expression of fungal diacylglycerol acyltransferase2
TRANSCRIPT

Expression of Fungal diacylglycerol acyltransferase2Genes to Increase Kernel Oil in Maize[OA]
Janette Oakes, Doug Brackenridge, Ron Colletti, Maureen Daley, Deborah J. Hawkins, Hui Xiong,Jennifer Mai, Steve E. Screen, Dale Val, Kathryn Lardizabal, Ken Gruys, and Jill Deikman*
Monsanto Company, Davis, California 95616 (J.O., M.D., D.J.H., H.X., J.M., D.V., K.L., K.G., J.D.);Monsanto Company, Chesterfield, Missouri 63017 (D.B.); and Monsanto Company, St. Louis,Missouri 63167 (R.C., S.E.S.)
Maize (Zea mays) oil has high value but is only about 4% of the grain by weight. To increase kernel oil content, fungaldiacylglycerol acyltransferase2 (DGAT2) genes from Umbelopsis (formerly Mortierella) ramanniana and Neurospora crassa wereintroduced into maize using an embryo-enhanced promoter. The protein encoded by the N. crassa gene was longer than that ofU. ramanniana. It included 353 amino acids that aligned to the U. ramanniana DGAT2A protein and a 243-amino acid sequenceat the amino terminus that was unique to the N. crassa DGAT2 protein. Two forms of N. crassa DGAT2 were tested: thepredicted full-length protein (L-NcDGAT2) and a shorter form (S-NcDGAT2) that encoded just the sequences that sharehomology with the U. ramanniana protein. Expression of all three transgenes in maize resulted in small but statisticallysignificant increases in kernel oil. S-NcDGAT2 had the biggest impact on kernel oil, with a 26% (relative) increase in oil inkernels of the best events (inbred). Increases in kernel oil were also obtained in both conventional and high-oil hybrids, andgrain yield was not affected by expression of these fungal DGAT2 transgenes.
The demand for vegetable oil is increasing both forhuman consumption and more recently for use in themanufacture of biodiesel. One way to meet this de-mand without devoting additional land to agricultureis to increase the yield of oil per acre. Although oilmakes up only a small proportion of maize (Zea mays)grain (approximately 4%), maize oil is the second involume vegetable oil produced in the United States,after soybean (Glycine max) oil (Ash and Dohlman,2006). Maize kernel oil is composed primarily oftriacylglycerol (TAG) and is used as an energy sourceby the plant during seed germination (Tan andMorrison,1979). Most of the oil in a maize seed is found in theembryo, which is about 33% oil by weight (Watson,1987). Breeding approaches to increase maize kerneloil have had some success, but high kernel oil concen-tration has been linked to a decrease in grain yield(Dudley et al., 1977; Misevic and Alexander, 1989).Transgenic approaches to increasing oil in the maizeembryo have shown promise in allowing increased oilproduction without yield drag (Zheng et al., 2008;Shen et al., 2010). For example, overexpression of theembryo development transcription factor LEAFY COT-
YLEDON1 (ZmLEC1) in maize increased kernel oil byup to 48% (Shen et al., 2010). Unfortunately, thistranscription factor also produced negative pleiotropiceffects, including reduced seed germination and leafgrowth. However, overexpression of a transcriptionfactor downstream of ZmLEC1, maize WRINKLED1(ZmWRI1), resulted in similar increases in kernel oilbut without any detectable off types (Shen et al., 2010).ZmWRI1 appears to control genes involved in glycol-ysis and fatty acid biosynthesis but not oil biosynthe-sis, and it is hypothesized that additional oil increasesmay be obtained by combining overexpression of thistranscription factor with genes that have a more directeffect on oil biosynthesis.
Diacylglycerol acyltransferase (DGAT) catalyzesthe final step of TAG biosynthesis by transferring anacyl group from acyl-CoA to the sn-3 position of 1,2-diacylglycerol (DAG; Kennedy, 1961). Two classes ofDGAT genes have been identified in eukaryotes,DGAT1 and DGAT2 (Lardizabal et al., 2001; Lungand Weselake, 2006). DGAT1 is related to the acyl-CoA:cholesterol acyltransferase family (Cases et al.,1998). DGAT2 has little sequence similarity to DGAT1and is part of a family of proteins that transfer acylgroups from CoA to neutral lipids (Lardizabal et al.,2001; Turkish and Sturley, 2007). Whether these twoclasses of DGATs have redundant or specific functionsin TAG biosynthesis in plants is not clear. They mayfunction at different times during development orprovide different fatty acid specificities (Lung andWeselake, 2006; Li et al., 2010a, 2010b). Ectopic expres-sion of DGAT transgenes from various plant and
* Corresponding author; e-mail [email protected] author responsible for distribution of materials integral to the
findings presented in this article in accordance with the policydescribed in the Instructions for Authors (www.plantphysiol.org) is:Jill Deikman ([email protected]).
[OA] Open Access articles can be viewed online without a sub-scription.
www.plantphysiol.org/cgi/doi/10.1104/pp.110.167676
1146 Plant Physiology�, March 2011, Vol. 155, pp. 1146–1157, www.plantphysiol.org � 2011 American Society of Plant Biologists www.plantphysiol.orgon April 12, 2019 - Published by Downloaded from
Copyright © 2011 American Society of Plant Biologists. All rights reserved.

fungal species has been shown to increase seed oilcontent in Arabidopsis (Arabidopsis thaliana; Jako et al.,2001), soybean (Lardizabal et al., 2008), and maize(Zheng et al., 2008).In addition to DGAT, other enzymes have also been
shown to be involved in TAG biosynthesis in plants,including phospholipid:diacylglycerol acyltransferase(PDAT) and DAG transacylase (Lung and Weselake,2006).Mutation of DGAT1 in Arabidopsis reduced seed oil
content 25% to 35% (relative to the wild type), support-ing a role for DGAT1 in oil deposition during seeddevelopment but also suggesting the participation ofother acyltransferases or transacylases (Routaboul et al.,1999; Zou et al., 1999). Work with Arabidopsis mutantsestablished that PDAT1 participates in the accumula-tion of TAG in Arabidopsis seeds and suggested thatDGAT1 and PDAT1 activities may account for all TAGbiosynthesis in Arabidopsis seeds (Zhang et al., 2009).DGAT2 appears to play an important role in oil
accumulation in seeds of other species. In both castorbean (Ricinus communis; Kroon et al., 2006) and tungtree (Vernicia fordii; Shockey et al., 2006), DGAT2 ismore highly expressed in seeds than DGAT1. Further-more, in tung tree, the DGAT2 form was shown tohave greater selectivity for eleostearic acid, an unusualfatty acid found at high concentration in tung treeseeds (Kroon et al., 2006). A survey of the expressionpatterns of DGAT1, DGAT2, and PDAT genes in avariety of species with seeds containing epoxy andhydroxyl fatty acids found that while all three en-zymes are important in oil accumulation in thesespecies, DGAT2 and PDAT are more highly expressedin developing seeds of these species than in Arabi-dopsis or soybean, supporting the concept that PDATand DGAT2 are involved in the accumulation of thesespecial fatty acids (Li et al., 2010b).In maize, an allele of a DGAT1 gene (DGAT1-2) was
shown to be responsible for a high-oil quantitative traitlocus on chromosome 6 (Zheng et al., 2008). Cloning ofthe high-oil DGAT1-2 allele revealed that a Phe ispresent at position 469 of the encoded protein that isabsent in DGAT1-2 from maize lines with oil contenttypical of commercial lines (normal-oil content). Thehigh-oil DGAT1-2 protein was shown to have greaterenzymatic activity comparedwith the normal-oil form.Overexpression of both the high-oil and normal-oilDGAT1-2 genes in transgenic maize was performedusing an embryo-enhanced promoter, and the trans-genic lines had increased kernel oil and increasedlevels of kernel oleic acid, a trait also associated withthe high-oil quantitative trait locus on chromosome 6(Zheng et al., 2008). Maize plants with the high-oilallele had larger increases in oil content and oleic acidlevels than lines that contained the normal-oil alleletransgene. The line with the largest increase had a41% (relative) increase in seed oil content and a 107%(relative) increase in oleic acid.Overexpression in soybean of a codon-optimized
version of the DGAT2A gene from the fungus Umbe-
lopsis ramanniana using a seed-specific promoter re-sulted in an increase in seed oil content of 1.5% byweight (an approximately 7.5% relative increase) with-out any effect on grain yield (Lardizabal et al., 2008).We introduced this same UrDGAT2A gene into maizeusing an embryo-enhanced promoter and demon-strated a small but significant increase in kernel oilof up to 0.7% by weight (a relative increase of 19%). Wealso cloned and overexpressed a DGAT2 gene fromNeurospora crassa and observed slightly higher DGATenzyme activities and oil increases up to 0.9% byweight (a 26% relative increase). We observed no effecton hybrid grain yield from overexpression of thesetransgenes. Importantly, oil increases were obtainedfrom overexpression of the fungal DGATs in high-oilhybrids, indicating the possibility of further enhancingoil biosynthesis in high-oil germplasm. An unexpectedseedling phenotype was observed in some of thetransgenic events and may provide insight into otherroles of DGAT proteins during development.
