expression of killer cell inhibitory receptors on human uterine natural killer cells
TRANSCRIPT

Eur. J. Immunol. 1997.27: 979-983 KIR on human uterine N K cells 979
Sanjay Verma, Ashley King and Yung Wai Loke
Research Group in Human Reproductive Immunobiology, Department of Pathology, University of Cambridge, Cambridge, GB
Expression of killer cell inhibitory receptors on human uterine natural killer cells
The establishment of the human placenta in early pregnancy is characterized by the presence of large numbers of natural killer (NK) cells within the maternal decidua in close proximity to the fetally-derived invading extravillous tropho- blast which expresses at least two HLA class I molecules, HLA-G and HLA-C. These NK cells have an unusual phenotype, CD56h"gh' CD16, distinguishing them from adult peripheral blood NK cells. They may control key events in tro- phoblast migration and therefore placentation. Human NK cells in peripheral blood express receptors for polymorphic HLA class I molecules. This family of receptors, known as killer cell inhibitory receptors (KIR), are expressed on overlapping subsets of NK cells to give an NK cell repertoire which differs between individuals. Using a panel of monoclonal antibodies to several mem- bers of the KIR family and analysis by flow cytometry, we have found that KIR are expressed by decidual NK cells. There is variation in both the percentage of cells expressing a particular receptor and the density of receptor expression between decidual NK cells from different individuals. Comparison of NK cells from decidua and peripheral blood of the same individual showed that NK cells from these two different locations express different repertoires of KIR. Recep- tors are present in individuals who do not possess the relevant class I ligand, raising the possibility that these NK receptors may be involved in recognition of the allogeneic fetus by the mother at the implantation site.
1 Introduction
Natural killer (NK) cells are present in large numbers in the human uterine mucosa (decidua) during early preg- nancy and account for -70% of the leukocytes present [l-31. These uterine NK cells differ phenoty ically from classical circulating NK cells, being CD56'ngh' CD16-. There are several observations which suggest maternal tis- sue NK cells have an important role to play in placenta- tion: they are hormonally regulated, increasing in number during the luteal phase of the menstrual cycle when implantation occurs [4]; they are present in the early part of gestation at the time placental trophoblast cells invade into decidua [5]; and they are particularly abundant around these infiltrating trophoblast cells [6]. Both over- and under-invasion have adverse clinical outcomes [6]. It has been proposed that uterine NK cells could control pla-
[I 164171
Received October 4, 1996; in revised form January 27, 1997; accepted January 27, 1997.
centation by influencing trophoblast invasion and differen- tiation [7].
Human NK cells circulating in peripheral blood express inhibitory receptors which have specificities for different polymorphic HLA class I molecules. This family of recep- tors, known as killer cell inhibitory receptors (KIR) are members of the Ig superfamily [8, 91. A particular KIR is expressed on a subset of NK cells and these subsets may overlap, indicating that individual NK cells can express one, two, or more receptors for class I molecules. There is heterogeneity in expression of KIR between different indi- viduals, but the pattern of expression in an individual remains stable over years [lo]. In addition, NK cells are found which express a particular KIR in individuals who do not possess the relevant class I ligand [lo, 111, indicat- ing that these receptors may be capable of allogeneic rec- ognition. The maternal recognition of the fetus during pregnancy is the only occasion when allogeneic cells natur- ally intermingle. Furthermore, the maternal NK cells in the placental bed are closely associated with invading trophoblast which expresses at least two HLA class I molecules, HLA-G and HLA-C [12-151.
Correspondence: Ashley King, Research Group in Human Reproductive Immunobiology, Department of Pathology, Univer- sity of Cambridge, Tennis Court Road, Cambridge CB2 IQP, GB Fax: 44-1223-333727; e-mail: [email protected]
It would be of interest to determine whether the distinctive CD56b'igh' CD16- NK cells in decidua also express KIR reactive with HLA class I molecules. Furthermore, it is not established whether the percentage of decidual NK cells expressing a particular KIR and-its surface density can differ between individuals. Finally, it is known whether the same exmession Datterns are found in blood
Abbreviations: KIR: Killer cell inhibitory receptor RLF Rela- tive linear fluorescence intensity
0 VCH Verlagsgesellschaft mbH, D-69451 Weinheim, 1997 0014-298Ol9710404-979$17.50 + .50/0

980 S. Verma et al. Eur. J. Immunol. 1997.27: 979-983
2 Materials and methods
2.1 Extraction of decidual and peripheral blood lymphocytes
Maternal decidual tissue was obtained from elective first trimester terminations of pregnancy (between 8 and 12 weeks) by suction curettage. Decidual lymphocytes were isolated as described [16]. Phenotypic characterization of these cells has been performed [l , 31. Peripheral blood lymphocytes were obtained from normal adults as before ~ 7 1 .
