evolutionary anticipation the human heart
TRANSCRIPT

Ann R Coll Surg Engl 2000; 82: 297-302
Review
Evolutionary anticipation of the human heart
Solomon Victor, Vijaya M Nayak
The Heart Institute, Chennai, India
We have studied the comparative anatomy of hearts from fish, frog, turtle, snake,crocodile, birds (duck, chicken, quail), mammals (elephant, dolphin, sheep, goat, ox,
baboon, wallaby, mouse, rabbit, possum, echidna) and man. The findings were analysedwith respect to the mechanism of evolution of the heart.
Key words: Comparative anatomy - Heart - Evolution - Anatomy
r7he origin of life is disputed.' Primordial soup - con-
sisting of basic chemicals such as oxygen, hydrogen,carbon, and methane - has been suggested as the pre-
cursor of self-replicating protein, the basic unit of life.'In the Vedic period, life was attributed to the Divine
Will.2 According to the Bible, creation was completed insix days. It is of interest that in the Vedic period, one
Divine day was considered equivalent to 4320 millionyears of human reckoning, based on astronomicalcalculation.3 It is, therefore, likely that a day alluded toin the Bible does not refer to one modem calendar day.The Holy Qur'an, states that creation was achieved insix periods.
Based on the study of fossils and natural history,Charles Darwin4 concluded that 'life was originallybreathed by the Creator into a few forms or into one ... andthat ... from so simple a beginning endless forms mostbeautiful and most wonderful have been and are beingevolved'.We have studied the comparative anatomy of hearts in
fish, frog, turtle, snake, crocodile, birds (duck, chicken,quail), mammals (elephant, dolphin, sheep, goat, ox,
baboon, wallaby, mouse, rabbit, possum, echidna) andman.-10 The findings were analysed to discover any duesrelating to the mechanism of evolution. The terms usedare as in human anatomy, imagining the animal heldhead up with its belly facing the observer.
Cartilaginous fish
In the cartilaginous fish such as the shark, the heart islodged in a pericardial cavity resting on a septumtransversum with a central tendinous and peripheralmuscular components. Anteriorly the parietal peri-cardium is adherent to the ventral cartilaginous chestwall. There is potential space for lungs on either side.
Externally there is a triangular sinus venosus. Itreceives a duct of Cuvier and a hepatic vein on eitherside. A large capacitance chamber is interposed bet-ween each lobe of the liver and the hepatic vein, beneaththe septum transversum. There is a common atriumwith bilateral isomeric atrial appendages (Fig. 1). Theventricular mass is symmetrical with right, left and
