estrÉs agudo en ratas y su efecto en la adquisiciÓn...
TRANSCRIPT

1
ESTRÉS AGUDO EN RATAS Y SU EFECTO EN LA ADQUISICIÓN,
CONSOLIDACIÓN Y EXTINCIÓN DE LA MEMORIA ESPACIAL: PAPEL DE
LA PROTEÍNA QUINASA ERK1/2 Y DE LAS PROTEÍNAS FOSFATASAS PP1
Y PP2B EN EL HIPOCAMPO.
Paola Ortiz Montero
Universidad Nacional de Colombia
Facultad de Medicina
Unidad de Fisiología.
Bogotá D.C.
2011

2
ESTRÉS AGUDO EN RATAS Y SU EFECTO EN LA ADQUISICIÓN,
CONSOLIDACIÓN Y EXTINCIÓN DE LA MEMORIA ESPACIAL: PAPEL DE
LA PROTEÍNA QUINASA ERK1/2 Y DE LAS PROTEÍNAS FOSFATASAS PP1
Y PP2B EN EL HIPOCAMPO.
Paola Ortiz Montero
Código: 598185
Trabajo de grado presentado para optar a título de magister en Fisiología
Dirigido por: Dr. Jean Paul Vernot, Ph.D.
Codirigido por: Francisco Alejandro Múnera Galarza, Ph.D.
Universidad Nacional de Colombia
Facultad de Medicina
Unidad de Fisiología.
Bogotá D.C.
2011

3
FORMATO ÚNICO DE PRESENTACIÓN DE TRABAJOS DE GRADO
UNIVERSIDAD NACIONAL DE COLOMBIA
TITULO
Estrés agudo en ratas y su efecto en la adquisición, consolidación y extinción de
la memoria espacial: papel de la proteína quinasa ERK1/2 y de las proteínas
fosfatasas PP1 y PP2B en el hipocampo.
TITLE
Acute stress in rats and their effect on acquisition, consolidation and extinction of
spatial memory: rol of the protein kinase ERK1/2 and protein phosphatases PP1
and PP2B in hippocampus.
RESUMEN
El estrés, dependiendo de la severidad y del contexto, puede causar efectos
negativos sobre el sistema nervioso central, los cuales varían desde cambios
cognoscitivos leves hasta el desencadenamiento de trastornos mentales. Los
efectos del estrés sobre el aprendizaje y la memoria han sido bien
documentados en un diverso arreglo de especies, desde peces a humanos.
Consecuentemente, los modelos animales pueden ser utilizados para entender
los cambios neurales que median los efectos del estrés. La prevención y el
tratamiento de los efectos nocivos del estrés sobre el sistema nervioso central
requieren de un conocimiento detallado de la naturaleza de la interacción entre
los mediadores de la respuesta de estrés y de los procesos neurales que
subyacen a los procesos cognoscitivos. El balance entre la actividad de
quinasas y fosfatasas en el cerebro, podría ser un posible mediador de ambos
procesos, puesto que la regulación del estado de fosforilación de proteínas,
constituye un mecanismo versátil de control de la función celular. La actividad de

4
diversas quinasas y fosfatasas en distintas regiones del cerebro, se ve
modificada de manera independiente por el estrés y por el aprendizaje; sin
embargo, hasta el momento no se han hecho estudios sistemáticos para
caracterizar la interacción de estos dos factores sobre el balance de actividad
entre estas proteínas en el sistema nervioso central.
Para caracterizar esta interacción se diseño un estudio, utilizando un modelo
animal, llevado a cabo por el grupo de aprendizaje y comportamiento animal, el
cual fue el encargado de inducir estrés agudo, corto y prolongado, mediante
restricción de movimientos, así como de la evaluación de los procesos de
aprendizaje, mediante el entrenamiento en una tarea espacial en el laberinto
circular de Barnes, en donde se evaluó la adquisición, recuperación y extinción
de memoria. Como indicador del balance de fosforilación en el sistema nervioso
central se determinó la concentración y la actividad enzimática de quinasas y
fosfatasas en el hipocampo. Finalmente se estableció una relación entre la
inducción de estrés y el entrenamiento espacial sobre la actividad de quinasas y
fosfatasas.
Los resultados de esta investigación sentarán las bases para nuevos estudios
destinados a comprender las interacciones moleculares desencadenadas por el
estrés y el aprendizaje en neuronas del sistema nervioso central.
PALABRAS CLAVES
Estrés, quinasas, fosfatasas, hipocampo, memoria espacial.

5
ABSTRACT
Stress, depending on its severity and context, can have negative effects on the
central nervous system, varying from mild cognitive changes to the onset of
mental disorders. The effects of stress on learning and memory have been well-
documented in a wide array of species from fish to humans. Consequently,
animal models can be used for understanding the neural changes mediating the
effects of stress. The prevention and treatment of the harmful effects of stress on
the central nervous system require a detailed knowledge of the nature of the
interaction between stress response mediators and knowledge of the neural
processes underlying cognitive events. The balance between kinase and
phosphatase activity in the brain could be a possible mediator of both processes,
since protein phosphorylation status regulation is a versatile control mechanism
for proper cell function. The activity of various kinases and phosphatases in
different regions of the brain is independently modified by stress and learning;
however, no systematic studies have been done so far to characterise these two
factors’ interaction regarding the balance of activity between these proteins in the
central nervous system.
A study using an animal model was thus designed for characterizing this
interaction. The study was carried out by the animal learning and behavior group
which was responsible for inducing short- and long-term acute stress by
restricting movement and evaluating learning through spatial task training in the
Barnes circular maze, where memory acquisition, retrieval and extinction were
assessed. Enzyme concentration and kinase and phosphatase activity in the
hippocampus was evaluated as an indicator of phosphorylation balance in the
central nervous system. A link between stress induction and spatial training
regarding kinase and phosphatase activity was established.
The results of this research will lay the foundations for further studies aimed at
understanding molecular interactions triggered by stress and learning in central
nervous system neurons.

6
FIRMA DEL DIRECTOR----------------------------------------------------------
Profesor: Jean Paul Vernot, Ph.D.
----------------------------------------------------------
Profesor: Alejandro Múnera, Ph.D.
NOMBRE COMPLETO DEL AUTOR Y AÑO DE NACIMIENTO:
Paola Esperanza Ortiz Montero (1984)

7
AGRADECIMIENTOS
Al Doctor Jean Paul Vernot, por su apoyo, enseñanza y excelentes sugerencias
durante el desarrollo de este trabajo de grado, por permitirme hacer parte de su
grupo de investigación y guiarme en mi formación investigativa.
Al Doctor Alejandro Múnera, por su enseñanza durante el desarrollo de este
trabajo.
A mis compañeros del laboratorio de Fisiología Celular y Molecular,
especialmente Ana María Perdomo, por acompañarme durante todo este
proceso, por sus enseñanzas y aportes en el desarrollo de esta tesis.
Al doctor Miguel Eduardo Martínez, por su apoyo durante mi formación como
Magister en Fisiología.
A Rocío Gonzales por su valiosa amistad y apoyo, a Diana Arias por toda su
ayuda y colaboración.

8
TABLA DE CONTENIDO.
Introducción
1. Planteamiento del problema
2. Marco teórico y estado del arte
2.1. Estrés y neurofisiología de la respuesta de estrés
2.2. Memoria Espacial
2.3. Efectos del estrés sobre la memoria espacial
2.4. Estado de fosforilación y control de la función celular
2.5. Memoria y fosforilación
2.6. Estrés y fosforilación
3. Justificación
4. Objetivos del estudio
4.1. General
4.2. Específicos
5. Metodología y diseño experimental
5.1. Animales
5.2. Preparación de extractos de hipocampo
5.2.1. Disección de hipocampos
5.2.2. Homogenización de los hipocampos
5.3. SDS-PAGE y Western blot
5.4. Ensayos de actividad enzimática
5.4.1. Actividad enzimática de proteínas fosfatasas
5.4.2. Actividad enzimática de ERK1/2
5.5. Diseño experimental
6. Resultados
6.1. Efecto del estrés por inmovilización en la expresión de ERK1/2 en el
hipocampo de ratas
6.2. Actividad y nivel de expresión de ERK1/2 durante los procesos de
formación de memoria
6.3. Efecto del estrés sobre la formación de memoria espacial y su relación
con la expresión de ERK1/2

9
6.4. Efecto del estrés por inmovilización en la expresión de PP2B en el
hipocampo de ratas
6.5. Actividad y nivel de expresión de PP2B durante los procesos de
formación de memoria
6.6. Efecto del estrés sobre la formación de memoria espacial y su relación
con la expresión de PP2B
6.7. Efecto del estrés por inmovilización en la expresión de PP1 en el
hipocampo de ratas
6.8. Actividad y nivel de expresión de PP1 durante los procesos de
formación de memoria
6.9. Efecto del estrés sobre la formación de memoria espacial y su relación
con la expresión de PP1
7. Discusión
7.1. Efecto del estrés por inmovilización sobre el nivel de expresión y la
actividad de ERK1/2, PP1 Y PP2B
7.2. Efecto del aprendizaje de una tarea de memoria espacial en el nivel
de expresión y la actividad de ERK1/2, PP1 Y PP2B
7.3. Efecto de la interacción del estrés agudo y los procesos de
aprendizaje en el nivel de expresión y la actividad de ERK1/2, PP1 Y
PP2B
7.4. Efectos comportamentales del estrés agudo inducido antes de la
adquisición de la memoria espacial
7.5. Efectos comportamentales del estrés agudo inducido antes de la
recuperación de la memoria espacial
7.6. Efectos comportamentales del estrés agudo inducido antes de la
extinción de la memoria espacial
8. Conclusiones
9. Anexos
10. Bibliografía

10
LISTA DE FIGURAS
Figura 1. Efectos fisiológicos del estrés en función de su nivel y duración. Figura 2. Corticosterona producida por estrés y sus blancos de acción en regiones críticas para la formación de memoria. Figura 3. Complejidad de las acciones de los glucocorticoides sobre la plasticidad sináptica. Figura 4. Receptores de glucocorticoides y procesamiento de información.
Figura 5. Rutas de transducción de señales que operan en el hipocampo durante los procesos de formación de memoria.
Figura 6. Cambios en la actividad enzimática de ERK1/2 provocados por el estrés por inmovilización.
Figura 7. Cambios en la actividad enzimática de ERK1/2 provocados por el estrés por inmovilización.
Figura 8. Cambios en la actividad enzimática de ERK1/2 inducidos por el entrenamiento de la adquisición de la memoria espacial.
Figura 9. Cambios en la actividad enzimática de ERK1/2 inducidos por el entrenamiento de la recuperación de la memoria espacial.
Figura 10. Cambios en la actividad enzimática de ERK1/2 inducidos por el entrenamiento de la extinción de la memoria espacial.
Figura 11. Cambios en la actividad enzimática de ERK1/2 inducidos durante las tres fases de la memoria espacial.
Figura 12. Cambios en el nivel de expresión de ERK1/2 provocados por el entrenamiento de la memoria espacial.
Figura 13. Cambios en la actividad enzimática de ERK1/2 inducidos por el estrés corto previo al entrenamiento en la adquisición de la memoria espacial.
Figura 14. Cambios en la actividad enzimática de ERK1/2 inducidos por el estrés prolongado previo al entrenamiento en la adquisición de la memoria espacial.
Figura 15. Cambios en la actividad enzimática de ERK1/2 inducidos por el estrés corto y prolongado previo al entrenamiento en la adquisición de la memoria espacial.

11
Figura 16. Cambios en la actividad enzimática de ERK1/2 inducidos por el estrés corto previo al entrenamiento en la recuperación de la memoria espacial.
Figura 17. Cambios en la actividad enzimática de ERK1/2 inducidos por el estrés prolongado previo al entrenamiento en la recuperación de la memoria espacial.
Figura 18. Cambios en la actividad enzimática de ERK1/2 inducidos por el estrés corto y prolongado previo al entrenamiento en la recuperación de la memoria espacial.
Figura 19. Cambios en el nivel de expresión de ERK1/2 inducidos por el estrés corto y prolongado previo al entrenamiento en la memoria espacial.
Figura 20. Cambios en la actividad enzimática de ERK1/2 inducidos por el estrés corto previo al entrenamiento en la extinción de la memoria espacial.
Figura 21. Cambios en la actividad enzimática de ERK1/2 inducidos por el estrés prolongado previo al entrenamiento en la extinción de la memoria espacial.
Figura 22. Cambios en la actividad enzimática de ERK1/2 inducidos por el estrés corto y prolongado previo al entrenamiento en la extinción de la memoria espacial.
Figura 23. Cambios en el nivel de expresión de ERK1/2 inducidos por el estrés corto previo al entrenamiento de la extinción de la memoria espacial.
Figura 24. Cambios en el nivel de expresión de ERK1/2 provocados por el estrés prolongado previo al entrenamiento en la extinción de la memoria espacial.
Figura 25. Cambios en el nivel de expresión de PP2B provocados por el estrés por inmovilización corto y prolongado.
Figura 26. Cambios en la actividad enzimática de PP2B provocados por el estrés por inmovilización.
Figura 27. Cambios en la actividad enzimática de PP2B provocados por el estrés por inmovilización.
Figura 28. Cambios en la actividad enzimática de PP2B inducidos por el entrenamiento de la adquisición de la memoria espacial. Figura 29. Cambios en la actividad enzimática de PP2B inducidos por el entrenamiento de la recuperación de la memoria espacial. Figura 30. Cambios en la actividad enzimática de PP2B inducidos por el entrenamiento de la extinción de la memoria espacial.

12
Figura 31. Cambios en la actividad enzimática de PP2B inducidos por el entrenamiento en la memoria espacial. Figura 32. Cambios en el nivel de expresión de PP2B inducidos por el entrenamiento en la recuperación de la memoria espacial. Figura 33. Cambios en la actividad enzimática de PP2B inducidos por el estrés previo al entrenamiento en la adquisición de la memoria espacial. Figura 34. Cambios en la actividad enzimática de PP2B inducidos por el estrés previo a la adquisición de la memoria. Figura 35. Cambios en el nivel de expresión de PP2B inducidos por el estrés corto y prolongado previo al entrenamiento de la adquisición de la memoria espacial. Figura 36. Cambios en la actividad enzimática de PP2B inducidos por el estrés previo al entrenamiento en la recuperación de la memoria espacial. Figura 37. Cambios en la actividad enzimática de PP2B inducidos por el estrés previo a la recuperación de la memoria. Figura 38. Cambios en el nivel de expresión de PP2B provocados por el estrés corto y prolongado previo al entrenamiento en la recuperación de la memoria espacial. Figura 39. Cambios en la actividad enzimática de PP2B inducidos por el estrés previo al entrenamiento en la extinción de la memoria espacial. Figura 40. Cambios en la actividad enzimática de PP2B inducidos por el estrés previo al entrenamiento en la extinción de la memoria espacial. Figura 41. Cambios en el nivel de expresión de PP2B inducidos por el estrés corto y prolongado previo al entrenamiento en la extinción de la memoria espacial. Figura 42. Cambios en la actividad enzimática de PP1 provocados por el estrés por inmovilización. Figura 43. Cambios en el nivel de expresión de PP2B provocados por el estrés por inmovilización corto y prolongado. Figura 44. Cambios en la actividad enzimática de PP1 inducidos por el entrenamiento de la memoria espacial. Figura 45. Cambios en la actividad enzimática de PP1 inducidos por el entrenamiento de la recuperación de la memoria espacial.

13
Figura 46. Cambios en la actividad enzimática de PP1 inducidos por el entrenamiento de una tarea espacial.
Figura 47. Cambios en el nivel de expresión de PP1 inducidos por el entrenamiento en una tarea espacial. Figura 48. Cambios en la actividad enzimática de PP1 inducidos por el estrés previo al entrenamiento en la adquisición de la memoria espacial.
Figura 49. Cambios en la actividad enzimática de PP1 inducidos por el estrés previo a la adquisición de la memoria.
Figura 50. Cambios en el nivel de expresión de PP1 inducidos por el estrés corto y prolongado previo al entrenamiento de la adquisición de la memoria espacial.
Figura 51. Cambios en la actividad enzimática de PP1 inducidos por el estrés previo al entrenamiento en la recuperación de la memoria espacial.
Figura 52. Cambios en la actividad enzimática de PP1 inducidos por el estrés previo a la recuperación de la memoria.
Figura 53. Cambios en el nivel de expresión de PP1 inducidos por el estrés corto y prolongado previo al entrenamiento de la recuperación de la memoria espacial.
Figura 54. Cambios en la actividad enzimática de PP1 inducidos por el estrés previo al entrenamiento en la extinción de la memoria espacial.
Figura 55. Cambios en la actividad enzimática inducidos por el estrés previo a la extinción de la memoria.
Figura 56. Cambios en el nivel de expresión de PP1 inducidos por el estrés corto y prolongado previo al entrenamiento en la extinción de la memoria espacial.
Figura 57. Rutas de señalización activadas en respuesta al estrés.
Figura 58. Activación de ERK1/2 en el hipocampo durante los procesos de aprendizaje y formación de memoria.
Figura 59. Rutas de señalización que conduce a un cambio en la actividad de PP1 en el hipocampo durante los procesos de aprendizaje y formación de memoria.
Figura 60. Efecto de los glucocorticoides en la transcripción génica.

14
LISTA DE ANEXOS
Figura A1. Inducción de estrés agudo por restricción de movimientos y entrenamiento de la memoria espacial. Figura A2. Cambios en el nivel de expresión de ERK1/2 provocados por el estrés por inmovilización. Figura A3. Nivel de expresión de ERK1/2 en condiciones de recuperación y extinción de la memoria y sacrificio inmediato. Figura A4. Nivel de expresión de ERK1/2 1h después de finalizado el entrenamiento en la recuperación y la extinción de la memoria. Figura A5. Nivel de expresión de ERK1/2 4h después de finalizado el entrenamiento en la recuperación y la extinción de la memoria. Detección de ERK1/2 por WB. Figura A6. Nivel de expresión de PP2B en condiciones de estrés. Figura A7. Nivel de expresión de PP2B en condiciones de recuperación de la memoria espacial. Figura A8. Nivel de expresión de PP2B en condiciones de estrés corto previo a la extinción de la memoria espacial. Figura A9. Nivel de expresión de PP2B en condiciones de estrés prolongado previo a la extinción. Figura A10. Diagrama de barras ilustrando la actividad enzimática de ERK1/2, PP1 y PP2B en cada uno de los tratamientos.
Tabla 1. Análisis de varianza de la actividad de ERK1/2 entre los grupos experimentales analizados.
Tabla 2. Análisis de varianza de la actividad de PP2B entre los grupos experimentales analizados.
Tabla 3. Análisis de varianza de la actividad de PP1 entre los grupos experimentales analizados.

15
INTRODUCCIÓN
1. PLANTEAMIENTO DEL PROBLEMA
El estrés es una respuesta sistémica y coordinada por la activación hormonal y
autonómica de la glándula suprarrenal, que en algunos casos, le permite al
organismo adaptarse a situaciones que amenazan su integridad, pero cuando se
vuelve severo puede lesionar distintos órganos, incluyendo el cerebro (Pacák y
Palkovits, 2001).
La memoria es el proceso mediante el cual el organismo adquiere, retiene y
recupera información relevante acerca de su entorno y se fundamenta en
cambios duraderos en el cerebro; se ha demostrado que el estrés puede alterar
los procesos de memoria en función de su intensidad, duración y el momento en
que ocurre (Olton, 1977). La estructura central del cerebro, encargada de la
formación, consolidación y recuperación de diversos tipos de memoria, así como
la memoria espacial, es el hipocampo, que además es susceptible a los efectos
deletéreos del estrés, lo que la hace una estructura blanco en estudios acerca
de la interacción entre el estrés y la memoria (Pacák y Palkovits, 2001).
Tanto el estrés como la adquisición de memorias provocan cambios duraderos
en la función celular en el sistema nervioso central, en gran medida mediante
cambios en el balance funcional entre quinasas y fosfatasas en el tejido
nervioso; según el estímulo, estas enzimas agregan (quinasas) o eliminan
(fosfatasas), grupos de fosfato a diversas proteínas, induciendo cambios en las
propiedades funcionales de está y de la célula (Malenka, 2003). El balance de la
actividad de estas enzimas esta finamente regulado y puede ser alterado por
diversos procesos fisiológicos, como el estrés o la memoria; sin embargo la
interacción entre el estrés y la memoria sobre el balance funcional entre
quinasas y fosfatasas ha sido poco estudiado, se sabe que la exposición al
estrés esta asociada con alteraciones en el aprendizaje y la formación de
memoria tanto en animales experimentales como en humanos.

16
Un conocimiento detallado de estos mecanismos permitirá señalar posibles
blancos para intervenciones terapéuticas dirigidas a prevenir o remediar los
efectos deletéreos del estrés sobre la neurobiología de los procesos
cognoscitivos.
2. MARCO TEÓRICO Y ESTADO DEL ARTE
2.1. Estrés y neurofisiología de la respuesta de estrés.
En la actualidad el estrés es un término muy popularizado, el cual es visto desde
diferentes perspectivas, como un estímulo, una respuesta o una consecuencia.
Según Selye H. (1936), se concibe como una respuesta adaptativa del
organismo ante los diversos agentes inductores de estrés. Selye también
describió el "síndrome general de adaptación" (SGA), conjunto de alteraciones
sistémicas que ocurren en respuesta a estímulos perturbadores de diversa
naturaleza y posteriormente, introdujo el término "estrés" para designar la
respuesta del organismo durante el SGA y el término "estímulo o agente
estresante" para nombrar el evento desencadenante. Por otra parte Cannon
(1929 a, b), con su definición de “homeostasis”, postuló que la médula
suprarrenal y el sistema nervioso autónomo ejercían un papel crucial para la
ejecución de este tipo de respuestas. Pacák y Palkovits (2001) plantearon que el
estrés podría concebirse como la respuesta a una amenaza a la homeostasis
producida por estímulos que ocasionan daño tisular, o por estímulos percibidos
que permiten prever la posibilidad de que se produzca lesión, los cuales
desencadena respuestas compensatorias en el organismo, las cuales ocurren
por activación de circuitos neuronales específicos.
La respuesta al estrés varía en función de: 1) la intensidad y duración del
estímulo; 2) el estado funcional del organismo; y, 3) la apreciación del sujeto y
de su capacidad para predecir y controlar tal estímulo (Jeansok y col., 1998)

17
(Figura 1). La variabilidad del estado funcional y de la apreciación del sujeto
conduce a que la respuesta de estrés ante un mismo estímulo sea diferente en
individuos distintos e, incluso, en un mismo individuo (López-Calderón, 1999).
Los estímulos estresantes, por su parte, pueden ser de diversa naturaleza: 1)
alteraciones del medio interno; 2) modificaciones ambientales; o 3) estados
emocionales. A nivel del sistema nervioso, estos eventos estresantes
desencadenan la activación del sistema simpático-adrenomedular, la cual
desencadena la liberación de la hormona liberadora de corticotropina (CRH) a
partir del núcleo paraventricular del hipotálamo, y la consecuente liberación
masiva de catecolaminas. La actividad del sistema simpático-adrenomedular
debe coordinarse con la actividad del sistema parasimpático, del eje hipotálamo-
hipófisis-suprarrenal y de otros sistemas de neuropéptidos. Probablemente, el
centro coordinador de estos sistemas efectores sea el hipotálamo. No obstante,
los mecanismos íntimos por los cuales se regula la respuesta global de estrés
no están completamente establecidos.
Figura 1. Efectos fisiológicos del estrés en función de su nivel y duración. Se muestran los
daños fisiológicos que pueden ocurrir en respuesta a diferentes niveles de estrés leve,
moderado, fuerte o crónico, dependiendo también de su duración, transitoria o permanente.
(tomado de Jeansok J. Kim and Kenneth S. Yoon, Trends Neurosci. (1998) 21, 505–09).