2.2 Immunofluorescent labeling and flow cytometry
The mAb to NK cell receptors used in the study were EB6 (p58.Up50.1); GL183 (p58.2/p50.2); HP3E4 (p58.l/p50.1/ p50.3); DX9 (p70); 5.133 (p70), and Hp3B1 (CD94). In addition, a number of isotype-matched mAb controls were also used. Immunofluorescent labeling and analysis was carried out as described [ 161.
3 Results
3.1 CD56' cells in blood in pregnancy
Comparison of CD56' cells in peripheral blood of 13 preg- nant women with 15 nonpregnant individuals (7 males and 8 females) showed no significant difference, with the range
'0 2 L, -
3 C L,
5.133 5.133
of CD56' cells in pregnant women being 10-34% (mean 19 YO) and in nonpregnant individuals 4.3-43 % (mean 22 YO) (p > 0.38). There was also no significant difference in the proportion of CD56b"gh' cells in male (range 0.3-1.3 %, mean 0.8 YO) and nonpregnant female (range 0.5-1.8 %, mean 0.9 YO) peripheral blood (p > 0.49). Interestingly, the CD56b"gh' subset was increased in preg- nant women (1-5 YO, mean 2.3 %) compared to nonpreg- nant females and in males (0.3-1.8%, mean 0.87%) (p < 0.0003). No significant difference was observed in the CD56dim population (p > 0.21).
3.2 Expression of KIR on blood and decidual CD56' cells
The KIR expression on peripheral blood NK cells was ana- lyzed for both the main CD56dim and the small CD56b"gh' subsets. Our results agree with those of others (Figs. 1 and 2). An important observation'was that the small CD56bngh' subset was found not to express KIR in any of the individu- als examined (Fig. 1).
To determine whether these KIR receptors for class I molecules are expressed by uterine tissue NK cells, lym- phocytes were isolated from the decidua of 19 women (gestational age 8-12 weeks). Decidual NK cells express receptors for HLA-A, -B, and -C gene products (Figs. 1 and 2). Similar to the results described for NK cells in blood, there are considerable differences between individ- uals, both in the percentage of cells expressing a particular
B l d CD56' cells Decidual CDW cells
51 C L,
I 23% 1
Figure 1. Flow cytometric analysis of expression of KIR by CD56+ cells from a single sample of peripheral blood leukocytes paired with decidual lymphocytes from the same individual. In both cases, anti-EGFR was used as a negative control to set quadrants defining posi- tive staining. The figure in the upper right quadrant shows the percentage of CD56' cells in the sample expressing the various KIR, while the figure in parentheses is the relative linear fluorescence intensity (RLF) for staining.