RESULTS
Expression of UrDGAT2A or NcDGAT2 in Seed IncreasedDGAT Activity and Kernel Oil Content
The identification of fungal DGAT2 sequences waspreviously reported for U. ramanniana (Lardizabalet al., 2001) and N. crassa (Lardizabal et al., 2004). TheU. ramanniana and N. crassa predicted protein se-quences were similar in length and shared 45% aminoacid identity and 66% amino acid similarity (Fig. 1). Asubsequent BLAST search of public databases withthe N. crassa amino acid sequence identified a sequenceannotated as “hypothetical protein NCU02665 [Neuros-pora crassa OR74A]” (National Center for Biotechnol-ogy Information [NCBI] accession no. 85118420;Galagan et al., 2003) that was 100% identical alongthe length of the originally identified NcDGAT2 se-quence. However, the hypothetical protein included anadditional 243 amino acids at the N terminus (Fig. 1).To learn more about the additional N-terminal part ofthe hypothetical N. crassa protein, we conducted aBLASTsearch of GenBankwith just this 243-amino acidfragment. The only sequences that were identifiedwereannotated “hypothetical protein” or “related to diacyl-glycerol acyl transferase” from N. crassa. Analysis ofthe N-terminal sequences using the Protean programindicated that they are hydrophilic (data not shown).The portion of the NcDGAT2 predicted protein thataligns with the UrDGAT2A sequence begins with asmall hydrophilic tail in front of a large hydrophobicdomain that is conserved structurally in the DGAT2family and probably anchors the protein to the mem-brane (data not shown; Shockey et al., 2006). Because itwas not clearwhether the predictedN-terminal portionof the NcDGAT sequence is really part of the native N.crassa protein, we decided to test both forms in maize.The longer NcDGAT2 coding sequence will be re-
Fungal DGAT2 Increases Maize Oil
Plant Physiol. Vol. 155, 2011 1147 www.plantphysiol.orgon April 12, 2019 - Published by Downloaded from
Copyright © 2011 American Society of Plant Biologists. All rights reserved.

ferred to as L-NcDGAT2, and the shorter form will bereferred to as S-NcDGAT2.
To determine whether expression of a fungalDGAT2can increase maize kernel oil, we constructed planttransformation vectors with the UrDGAT2A codingsequence and the two different lengths of the NcDGAT2protein-coding sequences. All constructs included anembryo-enhanced promoter, Per1, from barley (Hordeumvulgare; Stacy et al., 1996). Maize plants were trans-formed with these sequences, the regenerated trans-formed plants (R0 stage) were self-pollinated, and earswere harvested at maturity. Oil and fresh weight weredetermined for 36 kernels per ear. Kernels were thendissected into embryo and endosperm, and oil andweight were determined for the separate tissues.DNA was extracted from each embryo and analyzedby PCR to determine presence or absence of thetransgene. Oil and weight were compared for kernelson the same ear with or without the transgene. Forevents transformed with the UrDGAT2A transgene,nine of 11 events had a statistically significant (P, 0.05)increase in kernel oil ranging from +0.4% to +0.7% (byweight) oil (Fig. 2), a relative change of 9% to 19%. Onlyfour of seven events containing the long form of theNcDGAT2 transgene tested in this generation had anincrease in kernel oil, from +0.3% to +0.7% by weight(relative change of 8%–16%). However, the shortNcDGAT2 transgene produced increased kernel oilin 13 of 15 events, with a range of +0.4% to +0.9%
(relative change of 13%–26%). A construct-level anal-ysis of the dissected kernel data shows that each of theconstructs increased oil predominantly in the embryo(Table I) from about 12% (relative) for UrDGAT2A toabout 17% for S-NcDGAT2, as expected, since thetransgene was expressed using an embryo-enhancedpromoter. A small but significant increase in oil in theendosperm was observed for each construct and isprobably due to activity of the Per1 promoter in thealeurone. None of the constructs produced a signifi-cant change in total kernel weight or in the weight ofthe embryo or endosperm. The differences in oil andweight values for controls of the different constructsare likely due to the fact that the plants from eachconstruct were grown in the greenhouse at differenttimes. Environmental conditions are known to havesome influence on kernel growth and composition (Valet al., 2009).
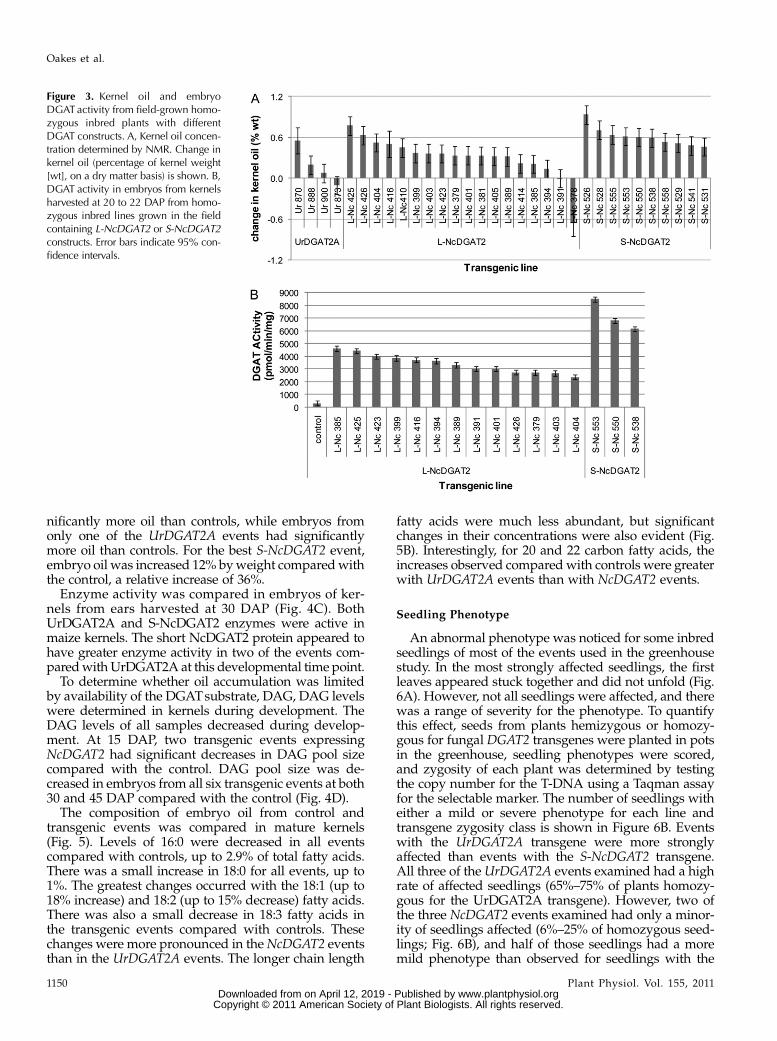
To more directly compare effects of the three con-structs, homozygous lines were identified from mul-tiple events for each construct and were grown in afield in Hawaii. Plants were self-pollinated, and earswere harvested at 22 to 24 d after pollination (DAP) orat maturity. In this experiment, only two of the fourUrDGAT2A events tested hadmore oil than the controlat maturity, and the best event increased oil by 0.6% byweight, resulting in a relative increase of 11% (Fig. 3A).Sixteen of 18 events containing L-NcDGAT2 increasedkernel oil, from +0.1% to +0.8%, giving a relative
Figure 1. Alignment of NcDGAT (Nc) and UrDGAT (Ur) proteins. Identical residues are highlighted in black. The arrow indicatesthe start of the short NcDGAT protein.
Oakes et al.
1148 Plant Physiol. Vol. 155, 2011 www.plantphysiol.orgon April 12, 2019 - Published by Downloaded from
Copyright © 2011 American Society of Plant Biologists. All rights reserved.
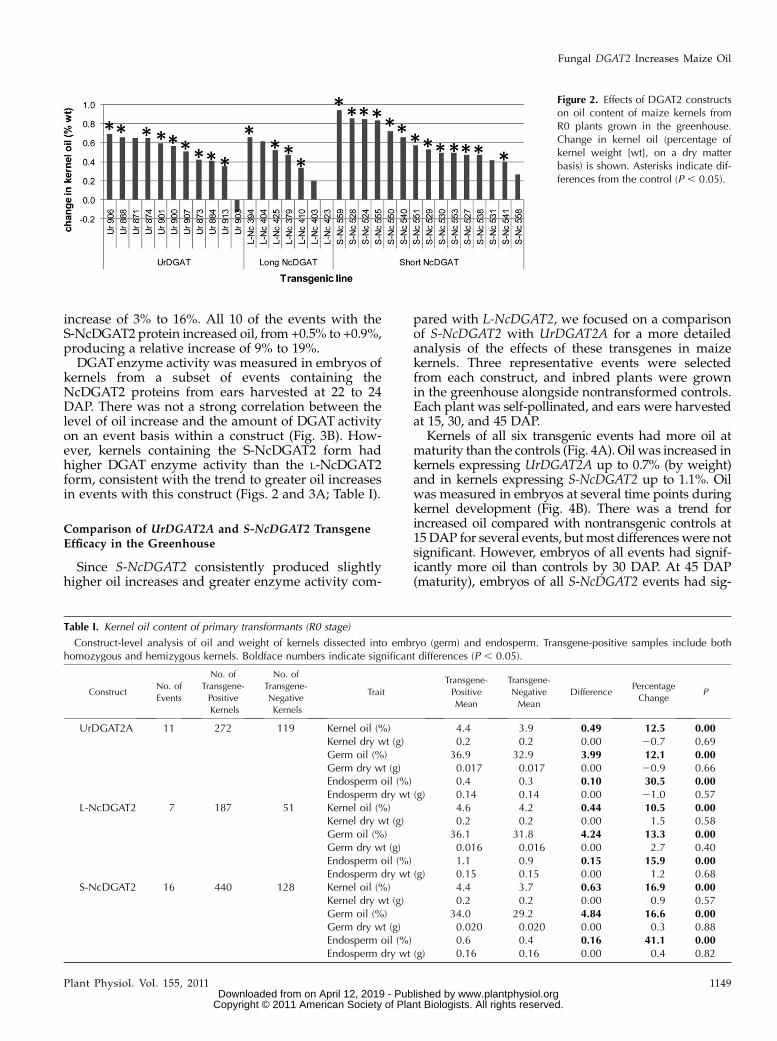
increase of 3% to 16%. All 10 of the events with theS-NcDGAT2 protein increased oil, from +0.5% to +0.9%,producing a relative increase of 9% to 19%.DGATenzyme activity was measured in embryos of
kernels from a subset of events containing theNcDGAT2 proteins from ears harvested at 22 to 24DAP. There was not a strong correlation between thelevel of oil increase and the amount of DGAT activityon an event basis within a construct (Fig. 3B). How-ever, kernels containing the S-NcDGAT2 form hadhigher DGAT enzyme activity than the L-NcDGAT2form, consistent with the trend to greater oil increasesin events with this construct (Figs. 2 and 3A; Table I).
