expression of stem cell markers in human astrocytomas of ... · expression of stem cell markers in...
TRANSCRIPT

LAB INVESTIGATION-HUMAN/ANIMAL TISSUE
Expression of stem cell markers in human astrocytomasof different WHO grades
Yue-Hui Ma Æ Rolf Mentlein Æ Friederike Knerlich ÆMarie-Luise Kruse Æ H. Maximilian Mehdorn ÆJanka Held-Feindt
Received: 11 May 2007 / Accepted: 11 June 2007
� Springer Science+Business Media B.V. 2007
Abstract According to new hypotheses astrocytomas/
gliomas either arise from or attract neural stem cells.
Biological markers, particularly antigenic markers, have
played a significant role for the characterization of these
tumour stem cells (TSCc). Because these studies have been
performed with single experimental samples mostly from
gliomas, we investigated the expression of the stem cell
markers CD133/Prominin, Nestin, Sox-2, Musashi-1,
CXCR4, Flt-4/VEGFR-3 and CD105/Endoglin in 72
astrocytomas of different WHO-grades and compared it to
normal adult human brain. Expression of their mRNA was
quantified by quantitative RT-PCR, of their protein by
counting immunopositive cells. In contrast to normal brain,
tumour samples showed a high variability for the expres-
sion of all markers. However, their mean expression was
significantly increased in astrocytomas, but this depended
on the WHO grade only for CD133, Nestin, Sox-2 and
Musashi-1. Confocal microscopy revealed that these
markers mostly could be co-stained with glial fibrillary
acidic protein, a marker for astoglial cells, but less fre-
quently with the proliferation marker Ki-67/MIB-1. These
markers sometimes, but not necessarily could be co-stained
with each other in complex patterns. Our results show that
most astrocytomas contain considerable portions of cells
expressing stem cell markers. It appears that some of these
cells originate from tumour genesis (supporting the stem
cell hypothesis) while others are attracted by the tumours.
Further functional markers are required to differentiate
these TSC-types.
Keywords Astrocytomas � CD133 � CXCR4 � Endoglin �Flt-4 � Musashi-1 � Nestin � SOX-2 � Stem cell markers �Tumour stem cells
Abbreviations
NSC Neural stem cell
TSC Tumour stem cell
HSC Hematopoietic stem cell
NBT Normal brain tissue
IHC Immunohistochemistry
RE Real-time reverse-transcription polymerase
chain reaction
RT-PCR Reverse-transcription polymerase chain
reaction
GFAP Glial fibrillary acidic protein
WHO World Health Organization
Introduction
Astrocytomas are the most common types of primary brain
tumours, accounting for 33.3% of all newly diagnosed
brain tumours [1]. These neuroepthelial tumours can occur
Y.-H. Ma � F. Knerlich � H. M. Mehdorn � J. Held-Feindt (&)
Department of Neurosurgery, University Medical Center
Schleswig-Holstein, Campus Kiel, Schittenhelmstr. 10, 24105
Kiel, Germany
e-mail: [email protected]
Y.-H. Ma
Department of Neurosurgery, the First Affiliated Hospital,
School of Medicine, Zhejiang University, Hangzhou, People’s
Republic of China
R. Mentlein
Department of Anatomy, University of Kiel, Kiel, Germany
M.-L. Kruse
Department of General Internal Medicine, University Medical
Center Schleswig-Holstein, Campus Kiel, Kiel, Germany
123
J Neurooncol
DOI 10.1007/s11060-007-9439-7

at any age, especially at young and middle ages, and are
more common in men. Based on specific histopathological
characteristics such as cellularity, nuclear atypia, mitotic
activity, microvascular proliferation and necrosis, astrocy-
tomas are subdivided into four grades of malignancy
[World Health Organization (WHO) grades I–IV] in order
of increasing malignancy [2]. By this, astrocytomas of
WHO grade IV = glioblastomas are the most malignant
forms.
It has traditionally been thought that astrocytomas
originate from astrocytes or their precursor cells. The
genesis of astrocytomas involves a number of character-
istic genetic alterations including activation of oncogenes
and inactivation of tumour suppressor genes [3, 4], which
lead to a progressive disorder of normal cellular growth,
apoptotic, cell motility and invasion pathways. However,
with discovery of neural stem cells (NSCs) in the central
nervous system and advances on molecular biology of
NSCs since early 1990s, there has been a wide speculation
that normal NSCs or progenitor cells might be principal
targets of mutations that lead to a brain tumour, probably
through dysregulation of self-renewal pathways. Tumour
cells with NSC properties are called tumour stem cells
(TSCs) [5–8].
The TSC hypothesis dictates that tumours arise from a
single, self-renewing cell type, which then gives rise to the
rest of the tumour cells, including a variety of ‘‘more dif-
ferentiated’’ cell types. It has been found that astrocytoma
cells share many characteristics with NSCs such as the
capacity of self-renewal and multi-lineage differentiation
[9–11], and similarities in phenotype and signalling [11,
12]. Imaging studies demonstrate that astrocytomas origi-
nate in two areas, the subventricular zone of the lateral
ventricle and the hippocampus, which have been previ-
ously shown to be reservoirs of normal NSCs [13]. Besides,
subependymal giant cell astrocytomas coexpressed several
neuronal markers (neuronal nuclear antigen, neuron spe-
cific enolase) and markers of NSCs (nestin) and glial cells
(glial fibrillary acidic protein) [14]. These studies suggest
that astrocytomas may result from transformation of NSCs.
The direct evidence supporting this notion comes from
transgenic mice models, which demonstrate that nestin-
expressing progenitors, excessively expressing oncogenes
such as c-Myc, can give rise to gliomas and medulloblas-
tomas [15–17].
Biological markers, particularly antigenic markers,
have played a significant role for the characterization of
various stem cells. Reported markers for detecting TSC
populations are Nestin (glioblastomas) [8], CD133 (glio-
blastomas; medulloblastomas, ependymomas and prostate
cancer) [5, 8, 18, 19], Musashi-1 (glioblastomas) [6],
SOX-2 (glioblastomas) [6], Bmi-1 (glioblastomas) [6],
CD44 (breast cancer; prostate cancer) [18, 20], CD20
(melanomas) [21], CD34 (leukaemia) [22], ABCG2 (ret-
inoblastoma, neuroblastoma) [23, 24] and stem cell anti-
gen-1 (Sca-1) (lung cancer) [25]. Moreover, it could be
shown that CXCR4 is expressed by NSCs [26], Flt-4/
VEGR-3 on neural progenitor cells [27], and CD105/
Endoglin, a specific marker for hematopoetic stem cells,
was found in neoplastic cells of sarcomas and ovary
carcinomas [28].
Tumour stem cells can be isolated and characterized in
many solid tumours, such as brain tumours [5–8, 19, 29–
31], breast cancers [20], melanomas [21], prostate cancers
[18] and lung cancers [25]. Nevertheless, TSCs were
primarily only identified in glioblastomas. However, most
studies were done with single samples and nothing is
known about the expression of these putative TCS
markers in a broader variety of gliomas and especially in
astrocytomas of lower WHO grades. Therefore, we anal-
ysed for the first time the expression of seven different
stem cell markers (CD133, Nestin, SOX-2, Musashi-1,
CXCR4, Flt-4 and Endoglin) in astrocytomas of all WHO
grades with different quantitative methods on the mRNA
and protein level. Moreover, we investigated the co-
expression of theses genes with markers for glial and
proliferating cells, and among themselves, and correlated
the results with the different malignant grades of the used
tumours.
Materials and methods
Patients and tumour specimens
Astrocytoma tissues of different WHO grades were col-
lected from 72 patients who were operated on between
January 1998 and August 2006 at the Department of
Neurosurgery, Kiel, Germany (see Table 1). Four normal
brain tissue (NBT) samples served as controls. The
patients included 28 women and 44 men. The mean age at
diagnosis was 37.6 ± 23.2 and 39.9 ± 21.5 years, respec-
tively. All samples were obtained in accordance with
approved ethical standards of the responsible committee
of the University of Kiel and with the Helsinki Declara-
tion of 1975, as revised in 1983. Tumours were classified
according to the WHO criteria into the various subtypes
of astrocytomas. Eighteen tumours corresponded to
astrocytomas WHO grade I, 12 tumours were astrocyto-
mas WHO grade II, 17 were astrocytomas WHO grade
III, and 25 tumours corresponded to astrocytomas WHO
grade IV (glioblastomas). If possible (enough material
available), for different experiments matched probes of
individual tumour samples were used (for details see
Table 1).
J Neurooncol
123

Real-time RT-PCR
For real-time RT-PCR solid astrocytoma tissues of different
WHO grades (13 astrocytoma I samples, 12 astrocytoma II
samples, 11 astrocytoma III samples and 18 astrocytoma IV
samples), and four NBT samples were chosen (see Table 1).