Ann R Coll Surg Engl 2000; 82
Correspondence to: Dr Solomon Victor, Director, The Heart Institute, 34 East Street, Kilpauk Garden Colony, Chennai - 600 010, IndiaTel: +91 44 483 1919; Fax: +91 44 484 4884; E-mail: [email protected]
The Royal College of Surgeons of England
297
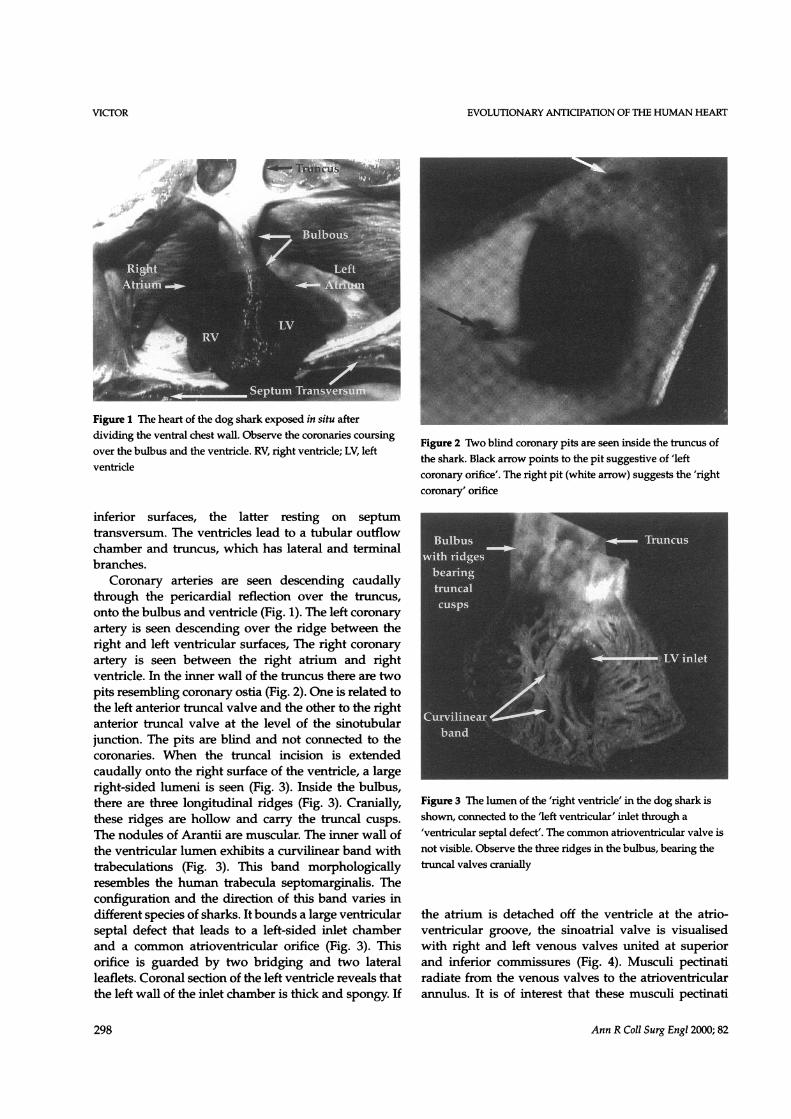
EVOLUTIONARY ANTICIPATION OF THE HUMAN HEART
Figure 1 The heart of the dog shark exposed in situ afterdividing the ventral chest wall. Observe the coronaries coursingover the bulbus and the ventride. RV, right ventricle; LV, leftventricle
inferior surfaces, the latter resting on septumtransversum. The ventricles lead to a tubular outflowchamber and truncus, which has lateral and terminalbranches.
Coronary arteries are seen descending caudallythrough the pericardial reflection over the truncus,onto the bulbus and ventricle (Fig. 1). The left coronaryartery is seen descending over the ridge between theright and left ventricular surfaces, The right coronaryartery is seen between the right atrium and rightventricle. In the inner wall of the truncus there are twopits resembling coronary ostia (Fig. 2). One is related tothe left anterior truncal valve and the other to the rightanterior truncal valve at the level of the sinotubularjunction. The pits are blind and not connected to thecoronaries. When the truncal incision is extendedcaudally onto the right surface of the ventride, a largeright-sided lumeni is seen (Fig. 3). Inside the bulbus,there are three longitudinal ridges (Fig. 3). Cranially,these ridges are hollow and carry the truncal cusps.The nodules of Arantii are muscular. The inner wall ofthe ventricular lumen exhibits a curvilinear band withtrabeculations (Fig. 3). This band morphologicallyresembles the human trabecula septomarginalis. Theconfiguration and the direction of this band varies indifferent species of sharks. It bounds a large ventricularseptal defect that leads to a left-sided inlet chamberand a common atrioventricular orifice (Fig. 3). Thisorifice is guarded by two bridging and two lateralleaflets. Coronal section of the left ventricle reveals thatthe left wall of the inlet chamber is thick and spongy. If
Figure 2 Two blind coronary pits are seen inside the truncus ofthe shark. Black arrow points to the pit suggestive of 'leftcoronary orifice'. The right pit (white arrow) suggests the 'rightcoronary' orifice
