evidence for two photoreactions possible involvement ... · ofphytochromein light-dependent...
TRANSCRIPT

Plant Physiol. (1973) 51, 543-548
Evidence for Two Photoreactions and Possible Involvementof Phytochrome in Light-dependent Stomatal Opening'
Received for publication July 21, 1972
HELEN M. HABERMANNDepartment of Biological Sciences, Goucher College, Baltimore, Maryland 21204
ABSTRACT
Leaves of the xantha mutant of Helianthus annuus have ahigher rate of transpiration and a lower diffusive resistance inthe light than in the dark. Stomates of this nonphotosyntheticmutant open in the light and close in the dark.Comparative studies of tobacco, xantha mutant, and wild-
type sunflower stomatal opening over a range of light intensi-ties in isolated portions of the spectrum reveal two patterns ofresponse: (a) a low intensity opening in the green and far redcharacterized by partial opening, absence of a threshold, andsaturation of the response at low light intensities; (b) a highintensity response in the blue characterized by a threshold(intensities greater than 100 microwatts per square centimeterneeded for opening) and a linear opening response at higherincident light intensities. In xantha mutant stomates only thelow intensity system appears to be operational, while both lowand high intensity systems are present in the wild-type sun-flower and tobacco.Red light has an inhibiting effect on stomatal opening in
both mutant and wild-type sunflowers. They require prior ex-posure to far red for opening to occur in red light. This red-far red antagonism suggests the involvement of phytochrome.
The stomatal apparatus controls not only transpirationalwater loss but also the exchange of gases between the interiorand exterior of leaves. Stomatal opening or closing is a conse-quence of turgor changes in the guard cells that are influencedby a number of environmental factors including CO2 and 02tension, water supply, temperature, and light. Of these con-trolling factors, the influence of light is perhaps the leastunderstood.
Intensity, quality, and periodicity of light influence stomatalopening (9, 10, 13). Most hypotheses concerning the mech-anism of light regulation of stomates postulate the involve-ment of the photosynthetic apparatus, an implication of thealmost universal presence of chloroplasts in the guard cellsand their absence in other epidermal cells. This circumstantialevidence based on the pattern of chloroplast distribution issupported by reports of the unresponsiveness to light of sto-mates in nonphotosynthetic mutants of higher plants and inetiolated leaves (19-21). However, some aspects of the photo-response of stomates are inconsistent with, or even contradic-
'This work was supported by National Science FoundationGrant GB 14816.
tory to, the hypothesis that photosynthesis is involved. Moststudies of wavelength effects indicate that blue light is manytimes more effective in promoting stomatal opening than redlight (9, 12, 13) while these regions of the spectrum are equallyefficient in promoting photosynthesis. There is no questionabout the controlling effects of stomatal aperture on the ratesof photosynthesis through regulating the movement of atmos-pheric CO2 to the leaf mesophyll cells. Evidence that photosyn-thesis controls stomatal aperature, on the other hand, is farless convincing and involves secondary effects such as de-pletion of CO2 from the interior of the leaf, condensation orhydrolysis of the carbohydrate products of photosynthesis (in-terconversion of sugar and starch), or utilization of ATPformed during photosynthesis as a source of energy for themovement of solutes across membranes against concentrationgradients.Two mutants of Helianthus annuus, the yellow xantha and
the white albina (22), unlike albino materials previously ex-amined, do exhibit light-dependent increases in transpirationrates and opening of stomates. In the dark, their transpirationrates decrease and stomates close. The response to light ofthese nonphotosynthetic mutants argues against the involve-ment of photosynthesis in light-dependent stomatal opening.The experiments summarized here suggest that phytochrome isinvolved in the stomatal response to light and that opening isa consequence of two photoreactions that have distinctly dif-ferent wavelength dependence and kinetics.Many recent studies have presented overwhelming evidence
that light-dependent stomatal opening is accompanied by a fluxof potassium ions into the guard cells (1, 2, 7, 8, 14, 18). Ifphytochrome mediates the change in membrane permeabilitythat must be associated with such an ion flux, then it appearsthat light-dependent stomatal opening is but one of the turgorphenomena in plants which have a common mechanism. Theother major category of responses with this mechanism is thenyctinastic movement of leaves (16, 17). In the latter system,light-dependent turgor changes in specific cells are accom-panied by movements of potassium ions and are mediated byphytochrome.