Comparison of UrDGAT2A and S-NcDGAT2 Transgene
Efficacy in the Greenhouse
Since S-NcDGAT2 consistently produced slightlyhigher oil increases and greater enzyme activity com-
pared with L-NcDGAT2, we focused on a comparisonof S-NcDGAT2 with UrDGAT2A for a more detailedanalysis of the effects of these transgenes in maizekernels. Three representative events were selectedfrom each construct, and inbred plants were grownin the greenhouse alongside nontransformed controls.Each plant was self-pollinated, and ears were harvestedat 15, 30, and 45 DAP.
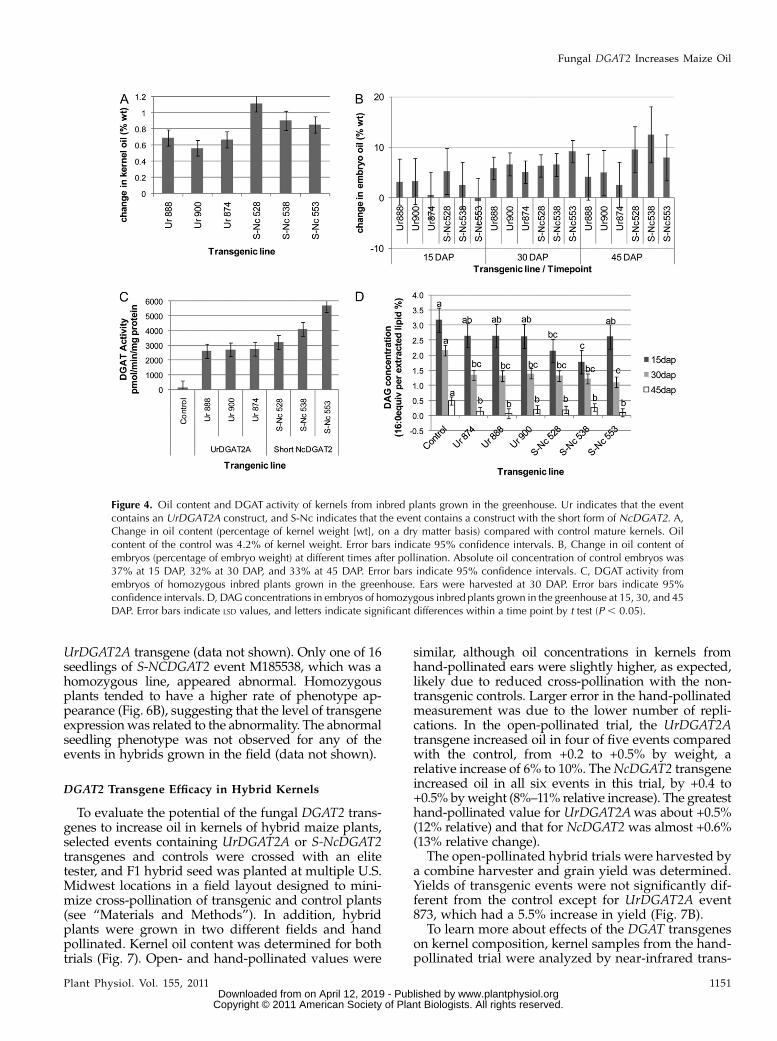
Kernels of all six transgenic events had more oil atmaturity than the controls (Fig. 4A). Oil was increased inkernels expressing UrDGAT2A up to 0.7% (by weight)and in kernels expressing S-NcDGAT2 up to 1.1%. Oilwas measured in embryos at several time points duringkernel development (Fig. 4B). There was a trend forincreased oil compared with nontransgenic controls at15 DAP for several events, butmost differences were notsignificant. However, embryos of all events had signif-icantly more oil than controls by 30 DAP. At 45 DAP(maturity), embryos of all S-NcDGAT2 events had sig-
Figure 2. Effects of DGAT2 constructson oil content of maize kernels fromR0 plants grown in the greenhouse.Change in kernel oil (percentage ofkernel weight [wt], on a dry matterbasis) is shown. Asterisks indicate dif-ferences from the control (P , 0.05).
Table I. Kernel oil content of primary transformants (R0 stage)
Construct-level analysis of oil and weight of kernels dissected into embryo (germ) and endosperm. Transgene-positive samples include bothhomozygous and hemizygous kernels. Boldface numbers indicate significant differences (P , 0.05).
ConstructNo. of
Events
No. of
Transgene-
Positive
Kernels
No. of
Transgene-
Negative
Kernels
Trait
Transgene-
Positive
Mean
Transgene-
Negative
Mean
DifferencePercentage
ChangeP
UrDGAT2A 11 272 119 Kernel oil (%) 4.4 3.9 0.49 12.5 0.00Kernel dry wt (g) 0.2 0.2 0.00 20.7 0.69Germ oil (%) 36.9 32.9 3.99 12.1 0.00Germ dry wt (g) 0.017 0.017 0.00 20.9 0.66Endosperm oil (%) 0.4 0.3 0.10 30.5 0.00Endosperm dry wt (g) 0.14 0.14 0.00 21.0 0.57
L-NcDGAT2 7 187 51 Kernel oil (%) 4.6 4.2 0.44 10.5 0.00Kernel dry wt (g) 0.2 0.2 0.00 1.5 0.58Germ oil (%) 36.1 31.8 4.24 13.3 0.00Germ dry wt (g) 0.016 0.016 0.00 2.7 0.40Endosperm oil (%) 1.1 0.9 0.15 15.9 0.00Endosperm dry wt (g) 0.15 0.15 0.00 1.2 0.68
S-NcDGAT2 16 440 128 Kernel oil (%) 4.4 3.7 0.63 16.9 0.00Kernel dry wt (g) 0.2 0.2 0.00 0.9 0.57Germ oil (%) 34.0 29.2 4.84 16.6 0.00Germ dry wt (g) 0.020 0.020 0.00 0.3 0.88Endosperm oil (%) 0.6 0.4 0.16 41.1 0.00Endosperm dry wt (g) 0.16 0.16 0.00 0.4 0.82
Fungal DGAT2 Increases Maize Oil
Plant Physiol. Vol. 155, 2011 1149 www.plantphysiol.orgon April 12, 2019 - Published by Downloaded from
Copyright © 2011 American Society of Plant Biologists. All rights reserved.
nificantly more oil than controls, while embryos fromonly one of the UrDGAT2A events had significantlymore oil than controls. For the best S-NcDGAT2 event,embryo oil was increased 12% byweight comparedwiththe control, a relative increase of 36%.
Enzyme activity was compared in embryos of ker-nels from ears harvested at 30 DAP (Fig. 4C). BothUrDGAT2A and S-NcDGAT2 enzymes were active inmaize kernels. The short NcDGAT2 protein appeared tohave greater enzyme activity in two of the events com-paredwithUrDGAT2A at this developmental time point.
To determine whether oil accumulation was limitedby availability of the DGATsubstrate, DAG, DAG levelswere determined in kernels during development. TheDAG levels of all samples decreased during develop-ment. At 15 DAP, two transgenic events expressingNcDGAT2 had significant decreases in DAG pool sizecompared with the control. DAG pool size was de-creased in embryos from all six transgenic events at both30 and 45 DAP compared with the control (Fig. 4D).