Figure 3 The lumen of the 'right ventricle' in the dog shark isshown, connected to the 'left ventricular' inlet through a'ventricular septal defect'. The common atrioventricular valve isnot visible. Observe the three ridges in the bulbus, bearing thetruncal valves cranially
the atrium is detached off the ventricle at the atrio-ventricular groove, the sinoatrial valve is visualisedwith right and left venous valves united at superiorand inferior commissures (Fig. 4). Musculi pectinatiradiate from the venous valves to the atrioventricularannulus. It is of interest that these musculi pectinati
Ann R Coll Surg Engl 2000; 82
VICTOR
298
cu
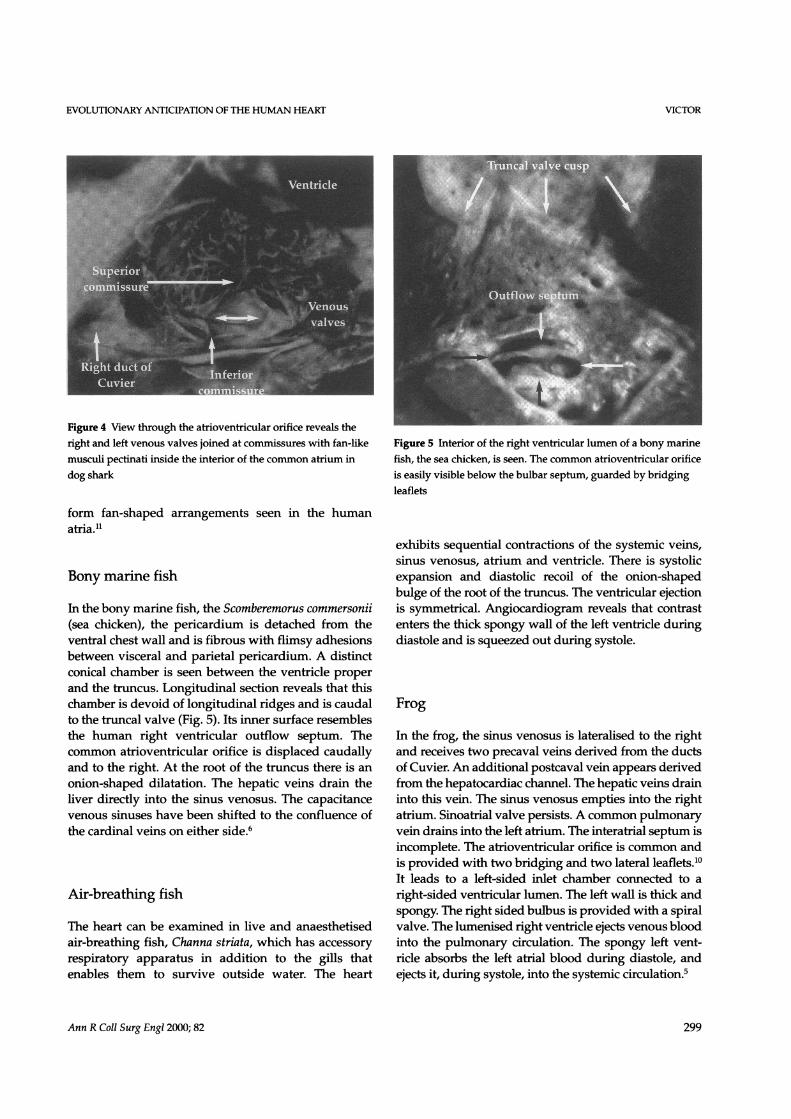
EVOLUTIONARY ANTICIPATION OF THE HUMAN HEART
Figure 4 View through the atrioventricular orifice reveals theright and left venous valves joined at commnissures with fan-like Figure 5 Interior of the right ventricular lumen of a bony marinemusculi pectinati inside the interior of the common atrium in fish, the sea chicken, is seen. The common atrioventricular orificedog shark is easily visible below the bulbar septum, guarded by bridging
leaflets
form fan-shaped arrangements seen in the humanatria."1
Bony marine fish
In the bony marine fish, the Scomberemorus commersonii(sea chicken), the pericardium is detached from theventral chest wall and is fibrous with flimsy adhesionsbetween visceral and parietal pericardium. A distinctconical chamber is seen between the ventricle properand the truncus. Longitudinal section reveals that thischamber is devoid of longitudinal ridges and is caudalto the truncal valve (Fig. 5). Its inner surface resemblesthe human right ventricular outflow septum. Thecommon atrioventricular orifice is displaced caudallyand to the right. At the root of the truncus there is anonion-shaped dilatation. The hepatic veins drain theliver directly into the sinus venosus. The capacitancevenous sinuses have been shifted to the confluence ofthe cardinal veins on either side.6
Air-breathing fish
The heart can be examined in live and anaesthetisedair-breathing fish, Channa striata, which has accessoryrespiratory apparatus in addition to the gills thatenables them to survive outside water. The heart
exhibits sequential contractions of the systemic veins,sinus venosus, atrium and ventricle. There is systolicexpansion and diastolic recoil of the onion-shapedbulge of the root of the truncus. The ventricular ejectionis symmetrical. Angiocardiogram reveals that contrastenters the thick spongy wall of the left ventricle duringdiastole and is squeezed out during systole.