MATERIALS AND METHODS
Plant Materials. The xantha and albina mutants of Heli-anthus annuus were propagated by grafting seedlings ontowild-type stocks according to procedures previously described(6, 22). Grafted mutant and control sunflowers were grown inpebbles irrigated with Hoagland's solution supplied with anautomatic nutrient cycling system.
Because large amounts of leaf material were needed, thexantha rather than the albina mutant was utilized for most ofthe experiments to be described. Xanthas can be grown in ourgreenhouse in any season, but the albina mutant must be grown
543
www.plantphysiol.orgon September 15, 2018 - Published by Downloaded from Copyright © 1973 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 51, 1973
5-15E
l00
3~
-X
400 500 600 700
Wavelength (nm)
FIG. 1. Spectral distribution of incident light intensity for lampand filter combinations used to control light quality. la: Blue light:General Electric blue fluorescent lamps plus 1 cm 10% CuSO4 filter;lb: blue light: Sylvania Gro-lux lamps plus 1 cm 10% CuS04 filter.2: Green light: General Electric green fluorescent lamps plus 1cm 10% CuS04 and one layer yellow cellophane filters. 3: Redlight: Sylvania Gro-lux lamps plus one layer red cellophane. 4: Farred light:50 w incandescent lamps plus one layer each red, blue,and green cellophane filters.
under filters (4) and therefore can be propagated only duringwinter months.
Tobacco plants (Nicotiana tabacum L., variety BroadleafJohn Williams) were grown in pots containing a soil, sand,and peat mixture. Some experiments with two additional varie-ties of tobacco (Florida Green and Florida Gold) indicatedthat there are no detectable varietal differences in stomatalresponse.
The guard cells of all experimental materials have beenexamined by phase contrast and fluorescence microscopy offreshly prepared epidermal strips from lower leaf surfaces.The presence or absence of chlorophyll fluorescence and struc-tural characteristics were noted. Observations were docu-mented by photomicrographs in which the same fields were
photographed first under phase contrast and then underfluorescence optics.
Leaves were harvested late in the afternoon on the day be-fore use. They were transferred (with petioles in water) to a
constant temperature (22 C) dark room. The next morningleaf discs (1.5 cm diameter) were cut with a No. 11 cork borer.There is no true "safelight" for this operation. A green filteredlamp was placed at least 1 m away from the work surface anddiscs were floated on water in 50-ml beakers and transferredto a light-tight box immediately after cutting.
Measurements. Transpiration rates were estimated as weightloss per unit time of single excised leaves mounted in vials ofwater. Leaves were equilibrated in the dark for 45 to 90 minbefore weighing was begun. The glass doors of a Mettleranalytical balance were covered with several layers of blackgreenhouse cloth during the dark periods. Daylight was sup-
plemented with a single 150-w incandescent bulb aligned withthe level of the leaf but placed 1.5 m away. Weights were re-
corded at 5-min intervals. Between readings in the light, theglass doors of the balance were opened. There were no meas-
urable changes in temperature within the balance chamberduring the light period. At the end of the sequence of readingswhich included an initial dark, a light, and final dark periods,the leaf blade was severed at the level of the stopper holding itin the vial. It was weighed, and its area was determined with a
planimeter. Transpiration rates could thus be expressed on a
weight or an area basis.Diffusive resistance was measured with a Lambda diffusive
resistance meter. Measurements with either the tubular or
horizontal sensors generally agreed. Readings were taken on
leaves of intact plants under the following conditions: in thegreenhouse (full sunlight) and in a constant temperature dark-
room (22 C) after dark equilibration and under a bank ofSylvania Gro-Lux lamps (approximately 500 ftw/cm2 incidentlight energy).
Stomatal aperture was estimated by means of the siliconeimpression technique(15). In all experiments in which aper-tures were measured, leaf discs were floated on a 1-cm layer ofwater. Samples exposed to the light (as well as dark controls)were maintained in constant temperature rooms (22-23 C). Thestandard duration of illumination was 2 hr. Stomatal aperaturewas measured under oil immersion with a calibrated ocularmicrometer. Samples examined were cellulose acetate replicasof silicone impressions. For estimates of per cent open sto-mates, stomates with apertures of 2,u or more were scored asopen, those with apertures of less than 2tp as closed.