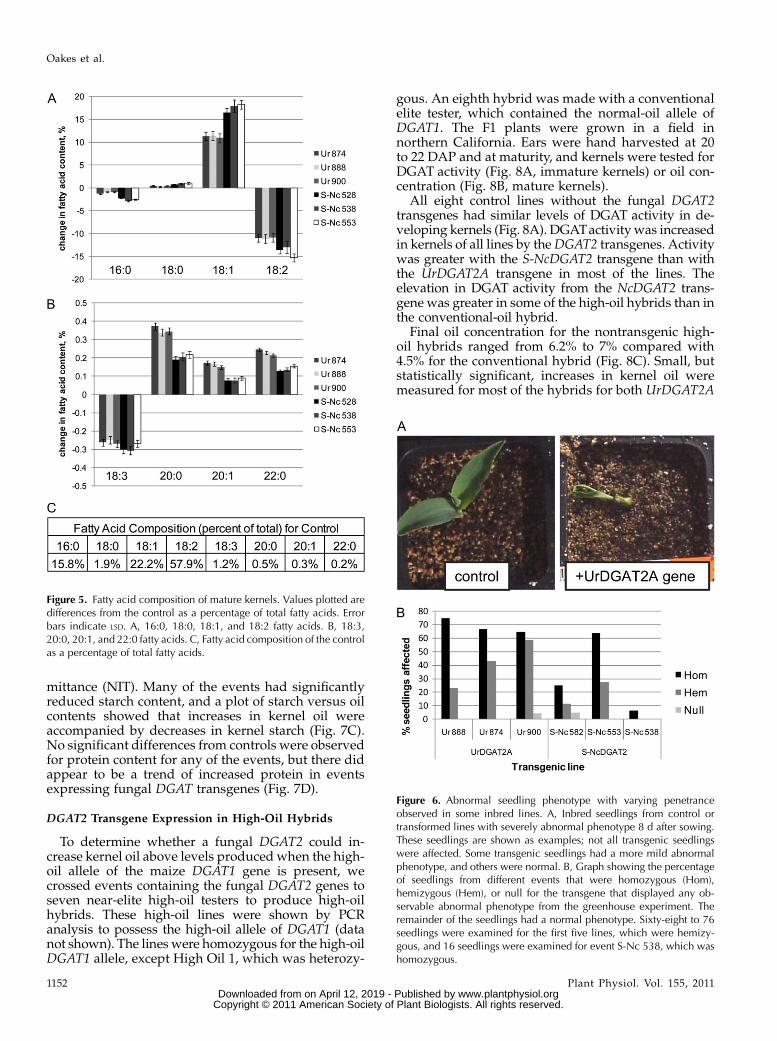
The composition of embryo oil from control andtransgenic events was compared in mature kernels(Fig. 5). Levels of 16:0 were decreased in all eventscompared with controls, up to 2.9% of total fatty acids.There was a small increase in 18:0 for all events, up to1%. The greatest changes occurred with the 18:1 (up to18% increase) and 18:2 (up to 15% decrease) fatty acids.There was also a small decrease in 18:3 fatty acids inthe transgenic events compared with controls. Thesechanges were more pronounced in theNcDGAT2 eventsthan in the UrDGAT2A events. The longer chain length
fatty acids were much less abundant, but significantchanges in their concentrations were also evident (Fig.5B). Interestingly, for 20 and 22 carbon fatty acids, theincreases observed comparedwith controls were greaterwith UrDGAT2A events than with NcDGAT2 events.
Seedling Phenotype
An abnormal phenotype was noticed for some inbredseedlings of most of the events used in the greenhousestudy. In the most strongly affected seedlings, the firstleaves appeared stuck together and did not unfold (Fig.6A). However, not all seedlings were affected, and therewas a range of severity for the phenotype. To quantifythis effect, seeds from plants hemizygous or homozy-gous for fungalDGAT2 transgenes were planted in potsin the greenhouse, seedling phenotypes were scored,and zygosity of each plant was determined by testingthe copy number for the T-DNA using a Taqman assayfor the selectable marker. The number of seedlings witheither a mild or severe phenotype for each line andtransgene zygosity class is shown in Figure 6B. Eventswith the UrDGAT2A transgene were more stronglyaffected than events with the S-NcDGAT2 transgene.All three of theUrDGAT2A events examined had a highrate of affected seedlings (65%–75% of plants homozy-gous for the UrDGAT2A transgene). However, two ofthe three NcDGAT2 events examined had only a minor-ity of seedlings affected (6%–25% of homozygous seed-lings; Fig. 6B), and half of those seedlings had a moremild phenotype than observed for seedlings with the
Figure 3. Kernel oil and embryoDGATactivity from field-grown homo-zygous inbred plants with differentDGAT constructs. A, Kernel oil concen-tration determined by NMR. Change inkernel oil (percentage of kernel weight[wt], on a dry matter basis) is shown. B,DGAT activity in embryos from kernelsharvested at 20 to 22 DAP from homo-zygous inbred lines grown in the fieldcontaining L-NcDGAT2 or S-NcDGAT2constructs. Error bars indicate 95% con-fidence intervals.
Oakes et al.
1150 Plant Physiol. Vol. 155, 2011 www.plantphysiol.orgon April 12, 2019 - Published by Downloaded from
Copyright © 2011 American Society of Plant Biologists. All rights reserved.
UrDGAT2A transgene (data not shown). Only one of 16seedlings of S-NCDGAT2 event M185538, which was ahomozygous line, appeared abnormal. Homozygousplants tended to have a higher rate of phenotype ap-pearance (Fig. 6B), suggesting that the level of transgeneexpressionwas related to the abnormality. The abnormalseedling phenotype was not observed for any of theevents in hybrids grown in the field (data not shown).
DGAT2 Transgene Efficacy in Hybrid Kernels
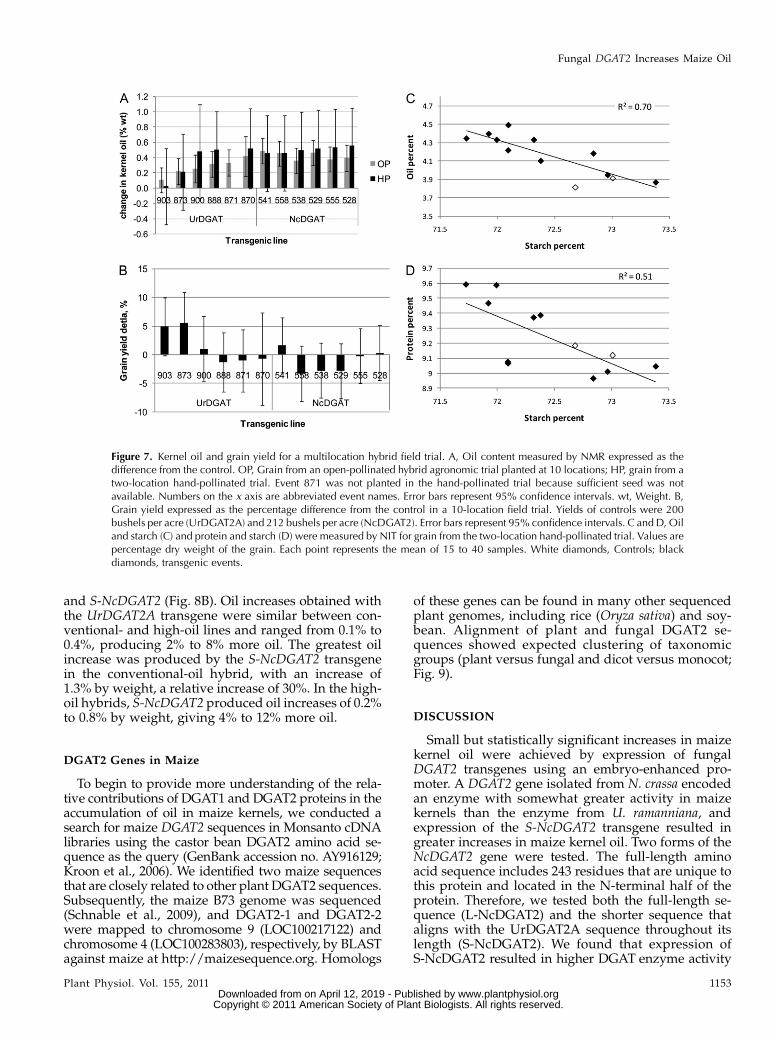
To evaluate the potential of the fungal DGAT2 trans-genes to increase oil in kernels of hybrid maize plants,selected events containing UrDGAT2A or S-NcDGAT2transgenes and controls were crossed with an elitetester, and F1 hybrid seed was planted at multiple U.S.Midwest locations in a field layout designed to mini-mize cross-pollination of transgenic and control plants(see “Materials and Methods”). In addition, hybridplants were grown in two different fields and handpollinated. Kernel oil content was determined for bothtrials (Fig. 7). Open- and hand-pollinated values were
similar, although oil concentrations in kernels fromhand-pollinated ears were slightly higher, as expected,likely due to reduced cross-pollination with the non-transgenic controls. Larger error in the hand-pollinatedmeasurement was due to the lower number of repli-cations. In the open-pollinated trial, the UrDGAT2Atransgene increased oil in four of five events comparedwith the control, from +0.2 to +0.5% by weight, arelative increase of 6% to 10%. TheNcDGAT2 transgeneincreased oil in all six events in this trial, by +0.4 to+0.5% byweight (8%–11% relative increase). The greatesthand-pollinated value for UrDGAT2Awas about +0.5%(12% relative) and that for NcDGAT2 was almost +0.6%(13% relative change).
The open-pollinated hybrid trials were harvested bya combine harvester and grain yield was determined.Yields of transgenic events were not significantly dif-ferent from the control except for UrDGAT2A event873, which had a 5.5% increase in yield (Fig. 7B).