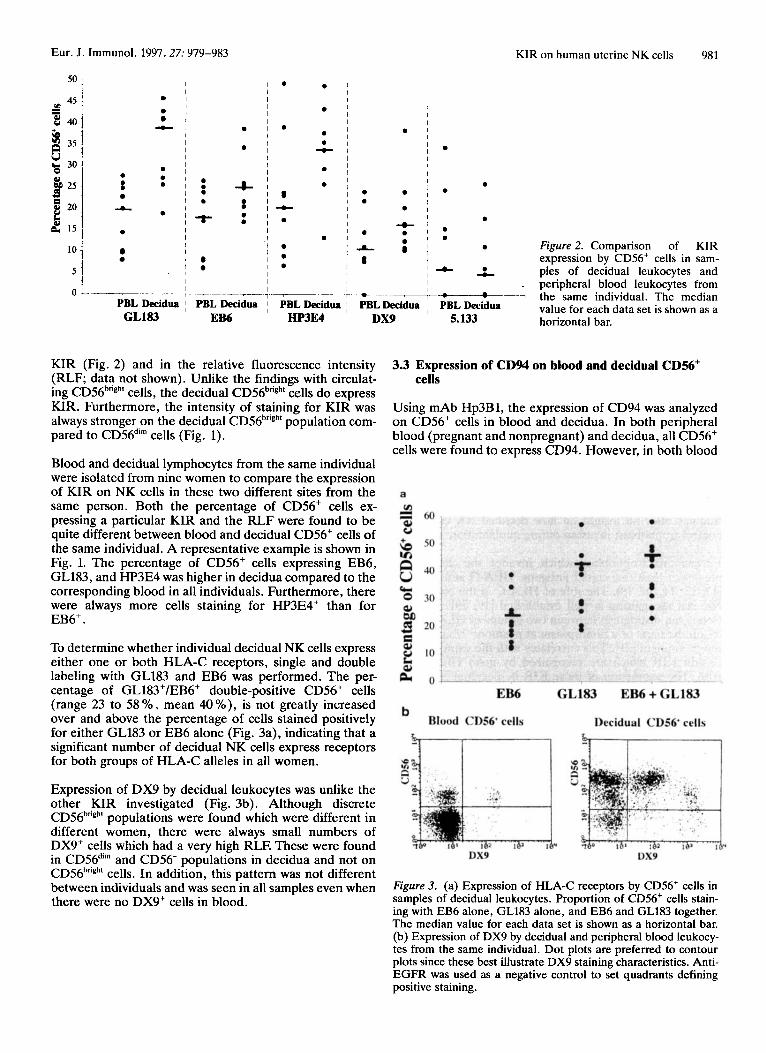
Eur. J. Immunol. 1997.27: 979-983 KIR on human uterine NK cells 981
50
I
0 i --
I 1
I m i
m i * I
- 1
1
* I
, -c I
0 I I
@ I
r ~ - . __ PBL Decidua 1
GL183
I
m i
I
I @ I
I
I
PBLDecidua I E M
~ _ - . + -- !.--*--- PBL Decidua ~ PBL Decidua I PBL Decidua
HP3E4 DX9 5.133
Figure 2. Comparison of KIR expression by CDS6+ cells in sam- ples of decidual leukocytes and
. peripheral blood leukocytes from 7 the same individual. The median
value for each data set is shown as a horizontal bar.
KIR (Fig. 2) and in the relative fluorescence intensity (RLF; data not shown). Unlike the findings with circulat- ing CD56'ngh' cells, the decidual CD56b"gh' cells do express KIR. Furthermore, the intensity of staining for KIR was always stronger on the decidual CD56hTigh' population com- pared to CD56di" cells (Fig. 1).
Blood and decidual lymphocytes from the same individual were isolated from nine women to compare the expression of KIR on NK cells in these two different sites from the same person. Both the percentage of CD56+ cells ex- pressing a particular KIR and the RLF were found to be quite different between blood and decidual CD56' cells of the same individual. A representative example is shown in Fig. 1. The percentage of CD56' cells expressing EB6, GL183, and HP3E4 was higher in decidua compared to the corresponding blood in all individuals. Furthermore, there were always more cells staining for HP3E4' than for EB6+.
3.3 Expression of CD94 on blood and decidual CD56' cells
Using mAb Hp3B1, the expression of CD94 was analyzed on CD56' cells in blood and decidua. In both peripheral blood (pregnant and nonpregnant) and decidua, all CD56' cells were found to express CD94. However, in both blood
To determine whether individual decidual NK cells express either one or both HLA-C receptors, single and double labeling with GL183 and EB6 was performed. The per- centage of GL183'/EB6+ double-positive CD56' cells (range 23 to 58 YO, mean 40 YO), is not greatly increased over and above the percentage of cells stained positively for either GL183 or EB6 alone (Fig. 3a), indicating that a significant number of decidual NK cells express receptors for both groups of HLA-C alleles in all women.
Expression of DX9 by decidual leukocytes was unlike the other KIR investigated (Fig. 3b). Although discrete CD56'ngh' populations were found which were different in different women, there were always small numbers of DX9+ cells which had a very high RLE These were found in CD56di" and CD56- populations in decidua and not on CD56'ngh' cells. In addition, this pattern was not different between individuals and was seen in all samples even when there were no DX9' cells in blood.
