200, Washington DC 20037, USA
Received 14 October 2002
Abstract
Pollen and charcoal records from two large, shallow lakes reveal that throughout most of the past 50,000 yr Noel Kempff Mercado
National Park, in northeastern lowland Bolivia (southwestern Amazon Basin), was predominantly covered by savannas and seasonally dry
semideciduous forests. Lowered atmospheric CO2 concentrations, in combination with a longer dry season, caused expansion of dry forests
and savannas during the last glacial period, especially at the last glacial maximum. These ecosystems persisted until the mid-Holocene,
although they underwent significant species reassortment. Forest communities containing a mixture of evergreen and semideciduous species
began to expand between 6000 and 3000 14C yr B.P. Humid evergreen rain forests expanded to cover most of the area within the past 200014C yr B.P., coincident with a reduction in fire frequencies. Comparisons between modern pollen spectra and vegetation reveal that the
Moraceae-dominated rain forest pollen spectra likely have a regional source area at least 2–3 km beyond the lake shore, whereas the grass-
and sedge-dominated savanna pollen spectra likely have a predominantly local source area. The Holocene vegetation changes are consistent
with independent paleoprecipitation records from the Bolivian Altiplano and paleovegetation records from other parts of southwestern
Amazonia. The progressive expansion in rain forests through the Holocene can be largely attributed to enhanced convective activity over
Amazonia, due to greater seasonality of insolation in the Southern Hemisphere tropics driven by the precession cycle according to the
Milankovitch Astronomical Theory.
D 2004 University of Washington. All rights reserved.
Keywords: Amazon; Bolivia; Pollen; Last glaciation; Quaternary; Holocene; Rain forest; Dry forest; Savanna
Introduction previously published summary paper (Mayle et al., 2000) to
The Quaternary environmental history of Amazonia is of
great scientific interest, but remains poorly understood due
to the paucity of data from this vast area. The aim of this
paper is to present a 50,000-yr paleoenvironmental recon-
struction of the ecosystems of Noel Kempff Mercado
National Park (NKMNP), northeastern Bolivia. The study
area (Fig. 1), located near the southwestern limit of Ama-
zonia, was chosen because of its climatically sensitive
ecotonal position between humid evergreen rain forests to
the north, semideciduous dry forests to the south, and
upland cerrado savannas to the east. We expand upon our
0033-5894/$ - see front matter D 2004 University of Washington. All rights rese
doi:10.1016/j.yqres.2003.12.004
* Corresponding author. Address beginning July 2004: Institute of
Geography, School of GeoSciences, The University of Edinburgh,
Drummond Street, Edinburgh EH8 9XP, UK. Fax: +44-116-252-3854.
E-mail address: [email protected] (F.E. Mayle).
show detailed pollen- and charcoal-based vegetation histo-
ries from two lacustrine sediment sequences and to discuss
the implications of these findings for the late Quaternary
paleoclimatology of southwestern Amazonia.
Study area
NKMNP is a 15,230-km2 protected area in northeastern
Bolivia near the southern margin of Amazonia, adjacent to
the Brazilian states of Rondonia and Mato Grosso (Fig. 1).
The Huanchaca Plateau, 600 to 900 m above sea level (asl),
dominates the eastern half of the park and is composed of
Pre-Cambrian sandstone and quartzite of the Brazilian
Shield (Litherland and Power, 1989). This table mountain
is dominated by edaphically derived upland savannas but
also contains patches of evergreen and deciduous forests
where soils are sufficiently deep and nutrient rich. The
rved.
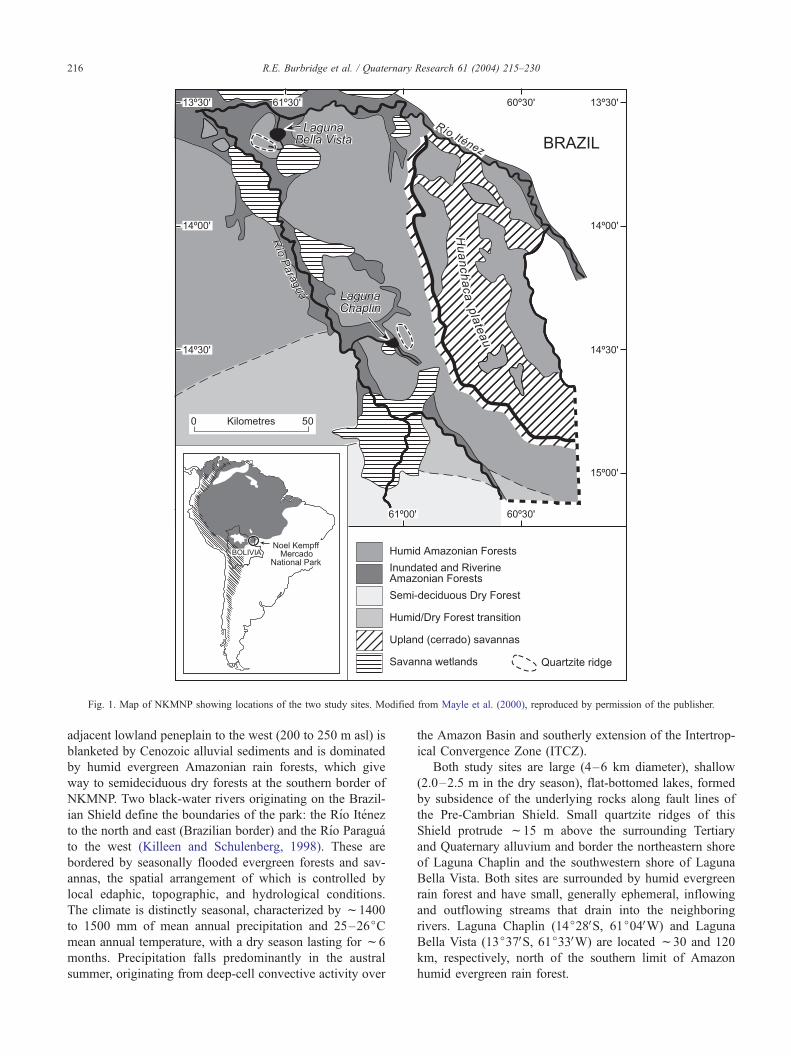
Fig. 1. Map of NKMNP showing locations of the two study sites. Modified from Mayle et al. (2000), reproduced by permission of the publisher.
R.E. Burbridge et al. / Quaternary Research 61 (2004) 215–230216
adjacent lowland peneplain to the west (200 to 250 m asl) is
blanketed by Cenozoic alluvial sediments and is dominated
by humid evergreen Amazonian rain forests, which give
way to semideciduous dry forests at the southern border of
NKMNP. Two black-water rivers originating on the Brazil-
ian Shield define the boundaries of the park: the Rıo Itenez
to the north and east (Brazilian border) and the Rıo Paragua
to the west (Killeen and Schulenberg, 1998). These are
bordered by seasonally flooded evergreen forests and sav-
annas, the spatial arrangement of which is controlled by
local edaphic, topographic, and hydrological conditions.