18
El eje Hipotalamo-Hipófisis-Suprarrenal (HPA, por la sigla inglesa de
Hypothalamus-Pituitary-Adrenal), se activa inmediatamente después de la
activación del sistema simpático-adrenomedular; la liberación de CRH, estimula
la liberación de hormona adrenocorticotrópica (ACTH, por la sigla inglesa de
Adreno-Cortico-Tropic Hormone). Una vez liberada, la ACTH actúa sobre la
corteza suprarrenal estimulando la secreción de glucocorticoides y,
parcialmente, de mineralocorticoides. Además de su acción en las
suprarrenales, la ACTH promueve la lipólisis y estimula la captación de glucosa
y aminoácidos por el tejido muscular. Así, la activación del eje HPA permite la
rápida movilización de energía almacenada y deprime procesos anabólicos e
inmunitarios; durante todo el periodo de estrés, se detectan niveles elevados de
CRH, ACTH y cortisol. (Jacobson y col., 1991). Por otra parte el sistema
nervioso central, participa de una manera global en el mantenimiento de la
homeostasis y en la organización de la respuesta de estrés. El sistema límbico
(especialmente amígdala e hipocampo) constituye, uno de los mayores circuitos
de regulación de la respuesta de estrés y de los comportamientos asociados
(Pacák y Palkovits, 2001).
El estrés, puede ser inducido experimentalmente por medio de dos
procedimientos: sistémicos o procesales (Anisman y col., 2001). La inducción
sistémica de estrés se logra mediante amenazas inmediatas a la homeostasis,
por ejemplo: desafíos inmunológicos, hemorragias, intoxicación alcohólica,
intoxicación con éter o hipoglicemia. Al parecer, estos eventos estresantes
estimulan al núcleo paraventricular del hipotálamo a través de proyecciones
monoaminérgicas del tallo cerebral, así mismo, la inducción procesal de estrés
requiere la integración e interpretación de eventos estresantes en centros
cerebrales superiores, con participación de circuitos límbico-prosencefálicos que
vinculan la corteza prefrontal, el hipocampo y la amígdala (Herman y Cullinan,
1997). Los eventos estresantes procesales pueden ser psicológico-
psicogénicos, como la exposición a un predador o un campo abierto novedoso,
o físico-neurogénicos como choques eléctricos en las patas, nado en agua fría o

19
inmovilización. Se ha demostrado que estas dos estrategias son igualmente
efectivas, en cuanto a la inducción de estrés.
2.2. Memoria espacial.
La memoria es el proceso cognoscitivo a través del cual los organismos
almacenan información relevante de sí mismos y del medio que los rodea
(Olton, 1977). Este proceso se encuentra estrechamente relacionado con el
aprendizaje, que es la modificación del comportamiento individual como
resultado de la experiencia (Thorpe, 1963). El aprendizaje, entonces, es el
proceso de adquisición de información que es almacenable y recuperable. La
formación de nuevas memorias sigue un curso temporal característico: 1)
Adquisición de información; 2) Consolidación o establecimiento de la
información recientemente adquirida; 3) Recuperación o reactivación de la
información previamente almacenada y de aquellas respuestas que demuestran
aprendizaje. Cada vez que se activa, la memoria entra en un período de
labilidad que puede concluir con el refuerzo (reconsolidación) o la extinción de
dicha memoria. Durante el proceso de reconsolidación, la memoria pasa de un
estado lábil a un estado permanente y duradero (Pedreira y Maldonado, 2003).
Durante el proceso de extinción hay una modificación del comportamiento en
donde el animal aprende activamente que una asociación de estímulos
previamente adquirida deja de tener validez. Por último, de no activarse, la
memoria es susceptible al olvido, que es la pérdida de la capacidad para activar
la información previamente aprendida (Torres, 2006). Así mismo, la formación y
el almacenamiento de memoria a largo plazo es el resultado de cambios
transitorios en la eficiencia neuronal seguido de modificaciones de más larga
duración a nivel morfológico y funcional, en varias áreas del cerebro, incluyendo
prominentemente el hipocampo (Izquierdo y col., 2006; Dudai, 2002).
La mayoría de los eventos moleculares que participan en la formación de
memorias han sido caracterizados, ellos involucran la activación de diferentes

20
sistemas de neurotransmisores y de rutas de señalización intracelular, las
cuales regulan los mecanismos celulares que controlan la expresión génica y
subsecuentemente la síntesis de nuevas proteínas que participan en la
remodelación y en el crecimiento de los procesos sinápticos. Estas
modificaciones ocurren como un resultado de un proceso dependiente de la
síntesis proteínas en donde la memoria llega a ser progresivamente estabilizada
(Izquierdo y col., 2006).
La memoria espacial es un tipo de memoria cuyo objetivo final es el
almacenamiento de la representación espacio-perceptual de los lugares a los
que se expone el organismo durante toda su vida (Carvajal, 2006). Evidencias
clínicas y experimentales indican que múltiples áreas cerebrales intervienen en
los procesos de memoria y aprendizaje. Una de estas áreas, el hipocampo, es
crucial para la adquisición de memorias declarativas, particularmente para la
adquisición de memorias espaciales (Mishkin, 1978). En este estudio, el
hipocampo fue la estructura escogida debido a que, no solo regula los procesos
de aprendizaje y formación de memoria, sino que también tiene un papel
importante en el aprendizaje de una tarea espacial. En efecto, se ha demostrado
que pequeñas lesiones en el hipocampo (30%) deterioran la memoria espacial
(Broadbent y col., 2004). En roedores, la memoria espacial, en general, y la
participación del hipocampo en la misma, en particular, han sido evaluadas
mediante diferentes paradigmas de aprendizaje: el laberinto acuático de Morris
(Morris, 1984); el laberinto de brazos radiales (Olton, 1977, 1979); el laberinto
circular de Barnes (Barnes, 1979).
2.3. Efectos del estrés sobre la memoria espacial
Durante las últimas dos décadas se ha encontrado que, en humanos y animales,
tanto la respuesta de estrés como la elevación de las hormonas relacionadas
con el estrés afectan la memoria dependiente del hipocampo, así, se sabe que
la memoria puede ser incrementada por experiencias estresantes agudas (Shors

21
y col., 1992; Radulovic y col., 1999; Roozendaal, 2002), incluso cuando el
entrenamiento es realizado después de que ha terminado la respuesta al estrés.
En personas con trastorno por estrés postraumático se ha encontrado
disminución del volumen del hipocampo asociado con deficiencias en tareas de
memoria dependientes del hipocampo (Utto y col., 1993).
En respuesta al estrés, los glucocorticoides entran al cerebro y se unen a 2 dos
subtipos de receptores, los receptores de mineralocorticoides (MR) y los
receptores de glucocorticoides (GR), los cuales se localizan en regiones del
cerebro que son importantes en los procesos de formación de memoria, como lo
es el hipocampo, la amígdala y la corteza prefrontal (de Kloet, 2005) (Figura 2).
Figura 2. Corticosterona producida por estrés y sus blancos de acción en regiones
críticas para la formación de memoria. Se ilustran las regiones del cerebro (Amígdala,
Hipocampo y Corteza prefrontal) que son susceptibles a ser modificadas por el estrés (Tomado
de de Kloet ER, Joels M, Holsboer F. Nature Rev Neurosci (2005). 6: 463-4).
La activación de los MRs, mantiene la transmisión glutaminergica en el área
CA1 del hipocampo y facilita la potenciación a largo plazo (LTP, long-term
potentiation) pero únicamente cuando la corticoesterona es presente alrededor
del tiempo en la que la LTP es inducida; la rapidez del efecto de la
corticoesterona indica que esto está relacionado con rutas no genómicas de

22
activación; mientras que altas cantidades de corticoesterona asociadas a un
estrés severo pueden suprimir la LTP, la transmisión sináptica y la plasticidad a
través de acciones genómicas mediadas por la activación de GRs, normalizando
de esta manera el aumento en la actividad neuronal inducida por el estrés (Joëls
y col., 2008) (Figura 3).
Figura 3. Complejidad de las acciones de los glucocorticoides sobre la plasticidad
sináptica. Se muestran los efectos a través del tiempo de la corticosterona y su acción sobre los
receptores de glucocorticoides (GR) y mineralocorticoides (MR) en la potenciación o depresión a
largo plazo, las cuales son medidas de la plasticidad sináptica. LTP (long term potentiation); LTD
(long term depression). (Tomado de: Krugers Harmen J, Casper C.Hoogenraad and Laurent Gro
Nature Reviews Neuroscienc. (2010). 11: 675-81).
A nivel comportamental, varios estudios en roedores, han mostrado que los MRs
ejercen un papel fundamental en la reactividad comportamental en situaciones
de novedad, en la apreciación de información y en la selección de una
respuesta, mientras que los GRs participan en la consolidación de información.
Se ha demostrado previamente, en diferentes tareas de aprendizaje (Laberinto
acuático de Morris, tareas de evitación inhibitoria y condicionamiento al miedo),
que los glucocorticoides promueven la consolidación a largo plazo de memoria
espacial dependiente de hipocampo y la información emocional a través de la

23
activación de GRs. También hay evidencia que sugiere que los GRs asociados a
membrana promueven la memoria a largo plazo en una tarea de reconocimiento
de objetos a través de modificaciones en la cromatina. Otros estudios, han
mostrado que los glucocorticoides incrementan la eficiencia sináptica mediante
la regulación del tráfico de receptores AMPA
(α-amino-3-hydroxyl-5-methyl-4-isoxazole-propionate) (Krugers y col., 2010).
En ratas, la inducción de estrés por restricción o la aplicación sistémica de
corticosterona produce deficiencias en tareas de memoria espacial (Luine y
col.,1993; Diamond y col., 1996; 1999). La aplicación de choques eléctricos a
ratas 30 min antes de la prueba de recuperación en el laberinto de Morris
deteriora su desempeño. Este efecto no se observa cuando los choques son
administrados 4 h o 2 min antes de la prueba. Esta ventana de susceptibilidad
está relacionada con el curso temporal de la elevación de la corticosterona en
respuesta a los choques (Diamond y Kim, 2002). Este mismo efecto se consigue
si se administra corticosterona 30 min antes de la prueba de recuperación. En
ambas condiciones, el efecto amnésico es bloqueado mediante la
administración de inhibidores de la síntesis de corticosterona (de Quervain y
col., 1998). Así mismo, se ha demostrado que el estrés afecta la plasticidad
sináptica, la morfología de las dendritas, la neuro-toxicidad y la neurogenesis
dentro del hipocampo, lo que potencialmente puede tener impactos sobre el
aprendizaje y la memoria (Jeansok y Kenneth, 1998; Sandi y col., 2005); varios
trabajos han mostrado que las situaciones estresantes perjudican algunas
formas de aprendizaje dependiente de hipocampo, así como tareas de memoria
espacial (Diamond y col., 1996; Luine y col., 1993).
El hipocampo es una de las estructuras del lobo temporal medio, mas vulnerable
a los efectos del estrés; dependiendo de la severidad y del contexto, el estrés
puede facilitar o perjudicar formas de aprendizajes dependientes de hipocampo
tanto en animales como en humanos, así mismo, estas evidencias indican que
las alteraciones en la memoria espacial inducidas por estrés se deben,
principalmente, a los efectos de los glucocorticoides sobre el hipocampo (Kim y

24
col., 2006). Está estructura expresa la mayor cantidad de receptores para
mineralocorticoides (MRs) y de receptores para glucocorticoides (GRs) en el
cerebro de los mamíferos. Además, el hipocampo participa en la
retroalimentación negativa de los corticoides sobre el HPA (Akirav y Richter-
Levin, 1999). En efecto, en ratas, el estrés por restricción o la administración
sistémica de corticosterona deteriora el metabolismo, la plasticidad sináptica y la
neurogénesis y provoca muerte neuronal en el hipocampo (McEwen y col.,
1995).
Los glucocorticoides afectan la adquisición de memorias, tanto en tareas
espaciales como no espaciales, con una relación dosis-efecto que sigue una
forma de U invertida. En dosis moderadas facilitan la adquisición, mientras que
en dosis bajas y altas producen el efecto contrario (Cordero y Sandi, 1998;
Roozendaal 2002). En contraste, los niveles altos de glucocorticoides deterioran
la recuperación de la memoria, este efecto es mediado por la activación de GRs
y MRs en el hipocampo (Roozendaal, 2002). Así mismo, dependiendo del tipo
de receptor activado (MRs o GRs), los glucocorticoides pueden afectar
diferentes aspectos del procesamiento de información, en una tarea de
aprendizaje espacial, la activación de MRs está relacionada con la interpretación
de la información recibida del medio ambiente y con la selección de una
adecuada respuesta comportamental, mientras que la activación de los GRs
esta relacionada directamente con el almacenamiento de nueva información (de
Kloet y col., 2004) (Figura 4)

25
Figura 4. Receptores de glucocorticoides y procesamiento de información. Se muestran las
tres fases de la formación de memoria que son afectadas de manera específica por el bloqueo
selectivo de los receptores de glucocorticoides (GR) y de los receptores de mineralocorticoides
(MR). (Tomado de Kloet E. Ron, Melly S. Oitzl and Marian Joëls (1999). Stress and cognition:
are corticosteroids good or bad guys? Trends Neurosci. 22: 422–26).
Aunque el estrés puede claramente regular la formación de memoria, los
sustratos neurales a través de los cuales esto es llevado a cabo, no han sido
completamente elucidados; el factor liberador de corticotropina (CRF) es uno de
los posibles candidatos. Este media el incremento en el aprendizaje dependiente
del hipocampo y ejerce sus funciones biológicas a través de dos receptores:
CRF1 y CRF2, los cuales han sido detectados en el hipocampo (Zorrilla, 2002).
Por otra parte las modificaciones inducidas por el estrés en el funcionamiento de
la amígdala también pueden participar en el deterioro de la memoria. Al parecer,
esto es debido a que la amígdala modula los efectos del estrés sobre el
hipocampo. La amígdala tiene una fuerte conectividad recíproca con casi todas
las regiones de la formación hipocámpica, por lo que, potencialmente, existen
interacciones funcionales complejas entre estas dos estructuras límbicas.

26
La actividad de la amígdala está estrechamente vinculada a la regulación de la
respuesta de estrés, pese a tener una densidad moderada de GRs; por otra
parte, la lesión o el bloqueo farmacológico de la amígdala previene los efectos
del estrés sobre la memoria espacial (Kim y col., 2001) y bloquea los efectos
facilitadores de la memoria de agonistas de glucocorticoides administrados
directamente en el hipocampo; por el contrario, la administración de agonistas
de glucocorticoides en la amígdala basolateral mejora el desempeño en la
prueba de recuperación en el laberinto de Morris (Roozendaal y col., 1996;
1997a; 1997b; 2001 y 2002).
Como se ha descrito, la determinación de los efectos del estrés sobre la
memoria se ha realizado en modelos animales, utilizando los diferentes tipos de
laberintos para evaluar la memoria espacial. El laberinto radial de Olton y el
laberinto acuático de Morris, tienen el inconveniente que utilizan situaciones
emocionales para facilitar el aprendizaje; sin embargo el laberinto de Barnes, el
cual consta de una plataforma circular elevada con agujeros en la periferia, uno
de los cuales conduce a una caja de escape, utiliza como motivación la aversión
de los roedores a los espacios abiertos e iluminados y su tendencia a
guarecerse en lugares cerrados y oscuros, este ofrece algunas ventajas: 1) Es
pequeño y fácil de manipular; 2) Es menos estresante para los animales que el
laberinto acuático de Morris; 3) A diferencia del laberinto radial no utiliza la
privación de alimento o agua como motivación, que puede verse alterada por la
acción de los corticoides sobre el hambre y la saciedad. (Barnes, 1979, McLay y
col., 1998; Olton 1979; Morris 1984).
Estudios llevados a cabo en el laboratorio de Neurofisiología Comportamental
de la Universidad Nacional de Colombia han evaluado los efectos del estrés
sobre la recuperación y la extinción de la memoria espacial, utilizando el
laberinto circular de Barnes (Carvajal, 2006; Torres, 2006). Los resultados de
estos estudios han demostrado que la inyección de 1 ó 3 mg/Kg de
corticosterona, no altera la recuperación de la tarea espacial, pero hace que la
memoria sea más susceptible a extinción (Carvajal, 2006).

27
Por otra parte, la inducción de estrés por restricción de movimientos por 1 ó 4
horas mejora la recuperación (Torres, 2006). Adicionalmente, la inducción de
estrés por restricción de movimientos por 4 h hace que la memoria sea más
susceptible a extinción (Torres, 2006). Los resultados anteriormente citados
muestran que el efecto del estrés y los glucocorticoides sobre la memoria han
sido estudiados de forma no sistemática.
2.4. Estado de fosforilación y control de la función celular
Las modificaciones postraduccionales de las proteínas tienen marcados y
variados efectos sobre su función. Existen diversos sistemas de modificaciones
covalentes de las proteínas: ribosilación, acetilación, miristoilación,
isoprenilación, carboximetilación, sulfatación, glicosilación y fosforilación entre
las principales. La fosforilación, constituye un mecanismo común de
modificación y tiene un papel importante en la mayoría de los eventos de
señalización; esta involucra la transferencia de un grupo fosfato desde el ATP a
residuos específicos de la proteína blanco (Van der Geer y col., 1994); es el
mecanismo más común (afecta al menos a un tercio de las proteínas de una
célula eucariótica) y mejor regulador del control molecular de la función proteíca
neuronal en respuesta a estímulos extracelulares. Casi todos los tipos de
señales (neurotransmisores, hormonas, factores neurotróficos, citoquinas e
incluso la luz) producen la mayoría de sus efectos fisiológicos mediante cambios
en el estado de fosforilación de proteínas específicas en las células receptoras.
Dado que casi todos los tipos de proteínas expresadas en neuronas cambian su
estado funcional dependiendo de su grado de fosforilación, es altamente
probable que los cambios plásticos en el sistema nervioso, incluida la memoria,
estén relacionados con cambios en el estado de fosforilación de diversas
proteínas. De hecho, se ha demostrado que la LTP, se asocia con aumento en
la fosforilación de los receptores para glutamato de tipo AMPA, mientras que la
LTD, se asocia con de-fosforilación de los mismos receptores (Malenka, 2003).

28
Proteínas quinasas y fosfatasas son las enzimas responsables de determinar el
estado de fosforilación de proteínas celulares, y de la traducción de señales
dentro de la célula. Cambios en los niveles, localización subcelular y actividad
de quinasas y fosfatasas tienen consecuencias para el funcionamiento normal y
el mantenimiento de la homeostasis celular (Denton y Tavare, 1995). Además, el
balance entre la fosforilación y la de-fosforilación de sustratos específicos es un
mecanismo que controla la plasticidad sináptica.
Las proteínas quinasas, pertenecen a una gran superfamilia de proteínas que se
caracterizan por poseer un dominio catalítico de 250-300 residuos de
aminoácidos. Son enzimas que agregan grupos fosfato sobre uno de los tres
residuos de aminoácidos que tienen un grupo hidroxilo libre; Constituyen una de
las familias más grandes de enzimas, en efecto, en humanos 1,7% de los genes
codifican para quinasas. La mayoría de las proteínas quinasas en eucariotas
comparten un centro catalítico conservado, además en el extremo N-terminal del
dominio catalítico se encuentra una región rica en aminoácidos de glicina que se
encuentra en vecindad con residuos de lisina, los cuales participan en la unión al
ATP; en la parte central del dominio catalítico, se encuentra un residuo de acido
aspartico conservado, el cual es importante para la actividad catalítica de estas
enzimas. Se agrupan en dos clases principales de acuerdo a su especificidad:
serina-treonina quinasas (que fosforilan residuos de serina y treonina) y tirosina
quinasas (que dan cuenta de menos del 1% de todas las reacciones de
fosforilación y fosforilan solo residuos de tirosina) (Hanks y Hunter, 1995).
Las tirosinas quinasas, participan en las vías de señalización asociadas con el
crecimiento y diferenciación celular. Su dominio catalítico les permite fosforilar
residuos de tirosina, el cual es importante para el procesamiento, vía un receptor
de superficie, de las señales extracelulares reguladoras del crecimiento
(Ziemiecki et al, 1994); se distinguen dos tipos de tirosina quinasas con base en
su localización subcelular: transmembrana que funcionan como receptor y
citoplasmáticas asociadas a un receptor. Las primeras, poseen un dominio
extracelular que les permite asociarse a ligandos y un dominio catalítico

29
intracelular con actividad de quinasa de tirosina. La unión del ligando produce
una variedad de efectos que incluyen la activación de otras tirosina quinasas,
elevación de los niveles intracelulares de calcio, activación de quinasas de
serina y treonina, fosfolipasa C, fosfatidilinositol-3-quinasa y, finalmente,
cambios en la expresión génica (Medema y Bos, 1993).
Por otro lado, las citoplasmáticas asociadas a receptores transmiten señales
desde la membrana interactuando con los dominios citoplasmáticos de proteínas
de membrana. En este grupo se incluyen las enzimas de las subfamilias de
quinasas Src y Jak (quinasas Janus) (Hunter, 1998). Las serina-treonina
quinasas se subdividen en: 1) Quinasas dependientes de segundos mensajeros
(PKA, PKC, PKG, CaMK); 2) Quinasas de proteínas activadas por mitógenos
(MAPKs: ERK, JNK, SAPK) y sus quinasas reguladoras (MEK, SEK, Raf, MEK
quinasas; 3) Quinasas dependientes de ciclinas (Cdk1, Cdk5) y su quinasas
reguladoras (CAK, CAK quinasa); 4) Quinasas de receptores acoplados a
proteína G (GRK) (Wei y col., 1994). Las proteínas quinasas activadas por
mitógenos (MAPK), son ampliamente distribuidas a través del sistema nervioso
central y juegan un papel importante en la plasticidad sináptica y en la formación
de memoria, las más comunes son las quinasas reguladas por señales
extracelulares ERK1 y ERK2 (p44 y p42 respectivamente), las cuales regulan
principalmente el crecimiento celular y la diferenciación (Roberson, 1999).
ERK1/2 integran señales que son recibidas por receptores acoplados a
proteínas G y factores de crecimiento presentes en la membrana celular y las
transfieren al núcleo (Chen y col., 1993; Cano y Mahadevan, 1995); la
activación y regulación de ERK1/2 es regulada por múltiples quinasas
intracelulares así como Mek1/2 (quinasas reguladas por señales extracelulares
activadas por mitogenos) y PKA, así como por auto-fosforilación (Impey y col.,
1998a; Roberson y col., 1999); subsecuentemente el acoplamiento de los
procesos bioquímicos citoplasmáticos a la expresión génica ocurre a través de
la fosforilación de blancos corriente abajo de ERK1/2, los cuales actúan como