Total RNAs from different samples were purified with
TRIzol Reagent (Invitrogen, Life Technologies, Karlsruhe,
Germany), treated with RNase free DNase (1 U/ll, Pro-
mega, Madison, WI, USA), and reverse transcribed by
RevertAidTM H Minus M-MuLV Reverse Transcriptase
(200 U/ll, Fermentas, Vilinius, Lithuania) according to the
manufacturer’s instruction.
Real-time RT-PCR was performed in three replicates of
each sample using a total reactive volume of 20 ll, which
contained 1 ll of 20 · Assays-on-DemandTM Gene
Expression Assay Mix (Hs00195682_m1 for CD133,
Hs00707120_s1 for Nestin, Hs00602736_s1 for SOX-2,
Hs00159291_m1 for Musashi-1, Hs00237052_m1 for
CXCR4, Hs00176607_m1 for Flt-4, Hs00164438_m1 for
Table 1 Tumour samples
Sample
number
Gender Age Histology Detection
1 Male 23 Astrocytoma I RE, IHC
2 Female 14 Astrocytoma I RE
3 Male 42 Astrocytoma I RE
4 Female 28 Astrocytoma I RE, IHC
5 Male 6 Astrocytoma I RE, IHC
6 Female 7 Astrocytoma I RE
7 Male 13 Astrocytoma I RE
8 Male 17 Astrocytoma I RE
9 Female 12 Astrocytoma I RE, IHC
10 Female 6 Astrocytoma I RE, IHC
11 Male 17 Astrocytoma I RE, IHC
12 Male 4 Astrocytoma I RE
13 Male 5 Astrocytoma I RE
14 Female 11 Astrocytoma I IHC
15 Female 15 Astrocytoma I IHC
16 Female 19 Astrocytoma I IHC
17 Male 48 Astrocytoma I IHC
18 Female 11 Astrocytoma I IHC
19 Male 34 Astrocytoma II RE, IHC
20 Male 26 Astrocytoma II RE, IHC
21 Female 40 Astrocytoma II RE, IHC
22 Male 29 Astrocytoma II RE, IHC
23 Female 43 Astrocytoma II RE, IHC
24 Male 21 Astrocytoma II RE, IHC
25 Female 25 Astrocytoma II RE
26 Male 58 Astrocytoma II RE
27 Female 42 Astrocytoma II RE
28 Male 21 Astrocytoma II RE, IHC
29 Male 8 Astrocytoma II RE, IHC
30 Female 18 Astrocytoma II RE, IHC
31 Female 41 Astrocytoma III RE, IHC
32 Female 65 Astrocytoma III RE, IHC
33 Male 36 Astrocytoma III RE
34 Male 12 Astrocytoma III RE
35 Male 32 Astrocytoma III RE
36 Female 37 Astrocytoma III RE, IHC
37 Male 44 Astrocytoma III RE
38 Male 57 Astrocytoma III RE, IHC
39 Female 51 Astrocytoma III RE
40 Male 7 Astrocytoma III RE
41 Male 32 Astrocytoma III RE, IHC
42 Female 31 Astrocytoma III IHC
43 Female 59 Astrocytoma III IHC
44 Male 43 Astrocytoma III IHC
45 Male 67 Astrocytoma III IHC
46 Male 45 Astrocytoma III IHC
47 Male 50 Astrocytoma III IHC
Table 1 continued
Sample
number
Gender Age Histology Detection
48 Female 63 Astrocytoma IV RE, IHC
49 Male 60 Astrocytoma IV RE
50 Male 71 Astrocytoma IV RE, IHC, Confocal
51 Male 49 Astrocytoma IV RE, IHC
52 Male 69 Astrocytoma IV RE, IHC
53 Male 57 Astrocytoma IV RE
54 Female 78 Astrocytoma IV RE
55 Male 61 Astrocytoma IV RE
56 Female 80 Astrocytoma IV RE
57 Male 45 Astrocytoma IV RE
58 Female 71 Astrocytoma IV RE
59 Male 28 Astrocytoma IV RE
60 Male 68 Astrocytoma IV RE
61 Male 49 Astrocytoma IV RE
62 Female 72 Astrocytoma IV RE
63 Male 34 Astrocytoma IV RE
64 Male 28 Astrocytoma IV RE
65 Male 61 Astrocytoma IV RE
66 Male 65 Astrocytoma IV IHC, Confocal
67 Male 73 Astrocytoma IV IHC
68 Male 75 Astrocytoma IV IHC
69 Female 25 Astrocytoma IV IHC, Confocal
70 Female 57 Astrocytoma IV IHC
71 Female 31 Astrocytoma IV IHC, Confocal
72 Male 68 Astrocytoma IV Confocal
RE real-time reverse transcription polymerase chain reaction, IHCimmunohistochemistry
J Neurooncol
123

Endoglin; Applied Biosystems, Foster City, CA, USA),
10 ll of 2 · TaqMan Universal PCR Master Mix and 100
or 10 ng of cDNA template (diluted in RNase-free water to
9 ll). After 2 min at 50�C and 10 min at 95�C, 40 cycles of
15 s at 95�C and 1 min at 60�C were run. Glyceraldehyde-
3-phosphate dehydrogenase (GAPDH; Hs99999905_m1,
Applied Biosystems) in each sample was tested as intrinsic
positive control. Each plate included at least three ‘‘No
Template Controls (NTC)’’. The reaction was carried out
with the MyiQTM Single Color Real-time PCR Detection
System (Bio-Rad, Munchen, Germany) and fluorescent
data were converted into cycle threshold (CT) measure-
ments. DCT-values of each sample were calculated as
CTgene of interest–CT GAPDH.
Immunohistochemistry
For immunohistochemistry (IHC) examination, fresh-fro-
zen astrocytoma tissues of different WHO grades (11
astrocytoma I samples, 9 astrocytoma II samples, 11
astrocytoma III samples and 10 astrocytoma IV samples)
and two NBT samples were cut in a cryostat into 6 lm
sections (see Table 1). The sections were air dried and
stored in –20�C freezer until used. IHC was performed
using the avidin-biotin-peroxidase complex (ABC) meth-
od. The sections were fixed with 4% para-formaldehyde in
phosphate buffered saline (PBS) for 30 min at room tem-
perature (RT). To block endogenous peroxidase and non-
specific binding, 3% H2O2 in 0.3% Triton·-100/PBS
(30 min at RT) and appropriate 10% normal blocking
serum in PBS (60 min at RT; normal horse serum for
Nestin and Musashi-1, normal rabbit serum for CD133, and
normal donkey serum for SOX-2, CXCR4, Flt-4 and En-
doglin; Jackson Immuno Research Laboratories, Mulkiteo,
WA, USA) were used, respectively. Sections were incu-
bated with appropriate dilutions of primary antibodies over
night at 4�C (anti-CD133, goat polyclonal, 1:100, Santa
Cruz, CA, USA; anti-Nestin, mouse monoclonal, 1:200,
R&D Systems, Wiesbaden, Germany; anti-Musashi-1,
mouse monoclonal, 1:200, R&D Systems; anti-SOX-2,
rabbit polyclonal, 1:100, Santa Cruz; anti-CXCR4, rabbit
polyclonal, 1:100, IMGENEX, San Diego, CA, USA; anti-
Flt-4, rabbit polyclonal, 1:200, Santa Cruz; anti-Endoglin,
rabbit polyclonal, 1:50; Santa Cruz). The antibodies were
diluted in 0.3% Triton·-100 and appropriate 2% normal
blocking serum in PBS. Primary antibodies were omitted
for negative controls. After washing steps with PBS
(10 min for two times), the sections were incubated with
corresponding second antibodies diluted 1:200 in 1.5%
blocking serum/PBS for 60 min at RT (biotinylated horse
anti-mouse IgG for Nestin and Musashi-1, Vector Labo-
ratories, Burlingame, CA, USA; biotinylated rabbit anti-
goat IgG for CD133, Vector Laboratories; biotinylated
donkey anti-rabbit for SOX-2, CXCR4, Flt-4 and Endoglin,
Jackson Immuno Research). Amplification of the signal
was carried out by ABC method with ABC Vectastain� kit
(Vector Laboratories, Burlingame, CA, USA). The signal
was visualized by incubation with 0.06% 3,3¢-diam-
inobenzidine-tetrahydrochloride (Sigma, Munchen, Ger-
many) and 0.003% H2O2 in 0.1 M Tris–HCl (pH 7.6) for
40–55 s. Finally, the sections were counterstained with
hematoxylin for 40 s, mounted with Eukitt� quick-harden
mounting medium (Fluka, Munchen, Germany) and cov-
erslipped for investigation. Brightfield microscopy with
digital photography was performed using a Zeiss micro-
scope (Zeiss, Oberkochen, Germany).
