evaluation of affymetrix array normalization procedures based on spiked crnas andrew hill expression...
TRANSCRIPT

Evaluation of Affymetrix array normalization procedures based on spiked cRNAs
Andrew Hill
Expression Profiling Informatics
Genetics Institute/Wyeth-Ayerst Research

October 11, 2001 2
Outline
• The GI/Harvard C. elegans array dataset as a normalization testbed
• Some general challenges of array data reduction• GeneChip Scaled Average Difference (ADs)
– the constant mean assumption• A purely spike-based normalization strategy
(Frequency)• A hybrid normalization (Scaled Frequency)• Conclusions

October 11, 2001 3
GI/Harvard C. elegans dataset• This data set used to evaluate several normalization procedures• Experiments:
– 8 developmental stages of the worm C. elegans were profiled, ranging from egg to adult worm
– n=2-4 replicate hybridizations for most array designs at most stages– 52 total arrays
• Arrays:– Three custom worm GeneChip designs (A, B, and C)– Each array monitors between 5700-6700 ORFs, in aggregate ~98% of the
worm genome– Chip A: ORFs with cDNA/EST matches in AceDB– Chips B/C: other ORFs– Several worm ORFs tiled on all 3 arrays for across-array-design comparisons
Science 290 809-812; Genome Biology (in the press)

October 11, 2001 4
Some challenges of Affymetrix GeneChip data reduction
• Array data from Affymetrix GeneChip sofware (pre-MAS 5.0):– negative low intensity signals
– lack of across-design normalization standard
– limited QC information
• Spike-based normalization methods can help to address each of these challengesNormalization: array scaling of average difference data from multiple
arrays/designs to minimize technical noise among arrays
• Current “standard” normalization procedure is a global scaling procedure: the GeneChip scaled average difference (ADs)

October 11, 2001 5
GeneChip Scaled Average Difference (ADs)
• The trimmed (2%) mean intensity of all probesets on all arrays is scaled to a constant target level.
• Works well in many cases (e.g. replicates)
• Some obvious situations where the “constant mean assumption” may not be well supported.

October 11, 2001 6
Constant mean assumption: problematic cases
•Chips monitoring a “small” fraction of transcriptome
•Non-random gene selection on arrays (e.g. C. elegans A vs. B/C)
•Large biological variation in expression

October 11, 2001 7
A cRNA spike-based normalization procedure (Frequency)
• Add 11 biotin-labeled cRNA spikes to each hybridization cocktail
• Construct a calibration curve• Use the Absent/Present calls for the
spikes to estimate array sensitivity• Dampen AD signals below the sensitivity
level to eliminate negative AD values.

October 11, 2001 8
Spiked Transcript ATCC Accession Affymetrix Gene Qualifier Final concentration (pMol) Final concentration (ppm)
DAPM 87826 AFFX-DapX-M_at 30 950
DAP5 87827 AFFX-DapX-5_at 10 317
CRE5 87832 AFFX-CreX-5_at 5 158
BIOB5 87825 AFFX-BioB-5_at 2.5 79
BIOD3 87830 AFFX-BioDn-3_at 1.2 38
BIOB3 87828 AFFX-BioB-3_at 0.6 19
CRE3 87835 AFFX-CreX-3_at 0.4 13
BIOC5 87833 AFFX-BioC-5_at 0.3 10
BIOC3 87834 AFFX-BioC-3_at 0.2 6
DAP3 87831 AFFX-DapX-3_at 0.15 5
BIOBM 87829 AFFX-BioB-M_at 0.1 3
Eleven spiked cRNAs

October 11, 2001 9
Figure 2Response to spikes over 2.5 log range
•Fit response with S-plus GLM, gamma error model, zero intercept.
•Power law fit AD=kFn yields n=0.93
•cRNA mass, scanner PMT gain are important determinants of response

October 11, 2001 10
Chip sensitivity calculation
0 1 2 3 4 5 6 7
0.0
0.2
0.4
0.6
0.8
1.0
log(frequency)
A/P
ca
ll
PPPPPPP
A
P
AA
•Consider A/P calls as binary response against log(known frequency)•Compute sensitivity as 70% likelihood level by either interpolation or logistic regression•“Dampen” computed frequencies below sensitivity:
•F < 0: F’ = avg(0,S)•0<F<S: F’=avg(F,S)

October 11, 2001 11
How well does it work?