30
factores de transcripción así como Elk-1 u otros reguladores transcripcionales
así como quinasa 1-3 S6 ribosomal (p90Rsk-1-3) (Chen y col.,1992).
Las proteínas fosfatasas sirven como reguladores negativos, que contrarrestan
las acciones de las proteínas quinasas mediante la inducción y el mantenimiento
de la LTD, regulando negativamente la fuerza sináptica. La de-fosforilación de
proteínas reguladoras es un mecanismo importante de modulación de las vías
de señalización que involucran proteína quinasas, esta se realiza por medio de
las fosfatasas, las cuales han sido clasificadas con base en su especificidad de
sustrato en fosfatasas de serina/treonina y fosfatasas de tirosina, existiendo
también fosfatasas de especificidad dual (Cohen, 1991).
Las fosfatasas de tirosina, remueven grupos fosfatos en residuos de tirosina y
participan en la regulación de la proliferación, diferenciación, migración celular,
transcripción de genes e incluso en la respuesta inmune (Burque y Zhang,
1998). Respecto a las fosfatasas de serina/treonina (Ser/Thr), el análisis
bioquímico ha permitido agrupar éstas en fosfatasas en: tipo 1 (PP1), 2A
(PP2A), 2B (PP2B, también conocida como calcineurina) y 2C (PP2C). Los tipos
PP1, PP2A y PP2B poseen alta homología en sus dominios catalíticos, pero
diferente especificidad de sustrato e interacción con moléculas reguladoras. Las
proteína fosfatasas PP2B y PP2C poseen un requerimiento absoluto de cationes
como Calcio y Magnesio, respectivamente, mientras que PP1 y PP2A no
requieren de estos (Cohen, 1989, 1997).
Aunque 418 genes en humanos codifican para proteínas Ser/Thr quinasas, solo
15 genes codifican para Ser/Thr fosfatasas. Entre las proteínas fosfatasas
expresadas por neuronas de mamíferos [PP1, PP2A, PP2B (calcineurina), PP2C
(PPM), PP4, PP5, PP6 and PP7], PP1, PP2A y PP2B son las mas abundantes y
de-fosforilan mas del 90% de las fosfoproteínas en neuronas. Las funciones de
estas enzimas han sido estudiadas usando dos clases de inhibidores
farmacológicos (1) acido ocadaico y caliculina, los cuales inhiben principalmente

31
PP1 y PP2A e (2) inmunosupresores como la ciclosporina A y FK506, los cuales
inhiben PP2B (Mansuy y Shenolikar, 2006).
La fosfatasa 2B (PP2B), es una Ser/Thr fosfatasa citosolica expresada
ubicuamente y altamente conservada en eucariotas. Consiste de dos
subunidades: la subunidad enzimática A y la subunidad reguladora B. La
subunidad A contiene un sitio de unión a calmodulina y un dominio auto-
inhibitorio, el cual bloquea el centro catalítico de la enzima. La unión de iones
calcio y calmodulina a la PP2B conduce a cambios de conformación y a un
subsecuente desenmascaramiento del centro activo. La actividad de la PP2B es
acoplada a los niveles citosolicos de calcio, la cual es una propiedad única de
esta enzima. Adicionalmente, la actividad y localización de la PP2B es modulada
por proteínas endógenas, así como reguladores de calcineurina (RCANs),
proteína 1 de unión a calcineurina (Cabin1) ó proteína 79 de anclaje a la quinasa
A (AKAP79). A parte de la regulación por calcio, la actividad catalítica de PP2B
puede ser regulada por miristoilación, una propiedad que puede marcar PP2B a
estructuras de membrana (Alistair y Sim, 1992).
PP1 es una fosfatasa dependiente de manganeso, de 37 kDa, la cual es
compuesta de un complejo oligomerico que comprende un centro enzimático,
denominado la subunidad catalítica (PP1c). PP1 carece de especificidad de
sustrato, sin embargo esta puede ser conferida por la interacción de PP1 con un
gran numero de subunidades reguladoras, las subunidades reguladores son
generalmente no relacionadas, pero la mayoría poseen un motivo RVxF de
unión a PP1 (Eto y col., 2004). Existe en 4 isoformas (α, β, γ, δ) las cuales se
expresan diferencialmente en el citoplasma y el núcleo, PP1α es principalmente
citoplasmática, y PP1 β y δ son tanto citoplasmáticas como nucleares y PP1 γ
es predominantemente nuclear (Ceulemans y Bollen 2004; Moorhead y col.,
2007). PP1 se encuentra normalmente inactiva en altos niveles en el citosol,
estado que es mantenido por su asociación con diferentes proteínas reguladoras
así como, la proteína inhibitoria NIPP-1 (inhibidor nuclear de PP1); la
fosforilación de NIPP-1 por proteínas quinasas dependientes de AMPc y por

32
quinasas de caseínas conlleva a la liberación de PP1 activa. Por otra parte la
quinasa GSK3 activadora de PP1 es predominantemente unida a membrana y
PP1 inactiva es ausente en las fracciones de membrana en el cerebro, lo que
sugiere que la regulación de esta forma de PP1 ocurre solamente en
compartimentos citosolicos que se aproximan a estructuras membranosas que
contienen GSK3 (Alistair y Sim, 1992).
Como es de esperarse, las vías de señalización que activan quinasas y
fosfatasas tienen un amplio grado de interacciones cruzadas. No obstante, es
posible determinar el grado de activación de estas enzimas a partir de su estado
de fosforilación.
2.5. Memoria y fosforilación
La mayoría de los eventos bioquímicos que soportan la consolidación de
memoria, son llevados a cabo por modificaciones en la fosforilación/activación
de diferentes proteínas quinasas localizadas a nivel sináptico y nuclear. La
proteína quinasa dependiente de AMP cíclico (PKA), es la mejor estudiada
(Bernabeu y col., 1997); está es regulada por receptores dopaminergicos D1,
adrenérgicos y serotonina 1A (5HT1A); estos receptores estimulan la
adenilciclasa y en consecuencia inducen rápidamente un incremento de los
niveles de AMPc, los cuales directamente estimulan PKA (Izquierdo y Medina,
1997). Se ha reportado en la literatura, diferentes relaciones entre los procesos
de fosforilación y la consolidación de memoria; así, por ejemplo, la activación
farmacológica de PKA en la región CA1 del hipocampo mejora el rendimiento en
tareas de memoria espacial (Sharifzadeh y col., 2007). Por el contrario, la
infusión, inmediatamente después del entrenamiento, de un inhibidor de la PKA
en el hipocampo deteriora la retención de la memoria espacial (Sharifzadeh y
col., 2006).

33
El estrés prenatal se asocia con disminución de la expresión de la PKC (proteína
quinasa dependiente de calcio, diacilglicerol y fosfolípidos) en el hipocampo y
con un rendimiento disminuido en el laberinto acuático de Morris (Wu y col.,
2007). La inhibición de la PKC después de la reactivación de una memoria
espacial induce una amnesia duradera, independientemente de la intensidad del
entrenamiento previo (Bonini y col., 2007). Por el contrario, la activación de la
PKC antes del entrenamiento favorece la formación de memoria a largo plazo de
la asociación entre un estímulo visual y uno vestibular en Hermissenda (Alkon y
col., 2005).
La expresión de una forma constitutivamente activa de la CaMKII (calcio
calmodulina quinasa II) inducida inmediatamente después de un entrenamiento
espacial exitoso impide la consolidación de esta memoria (Yasuda y Mayford,
2006). Por el contrario, ratones transgénicos con una forma de CaMKII
independiente de calcio no pueden adquirir memorias espaciales en el laberinto
de Barnes pero si pueden adquirir miedo contextual condicionado (Bach y col.,
1995).
La activación de SGK (quinasa inducible por suero y glucocorticoides) facilita la
formación de memoria de miedo contextual (Lee y col., 2007) y de memorias
espaciales (Lee y col., 2006). Más aún, los animales que adquieren más
rápidamente memorias espaciales se caracterizan por una mayor expresión de
la SGK (Tsai y col., 2002). En contraste, la inactivación de SGK por la CK2
(casina quinasa II) deteriora la formación de memorias espaciales (Chao y col.,
2007).
La activación de la PKG (proteína quinasa dependiente de GMP cíclico)
mediante la estimulación farmacológica de la guanilato-ciclasa dependiente de
oxido nítrico, facilita la potenciación a largo término y mejora el rendimiento en el
laberinto acuático de Morris (Chien y col., 2005). En contraste, en una tarea de
evitación inhibitoria, la inhibición de la PKG produce un deterioro de la memoria
a largo plazo, pero no de la memoria a corto plazo (Izquierdo y col., 2000); por

34
otra parte, se sugiere un papel regulador de PKG en el hipocampo, en la
formación de memoria en los primero estados, de una tarea de aprendizaje,
aversivamente motivada en ratas (Bernabeu y col., 1997).
La degradación proteolítica de SCOP (proteína oscilatoria supraquiasmática)
desinhibe la transcripción mediada por ERK1/2 (quinasa regulada por señales
extracelulares) y mejora el rendimiento en una tarea de reconocimiento de
objetos (Shimizu y col., 2007). Tras la inducción de LTP y el entrenamiento en el
laberinto acuático de Morris se observa un incremento en la fosforilación de la
ERK1/2 en la circunvolución dentada del hipocampo (Gooney y col., 2002). Por
el contrario, la inhibición de la ERK1/2 en el hipocampo deteriora la memoria
espacial a largo plazo (Hebert y Dash, 2004) sin afectar su adquisición (Blum y
col., 1999). También, se ha mostrado que la ruta MAPK quinasa es requerida
tanto para la consolidación como para la reconsilidación de memoria de
reconocimiento a largo plazo, y que está asociada con hiperfosforilación de
ERK1/2 en diferentes subregiones de la corteza entorrinal-circuito hipocampal
(Aine y col., 2003). La quinasa regulada por señales extracelulares, tiene un
papel importante en la plasticidad sináptica, y en varias formas de memoria
dependientes de hipocampo. Una vez activada, ERK1/2 puede translocarse al
núcleo y fosforilar ciertos factores de transcripción así como CREB (proteína de
unión al elemento de respuesta al AMP cíclico) y Elk, lo cual desencadena
expresión de genes asociados con los procesos de memoria (Sweatt, 2001;
2004; Davis y Laroche, 2006; Giovannini, 2006).
Por otra parte, la inhibición parcial de la PP1 (proteína fosfatasa 1) mediante la
expresión inducida de inhibidor 1 facilita la LTP (Jouvenceau y col., 2006) y la
formación de memorias espaciales en el laberinto de Morris (Genoux y col.,
2002; Waddell, 2003). Por estos motivos, se ha sugerido, aunque aún no se ha
comprobado que esta enzima está relacionada con fenómenos de debilitamiento
de la memoria. De la misma manera, la inhibición de la PP2A (proteína fosfatasa
2A) en el prosencéfalo basal causa una disminución de la concentración de
acetilcolina acompañada de deficiencias para el aprendizaje espacial (Tian y

35
col., 2004). Por otra parte, la desinhibición sostenida de la PP2A en ratones
“knock out” para alfa4 (una proteína inhibidora de la PP2A) se asocia con
aumento de la actividad de la CaMKII-alfa en el hipocampo y con deficiencias
para el aprendizaje de evitación activa y para el aprendizaje espacial (Yamashita
y col., 2006). De nuevo, la escasa evidencia indica que la activación de la PP2A
en el hipocampo debilita el aprendizaje, pero este hecho aún no ha sido
plenamente demostrado.
La actividad de la PP2B disminuye durante la adquisición de una tarea espacial,
pero aumenta durante la reversión de tal aprendizaje (Havekes y col., 2006). En
ratas viejas la expresión de PP2B está aumentada y no se observa disminución
durante el entrenamiento espacial, lo cual se asocia con dificultades para la
retención de la tarea (Monti y col., 2005). Durante la extinción se observa un
aumento en la expresión de la calcineurina que puede ser bloqueado por
inhibidores de la síntesis de proteína, antagonistas del receptor NMDA y por
inhibidores de la fosfatidil-inositol 3 quinasa y de la MAPK (Lin y col., 2003).
Según los antecedentes citados, la adquisición de memorias espaciales requiere
de la inhibición de la calcineurina, mientras que las modificaciones de dichas
memorias requieren del aumento de la actividad de calcineurina secundaria a
síntesis proteica, además, en términos generales, un aumento en la actividad de
quinasas esta relacionada con una regulación positiva de los procesos de
aprendizaje, mientras que un aumento en la actividad de fosfatasas está más
relacionado con una regulación negativa (Figura 5).

36
Figura 5. Rutas de transducción de señales que operan en el hipocampo durante los procesos de formación de memoria. Activación diferencial de proteínas quinasas y fosfatasas durante el proceso de formación de memoria. La activación de receptores metabotrópicos de glutamato constituye el paso inicial en la formación de memoria, que conduce a un aumento en la actividad de varias proteínas quinasas; este proceso inicia con la activación dependiente de calcio y calmodulina de la adenilciclasa, la producción de AMP cíclico (AMPc) y la activación de proteína quinasa dependiente de AMPc (PKA) y de proteína quinasa C (PKC), las cuales posteriormente desencadenan la activación de quinasas reguladas por señales extracelulares (ERK), la cual se trasloca al núcleo y activa varias proteínas efectoras como lo son la quinasa 2 ribosomal S6 (RSK2), y CREB; Contrarrestando este proceso, las proteínas fosfatasas PP1 y PP2A, actúan sobre ERK, modulando su actividad y generando un balance característico de la plasticidad sináptica. A/K, receptors AMPA/KA receptors; CaM, calmodulina; DAR, receptor de dopamina D1/D5;βAR, receptor β-adrenergico; mGLUR, receptor metabotropico de glutamato; mAChR, receptor muscarinico acetylcolina; PLC, fosfolipasa C;AC, adenilil ciclasa; DAG, diacilglicerol; PKC, proteína quinasa C; RSK2, quinasa S6 ribosomal, también conocida como quinasa CREB; CaMKII o CaMKIV, quinasas reguladas por calmodulina.(Tomado de Roberson Erik D, English Joey D, Adams J. Paige, Selcher Joel C, Kondratick Christine and Sweatt J. David. (1999) The Journal of Neuroscience. 19; 11:4337-48 y Eric Klann (2002). Metaplastic Protein Phosphatases. Learning & Memory. 9:153-55.
2.6. Estrés y fosforilación
La exposición crónica a una serie de eventos estresantes leves provoca una
disminución significativa de la expresión de PKA y de pCREB (Wang y col.,
2006). Tras la inducción de desesperanza aprendida mediante la aplicación
repetida de choques eléctricos inescapables se ha encontrado disminución de la
actividad de la PKA y de la expresión de las subunidades RIIbeta, Calfa y Cbeta
en la corteza frontal y el hipocampo (Dwivedi y col., 2004) y un aumento de la
actividad de PKA en el núcleo accumbens (Araujo y col., 2003).

37
El estrés por restricción induce disminución de la expresión de PKC en el
hipocampo cuando es aplicado repetidamente durante la vida prenatal (Wu y
col., 2007) o postnatal (Alfonso y col., 2006). Cuando el estrés por restricción es
aplicado de forma aguda disminuye la expresión de la PKC gamma en el
hipocampo (McNamara y Lenox, 2004), pero aumenta su actividad en la
amígdala (Shors y col., 1997).
Aún no se ha probado el efecto del estrés inducido por tratamientos
comportamentales sobre la actividad de la PKG en el sistema nervioso central.
No obstante, se ha encontrado que la activación de la PKG podría favorecer la
supervivencia celular frente al estrés oxidativo (Andoh y col., 2002; Li y col.,
2005).
La fosforilación de la CaMKII en el hipocampo aumenta tras la aplicación crónica
de estrés por inmovilización (Suenaga y col., 2004). Por otro lado, la exposición
aguda a estrés por inmovilización produce un aumento de la activación de la
CaMKII (Blank et al., 2002), pero disminuye su autofosforilación en el hipocampo
(Blank y col., 2003).
El estrés crónico por nado forzado se asocia con disminución de la fosforilación
de ERK2 en el hipocampo y la corteza prefrontal (Qi y col., 2006). El estrés
agudo por inmovilización, por el contrario, provoca un aumento de la
fosforilación de ERK1/2 en el núcleo paraventricular del hipotálamo (Sasaguri y
col., 2005), el hipocampo, la neocorteza, la amígdala y el estriado (Shen y col.,
2004).
La exposición al laberinto en cruz elevado desencadena respuestas
comportamentales defensivas y se asocia con inducción de la SGK en el núcleo
tegmental ventral (Koya y col., 2005). Tras un estrés oxidativo por isquemia se
produce un incremento de la expresión de SGK1 en el hipocampo (Nishida y
col., 2004). La inhibición de la expresión de SGK se asocia con aumento de la

38
supervivencia de células sometidas estrés oxidativo (Schoenebeck y col., 2005).
No obstante, aún no se ha probado el efecto del estrés inducido por
intervenciones comportamentales sobre la expresión de SGK en el sistema
nervioso central.
No hay estudios acerca de los efectos del estrés provocado por tratamientos
comportamentales sobre la actividad de PP1 en el sistema nervioso. No
obstante, existen evidencias que sugieren que esta fosfatasa ejerce una función
protectiva durante la respuesta de estrés celular (Kim, 2006). El estrés por
inmovilización administrado en forma aislada o repetida se asocia con
incremento de la actividad de la PP2A, sin aumento de su concentración en el
hipocampo, la corteza frontal (Morinobu y col., 2003) el núcleo accumbens y el
área tegmental ventral (Maeda y col.,2006). La exposición única (Takahashi y
col., 2001) o repetida (Gerges y col., 2004) a estrés por inmovilización provoca
aumento de la actividad de PP2B en el hipocampo.
Los efectos del estrés en los procesos de aprendizaje y formación de nuevas
memorias han sido ampliamente estudiados. En animales y en humanos, se ha
mostrado que las hormonas adrenales de estrés, catecolaminas y
glucocorticoides, tienen profundos efectos sobre la memoria. Los resultados al
respecto han sido muy variables, dependiendo del tipo de estrés, el tipo de
aprendizaje y el modelo experimental. Algunos estudios han mostrado un papel
facilitador de la corticosterona sobre los mecanismos neurales que determinan
la capacidad de almacenamiento de la memoria (Akirav, 2004). En la memoria
espacial, se ha reportado que dentro de los efectos agudos de los
glucocorticoides están, un aumento en la formación de nuevas memorias
(adquisición y consolidación) y un daño en la capacidad de recordar lo que
previamente había sido aprendido (recuperación) (de Quervain, 1998). Estos
estudios se han centrado en el hipocampo, aportando considerable evidencia
que soporta el papel de esta estructura como mediador del incremento en la
consolidación de la memoria inducida por los glucocorticoides (Roozendaal,
2003). En relación a los efectos de los glucocorticoides en los procesos de

39
formación de memoria, el modelo mas conocido es el de la relación dosis-
respuesta en forma de U invertida, que sustenta que dosis moderadas de
glucocorticoides incrementan la memoria, mientras que dosis altas son menos
efectivas e incluso pueden perjudicar los procesos de formación de memoria.
(Roozendaal y col., 1999).
De lo expuesto anteriormente, queda claro que la integridad funcional del
hipocampo es un requisito indispensable para la adquisición de memorias
espaciales. Una de las regiones cerebrales más susceptibles al estrés es
precisamente el hipocampo. Además, el estrés produce modificaciones variadas
en los procesos de memoria (facilitación o deterioro) en función de su
intensidad, de su duración y del momento en que ocurra. A su vez, la formación
de memorias a largo plazo está mediada por cambios duraderos en la función
neuronal que, a su vez, dependen de la síntesis de proteínas y de cambios en
su estado de fosforilación. Por lo tanto, es altamente probable que la
complejidad de las interacciones entre la memoria y el estrés esté mediada por
los efectos de cada uno de estos procesos sobre el estado de activación de
quinasas y fosfatasas en regiones cerebrales críticas para la formación de
memorias, como el hipocampo.
Dada la amplitud del grupo de las serina-treonina quinasas y fosfatasas, para el
presente trabajo se han seleccionado la PP1 (por ejercer un rol crucial en el
deterioro de la memoria espacial y por no haber sido estudiada su actividad en
el tejido nervioso durante la respuesta de estrés) y la PP2B (por su vinculación
con las dificultades de memoria asociadas con el envejecimiento y por existir
evidencia inicial de su activación durante la respuesta de estrés) y ERK1/2
(porque su actividad aumenta tanto durante el estrés como durante la
consolidación de la memoria).

40
3. JUSTIFICACIÓN
En Colombia, el conflicto persistente y los problemas socioeconómicos hacen
que una proporción amplia de la población este sometida a eventos estresantes
de intensidad moderada a severa; esto implica que potencialmente pueden estar
en riesgo de sufrir los efectos deletéreos del estrés sobre el sistema nervioso
central, incluyendo el desencadenamiento de trastornos mentales persistentes e
incapacitantes; por ello es necesario no solo mejorar la capacidad de detección
temprana y tratamiento precoz de los trastornos mentales desencadenados por
estrés, sino también generar estrategias terapéuticas novedosas para la
prevención y, eventualmente la reversión de los efectos deletéreos del estrés
sobre procesos cognitivos. Para ello, es necesario comprender los mecanismos
por los cuales el estrés modifica la neurobiología de la cognición. Por otra parte
los mecanismos moleculares y las rutas de señalización que se activan en
respuesta al estrés y al aprendizaje son poco conocidos, por esta razón este
estudio sentaran las bases para el entendimiento a nivel molecular de los
procesos que subyacen durante el estrés y la formación de memoria.

41
4. OBJETIVOS DEL ESTUDIO
4.1. General
Establecer la relación entre los cambios inducidos por estrés agudo en la
adquisición, recuperación y extinción de la memoria espacial y las
modificaciones en la expresión y actividad de la quinasa ERK1/2 y las fosfatasas
PP1 y PP2B en el hipocampo de ratas.
4.2. Específicos
1. Determinar la actividad y el nivel de expresión basal de la quinasa
ERK1/2 y las fosfatasas PP1 y PP2B en el hipocampo de ratas.
2. Determinar los efectos del estrés agudo sobre la actividad y el nivel de
expresión de la quinasa ERK1/2 y las fosfatasas PP1 y PP2B en el
hipocampo de ratas.
3. Identificar si el entrenamiento en una tarea espacial ejerce algún efecto
sobre la actividad de la quinasa ERK1/2 y las fosfatasas PP1 y PP2B en
el hipocampo de ratas.
4. Identificar si la interacción entre el estrés agudo y el aprendizaje de una
tarea espacial tienen algún efecto en la actividad de la quinasa ERK1/2 y
las fosfatasas PP1 y PP2B en el hipocampo de ratas.