Confocal microscopy
Cryostat sections of five different astrocytoma tissues
WHO grade IV (see Table 1) were fixed in acetone/meth-
anol (1:1; 10 min) at -20�C, washed with Tris-buffered
saline plus 0.1% Tween 20 (TBS-T, 3·, RT), washed with
20%, then 70% ethanol (each 2 min), blocked with Sudan
black (1% in 70% ethanol) for 10 min, rinsed with 70%
ethanol until dye free, then for 2 min with 20% ethanol,
washed with TBS-T (3·), blocked with 0.1% bovine serum
albumin and 0.2% glycine in TBS (1 h), then without
washing incubated in with primary antibodies in TBS-T at
4�C overnight. Primary antibodies were omitted for nega-
tive controls. After a washing step (3 · TBS-T, 10 min)
the first secondary antibody was incubated for 1 h at 37�C
in darkness. The sections were washed with TBS-T
(3 · 10 min). When both primary antibodies were obtained
from mouse, an additional blocking step with goat anti-
mouse FAB-fragments (1:1,000 1 h, RT, from Dianova,
Hamburg, Germany) was necessary. The second primary
antibody was applied after another washing step (3 · TBS-
T) and incubated over night at 4�C. Second primary anti-
bodies were omitted for negative controls. The slides were
washed again (3 · TBS-T) and incubated with the second
secondary antibody for 1 h at 37�C. After washing with
TBS-T (1 · 10 min), TBS (2 · 10 min), nuclei were
stained with diamidino-2-phenylindole (DAPI; Molecular
Probes/Invitrogen, Life Technologies, Karlsruhe, Ger-
many; 1:30,000, 30 min RT), washed with TBS (3·) and
finally distilled water. After embedding in Immu-Mount
(Shandon, Pittsburgh, PA, USA) data were observed with a
confocal microscope (Zeiss).
In combination with anti-GFAP (rabbit polyclonal,
1:100, Santa Cruz), anti-MIB-1/Ki-67 (rabbit polyclonal,
1:2,000, Santa Cruz), and anti-SOX-2 (1:500, rabbit poly-
clonal, Santa Cruz) anti-Nestin (mouse monoclonal, 1:750,
R&D Systems) and anti-Musashi-1 (mouse monoclonal,
1:250, R&D Systems) were always stained first with
anti-mouse Alexa Fluor 555 (goat polyclonal, 1:1,000,
J Neurooncol
123

Invitrogen) as secondary antibody, the second secondary
antibody was anti-rabbit Alexa Fluor 488 (goat polyclonal,
1:1,000, Invitrogen). In combination with anti-Musashi-1
(1:250) anti-Nestin (1:750) was stained first with anti-
mouse Alexa Fluor 555 (goat polyclonal, 1:1,000) as
secondary antibody, the second secondary antibody was
anti-mouse Alexa Fluor 488 (goat polyclonal, 1:1,000). In
combination with anti-GFAP (mouse monoclonal, 1:500;
DAKO, Glostrup, Denmark), and anti-MIB-1/Ki-67
(mouse monoclonal, 1:500, DAKO) anti-SOX-2 (1:500)
was always stained first with anti-rabbit Alexa Fluor 555
(goat polyclonal, 1:1,000) as secondary antibody, the sec-
ond secondary antibody was anti-mouse Alexa Fluor 488
(goat polyclonal, 1:1,000). In combination with anti-GFAP
(mouse monoclonal, 1:500), anti-Nestin (1:750), and anti-
Musashi-1 (1:250) anti-CD133 (goat polyclonal, 1:100,
Santa Cruz) was always stained first with anti-goat Alexa
Fluor 488 (donkey polyclonal, 1:1,000, Invitrogen) as
secondary antibody, the second secondary antibody was
anti-mouse Alexa Fluor 555 (goat polyclonal, 1:1,000). In
combination with anti-SOX-2 (1:500) and anti-MIB-1/Ki-
67 (rabbit polyclonal, 1:2,000) anti-CD133 (1:100) was
first stained with anti-goat Alexa Fluor 488 (donkey
polyclonal, 1:1,000) as secondary antibody, the second
secondary antibody was anti-rabbit Alexa Fluor 488
(donkey polyclonal, 1:1,000, Invitrogen).
Statistical analysis
Statistical methods of Student’s t-test with independent
samples and bivariate correlation analysis (Pearson corre-
lation coefficients) were used. Significance levels ranged
between P < 0.05 (indicated by *), P < 0.01 (indicated by
**) and P < 0.005 (indicated by ***).
Results
Expression of CD133, Nestin, SOX-2 and Musashi-1,
but not of CXCR4, Flt-4 and Endoglin correlated with
malignant grades of astrocytomas
To evaluate mRNA and protein expression levels of the
investigated stem cell markers in astrocytomas of different
WHO grades real-time RT-PCR and IHC of 72 different
tumour samples (see Table 1) and four different NBT
samples were performed. Results are shown in Figs. 1–4.
Although a wide range between single samples oc-
curred, in relation to NBT in astrocytomas of WHO grades
II–IV a clear mRNA over-expression of CD133, Nestin,
SOX-2 and Musashi-1 could be detected (Figs. 1a–4a). The
normalized averaged DCT-values were 9.8, 10.6, 8.5 for
CD133 (NBT 12.5); 1.6, 2.3, 0.6 (NBT 4.6) for Nestin; 6.4,
5.9, 7.1 (NBT 9.5) for SOX-2 and 4.2, 2.3, 4.7 (NBT 6.4)
for Musashi-1 in astrocytomas of WHO grades II, II and
IV, respectively. The statistical differences ranged between
P < 0.05 and 0.005 (see Figs. 1a–4a, indicated by stars). In
contrast to this, with exception of CD133 in astrocytomas
of WHO grade I mean mRNA expression levels of Nestin,
SOX-2 and Musashi-1 were comparable to those of NBT.
These results could be confirmed by IHC. Even if in
relation to NBT not in all cases statistically significant
differences were evaluated and results differed from sam-
ple to sample, in astrocytomas of WHO grade IV the
highest amounts of positive stained cells of CD133, Nestin,
SOX-2 and Musashi-1 were found (Figs. 1b–4b, 1c–4c).
Amounts of positive stained cells were 15.6% for CD133
(NBT 2.3%), 25.3% for Nestin (NBT 4.2%), 9.7% for
SOX-2 (NBT 2.3%) and 9.8% for Musashi-1 (NBT 2.1%),
respectively.
Moreover, a comparison between the averaged DCT-
values and percentages of positive stained cells of CD133,
Nestin, SOX-2 and Musashi-1, especially in astrocytomas
of WHO grade IV, gave interesting results (Figs. 1–4).
While the mean DCT-value of Nestin was the lowest (0.6;
means highest expression) among these four markers, and
similarly, the percentage of nestin positive cells was the
highest (25.3%), the DCT-value of CD133 was the highest
(8.5; means lowest expression) among all markers ob-
served. In contrast to this, the percentage of CD133 posi-
tive cells was higher than that of SOX-2 and Musashi-1
(15.6% for CD133; 9.7% for SOX-2; 9.8% for Musashi-1).
DCT-values of SOX-2 and Musashi-1 were higher than that
of Nestin but lower than that of CD133 (7.1 for SOX-2; 4.7
for Musashi-1).
To evaluate whether a correlation between the expres-
sion of CD133, Nestin, SOX-2 and Musashi-1 and the
pathological grades of astrocytomas could be observed a
bivariate correlation analysis (Pearson correlation coeffi-
cients) was done. Statistically significant positive correla-
tions between the percentages of immunostained cells and
pathological grades of astrocytomas were found for CD133
(cp = 0.318, P < 0.05), Nestin (cp = 0.599, P < 0.001),
SOX-2 (cp = 0.331, P < 0.05) and Musashi-1 (cp = 0.327,
P < 0.05). DCT-values of these four stem cell markers had
a negative correlation with the pathological grades of
astrocytomas, which means that with higher malignant
grades of astrocytomas, higher mRNA amount (low-DCT-
values) could be found. Statistically significant correlations
appeared for Nestin (cp = -0.465, P < 0.005), and SOX-2
(cp = -0.324, P < 0.05).