The climate is distinctly seasonal, characterized by f1400
to 1500 mm of mean annual precipitation and 25–26jCmean annual temperature, with a dry season lasting for f6
months. Precipitation falls predominantly in the austral
summer, originating from deep-cell convective activity over
the Amazon Basin and southerly extension of the Intertrop-
ical Convergence Zone (ITCZ).
Both study sites are large (4–6 km diameter), shallow
(2.0–2.5 m in the dry season), flat-bottomed lakes, formed
by subsidence of the underlying rocks along fault lines of
the Pre-Cambrian Shield. Small quartzite ridges of this
Shield protrude f15 m above the surrounding Tertiary
and Quaternary alluvium and border the northeastern shore
of Laguna Chaplin and the southwestern shore of Laguna
Bella Vista. Both sites are surrounded by humid evergreen
rain forest and have small, generally ephemeral, inflowing
and outflowing streams that drain into the neighboring
rivers. Laguna Chaplin (14j28VS, 61j04VW) and Laguna
Bella Vista (13j37VS, 61j33VW) are located f30 and 120
km, respectively, north of the southern limit of Amazon
humid evergreen rain forest.

R.E. Burbridge et al. / Quaternary Research 61 (2004) 215–230 217
Methodology
Laguna Bella Vista (LBV) was cored in August, 1995,
with a modified square-rod Livingstone piston corer
(Wright, 1967). The overlapping cores were taken near the
center of the lake. The inorganic clay sediments at the base
of these cores proved too stiff to penetrate further. Logistical
constraints prevented exploration of the sediment stratigra-
phy in other parts of the lake basin. Laguna Chaplin (LCH)
was cored in August, 1998, using a Geocore ‘‘drop-ham-
mer’’ piston corer (Colinvaux et al., 1999), f80 m from the
southern shore. This shoreline position was chosen because
reconnaissance cores with an auger revealed that the thick-
ness of the upper organic unit increased toward the south
shore. At both sites, uppermost surface sediments were
recovered with a clear Perspex plastic tube and piston. At
LCH, another two surface cores were taken farther from the
shore to allow determination of the extent of spatial varia-
tion in the surface pollen spectra across the basin.
Percentage loss-on-ignition (LOI) was undertaken to
determine organic content and facilitate correlation of over-
lapping cores. The scarcity of terrestrial plant macrofossils
(even after systematic sieving with a 250-Am mesh sieve)
necessitated accelerator mass spectrometer 14C (AMS) dat-
ing of bulk sediment samples. The absence of carbonate
deposits in the area meant that any dating anomalies due to a
‘‘bedrock hard-water effect’’ were likely to be negligible.
One-cubic-centimeter samples were processed for pollen
using standard techniques (Faegri and Iverson, 1989).
Pollen types were identified using published pollen floras
(Roubik and Moreno, 1991; Colinvaux et al., 1999) and
the modern pollen reference collection (f1000 taxa) at the
Fig. 2. Surface pollen spectra (sediment/water interface) of selected ta
University of Leicester. The latter was compiled from
herbarium material housed at the Noel Kempff Mercado
Natural History Museum, Santa Cruz, Bolivia, collected
from NKMNP. Pollen percentage, concentration, and
accumulation rate (PAR) diagrams were prepared using
the Psimpoll program (Bennett, 1997). Percentage calcu-
lations were based on the sum of total land pollen (TLP),
which included terrestrial Pteridophyte spores, Cyperaceae
pollen, and unidentified pollen types. Aquatic taxa and
indeterminable pollen types were excluded from the
terrestrial pollen sum. Pollen zones were produced by
comparing different statistical outputs (using binary and
optimal splitting and cluster analysis) as determined by
Psimpoll.
Concentrations of pollen-slide charcoal particles >12 Amwere determined. Since particle fragmentation occurs during
pollen processing, a sieving-based methodology, modified
from Rhodes (1998), was adopted to minimize fragmenta-
tion and determine the size-class distribution of charcoal
particles to obtain insights into differing charcoal source
areas (e.g., local vs regional). Particles were sorted into
categories of 54–180, 181–250, 251–500, and >500 Am.
Modern pollen and charcoal
Modern pollen percentages, both between the three
LCH surface samples and between the surface samples
of the two sites, show similarities (Fig. 2). All four modern
pollen spectra are dominated by 40–55% Moraceae/Urti-
caceae pollen, providing a signature indicative of the
surrounding seasonally flooded, riparian, evergreen rain
xa from Laguna Chaplin (LCH) and Laguna Bella Vista (LBV).

R.E. Burbridge et al. / Quaternary Research 61 (2004) 215–230218
forests and terra firme, humid, evergreen rain forests on
higher ground (Fig. 1). The dominance of the Moraceae
family (in both abundance and species richness) in these
forests (Killeen, 1998; Killeen et al., 2001) suggests that
the majority of grains identified as Moraceae/Urticaceae
type can be attributed to the Moraceae rather than the
Urticaceae family. Although the study area is predominantly
covered by rain forest, LBV lies within 5–10 km of several
areas of seasonally flooded savanna. LCH is bordered to the
south by a patch of savanna marsh and is only 20 km away
from extensive cerrado savannas on the Huanchaca Plateau
to the east and seasonally flooded savannas to the south.
However, pollen from these Poaceae- and Cyperaceae-
dominated ecosystems appears to be poorly dispersed and
significantly underrepresented, since the surface pollen
spectra of both sites contain <10% Poaceae and Cyper-
Fig. 3. Age–depth plot for (A) Laguna C
aceae pollen. These findings are supported by preliminary
percentage and PAR data (W.D. Gosling, personal commu-
nication, 2002) from artificial funnel pollen traps (Gosling
et al., 2003) from NKMNP, which show that seasonally
flooded savannas surprisingly contain more Moraceae than
Poaceae pollen (32% vs 18%, respectively) even though
the nearest Moraceae populations were f3 km away.
Clearly, the high Moraceae pollen percentages in the
surface sediment samples can be attributed to both local
populations growing around the lake shores and regional
populations several kilometers away. In contrast, the Poa-
ceae and Cyperaceae pollen have a predominantly local
source area, possibly no farther than 2–3 km from the lake
shore.
The complete absence of charcoal in the LBV surface
sediments (Fig. 9) attests to the rarity of fires in rain forest
haplin and (B) Laguna Bella Vista.

Table 2
AMS radiocarbon dates for Laguna Bella Vista
Laboratory
code
Sample depth
(cm)
AMS 14C yr B.P.
(F 1 j)Calibrated yr B.P.
(1 j)
AA39713 28–32 1,650 F 40 1,510 (1,530) 1,550
AA33345 40.5–41.5 2,740 F 50 2,860 (2,790) 2,770
AA39714 50.5–51.5 3,220 F40 3,460 (3,400) 3,370
R.E. Burbridge et al. / Quaternary Research 61 (2004) 215–230 219
ecosystems. It further demonstrates that the local rain forests
surrounding this lake serve as an effective barrier to char-
coal generated by the frequent fires we observed in the patch
of seasonally flooded savanna 5 km away. The presence of
charcoal (albeit negligible) in the surface sediments of LCH
(Fig. 6) is likely due to its closer proximity to the upland
savannas on the Huanchaca Plateau.