42
5. METODOLOGÍA Y DISEÑO EXPERIMENTAL
5.1. Animales.
Se utilizaron Ratas Wistar macho (300 ± 20 g) suministradas por el bioterio
central de la Universidad Nacional de Colombia; se incluyeron 5 ratas por grupo
experimental; Las sesiones experimentales se llevaron a cabo en horas de la
mañana, durante el ciclo de luz, entre las 7:00 y las 12:30 horas en el bioterio
central; los experimentos de inducción de estrés agudo por inmovilización y de
aprendizaje en el laberinto circular de Barnes los realizó el grupo de
Neurofisiología del comportamiento animal (Anexo 1).
5.2. Preparación de extractos de hipocampo
5.2.1. Disección de hipocampos
Las ratas se sacrificaron por decapitación a las 0, 1 y 4 h después del estrés por
inmovilización y de las pruebas de aprendizaje; los cerebros se removieron
rápidamente sobre hielo; inmediatamente se practicó una disección para extraer
ambos hipocampos los cuales se almacenaron en nitrógeno líquido hasta el
momento del análisis (Grupo de Neurofisiología del comportamiento animal ).
5.2.2. Homogenización de los hipocampos
Debido a que las soluciones de extracción de proteínas fosfatasas y quinasas
diferían en algunos de sus componentes, un hipocampo de cada animal se
utilizó para determinar el nivel de expresión y la actividad enzimática de las
proteínas fosfatasas y el otro para las proteínas quinasas. Los hipocampos se
homogenizaron sobre hielo mediante lisis mecánica en buffer de lisis frío
haciendo uso de un homogenizador Dounce. Para la determinación de las
fosfatasas se utilizó 250 µl de 50 mM Tris-HCl pH 7.5, 50 mM NaCl, 10 mM
EGTA, 5 mM EDTA, 1 mM PMSF, 20 μg/mL leupeptina, and 4 μg/mL aprotinina,

43
1mM ortovanadato de sodio y para la determinación de quinasas 250 µl de 50
mM Tris-HCl pH 7.5, 150 mM NaCl, 1 mM EGTA, 2 mM EDTA, 1 mM PMSF, 20
μg/mL leupeptina, and 20 μg/mL aprotinina, 1 mM ortovanadato de sodio, 20 µg
/ml pepstatina, 50 mM fluoruro de sodio, 1 mM ácido okadáico y Triton X-100
(1%). Las muestras homogenizadas se centrifugaron 15 min a 30,000 g, 4°C; se
recogió el sobrenadante y se almacenó a -80°C. El contenido de proteína en los
lisados se determinó con el sistema Qubit (Invitrogen).
5.3. SDS-PAGE y Western blot
Las muestras (~30 µg de proteína) se separaron en electroforesis en
condiciones denaturantes (SDS-PAGE) en geles de poliacrilamida al 10%
siguiendo procedimientos estándar (García, 2000; Burnette, 1981). Un carril en
cada gel contenía marcadores de peso molecular de amplio rango pre-teñidos
(BioRad). Se hicieron geles por duplicado, de tal manera que uno de los geles
se tiño con azul brillante de Coomasie y el otro se transfirió a membranas PVDF.
Brevemente, las membranas se incubaron en solución de bloqueo (leche en
polvo descremada, preparada en TBS-Tween (Tris Cl 10x, 1M pH 7.5, NaCl
(0.9%) y Tween (0.1%)) por 1 h a temperatura ambiente (TA); posteriormente se
incubaron toda la noche a 4 °C con los anticuerpos primarios específicos para
PP1 (Mouse monoclonal IgG2b, Santa Cruz Biotechnology; lote # G0908.
Dilución 1:2000); PP2B (Pan Calcineurina; Cell signalling technology; lote # 1.
Dilución 1:5000); y ERK1/2 (Rabbit Pab Anti-ERK1/2, Invitrogen; lote # 534972A.
Dilución 1:5000). Al día siguiente las membranas se dejaron temperar 1 h a TA,
se lavaron 5 veces durante 5 min en TBS-Tween y se incubaron por 1 h a TA
con los anticuerpos secundarios apropiados, anti-mouse IgG AP (dilución
1:3500) para PP1 y anti-rabbit IgG AP (dilución 1:2000) para PP2B y ERK1/2;
finalmente, se hicieron 5 lavados de 5 min cada uno y las membranas se
incubaron por 10 minutos con solución activadora.
La detección se realizó utilizando el sistema BCIP/NBT (Sustrato cromogénico
para la enzima fosfatasa alcalina). La densidad de cada una de las bandas fue

44
determinada y cuantificada haciendo uso del equipo de análisis y foto-
documentación de geles (CHEMI GENIUS-BIO IMAGING SYSTEM).
5.4. Ensayos de actividad enzimática.
5.4.1. Actividad enzimática de proteínas fosfatasas.
Para medir la actividad enzimática de PP1 y PP2B en los lisados celulares, se
empleó un ensayo fluorométrico, ProFluor Ser/Thr Phosphatase assay kit
(Promega, Madison, WI). Se tomó 25 µl (~70 µg de proteína) del lisado celular y
se le adicionó 25 µl de la solución de dilución de fosfatasas (buffer de reacción
5X, más los cofactores necesarios para la activación de PP1: 1 mM de MgCl2 y
20 mM de MnCl2 y para la activación de PP2B: 100 mM CaCl2, 100 mM NiCl2 y
Calmodulina 0,5 Mm); se sirvieron 25 µl de la dilución en placas de 96 pozos y
se adicionó 25 µl de solución péptido (buffer de reacción 5X, 10 mM de sustrato
péptido fosforilado bisamida rodamina-110 y 10 mM de sustrato AMC (control
interno)), se incubó por 10 min a TA. Posteriormente se adicionó 25 µl de
solución de proteasa (solución de terminación, reactivo proteasa (kit) y ácido
ocadáico solo para PP1) y se incubó por 90 min a TA, finalmente, se adicionó 25
µl de solución estabilizadora. La actividad de PP1y PP2B se determinó en
términos de fluorescencia (485/530, excitación/emisión sustrato fosfopeptido
R110) la cual se leyó en un lector de placas FLX80; El aumento en la intensidad
de fluorescencia se correlacionó con un aumento en la actividad enzimática de
PP1 ó PP2B.
5.4.2. Actividad enzimática de ERK1/2
La actividad enzimática de ERK1/2 en los lisados celulares se determinó con el
kit ELISA [pTpY185/187] (Invitrogen), el cual detecta y cuantifica la cantidad de
ERK1/2 fosforilado, el nivel de fosforilación de ERK1/2 es un indicativo de la
actividad enzimática de esta enzima. Las muestras se prepararon de la siguiente
manera: a 20 µl de muestra (100 µg de proteína), se les adicionó 20 µl de buffer

45
de tratamiento de muestra, se incubó por 20 min en hielo y se adicionó 160 µl de
solución de dilución del estándar. El ensayo se inició al adicionar 100 µl de
solución de dilución del estándar al pozo cero (blanco cromogénico) y 100 µl del
estándar y de las muestras a las placas cubiertas con el anticuerpo monoclonal
específico para ERK1/2; se dejó incubar por 2 h seguido de 4 lavados con buffer
de lavado (kit), se adicionó 100 µl de anti-ERK1/2 [pTpY185/187] el cual se dejó
incubar por 1 h, nuevamente se hicieron 4 lavados y se adicionó 100 µl de anti-
rabbit IgG-HRP, se incubó por 30 min y se lavo 4 veces, finalmente se adicionó
100 µl del cromógeno estabilizador, se incubó por 30 min y se adicionó 100 µl de
solución de parada e inmediatamente se hizo la lectura de absorbancia a 450
nm. La absorbancia obtenida es directamente proporcional a la concentración de
ERK1/2 fosforilado presente en la muestra.
5.5. Diseño experimental.
Los siguientes grupos experimentales se establecieron con el fin de determinar
la interacción entre el estrés agudo y el aprendizaje espacial sobre la actividad
de quinasas y fosfatasas en el hipocampo.
1. Estado basal de activación de quinasas y fosfatasas en el hipocampo.
2. Efecto del estrés agudo corto (1 h) sobre el nivel de expresión y la actividad
de quinasas y fosfatasas en el hipocampo.
3. Efecto del estrés agudo prolongado (4 h) sobre el nivel de expresión y la
actividad de quinasas y fosfatasas en el hipocampo.
4. Efecto de la adquisición sobre el nivel de expresión y la actividad de quinasas
y fosfatasas en el hipocampo.
5. Efecto de la recuperación sobre el nivel de expresión y la actividad de
quinasas y fosfatasas en el hipocampo.
6. Efecto de la extinción sobre el nivel de expresión y la actividad de quinasas y
fosfatasas en el hipocampo.
7. Interacción entre el estrés corto y la adquisición sobre el nivel de expresión y
la actividad de quinasas y fosfatasas en el hipocampo.

46
8. Interacción entre el estrés prolongado y la adquisición sobre el nivel de
expresión y la actividad de quinasas y fosfatasas en el hipocampo.
9. Interacción entre el estrés corto y la recuperación sobre el nivel de expresión
y la actividad de quinasas y fosfatasas en el hipocampo.
10. Interacción entre el estrés prolongado y la recuperación sobre el nivel de
expresión y la actividad de quinasas y fosfatasas en el hipocampo.
11. Interacción entre el estrés corto y la extinción sobre el nivel de expresión y la
actividad de quinasas y fosfatasas en el hipocampo
12. Interacción entre el estrés prolongado y la extinción sobre el nivel de
expresión y la actividad de quinasas y fosfatasas en el hipocampo
Para los análisis inmunoquímicos se utilizaron quince hipocampos provenientes
de cada uno de los grupos experimentales (15/grupo experimental), los cuales
se obtuvieron en diferentes condiciones 1) cinco hipocampos provenientes de
animales que se sacrificaron inmediatamente finalizada la prueba en cuestión; 2)
cinco hipocampos provenientes de animales que se sacrificaron 1 h después de
finalizada la prueba; y 3) cinco hipocampos provenientes de animales que se
sacrificaron 4 h después de finalizada la prueba.
Para cada individuo, en cada condición experimental, para cada enzima
analizada en el hipocampo y para cada tiempo entre el final del experimento y el
sacrificio, se evaluaron las siguientes variables:
A. Concentración total de proteína en el homogenizado.
B. determinación del nivel de expresión de quinasas y fosfatasas por western
blot.
C. Actividad quinasa, expresada como densidad óptica a 450 nm.
D. Actividad fosfatasa: expresada en unidades de fluorescencia,
Se explorará el efecto del entrenamiento, el tratamiento y el tiempo entre el final
del experimento y el sacrificio sobre la actividad de cada enzima mediante un

47
análisis de varianza de tres factores (entrenamiento, tratamiento, tiempo entre el
final del experimento y el sacrificio) haciendo uso del paquete estadístico
SigmaStat 3.01 (SPSS Inc.), con un valor crítico de significatividad de 0,05.
Se establecerán correlaciones entre la actividad de cada enzima y las variables
comportamentales mediante correlación de Spearman seguida de una
transformada z de Fisher haciendo uso del paquete estadístico SigmaStat 3.01
(SPSS Inc.), con un valor crítico de significancia de 0,05. En caso de encontrar
correlaciones significativas se procederá a un análisis de regresión por el
método de los mínimos cuadrados haciendo uso del mismo paquete estadístico.

48
6. RESULTADOS
6.1. Efecto del estrés por inmovilización en la expresión de ERK1/2
en el hipocampo de ratas
Para evaluar el efecto del estrés en la expresión de ERK1/2, las ratas se
inmovilizaron por 1 h (estrés corto) y 4 h (estrés prolongado) y se sacrificaron
inmediatamente, 1 y 4 h después del estrés, con el fin de detectar cambios tanto
a nivel de expresión de ERK1/2 como en su actividad enzimática. Los resultados
de estos ensayos se compararon también con los niveles basales (animales no
estresados).
De acuerdo con los resultados obtenidos por Western blot (WB), no hubo
diferencias en el nivel de expresión de ERK1/2 al comparar el estrés corto y el
prolongado con el basal, en los tres tiempos estudiados (Figura A2 en anexos);
sin embargo, los análisis de la actividad enzimática mostraron un aumento
estadísticamente significativo de ERK1/2 en el hipocampo durante el estrés
corto y el prolongado comparativamente con el basal (t(8) = -5,791, p<0,001; E1-
1H>BASAL; t(8) = -2,866, p = 0,021; E1-4H>BASAL; t(8) = -14,616 E4-
0H>BASAL; t(8) = -5,540, p<0,001; E4-1H>BASAL; t(8) = -3,314, p= 0,011, E4-
4H>BASAL) (Figura 6); al comparar los grupos de estrés, se observó que el
principal aumento en la actividad enzimática de ERK1/2 se obtuvo en
condiciones de estrés prolongado y sacrificio inmediato (Figura 7).
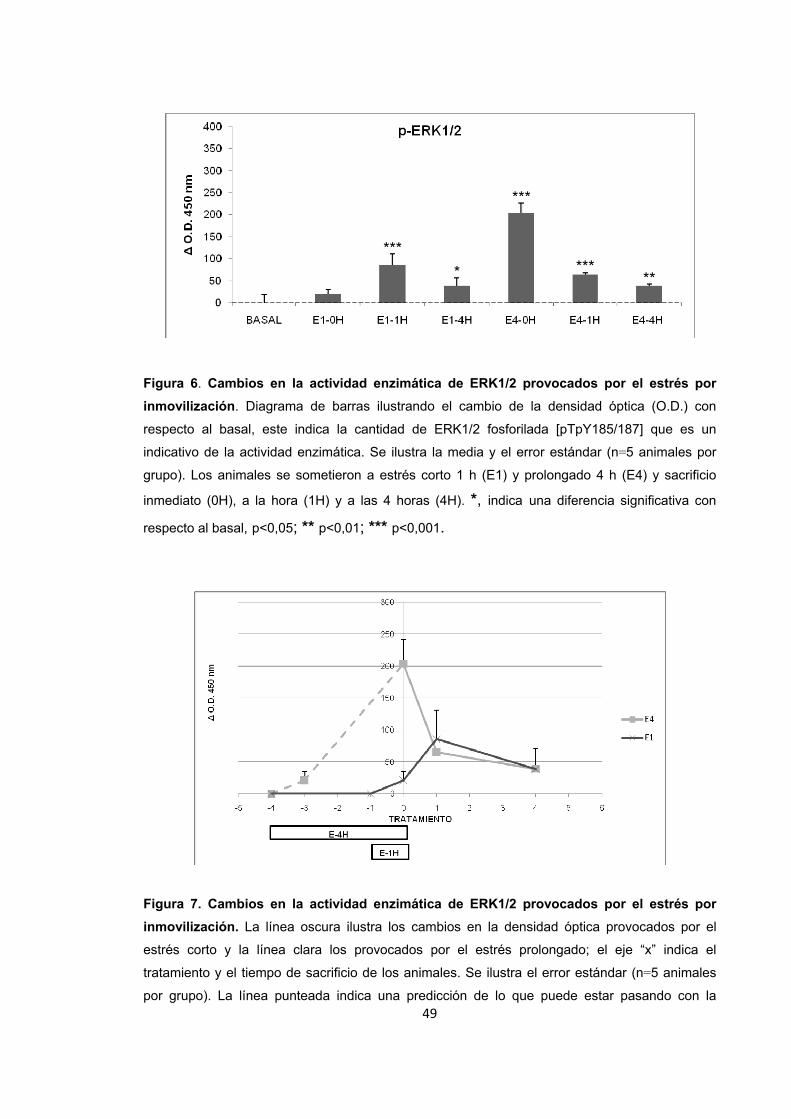
49
Figura 6. Cambios en la actividad enzimática de ERK1/2 provocados por el estrés por
inmovilización. Diagrama de barras ilustrando el cambio de la densidad óptica (O.D.) con
respecto al basal, este indica la cantidad de ERK1/2 fosforilada [pTpY185/187] que es un
indicativo de la actividad enzimática. Se ilustra la media y el error estándar (n=5 animales por
grupo). Los animales se sometieron a estrés corto 1 h (E1) y prolongado 4 h (E4) y sacrificio
inmediato (0H), a la hora (1H) y a las 4 horas (4H). *, indica una diferencia significativa con
respecto al basal, p<0,05; ** p<0,01; *** p<0,001.
Figura 7. Cambios en la actividad enzimática de ERK1/2 provocados por el estrés por
inmovilización. La línea oscura ilustra los cambios en la densidad óptica provocados por el
estrés corto y la línea clara los provocados por el estrés prolongado; el eje “x” indica el
tratamiento y el tiempo de sacrificio de los animales. Se ilustra el error estándar (n=5 animales
por grupo). La línea punteada indica una predicción de lo que puede estar pasando con la
* *****
***
***

50
actividad enzimática de ERK1/2 durante la inmovilización por 4 h, teniendo en cuenta el
comportamiento de esta en respuesta al estrés por 1 h.
6.2. Actividad y nivel de expresión de ERK1/2 durante los procesos
de formación de memoria
Comparativamente con la actividad de ERK1/2 en condiciones basales, los
resultados obtenidos indican que el entrenamiento en una tarea espacial
provocó un aumento significativo en la actividad enzimática de ERK1/2 durante
la adquisición (t(7) = -11,251, p <0,001, ADQ-OH>BASAL; t(8) = -14,545, p
<0,001, ADQ-1H>BASAL; t(8) = -16,413, p <0,001, ADQ-4H>BASAL y la
recuperación (t(8) = -12,486, p <0,001, REC-0H>BASAL; t(8) = -19,996, p <0,001,
REC-1H>BASAL; t(8) = -18,540, p <0,001, REC-4H>BASAL) de la memoria en
los tres tiempos estudiados como se observa en la figuras 8 y 9.
Figura 8. Cambios en la actividad enzimática de ERK1/2 inducidos por el entrenamiento
de la adquisición de la memoria espacial. Diagrama de barras ilustrando el cambio de la O.D.
con respecto al basal. Se ilustra la media y el error estándar (n=5 animales por grupo). ADQ-0H,
animales sacrificados inmediatamente después del entrenamiento de adquisición; ADQ-1H,
sacrificados 1 h después del; ADQ-4H, sacrificados 4 h después; ***, p<0,001.
*** ***
***
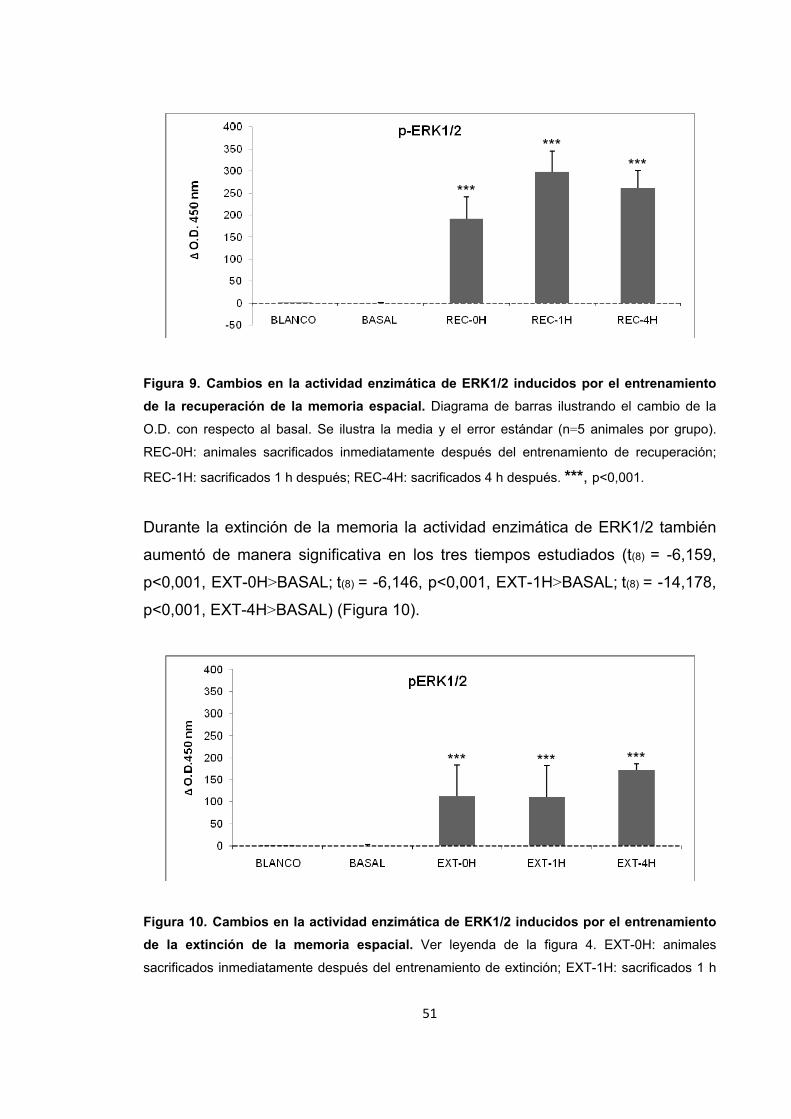
51
Figura 9. Cambios en la actividad enzimática de ERK1/2 inducidos por el entrenamiento
de la recuperación de la memoria espacial. Diagrama de barras ilustrando el cambio de la
O.D. con respecto al basal. Se ilustra la media y el error estándar (n=5 animales por grupo).
REC-0H: animales sacrificados inmediatamente después del entrenamiento de recuperación;
REC-1H: sacrificados 1 h después; REC-4H: sacrificados 4 h después. ***, p<0,001.
Durante la extinción de la memoria la actividad enzimática de ERK1/2 también
aumentó de manera significativa en los tres tiempos estudiados (t(8) = -6,159,
p<0,001, EXT-0H>BASAL; t(8) = -6,146, p<0,001, EXT-1H>BASAL; t(8) = -14,178,
p<0,001, EXT-4H>BASAL) (Figura 10).
Figura 10. Cambios en la actividad enzimática de ERK1/2 inducidos por el entrenamiento
de la extinción de la memoria espacial. Ver leyenda de la figura 4. EXT-0H: animales
sacrificados inmediatamente después del entrenamiento de extinción; EXT-1H: sacrificados 1 h
***
*** ***
*** *** ***
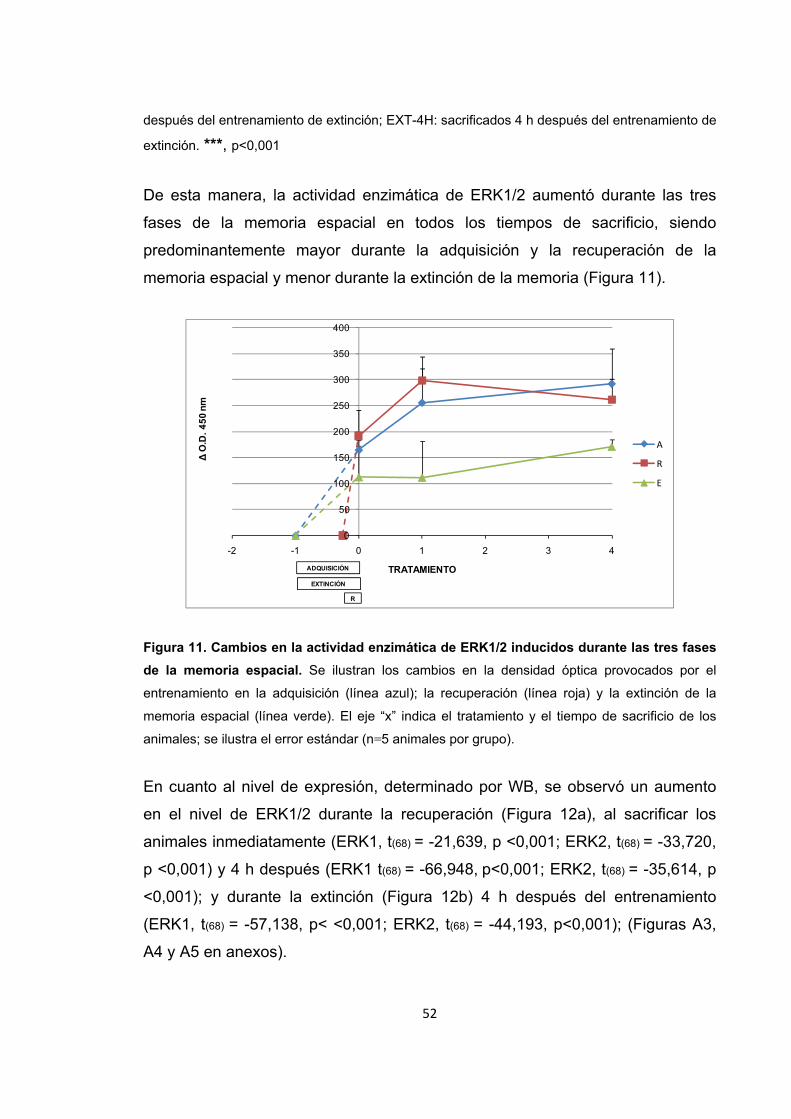
52
después del entrenamiento de extinción; EXT-4H: sacrificados 4 h después del entrenamiento de
extinción. ***, p<0,001
De esta manera, la actividad enzimática de ERK1/2 aumentó durante las tres
fases de la memoria espacial en todos los tiempos de sacrificio, siendo
predominantemente mayor durante la adquisición y la recuperación de la
memoria espacial y menor durante la extinción de la memoria (Figura 11).
0
50
100
150
200
250
300
350
400
-2 -1 0 1 2 3 4
ΔO
.D.
45
0 n
m
TRATAMIENTO
A
R
E
EXTINCIÓN
R
ADQUISICIÓN
Figura 11. Cambios en la actividad enzimática de ERK1/2 inducidos durante las tres fases
de la memoria espacial. Se ilustran los cambios en la densidad óptica provocados por el
entrenamiento en la adquisición (línea azul); la recuperación (línea roja) y la extinción de la
memoria espacial (línea verde). El eje “x” indica el tratamiento y el tiempo de sacrificio de los
animales; se ilustra el error estándar (n=5 animales por grupo).
En cuanto al nivel de expresión, determinado por WB, se observó un aumento
en el nivel de ERK1/2 durante la recuperación (Figura 12a), al sacrificar los
animales inmediatamente (ERK1, t(68) = -21,639, p <0,001; ERK2, t(68) = -33,720,
p <0,001) y 4 h después (ERK1 t(68) = -66,948, p<0,001; ERK2, t(68) = -35,614, p
<0,001); y durante la extinción (Figura 12b) 4 h después del entrenamiento
(ERK1, t(68) = -57,138, p< <0,001; ERK2, t(68) = -44,193, p<0,001); (Figuras A3,
A4 y A5 en anexos).