Figure 3. (a) Expression of HLA-C receptors by CDS6' cells in samples of decidual leukocytes. Proportion of CDS6' cells stain- ing with EB6 alone, GL183 alone, and EB6 and GL183 together. The median value for each data set is shown as a horizontal bar. (b) Expression of DX9 by decidual and peripheral blood leukocy- tes from the same individual. Dot plots are preferred to contour plots since these best illustrate DX9 staining characteristics. Anti- EGFR was used as a negative control to set quadrants defining positive staining.

982 S. Verma et al. Eur. J. Immunol. 1997.27: 979-983
and decidua in all CD56bngh' cells, the RLF of CD94 was at least twice as high as that on CD56di" cells (data not shown).
3.4 Expression of NK receptors on blood and decidual CD3' cells
Considering CD3' cells in peripheral blood, our results agree with those of others in finding expression of KIR on a minor subset of CD3' cells. When CD3' cells from the corresponding decidua from the same individual were ana- lyzed using 5.133, GL183, EB6, and CD94, they showed the same results as CD3' cells in blood. However, n o HP3E4' decidual CD3' Tcells were ever found despite the presence of HP3E4+ CD3' cells in peripheral blood of the same individual.
4 Discussion
This study shows that the distinctive tissue CD56h'igh' NK cells which populate the uterine mucosa at implantation and during early pregnancy express KIR for polymorphic class I HLA molecules. Using a panel of mAb directed at several members of the KIR family, we have shown by analyzing the decidual NK cells from individual pregnant women that there is a distinct but overlapping NK cell receptor repertoire for each woman. Furthermore, the pat- tern of NK receptors expressed is different between blood and decidual NK cells from the same individual. These findings offer an insight on how decidual NK cells may control trophoblast invasion during implantation.
Extravillous trophoblast which invades the uterus ex- presses at least two class I antigens, HLA-G and HLA-C [12, 13, 14, 15, 181. The role of HLA-C in the context of NK cell recognition is well established [19, 201. NK cell clones can distinguish between two groups of HLA-C alle- les on the basis of a dimorphism at positions 77 and 80 on the a1 domain [21]. This correlates with the expression of certain KTR which are recognized by mAb GL183 and EB6. The expression of these KIR by decidual NK cells, therefore, could provide a basis for maternal NK cell rec- ognition of HLA-C on fetal trophoblast. Interestingly, the proportion of NK cells expressing receptors for HLA-C alleles was higher in decidua than in peripheral blood from the same individual. Perhaps decidual NK cells are more biased towards HLA-C recognition than blood NK cells. In addition, comparing the data of singly labeled EB6' or GL183' cells with those double labeled with these anti- bodies, it is clear that there are appreciable numbers of EB6' GL183' double-positive NK cells in the decidua of all women. These NK cells will recognize all HLA-C alle- les, and such cells are known to be unable to lyse any allo- geneic cells [22]. This could be a mechanism for trophob- last survival and means that the paternal HLA-C allele expressed by trophoblast is perhaps of less importance than it might intuitively appear to be.
HLA-G can partially inhibit cytolysis of both decidual and blood polyclonal NK cell populations as well as CD3- deci- dual clones [23, 241. Two recent reports suggest that the HLA-G receptor(s) may also belong to the KIR family ([25] and Munz et al., submitted), but it cannot be ruled
out that HLA-G may be a target ligand for other NK receptor families. Indeed, - 50 'YO of decidual NK cells are unstained for both GL183 and EB6. However, all decidual CD56hr'gh' cells are also CD94b'igh'. CD94 is a member of the C-type lectin superfamily encoding type I1 membrane glycoproteins which is preferentially expressed by NK cells [26,27] in association with another NK lectin, NKG2, and is implicated in HLA recognition by these cells [28, 291. HLA-A, -B, and -C genes can all inhibit killing by CD94hr'gh' NK clones, but the exact HLA ligands of CD94 are still unknown and may possibly include HLA-G [29]. The CD94'ngh' expression by decidual CD56bngh' cells can, therefore, provide an alternative mechanism for GL183-, EB6- NK cells to interact with trophoblast HLA class I molecules.
A proportion of decidual NK cells express KIR which have been shown to bind HLA-A or HLA-B alleles (DX9' and 5.133' cells). The pattern of DX9 staining of decidual lym- phocytes was unusual. While different discrete proportions of DX9' CD56b"gh' cells were present in different individu- als, all women had a few CD56di" and CD56- cells staining at high intensity with DX9. These findings were never seen with peripheral blood lymphocytes and their significance is unknown.