Control of Light Quality and Intensity. Light intensity wasvaried by controlling the distance between samples and thesource of illumination. The range of distances available wasapproximately 10 to 100 cm. Light quality was controlled byselecting appropriate lamp and filter combinations. Spectraldistribution of incident light intensity was measured for everyexperimental condition with an ISCO model SR spectrora-diometer. Representative curves for each of the lamp and filtercombinations used to control light quality are shown in Figure1.
RESULTS
Guard Cell Structure and Chlorophyll Content. The xanthaand albina mutants of Helianthus annuus L. are nonphotosyn-thetic (22). The complete absence of detectable chlorophylland carotenoids in the albina and the presence of only tracelevels of chlorophyll in the xantha (4, 22) are consistent withthe absence of light-dependent changes in 02 uptake and CO2evolution. But are the trace amounts of chlorophyll in thexantha mutant distributed through all of the normally photo-synthetic cells and tissues of the leaf or localized within asingle cell type such as the guard cells? Fluorescence micros-copy, which provides a sensitive test for the presence of chloro-phyll, and observations of the same fields under phase contrastoptics were utilized to answer this critical question and to makegeneral comparisons of cell structure in the epidermal layers ofall experimental materials used in these studies.The guard cell chloroplasts of both wild-type sunflower and
tobacco leaf epidermal strips exhibit the bright red fluorescencecharacteristic of chlorophyll. Under fluorescence optics, albinaguard cells exhibit no traces of red fluorescence. In the xantha,there is a barely perceptible pink fluorescence associated withthe poorly developed plastids in some preparations and anabsence of detectable fluorescence in others. Thus, fluorescencemicroscopy is consistent with chemical analyses of the pigmentcontent of mutant leaves and provides no evidence that thepersistent traces of chlorophyll in the xantha mutant are lo-calized within the guard cells.
Structural peculiarities were evident only in the epidermalcells of the albina mutant. In this complete albino, there iswide variation in size of the stomatal apparatus with unusuallysmall to exceptionally large pairs of guard cells. Xantha mutantand wild-type stomates are structurally almost identical, themost distinguishing feature being the absence or presence ofstrong chlorophyll fluorescence in guard cell plastids.
Transpiration Rates and Diffusive Resistance of Mutant andWild-type Sunflowers. Transpiration rates in light and darkmeasured as weight loss from single excised leaves are sum-marized in Table I. In both xantha and albina mutants thereis a measurable response to illumination. Although individualleaves exhibit wide variations in rate, there is clearly a morerapid weight loss in the light than in the dark. Mutant leaves
544 HABERMANN
www.plantphysiol.orgon September 15, 2018 - Published by Downloaded from Copyright © 1973 American Society of Plant Biologists. All rights reserved.

TWO PHOTOREACTIONS IN STOMATAL OPENING
Table I. Summary of Transpiration Rates anzd Diffusive Resistantce Measutrements for Mutantt and Wild-type Helianthus annftuus L.
Experimental MfaterialConditions Units
Albina Xantha Wild-type
A. Fresh weight per unzit areamg cm-2 33.8 it 7.6 26.1 i 6.0 | 18.0 + 5.5
B. Transpiration rates
Excised leavesDark mg g-1 min-' 5.8 i 3.0 2.5 ± 1.4 3.3 i 2.2
mg cm-2 min-' 0.20 0.06 0.06Light;(room light plus incandescent mg g-1 min-' 8.6 ±4 4.7 5.0 4 2.1 8.5 ± 3.8supplement) mg cm72 min'1 0.29 0.13 0.15
C. Diffusive resistanceLeaves on intact plantsLight (greenhouse) cm sec-1 2.8 i 0.5 2.7 4i 0.2Dark (22 C) cm sec-1 11.0 4± 1.3 11.7 ± 1.9Light (22 C, Gro-Lux lamps) cm sec'1 3.1 ± 0.3 5.9 ± 1.2
tend to have more rapid rates of transpiration in the dark thando wild-type leaves because their stomates tend to remain par-tially open in the dark.