53
a.
b.
Figura 12. Cambios en el nivel de expresión de ERK1/2 provocados por el entrenamiento
de la memoria espacial. Diagrama de barras ilustrando el cambio en el nivel de expresión de
ERK1/2 con respecto al basal durante la recuperación (a) y la extinción (b). Se ilustra la media y
el error estándar (n=5 animales por grupo). ***, p<0,001.
***
***
***
***
*** ***

54
6.3. Efecto del estrés sobre la formación de memoria espacial y su
relación con la expresión de ERK1/2
Finalmente, se pretendía caracterizar el efecto del estrés en la formación de la
memoria espacial en términos de la actividad y expresión de ERK1/2; para esto,
se empleó un paradigma de aprendizaje espacial en el laberinto circular de
Barnes, y se diseñaron experimentos en los que se indujo estrés por restricción
de movimientos durante 1 y 4 h antes de las fases críticas del aprendizaje, los
tiempos de sacrificio fueron los mismos en todas las condiciones
experimentales.
Comparativamente con el grupo basal, la actividad enzimática de ERK1/2
aumentó significativamente con el estrés corto previo a la adquisición de la
memoria espacial (t(8) = -4,513, p = 0,002, E1-ADQ-0H>BASAL; t(8) = -5,069,
p<0,001, E1-ADQ-1H>BASAL; t(8) = -5,934, p<0,001; E1-ADQ-4H>BASAL); sin
embargo, no permitió el aumento en la actividad enzimática observado durante
la adquisición (t(3) = 4,031, p= 0,027, E1-ADQ-0H<ADQ-0H; t(4) = 5,756, p =
0,005, E1-ADQ-1H<ADQ-1H; t(4) = 2,756, p= 0,051, E1-ADQ-4H<ADQ-4H)
(Figura 13 y 15).
0
50
100
150
200
250
300
350
400
BASAL E1-0H E1-1H E1-4H ADQ-0H ADQ-1H ADQ-4H E1-ADQ-0H E1-ADQ-1H E1-ADQ-4H
ΔO
.D.
450 n
m
pERK1/2
Figura 13. Cambios en la actividad enzimática de ERK1/2 inducidos por el estrés corto
previo al entrenamiento en la adquisición de la memoria espacial. Diagrama de barras
***
*
*****
***
***
*
***
***
***
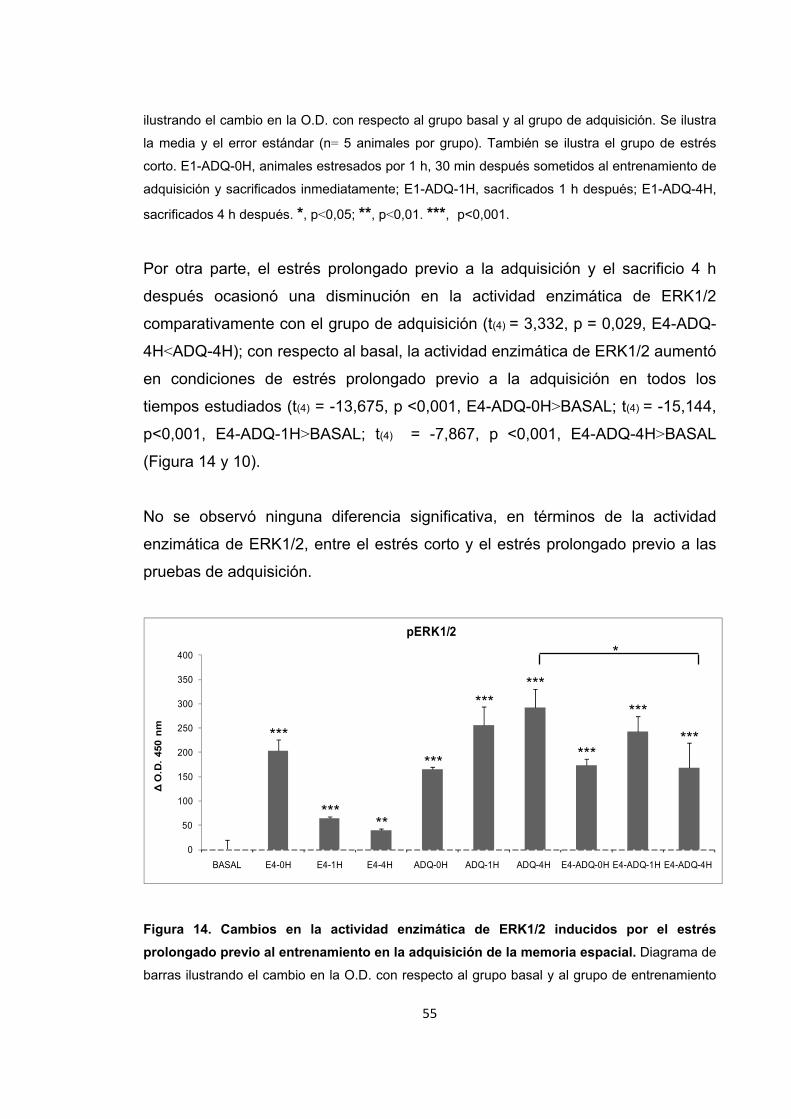
55
ilustrando el cambio en la O.D. con respecto al grupo basal y al grupo de adquisición. Se ilustra
la media y el error estándar (n= 5 animales por grupo). También se ilustra el grupo de estrés
corto. E1-ADQ-0H, animales estresados por 1 h, 30 min después sometidos al entrenamiento de
adquisición y sacrificados inmediatamente; E1-ADQ-1H, sacrificados 1 h después; E1-ADQ-4H,
sacrificados 4 h después. *, p<0,05; **, p<0,01. ***, p<0,001.
Por otra parte, el estrés prolongado previo a la adquisición y el sacrificio 4 h
después ocasionó una disminución en la actividad enzimática de ERK1/2
comparativamente con el grupo de adquisición (t(4) = 3,332, p = 0,029, E4-ADQ-
4H<ADQ-4H); con respecto al basal, la actividad enzimática de ERK1/2 aumentó
en condiciones de estrés prolongado previo a la adquisición en todos los
tiempos estudiados (t(4) = -13,675, p <0,001, E4-ADQ-0H>BASAL; t(4) = -15,144,
p<0,001, E4-ADQ-1H>BASAL; t(4) = -7,867, p <0,001, E4-ADQ-4H>BASAL
(Figura 14 y 10).
No se observó ninguna diferencia significativa, en términos de la actividad
enzimática de ERK1/2, entre el estrés corto y el estrés prolongado previo a las
pruebas de adquisición.
0
50
100
150
200
250
300
350
400
BASAL E4-0H E4-1H E4-4H ADQ-0H ADQ-1H ADQ-4H E4-ADQ-0H E4-ADQ-1H E4-ADQ-4H
ΔO
.D.
450 n
m
pERK1/2
Figura 14. Cambios en la actividad enzimática de ERK1/2 inducidos por el estrés
prolongado previo al entrenamiento en la adquisición de la memoria espacial. Diagrama de
barras ilustrando el cambio en la O.D. con respecto al grupo basal y al grupo de entrenamiento
*
***
***
******
***
*****
***
***
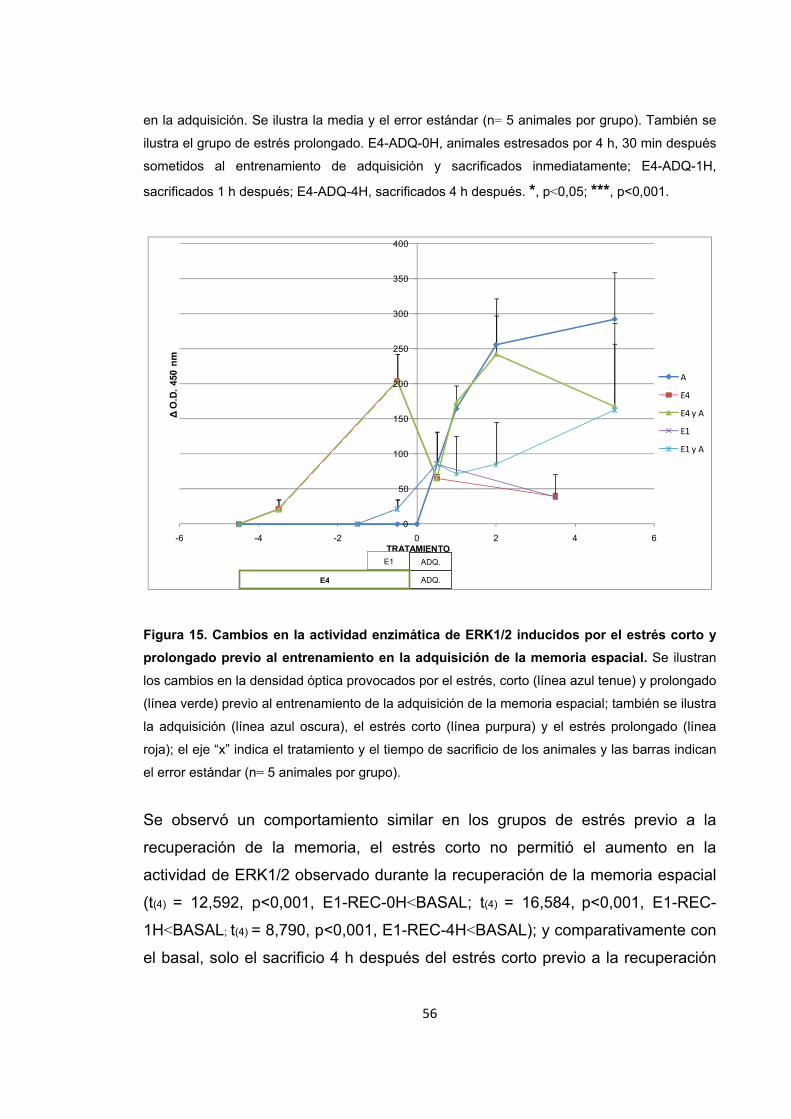
56
en la adquisición. Se ilustra la media y el error estándar (n= 5 animales por grupo). También se
ilustra el grupo de estrés prolongado. E4-ADQ-0H, animales estresados por 4 h, 30 min después
sometidos al entrenamiento de adquisición y sacrificados inmediatamente; E4-ADQ-1H,
sacrificados 1 h después; E4-ADQ-4H, sacrificados 4 h después. *, p<0,05; ***, p<0,001.
0
50
100
150
200
250
300
350
400
-6 -4 -2 0 2 4 6
ΔO
.D.
450
nm
TRATAMIENTO
A
E4
E4 y A
E1
E1 y A
Figura 15. Cambios en la actividad enzimática de ERK1/2 inducidos por el estrés corto y
prolongado previo al entrenamiento en la adquisición de la memoria espacial. Se ilustran
los cambios en la densidad óptica provocados por el estrés, corto (línea azul tenue) y prolongado
(línea verde) previo al entrenamiento de la adquisición de la memoria espacial; también se ilustra
la adquisición (línea azul oscura), el estrés corto (línea purpura) y el estrés prolongado (línea
roja); el eje “x” indica el tratamiento y el tiempo de sacrificio de los animales y las barras indican
el error estándar (n= 5 animales por grupo).
Se observó un comportamiento similar en los grupos de estrés previo a la
recuperación de la memoria, el estrés corto no permitió el aumento en la
actividad de ERK1/2 observado durante la recuperación de la memoria espacial
(t(4) = 12,592, p<0,001, E1-REC-0H<BASAL; t(4) = 16,584, p<0,001, E1-REC-
1H<BASAL; t(4) = 8,790, p<0,001, E1-REC-4H<BASAL); y comparativamente con
el basal, solo el sacrificio 4 h después del estrés corto previo a la recuperación
E1 ADQ.
ADQ.E4
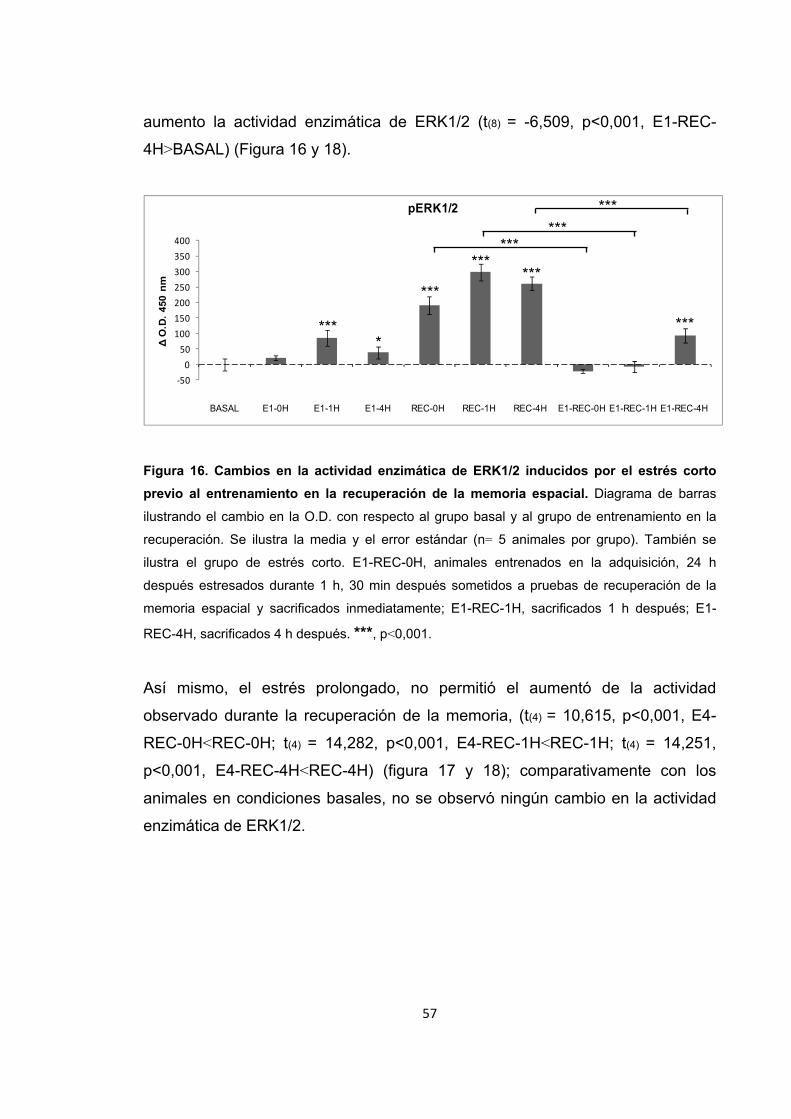
57
aumento la actividad enzimática de ERK1/2 (t(8) = -6,509, p<0,001, E1-REC-
4H>BASAL) (Figura 16 y 18).
‐50
0
50
100
150
200
250
300
350
400
BASAL E1-0H E1-1H E1-4H REC-0H REC-1H REC-4H E1-REC-0H E1-REC-1H E1-REC-4H
ΔO
.D.
450 n
m
pERK1/2
Figura 16. Cambios en la actividad enzimática de ERK1/2 inducidos por el estrés corto
previo al entrenamiento en la recuperación de la memoria espacial. Diagrama de barras
ilustrando el cambio en la O.D. con respecto al grupo basal y al grupo de entrenamiento en la
recuperación. Se ilustra la media y el error estándar (n= 5 animales por grupo). También se
ilustra el grupo de estrés corto. E1-REC-0H, animales entrenados en la adquisición, 24 h
después estresados durante 1 h, 30 min después sometidos a pruebas de recuperación de la
memoria espacial y sacrificados inmediatamente; E1-REC-1H, sacrificados 1 h después; E1-
REC-4H, sacrificados 4 h después. ***, p<0,001.
Así mismo, el estrés prolongado, no permitió el aumentó de la actividad
observado durante la recuperación de la memoria, (t(4) = 10,615, p<0,001, E4-
REC-0H<REC-0H; t(4) = 14,282, p<0,001, E4-REC-1H<REC-1H; t(4) = 14,251,
p<0,001, E4-REC-4H<REC-4H) (figura 17 y 18); comparativamente con los
animales en condiciones basales, no se observó ningún cambio en la actividad
enzimática de ERK1/2.
***
***
******
*
***
******
***
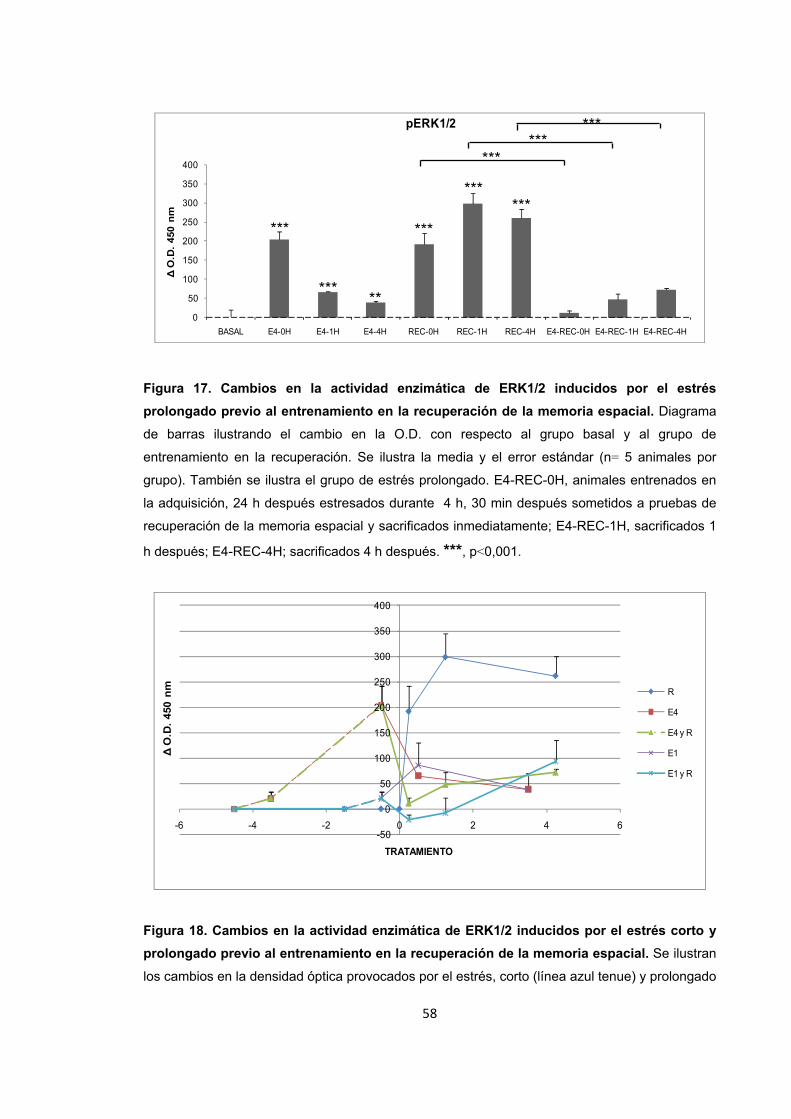
58
0
50
100
150
200
250
300
350
400
BASAL E4-0H E4-1H E4-4H REC-0H REC-1H REC-4H E4-REC-0H E4-REC-1H E4-REC-4H
ΔO
.D.
450 n
m
pERK1/2
Figura 17. Cambios en la actividad enzimática de ERK1/2 inducidos por el estrés
prolongado previo al entrenamiento en la recuperación de la memoria espacial. Diagrama
de barras ilustrando el cambio en la O.D. con respecto al grupo basal y al grupo de
entrenamiento en la recuperación. Se ilustra la media y el error estándar (n= 5 animales por
grupo). También se ilustra el grupo de estrés prolongado. E4-REC-0H, animales entrenados en
la adquisición, 24 h después estresados durante 4 h, 30 min después sometidos a pruebas de
recuperación de la memoria espacial y sacrificados inmediatamente; E4-REC-1H, sacrificados 1
h después; E4-REC-4H; sacrificados 4 h después. ***, p<0,001.
-50
0
50
100
150
200
250
300
350
400
-6 -4 -2 0 2 4 6
ΔO
.D.
450 n
m
TRATAMIENTO
R
E4
E4 y R
E1
E1 y R
Figura 18. Cambios en la actividad enzimática de ERK1/2 inducidos por el estrés corto y
prolongado previo al entrenamiento en la recuperación de la memoria espacial. Se ilustran
los cambios en la densidad óptica provocados por el estrés, corto (línea azul tenue) y prolongado
*** ***
*****
***
******
***
***
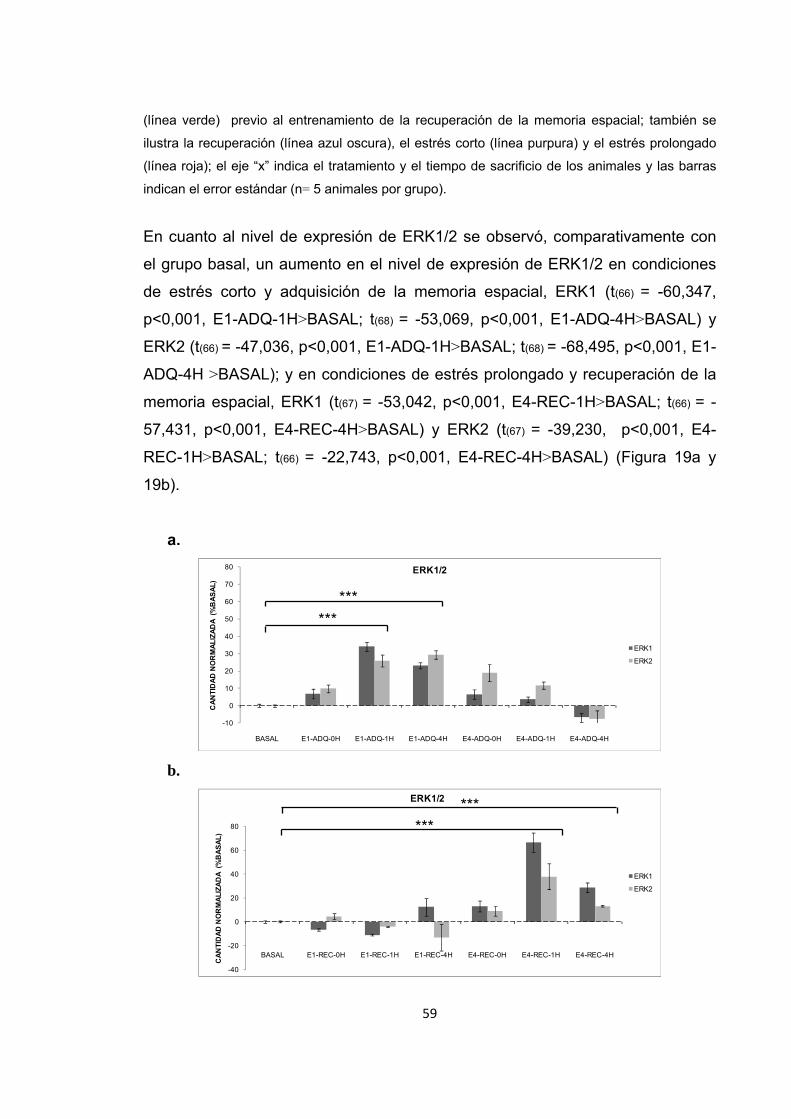
59
(línea verde) previo al entrenamiento de la recuperación de la memoria espacial; también se
ilustra la recuperación (línea azul oscura), el estrés corto (línea purpura) y el estrés prolongado
(línea roja); el eje “x” indica el tratamiento y el tiempo de sacrificio de los animales y las barras
indican el error estándar (n= 5 animales por grupo).
En cuanto al nivel de expresión de ERK1/2 se observó, comparativamente con
el grupo basal, un aumento en el nivel de expresión de ERK1/2 en condiciones
de estrés corto y adquisición de la memoria espacial, ERK1 (t(66) = -60,347,
p<0,001, E1-ADQ-1H>BASAL; t(68) = -53,069, p<0,001, E1-ADQ-4H>BASAL) y
ERK2 (t(66) = -47,036, p<0,001, E1-ADQ-1H>BASAL; t(68) = -68,495, p<0,001, E1-
ADQ-4H >BASAL); y en condiciones de estrés prolongado y recuperación de la
memoria espacial, ERK1 (t(67) = -53,042, p<0,001, E4-REC-1H>BASAL; t(66) = -
57,431, p<0,001, E4-REC-4H>BASAL) y ERK2 (t(67) = -39,230, p<0,001, E4-
REC-1H>BASAL; t(66) = -22,743, p<0,001, E4-REC-4H>BASAL) (Figura 19a y
19b).
a.
-10
0
10
20
30
40
50
60
70
80
BASAL E1-ADQ-0H E1-ADQ-1H E1-ADQ-4H E4-ADQ-0H E4-ADQ-1H E4-ADQ-4H
CA
NT
IDA
D N
OR
MA
LIZ
AD
A (
%B
AS
AL
)
ERK1/2
ERK1
ERK2
b.
-40
-20
0
20
40
60
80
BASAL E1-REC-0H E1-REC-1H E1-REC-4H E4-REC-0H E4-REC-1H E4-REC-4H
CA
NT
IDA
D N
OR
MA
LIZ
AD
A (
%B
AS
AL
)
ERK1/2
ERK1
ERK2
******
******