To learn more about effects of the DGAT transgeneson kernel composition, kernel samples from the hand-pollinated trial were analyzed by near-infrared trans-
Figure 4. Oil content and DGAT activity of kernels from inbred plants grown in the greenhouse. Ur indicates that the eventcontains an UrDGAT2A construct, and S-Nc indicates that the event contains a construct with the short form of NcDGAT2. A,Change in oil content (percentage of kernel weight [wt], on a dry matter basis) compared with control mature kernels. Oilcontent of the control was 4.2% of kernel weight. Error bars indicate 95% confidence intervals. B, Change in oil content ofembryos (percentage of embryo weight) at different times after pollination. Absolute oil concentration of control embryos was37% at 15 DAP, 32% at 30 DAP, and 33% at 45 DAP. Error bars indicate 95% confidence intervals. C, DGAT activity fromembryos of homozygous inbred plants grown in the greenhouse. Ears were harvested at 30 DAP. Error bars indicate 95%confidence intervals. D, DAG concentrations in embryos of homozygous inbred plants grown in the greenhouse at 15, 30, and 45DAP. Error bars indicate LSD values, and letters indicate significant differences within a time point by t test (P , 0.05).
Fungal DGAT2 Increases Maize Oil
Plant Physiol. Vol. 155, 2011 1151 www.plantphysiol.orgon April 12, 2019 - Published by Downloaded from
Copyright © 2011 American Society of Plant Biologists. All rights reserved.
mittance (NIT). Many of the events had significantlyreduced starch content, and a plot of starch versus oilcontents showed that increases in kernel oil wereaccompanied by decreases in kernel starch (Fig. 7C).No significant differences from controls were observedfor protein content for any of the events, but there didappear to be a trend of increased protein in eventsexpressing fungal DGAT transgenes (Fig. 7D).
DGAT2 Transgene Expression in High-Oil Hybrids
To determine whether a fungal DGAT2 could in-crease kernel oil above levels produced when the high-oil allele of the maize DGAT1 gene is present, wecrossed events containing the fungal DGAT2 genes toseven near-elite high-oil testers to produce high-oilhybrids. These high-oil lines were shown by PCRanalysis to possess the high-oil allele of DGAT1 (datanot shown). The lines were homozygous for the high-oilDGAT1 allele, except High Oil 1, which was heterozy-
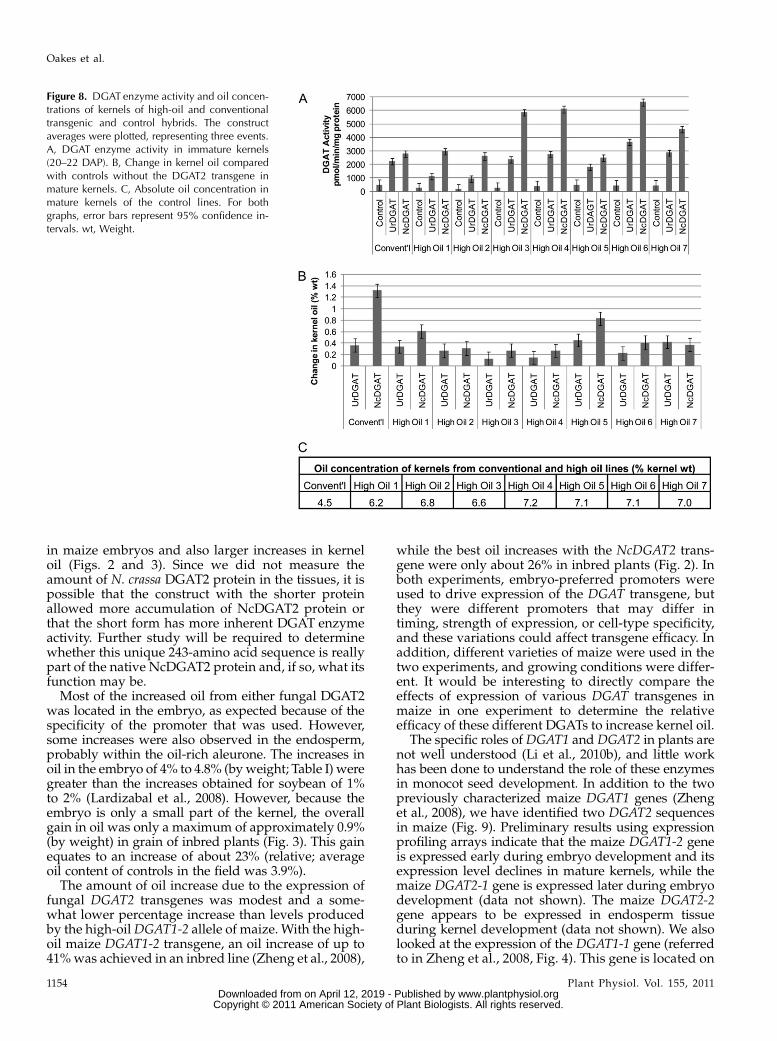
gous. An eighth hybrid was made with a conventionalelite tester, which contained the normal-oil allele ofDGAT1. The F1 plants were grown in a field innorthern California. Ears were hand harvested at 20to 22 DAP and at maturity, and kernels were tested forDGAT activity (Fig. 8A, immature kernels) or oil con-centration (Fig. 8B, mature kernels).
All eight control lines without the fungal DGAT2transgenes had similar levels of DGAT activity in de-veloping kernels (Fig. 8A). DGATactivitywas increasedin kernels of all lines by theDGAT2 transgenes. Activitywas greater with the S-NcDGAT2 transgene than withthe UrDGAT2A transgene in most of the lines. Theelevation in DGAT activity from the NcDGAT2 trans-gene was greater in some of the high-oil hybrids than inthe conventional-oil hybrid.
Final oil concentration for the nontransgenic high-oil hybrids ranged from 6.2% to 7% compared with4.5% for the conventional hybrid (Fig. 8C). Small, butstatistically significant, increases in kernel oil weremeasured for most of the hybrids for both UrDGAT2A
Figure 5. Fatty acid composition of mature kernels. Values plotted aredifferences from the control as a percentage of total fatty acids. Errorbars indicate LSD. A, 16:0, 18:0, 18:1, and 18:2 fatty acids. B, 18:3,20:0, 20:1, and 22:0 fatty acids. C, Fatty acid composition of the controlas a percentage of total fatty acids.
Figure 6. Abnormal seedling phenotype with varying penetranceobserved in some inbred lines. A, Inbred seedlings from control ortransformed lines with severely abnormal phenotype 8 d after sowing.These seedlings are shown as examples; not all transgenic seedlingswere affected. Some transgenic seedlings had a more mild abnormalphenotype, and others were normal. B, Graph showing the percentageof seedlings from different events that were homozygous (Hom),hemizygous (Hem), or null for the transgene that displayed any ob-servable abnormal phenotype from the greenhouse experiment. Theremainder of the seedlings had a normal phenotype. Sixty-eight to 76seedlings were examined for the first five lines, which were hemizy-gous, and 16 seedlings were examined for event S-Nc 538, which washomozygous.
Oakes et al.
1152 Plant Physiol. Vol. 155, 2011 www.plantphysiol.orgon April 12, 2019 - Published by Downloaded from
Copyright © 2011 American Society of Plant Biologists. All rights reserved.
and S-NcDGAT2 (Fig. 8B). Oil increases obtained withthe UrDGAT2A transgene were similar between con-ventional- and high-oil lines and ranged from 0.1% to0.4%, producing 2% to 8% more oil. The greatest oilincrease was produced by the S-NcDGAT2 transgenein the conventional-oil hybrid, with an increase of1.3% by weight, a relative increase of 30%. In the high-oil hybrids, S-NcDGAT2 produced oil increases of 0.2%to 0.8% by weight, giving 4% to 12% more oil.
DGAT2 Genes in Maize
To begin to provide more understanding of the rela-tive contributions of DGAT1 and DGAT2 proteins in theaccumulation of oil in maize kernels, we conducted asearch for maize DGAT2 sequences in Monsanto cDNAlibraries using the castor bean DGAT2 amino acid se-quence as the query (GenBank accession no. AY916129;Kroon et al., 2006). We identified two maize sequencesthat are closely related to other plant DGAT2 sequences.Subsequently, the maize B73 genome was sequenced(Schnable et al., 2009), and DGAT2-1 and DGAT2-2were mapped to chromosome 9 (LOC100217122) andchromosome 4 (LOC100283803), respectively, by BLASTagainst maize at http://maizesequence.org. Homologs
of these genes can be found in many other sequencedplant genomes, including rice (Oryza sativa) and soy-bean. Alignment of plant and fungal DGAT2 se-quences showed expected clustering of taxonomicgroups (plant versus fungal and dicot versus monocot;Fig. 9).