Our present finding that the receptor repertoire of NK cells from blood and decidua is different within each indi- vidual is interesting in the light of the previous demonstra- tion that NKBl+ expression is genetically regulated [lo]. This dichotomy raises the question as to the developmen- tal relationship of the blood (CD56hrigh' and CD56di") and decidual CD56hngh' cells to each other [6].
Whatever the explanation for the differences found between blood and decidual NK cells, our observations indicate that the decidual NK cell repertoire cannot be determined by analysis of the corresponding peripheral blood NK cells from the same individual. This precludes any prospective studies attempting to link pregnancy out- come, and in particular disorders such as pre-eclampsia, which results from aberrant trophoblast invasion, to a par- ticular maternal decidual NK cell repertoire by monitoring peripheral blood NK cells.
KIR have been detected on a minor subset of T cells in peripheral blood where they have been shown to affect cytokine production and cytolysis [30-331. Although CD3' T cells represent only about 10 'YO of decidual leuko- cytes, a minor subset of these similarly did express KIR. The exception was that decidual CD3' HP3E4' cells were never found. PHA-stimulated T cell blasts from blood were also reported to be unreactive with HP3E4 [31]. There are other features of decidual CD3' T cells which indicate they may be polyclonally activated [6].
KIR are characterized by two immune receptor tyrosine- based inhibitory motifs (ITIM) in the cytoplasmic tail. Alternative forms have also been described as killer cell activatory receptors (KAR), which are stimulatory recep- tors [34-361. In a parallel study, we detected both activa- tory and inhibitory NK receptors by RT-PCR of cDNA iso- lated from decidual NK cells (Hiby et al., submitted). Thus, the outcome of maternal decidual NK cell interac- tion with fetal trophoblast in vivo is likely to be complex

Eur. J. Immunol. 1997.27: 979-983 KIR on human uterine NK cells 983
a n d will be dependent on t h e balance between positive a n d negative signals generated.
We thank our obstetric colleagues and staff at Addenbrookek Hos- pital, Cambridge, for collecting the placental material. We acknowledge financial support from the Medical Research Coun- cil, Tommy5 Campaign and the Special Programme of Research, Development and Research Training in Human Reproduction, World Health Organization. A. K . is in receipt of the Meres Senior Studentship for Medical Research at St. John’s College, Cum- bridge, and S. V k a Wellcome MB Ph. D Student. We are grateful for kind gifts of antibodies from Professor M . Ldpez-Botet, Dr. L. Lanier, Professor L. Moretta, and Dr. M. Colonna.
5 References
1
2
3
4
5
6
7 8 9
10
11
12
13
14
King, A., Balendran, N., Wooding, P., Carter, N. P. and Loke, Y. W., Dev. Immunol. 1991. I : 169. Bulmer, J. N., Morrison, L., Longfellow, M., Ritson, A. and Pace, D., Hum. Reprod. 1991. 6: 791. Nishikawa, K., Saito, S., Morii, T., Hamada, K., Ako, H., Narita, N., Ichijo, M., Kurahayashi, M. and Sugamura, K., Int. Immunol. 1991. 3: 743. King, A., Wellings, V., Gardner, L. and Loke, Y. W., Hum. lmmunol. 1989.24: 195. Bulmer, J. N., Johnson, P. M. and Bulmer, D., in Gill, T. J. and Wegmann, T. G. (Eds.), Immunoregulation and Fetal Sur- vival, Oxford University Press 1987, p. 111. Loke, Y. W. and King, A., Human Implantation: Cell Biology and Immunology, Cambridge University Press 1995. King, A. and Loke, Y. W., Immunol. Today 1991. 12: 432. Gumperz, J. E. and Parham, P., Nature 1995. 378: 245. Lanier, L. L. and Phillips, J. H., Immunol. Today 1996.17: 86. Gumperz, J. E., Valiante, N. M., Parham, P., Lanier, L. L. andTyan, D. , J. Exp. Med. 1996. 183: 1817. Vitale, M., Sivori, S., Pende, D., Augugliaro, R., Di Donato, C., Amoroso, A., Malnati, M., Botting, C., Moretta, L. and Moretta, A., Proc. Natl. Acad. Sci. USA 1996. 93: 1453. Kovats, A., Main, E. K., Librach, C., Stubblebine, M., Fisher, S. J. and DeMars, R., Science 1990.248: 220. Ellis, S. A., Palmer, M. S. and McMichael, A. J., J. Immunol. 1990. 144: 731. Ellis, S. A., Strachan, T., Palmer, M. S. and McMichael, A. J.,
17 Burrows, T. D., King, A. and Loke, Y. W., Cell. Immunol. 1993. 147: 81.
18 McMaster, M. T., Librach, C. L., Zhou, Y., Lim, K.-H., Janatpour, M. J., DeMars, R., Kovats, S. , Damsky, C. and Fisher, S. J., J. Immunol. 1995. 154: 3771.23.