60
Figura 19. Cambios en el nivel de expresión de ERK1/2 inducidos por el estrés corto y
prolongado previo al entrenamiento en la memoria espacial. Diagrama de barras ilustrando
el cambio en el nivel de expresión de ERK1/2 con respecto al basal durante el estrés previo a la
adquisición (a) y a la recuperación de la memoria espacial (b). Se ilustra la media y el error
estándar (n=5 animales por grupo). ***, p<0,001.
Por ultimo, la actividad enzimática de ERK1/2 aumentó con el estrés corto previo
a la extinción de la memoria en todos los tiempos estudiados (t(8) = -10,011,
p<0,001, E1-EXT-0H>BASAL; t(8) = -8,529, p<0,001, E1-EXT-1H>BASAL; t(8) = -
13,076, p<0,001, E1-EXT-4H>BASAL) (Figura 20).
0
50
100
150
200
250
300
350
400
BASAL E1-0H E1-1H E1-4H EXT-0H EXT-1H EXT-4H E1-EXT-0H E1-EXT-1H E1-EXT-4H
ΔO
.D.
450 n
m
pERK1/2
Figura 20. Cambios en la actividad enzimática de ERK1/2 inducidos por el estrés corto
previo al entrenamiento en la extinción de la memoria espacial. Diagrama de barras
ilustrando el cambio en la O.D. con respecto al grupo basal y al grupo de entrenamiento en la
extinción. Se ilustra la media y el error estándar (n= 5 animales por grupo). También se ilustra el
grupo de estrés corto. E1-EXT-0H, animales entrenados en la adquisición, 24 h después
estresados durante 1 h, 30 min después sometidos a pruebas de recuperación y extinción de la
memoria espacial y sacrificados inmediatamente; E1-EXT-1H, sacrificados 1 h después; E1-
EXT-4H, sacrificados 4 h después. ***, p<0,001.
Por otra parte, la actividad de ERK1/2, en condiciones de estrés prolongado
previo a la extinción, aumentó significativamente en las siguientes condiciones
(t(8) = -10,028, p<0,001, E4-EXT-0H>BASAL; t(8) = -4,462, p = 0,002, E4-EXT-
1H>BASAL), pero comparativamente con el grupo de extinción, la actividad de
*** *********
******
*
***

61
ERK1/2 disminuyó significativamente con el estrés prolongado previo a la
extinción y el sacrificio 4 h después (t(4) = 24,505, p<0,001, E4-EXT-4H<EXT-4H)
(Figura 21 y 22).
0
50
100
150
200
250
300
350
400
BASAL E4-0H E4-1H E4-4H EXT-0H EXT-1H EXT-4H E4-EXT-0H E4-EXT-1H E4-EXT-4H
ΔO
.D.
450 n
m
pERK1/2
Figura 21. Cambios en la actividad enzimática de ERK1/2 inducidos por el estrés
prolongado previo al entrenamiento en la extinción de la memoria espacial. Diagrama de
barras ilustrando el cambio en la O.D. con respecto al grupo basal y al grupo de entrenamiento
en la extinción. Se ilustra la media y el error estándar (n= 5 animales por grupo). También se
ilustra el grupo de estrés prolongado. E4-EXT-0H, animales entrenados en la adquisición, 24 h
después estresados durante 4 h, 30 min después sometidos a pruebas de recuperación y
extinción de la memoria espacial y sacrificados inmediatamente; E4-EXT-1H, sacrificados 1 h
después; E4-EXT-4H, sacrificados 4 h después. **, p<0,01; ***,p<0,001.
**
***
***
*** ******
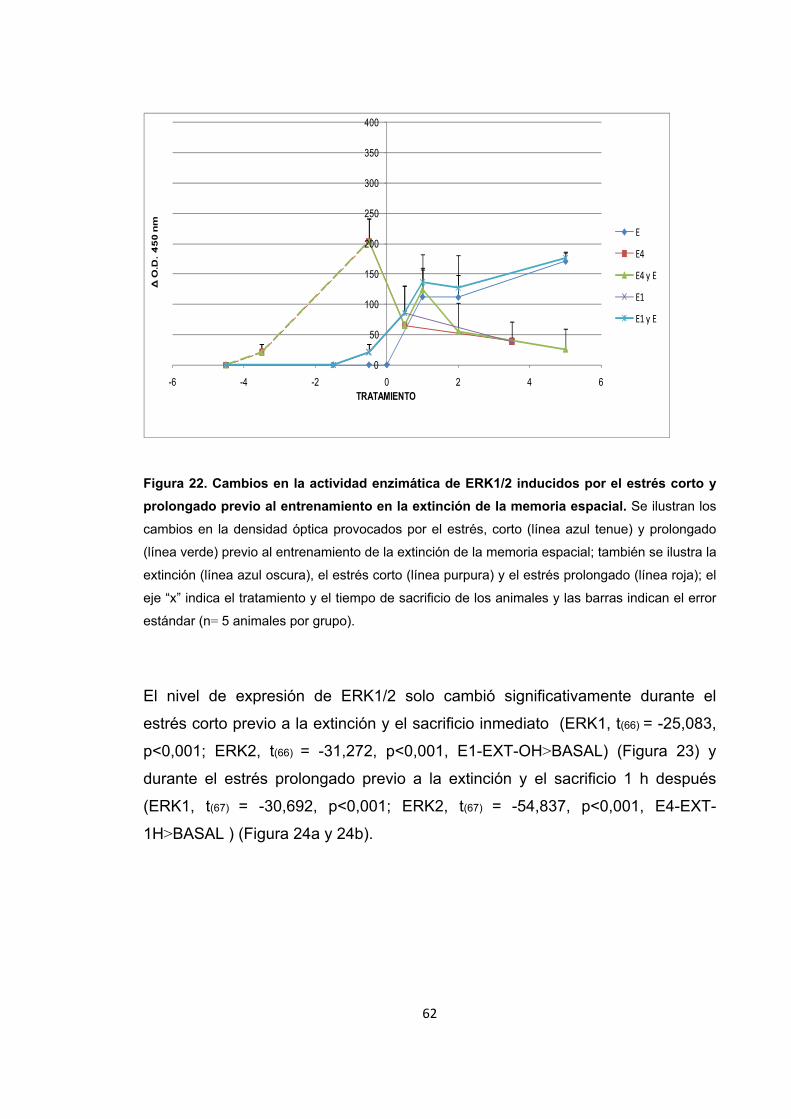
62
0
50
100
150
200
250
300
350
400
-6 -4 -2 0 2 4 6
ΔO
.D.
45
0 n
m
TRATAMIENTO
E
E4
E4 y E
E1
E1 y E
Figura 22. Cambios en la actividad enzimática de ERK1/2 inducidos por el estrés corto y
prolongado previo al entrenamiento en la extinción de la memoria espacial. Se ilustran los
cambios en la densidad óptica provocados por el estrés, corto (línea azul tenue) y prolongado
(línea verde) previo al entrenamiento de la extinción de la memoria espacial; también se ilustra la
extinción (línea azul oscura), el estrés corto (línea purpura) y el estrés prolongado (línea roja); el
eje “x” indica el tratamiento y el tiempo de sacrificio de los animales y las barras indican el error
estándar (n= 5 animales por grupo).
El nivel de expresión de ERK1/2 solo cambió significativamente durante el
estrés corto previo a la extinción y el sacrificio inmediato (ERK1, t(66) = -25,083,
p<0,001; ERK2, t(66) = -31,272, p<0,001, E1-EXT-OH>BASAL) (Figura 23) y
durante el estrés prolongado previo a la extinción y el sacrificio 1 h después
(ERK1, t(67) = -30,692, p<0,001; ERK2, t(67) = -54,837, p<0,001, E4-EXT-
1H>BASAL ) (Figura 24a y 24b).

63
-30
-20
-10
0
10
20
30
40
50
60
BASAL E1-EXT-0H E1-EXT-1H E1-EXT-4H
CA
NT
IDA
D N
OR
MA
LIZ
AD
A (
%B
AS
AL
)
ERK1/2
ERK 1
ERK2
Figura 23. Cambios en el nivel de expresión de ERK1/2 inducidos por el estrés corto
previo al entrenamiento de la extinción de la memoria espacial. Diagrama de barras
ilustrando el cambio en el nivel de expresión de ERK1/2 con respecto al basal. Se ilustra la
media y el error estándar (n=5 animales por grupo). ***, p<0,001.
a.
-20
-10
0
10
20
30
40
50
60
BASAL E4-EXT-0H E4-EXT-1H E4-EXT-4H
CA
NT
IDA
D N
OR
MA
LIZ
AD
A (
%B
AS
AL
)
ERK1/2
ERK 1
ERK2
b. 1 2 3 4 5 6 7 8 MW (kDa) 9 10 11 12 13 14 15 16 MW
198,844 115,7 96,743 53,541
*** ***
*** ***
ERK1 ERK2
(1)
(2)

64
Figura 24. Cambios en el nivel de expresión de ERK1/2 provocados por el estrés
prolongado previo al entrenamiento en la extinción de la memoria espacial. a). Diagrama
de barras ilustrando el cambio en el nivel de expresión de ERK1/2 con respecto al basal. Se
ilustra la media y el error estándar (n=5 animales por grupo). ***, p<0,001. b). (1). SDS-PAGE y
(2). WB; Carriles 1, 2, 9 y 10 muestras en condiciones basales; 3, 4, 11 y 12 en condiciones de
estrés severo, extinción y sacrificio inmediato; 5, 6, 13 y 14 en condiciones de estrés severo,
extinción y sacrificio 1 h después y 7, 8, 15 y 16 estrés severo, extinción y sacrificio 4 h después.
MW, marcador de peso molecular (Estándar pre-teñido de amplio rango, BIO-RAD).
6.4. Efecto del estrés por inmovilización en la expresión de PP2B
en el hipocampo de ratas
Los resultados obtenidos por WB, mostraron que el estrés por inmovilización,
tanto corto como prolongado, en términos generales, no afectó el nivel de
expresión de PP2B, sin embargo, como se muestra en la figura 25, hubo una
disminución significativa en el nivel de expresión de PP2B 4 h después de la
inmovilización corta (t(69) = 6,449, p<0,001 E1-4H<BASAL) (figura A6 en anexos).
-40
-30
-20
-10
0
10
20
30
BASAL E1-0H E1-1H E1-4H E4-0H E4-1H E4-4H
CA
NT
IDA
D N
OR
MA
LIZ
AD
A (
%B
AS
AL
)
PP2B
Figura 25. Cambios en el nivel de expresión de PP2B provocados por el estrés por
inmovilización corto y prolongado. Diagrama de barras ilustrando el cambio en el nivel de
expresión de PP2B con respecto al basal; se ilustra la media y el error estándar (n=5 animales
por grupo). ***, p<0,001.
***
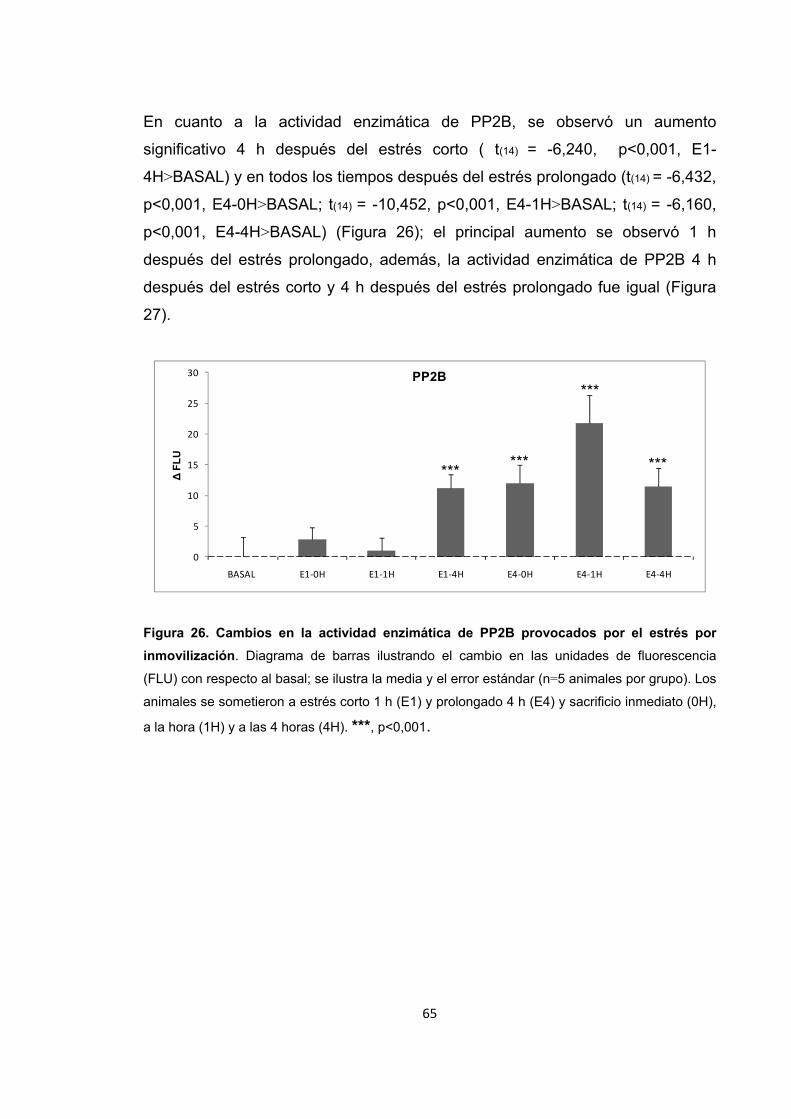
65
En cuanto a la actividad enzimática de PP2B, se observó un aumento
significativo 4 h después del estrés corto ( t(14) = -6,240, p<0,001, E1-
4H>BASAL) y en todos los tiempos después del estrés prolongado (t(14) = -6,432,
p<0,001, E4-0H>BASAL; t(14) = -10,452, p<0,001, E4-1H>BASAL; t(14) = -6,160,
p<0,001, E4-4H>BASAL) (Figura 26); el principal aumento se observó 1 h
después del estrés prolongado, además, la actividad enzimática de PP2B 4 h
después del estrés corto y 4 h después del estrés prolongado fue igual (Figura
27).
0
5
10
15
20
25
30
BASAL E1‐0H E1‐1H E1‐4H E4‐0H E4‐1H E4‐4H
ΔF
LU
PP2B
Figura 26. Cambios en la actividad enzimática de PP2B provocados por el estrés por
inmovilización. Diagrama de barras ilustrando el cambio en las unidades de fluorescencia
(FLU) con respecto al basal; se ilustra la media y el error estándar (n=5 animales por grupo). Los
animales se sometieron a estrés corto 1 h (E1) y prolongado 4 h (E4) y sacrificio inmediato (0H),
a la hora (1H) y a las 4 horas (4H). ***, p<0,001.
****** ***
***