In relation to NBT mRNA and protein expression levels
of CXCR4, Flt-4 and Endoglin did not correlate signifi-
cantly with malignant grades of astrocytomas (data not
shown). In detail, averaged DCT-values of CXCR4 were
6.82, 6.84, 7.28 and 6.23 in astrocytomas of WHO grades
J Neurooncol
123

I–IV, respectively, and 7.46 in NBT. The amounts of
positive stained CXCR4 cells were 6.17, 5.48, 6.67 and
11.06% in grades I–IV, and 0.97% in NBT. By this,
whereas in astrocytomas of WHO grade IV a notable in-
crease of positive stained CXCR4 cells was detected, and
this difference was statistical significant (P < 0.01), no
correlation between the expression of CXCR4 and the
pathological grades of astrocytomas was found. Same
results were observed for Flt-4/VEGFR-3 and Endoglin/
CD105 mRNA and protein expression levels. Averaged
DCT-values were 8.22, 7.15, 6.96 and 6.77 for Flt-4 (NBT
8.33), and 5.29, 5.15, 5.18 and 3.77 for Endoglin (NBT
5.02) in astrocytomas WHO grades I–IV, respectively. The
amounts of positive stained cells ranged between 1.33, 6.9,
9.71 and 7.91% for Flt-4 (NBT 5.17%) and 3.38, 4.89,
8.85 and 5.71% for Endoglin (NBT 2.27%). Although
Flt-4 and Endoglin mRNA and protein expression levels
increased from low-grade to high-grade astrocytomas, no
statistically significant differences between expression
levels of NBT and tumour tissue samples and no correla-
tion with the pathological grades of astrocytomas was
found.
Fig. 1 Expression of CD133 in
astrocytomas (AC) of different
WHO grades and normal brain
tissue (NBT) evaluated by areal-time RT-PCR (logarithmic
scale, DCT = 3.3 corresponds to
a tenfold difference) and (b, c)
immunohistochemistry (IHC); bgraphical representation of
percentages of CD133 positive
stained cells, c examples of
immunohistochemical staining
of CD133 (original
magnification · 200); inset:
negative control (AC IV)
without primary antibody.
Although a wide range between
single samples occurred,
compared with NBT in high-
grade astrocytomas elevated
mRNA expression levels and
amounts of CD133 positive
cells were found. Normalized
averaged DCT-values were 9.7,
9.8, 10.6 and 8.5 (NBT 12.5),
averaged percentages were 6.9,
6.5, 15 and 15.6% (NBT 2.3%)
in astrocytomas of WHO grades
I–IV, respectively. Results
correlated with malignant
grades of astrocytomas and
were statistical significant
(*P < 0.05; **P < 0.01); UNundetectable
J Neurooncol
123

In summary, the mRNA and protein expression of the
stem cell markers CD133, Nestin, SOX-2 and Musashi-1
correlated with malignant grades of human astrocytomas,
whereas those of CXCR4, Flt-4 and Endoglin did not.
CD133, Nestin, SOX-2 and Musashi-1 could
be co-stained with GFAP and among themselves
in a complex pattern
To analyse whether CD133, Nestin, SOX-2 and Musashi-1
could be co-stained with GFAP (glial fibrillary acidic
protein), Ki-67/MIB-1 (a marker of proliferating cells), or
among themselves confocal microscopy was performed.
Astrocytomas of WHO grade IV were used for investiga-
tions. Results are summarized in Table 2 and exemplary
data are shown in Figs. 5 and 6.
Initially it should be mentioned that both staining
intensities and amounts of positive regions differed among
single samples and within the same sample. Although
especially in tumour regions with high amounts of positive
stained cells results were difficult to interpret, for all
investigated stem cell markers no clearly co-staining with
Fig. 2 Expression of Nestin in
astrocytomas (AC) of different
WHO grades and normal brain
tissue (NBT) evaluated by areal-time RT-PCR (logarithmic
scale, DCT = 3.3 corresponds to
a tenfold difference) and (b, c)
immunohistochemistry (IHC); bgraphical representation of
percentages of Nestin positive
stained cells, c examples of
immunohistochemical staining
of Nestin (original
magnification · 200); inset:
negative control (AC IV)
without primary antibody.
While mean normalized DCT-
values were 3.9, 1.6, 2.3 and 0.6
(NBT 4.6), and averaged
percentages were 7.9, 7.7, 9.6
and 25.3% (NBT 4.2%) in
astrocytomas of WHO grades
I–IV, respectively, an increase
of Nestin expression could be
observed. Results correlated
with malignant grades of
astrocytomas and were
statistical significant
(*P < 0.05; ***P < 0.005)
J Neurooncol
123

Ki-67/MIB-1 could be observed. While the nuclear antigen
Ki-67/MIB-1 is present during all active phases of the cell
cycle (G1, S, G2 and M-phases), but absent in resting cells
(G0-phases), it seems that cells which were going through
the active cell cycle phases did not express CD133, Nestin,
SOX-2 and Musashi-1 in detectable amounts in parallel
(for example see Figs. 5b, 6b).
In contrast to this, we could show that all investigated
stem cell markers could be co-stained with GFAP. Whilst
the intermediate filament protein GFAP is an immunohis-
tochemical marker for localizing benign astrocyte and
neoplastic cells of glial origin in the central nervous sys-
tem, this means that astroglia also expressed CD133,
Nestin, SOX-2 and Musashi-1 (examples for merged
regions are shown in Figs. 5a, 6d). Additionally, cells
which were solely positive for GFAP or the individual stem
cell marker (or negative for both) existed within the tumour
sections (see Figs. 5a, 6d; Table 2).
Expression of both CD133 and Nestin was found in
combination with SOX-2, Musashi-1 and between each
other, respectively (for example see Fig. 5c, d, 6a, c;
Table 2; since different markers are not localized within
the same structures in the cells, signals did not merged in
all cases, but were found in the same regions). Thereby, it
seems that amounts of CD133 and Nestin positive stained
cells were higher than that of SOX-2 and Musashi-1 (see
Fig. 5d, 6a). This was in accordance with the results
obtained by IHC (in astrocytomas of grade IV percentages
of CD133 and Nestin positive stained cells were clearly
higher that that of SOX-2 and Musashi-1, Figs. 1–4).
Fig. 3 Expression of SOX-2 in
astrocytomas (AC) of different
WHO grades and normal brain
tissue (NBT) evaluated by areal-time RT-PCR (logarithmic
scale, DCT = 3.3 corresponds to
a tenfold difference) and (b, c)
immunohistochemistry (IHC); bgraphical representation of
percentages of SOX-2 positive
stained cells, c examples of
immunohistochemical staining
of SOX-2 (original
magnification · 200); inset:
negative control (AC IV)
without primary antibody.
Especially on mRNA level a
clear SOX-2 over-expression
was detected. Mean normalized
DCT-values were 8.8, 6.4, 5.9
and 7.1 (NBT 9.5) in
astrocytomas of WHO grades I-
IV, respectively (*P < 0.05).
Although, the amounts of SOX-
2 positive cells showed no
statistically significant
differences in relation to NBT,
the percentages of SOX-2
labelled cells correlated with
grades of malignancy (1.9, 6.6,
6.4 and 9.7% in astrocytomas
grades I–IV and 2.3% in NBT);
UN undetectable
J Neurooncol
123
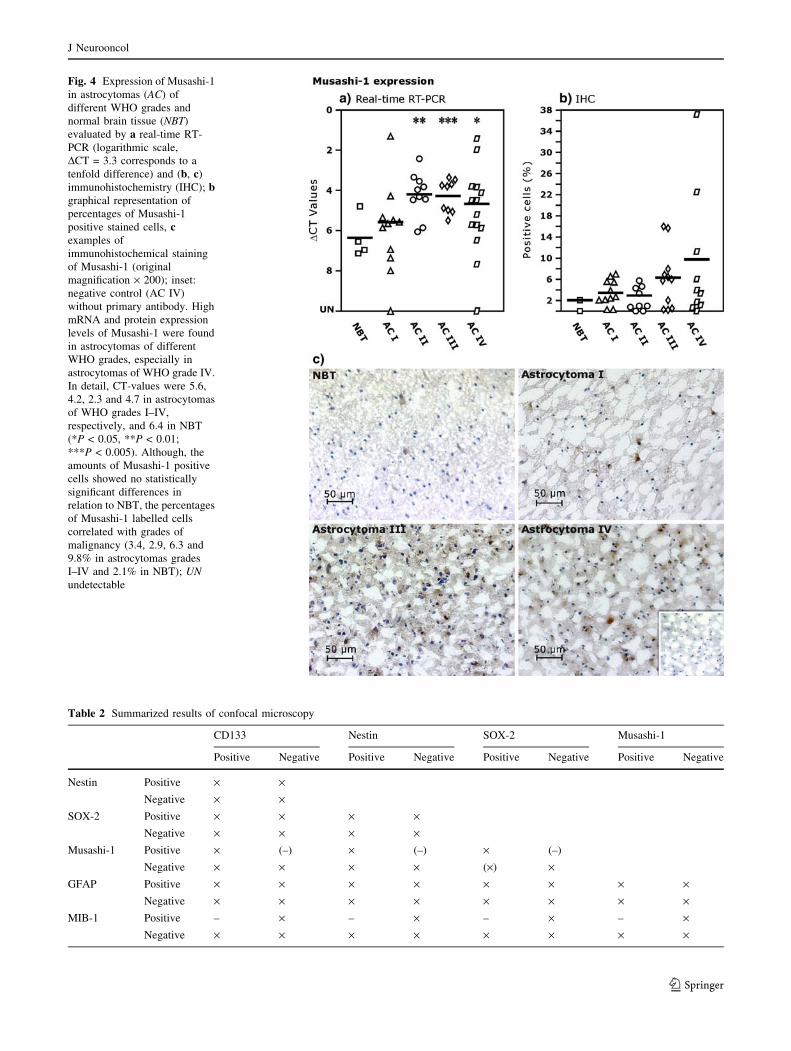
Fig. 4 Expression of Musashi-1
in astrocytomas (AC) of
different WHO grades and
normal brain tissue (NBT)
evaluated by a real-time RT-
PCR (logarithmic scale,
DCT = 3.3 corresponds to a
tenfold difference) and (b, c)
immunohistochemistry (IHC); bgraphical representation of
percentages of Musashi-1
positive stained cells, cexamples of
immunohistochemical staining
of Musashi-1 (original
magnification · 200); inset:
negative control (AC IV)
without primary antibody. High
mRNA and protein expression
levels of Musashi-1 were found
in astrocytomas of different
WHO grades, especially in
astrocytomas of WHO grade IV.