Transpiration rates have been expressed on a weight and onan area basis. There are differences between mutant and wild-type leaves in weight per unit area that are related to differ-ences in leaf thickness. The albina has an abnormal leafdevelopment with little differentiation of the mesophyll and in-hibited lamina expansion (4), while the xantha more nearlyapproximates the wild type in leaf development (22). Whentranspiration rates are expressed as weight loss per cm2 leafarea, the albina has the highest rate of transpiration in boththe light and dark with a smaller difference in rate between lightand dark than exhibited by the xantha and wild type.As would be anticipated, there are measurable differences in
values for diffusive resistance measured in the light and thedark (Table I). Only data for the xantha and wild type sun-flower are available because leaves of the albina mutant are toosmall and irregular to fit in the sensors of the diffusive resist-ance meter.The comparative data summarized in Table I indicate that
the guard cells of mutant leaves do respond to light and that aphotoactive stomatal opening can occur in the absence ofphotosynthesis.
Responses to Light Intensity. Preliminary studies with leafdiscs exposed to low intensity monochromatic light obtainedby means of interference filters presented a very confusing pic-ture of stomatal response to light as a function of wavelength.With xantha mutant and wild-type sunflower leaf discs, sto-matal opening was observed not only at 460 and 600 but alsoat 720 nm. Parallel runs with tobacco leaf discs revealednegligible opening at 460 and 600 nm with only partial open-ing at 720 nm. It seemed possible that the light intensities usedwere too low for significant opening to be observed and there-fore a series of light intensity-response curves were run be-ginning with "white" light from Sylvania Gro-Lux lamps.
White Light. The spectral output of Gro-Lux lamps is pre-dominantly in the blue and red, regions of the spectrum thatare most effective for photosynthesis. In "white" light obtainedfrom these lamps without filters, two patterns of stomatal re-sponse were evident (Fig. 2). With xantha mutant leaf discs,maximal stomatal aperture and the highest per cent of openstomates were found in leaf discs exposed to 300 ,uw/cmn. Athigher intensities, stomates closed. Wild-type sunflower and
E
aw
3.0
.o
i2.0 -.
.. b...
)800 0 200 400 600
Light intensity (,iW/cm2)
FIG. 2. Light intensity response curves to white light. SylvaniaGro-lux lamps (unfiltered). Xantha mutant sunflower (0); wild-type sunflower (0); tobacco (A).
tobacco stomates responded to low light intensities, as did thexantha mutant. In the range of 200 to 300 puw/cm' both ex-hibited a plateau in their response with greater opening athigher intensities. Without knowledge of the pattern of re-sponse in the xantha mutant, the nonlinearity in wild-type sun-flower and tobacco light intensity-response curves might havebeen attributed to experimental error. However, a similar pat-tern of response has been described by Liebig (10) in whichopening occurred at very low intensities, stomates closed par-tially at higher intensities, but at still higher intensities openedfurther. With comparative data for the nonphotosynthetic mu-tant available, a plausible hypothesis for this strange behavioris that in wild-type stomates two light-dependent reactions areinvolved in opening. Only one of these reactions (a low in-tensity response) is operational in the mutant. By next runninglight intensity-response curves in isolated portions of thespectrum, it was possible to demonstrate that one or the otherof the two postulated photoreactions could be activated selec-tively by utilizing the appropriate light quality.
Blue Light. The intensity-response curves of wild-type sun-flower and tobacco in blue light exhibit a pattern that will bereferred to here as the "high intensity" photoreaction of sto-mates, a response that is not seen in the xantha mutant (Fig. 3).There is a distinct threshold with a minimal light intensity ofapproximately 100 ttw/cm2 needed for opening. At higher in-tensities there is a fairly linear increase in stomatal aperture
Plant Physiol. Vol. 51, 1973 545
www.plantphysiol.orgon September 15, 2018 - Published by Downloaded from Copyright © 1973 American Society of Plant Biologists. All rights reserved.
HABERMANN
Light intensity (pW/cm2)
4.0
c 30o
E2.0
< 1.0 .