AA33344 60.5–61.5 7,980 F 70 9,000 (8,940, 8,930,8,860, 8,840, 8,780)
8,640
AA39715 80.5–81.5 9,380 F 70 10,650 (10,570) 10,490
AA39716 100.5–101.5 9,590 F 70 11,110 (11,060, 10,950,
10,770) 10,710
Beta-100829 121.5–122.5 9,720 F 60 11,180 (11,160) 11,100
AA39717 125.5–126.5 9,820 F 70 11,230 (11,200) 11,170
AA39718 133–133.5 11,030 F 80 13,130 (13,000) 12,930
AA33343 137.5–138.5 38,600 F 1000 —
AA39719 147.5–148.5 46,600 F 2900 —
AA39720 187.5–188.5 41,400 F 1500 —
AA33342 197.5–198.5 43,900 F 1900 —
AA33341 247.5–248.5 43,700 F 2000 —
Beta-100828 279–282 >50,850 —
All samples are carbonate-free bulk sediment, except for sample Beta-
100828, which consists of unidentified twig fragments. See Table 1 for
explanation.
Sediment chronology and stratigraphy
Sedimentation rates varied markedly, both within and
between LCH-3 and LBV (Fig. 3; Tables 1 and 2). At LCH
a sudden change occurred near 6000 14C yr B.P., with slow
sedimentation (0.02 mm yr�1) during the early Holocene
and much faster sedimentation (by a factor of 10) by the late
Holocene (0.2 mm yr�1). Deposition of Holocene sediments
at LBV was more complex, with a period of slow sedimen-
tation in the mid-Holocene (0.02 mm yr�1, 7980–3220 14C
yr B.P.) bracketed by more rapid deposition in the early and
late Holocene (1.0 and 0.2 mm yr�1, respectively). LBV
was completely dry between 11,030 and 38,600 14C yr B.P.,
but LCH apparently never dried up for any significant
period (i.e., millennia), although sedimentation was ex-
tremely slow throughout the Pleistocene. In contrast to
LBV and lakes in other parts of Amazonia (Ledru et al.,
1998), the date of 17,820 F 140 14C yr B.P. indicates
deposition during the last glacial maximum at LCH.
At both sites, the reliability of the chronology, and
therefore sedimentation rates, of the clays and sands older
Table 1
AMS radiocarbon dates for Laguna Chaplin
Laboratory
code
Sample depth
(cm)
AMS 14C yr B.P.
(F 1 j)Calibrated yr B.P.
(1 j)
Beta-137570 36.5–37.5 710 F 50 670 (660) 570
AA39700 51.5–52.5 2,240 F 40 2,320 (2,300, 2,240,
2,170) 2,150
AA39701 69.5–70.5 2,740 F 40 2,850 (2,790) 2,770
AA39702 85–86 3,870 F 50 4,350 (4,250) 4,160
AA39703 100–101 4,330 F 80 4,960 (4,850) 4,830
AA39704 125–126 6,040 F 50 6,900 (6,860, 6,830,
6,800) 6,750
AA39705 135–136 9,000 F 100 10,230 (10,180)
9,930
AA39706 155–156 17,820 F 140 21,470 (21,210)
20,960
AA39707 175–176 31,060 F 440 —
AA39708 195–196 34,820 F 700 —
AA39709 213–214 37,750 F 970 —
AA39710 250–251 43,400 F 1900 —
AA39711 285–286 41,200 F 1400 —
AA39712 296–297 38,100 F 1000 —
All samples are carbonate-free bulk sediment. Radiocarbon ages less than
21,000 14C yr B.P. have been calibrated into years before present (cal yr
B.P.) (Stuiver et al., 1998). The radiocarbon dates have been calibrated by a
simple intercept with a linear interpolation of the calibration data points.
The 1 j cal age ranges are shown, with the cal age intercepts in parentheses.
All ages have been rounded to the nearest 10 years.
than f38,000 14C yr B.P. is more difficult to ascertain.
Since the 14C dates all overlap at 2 standard deviations (2 j),these sediments might be the same age, having been depos-
ited extremely rapidly, perhaps as a series of discrete facies.
However, the marked pollen changes through these sedi-
ments, many of which do not coincide with lithological
boundaries, argue against such an interpretation. More likely
these dates overlap because they are close to, or at, the
effective limit of radiocarbon dating. Some of these dates
appear anomalously young. Although there was no indica-
tion of contamination, during either field or laboratory
sampling, such old samples would be highly susceptible to
even the slightest introduction of younger carbon. Perhaps
this was the case for the two lowermost dates of 41,200 F1400 and 38,100 F 1000 14C yr B.P. at LCH (Fig. 3A),
because they are seemingly the only outliers on the age–
depth curve. We therefore excluded these two ages for
estimating the sedimentation rate between 176 and 251 cm
(31,060–43,400 14C yr B.P.). We extrapolated the rate of
0.06 mm yr�1, giving age estimates of 48,000 14C yr B.P. for
the LCH-1/LCH-2 boundary and 51,000 14C yr B.P. for the
base of LCH-1. All other dates from LCH are in chronolog-
ical order and conform to a consistent age-depth relationship,
strengthening our argument that the entire sedimentary
sequence was deposited continuously, albeit at varying
sedimentation rates, over the past 50,000 yr. Ages for all
pollen zone boundaries at LCH (notwithstanding the base of
LCH-1 and LCH-1/LCH-2) were therefore calculated by
linear interpolation between consecutive radiocarbon dates.
Dates for the LBV Holocene pollen zone boundaries were
calculated similarly, although the absence of any age–depth
relationship for the older dates of this site precludes calcu-
lation of the chronology of Pleistocene pollen changes.

Fig. 4. Laguna Chaplin summary pollen percentage diagram, showing the most common taxa from the full complement of 290 pollen types. Dots on the curves denote <0.5%. Curves showing�10 exaggeration are
depicted for selected taxawhich have low percentages. TLP sums are > 300 grains per sample, except for the four pollen spectra in zone LCH-1which had lower TLP sums of 100–160, due to low pollen concentrations.
R.E.Burbrid
geet
al./Quatern
ary
Resea
rch61(2004)215–230
220

R.E. Burbridge et al. / Quaternary Research 61 (2004) 215–230 221
In both records, the Pleistocene sediments are predom-
inantly inorganic clays and sands, whereas the Holocene
sediments consist of organic muds (see Figs. 4 and 7),
indicating fundamentally different depositional environ-
ments in these two periods. Since the lakes currently are
located in a seasonally flooded landscape, connected to
neighboring rivers by small streams (Fig. 1), the Pleistocene
clastic sediments in these basins possibly are fluvial,
originating from floodwaters of the neighboring rivers in
the rainy season. However, we think this is unlikely because
the Rıo Paragua and Rıo Itenez are not white-water rivers,
laden with clastic material eroded from the Andes. Instead,
they are sediment-free, black-water rivers with headwaters
to the southeast on the Brazilian Shield. By contrast, the
lakes in the seasonally flooded forests and savannas of the
Beni, west of the Brazilian Shield, are entirely filled with
clay sediments deposited from flood waters of the white-
water rivers which drain the Andes. We suggest that the
Pleistocene sediments at our study sites (especially the
quartzite sands) were deposited by erosion from the adja-
cent quartzite ridges. The lack of organics in these sedi-
ments perhaps is due to rapid oxidation associated with
lowered water levels caused by a longer dry season.