In detail, CT-values were 5.6,
4.2, 2.3 and 4.7 in astrocytomas
of WHO grades I–IV,
respectively, and 6.4 in NBT
(*P < 0.05, **P < 0.01;
***P < 0.005). Although, the
amounts of Musashi-1 positive
cells showed no statistically
significant differences in
relation to NBT, the percentages
of Musashi-1 labelled cells
correlated with grades of
malignancy (3.4, 2.9, 6.3 and
9.8% in astrocytomas grades
I–IV and 2.1% in NBT); UNundetectable
Table 2 Summarized results of confocal microscopy
CD133 Nestin SOX-2 Musashi-1
Positive Negative Positive Negative Positive Negative Positive Negative
Nestin Positive · ·Negative · ·
SOX-2 Positive · · · ·Negative · · · ·
Musashi-1 Positive · (–) · (–) · (–)
Negative · · · · (·) ·GFAP Positive · · · · · · · ·
Negative · · · · · · · ·MIB-1 Positive – · – · – · – ·
Negative · · · · · · · ·
J Neurooncol
123

Consequently, CD133 and Nestin positive cells existed not
only in combination with SOX-2 or Musashi-1, but also
without any co-staining (see Table 2). Moreover, as ini-
tially mentioned in different tumour sections regions with
high and low amounts of positive stained cells existed in
parallel (for CD133 see for example Figs. 5b, 6c).
SOX-2 positive signals were found alone or in combi-
nation with CD133 and Nestin (for example see Fig. 6a;
Fig. 5 Examples of co-staining of different stem cell markers in
astrocytomas grade IV determined by confocal microscopy. Upper
panel: CD133 (green) stained in combination with a GFAP (red) or bKi-67/MIB-1 (red). Beside cells which were solely positive for
CD133 or GFAP also clearly CD133/GFAP double positive cells
were observed (merged regions, yellow). No co-expression of CD133
and Ki-67/MIB-1 could be detected. Lower panel: Nestin (red)
stained in combination with c CD133 (green) and d Musashi-1
(green). Since different markers for Nestin/CD133 or Nestin/
Musashi-1 are not localized within the same structures in the cells,
signals did not merged in all cases, but were found in the same
regions. Whereas, Nestin and CD133 solely stained cells were found,
Musashi-1 seemed to be expressed only in combination with Nestin
Fig. 6 Examples of co-staining of different stem cell markers in
astrocytomas grade IV determined by confocal microscopy. Upper
panel: SOX-2 (red) stained in combination with a CD133 (green) or bKi-67/MIB-1 (green). Beside many CD133 and single SOX-2 positive
cells also some cells which could be co-stained with CD133 and
SOX-2 exited within the tumour section. By this, signals did not
merge, but were found in the same regions. No co-expression of SOX-
2 and Ki-67/MIB-1 could be detected. Lower panel: Musashi-1 (red)
stained in combination with c CD133 (green) and d GFAP (green).
Whereas, Musashi-1 seemed to be expressed only in combination
with CD133, it could be found alone or in combination with GFAP
(merged regions, yellow). Moreover, with this example it should be
shown that in astrocytomas of WHO grade IV regions with high
amounts of CD133 positive cells (c) and those with low amounts (d)
exited
J Neurooncol
123

Table 2). By this, since different markers are not localized
within the same structures in the cells, fluorescence signals
did not merged in all cases, but were found in the same
regions (e.g. see Fig. 6a). If a co-staining of SOX-2 and
Musashi-1 was performed, it seems that SOX-2 positive
cells were found with and without Musashi-1 expression in
parallel. Since in astrocytomas WHO grade IV averaged
percentages of SOX-2 and Musashi-1 positive cells were
only 9.7 and 9.8%, respectively, and sometimes single
structures were hard to investigate, it was very difficult to
evaluate whether SOX-2 positive/Musashi-1 negative cells
exist. Thereby, in Table 2 this result was demonstrated
with brackets. Moreover, it seems that Musashi-1 positive
could be only evaluated in combination with CD133,
Nestin and SOX-2 (for example see Figs. 5d, 6c, d). Flu-
orescence signals did not merged in all cases, but were
found in the same regions. For same reasons as mentioned
above in Table 2 results were shown with brackets.
In summary, CD133, Nestin, SOX-2 and Musashi-1
could be co-stained with GFAP and among themselves in a
complex pattern. In relation to CD133 and Nestin, SOX-2
and Musashi-1 were found in lower amounts in astrocytoma
grade IV tumour sections, and Musashi-1 seems to be ex-
pressed only in combination with other stem cell markers.
Discussion
Several recent reports suggest that TSCs exist in high-grade
astrocytomas and could be isolated from these tumours.
Nevertheless, little is known about the expression of these
markers in solid astrocytomas, especially in relation with
the malignant grades of these tumours. Herein, the aim of
this study was to investigate the expression of CD133,
Nestin, SOX-2, Musashi-1, CXCR4, Flt-4 and Endoglin on
mRNA and protein level, to analyse their relation with the
malignant grades of different astrocytomas, and to study the
co-expression of theses genes with markers for glial and
proliferating cells, and among themselves.
CD133/Prominin
Which belongs to the pentaspan membrane glycoproteins
family, was originally described to be expressed in neu-
roepithelial and hematopoietic stem cells (HSCs) [32, 33],
CD133 could be used to identify and isolate human stem
cells from various sources including CNS [33, 34]. We
could show that in relation to NBT in astrocytomas
amounts of CD133 positive stained cells were higher
(15.6% in astrocytomas of WHO grade IV; 2.3% in NBT)
and correlated with the malignant grades of these tumours.
Results were confirmed by real-time RT-PCR (DCT-values
were 12.5 and 8.5 in astrocytomas grade IV and NBT,
respectively). The high amount of CD133 positive cells
exceeds our expectation; however it is similar to the result
that 6–29% of putative TSCs (CD133 positive cells) were
sorted from glioblastomas [8]. It was found that more than
10,000 unsorted cancer cells were needed to initiate tu-
mours [19], which implied that the TSCs fraction was less
than 0.01% of the unsorted population. Even if as low as
100 CD133 positive glioblastoma cells were necessary to
form tumours in immunodeficient mice [8], theoretically,
the proportion of TSCs (sorted CD133 positive cell popu-
lation) in solid tumour should be only 1%. But, in contrast
to that and in accordance with our result, 11–35% cells that
expressed the stem cell markers CD44+/CD24– were sorted
from breast cancer in another study [20]. These data sug-
gest that not all of the CD133 positive cells are true TSCs,
even though the CD133 positive cell portion represents
undifferentiated cells. In fact, primary tumour sphere-de-
rived cells (CD133 positive) formed secondary spheres
with a range of 0.53–1.0 secondary spheres/100 primary
sphere cells [19]. So, CD133 positive cells are not always
linked with the ability of self-renewal and tumourigenesis.
Nestin
An intermediate filament protein typical for neural pre-
cursor cells, has been extensively used as a marker for
NSCs [35, 36]. Based on the observations of a correlation
of Nestin expression with NSCs in vivo, now Nestin is also
used as a typical marker for identifying TSCs from CNS
tumours [5–8, 19, 29, 37]. In primary malignant CNS tu-
mours high amounts (more than 50%) of Nestin positive
cells have been reported [38, 39]. Our results are in
agreement with these reports. Astrocytomas were abun-
dantly positive for Nestin, and the amounts of Nestin po-
sitive cells increased with malignant grades of these
tumours. The highest proportion of Nestin positive cells
(average 25.3%) was found in astrocytomas of WHO grade
IV with a statistically significant difference in relation to
NBT (4.2%). Moreover, compared with NBT in astrocy-
tomas of WHO grades II, III and IV a statistically signifi-
cant up-regulated expression of Nestin mRNA was found
(DCT-values were 1.6, 2.3, 0.6, respectively, and 4.6 in
NBT).