Frog
In the frog, the sinus venosus is lateralised to the rightand receives two precaval veins derived from the ductsof Cuvier. An additional postcaval vein appears derivedfrom the hepatocardiac channel. The hepatic veins draininto this vein. The sinus venosus empties into the rightatrium. Sinoatrial valve persists. A common pulmonaryvein drains into the left atrium. The interatrial septum isincomplete. The atrioventricular orifice is common andis provided with two bridging and two lateral leaflets.10It leads to a left-sided inlet chamber connected to aright-sided ventricular lumen. The left wall is thick andspongy. The right sided bulbus is provided with a spiralvalve. The lumenised right ventricle ejects venous bloodinto the pulmonary circulation. The spongy left vent-ricle absorbs the left atrial blood during diastole, andejects it, during systole, into the systemic circulation.5
Ann R Coll Surg Engl 2000; 82
VICTOR
299

EVOLUTIONARY ANTICIPATION OF THE HUMAN HEART
Turtle
There is sequential contraction of the systemic andpulmonary veins, sinus venosus atrium and ventricles.There is a complete interatrial septum. The atrio-ventricular orifices are D-shaped with a commonstraight segment.9'10 Each orifice is provided with atilting-disc type of valve hanging from the interatrialseptum.9'10 The curved segment of each annuluscontracts during systole. There is no curved leaflet.Anatomically, the triple outlet right ventricle has abulbo trabecular septum with a pulmonary arteryventrally, and two aortae facing a ventricular septaldefect. The left ventricle, which is largely spongy, ejectsthrough the ventricular septal defect into the twoaortae connected to the right ventricle. The rightventricle ejects into the pulmonary circulation.5
Snake
The atrioventricular valves and circulation are similarto the turtles.5 The left ventricle is largely spongy witha small inlet chamber. It ejects through a ventricularseptal defect into the two aortae, anatomically con-nected to the right ventricle.
Crocodile
In the crocodile, interventricular and interatrial sept-ation is complete. The sinus venosus remains con-nected to the right atrium with persistence of thesinoatrial valve. The right aorta is connected to the leftventricle with consequent increase in the lumen of theleft ventricle.10 However, the stratum spongiosum ofthe left ventricle is thick. This right aorta displaces thestraight leaflet of the left atrioventricular valve fromthe septal location to an aortic position.10 The valveacquires a curved mural leaflet in relation to the muralwall of the left ventricle. Anterolateral and postero-medial muscles are seen inside the left ventricledirectly attached to the corresponding leaflets with nointervening chordae.10 The venous atrioventricularvalve has a straight septal and a curved mural leaflet.There is an anterior papillary muscle inside the rightventricle.10
Interestingly, during angiocardiography the rightventricle ejects only into the pulmonary circulation.The left aorta connected to the right ventricle receivesblood from the right. aorta through an aorto-aorticforamen.5 If the pulmonary artery is occluded, there isno distention of the right ventricle. A cog-wheel type of
muscular infundibular valve below the pulmonaryvalve shuts off the pulmonary circulation when thecrocodile is under water.5 The right ventricle then ejectsinto the left aorta.5
Birds
In the birds, the sinus venosus is amalgamated with theright atrium. The right venous valve is seen as a ridgeinside the right atrium.12 There is a similar ridge insidethe left atrium. Both of these ridges are connected by atransatrial band. Both ridges yield musculi pectinatileading to the atrioventricular annulus.7 Curiously, theright atrioventricular valve has only a curved muralleaflet in relation to mural wall of the right ventricle.9'10The septal leaflet is rudimentary or absent. The entireaorta is connected to the left ventricle with twocoronaries. The base of the aortic leaflets are muscular.The muscle in the aortic cusps and intercusp area forman annular band of musculature. A segment of thisband is interposed between the left and non-coronarycusps of the aortic valve and the aortic leaflet of themitral valve, and causes aorto-mitral discontinuity. Theleft atrioventricular valve has an aortic leaflet, abiscalloped mural leaflet, anterolateral and postero-medial chordopapillary support.9'10 The two scallops ofthe mural leaflet have an additional chordopapillarysupport.9'10
Egg-laying mammal
It is interesting that the atrioventricular valves ofechidna, an Australian egg-laying mammal, sharefeatures seen in the reptiles and birds.10 The muralleaflet of the left atrioventricular valve is biscalloped asin birds.10 However, the papillary muscles are directlyattached to the leaflets as in the crocodile.10 In the rightatrioventricular valve, the leaflet has becomemembranous.10 Additional posterior papillary musclesdevelop.10 The septal leaflet re-appears inferiorly.Superiorly, there is a gap between the septal leaflet andthe mural leaflet.10
Other mammals
The hearts of mouse, elephant, wallaby, possum,panther, pig, horse, cow, sheep, baboon and dolphinexhibit common features.10 The hearts are fourchambered. The right atrioventricular valve exhibitselaborate chordopapillary support and varying scallops
Ann R Coll Surg Engl 2000; 82
VICTOR
300

EVOLUTIONARY ANTICIPATION OF THE HUMAN HEART
of the mural leaflet. However, all of them essentiallyconsist of a straight leaflet and a curved mural leaflet.9The dominant scallop of the mural leaflet is equivalentto the anterior leaflet of the human tricuspid valvewith conus papillary support superiorly and anteriorpapillary support inferiorly. The chordopapillarysupport of the right or venous atrioventricular valvecan be separated into superior and inferior groups by aline bisecting the anterior and septal leaflets.13'14 Theleft atrioventricular valve is also similar to humanmorphology15 with a straight aortic leaflet, curvedmural leaflet with varying scallops, anterolateral andposteromedial papillary muscle groups.