0I 200 400 600 800
Light intensity (pW/cm2)
-~3.0-2 A
. O2
E
20
800 0 200 400 600 800
Light intensity (p.W/cm2)
1 4.0
-~3.0-
E
2.0
.0
-A-- -8
0 200 400 600 800
Light intensity (PW/cm2)
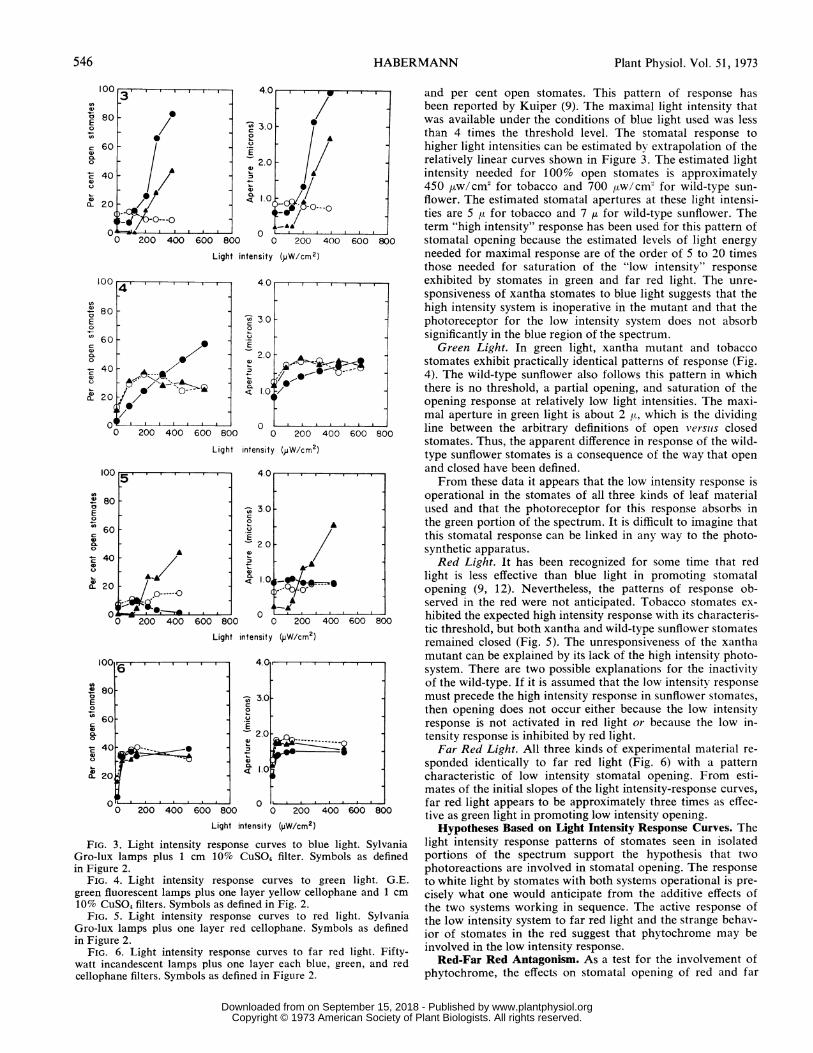
FIG. 3. Light intensity response curves to blue light. SylvaniaGro-lux lamps plus 1 cm 10% CuS04 filter. Symbols as definedin Figure 2.
FIG. 4. Light intensity response curves to green light. G.E.green fluorescent lamps plus one layer yellow cellophane and 1 cm10% CUS04 filters. Symbols as defined in Fig. 2.
FIG. 5. Light intensity response curves to red light. SylvaniaGro-lux lamps plus one layer red cellophane. Symbols as definedin Figure 2.
FIG. 6. Light intensity response curves to far red light. Fifty-watt incandescent lamps plus one layer each blue, green, and redcellophane filters. Symbols as defined in Figure 2.
Plant Physiol. Vol. 51, 1973
and per cent open stomates. This pattern of response hasbeen reported by Kuiper (9). The maximal light intensity thatwas available under the conditions of blue light used was lessthan 4 times the threshold level. The stomatal response tohigher light intensities can be estimated by extrapolation of therelatively linear curves shown in Figure 3. The estimated lightintensity needed for 100% open stomates is approximately450 /Aw/cm2 for tobacco and 700 fUw/cm2 for wild-type sun-flower. The estimated stomatal apertures at these light intensi-ties are 5 ,u for tobacco and 7 ,u for wild-type sunflower. Theterm "high intensity" response has been used for this pattern ofstomatal opening because the estimated levels of light energyneeded for maximal response are of the order of 5 to 20 timesthose needed for saturation of the "low intensity" responseexhibited by stomates in green and far red light. The unre-sponsiveness of xantha stomates to blue light suggests that thehigh intensity system is inoperative in the mutant and that thephotoreceptor for the low intensity system does not absorbsignificantly in the blue region of the spectrum.