Nestin is expressed in primitive neuroepithelial cells of
all regions of the CNS during development [40], whereas in
the adult CNS, Nestin is a marker for NSCs lining in
the ventricular wall and the central canal [41]. However,
other studies showed that Nestin was also detected in
NBT ?tul?> outside of the subependymal zone [42–45],
and could be re-expressed in astrocytes of the adult CNS in
response to cellular stress, including brain trauma/trans-
plantation [46–48], CNS ischemia [48, 49], neurotoxicant
[50], neoplastic transformation [38, 39] and even altered
J Neurooncol
123

trophic stimulation [51]. The relation between the ability of
neurosphere formation and Nestin expression in adult brain
is different from that in embryonic brain [52]. So, Nestin
expression is complex at every stage of stem cell devel-
opment and differentiation.
SOX-2
Encodes a SRY-related high mobility group of box tran-
scription factors, which act as transcriptional activators for
a variety of genes that are developmentally regulated [53,
54]. SOX-2 expression is found in at least three types of
stem cells (NSCs, embryonic and trophoblast stem cells)
[54, 55]. Glioma-derived neurospheres can express SOX-2,
and SOX-2 mRNA expression could be detected in brain
tumours [6]. In this study we could show that SOX-2 was
expressed in astrocytomas, and its expression was up reg-
ulated in relation to NBT. Although no statistically sig-
nificant differences of SOX-2 protein expression could be
detected by IHC between astrocytomas and NBT (9.7% in
astrocytomas grade IV, 2.3% in NBT), SOX-2 mRNA
expression was really significantly up regulated in astro-
cytomas of WHO grades II, III and IV (DCT-values were
6.4, 5.6, 7.1, respectively, and 9.5 in NBT) Moreover,
amounts of SOX-2 positive cells and mRNA expression
levels increased with malignant grades of these tumours.
SOX-2 appears to be a common marker for all neural
cells within heterogeneous neurospheres derived from
different parts of the brain [56]. However, that may be not
right for tumour-derived spheres. Tumour-derived spheres
could acquire SOX-2 mRNA expression that was absent in
their parent tumours or they could lose SOX-2 expression
[6].
Musashi-1
Encodes an evolutionarily conserved group of RNA-bind-
ing proteins, which are selectively expressed in neural
progenitor/stem cells, and play a key role in the mainte-
nance of the stem cell state during embryonic CNS
development. Musashi-1 is also an excellent marker for
neural progenitor/stem cells in adult brains [57], and is
expressed in neuronal precursors of the cerebellum and
subventricular zone [58, 59]. In our study, high mRNA and
protein expression levels of Musashi-1 were found in
astrocytomas of different WHO grades, especially in
astrocytomas of WHO grade IV. In detail, DCT-values
were 4.2, 2.3 and 4.7 in astrocytomas of WHO grades II–
IV, respectively, and 6.4 in NBT. Although, the amounts of
Musashi-1 positive cells showed no statistically significant
differences in relation to NBT, the percentages of Musashi-
1 labelled cells correlated with grades of malignancy (2.9,
6.3 and 9.8% in astrocytomas grade II–IV and 2.1% in
NBT). In accordance with these results, previous studies
showed that the expression of Musashi-1 could be corre-
lated to the proliferating activity of gliomas [57]. No
relationship between the grades of malignancy and the
degree of Musashi-1 expression could be observed in hu-
man hepatoma cells [60]. Tumour-derived spheres (undif-
ferentiated and differentiated) could acquire Musashi-1
mRNA expression which was absent in their parent tu-
mours, or they could loss Musashi-1 expression [6]. So,
mRNA expression of Musashi-1 appears to be dynamic.
CXCR4
A chemokine receptor, belongs to the family of G protein-
coupled receptors and is selective for a single chemokine,
the stromal cell-derived factor 1 (SDF-1, CXCL12).
CXCR4 is expressed by NSCs of the cerebellar external
granular layer [26], and CXCR4 positive NSCs could be
isolated from mouse neurospheres. Moreover, the intrace-
rebral administration of SDF-1 enabled intravenously in-
jected CXCR4 positive NSCs to migrate into the CNS and
differentiate into neurons [61]. Although in our study no
correlation between the expression of CXCR4 and the
pathological grades of astrocytomas was found, in astro-
cytomas of WHO grade IV a notable increase of positive
stained CXCR4 cells was detected, and this difference was
statistical significant (P < 0.01).
Flt-4/VEGFR-3 (fms-like-tyrosine kinase-4, also
named vascular endothelial growth factor receptor-3)
A receptor for VEGF-C and VEGF-D, is a member of the
tyrosine protein kinase family, and was first observed in
endothelial cell precursors of mouse embryos [62]. CD34
positive cells, isolated from human fetal liver, were posi-
tive for Flt-4 [63]. These CD34+/Flt-4+ cells, which also
co-expressed CD133, were functionally endothelial cell
precursors, which demonstrated that Flt-4 might be a po-
tential stem cell marker. Furthermore, murine neural pro-
genitor cells express VEGFR-3, and VEGF-C is a trophic
factor for these cells [27].
Endoglin (CD105)
Is a homodimeric cell surface component of the trans-
forming growth factor-beta (TGF-b) type I receptor com-
plex [64]. Several reports showed that Endoglin was a
highly specific marker for HSCs and could be used to
purify HSCs from bone marrow [65–67]. TGF-b1 induces
Endoglin expression and is a crucial factor for maintaining
immaturity of Endoglin positive HSCs [67].
Although Flt-4 and Endoglin mRNA and protein
expression levels increased from low-grade to high-grade
J Neurooncol
123

astrocytomas, in our study and no correlation with the
pathological grades of astrocytomas was found.
To evaluate whether in astrocytomas of WHO grade IV
CD133, Nestin, SOX-2 and Musashi-1 could be co-stained
with GFAP, Ki-67/MIB-1 or among themselves, confocal
microscopy was performed. Whereas, in our study none of
these stem cell markers were co-expressed with Ki-67/
MIB-1, a co-staining of CD133, Nestin, SOX-2 and Mus-
ashi-1 with GFAP could be observed in some cells. These
results are in accordance with findings that Nestin positive
cells can co-express GFAP or beta III-tubulin [44–47]. In
contrast to this, Kanemura et al. demonstrated that Mus-
ashi-1 was expressed in GFAP-negative tumour cells that
formed foci and were surrounded by GFAP-positive cells
[57]. While the expression of Musashi-1 could be corre-
lated with the proliferating activity of gliomas [68], and
this molecule could be found in proliferating cells of the
neural retina [69], some studies demonstrated that Mus-
ashi-1 positive tumour cells did not co-express PCNA or
Ki-67 [70]. Now we could show that in astrocytomas grade
IV CD133, Nestin, SOX-2 and Musashi-1 could be co-
stained with each other, and Musashi-1 seems to be ex-
pressed only in combination with other stem cell markers.
Our results reveal a high heterogeneity of stem cell
marker expression within astrocytomas. Furthermore, the
expression pattern is rather complex, and the different
markers do not necessarily conincide with each other. Also,
cells positive for stem cell marker are not always highly
proliferative ones. It appears that some of the markers, e.g.
Nestin, result from dedifferentiation or from activated state
of the glial tumour cells. Clearly, there is a need for
defining TSCs by more appropriate functional markers, e.g.
by receptors for growth or chemotactic factors that are
involved in stem cell maintenance or differentiation.
Assuming that the most of the investigated markers
represent more or less NSCs or TSCs, astrocytomas in situ
contain a considerable amount of cells with stem cell
character. With increasing WHO grade some of these
markers, namely, CD133, Nestin, Sox-2 and Musashi-1
increase, while others (CXCR4, Flt-4, Endoglin) are ele-
vated over NBT, but are independent for the tumour grade.
It cannot be judged from the in situ expression data whe-
ther these stem cells are the origin of ‘‘differentiated’’
astrocytoma cells or whether astroglial tumours attract
adult stem cells from other parts of the brain. The latter
hypothesis could be supported by an increase of cells po-
sitive for stem cell markers with the WHO grade, whereas
for the first (TSC) hypothesis this would not be required.
Perhaps, both possibilities exist side by side, and mixed
populations of NSCs and (more differentiated) TSCs exist
together, especially in gliomas. Therefore, further investi-
gations on stem cells in astrocytomas/gliomas should
consider their heterogeneity and focus more on functional
markers and properties.
Acknowledgements We thank Brigitte Rehmke, Ute Malcus-Cos-
kun and Jorg Krause for expert technical assistance and Clemens
Franke for drawing pictures. This work was supported by an intra-
mural grant of the Universitatsklinikum Schleswig-Holstein, Campus
Kiel (JH-F & RM) ‘‘Stem cells in brain tumours’’.