Furthermore, the square-like morphometry of these basins,
and the substantial distance (15 km) of LCH from the Rıo
Paragua (Fig. 1), clearly shows they are not ox-bow lakes.
Instead, they are most likely pseudo-karst basins formed by
solution of silica from the underlying rocks of the Pre-
Cambrian Shield.
Vegetation and climate history of Noel Kempff Mercado
National Park
The last glaciation (f51,000–10,000 14C yr B.P.)
Our discussion refers predominantly to the LCH record,
since much of the glacial sequence is missing from LBV.
Pollen zone LCH-1 (51,000–48,000 14C yr B.P.) is indic-
ative of a catchment dominated by seasonally flooded
‘‘palm marsh’’ comprising almost exclusively two closely
related species, Mauritia flexuosa and Mauritiella armata
(Fig. 4). Concentration and PAR data (Fig. 5) reveal that the
exceedingly high pollen percentages of these palm taxa
(over 80% TLP) are primarily due to the negligible abun-
dance of other pollen types in this zone. These palms, which
form large, mixed colonies in such habitats today, typically
experience seasonal floods up to 2 m deep (Killeen, 1998).
The trace abundances of charcoal (Fig. 6) indicate that this
palm-dominated vegetation was not sufficiently dry to burn
frequently.
In zones LCH-2–LCH-5 (48,000–10,760 14C yr B.P.),
the dominance of Poaceae (>40%) and Cyperaceae pollen
(f15%) and the continuous appearance of Mauritia/Maur-
itiella pollen, especially between 37,750 F 970 and 17,820
F 140 14C yr B.P (zones LCH-3 and LCH-4), indicate the
presence of local low-lying areas covered by seasonally
flooded termite savannas and savanna marsh. This vegeta-
tion most likely occupied those areas that are currently
covered by seasonally inundated evergreen forest. The
consistently low values of Moraceae pollen (<10%) indicate
that humid evergreen forest communities were scarce
regionally, possibly existing only as gallery forests along
levees bordering the Rıo Paragua and in suitable micro-
habitats around the lake shore.
Consistent presence of Paullinia/Roupala, Celtis,
Machaerium/Dalbergia/Byrsonima, and Erythroxylum pol-
len throughout the Pleistocene record suggests that local
upland areas, such as the neighboring quartzite ridge, may
have supported stands of dry forest and/or mixed liana
forest. However, the pollen of Anadenanthera, a key dry
forest indicator (Prado and Gibbs, 1993; Mayle et al., 2004),
is largely absent from glacial sediments of both sites. The
composition of these forest communities probably differed
from those of the contemporary deciduous and semidecid-
uous dry forests.
Clear floristic changes in the community composition of
these savanna and dry forest habitats occurred through the
Pleistocene. The vegetation changes also are accompanied
by variations in charcoal concentrations and inferred fire
dynamics. For example, Machaerium/Dalbergia/Byrsonima
and Erythroxylum pollen are important components of zone
LCH-2 and are associated with peaks in charcoal concen-
trations, suggesting that these savanna and dry forest eco-
systems may have exhibited the highest fire frequencies of
the entire record. Peaks in macroscopic charcoal (e.g., >500
Am) likely represent local fires, whereas microscopic char-
coal noted on pollen slides reflects regional fires. In con-
trast, the peak in Mauritia/Mauritiella pollen and minimal
charcoal concentrations in zones LCH-3 and LCH-4 suggest
a greater coverage of seasonally flooded savanna and
savanna marsh that contained a relatively smaller arboreal
component and hence less fuel for fires. However, interpre-
tation of the charcoal record is complicated by the likeli-
hood that differences in charcoal concentrations could at
least partly be attributable to taphonomic changes associated
with the switch from relatively higher energy clay/sand
facies to lower energy clay facies. The abundance of Isoetes
spores throughout the Pleistocene sediments (40–80% of
the total pollen sum) suggests that water depth was lower
than today, allowing populations of the emergent semiaquat-
ic fern to expand throughout much of the lake. This
vegetation reconstruction is generally corroborated by pol-
len concentrations and PARs. However, PARs of all taxa are
unexpectedly low in zones LCH-4 and LCH-5 (f29,950–
10,760 14C yr B.P.). Perhaps pollen grains oxidized under
low sedimentation rates (f0.02 mm yr�1) and/or within
environments in which sedimentation was at least annually
discontinuous.
These terrestrial and aquatic ecosystem changes indicate
vegetation responses to increased aridity and lowered atmo-
spheric CO2 concentrations (Monnin et al., 2001). Carbon-
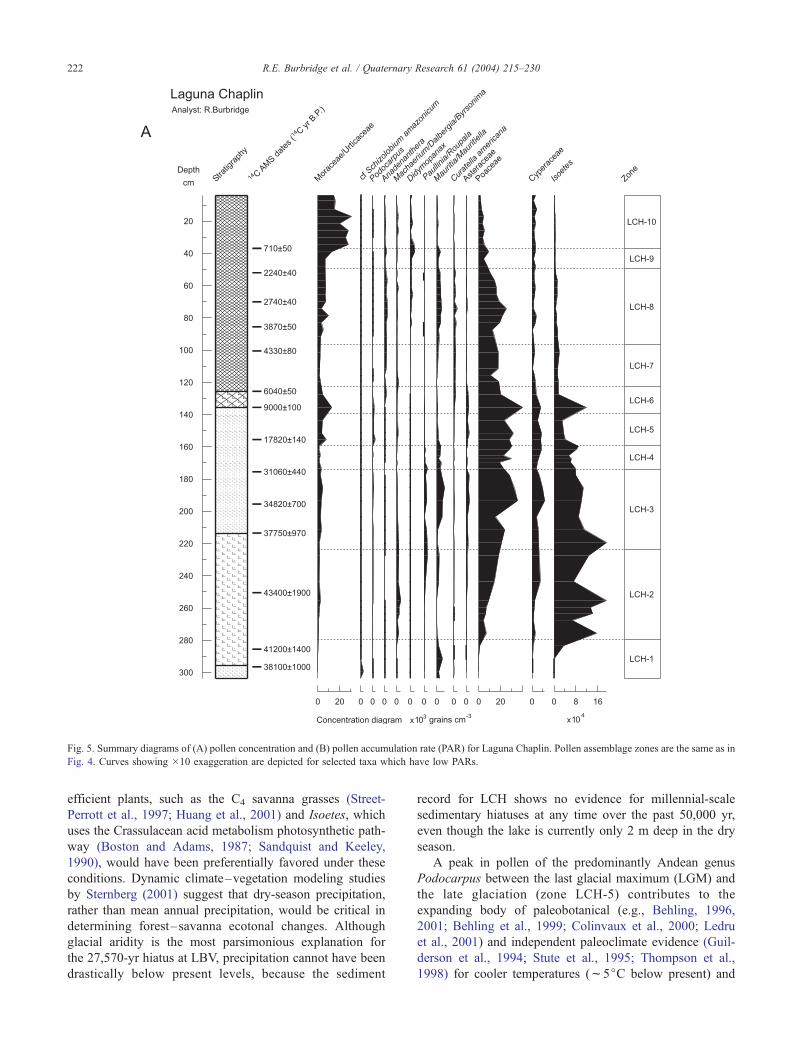
Fig. 5. Summary diagrams of (A) pollen concentration and (B) pollen accumulation rate (PAR) for Laguna Chaplin. Pollen assemblage zones are the same as in
Fig. 4. Curves showing �10 exaggeration are depicted for selected taxa which have low PARs.