Green Light. In green light, xantha mutant and tobaccostomates exhibit practically identical patterns of response (Fig.4). The wild-type sunflower also follows this pattern in whichthere is no threshold, a partial opening, and saturation of theopening response at relatively low light intensities. The maxi-mal aperture in green light is about 2 [,u which is the dividingline between the arbitrary definitions of open versus closedstomates. Thus, the apparent difference in response of the wild-type sunflower stomates is a consequence of the way that openand closed have been defined.From these data it appears that the low intensity response is
operational in the stomates of all three kinds of leaf materialused and that the photoreceptor for this response absorbs inthe green portion of the spectrum. It is difficult to imagine thatthis stomatal response can be linked in any way to the photo-synthetic apparatus.Red Light. It has been recognized for some time that red
light is less effective than blue light in promoting stomatalopening (9, 12). Nevertheless, the patterns of response ob-served in the red were not anticipated. Tobacco stomates ex-hibited the expected high intensity response with its characteris-tic threshold, but both xantha and wild-type sunflower stomatesremained closed (Fig. 5). The unresponsiveness of the xanthamutant can be explained by its lack of the high intensity photo-system. There are two possible explanations for the inactivityof the wild-type. If it is assumed that the low intensity responsemust precede the high intensity response in sunflower stomates,then opening does not occur either because the low intensityresponse is not activated in red light or because the low in-tensity response is inhibited by red light.Far Red Light. All three kinds of experimental material re-
sponded identically to far red light (Fig. 6) with a patterncharacteristic of low intensity stomatal opening. From esti-mates of the initial slopes of the light intensity-response curves,far red light appears to be approximately three times as effec-tive as green light in promoting low intensity opening.
Hypotheses Based on Light Intensity Response Curves. Thelight intensity response patterns of stomates seen in isolatedportions of the spectrum support the hypothesis that twophotoreactions are involved in stomatal opening. The responseto white light by stomates with both systems operational is pre-cisely what one would anticipate from the additive effects ofthe two systems working in sequence. The active response ofthe low intensity system to far red light and the strange behav-ior of stomates in the red suggest that phytochrome may beinvolved in the low intensity response.Red-Far Red Antagonism. As a test for the involvement of
phytochrome, the effects on stomatal opening of red and far
546
E 80E0
c 60Ck-0
-c 40
a- 20
0O'
100
c) 80E0
U) 60c
CL0
- 40
X¶ 20
is __-i.----Ijl f
10
8
6
4
2
U)
20E.2cvCL0
c4Du
-Va.
)0 5
30
'0 A
A-A0 4
0
0 200 400 600
E00
0
l-a!
www.plantphysiol.orgon September 15, 2018 - Published by Downloaded from Copyright © 1973 American Society of Plant Biologists. All rights reserved.

TWO PHOTOREACTIONS IN STOMATAL OPENING
red light in sequence were investigated. The experiments sum-marized in Table II were done with xantha mutant and wild-type sunflower leaf discs. The protocol involved exposure tored or far red light for a variable time (0-120 min) followed bythe standard 2-hr period in light of the other color. The con-ditions of illumination used (204 ,ew/cm2 red and 140 ,uw/cm2far red) were saturating for the low intensity opening in farred but only twice the threshold level of the high intensity re-sponse to red light exhibited by tobacco stomates. The resultsof these experiments can be summarized briefly as follows. (a)Red preceding far red inhibits opening. The greatest inhibitionis achieved by relatively short exposures to red (5-10 min)preceding 2 hr in far red. Longer preillumination (which sig-nificantly lengthens the total light period) is less inhibitory butmaximal opening is never significantly greater than in far redalone. (b) Far red preceding red promotes opening. The extentof opening after the final 2 hr of red light is proportional to thelength of prior exposure to far red. The greatest opening re-sponse occurred in 2 hr far red plus 2 hr red.