References
1. Mahaley MS Jr, Mettlin C, Natarajan N, Laws ER Jr, Peace B
(1989) National survey of patterns of care for brain-tumour pa-
tients. J Neurosurg 71:826–836
2. Kleihues P, Soylemezoglu F, Schauble B, Scheithauer BW,
Burger PC (1995) Histopathology, classification, and grading of
gliomas. Glia 15:211–221
3. Nozaki M, Tada M, Kobayashi H, Zhang CL, Sawamura Y, Abe
H, Ishii N, Van Meir EG (1999) Roles of the functional loss of
p53 and other genes in astrocytoma tumourigenesis and pro-
gression. Neuro-Oncol 1:124–137
4. von Deimling A, Louis DN, Wiestler OD (1995) Molecular
pathways in the formation of gliomas. Glia 15:328–338
5. Singh SK, Clarke ID, Terasaki M, Bonn VE, Hawkins C, Squire
J, Dirks PB (2003) Identification of a cancer stem cell in human
brain tumours. Cancer Res 63:5821–5828
6. Hemmati HD, Nakano I, Lazareff JA, Masterman-Smith M,
Geschwind DH, Bronner-Fraser M, Kornblum H (2003) Can-
cerous stem cells can arise from pediatric brain tumours. Proc
Natl Acad Sci USA 100:15178–15183
7. Galli R, Binda E, Orfanelli U, Cipelletti B, Gritti A, De Vitis S,
Fiocco R, Foroni C, Dimeco F, Vescovi A (2004) Isolation and
characterization of tumourigenic, stem-like neural precursors
from human glioblastoma. Cancer Res 64:7011–7021
8. Singh SK, Hawkins C, Clarke ID, Squire JA, Bayani J, Hide T,
Henkelman RM, Cusimano MD, Dirks PB (2004) Identification
of human brain tumour initiating cells. Nature 432:396–401
9. Dahmane N, Sanchez P, Gitton Y, Palma V, Sun T, Beyna M,
Weiner H, Ruiz i Altaba A (2001) The Sonic Hedgehog-Gli
pathway regulates dorsal brain growth and tumourigenesis.
Development 128:5201–5212
10. Ruiz i Altaba A, Sanchez P, Dahmane N (2002) Gli and hedgehog
in cancer: tumours, embryos and stem cells. Nat Rev Cancer
2:361–372
11. Pardal R, Clarke MF, Morrison SJ (2003) Applying the principles
of stem-cell biology to cancer. Nat Rev Cancer 3:895–902
12. Reya T, Morrison SJ, Clarke MF, Weissman IL (2001) Stem
cells, cancer, and cancer stem cells. Nature 414:105–111
13. Zhu Y, Guignard F, Zhao D, Liu L, Burns DK, Mason RP,
Messing A, Parada LF (2005) Early inactivation of p53 tumor
suppressor gene cooperating with NF1 loss induces malignant
astrocytoma. Cancer Cell 8:119–130
14. You H, Kim YI, Im SY, Suh-Kim H, Paek SH, Park SH, Kim DG,
Jung HW (2005) Immunohistochemical study of central neuro-
cytoma, subependymoma, and subependymal giant cell astrocy-
toma. J Neuro-Oncol 74:1–8
15. Rao G, Pedone CA, Coffin CM, Holland EC, Fults DW (2003) c-
Myc enhances sonic hedgehog-induced medulloblastoma forma-
tion from nestin-expressing neural progenitors in mice. Neoplasia
5:198–204
16. Holland EC, Celestino J, Dai C, Schaefer L, Sawaya RE, Fuller
GN (2000) Combined activation of Ras and Akt in neural pro-
J Neurooncol
123

genitors induces glioblastoma formation in mice. Nat Genet
25:55–57
17. Uhrbom L, Dai C, Celestino JC, Rosenblum MK, Fuller GN,
Holland EC (2002) Ink4a-Arf loss cooperates with KRas acti-
vation in astrocytes and neural progenitors to generate glioblas-
tomas of various morphologies depending on activated Akt.
Cancer Res 62:5551–5558
18. Collins AT, Berry PA, Hyde C, Stower MJ, Maitland NJ (2005)
Prospective identification of tumourigenic prostate cancer stem
cells. Cancer Res 65:10946–10951
19. Taylor MD, Poppleton H, Fuller C, Su X, Liu Y, Jensen P,
Magdaleno S, Dalton J, Calabrese C, Board J, Macdonald T,
Rutka J, Guha A, Gajjar A, Curran T, Gilbertson RJ (2005) Ra-
dial glia cells are candidate stem cells of ependymoma. Cancer
Cell 8:323–335
20. Al-Hajj M, Wicha MS, Benito-Hernandez A, Morrison SJ, Clarke
MF (2003) Prospective identification of tumourigenic breast
cancer cells. Proc Natl Acad Sci USA 100:3983–3988
21. Fang D, Nguyen TK, Leishear K, Finko R, Kulp AN, Hotz S, Van
Belle PA, Xu X, Elder DE, Herlyn M (2005) A tumourigenic
subpopulation with stem cell properties in melanomas. Cancer
Res 65:9328–9337
22. Nilsson L, Astrand-Grundstrom I, Arvidsson I, Jacobsson B,
Hellstrom-Lindberg E, Hast R, Jacobsen SE (2000) Isolation and
characterization of hematopoietic progenitor/stem cells in 5q-
deleted myelodysplastic syndromes: evidence for involvement at
the hematopoietic stem cell level. Blood 96:2012–2021
23. Seigel GM, Campbell LM, Narayan M, Gonzalez-Fernandez F
(2005) Cancer stem cell characteristics in retinoblastoma. Mol
Vis 11:729–737
24. Hirschmann-Jax C, Foster AE, Wulf GG, Nuchtern JG, Jax TW,
Gobel U, Goodell MA, Brenner MK (2004) A distinct ‘‘side
population’’ of cells with high drug efflux capacity in human
tumour cells. Proc Natl Acad Sci USA 101:14228–14233
25. Kim CF, Jackson EL, Woolfenden AE, Lawrence S, Babar I,
Vogel S, Crowley D, Bronson RT, Jacks T (2005) Identification
of bronchioalveolar stem cells in normal lung and lung cancer.
Cell 121:823–835
26. Reiss K, Mentlein R, Sievers J, Hartmann D (2002) Stromal cell-
derived factor 1 is secreted by meningeal cells and acts as che-
motactic factor on neuronal stem cells of the cerebellar external
granular layer. Neuroscience 115:295–305
27. Le Bras B, Barallobre MJ, Homman-Ludiye J, Ny A, Wyns S,
Tammela T, Haiko P, Karkkainen MJ, Yuan L, Muriel MP,
Chatzopoulou E, Breant C, Zalc B, Carmeliet P, Alitalo K,
Eichmann A, Thomas JL (2006) VEGF-C is a trophic factor for
neural progenitors in the vertebrate embryonic brain. Nat Neu-
rosci 9:340–348
28. Henriksen R, Gobl A, Wilander E, Oberg K, Miyazono K, Funa K
(1995) Expression and prognostic significance of TGF-beta iso-
types, latent TGF-beta 1 binding protein, TGF-beta type I and
type II receptors, and endoglin in normal ovary and ovarian
neoplasms. Lab Invest 73:213–220
29. Yuan X, Curtin J, Xiong Y, Liu G, Waschsmann-Hogiu S, Farkas
DL, Black KL, Yu JS (2004) Isolation of cancer stem cells from
adult glioblastoma multiforme. Oncogene 23:9392–9400
30. Ignatova TN, Kukekov VG, Laywell ED, Suslov ON, Vrionis FD,
Steindler DA (2002) Human cortical glial tumours contain neural
stem-like cells expressing astroglial and neuronal markers in vi-
tro. Glia 39:193–206
31. Salmaggi A, Boiardi A, Gelati M, Russo A, Calatozzolo C, Ci-
usani E, Sciacca FL, Ottolina A, Parati EA, La Porta C, Aless-
andri G, Marras C, Croci D, De Rossi M (2006) Glioblastoma-
derived tumourspheres identify a population of tumour stem-like
cells with angiogenic potential and enhanced multidrug resistence
phenotype. Glia 54:850–860
32. Weigmann A, Corbeil D, Hellwig A, Huttner WB (1997)
Prominin, a novel microvilli-specific polytopic membrane protein
of the apical surface of epithelial cells, is targeted to plasma-
lemmal protrusions of non-epithelial cells. Proc Natl Acad Sci
USA 94:12425–12430
33. Yin AH, Miraglia S, Zanjani ED, Almeida-Porada G, Ogawa M,
Leary AG, Olweus J, Kearney J, Buck DW (1997) AC133, a
novel marker for human hematopoietic stem and progenitor cells.