R.E. Burbridge et al. / Quaternary Research 61 (2004) 215–230222
efficient plants, such as the C4 savanna grasses (Street-
Perrott et al., 1997; Huang et al., 2001) and Isoetes, which
uses the Crassulacean acid metabolism photosynthetic path-
way (Boston and Adams, 1987; Sandquist and Keeley,
1990), would have been preferentially favored under these
conditions. Dynamic climate–vegetation modeling studies
by Sternberg (2001) suggest that dry-season precipitation,
rather than mean annual precipitation, would be critical in
determining forest–savanna ecotonal changes. Although
glacial aridity is the most parsimonious explanation for
the 27,570-yr hiatus at LBV, precipitation cannot have been
drastically below present levels, because the sediment
record for LCH shows no evidence for millennial-scale
sedimentary hiatuses at any time over the past 50,000 yr,
even though the lake is currently only 2 m deep in the dry
season.
A peak in pollen of the predominantly Andean genus
Podocarpus between the last glacial maximum (LGM) and
the late glaciation (zone LCH-5) contributes to the
expanding body of paleobotanical (e.g., Behling, 1996,
2001; Behling et al., 1999; Colinvaux et al., 2000; Ledru
et al., 2001) and independent paleoclimate evidence (Guil-
derson et al., 1994; Stute et al., 1995; Thompson et al.,
1998) for cooler temperatures (f5jC below present) and

Fig. 5 (continued).
R.E. Burbridge et al. / Quaternary Research 61 (2004) 215–230 223
nonanalog plant communities at this time. Cooling would
reduce evapotranspiration rates and therefore offset, to
some degree, the increased water stress expected by a
reduction in precipitation and/or atmospheric CO2 during
the LGM.
Given that the Pleistocene assemblages are dominated
by pollen types that are poorly dispersed and/or produced
in low quantities relative to wind-pollinated taxa, such as
Moraceae, vegetation interpretations on a regional scale
(i.e., beyond f2 km from the lake shore) are speculative.
In nonflooded areas, however, increased water stress,
caused by a combination of reduced precipitation and
lowered CO2 concentrations, would likely result in in-
creased populations of dry forest species, lianas, and
cerrado (savanna woodland). Certainly, regional differen-
ces existed in the vegetation of NKMNP ca. 40,000–
44,000 14C yr B.P. For example, Alchornea was abundant
around LBV (20–50% pollen, LBV-2) (Fig. 7) but was
largely absent from LCH. Unfortunately, the paleoecolog-
ical significance of this genus for Amazonian Bolivia is
unclear, because it grows in humid evergreen forest,
gallery forest, and seasonally flooded savannas (Killeen
et al., 1993; Killeen, 1998). However, coincident peaks in
macroscopic charcoal (Fig. 9) and Alchornea pollen in
zone LBV-2 suggest a frequently burnt landscape (e.g., a
combination of gallery forests and savanna).

Fig. 6. Microscopic and macroscopic charcoal concentrations for Laguna Chaplin. Pollen assemblage zones are the same as in Fig. 4.
R.E. Burbridge et al. / Quaternary Research 61 (2004) 215–230224
The Holocene (10,000 14C yr B.P.–present)
For the Holocene zones (LCH-6–LCH-8), continued
high percentages of Poaceae pollen (f40%), low percen-
tages of Moraceae pollen (<20%), and similar trends in
pollen concentration and PAR (Figs. 4 and 5) show that
seasonally flooded savannas continued to dominate low-
lying areas around LCH untilf2000 14C yr B.P. Significant
amounts of pollen of Mauritia flexuosa and/or Mauritiella
armata palms and the obligate savanna species, Curatella
americana, which commonly grow together on termite
mounds characteristic of such habitats (Killeen, 1998),
provide further support for this interpretation. Semidecidu-
ous dry forest bordered the lake (most likely in upland,
nonflooded areas such as the quartzite ridge) as indicated by
the presence of Anadenanthera, Astronium fraxinifolium,
Astronium urundeuva, and Gallesia pollen, which are abun-
dant components of the modern Chiquitano Dry Forest,
located 30 km south of LCH (Killeen, 1998; Killeen et al.,
1998). Additional evidence for the establishment of these
dry forest and savanna communities comes from macrofos-
sil charcoal (Fig. 6), which suggests the presence of fire-
prone vegetation. Temporal differences in peak abundances
of these taxa through the Holocene attest to the dynamic

Fig. 7. Laguna Bella Vista summary pollen percentage diagram, showing the most common taxa from the full complement of 263 pollen types. Dots on the curves denote <0.5%. Curves showing �10
exaggeration are depicted for selected taxa which have low percentages. TLP sums are >300 grains per sample, except for sample numbers 92, 247, 257, 289, and 297, which had lower TLP sums of 150–280, due
to low pollen concentrations.
R.E.Burbrid
geet
al./Quatern
ary
Resea
rch61(2004)215–230
225

R.E. Burbridge et al. / Quaternary Research 61 (2004) 215–230226
nature of these ecosystems (e.g., Alchornea in zone LCH-6,
Tapirira guianensis and Erythroxylum in zones LCH-6 and
LCH-7, Mauritia/Mauritiella and Didymopanax in zone
LCH-8).
Between f2000 and 710 F 50 14C yr B.P. (zone LCH-
9), declining Poaceae and increasing Moraceae pollen (Figs.
4 and 5) indicate a reduction in open savannas and an
expansion of forest, the latter likely a mixture of rain forest
and dry forest species. At 710 F 50 14C yr B.P. (zone LCH-
9/LCH-10 boundary), the sharp increase in Moraceae pollen
percentages to present-day levels of 40% and the decline in
Fig. 8. Summary pollen concentration data for Laguna Bella Vista. Pollen assemb
depicted for selected taxa which have low concentrations.
savanna and dry forest taxa to negligible levels mark the
establishment of the modern rain forest-dominated land-
scape around LCH.