66
0
5
10
15
20
25
30
‐5 ‐4 ‐3 ‐2 ‐1 0 1 2 3 4
ΔF
LU
TRATAMIENTO
E4
E1
Figura 27. Cambios en la actividad enzimática de PP2B provocados por el estrés por
inmovilización. Se ilustran los cambios en las unidades de fluorescencia (FLU) provocados por
el estrés corto (línea oscura) y el estrés prolongado (línea clara); el eje “x” indica el tratamiento y
el tiempo de sacrificio de los animales. Se ilustra el error estándar (n=5 animales por grupo).
6.5. Actividad y nivel de expresión de PP2B durante los procesos
de formación de memoria
El entrenamiento en una tarea espacial en el laberinto circular de Barnes
provocó cambios en la actividad enzimática de PP2B durante la adquisición, la
recuperación y la extinción de la memoria, comparativamente con el grupo
basal. Este cambio se caracterizó principalmente por un aumento en todos los
tiempos estudiados (t(14) = -9,735, p <0,001, ADQ-0H>BASAL; t(14) = -8,106, p
<0,001, ADQ-1H>BASAL; t(14) = -7,283, p <0,001, ADQ-4H>BASAL. Figura 28);
(t(14) = -12,973, p <0,001, REC-0H>BASAL; t(14) = -5,558, p <0,001; REC-
1H>BASAL; t(14) = -6,965, p <0,001, REC-4H>BASAL. Figura 29); (t(14) = -8,516,
p <0,001, EXT-0H>BASAL; t(14) = -8,749, p <0,001, EXT-1H>BASAL ; t(14) = -
6,835, p <0,001, EXT-4H>BASAL. Figura 30).
E4H
E1H
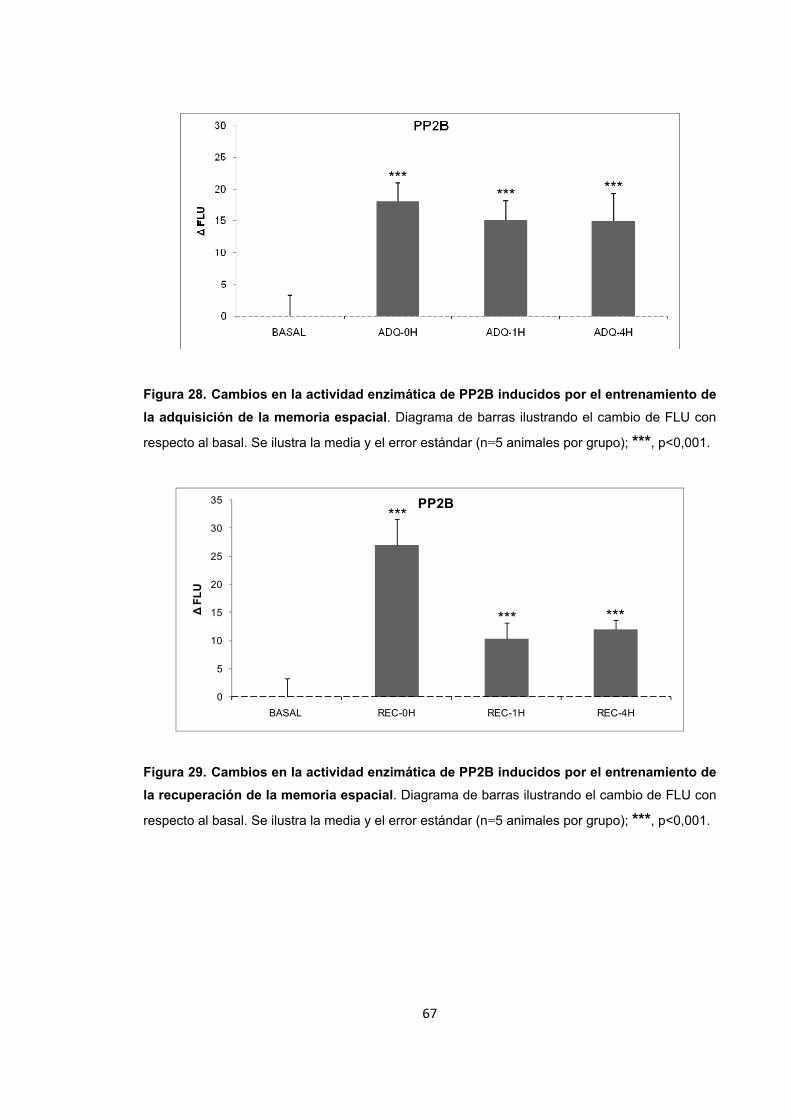
67
Figura 28. Cambios en la actividad enzimática de PP2B inducidos por el entrenamiento de
la adquisición de la memoria espacial. Diagrama de barras ilustrando el cambio de FLU con
respecto al basal. Se ilustra la media y el error estándar (n=5 animales por grupo); ***, p<0,001.
0
5
10
15
20
25
30
35
BASAL REC-0H REC-1H REC-4H
ΔF
LU
PP2B
Figura 29. Cambios en la actividad enzimática de PP2B inducidos por el entrenamiento de
la recuperación de la memoria espacial. Diagrama de barras ilustrando el cambio de FLU con
respecto al basal. Se ilustra la media y el error estándar (n=5 animales por grupo); ***, p<0,001.
****** ***
***
*** ***
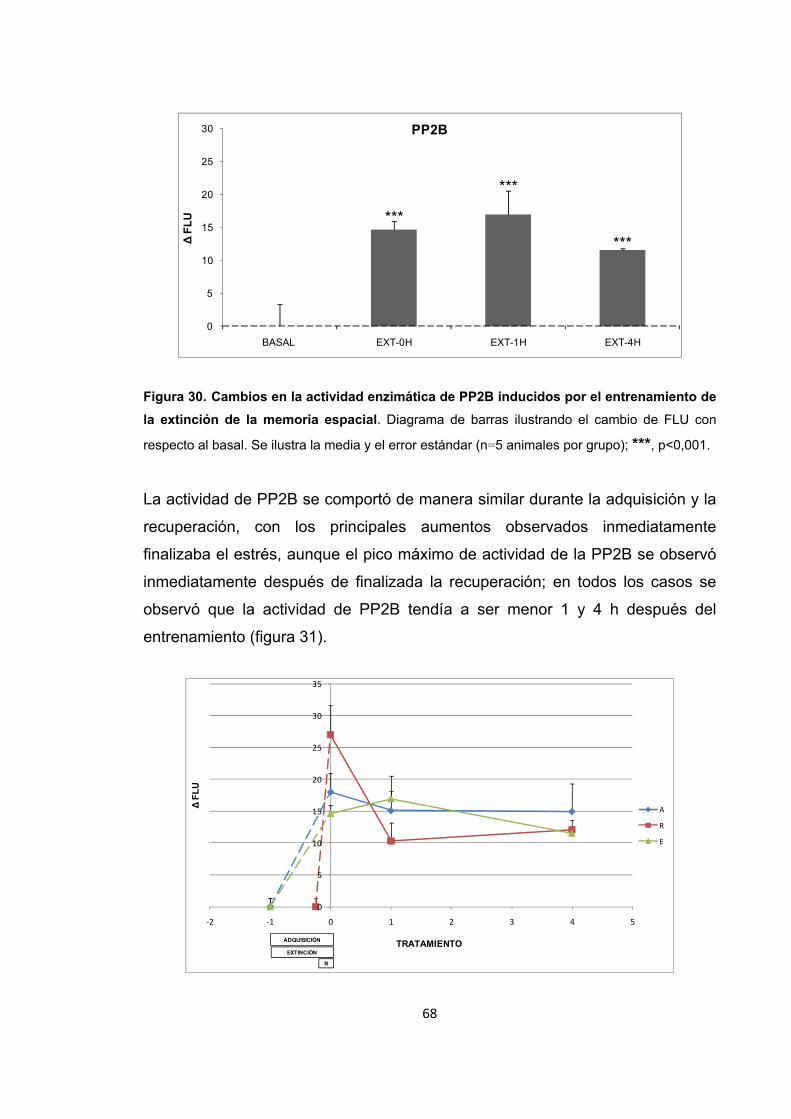
68
0
5
10
15
20
25
30
BASAL EXT-0H EXT-1H EXT-4H
ΔF
LU
PP2B
Figura 30. Cambios en la actividad enzimática de PP2B inducidos por el entrenamiento de
la extinción de la memoria espacial. Diagrama de barras ilustrando el cambio de FLU con
respecto al basal. Se ilustra la media y el error estándar (n=5 animales por grupo); ***, p<0,001.
La actividad de PP2B se comportó de manera similar durante la adquisición y la
recuperación, con los principales aumentos observados inmediatamente
finalizaba el estrés, aunque el pico máximo de actividad de la PP2B se observó
inmediatamente después de finalizada la recuperación; en todos los casos se
observó que la actividad de PP2B tendía a ser menor 1 y 4 h después del
entrenamiento (figura 31).
0
5
10
15
20
25
30
35
‐2 ‐1 0 1 2 3 4 5
ΔF
LU
TRATAMIENTO
A
R
E
ADQUISICIÓN
EXTINCIÓN
R
***
***
***
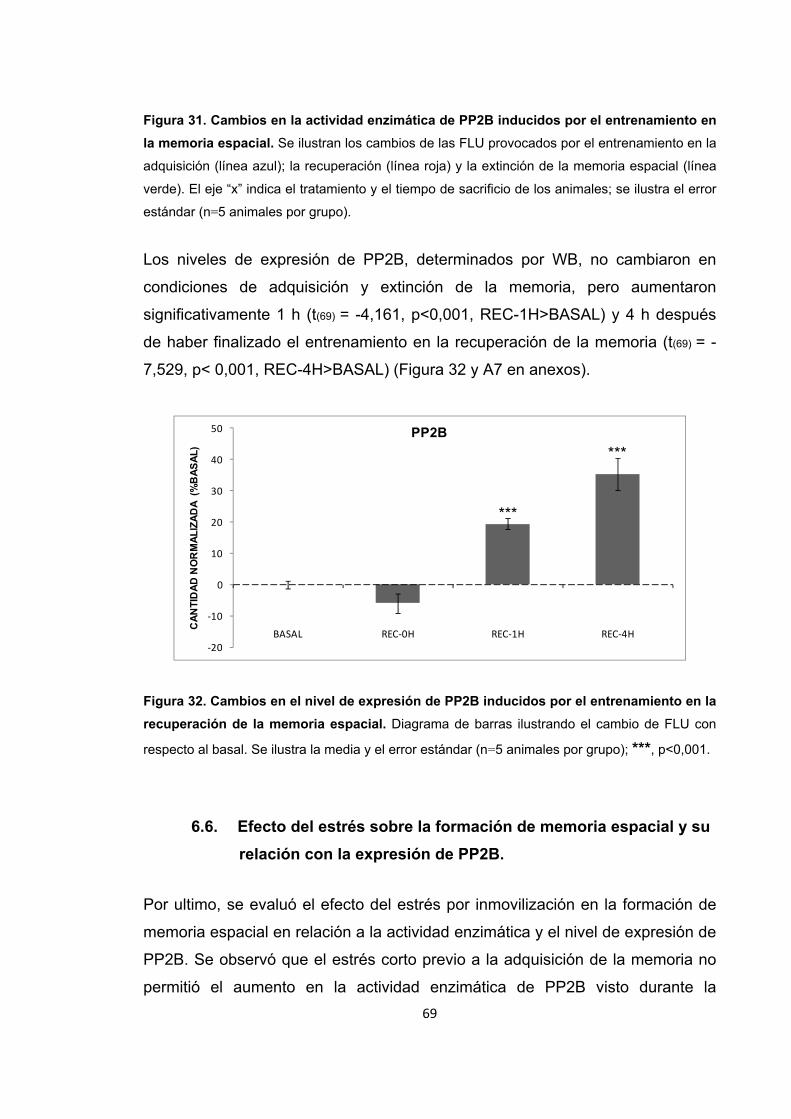
69
Figura 31. Cambios en la actividad enzimática de PP2B inducidos por el entrenamiento en
la memoria espacial. Se ilustran los cambios de las FLU provocados por el entrenamiento en la
adquisición (línea azul); la recuperación (línea roja) y la extinción de la memoria espacial (línea
verde). El eje “x” indica el tratamiento y el tiempo de sacrificio de los animales; se ilustra el error
estándar (n=5 animales por grupo).
Los niveles de expresión de PP2B, determinados por WB, no cambiaron en
condiciones de adquisición y extinción de la memoria, pero aumentaron
significativamente 1 h (t(69) = -4,161, p<0,001, REC-1H>BASAL) y 4 h después
de haber finalizado el entrenamiento en la recuperación de la memoria (t(69) = -
7,529, p< 0,001, REC-4H>BASAL) (Figura 32 y A7 en anexos).
‐20
‐10
0
10
20
30
40
50
BASAL REC‐0H REC‐1H REC‐4H
CA
NT
IDA
D N
OR
MA
LIZ
AD
A (
%B
AS
AL
)
PP2B
Figura 32. Cambios en el nivel de expresión de PP2B inducidos por el entrenamiento en la
recuperación de la memoria espacial. Diagrama de barras ilustrando el cambio de FLU con
respecto al basal. Se ilustra la media y el error estándar (n=5 animales por grupo); ***, p<0,001.
6.6. Efecto del estrés sobre la formación de memoria espacial y su
relación con la expresión de PP2B.
Por ultimo, se evaluó el efecto del estrés por inmovilización en la formación de
memoria espacial en relación a la actividad enzimática y el nivel de expresión de
PP2B. Se observó que el estrés corto previo a la adquisición de la memoria no
permitió el aumento en la actividad enzimática de PP2B visto durante la
***
***
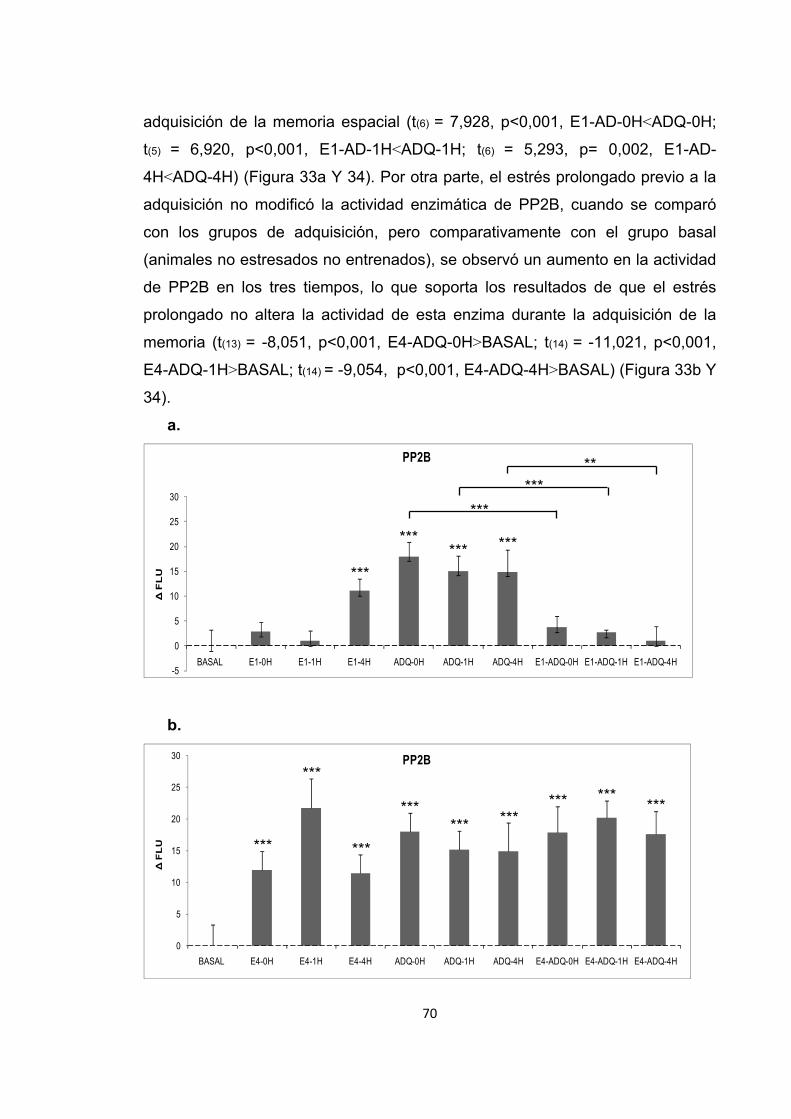
70
adquisición de la memoria espacial (t(6) = 7,928, p<0,001, E1-AD-0H<ADQ-0H;
t(5) = 6,920, p<0,001, E1-AD-1H<ADQ-1H; t(6) = 5,293, p= 0,002, E1-AD-
4H<ADQ-4H) (Figura 33a Y 34). Por otra parte, el estrés prolongado previo a la
adquisición no modificó la actividad enzimática de PP2B, cuando se comparó
con los grupos de adquisición, pero comparativamente con el grupo basal
(animales no estresados no entrenados), se observó un aumento en la actividad
de PP2B en los tres tiempos, lo que soporta los resultados de que el estrés
prolongado no altera la actividad de esta enzima durante la adquisición de la
memoria (t(13) = -8,051, p<0,001, E4-ADQ-0H>BASAL; t(14) = -11,021, p<0,001,
E4-ADQ-1H>BASAL; t(14) = -9,054, p<0,001, E4-ADQ-4H>BASAL) (Figura 33b Y
34).
a.
-5
0
5
10
15
20
25
30
BASAL E1-0H E1-1H E1-4H ADQ-0H ADQ-1H ADQ-4H E1-ADQ-0H E1-ADQ-1H E1-ADQ-4H
ΔF
LU
PP2B
b.
0
5
10
15
20
25
30
BASAL E4-0H E4-1H E4-4H ADQ-0H ADQ-1H ADQ-4H E4-ADQ-0H E4-ADQ-1H E4-ADQ-4H
ΔF
LU
PP2B
***
*****
***
*** ******
***
*** ***
******
***
***
***
***
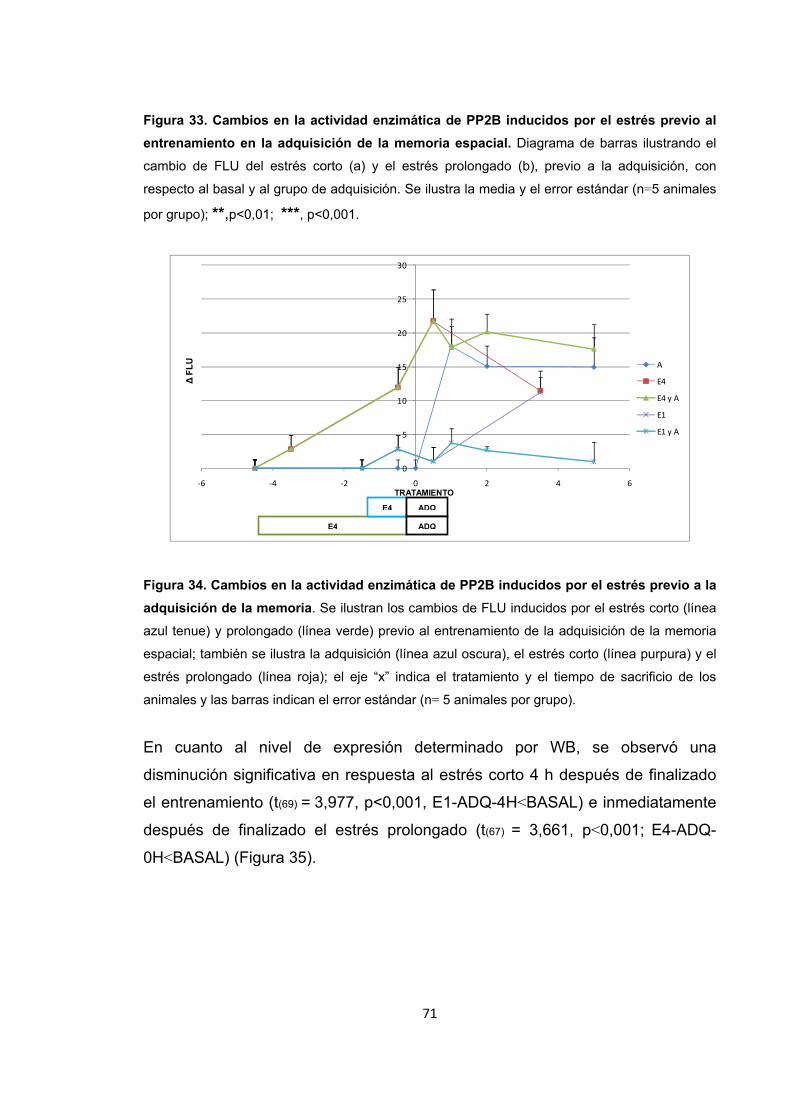
71
Figura 33. Cambios en la actividad enzimática de PP2B inducidos por el estrés previo al
entrenamiento en la adquisición de la memoria espacial. Diagrama de barras ilustrando el
cambio de FLU del estrés corto (a) y el estrés prolongado (b), previo a la adquisición, con
respecto al basal y al grupo de adquisición. Se ilustra la media y el error estándar (n=5 animales
por grupo); **,p<0,01; ***, p<0,001.
0
5
10
15
20
25
30
‐6 ‐4 ‐2 0 2 4 6
ΔF
LU
TRATAMIENTO
A
E4
E4 y A
E1
E1 y A
Figura 34. Cambios en la actividad enzimática de PP2B inducidos por el estrés previo a la
adquisición de la memoria. Se ilustran los cambios de FLU inducidos por el estrés corto (línea
azul tenue) y prolongado (línea verde) previo al entrenamiento de la adquisición de la memoria
espacial; también se ilustra la adquisición (línea azul oscura), el estrés corto (línea purpura) y el
estrés prolongado (línea roja); el eje “x” indica el tratamiento y el tiempo de sacrificio de los
animales y las barras indican el error estándar (n= 5 animales por grupo).
En cuanto al nivel de expresión determinado por WB, se observó una
disminución significativa en respuesta al estrés corto 4 h después de finalizado
el entrenamiento (t(69) = 3,977, p<0,001, E1-ADQ-4H<BASAL) e inmediatamente
después de finalizado el estrés prolongado (t(67) = 3,661, p<0,001; E4-ADQ-
0H<BASAL) (Figura 35).
E4
E4 ADQ
ADQ

72
-30
-20
-10
0
10
20
30
BASAL E1-ADQ-0H E1-ADQ-1H E1-ADQ-4H E4-ADQ-0H E4-ADQ-1H E4-ADQ-4H
CA
NT
IDA
D N
OR
MA
LIZ
AD
A (%
BA
SA
L)
PP2B
Figura 35. Cambios en el nivel de expresión de PP2B inducidos por el estrés corto y
prolongado previo al entrenamiento de la adquisición de la memoria espacial. Diagrama de
barras ilustrando el cambio en el nivel de expresión de PP2B con respecto al basal durante el
estrés previo a la adquisición. Se ilustra la media y el error estándar (n=5 animales por grupo).
***, p<0,001.
Se observó un comportamiento similar al evaluar los efectos del estrés sobre la
recuperación de la memoria, el estrés corto no permitió el incremento en la
actividad de PP2B visto durante la recuperación de la memoria espacial (t(6) =
5,850, p = 0,001, E1-REC-0H<REC-OH; t(6) = 3,157, p = 0,020, E1-REC-
1H<REC-1H; t(6) = 6,772, p<0,001, E1-REC-4H<REC-4H. Figura 36a y 37); así
mismo, el estrés prolongado previo a la recuperación de la memoria no altero la
actividad enzimatica de PP2B, pero comparativamente con el grupo basal, la
actividad de esta enzima aumento significativamente (t(14) = -8,074, p<0,001, E4-
REC-0H>BASAL; t(14) = -4,896, p <0,001, E4-REC-1H>BASAL; t(14) = -9,597, p
<0,001, E4-REC-4H>BASAL. Figura 36b y 37).
******

73
a.
0
5
10
15
20
25
30
35
BASAL E1-0H E1-1H E1-4H REC-0H REC-1H REC-4H E1-REC-0H E1-REC-1H E1-REC-4H
ΔF
LU
PP2B
b.
0
5
10
15
20
25
30
35
BASAL E4-0H E4-1H E4-4H REC-0H REC-1H REC-4H E4-REC-0H E4-REC-1H E4-REC-4H
ΔF
LU
PP2B
Figura 36. Cambios en la actividad enzimática de PP2B inducidos por el estrés previo al
entrenamiento en la recuperación de la memoria espacial. Diagrama de barras ilustrando el
cambio de FLU del estrés corto (a) y el prolongado (b), previo a la recuperación, con respecto al
basal y al grupo de recuperación. Se ilustra la media y el error estándar (n=5 animales por
grupo); *, p<0,05; ***, p<0,001.
****
***
***
***
***
*** ******
***
***
***
***
***
*** ***