October 11, 2001 12
Reproducibility of F metric (A array)
Absent Present0
0.5
1ADADs
F
ADADs F
0hM
ED
AC
V
Absent Present0
0.5
1
ADADs
F
ADADs
F
36h
ME
DA
CV
Absent Present0
0.5
1 ADADs
F
ADADs
F
48h
ME
DA
CV
Absent Present0
0.5
1ADADs
FAD
ADsF
60h
ME
DA
CV

October 11, 2001 13
Example of spike-skewed hybridization (36 hr sample)
0 500 1000 1500 20000
200
400
600
800
1000
1200
1400
1600
1800
2000
frequency 36h
freq
uenc
y 36
h
Worm GenescRNA spikes •cRNA spikes are
well normalized at the expense of worm genes
• Suggests inconsistency between ratio of spikes to worm cRNA across samples: spike skew

October 11, 2001 14
Sources of spike skew• Actual concentration of spikes may not be
nominal due to variation in cRNA “purity”
• Causes: liquid handling of small microlitre volumes, side reactions in cDNA/IVT process produce UV-absorbing, non-hybridizable contaminants
• Result: random per-hybe noise term introduced into normalized frequencies

October 11, 2001 15
An alternative hybrid normalization:
Scaled frequency (Fs)
• Need to reduce or eliminate spike skew as a source of experimental variation in normalized frequencies
• Average the globally scaled spike response over a complete set of arrays

October 11, 2001 16
Scaled frequency description
• Define a set of arrays
• Compute ADs for all arrays
• Pool spike responses and fit single model to pooled response
• Calibrate all arrays with single calibration factor
• Compute array sensitivity and dampen frequencies as in the frequency approach.