A similar pattern of Holocene vegetation changes is
evident at LBV, although significant differences occur in
the times of change. The rain forest/dry forest expansion
began between f6000 (onset of zone LBV-6) and 3000 14C
yr B.P., although the extremely slow sedimentation rate over
this interval (0.02 mm yr�1) precludes a more precise
estimate of the age of this vegetation change. Percentage
data (Fig. 7) suggest that the modern rain forest communi-
lage zones are the same as in Fig. 7. Curves showing �10 exaggeration are

R.E. Burbridge et al. / Quaternary Research 61 (2004) 215–230 227
ties appeared f1650 F 40 14C yr B.P. (zone LBV-6/LBV-7
boundary), although pollen concentration data (Fig. 8)
indicate a somewhat earlier arrival at f2000 14C yr B.P.
At both LBVand LCH, reduced concentrations and PARs of
all taxa in the uppermost sediments can be attributed to the
high water content that typifies samples near the sediment/
water interface.
Discerning the relative contribution of atmospheric CO2
concentrations, precipitation, and temperature as controls of
glacial-age vegetation is difficult for NKMNP. Nonetheless,
precipitation changes were most certainly the primary factor
behind the observed Holocene vegetation dynamics, be-
cause atmospheric CO2 concentrations and temperature
Fig. 9. Microscopic and macroscopic charcoal concentrations for Lagun
exhibited only minor fluctuations through the Holocene
(Indermuhle et al., 1999) relative to those of the LGM–
Holocene transition (Monnin et al., 2001). The progressive
vegetation succession through the Holocene in NKMNP
from savanna/semideciduous forest to semideciduous/ever-
green forest and finally to evergreen rain forest can be
attributed to a gradual increase in mean annual precipitation
and reduction in the length/severity of the dry season since
the mid-Holocene.
Evidence of dense populations of indigenous peoples
throughout much of the Bolivian Amazon prior to the
arrival of Europeans (e.g., Denevan, 1966) raises the
possibility that human activity may have contributed to
a Bella Vista. Pollen assemblage zones are the same as in Fig. 7.

R.E. Burbridge et al. / Quaternary Research 61 (2004) 215–230228
these vegetation changes. Although there is no conclusive
pollen evidence (e.g., Zea) of paleo-Indian agriculture
around either of these lakes, ceramics have been recovered
from a soil pit in an interfluve f25 km northwest of LCH
(T.J. Killeen, personal communication, 2002) and abundant
ceramics mixed with charcoal (dated to f400 14C yr B.P.)
were recovered from anthosols (terra preta) throughout La
Chonta (Guarayos province) f150 km west of NKMNP
(C. Paz, personal communication, 2002). Furthermore,
well-documented evidence (e.g., Denevan, 1966; Erickson,
1995, 2000; Langstroth, 1999) indicates that paleo-Indians
caused major landscape modification to the seasonally
flooded savannas of the alluvial plains throughout the
Bolivian Beni, which encompass much of the Bolivian
Amazon northwest of NKMNP. Although many forest
islands in the Moxos plains of the Beni are natural remnants
or fragments of gallery forests on ancient river levees
(Langstroth, 1999), other forest islands are the result of
human activity, whereby forests invaded artificial earth
mounds following abandonment by paleo-Indians (e.g.,
Denevan, 1966, Erickson, 1995). Although an anthropo-
genic explanation for the recent expansion of rain forest in
NKMNP may seem plausible, we discount this possibility
for several reasons. First, in contrast to the forest islands in
the Beni savannas, rain forests in our study area are not
restricted to terra firme environments, but occur in season-
ally flooded habitats. Second, no archaeological earth-
works, such as earth mounds or ridges, have been
discovered in savanna or forested environments. Finally,
if rain forest expansion occurred simply as secondary
succession following human population collapse in the
area, rain forests should have dominated the area prior to
human arrival. Our pollen data clearly discount this hy-
pothesis. Although ceramic evidence shows that paleo-
Indians lived in NKMNP prior to the Spanish Conquest,
possibly around the shores of the two sites, they appear not
to have had any significant impact upon the ecosystems of
our study area. We therefore infer that shifts in precipitation
were the predominant drivers of vegetation change through
the Holocene.
Implications for understanding paleoclimatology of
southwestern Amazonia
Stable carbon isotope data from soil samples taken
along a 200-km transect across savanna ‘‘islands’’ sur-
rounded by rain forests between Porto Velho (Rondonia
State) and Humaita (Amazonas State), Brazil (8j43VS,63j58VW–7j38VS, 63j04VW), show no evidence for savan-
na expansion during the LGM (de Freitas et al., 2001).
These results are in contrast to our findings from NKMNP,
even though the transect lies only 450 km north of
NKMNP. Furthermore, lake-level reconstructions from
Lake Titicaca (Baker et al., 2001a) and the Salar de Uyuni
salt flats (Baker et al., 2001b) and ice-core data from
Sajama Mountain (Thompson et al., 1998), all sites whose
modern source of precipitation is primarily from the
Amazon Basin, demonstrate that LGM precipitation on
the Bolivian Altiplano was perhaps 20–75% higher than
today (Hastenrath and Kutzbach, 1985; Blodgett et al.,
1997). The difference between a wet Altiplano and rela-
tively drier Bolivian Amazon lowlands is supported by
general circulation modeling studies (Hostetler and Mix,
1999), which clearly show that, in contrast to the present
day, these two regions received distinctly different moisture
sources during the LGM.
How can these regional precipitation differences in
southwestern Amazonia and the Bolivian Altiplano be
explained? Predictions based on orbital forcing by the
19,000- to 22,000-yr precession cycle of the Milankovitch
Astronomical Theory (Berger and Loutre, 1991) would
maximize LGM precipitation in southwestern Amazonia
due to maximum austral summer insolation at 10jS and
hence maximum latitudinal, seasonal migration of the
ITCZ. However, expansion of the Antarctic ice sheet
and Patagonian glaciers would likely steepen the latitudi-
nal temperature gradient across South America despite the
5jC equatorial cooling. Wet-season convective activity
over southern Amazonia would consequently lessen,
thereby reducing precipitation in southernmost Amazonia
relative to today. Furthermore, reduced sea-surface temper-
atures in the equatorial Atlantic would curtail oceanic
moisture transfer to the Basin. These factors may explain
why there was an ecotonal response in the Bolivian
Amazonian lowlands but no ecotonal shift farther north
at Porto Velho/Humaita. The precipitation contrast be-
tween the Altiplano and lowland Bolivia could perhaps
be explained by the dominant moisture source for the
Altiplano coming from winter anticyclones off the eastern
Pacific (as simulated by Hostetler and Mix, 1999) rather
than from the Amazon summer monsoons associated with
the ITCZ.
In contrast to the marked regional differences during the
LGM, climatic changes through the Holocene are broadly
consistent and synchronous throughout southwestern Ama-
zonia and the Altiplano. The early to mid-Holocene arid
phase at NKMNP correlates with: (1) the expansion of
savanna islands 450 km to the north at the Rondonia/
Amazonas border between 9000 and 3000 14C yr B.P. (de
Freitas et al., 2001); (2) replacement of cloud forest by open,
grass-dominated ecosystems in the eastern Bolivian Andes
(17j50VS, 64j43V08WW) between f11,000 and 4000 14C yr
B.P. (Mourguiart and Ledru, 2003); and (3) a prolonged
lake-level low-stand in Lake Titicaca between 8500 and
3500 cal yr B.P. (7750–3300 14C yr B.P.) (Baker et al.,
2001a). These correlations point to a common climatic
forcing affecting all these sites. The most likely explanation
for this arid episode is that the orbitally driven minimum in
January insolation at 15–10jS (Berger and Loutre, 1991)
would have restricted the southerly penetration of the ITCZ
and deep cell convective activity over the Amazon Basin,

R.E. Burbridge et al. / Quaternary Research 61 (2004) 215–230 229
thus reducing the length of the summer rainy season.