DISCUSSION
Small but statistically significant increases in maizekernel oil were achieved by expression of fungalDGAT2 transgenes using an embryo-enhanced pro-moter. A DGAT2 gene isolated from N. crassa encodedan enzyme with somewhat greater activity in maizekernels than the enzyme from U. ramanniana, andexpression of the S-NcDGAT2 transgene resulted ingreater increases in maize kernel oil. Two forms of theNcDGAT2 gene were tested. The full-length aminoacid sequence includes 243 residues that are unique tothis protein and located in the N-terminal half of theprotein. Therefore, we tested both the full-length se-quence (L-NcDGAT2) and the shorter sequence thataligns with the UrDGAT2A sequence throughout itslength (S-NcDGAT2). We found that expression ofS-NcDGAT2 resulted in higher DGAT enzyme activity
Figure 7. Kernel oil and grain yield for a multilocation hybrid field trial. A, Oil content measured by NMR expressed as thedifference from the control. OP, Grain from an open-pollinated hybrid agronomic trial planted at 10 locations; HP, grain from atwo-location hand-pollinated trial. Event 871 was not planted in the hand-pollinated trial because sufficient seed was notavailable. Numbers on the x axis are abbreviated event names. Error bars represent 95% confidence intervals. wt, Weight. B,Grain yield expressed as the percentage difference from the control in a 10-location field trial. Yields of controls were 200bushels per acre (UrDGAT2A) and 212 bushels per acre (NcDGAT2). Error bars represent 95% confidence intervals. C and D, Oiland starch (C) and protein and starch (D) were measured by NIT for grain from the two-location hand-pollinated trial. Values arepercentage dry weight of the grain. Each point represents the mean of 15 to 40 samples. White diamonds, Controls; blackdiamonds, transgenic events.
Fungal DGAT2 Increases Maize Oil
Plant Physiol. Vol. 155, 2011 1153 www.plantphysiol.orgon April 12, 2019 - Published by Downloaded from
Copyright © 2011 American Society of Plant Biologists. All rights reserved.
in maize embryos and also larger increases in kerneloil (Figs. 2 and 3). Since we did not measure theamount of N. crassa DGAT2 protein in the tissues, it ispossible that the construct with the shorter proteinallowed more accumulation of NcDGAT2 protein orthat the short form has more inherent DGAT enzymeactivity. Further study will be required to determinewhether this unique 243-amino acid sequence is reallypart of the native NcDGAT2 protein and, if so, what itsfunction may be.
Most of the increased oil from either fungal DGAT2was located in the embryo, as expected because of thespecificity of the promoter that was used. However,some increases were also observed in the endosperm,probably within the oil-rich aleurone. The increases inoil in the embryo of 4% to 4.8% (byweight; Table I) weregreater than the increases obtained for soybean of 1%to 2% (Lardizabal et al., 2008). However, because theembryo is only a small part of the kernel, the overallgain in oil was only a maximum of approximately 0.9%(by weight) in grain of inbred plants (Fig. 3). This gainequates to an increase of about 23% (relative; averageoil content of controls in the field was 3.9%).
The amount of oil increase due to the expression offungal DGAT2 transgenes was modest and a some-what lower percentage increase than levels producedby the high-oilDGAT1-2 allele of maize. With the high-oil maize DGAT1-2 transgene, an oil increase of up to41%was achieved in an inbred line (Zheng et al., 2008),
while the best oil increases with the NcDGAT2 trans-gene were only about 26% in inbred plants (Fig. 2). Inboth experiments, embryo-preferred promoters wereused to drive expression of the DGAT transgene, butthey were different promoters that may differ intiming, strength of expression, or cell-type specificity,and these variations could affect transgene efficacy. Inaddition, different varieties of maize were used in thetwo experiments, and growing conditions were differ-ent. It would be interesting to directly compare theeffects of expression of various DGAT transgenes inmaize in one experiment to determine the relativeefficacy of these different DGATs to increase kernel oil.
The specific roles ofDGAT1 andDGAT2 in plants arenot well understood (Li et al., 2010b), and little workhas been done to understand the role of these enzymesin monocot seed development. In addition to the twopreviously characterized maize DGAT1 genes (Zhenget al., 2008), we have identified two DGAT2 sequencesin maize (Fig. 9). Preliminary results using expressionprofiling arrays indicate that the maize DGAT1-2 geneis expressed early during embryo development and itsexpression level declines in mature kernels, while themaize DGAT2-1 gene is expressed later during embryodevelopment (data not shown). The maize DGAT2-2gene appears to be expressed in endosperm tissueduring kernel development (data not shown). We alsolooked at the expression of the DGAT1-1 gene (referredto in Zheng et al., 2008, Fig. 4). This gene is located on
Figure 8. DGAT enzyme activity and oil concen-trations of kernels of high-oil and conventionaltransgenic and control hybrids. The constructaverages were plotted, representing three events.A, DGAT enzyme activity in immature kernels(20–22 DAP). B, Change in kernel oil comparedwith controls without the DGAT2 transgene inmature kernels. C, Absolute oil concentration inmature kernels of the control lines. For bothgraphs, error bars represent 95% confidence in-tervals. wt, Weight.
Oakes et al.
1154 Plant Physiol. Vol. 155, 2011 www.plantphysiol.orgon April 12, 2019 - Published by Downloaded from
Copyright © 2011 American Society of Plant Biologists. All rights reserved.

chromosome 6 (LOC100383910), as is theDGAT1-2 gene(LOC100283803), and is also expressed early in embryodevelopment, although at lower levels than DGAT1-2.The expression of DGAT1-1 appears to be less specificthan DGAT1-2, with expression in shoot apical meri-stem and tassels (based on analysis of the NCBI ESTprofile data by Pontius et al. [2003]). Thus, these DGATgenes are differentiated by timing and tissue specificity.Whether the enzymatic activities of the two DGATforms are distinct in maize remains to be determined.To better understand the potential benefit of addi-
tional DGAT activity on increasing kernel oil, wemeasured the level of the substrate for DGAT, DAG,in wild-type and transgenic maize embryos duringkernel development. The DAG pool was reduced byexpression of the fungal DGATs, and very little DAGwas present in the embryos at maturity (Fig. 4D).However, it seems possible that sufficient DAG wouldbe present at 30 DAP to support higher levels of oilbiosynthesis if DGAT activity were further increased.As with overexpression of DGAT1 in maize (Zheng
et al., 2008), overexpression of the fungal DGAT2 genesresulted in changes in fatty acid composition. In par-ticular, there were increases in 18:1 (up to 18%) anddecreases in 18:2 (up to 15%). In agreement with itsgreater enzyme activity, larger changes in fatty acidcomposition were observed from overexpression ofS-NcDGAT2 compared with UrDGAT2A (Fig. 5). Thefatty acid composition changes that were observed inevents expressing S-NcDGAT2 are similar in type andmagnitude to those reported for overexpression of thehigh-oil DGAT1 gene in maize (Zheng et al., 2008), andthese values are within the range reported for non-transgenic maize (Reynolds et al., 2005). This effect onfatty acid composition has been hypothesized to resultfrom competition for DAG as a substrate by bothcholine phosphotransferase and by DGAT (Zhenget al., 2008). Higher levels of DGATactivity could result
in reduced levels of oleoyl-phosphatidylcholine madeby choline phosphotransferase, which is a substrate forthe FAD2 desaturase that is responsible for the produc-tion of 18:2 fatty acids. Thus, increased competition atthis juncture in the pathway could have an impact onthe production of polyunsaturated fatty acids.
We observed a phenotype in seedlings of someplants overexpressing the fungal DGAT genes in whichthe first leaves appeared to be stuck together and failedto expand normally (Fig. 6). It is interesting that inArabidopsis, the highest levels of expression of theDGAT1 gene are in germinating seeds and youngseedlings (Zou et al., 1999), although the need forhigh levels of DGAT expression at this stage of de-velopment is not clear. In addition, study of Brassicanapus plants with reduced DGAT1 expression byantisense suppression revealed the presence of somedevelopmental abnormalities, including effects on seeddevelopment (Lock et al., 2009). These researchersconcluded that DGAT1may have a specific role in seeddevelopment of B. napus in addition to control of TAGaccumulation. Mutants of wax production can havefused organs (Chen et al., 2003), showing the impor-tance of cuticular waxes in organ development. It ispossible that excess DGAT activity may lead to theproduction of extra or abnormal cuticular waxes andcause the fusion of seedling leaves. However, the ab-normal phenotype does not appear to be directly cor-related with the amount of transgenic DGAT2 activity,since the phenotype was more common in lines withUrDGAT2A thanwithNcDGAT2, even thoughNcDGAT2lines tended to have higher DGAT activity (Fig. 3B).Perhaps differences in the enzyme activities of thesetwo DGATs may include different specificities for sub-strates of wax biosynthesis.
UrDGAT2A and S-NcDGAT2 activities demon-strated some interesting differences that cannot beexplained simply by the greater activity of S-NcDGAT2.The abnormal seedling phenotype was greater withUrDGAT2A, and the two DGATs had different effectson fatty acid composition (Fig. 5). S-NcDGAT2 expres-sion resulted in larger increases in 18:1 and largercorresponding decreases in 18:2 and 18:3. But expres-sion ofUrDGAT2A had bigger effects on levels of longerchain fatty acids (20:0, 20:1, and 22:0). It would bevaluable to test DGAT2 genes from additional sourcesto gain a better understanding of the fatty acid compo-sitional changes and to have an opportunity to furtheroptimize the increase in kernel oil.