Blood 90:5002–5012
34. Uchida N, Buck DW, He D, Reitsma MJ, Masek M, Phan TV,
Tsukamoto AS, Gage FH, Weissman IL (2000) Direct isolation of
human central nervous system stem cells. Proc Natl Acad Sci
USA 97:14720–14725
35. Cattaneo E, McKay R (1990) Proliferation and differentiation of
neuronal stem cells regulated by nerve growth factor. Nature
347:762–765
36. Lendahl U, Zimmerman LB, McKay RD (1990) CNS stem cells
express a new class of intermediate filament protein. Cell 60:585–
595
37. Kondo T, Setoguchi T, Taga T (2004) Persistence of a small
subpopulation of cancer stem-like cells in the C6 glioma cell line.
Proc Natl Acad Sci USA 101:781–786
38. Almqvist PM, Mah R, Lendahl U, Jacobsson B, Hendson G
(2002) Immunohistochemical detection of nestin in pediatric
brain tumours. J Histochem Cytochem 50:147–158
39. Tohyama T, Lee VM, Rorke LB, Marvin M, McKay RD, Tro-
janowski JQ (1992) Nestin expression in embryonic human
neuroepithelium and in human neuroepithelial tumour cells. Lab
Invest 66:303–313
40. Hockfield S, McKay RD (1985) Identification of major cell
classes in the developing mammalian nervous system. J Neurosci
5:3310–3328
41. Johansson CB, Momma S, Clarke DL, Risling M, Lendahl U,
Frisen J (1999) Identification of a neural stem cell in the adult
mammalian central nervous system. Cell 96:25–34
42. Gu H, Wang S, Messam CA, Yao Z (2002) Distribution of nestin
immunoreactivity in the normal adult human forebrain. Brain Res
943:174–180
43. Lu G, Kwong WH, Li Q, Wang X, Feng Z, Yew DT (2005) bcl2,
bax, and nestin in the brains of patients with neurodegeneration
and those of normal aging. J Mol Neurosci 27:167–174
44. Wei LC, Shi M, Chen LW, Cao R, Zhang P, Chan YS (2002)
Nestin-containing cells express glial fibrillary acidic protein in
the proliferative regions of central nervous system of postnatal
developing and adult mice. Brain Res Dev Brain Res 139:9–17
45. Mignone JL, Kukekov V, Chiang AS, Steindler D, Enikolopov G
(2004) Neural stem and progenitor cells in nestin-GFP transgenic
mice. J Comp Neurol 469:311–324
46. Holmin S, Almqvist P, Lendahl U, Mathiesen T (1997) Adult
nestin-expressing subependymal cells differentiate to astrocytes
in response to brain injury. Eur J Neurosci 9:65–75
47. Krum JM, Rosenstein JM (1999) Transient coexpression of nes-
tin, GFAP, and vascular endothelial growth factor in mature
reactive astroglia following neural grafting or brain wounds. Exp
Neurol 160:348–360
48. Lin RC, Matesic DF, Marvin M, McKay RD, Brustle O (1995)
Re-expression of the intermediate filament nestin in reactive as-
trocytes. Neurobiol Dis 2:79–85
49. Li Y, Chopp M (1999) Temporal profile of nestin expression after
focal cerebral ischemia in adult rat. Brain Res 838:1–10
50. Yoo YM, Lee U, Kim YJ (2005) Apoptosis and nestin expression
in the cortex and cultured astrocytes following 6-OHDA admin-
istration. Neurosci Lett 382:88–92
51. Schmidt-Kastner R, Humpel C (2002) Nestin expression persists
in astrocytes of organotypic slice cultures from rat cortex. Int J
Dev Neurosci 20:29–38
J Neurooncol
123

52. Kawaguchi A, Miyata T, Sawamoto K, Takashita N, Murayama
A, Akamatsu W, Ogawa M, Okabe M, Tano Y, Goldman SA,
Okano H (2001) Nestin-EGFP transgenic mice: visualization of
the self-renewal and multipotency of CNS stem cells. Mol Cell
Neurosci 17:259–273
53. van de Wetering M, Oosterwegel M, van Norren K, Clevers H
(1993) Sox-4, an Sry-like HMG box protein, is a transcriptional
activator in lymphocytes. EMBO J 12:3847–3854
54. Yuan H, Corbi N, Basilico C, Dailey L (1995) Developmental-
specific activity of the FGF-4 enhancer requires the synergistic
action of Sox2 and Oct-3. Genes Dev 9:2635–2645
55. Avilion AA, Nicolis SK, Pevny LH, Perez L, Vivian N, Lovell-
Badge R (2003) Multipotent cell lineages in early mouse devel-
opment depend on SOX2 function. Genes Dev 17:126–140
56. Brazel CY, Limke TL, Osborne JK, Miura T, Cai J, Pevny L, Rao
MS (2005) Sox2 expression defines a heterogeneous population
of neurosphere-forming cells in the adult murine brain. Aging
Cell 4:197–207
57. Kanemura Y, Mori K, Sakakibara S, Fujikawa H, Hayashi H,
Nakano A, Matsumoto T, Tamura K, Imai T, Ohnishi T, Fushiki
S, Nakamura Y, Yamasaki M, Okano H, Arita N (2001) Mus-
ashi1, an evolutionarily conserved neural RNA-binding protein,
is a versatile marker of human glioma cells in determining their
cellular origin, malignancy, and proliferative activity. Differen-
tiation 68:141–152
58. Sakakibara S, Imai T, Hamaguchi K, Okabe M, Aruga J, Nak-
ajima K, Yasutomi D, Nagata T, Kurihara Y, Uesugi S, Miyata T,
Ogawa M, Mikoshiba K, Okano H (1996) Mouse-Musashi-1, a
neural RNA-binding protein highly enriched in the mammalian
CNS stem cell. Dev Biol 176:230–242
59. Sakakibara S, Okano H (1997) Expression of neural RNA-bind-
ing proteins in the postnatal CNS: implications of their roles in
neuronal and glial cell development. J Neurosci 17:8300–8312
60. Shu HJ, Saito T, Watanabe H, Ito JI, Takeda H, Okano H, Kawata
S (2002) Expression of the Musashi1 gene encoding the RNA-
binding protein in human hepatoma cell lines. Biochem Biophys
Res Commun 293:150–154
61. Corti S, Locatelli F, Papadimitriou D, Donadoni C, Del Bo R,
Fortunato F, Strazzer S, Salani S, Bresolin N, Comi GP (2005)
Multipotentiality, homing properties, and pyramidal neurogenesis
of CNS-derived LeX (ssea-1)+/CXCR4+ stem cells. FASEB J
19:1860–1862
62. Kaipainen A, Korhonen J, Mustonen T, van Hinsbergh VW, Fang
GH, Dumont D., Breitman M, Alitalo K (1995) Expression of the
fms-like tyrosine kinase 4 gene becomes restricted to lymphatic
endothelium during development. Proc Natl Acad Sci USA
92:3566–35670
63. Salven P, Mustjoki S, Alitalo R, Alitalo K, Rafii S (2003)
VEGFR-3 and CD133 identify a population of CD34+ lymphatic/
vascular endothelial precursor cells. Blood 101:168–172
64. Cheifetz S, Bellon T, Cales C, Vera S, Bernabeu C, Massague J,
Letarte M (1992) Endoglin is a component of the transforming
growth factor-beta receptor system in human endothelial cells. J
Biol Chem 267:19027–19030
65. Chen CZ, Li L, Li M, Lodish HF (2003) The endoglin (positive)
sca-1 (positive) rhodamine (low) phenotype defines a near-
homogeneous population of long-term repopulating hematopoi-
etic stem cells. Immunity 19:525–533
66. Chen CZ, Li M, de Graaf D, Monti S, Gottgens B, Sanchez MJ,
Lander ES, Golub TR, Green AR, Lodish HF (2002) Identifica-
tion of endoglin as a functional marker that defines long-term
repopulating hematopoietic stem cells. Proc Natl Acad Sci USA
99:15468–15473
67. Pierelli L, Bonanno G, Rutella S, Marone M, Scambia G, Leone
G (2001) CD105 (endoglin) expression on hematopoietic stem/
progenitor cells. Leuk Lymphoma 42:1195–1206
68. Toda M, Iizuka Y, Yu W, Imai T, Ikeda E, Yoshida K, Kawase T,
Kawakami Y, Okano H, Uyemura K (2001) Expression of the
neural RNA-binding protein Musashi1 in human gliomas. Glia
34:1–7
69. Wilson JM, Sato K, Chernoff EA, Belecky-Adams TL (2007)
Expression patterns of chick Musashi-1 in the developing nervous
system. Gene Expr Patterns Apr 21, Epub ahead of print
70. Akasaka Y, Saikawa Y, Fujita K, Kubota T, Ishikawa Y, Fu-
jimoto A, Ishii T, Okano H, Kitajima M (2005) Expression of a
candidate marker for progenitor cells, Musashi-1, in the prolif-
erative regions of human antrum and its decreased expression in
intestinal metaplasia. Histopathology 47:348–356
J Neurooncol
123