Although the close correlation between the Lake Titicaca
low-stand and the sedimentation minimum between 7980
and 3220 14C yr B.P. at LBV (Fig. 3b) suggests that organic
sedimentation in the lake was tightly coupled to climate, the
poor correlation with the sedimentation trends at LCH raises
doubts about the strength of such a causal relationship,
suggesting that local catchment processes were perhaps the
overriding factor.
The trend of increasing precipitation in our study area,
beginning 6000–3000 14C yr B.P., is similarly part of a
widespread phenomenon throughout southwestern Amazo-
nia and the Altiplano, corroborated by: (1) rising water
levels in Lake Titicaca, beginning f4500 cal yr B.P.
(4000 14C yr B.P.) and reaching modern levels byf2000 cal yr B.P. (2000 14C yr B.P.) (Baker et al.,
2001a); (2) increased snow accumulation on Sajama
Mountain at f3000 14C yr B.P. (Thompson et al.,
1998); (3) expansion of cloud forest in the Bolivian Andes
after 4000 14C yr B.P. (Mourguiart and Ledru, 2003); and
(4) contraction of savanna islands in Rondonia/Amazonas
beginning 3000 14C yr B.P. (de Freitas et al., 2001). This
late Holocene increase in humidity reflects progressive
increases in January insolation at 10–15jS since the early
Holocene minimum (Berger and Loutre, 1991) and conse-
quent greater southerly migration of the ITCZ. The 1300-
yr delay in rain forest expansion at LCH compared with
LBV is likely caused by the time-transgressive nature of
precipitation change through the Holocene and/or species’
dispersal lagging behind climatic change. The Holocene
lake-level record from the Bolivian Andes shows that this
rain forest expansion was a function, not only of a
lengthening of the rainy season, but also an absolute
increase in mean annual precipitation.
Acknowledgments
We thank the following: M. Suarez Riglos and M.
Saldias for use of the herbarium at the Natural History
Museum, Santa Cruz, Bolivia; J. Ratter for use of the
herbarium at the Royal Botanic Garden Edinburgh; P.
Colinvaux for use of the Amazon pollen reference collection
at the Smithsonian Tropical Research Institute, Panama; R.
Guillen, L. Sanchez, M. Siles, J. Surubi, and P. Soliz for
assistance with fieldwork; the Fundacion Amigos de la
Naturaleza and the Bolivian Government for permission to
work in NKMNP; and Mark Bush and an anonymous
reviewer, whose comments greatly improved the original
manuscript. This work was funded by a Royal Society
research grant (F.E.M.), two Royal Society Overseas Study
Visits (F.E.M.), a University of Leicester Research Grant
(F.E.M.), and a University of Leicester Ph.D. Studentship
(R.B.). Funding for radiocarbon dates was provided by the
Natural Environment Research Council (26 dates) and the
University of Leicester (1 date).
References
Baker, P.A., Seltzer, G.O., Fritz, S.C., Dunbar, R.B., Grove, M.J., Tapia,
P.M., Cross, S.L., Rowe, H.D., Broda, J.P., 2001a. The history of South
American tropical precipitation for the past 25,000 years. Science 291,
640–643.
Baker, P.A., Rigsby, C.A., Seltzer, G.O., Fritz, S.C., Lowenstein, T.K.,
Bacher, N.P., Veliz, C., 2001b. Tropical climate changes at millennial
and orbital timescales on the Bolivian Altiplino. Nature 409, 698–701.
Behling, H., 1996. First report on new evidence for the occurrence of
Podocarpus and possible human presence at the mouth of the Amazon
during the Late-glacial. Vegetation History and Archaeobotany 5 (3),
241–246.
Behling, H., 2001. Late Quaternary environmental changes in the Lagoa da
Curuc�a region (eastern Amazonia, Brazil) and evidence of Podocarpus
in the Amazon lowland. Vegetation History and Archaeobotany 10,
175–183.
Behling, H., Berrio, J.C., Hooghiemstra, H., 1999. Late Quaternary pollen
records from the middle Caqueta river basin in central Colombian
Amazon. Palaeogeography, Palaeoclimatology, Palaeoecology 145,
193–213.
Bennett, K.D., 1997. Documentation for Psimpoll2.30 and Pscomb 1.02; C
Programs for Plotting Pollen Diagrams and Analysing Pollen Data.
Univ. of Cambridge, Cambridge, UK.
Berger, A., Loutre, M.F., 1991. Insolation values for the climate of the last
10 million years. Quaternary Science Reviews 10, 297–317.
Blodgett, T., Lenters, J., Isacks, B., 1997. Constraints on the origin of
palaeolake expansions in the central Andes. Earth Interactions 1 (1),
1–28.
Boston, H.L., Adams, M.S., 1987. Productivity, growth and photosynthesis
of two small ‘‘isoetid’’ plants, Littorella uniflora and Isoetes macro-
spora. Journal of Ecology 75, 333–350.
Colinvaux, P., de Oliveira, P.E., Moreno, J.E., 1999. Amazon Pollen Man-
ual and Atlas. Harwood Academic, The Netherlands.
Colinvaux, P.A., de Oliveira, P.E., Bush, M.B., 2000. Amazonian and
neotropical plant communities on glacial time-scales: the failure of
the aridity and refuge hypotheses. Quaternary Science Reviews 19,
141–169.
de Freitas, H.A., Pessenda, L.C.R., Aravena, R., Gouveia, S.E.M., Ribeiro,
A., Boulet, R., 2001. Late Quaternary vegetation dynamic record in
southern Amazon inferred from carbon isotopes in soil organic matter.
Quaternary Research 55, 39–46 (doi:10.1006/qres.2000.2192).
Denevan, W.M., 1966. The aboriginal cultural geography of the Llanos de
Mojos of Bolivia. Ibero-Americana 48, 1–60.
Erickson, C.L., 1995. Archaeological methods for the study of ancient
landscapes of the Llanos de Mojos in the Bolivian Amazon. In: Stahl,
P. (Ed.), Archaeology in the American Tropics: Current Analytical
Methods and Applications. Cambridge Univ. Press, Cambridge,
UK, pp. 66–95.
Erickson, C.L., 2000. An artificial landscape-scale fishery in the Bolivian
Amazon. Nature 408, 190–193.
Faegri, K., Iverson, I., 1989. Textbook of Pollen Analysis. Blackwell
Sci., Oxford.
Gosling, W.D., Mayle, F.E., Killeen, T.J., Siles, M., Sanchez, L.,
Boreham, S., 2003. A simple and effective methodology for sam-
pling modern pollen rain in tropical environments. The Holocene
13 (4), 613–618.