October 11, 2001 17
A pooled, scaled spike response
P
PP
PPP
P
PP
P
PP
PPP
P
P
P
PP
P
PP
PP
P
P
P
PP
P
PP
P
PP
P
P
P
PP
P
P
PP
PP
P
P
P
PP
PP
PPP
PP
P
P
PP
P
P
P
PPP
P
PP
P
P
P
P
PP
P
P
PP
P
P
PPPP
PP
P
PP
P
PP
PP
P
P
P
PP
P
PP
PPP
P
P
P
PP
P
P
P
PPP
P
P
PP
P
P
PP
PP
P
P
P
PP
P
P
PPP
P
P
P
P
PP
P
PP
PPP
PP
P
PP
P
P
PPP
P
P
P
PP
P
P
PPPP
P
P
PP
P
P
PPPP
P
P
P
PP
P
PP
P
PP
P
PP
P
PP
PPP
P
P
P
PP
P
PP
PPP
P
P
1 2 3
12
34
log10 ppm
log
10
ave
rag
e_
diff
ere
nce
fitted slope: 0.146162419368372
•Fit response with S-plus GLM, gamma error model, zero intercept.
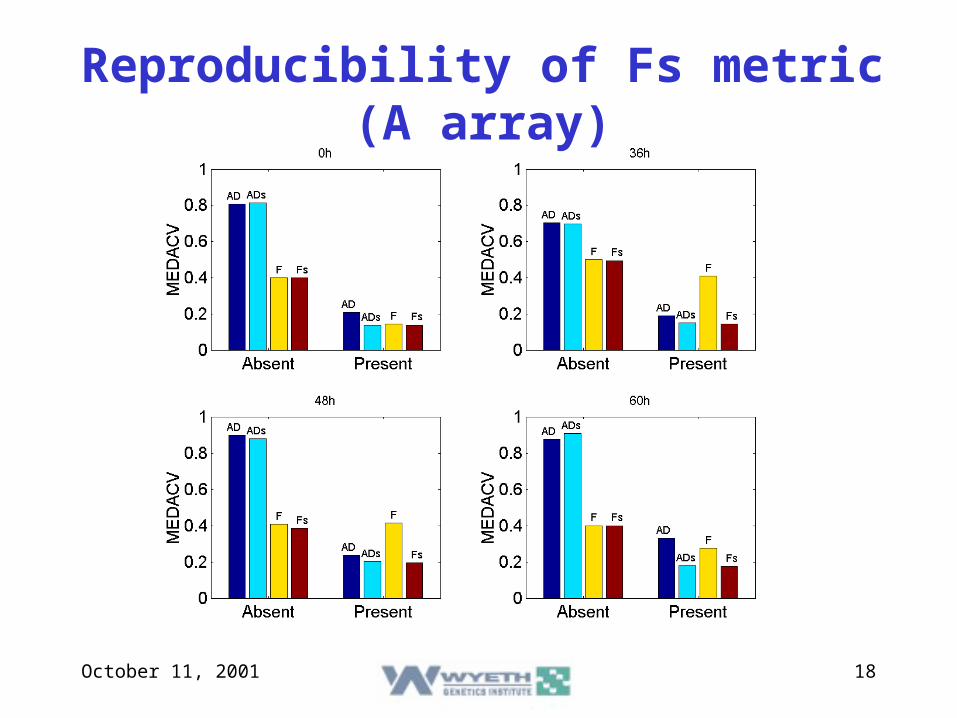
October 11, 2001 18
Reproducibility of Fs metric (A array)
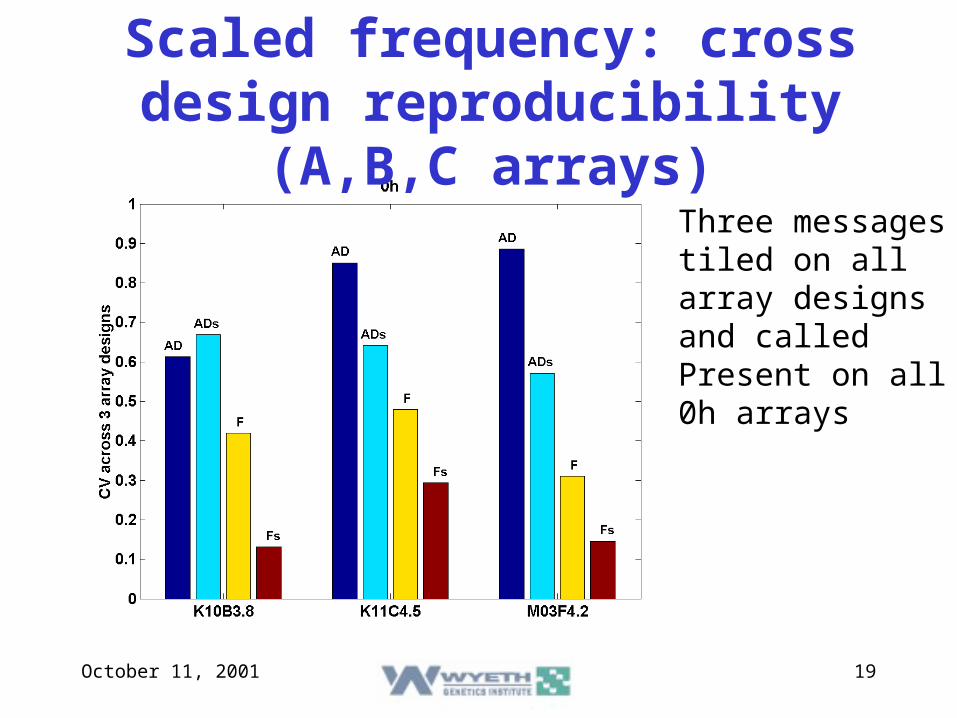
October 11, 2001 19
Scaled frequency: cross design reproducibility (A,B,C arrays)
Three messages tiled on all array designs and called Present on all 0h arrays

October 11, 2001 20
Conclusions
• Array response to spiked cRNAs can be close to linear over 2.5 logs of concentration.
• A chip sensitivity metric can be computed from Absolute Decisions associated with spikes; a very useful QC metric.
• Normalization based only on spikes performs inconsistently in some cases due to ill-quantitation of cRNAs, but can still be valuable when constant-mean assumption is violated. Better cRNA quantitation and process control will help.
• A hybrid approach based on global scaling and spikes performs the same as global AD scaling for single designs, and also allows cross-design comparisons

October 11, 2001 21
Acknowledgements
• Donna Slonim
• Maryann Whitley
• Yizheng Li
• Bill Mounts
• Scott Jelinsky
• Gene Brown
Harvard University:•Craig Hunter•Ryan Baugh

October 11, 2001 22
Extra slides follow ( not part of presentation)

October 11, 2001 23
Simulations (description)
• Simulations were performed
• Governing equation:
ijijijjiijij r s m a ADB bAD

October 11, 2001 24
Figure 4
CV characteristics of simulated data

October 11, 2001 25
Simulations: spike skew degrades reproducibility of frequency (A array)

October 11, 2001 26
Figure 7
Simulations: spike skew degrades accuracy of frequency