Mechanism of evolution
Comparative anatomy of the heart reveals the heart ofthe fish has all the ingredients necessary for the humanheart. In particular there are two atria, two ventricles,and two coronaries. Blind coronary ostial pits are alsoseen.
'Nothing is superfluous in nature' said Leonardo daVinci. We need to speculate on why the heart of thefish that needs to circulate only venous blood has beenprovided with two atria and two ventricles. Obviously,there is a master plan for the heart. The eventual needfor the four-chambered heart was foreseen, and thenecessary basic framework established in the heart ofthe fish itself about 500 million years before Homo-sapiens sapiens appeared.
Step-by-step logical update of this basic design isobvious in comparative biology. The ducts of Cuviereventually become the coronary sinus and rightsuperior vena cava. The hepatic veins drain through ahepatocardiac channel, which becomes the inferiorvena cava. The sinus venosus lateralises to the rightinitially and later amalgamates with the right atrium.The right venous valve becomes the crista terminalis,Eustachian and Thebesian valves. The superior com-missure becomes the septum spurium, and the inferiorcommissure the tendon of Todaro.
The fate of left venous valve is disputed.7 We haveobserved that even though macroscopically there areno ridges in the body of the human left atrium,transillumination reveals a pattern of musculi pectinatiin the left atrium, identical with that of the rightatrium.11 This observation along with the observationof the left atrial bands in birds and mammals7 suggeststhat there is a left atrial crista terminalis in the humanheart possibly derived from the left venous valve.
The common atrioventricular orifice in fish andfrogs gets divided into two symmetrical D-shaped
orifices in the turtle and snake.9'10 This symmetry is lostwhen the septal leaflet moves to aortic position in thecrocodile. Later, the entire aorta is shifted to the leftventricle. The oval contractile orifice is replaced withtwo D-shaped orifices. The 'C' component of each ofthese orifices is related to the atrioventricular mus-culature and subject to deformation during systole. Therelated curved leaflet adjusts to this by developingscallops.
In phylogeny, the left ventricle has initially a smallinlet chamber and thick spongy wall. Later it getsprogressively cavitated.5 The left bundle of His seenprominently in mammals is replaced by left ventriculartendons in man.16
In man, we have observed contractions of thesuperior and inferior vena cava and the adjacent sinusvenareal part of the right atrium. This is in consonancewith the sequential contractions of the systemic veins,right atrium and right ventricle in the pre-humanspecies. We have also observed the terminal vena cavafibrillating along with the atrium in a patient withatrial fibrillation. The contraction of the terminalpulmonary veins precedes the atrium in snake, turtleand crocodile. We have observed, in a patient withlong intrapericardial course of all the pulmonaryveins, that the contractions of the pulmonary veinsprecede those of the left atrium.
Thus, comparative anatomy reveals a master plan,with logical step-by-step upgradation in variousspecies. It is also amazing that all the ingredientsnecessary for the human heart have been provided inthe heart of the shark. The question arises as towhether this master plan and anticipation of thedesign characteristics of the human heart, aeons oftime before Homosapiens sapiens appeared,17'18 is theresult of mere struggle for existence and survival of thefittest or whether there is a Designer behind thedesign?
While we give credit to the human intellect forhuman inventions, we are often hesitant to acknow-ledge an intellect behind nature's creation.17'18 We takerefuge behind terms like 'nature' or assume that lifecan evolve given a long period of time.