Guilderson, T.P., Fairbanks, R.G., Rubenstone, J.L., 1994. Tropical tem-
perature variations since 20,000 years ago: modulating interhemispheric
climate change. Science 263, 663–665.
Hastenrath, S., Kutzbach, J., 1985. Late Pleistocene climate andwater budget
of the South American Altiplano. Quaternary Research 24, 249–256.
Hostetler, S.W., Mix, A.C., 1999. Reassessment of ice-age cooling of the
tropical ocean and atmosphere. Nature 399, 673–676.
Huang, Y., Street-Perrott, F.A., Metcalfe, S.E., Breener, M., Moreland, M.,
Freeman, K.H., 2001. Climate change as the dominant control on Gla-

R.E. Burbridge et al. / Quaternary Research 61 (2004) 215–230230
cial– Interglacial variations in C3 and C4 plant abundance. Science 293,
1647–1651.
Indermuhle, A., Stocker, T.F., Joos, F., Fischer, H., Smith, H.J., Wahlen,
M., Deck, B., Mastroianni, D., Tschumi, J., Blunier, T., Meyer, R.,
Stauffer, B., 1999. Holocene carbon-cycle dynamics based on CO2
trapped in ice at Taylor Dome, Antarctica. Nature 398, 121–126.
Killeen, T.J., 1998. Vegetation and flora of Noel Kempff Mercado National
Park. In: Killeen, T.J., Schulenberg, T.S. (Eds.), A Biological Assessment
of Parque Nacional Noel Kempff Mercado, Bolivia. RAP Working
Papers, vol. 10. Conservation International,Washington, DC, pp. 61–85.
Killeen, T.J., Schulenberg, T.S., 1998. A Biological Assessment of Parque
Nacional Noel Kempff Mercado, Bolivia. RAPWorking Papers, vol. 10.
Conservation International, Washington, DC.
Killeen, T.J., Garcia, E., Beck, S.G., Eds. (1993). ‘‘Guia de Arboles de
Bolivia.’’ Herbario Nacional de Bolivia, La Paz, Bolivia, and Missouri
Botanical Garden, St. Louis, MO.
Killeen, T.J., Jardim, A., Mamani, F., Rojas, N., 1998. Diversity, composi-
tion and structure of tropical semi deciduous forest in the Chiquitania
region of Santa Cruz, Bolivia. Journal of Tropical Ecology 14, 803–827.
Killeen, T.J., Siles, T.M., Grimwood, T., Tieszen, L.L., Steininger, M.K.,
Tucker, C.J., Panfil, S.N., 2001. Habitat heterogeneity on a forest –
savanna ecotone in Noel Kempff Mercado National Park (Santa Cruz,
Bolivia): implications for the long-term conservation of biodiversity in a
changing climate. In: Bradshaw, G.A., Marquet, P.A. (Eds.), How
Landscapes Change: Human Disturbance and Ecosystem Disruptions
in the Americas. Ecological Studies, vol. 162. Springer-Verlag, Berlin,
pp. 285–312.
Langstroth, R.P., 1999. Forest islands in an Amazonian savanna of north-
eastern Bolivia. Ph.D. dissertation, Univ. of Wisconsin, Madison.
Ledru, M.-P., Bertaux, J., Sifeddine, A., and Suguio, K. (1998). Absence of
last glacial maximum records in lowland tropical forests. Quaternary
Research 49, 233–237.
Ledru, M., Cordeiro, R.C., Dominguez, J.M.L., Martin, L., Mourguiart, P.,
Sifeddine, A., and Turcq, B. (2001). Late glacial cooling in Amazonia
inferred from pollen at Lagoa do Caco, northern Brazil. Quaternary
Research 55, 47–56, doi:10.1006.qres.2000.2187.
Litherland, M., Power, G., 1989. The geological and geomorphic evolution
of Serrania Huanchaca (Eastern Bolivia): The Lost World. Journal of
South American Earth Sciences 2, 1–17.
Mayle, F.E., Burbridge, R., Killeen, T., 2000. Millennial-scale dynamics of
southern Amazonian rain forests. Science 290, 2291–2294.
Mayle, F.E., Beerling, D.J., Gosling, W.D., Bush, M.B., 2004. Responses
of Amazonian ecosystems to climatic and atmospheric CO2 changes
since the last glacial maximum. Philosophical Transactions of the Royal
Society. London, Series B (in press).
Monnin, E., Indermuhle, A., Dallenbach, A., Fluckiger, J., Stauffer, B.,
Stocker, T.F., Raynaud, D., Barnola, J., 2001. Atmospheric CO2 con-
centrations over the last glacial termination. Science 291, 112–114.
Mourguiart, P., Ledru, M.-P., 2003. Last glacial maximum in an Andean
cloud forest environment (Eastern Cordillera, Bolivia). Geology 31 (3),
195–198.
Prado, D.E., Gibbs, P.E., 1993. Patterns of species distributions in the dry
seasonal forests of South America. Annals of the Missouri Botanical
Garden 80 (4), 902–927.
Rhodes, A.N., 1998. A method for the preparation and quantification of
microscopic charcoal from terrestrial and lacustrine sediment cores. The
Holocene 8, 113–117.
Roubik, D.W., Moreno, J.E.P., 1991. Pollen and Spores of Barro Colorado
Island. Missouri Botanical Garden, St. Louis, MO.
Sandquist, D.R., Keeley, J.E., 1990. Carbon uptake characteristics in two
high elevation populations of the aquatic CAM plant Isoetes bolanderi
(Isoetacae). American Journal of Botany 77, 682–688.
Sternberg, L.S.L., 2001. Savanna–forest hysteresis in the tropics. Global
Ecology and Biogeography 10, 369–378.
Street-Perrott, F.A., Huang, Y., Perrott, R.A., Eglinton, G., Barker, P.,
Khelifa, L.B., Harkness, D.D., Olago, D., 1997. Impact of lower atmo-
spheric carbon dioxide on tropical mountain ecosystems. Science 278,
1422–1426.
Stute, M., Forster, M., Frischkorn, H., Serejo, A., Clark, J.F., Schlosser, P.,
Broecker, W.S., Bonani, G., 1995. Cooling of tropical Brazil (5jC)during the last glacial maximum. Science 269, 379–383.
Stuiver, M., Reimer, P.J., Bard, E., Beck, J.W., Burr, G.S., Hughen, K.A.,
Kromer, B., McCormac, G., Van der Plicht, J., Spurk, M., 1998.
INTCAL98 Radiocarbon Age Calibration 24,000–0 cal BP. Radiocar-
bon 40 (3), 1041–1083.
Thompson, L.G., Davis, M.E., Mosley-Thompson, E., Sowers, T.A.,
Henderson, K.A., Zagorodnov, V.S., Lin, P.-N., Mikhalenko, V.N.,
Campen, R.K., Bolzan, J.F., Cole-Dai, J.A., 1998. 25,000 year
tropical climate history from Bolivian ice cores. Science 282,
1858–1864.
Wright, H.E., 1967. A square-rod piston sampler for lake sediments. Jour-
nal of Sedimentary Petrology 27, 956–957.