DISCUSSION
The results of these experiments clearly demonstrate a red-far red antagonism in the light-dependent opening of bothxantha and wild-type sunflower stomates. They suggest the in-volvement of phytochrome and also imply that there may be asingle mechanism for turgor movements in plants. The nycti-nastic movements of leaves were first shown to be underphytochrome control when Fondeville et al. (3) demonstratedthat a period of red prior to darkness hastens the closing move-
Table II. Effects ofRed and Far Red Light in Sequenzce on StomatalOpenin2g in Xanttha anid Wild-type Sunlflowers
Stomatal Response'
Length ofFirst Light, Open stomates Aperture
Period
Xantha Wild type Xantha I Wild-type
min % p|
A. Red preceding 2 hr far red2
0 39.8 + 3.3 33.8 i 2.3 1.8 0.1 1.5 4 0.15 14.8 i 4.7 4.8 i 1.3 1.2 ± 0.2 0.8 i 0.110 18.2 i 5.9 6.2 -+- 2.8 1.3 ±0.2 0.8 ± 0.215 22.2 ± 7.6 10.8 + 6.3 1.3 4 0.2 0.9 i 0.330 30.5 ± 12.6 11.0 4 6.0 1.5 + 0.2 1.0 4 0.260 23.8 ± 10.8 14.2 i 9.4 1.3 ± 0.2 '1.0 i 0.390 36.8 ± 2.6 38.5 ±4- 13.5 1.6 ± 0.1 1.5 ±4- 0.2120 52.0 + 8.5 34.5 + 14.9 2.0 + 0.3 1.5 i 0.3
B. Far red precedinzg 2 hr red2
0 8.8 ± 8.0 5.5 3.4 10.7 4 0.3 0.9 0.15 16.0± 6.31 5.2 1.6 1.240.2 0.8±0.1
10 21.8 9.5 10.0 5.7 1.3 + 0.2 0.9 4 0.215 38.8 i 8.9 18.2 ± 9.1 1.6 ± 0.2 1.1 ± 0.230 43.5 it 9.9 26.8 == 3.6 1.7 - 0.2 1.3 i 0.160 54.0 -- 10.6 26.2 it 1.9 1.9 = 0.3 1.3 - 0.190 61.0 -- 13.6 34.8 ±= 9.4 2.2 ± 0.3 1.5 -- 0.2120 76.5 == 5.2 51.2 ±t 15.7 2.7 ± 0.4 1.8 ± 0.3
C. Dark conttrols0 11.7 ±= 6.6 4.8 -- 0.8 0.9 ±t 0.4 0.9±0.1
1 Mean±
SD for four runs.2 Intensity of illumination: red, 209 MAw/cm2; far red, 140 4w/
cm2.
ment of Mimosa pudica leaves while far red prior to darknessdelays this movement. Satter et al. (16, 17) have shown thatin the nyctinastic movements of Albizzia leaflets opening is ac-companied by a flux of potassium ions from dorsal to ventralpulvinule cells while in closing there is a movement of potas-sium ions in the opposite direction.
Both the nyctinastic movements of leaves and stomatal open-ing and closing are complicated by the fact that they exhibitendogenous rhythms which are synchronized to diurnal cyclesof light and dark but can persist through periods of experimen-tally imposed constant light or constant dark conditions. It wasfrom studies of such rhythmic stomatal behavior that Mansfield(11) first demonstrated a low intensity light effect which causeda phase shift in the rhythm of stomatal opening ability. Themost effective wavelength for induction of such phase shiftswas found to be 703 nm, while comparable effects with redlight could be obtained only by continuous low intensity orintermittent high intensity interruption of the dark period.Mansfield considered the involvement of phytochrome in thelow intensity phase shift phenomenon, but he rejected thishypothesis because of the absence of a reversal of red effectsby far red and the requirement for frequent interruptions ofthe dark period for a phase shift response to be elicited.The interactions of red and far red on the low intensity re-
sponse are complex. There is no simple way to explain why along exposure to red preceding far red should be less inhibitorythan shorter exposures. However, sufficient interaction hasbeen demonstrated to support the proposed involvement ofphytochrome.The experiments reported here provide little information
about the photoreceptor for the high intensity photoresponseof stomates. Yet they do provide an explanation for the gen-erally poor response to red as compared to blue light. If thefunction of the low intensity response is, as Mansfield (11) hassuggested, one of effecting a "readiness to open" or, as we pre-fer to view it, to serve as an "unlocking mechanism," and ifthis response is inhibited by red light, then the generallygreater response of stomates to blue light is understandable.
Acknzowledgments-The author gratefully acknowledges the technical assistanceof Mrs. Penelope IMcKellar Wolkow, Mrs. Jennifer Chambers, MIrs. Doreen B.Sekulow, and 'Miss Shannon Beatty.
LITERATURE CITED
1. FISCHER, R. A. 1971. Role of potassium in stomatal opening in the leaf ofVicia faba. Plant Physiol. 47: 555-558.
2. FISCHER, R. A. AND T. C. HsIAo. 1968. Stomatal opening in isolated epidermalstrips of Vicia faba. II. Response to KCl concentration and the role ofpotassium absorption. Plant Physiol. 43: 1953-1958.