With all the human intellect, man is unable todesign a lung, valve, or blood vessel anywhere close tothat which occurs in nature. Do the species like fish,frog and reptiles know how to change their genome toredesign their cardiovascular and other systems,resulting in formation of a new species? Also,obviously, many species remain as they were, incountless varieties for millions of years. Which varietyof fish led to which variety of frog? Did the varietywhich was converted to a frog become obsolete or did
Ann R Coll Surg Engl 2000; 82
VICTOR
301

VICTOR EVOLUTIONARY ANTICIPATION OF THE HUMAN HEART
only some rebels in a particular species opt for achange. Even if a change occurred, the change has to beincorporated by microprocessor technology into thegerm cells for sustaining the changes in the offspring.Do various forms of life have access to this technology?There is no scientific proof that one form of life can orhas transformed itself into another form.
Comparative anatomy points to a design and aDesigner. Surgeons, anatomists and anyone studyingthe human form and function have an unsurpassedopportunity to ponder over the wonders of creationand contemplate the basic questions: where did wecome from? why are we here? and where are we going?
Acknowledgements
This article is based on the Arnott DemonstratorshipLecture delivered during a meeting of the Cardio-thoracic Section of the Royal Society of Medicine heldin London on 25 June 1999.
Our grateful thanks to Mr Raveen, Department ofZoology, Madras Christian College, Tambaram forassistance in dissecting various animals. We are alsothankful to Professor Franklin Rosenfeldt, BakerResearch Institute, Melbourne, and the staff of the Mel-bourne Zoo, for providing the hearts of the echidna,wallaby and possum for study. We are grateful toProfessor Vijayaragavan, Head of the Department ofAnatomy, Madras Veterinary College and his facultyfor the opportunity to study the heart of an elephant.Dr AN Subramanium, of the Marine Biology Depart-ment of Annamalai University, Parangipettai, TamilNadu, kindly provided dolphins for study.
References1. Monastersky R. The rise of life on earth. Natl Geog 1998; 193: 54-81.2. Vidyalankar S. The Holy Vedas, 2nd edn. New Delhi: Clarion, 1998;
30.3. Zimmer H. Myths and Symbols in Indian Art and Civilization.
Campbell J. (ed). Princeton, NJ: Princeton University Press, 1972; 16.4. Darwin C. The Origin of Species. New York, NY: The Modem Library,
1998.5. Victor S, Nayak VM, Raveen R. Evolution of the ventricle. Texas
Heart Inst J 1999; 26: 168-75.6. Victor S, Nayak VM, Raveen R. Evolution of the liver and the
hepatic venous oufflow [abstract]. The Third Congress of the AsianVascular Society, Beijing May 1998; 100.
7. Victor S, Nayak VM, Raveen R. Evolution of the venous connectionsof the heart [abstract]. Asia Pacific Heart J 1998; 7: 59.
8. Victor S, Nayak VM, Raveen R. Unfolding of the master plan forevolution of the great arteries [abstract]. The Third Congress of theAsian Vascular Society, Beijing May 1998; 129.
9. Victor S, Nayak VM. Bicuspid evolution of the arterial and venousatrioventricular valves. J Heart Valve Dis 1995; 4: 78-87.
10. Victor S, Nayak VM, Raveen R. Evolution of atrioventricular valvesspanning 600 million years [abstract]. Seventh Annual Conference ofAsian Societyfor Cardiovascular Surgery. Singapore May 1999; 419.
11. Victor S, Nayak VM. Musculi pectinati: design, function and surgicalconsiderations [abstract]. 13th Biennial Asian Congress on Thoracic andCardiovascular Surgery, Sydney, October 1997;107.
12. Victor S, Nayak VM. An anomalous muscle bundle inside the rightatrium related to the right venous valve. I Heart Valve Dis 1997; 6:439-40.
13. Victor S, Nayak VM. The tricuspid valve is bicuspid. J Heart ValveDis 1994; 3:27-36.
14. Victor S, Nayak VM. Bicuspidization for tricuspid stenosis. AnnThorac Surg 1998; 65: 1468-70.
15. Victor S, Nayak VM. Definition and function of commissures, slitsand scallops of the mitral valve: analysis in 100 hearts. Asia Pacific JThorac Cardiovasc Surg 1994; 3: 10-6.
16. Victor S, Nayak VM. Vestigial bands in the left ventricle. Asia PacificHeart J 1997; 6: 121-3.
17. Victor S. Anticipation and planning in evolution. Bhavan's J 1999; 45:63-6.
18. Victor S. Acknowledging the intellect. 7he Hindu December 71997; 2.
302 Ann R Coll Surg Engl 2000; 82