We have shown that fungal DGAT2 genes have thepotential to be useful in the development of maize withhigher oil content. These transgeneswere able to increasekernel oil in hybrids in the field and appeared not tohave any negative effect on grain yield (Fig. 7). Theywere also able to increase total oil content in high-oilhybrids, which contain the high-oil allele of DGAT1(Zheng et al., 2008; Fig. 8). Additional increases in kerneloil may be achievable with alternativeDGAT genes or byvarying the timing of gene expression during embryodevelopment and maturation. In addition, engineering
Figure 9. Alignment of plant and fungal DGAT2 deduced amino acidsequences using the MegAlign program (ClustalV algorithm). Speciesidentifications with GenBank accession numbers are as follows: Rc,Ricinus communis, AY916129; Vf, Vernicia fordii, ABC94473; Vg,Vernonia galamensis, FJ652577; At, Arabidopsis, NP566952.1; Ur, U.ramanniana, AF391089; Nc, N. crassa, XP965438.2, including onlythe amino acid sequences that align with the UrDGAT2A protein (Fig.1); Sc, Saccharomyces cerevisiae, EDN63574.1. Maize sequenceZmDGAT2-1 is ACL54036.1 and ZmDGAT2-2 is ACG40288.1.Rice sequence OsDGAT2-1 is NM_001064065 and OsDGAT2-2 isNM_001054452.
Fungal DGAT2 Increases Maize Oil
Plant Physiol. Vol. 155, 2011 1155 www.plantphysiol.orgon April 12, 2019 - Published by Downloaded from
Copyright © 2011 American Society of Plant Biologists. All rights reserved.

upstream pathways to provide additional DAG sub-stratewould likely be a productive approach to obtainingfurther oil increases. Overexpression of the transcrip-tion factor ZmWRI1 has been shown to up-regulategenes involved in fatty acid biosynthesis (Shen et al.,2010), so combining overexpression of fungal DGAT2with ZmWRI1 would likely produce a synergistic ef-fect on oil accumulation in maize kernels.
MATERIALS AND METHODS
Cloning L-NcDGAT2 cDNA
The L-NcDGAT2 sequence was identified from a BLAST search of public
databases using the previously cloned NcDGAT2 sequence (Lardizabal et al.,
2004) as the query. A sequence identified as “hypothetical protein NCU02665
[Neurospora crassaOR74A]” (NCBI accession no. 85118420; Galagan et al., 2003)
was 100% identical along the length of the originally identified NcDGAT2
sequence but also included an upstream sequence encoding an additional
243 amino acids not found in the UrDGAT2A protein (Fig. 1). To clone this
sequence, RNA was isolated from N. crassa mating type A (Fungal Genetics
Stock Center) mycelium using Tri-Reagent (Sigma) according to the manu-
facturer’s protocol. First-strand cDNA synthesis was completed using the
SMART cDNA Amplification kit (Clontech). Oligonucleotide primers were
designed to amplify the full-length coding region of the NcDGAT2 sequence,
and the PCR products were cloned into plasmid pCR2.1 according to the
manufacturer’s protocol (Invitrogen).
Vector Construction and Plant Transformation
Maize (Zea mays) transformation vectors were made using the barley
(Hordeum vulgare) Per1 promoter and 5# untranslated region, which contained
an intron from maize (ZmHsp70) to control the expression of each of the three
fungal DGAT2 genes. The sequence for UrDGAT2Awas the codon-optimized
form described previously (Lardizabal et al., 2008), but the native N. crassa
sequences were used for both NcDGAT2 cDNAs. Either the 3# nontranslatedregion of the potato (Solanum tuberosum) proteinase inhibitor II gene (Keil
et al., 1986; UrDGAT2A and S-NcDGAT2 constructs) or the 3# nontranslated
region from the wheat (Triticum aestivum) hsp17 gene (L-NcDGAT2 construct)
was fused to the 3# end of the coding sequence to provide a signal for
polyadenylation. The selectable marker cassette for these vectors included
Agrobacterium tumefaciens CP4 5-enol-pyruvylshikimate-3-phosphate synthase
driven by the rice (Oryza sativa) actin1 promoter and completed by the 3# endwith the Agrobacterium nopaline synthase termination sequence. Both the gene
cassettes were within the T-DNA borders.
A near-elite maize inbred line was transformed by the method described
by Armstrong and Rout (2003). Transgenic maize lines containing one or two
copies of the construct were selected for further study. R0 plants were self-
pollinated and grown to maturity in the greenhouse. Subsequent inbred
generations were grown in the field in Hawaii.
Greenhouse Experiment
The greenhouse wasmaintained at a day temperature of 30�C to 31�C and a
night temperature of 20�C to 21�C. The photoperiod was maintained at 18 h
minimum with a light intensity of 900 to 1,400 mE m22 s21. The soil was
Sunshine No. 1 with 14-14-14 NPK fertilizer and Micromax micronutrient
incorporated at a moderate rate. Plants were fed by continuous liquid
fertilization via irrigation with calcium nitrate, Epsom salts, and 0-10-30
fertilizer with micronutrients. All plants were self-pollinated. Sampling of
ears was conducted at 15, 30, and 45 DAP. At 15 and 30 DAP, ears were
harvested and immediately frozen in liquid nitrogen. Ears were stored at
270�C, and subsequent manipulations and dissections were done on dry ice.
Dissected embryos were hand ground in liquid nitrogen with a mortar and
pestle. Approximately 50 mL of material was subsampled for DGAT assays.
The remaining ground embryo sample was freeze dried for lipid analysis.
Mature ears (45 DAP) were maintained at room temperature until dissection.
Kernels were softened for dissection by boiling in water for 10 min. Dissected
embryos were then frozen at 270�C, hand ground in liquid nitrogen with a
mortar and pestle, and freeze dried for lipid analysis.
Enzyme Assays
UrDGAT2A and NcDGAT2 enzyme activities were measured as described
(Lardizabal et al., 2006), with the following changes. The assay buffer for plant
samples substituted 5 mM Tricine, pH 7.5, for the 10 mM potassium phosphate,
280 mM NaCl for the 150 mM KCl, and 0.06% CHAPS for the 0.1% Triton X-100.
Assays were performed at 25�C for 10 min and were immediately processed
after assay termination. Twenty-six percent of the final organic phase was
liquid scintillation counted, and thin-layer chromatography was not per-
formed.
Seed Composition
NMR
Seed oil and moisture content for both whole maize kernels and dissected
embryos and endospermwere determined by pulsed NMR (Tiwari et al., 1974;
Rubel, 1994) using a Maran Ultra-20 Benchtop NMR spectrometer (Resonance
Instruments) operated at 23.4 MHz. The NMR spectrometer was calibrated
using 100% maize oil, and results were reported on a dry matter basis.
NIT Spectroscopy
Bulk quantities of maize consisting of 30 to 50 mL of maize kernels (i.e.
100–150 kernels) were analyzed for average oil, starch, and protein contents
with a NIT method using a Tecator Infratec 1221 scanning monochromator
(Foss North America) with a 30-mm path-length sample cuvette. Absorbance
values, as log (1/T) where T represents transmission, were recorded at 2-nm
intervals between 850 and 1,048 nm. These absorbance values were translated
to oil, starch, and protein content measurements using Infratec Corn Analysis
model CO980811. The commercial Analysis model and standard operating
procedure were validated relative to Accelerated Soxhlet Extraction results
prior to this analysis; the SE of accuracy (absolute errors of differences) was
shown to be 0.6% for moisture, 0.4% for oil, 0.3% for protein, 1.2% for starch,
and 1.5% for extractible starch relative to dry sample weight.
Lipid Analysis
Oil fatty acyl composition was analyzed by gas-liquid chromatography of
methyl esters (Browse et al., 1986). DAG and free fatty acid pool sizes were
determined by a modification of a HPLC-evaporative light scattering detector
method using a silica column instead of a cyanopropyl column (El-Hamdy
and Christie, 1993; American Oil Chemists’ Society, 1999).
Hybrid Field Testing
Yield trials with elite testers were conducted at multiple locations in Iowa
and Illinois using two-row, 22-foot-long plots. Transgenic lines and their
controls were separated by two plots of sterile hybrids to minimize cross-
pollination. Harvest and yield determination was done using a combine. In
addition, the same hybrid entries were planted at two locations in the
Midwest and one in California and hand pollinated. Ears were hand-
harvested at maturity for analysis of oil content.
Genbank accession numbers for DGAT2 sequences in this article are as
follows: castor bean (Ricinus communis), AY916129; tung tree (Vernicia fordii),
ABC94473; Vernonia galamensis, FJ652577; Arabidopsis (Arabidopsis thaliana),
NP566952.1; Umbelopsis ramanniana, AF391089; N. crassa, XP965438.2; Sac-
charomyces cerevisiae, EDN63574.1.; maize DGAT2-1, ACL54036.1; maize
DGAT2-2, ACG40288.1; rice DGAT2-1, NM_001064065; and rice DGAT2-2,
NM_001054452.
ACKNOWLEDGMENTS
We thank the many individuals from Monsanto who contributed to this
work, including the cloning team, the vector archiving team, the maize
transformation team, the molecular analysis team, and the nursery and field
teams. Analytics support was provided by Chingying Li, Tom Hayes, Tim
Hickman, and Martin Ruebelt. Mohammadreza Ghaffarzadeh, Trenton
Stanger, and the Monsanto farm teams provided field expertise. Martin
Oakes et al.
1156 Plant Physiol. Vol. 155, 2011 www.plantphysiol.orgon April 12, 2019 - Published by Downloaded from
Copyright © 2011 American Society of Plant Biologists. All rights reserved.

Stoecker, Eliza Anderson, Audrey Vaughn, and Rebecca Ryan contributed to
line advancement and field trials. Statistical analysis was conducted by
Beiyan Zeng and Jay Harrison. Greenhouse support was provided by Dan
Ovadya and his team. High-oil maize lines were supplied by Tom Carlson.