19 Moretta, A., Vitale, M., Bottino, C., Orengo, A. M., Morelli, L., Augugliaro, R., Barbaresi, M., Ciccone, E. and Moretta, L., J. Exp. Med. 1993. 178: 597.
20 Colonna, M., Borsellino, G., Falco, M., Battista Ferrara, G. and Strominger, J. L., Proc. Natl. Acad. Sci. USA 1993. 90: 12000.
21 Colonna, M., Spies, T., Strominger, J . L., Ciccone, E., Moretta, A., Moretta, L., Pende, D. and Viale, O., Proc. Natl. Acad. Sci. USA 1992. 89: 7983.
22 Wale, M., Sivori, S., Pende, D., Moretta, L. and Moretta, A. , Proc. Natl. Acad. Sci. USA 1995. 92: 3536.
23 Chumbley, G., King, A., Robertson, K., Holmes, N. and Loke, Y. W., Cell. Immunol. 1994. 155: 312.
24 Deniz, G., Christmas, S. E., Brew, R. and Johnson, P. M., J. Immunol. 1994. 152: 4255.
25 Pazmany, L., Mandelboim, O., Vales-Gomez, M., Davis, D. M., Reyburn, H. T. and Strominger, J. L., Science 1996. 274: 792.
26 Aramburu, J., Balboa, M. A., Izquierdo, M. and Lopez- Botet, M., J. Immunol. 1991. 147: 714.
27 Chang, C., Rodriguez, A., Carretero, M., Lopez-Botet, M., Phillips, J. H. and Lanier, L. L., Eur. J. Immunol. 1995. 25: 2433.
28 Lazetic, S., Chang, C., Houchins, J . P., Lanier, L. L. and Phil- lips, J. H., J. Immunol. 1996. 157: 4741.
29 Phillips, J. H., Chang, C., Mattson, J., Gumperz, J. E., Par- ham, P. and Lanier, L. L., Immunity 1996. 5: 163.
30 Ferrini, S., Cambiaggi, A., Meazza, R., Sforzini, S., Marci- ano, S. , Mingari, M. C. and Moretta, L., Eur. J. Immunol. 1994.24: 2294.
31 Melero, I., Salmeron, A., Balboa, M. A. , Aramburu, J. and Lbpez-Botet, J., J . Immunol. 1994. 152: 1662.
32 Mingari, M. C., Vitale, C., Cambiaggi, A., Schiavetti, F., Melioli, G., Ferrini, S. and Poggi, A., Int. Immunol. 1995. 7: 697.
33 Phillips, J . H., Gumperz, J. E., Parham, P. and Lanier, L. L., Science 1995.268: 403.
34 Biassoni, R., Cantoni, C., Falco, M., Verdiani, S. , Bottino, C., Vitale, M., Conte, R., Poggi, A., Moretta, A . and Moretta, L., J . Exp. Med. 1996. 183: 645.
35 Moretta, A., Sivori, S., Vitale, M., Pende, D. , Morelli, L., J. Immunol. 1989. 142: 3281.
15 King, A., Boocock, C., Sharkey, A. M., Gardner, L., Beretta, A., Siccardi, A. G. and Loke, Y. W., J. Immunol. 1996. 156: 2068.
16 Jokhi, P., Chumbley, G., Gardner, L., King, A. and Lake, Y. W., Lab. Invest. 1993. 68: 308.
Augugliaro, R., Bottino, C. and Moretta, L., J . Exp. Med. 1995. 182: 875.
36 Bottino, c . , Sivon, S . , Vitale, M., Cantoni, C., Falco, M., Pende, D., Morelli, L., Augugliaro, R., Semenzato, G., Bias- soni, R., Moretta, L. and Moretta, A., Eur. J . Immunol. 1996. 26: 1816.