3. FONDEVILLE, J. C., H. A. BORTHWICK, AND S. B. HENDRICKS. 1966. Leafletmovement of Mimosa pudica L. indicative of phytochrome action. Planta69: 357-364.
4. HABERMANIN, H. MI. 1966. Light-inhibited leaf development in a whitemutant: resemblance to effects of 2-thiouracil in normally pigmentedHelianthus annuus. Physiol. Plant. 19: 122-127.
5. HABERNIANN, H. M. 1960. Spectra of normal and pigment-deficient mutantleaves of Helianthus annuus L. Physiol. Plant. 13: 718-725.
6. HABERMANN, H. MI. AND R. H. WALLACE. 1958. Transfer of flowering stimulusfrom stock to scion in grafted Helianthus annuus L. Amer. J. Bot. 45:479-482.
7. HUMIBLE, G. D. AND T. C. HSIAO. 1970. Light-dependent influx and effluxof potassium of guard cells during stomatal opening and closing. PlantPhysiol. 46: 483-487.
8. HuNIBLE, G. D. ANsD K. RASCHEKE. 1971. Stomatal opening quantitativelyrelated to potassium transport. Plant Physiol. 48: 447-453.
9. KTIPER, P. J. C. 1964. Dependence upon wavelength of stomatal movementin epidermal tissue of Senecio odoris. Plant Physiol. 39: 952-955.
10. LIEBIG, M. 1942. Untersuchungen uiber die Abha.ngigkeit der Spaltweite derStomata von IntensitSt und Qualitat der Strahlung. Planta 33: 206-257.
11. MIANSFIELD, T. A. 1965. The low intensity light reaction of stomata: effects ofred light on rhythmic stomatal behavior in Xanthium pennsylvanicum.Proc. Roy. Soc. Ser. B Biol. Sci. 162: 567-574.
Plant Physiol. Vol. 51, 1973 547
www.plantphysiol.orgon September 15, 2018 - Published by Downloaded from Copyright © 1973 American Society of Plant Biologists. All rights reserved.

HABERMANN
12. MEIDNER, H. 1968. The comparative effects of blue and red light on thestomata of Allium cepa L. and Xanthium pennsylvanicum. J. Exp. Bot.19: 146-151.
13. MEIDNER, H. AND T. A. MANSFIELD. 1965. Stomatal responses to illumination.Biol. Rev. 40: 483-509.
14. RAsCHKE, K. AND M. P. FELLOWS. 1971. Stomatal movement in Zea mays:
shuttle of potassium and chloride between guard cells and subsidiary cells.Planta 101: 296-316.
15. SAMPSON, J. 1961. A method of replicating dry or moist surfaces for examina-tion by light microscopy. Nature 191: 932-933.
16. SATTER, R. L. AND A. W. GALSTON. 1971. Potassium flux: a common featureof Albizzia leaflet movement controlled by phytochrome and by an endog-enous rhythm. Science 174: 518-520.
17. SATTER, R. L., P. MARINOFF, AND A. W. GALSTON. 1970. Phytochrome con-
Plant Physiol. Vol. 51, 1973
trolled nyctinasty in Albizzia julibrissin. II. Potassium flux as a basis forleaflet movement. Amer. J. Bot. 57: 916-926.
18. SAWHNEY, B. L. AND I. ZELITCH. 1969. Direct determination of potassium ionaccumulation in guard cells in relation to stomatal opening in the light.Plant Physiol. 44: 1350-1354.
19. SHAw, M. 1958. The physiology of stomata. II. Can. J. Bot. 36: 575-579.20. VIRGaN, H. I. 1956. Stomatal transpiration of etiolated wheat leaves as
related to chlorophyll a content. Physiol. Plant. 9: 482-493.21. VIRGIN, H. I. 1957. Stomatal transpiration of some variegated plants and of
chlorophyll-deficient mutants of barley. Physiol. Plant. 10: 170-186.22. WALLACE, R. H. AND H. M. HABERMANN. 1959. Genetic history and general
comparison of two albino mutations of Helianthus annuus. Amer. J. Bot.46: 157-162.
548
www.plantphysiol.orgon September 15, 2018 - Published by Downloaded from Copyright © 1973 American Society of Plant Biologists. All rights reserved.