We thank Dangyang Ke and Steve Schwartz for many helpful discussions and
Benjamin Moll for critical review of the manuscript.
Received October 20, 2010; accepted January 14, 2011; published January 18,
2011.
LITERATURE CITED
American Oil Chemists’ Society (1999) Mono- and diglycerides determi-
nation by HPLC-ELSD. In Sampling and Analysis of Commercial Fats
and Oils. AOCS Official Method Cd 11d-96. American Oil Chemists’
Society, Champaign, IL
Armstrong CL, Rout JR (2003) Novel Agrobacterium-mediated plant trans-
formation method. U.S. Patent Application No. US2003/0204875A1
Ash M, Dohlman E (2006) USDA Oil Crops Situation and Outlook Year-
book. Market and Trade Economics Division, Economic Research Ser-
vice, U.S. Department of Agriculture, Washington, DC
Browse J, McCourt P, Somerville C (1986) A mutant of Arabidopsis
deficient in c(18:3) and c(16:3) leaf lipids. Plant Physiol 81: 859–864
Cases S, Smith SJ, Zheng YW, Myers HM, Lear SR, Sande E, Novak S,
Collins C, Welch CB, Lusis AJ, et al (1998) Identification of a gene
encoding an acyl CoA:diacylglycerol acyltransferase, a key enzyme in
triacylglycerol synthesis. Proc Natl Acad Sci USA 95: 13018–13023
Chen X, Goodwin SM, Boroff VL, Liu X, Jenks MA (2003) Cloning and
characterization of the WAX2 gene of Arabidopsis involved in cuticle
membrane and wax production. Plant Cell 15: 1170–1185
Dudley JW, Lambert RJ, de la Roche IA (1977) Genetic analysis of crosses
among corn strains divergently selected for percent oil and protein.
Crop Sci 17: 111–117
El-Hamdy AH, Christie WW (1993) Separation of non-polar lipids by high
performance liquid chromatography on a cyanopropyl column. J High
Resolut Chromatogr 16: 55–57
Galagan JE, Calvo SE, Borkovich KA, Selker EU, Read ND, Jaffe D,
FitzHugh W, Ma LJ, Smirnov S, Purcell S, et al (2003) The genome
sequence of the filamentous fungus Neurospora crassa. Nature 422:
859–868
Jako C, Kumar A, Wei Y, Zou J, Barton DL, Giblin EM, Covello PS, Taylor
DC (2001) Seed-specific over-expression of an Arabidopsis cDNA en-
coding a diacylglycerol acyltransferase enhances seed oil content and
seed weight. Plant Physiol 126: 861–874
Keil M, Sanchez-Serrano J, Schell J, Willmitzer L (1986) Primary structure
of a proteinase inhibitor II gene from potato (Solanum tuberosum).
Nucleic Acids Res 14: 5641–5650
Kennedy EP (1961) Biosynthesis of complex lipids. Fred Proc Am Soc Exp
Biol 20: 934–940
Kroon JTM, Wei W, Simon WJ, Slabas AR (2006) Identification and
functional expression of a type 2 acyl-CoA:diacylglycerol acyltransfer-
ase (DGAT2) in developing castor bean seeds which has high homology
to the major triglyceride biosynthetic enzyme of fungi and animals.
Phytochemistry 67: 2541–2549
Lardizabal K, Effertz R, Levering C, Mai J, Pedroso MC, Jury T, Aasen E,
Gruys K, Bennett K (2008) Expression of Umbelopsis ramanniana DGA-
T2A in seed increases oil in soybean. Plant Physiol 148: 89–96
Lardizabal KD, Bennett KA, Wagner NW (2004) Diacylglycerol acyltrans-
ferase nucleic acid sequences and associated products. U.S. Patent
Application No. US200440107459A1
Lardizabal KD, Hawkins D, Thompson G (2006) Diacylglycerol acyltrans-
ferase proteins. U.S. Patent Application No. US135617
Lardizabal KD, Mai JT, Wagner NW, Wyrick A, Voelker T, Hawkins DJ
(2001) DGAT2 is a new diacylglycerol acyltransferase gene family:
purification, cloning, and expression in insect cells of two polypeptides
from Mortierella ramanniana with diacylglycerol acyltransferase activity.
J Biol Chem 276: 38862–38869
Li R, Yu K, Hatanaka T, Hildebrand DF (2010a) Vernonia DGATs increase
accumulation of epoxy fatty acids in oil. Plant Biotechnol J 8: 184–195
Li R, Yu K, Hildebrand DF (2010b) DGAT1, DGAT2 and PDAT expression
in seeds and other tissues of epoxy and hydroxy fatty acid accumulating
plants. Lipids 45: 145–157
Lock YY, Snyder CL, Zhu W, Siloto RMP, Weselake RJ, Shah S (2009)
Antisense suppression of type 1 diacylglycerol acyltransferase ad-
versely affects plant development in Brassica napus. Physiol Plant 137:
61–71
Lung SC, Weselake RJ (2006) Diacylglycerol acyltransferase: a key medi-
ator of plant triacylglycerol synthesis. Lipids 41: 1073–1088
Misevic D, Alexander DE (1989) Twenty-four cycles of phenotypic recur-
rent selection for percent oil in maize. I. Per se and test-cross perfor-
mance. Crop Sci 29: 320–324
Pontius JU, Wagner L, Schuler GD (2003) UniGene: a unified view of the
transcriptome. In The NCBI Handbook. National Center for Biotechnol-
ogy Information, Bethesda, MD
Reynolds TL, Nemeth MA, Glenn KC, Ridley WP, Astwood JD (2005)
Natural variability of metabolites in maize grain: differences due to
genetic background. J Agric Food Chem 53: 10061–10067
Routaboul JM, Benning C, Bechtold N, Caboche M, Lepiniec L (1999) The
TAG1 locus of Arabidopsis encodes for a diacylglycerol acyltransferase.
Plant Physiol Biochem 37: 831–840
Rubel G (1994) Simultaneous determination of oil and water contents in
different oilseeds by pulsed nuclear magnetic resonance. J Am Oil Chem
Soc 71: 1057–1062
Schnable PS, Ware D, Fulton RS, Stein JC, Wei F, Pasternak S, Liang C,
Zhang J, Fulton L, Graves TA, et al (2009) The B73 maize genome:
complexity, diversity, and dynamics. Science 326: 1112–1115
Shen B, Allen WB, Zheng P, Li C, Glassman K, Ranch J, Nubel D,
Tarczynski MC (2010) Expression of ZmLEC1 and ZmWRI1 increases
seed oil production in maize. Plant Physiol 153: 980–987
Shockey JM, Gidda SK, Chapital DC, Kuan JC, Dhanoa PK, Bland JM,
Rothstein SJ, Mullen RT, Dyer JM (2006) Tung tree DGAT1 and DGAT2
have nonredundant functions in triacylglycerol biosynthesis and are
localized to different subdomains of the endoplasmic reticulum. Plant
Cell 18: 2294–2313
Stacy RA, Munthe E, Steinum T, Sharma B, Aalen RB (1996) A peroxi-
redoxin antioxidant is encoded by a dormancy-related gene, Per1,
expressed during late development in the aleurone and embryo of
barley grains. Plant Mol Biol 31: 1205–1216
Tan SL, Morrison WR (1979) Lipids in the germ, endosperm and pericarp
of the developing maize kernel. J Am Oil Chem Soc 56: 759–764
Tiwari PN, Gambhir PM, Rajan TS (1974) Rapid and nondestructive
determination of seed oil by pulsed nuclear magnetic resonance tech-
nique. J Am Oil Chem Soc 51: 104–109
Turkish A, Sturley SL (2007) Regulation of triglyceride metabolism. I.
Eukaryotic neutral lipid synthesis: “many ways to skin ACAT or a
DGAT.” Am J Physiol Gastrointest Liver Physiol 292: G953–G957
Val DL, Schwartz SH, Kerns MR, Deikman J (2009) Development of a high
oil trait for maize. In AL Kriz, BA Larkins, eds, Biotechnology in
Agriculture and Forestry: Molecular Genetic Approaches to Maize
Improvement, Vol 63. Springer-Verlag, Berlin, pp 303–323
Watson SA (1987) Structure and composition. In SA Watson, PE Ramstad,
eds, Corn Chemistry and Technology. American Association of Cereal
Chemists, St. Paul, pp 53–82
Zhang M, Fan J, Taylor DC, Ohlrogge JB (2009) DGAT1 and PDAT1
acyltransferases have overlapping functions in Arabidopsis triacylglyc-
erol biosynthesis and are essential for normal pollen and seed devel-
opment. Plant Cell 21: 3885–3901
Zheng P, Allen WB, Roesler K, Williams ME, Zhang S, Li J, Glassman K,
Ranch J, Nubel D, Solawetz W, et al (2008) A phenylalanine in DGAT is
a key determinant of oil content and composition in maize. Nat Genet
40: 367–372
Zou J, Wei Y, Jako C, Kumar A, Selvaraj G, Taylor DC (1999) The
Arabidopsis thaliana TAG1 mutant has a mutation in a diacylglycerol
acyltransferase gene. Plant J 19: 645–653
Fungal DGAT2 Increases Maize Oil
Plant Physiol. Vol. 155, 2011 1157 www.plantphysiol.orgon April 12, 2019 - Published by Downloaded from
Copyright © 2011 American Society of Plant Biologists. All rights reserved.