74
0
5
10
15
20
25
30
35
-6 -4 -2 0 2 4 6
ΔF
LU
TRATAMIENTO
R
E4
E4 y R
E1
E1 y R
Figura 37. Cambios en la actividad enzimática de PP2B inducidos por el estrés previo a la
recuperación de la memoria. Se ilustran los cambios de FLU provocados por el estrés, corto
(línea azul tenue) y prolongado (línea verde) previo al entrenamiento de la recuperación de la
memoria espacial; también se ilustra la recuperación (línea azul oscura), el estrés corto (línea
purpura) y el estrés prolongado (línea roja); el eje “x” indica el tratamiento y el tiempo de
sacrificio de los animales y las barras indican el error estándar (n= 5 animales por grupo).
En cuanto al nivel de la expresión de PP2B, los análisis por WB mostraron una
disminución significativa en los grupos de estrés prolongado previo a la
recuperación (t(68) = 4,743, p<0,001, E4-REC-1H<BASAL; t(68) = 4,879, p<0,001,
E4-REC-4H<BASAL) (Figura 38).
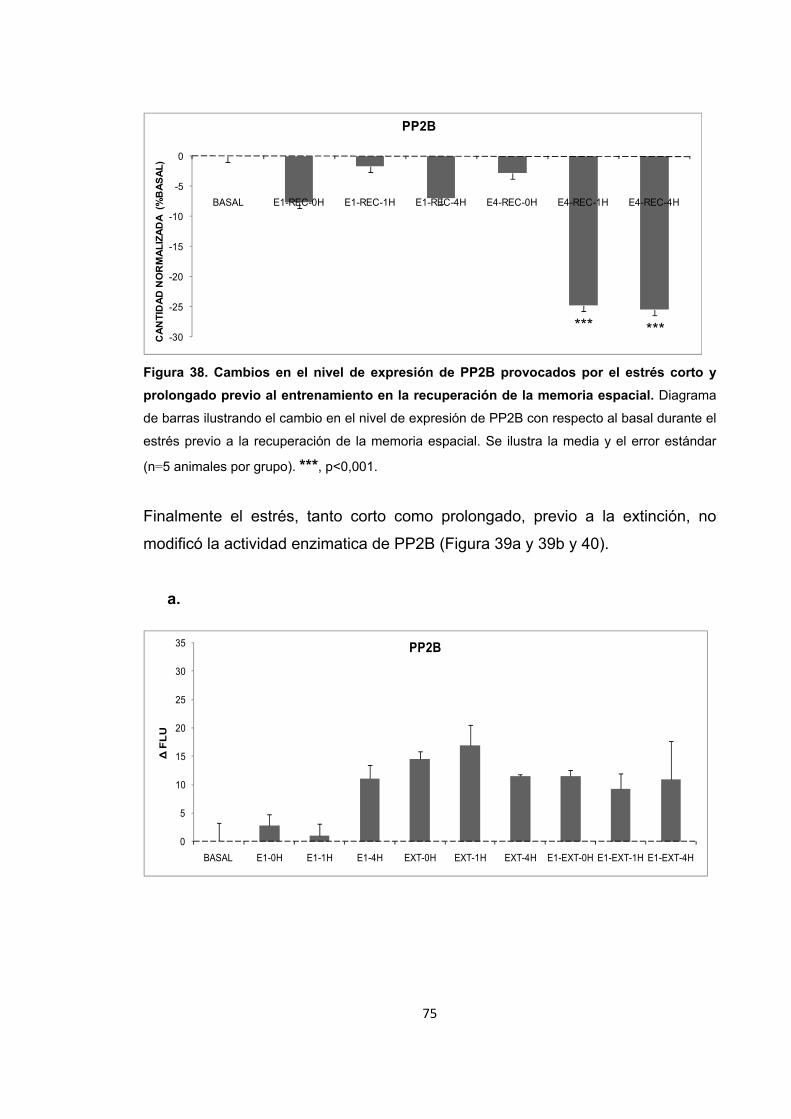
75
-30
-25
-20
-15
-10
-5
0
BASAL E1-REC-0H E1-REC-1H E1-REC-4H E4-REC-0H E4-REC-1H E4-REC-4H
CA
NT
IDA
D N
OR
MA
LIZ
AD
A (
%B
AS
AL
)
PP2B
Figura 38. Cambios en el nivel de expresión de PP2B provocados por el estrés corto y
prolongado previo al entrenamiento en la recuperación de la memoria espacial. Diagrama
de barras ilustrando el cambio en el nivel de expresión de PP2B con respecto al basal durante el
estrés previo a la recuperación de la memoria espacial. Se ilustra la media y el error estándar
(n=5 animales por grupo). ***, p<0,001.
Finalmente el estrés, tanto corto como prolongado, previo a la extinción, no
modificó la actividad enzimatica de PP2B (Figura 39a y 39b y 40).
a.
0
5
10
15
20
25
30
35
BASAL E1-0H E1-1H E1-4H EXT-0H EXT-1H EXT-4H E1-EXT-0H E1-EXT-1H E1-EXT-4H
ΔF
LU
PP2B
*** ***
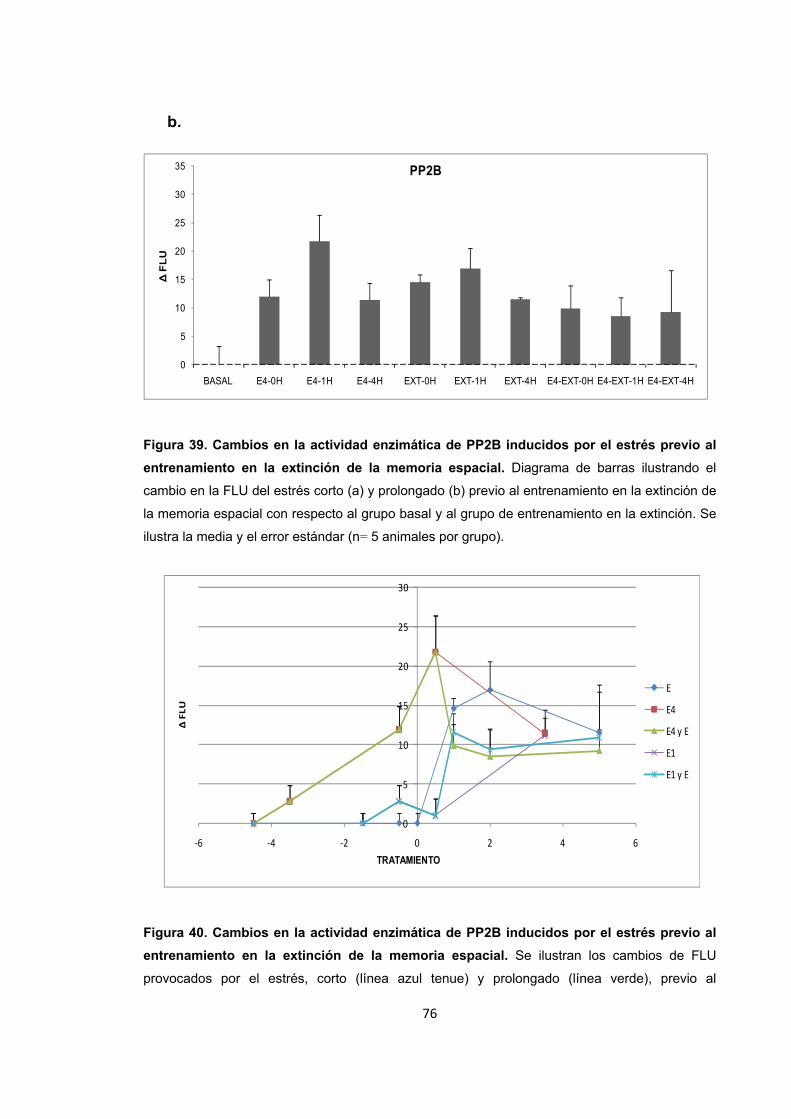
76
b.
0
5
10
15
20
25
30
35
BASAL E4-0H E4-1H E4-4H EXT-0H EXT-1H EXT-4H E4-EXT-0H E4-EXT-1H E4-EXT-4H
ΔF
LU
PP2B
Figura 39. Cambios en la actividad enzimática de PP2B inducidos por el estrés previo al
entrenamiento en la extinción de la memoria espacial. Diagrama de barras ilustrando el
cambio en la FLU del estrés corto (a) y prolongado (b) previo al entrenamiento en la extinción de
la memoria espacial con respecto al grupo basal y al grupo de entrenamiento en la extinción. Se
ilustra la media y el error estándar (n= 5 animales por grupo).
0
5
10
15
20
25
30
‐6 ‐4 ‐2 0 2 4 6
ΔFLU
TRATAMIENTO
E
E4
E4 y E
E1
E1 y E
Figura 40. Cambios en la actividad enzimática de PP2B inducidos por el estrés previo al
entrenamiento en la extinción de la memoria espacial. Se ilustran los cambios de FLU
provocados por el estrés, corto (línea azul tenue) y prolongado (línea verde), previo al
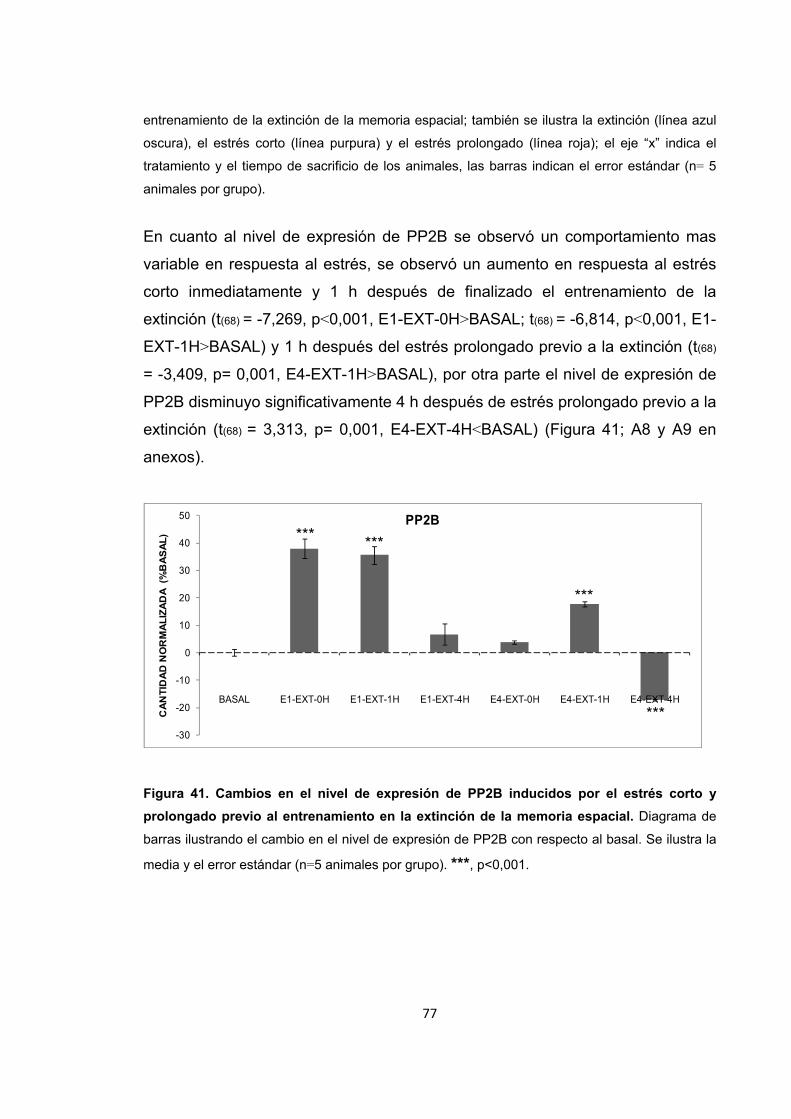
77
entrenamiento de la extinción de la memoria espacial; también se ilustra la extinción (línea azul
oscura), el estrés corto (línea purpura) y el estrés prolongado (línea roja); el eje “x” indica el
tratamiento y el tiempo de sacrificio de los animales, las barras indican el error estándar (n= 5
animales por grupo).
En cuanto al nivel de expresión de PP2B se observó un comportamiento mas
variable en respuesta al estrés, se observó un aumento en respuesta al estrés
corto inmediatamente y 1 h después de finalizado el entrenamiento de la
extinción (t(68) = -7,269, p<0,001, E1-EXT-0H>BASAL; t(68) = -6,814, p<0,001, E1-
EXT-1H>BASAL) y 1 h después del estrés prolongado previo a la extinción (t(68)
= -3,409, p= 0,001, E4-EXT-1H>BASAL), por otra parte el nivel de expresión de
PP2B disminuyo significativamente 4 h después de estrés prolongado previo a la
extinción (t(68) = 3,313, p= 0,001, E4-EXT-4H<BASAL) (Figura 41; A8 y A9 en
anexos).
-30
-20
-10
0
10
20
30
40
50
BASAL E1-EXT-0H E1-EXT-1H E1-EXT-4H E4-EXT-0H E4-EXT-1H E4-EXT-4H
CA
NT
IDA
D N
OR
MA
LIZ
AD
A (
%B
AS
AL
)
PP2B
Figura 41. Cambios en el nivel de expresión de PP2B inducidos por el estrés corto y
prolongado previo al entrenamiento en la extinción de la memoria espacial. Diagrama de
barras ilustrando el cambio en el nivel de expresión de PP2B con respecto al basal. Se ilustra la
media y el error estándar (n=5 animales por grupo). ***, p<0,001.
*** ***
***
***
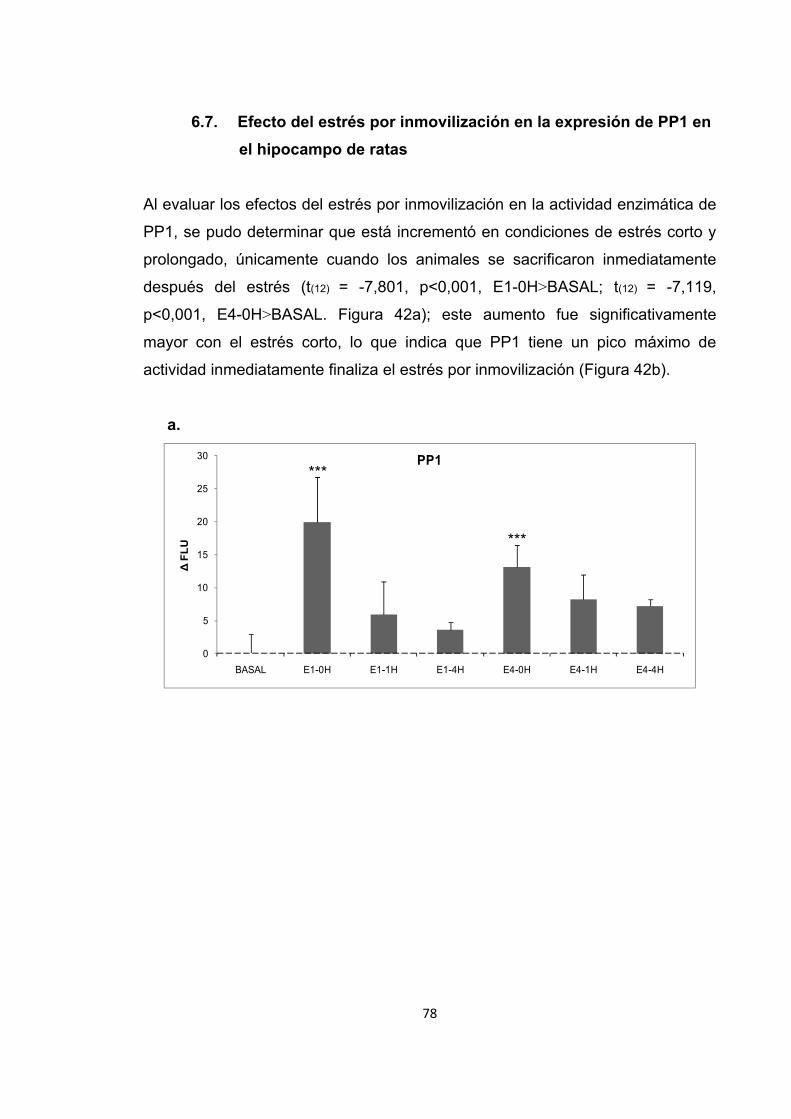
78
6.7. Efecto del estrés por inmovilización en la expresión de PP1 en
el hipocampo de ratas
Al evaluar los efectos del estrés por inmovilización en la actividad enzimática de
PP1, se pudo determinar que está incrementó en condiciones de estrés corto y
prolongado, únicamente cuando los animales se sacrificaron inmediatamente
después del estrés (t(12) = -7,801, p<0,001, E1-0H>BASAL; t(12) = -7,119,
p<0,001, E4-0H>BASAL. Figura 42a); este aumento fue significativamente
mayor con el estrés corto, lo que indica que PP1 tiene un pico máximo de
actividad inmediatamente finaliza el estrés por inmovilización (Figura 42b).
a.
0
5
10
15
20
25
30
BASAL E1-0H E1-1H E1-4H E4-0H E4-1H E4-4H
ΔF
LU
PP1
***
***
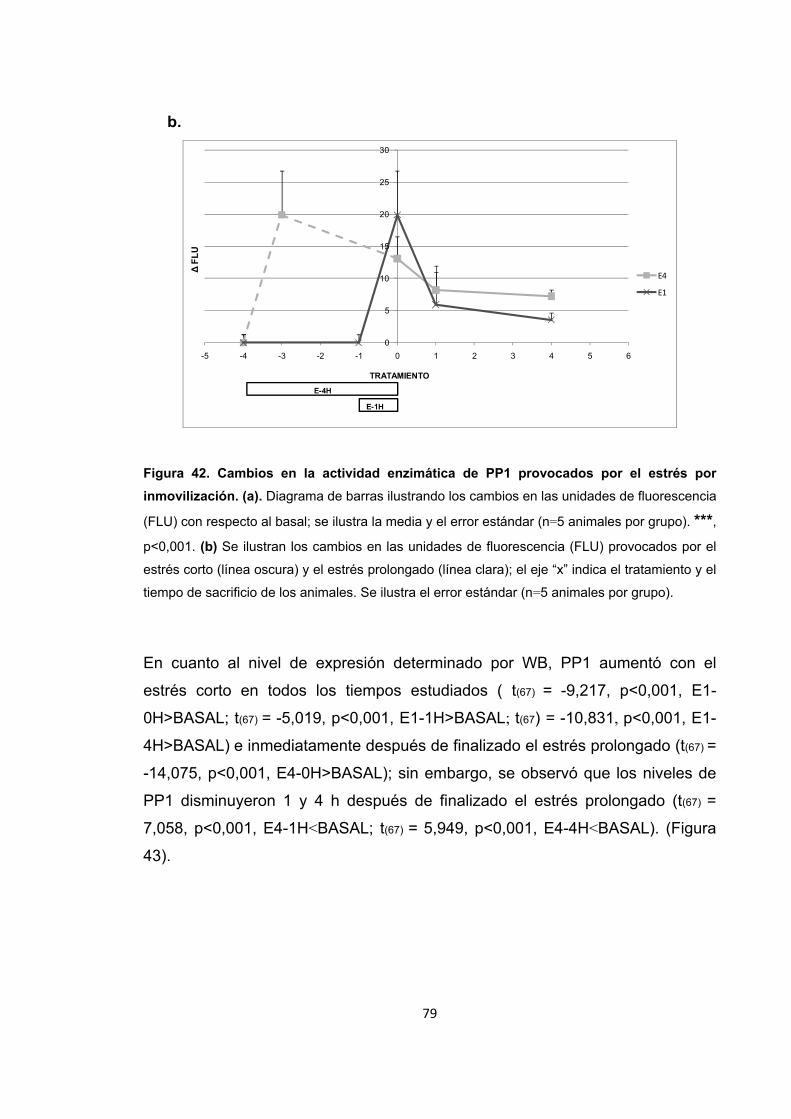
79
b.
0
5
10
15
20
25
30
-5 -4 -3 -2 -1 0 1 2 3 4 5 6
ΔF
LU
TRATAMIENTO
E4
E1
E-4H
E-1H
Figura 42. Cambios en la actividad enzimática de PP1 provocados por el estrés por
inmovilización. (a). Diagrama de barras ilustrando los cambios en las unidades de fluorescencia
(FLU) con respecto al basal; se ilustra la media y el error estándar (n=5 animales por grupo). ***,
p<0,001. (b) Se ilustran los cambios en las unidades de fluorescencia (FLU) provocados por el
estrés corto (línea oscura) y el estrés prolongado (línea clara); el eje “x” indica el tratamiento y el
tiempo de sacrificio de los animales. Se ilustra el error estándar (n=5 animales por grupo).
En cuanto al nivel de expresión determinado por WB, PP1 aumentó con el
estrés corto en todos los tiempos estudiados ( t(67) = -9,217, p<0,001, E1-
0H>BASAL; t(67) = -5,019, p<0,001, E1-1H>BASAL; t(67) = -10,831, p<0,001, E1-
4H>BASAL) e inmediatamente después de finalizado el estrés prolongado (t(67) =
-14,075, p<0,001, E4-0H>BASAL); sin embargo, se observó que los niveles de
PP1 disminuyeron 1 y 4 h después de finalizado el estrés prolongado (t(67) =
7,058, p<0,001, E4-1H<BASAL; t(67) = 5,949, p<0,001, E4-4H<BASAL). (Figura
43).
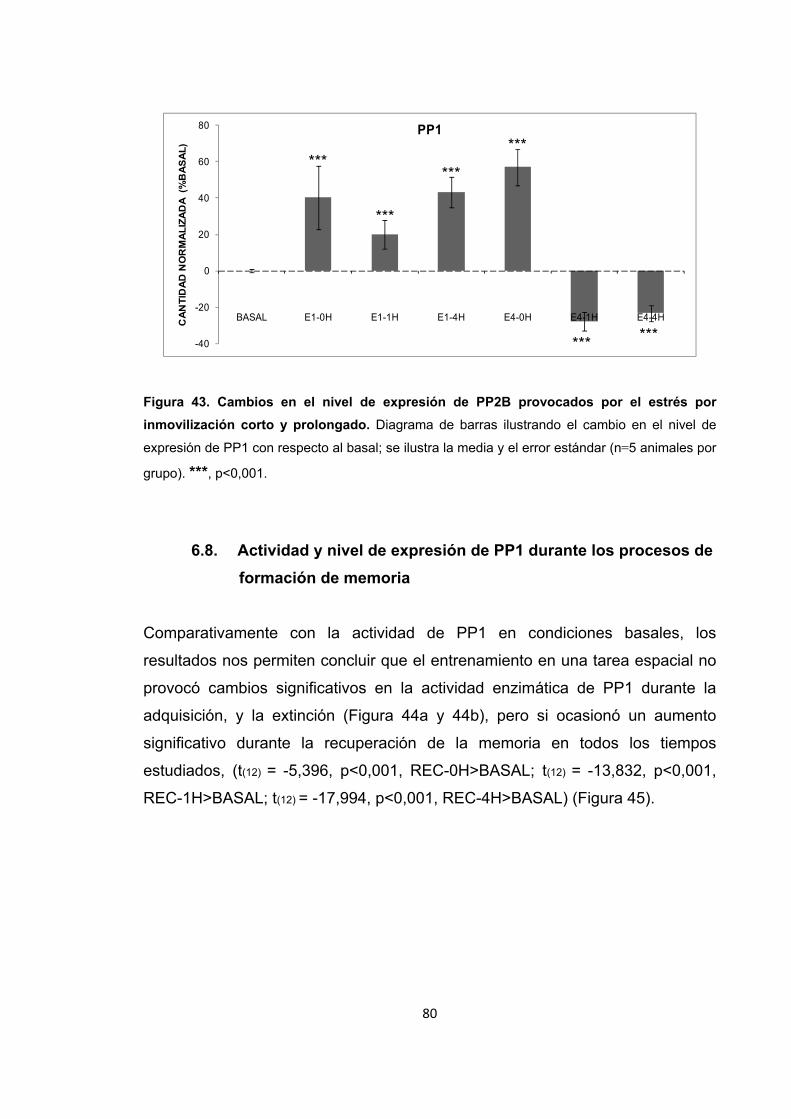
80
-40
-20
0
20
40
60
80
BASAL E1-0H E1-1H E1-4H E4-0H E4-1H E4-4H
CA
NT
IDA
D N
OR
MA
LIZ
AD
A (
%B
AS
AL
)
PP1
Figura 43. Cambios en el nivel de expresión de PP2B provocados por el estrés por
inmovilización corto y prolongado. Diagrama de barras ilustrando el cambio en el nivel de
expresión de PP1 con respecto al basal; se ilustra la media y el error estándar (n=5 animales por
grupo). ***, p<0,001.
6.8. Actividad y nivel de expresión de PP1 durante los procesos de
formación de memoria
Comparativamente con la actividad de PP1 en condiciones basales, los
resultados nos permiten concluir que el entrenamiento en una tarea espacial no
provocó cambios significativos en la actividad enzimática de PP1 durante la
adquisición, y la extinción (Figura 44a y 44b), pero si ocasionó un aumento
significativo durante la recuperación de la memoria en todos los tiempos
estudiados, (t(12) = -5,396, p<0,001, REC-0H>BASAL; t(12) = -13,832, p<0,001,
REC-1H>BASAL; t(12) = -17,994, p<0,001, REC-4H>BASAL) (Figura 45).
***
***
***
***
******
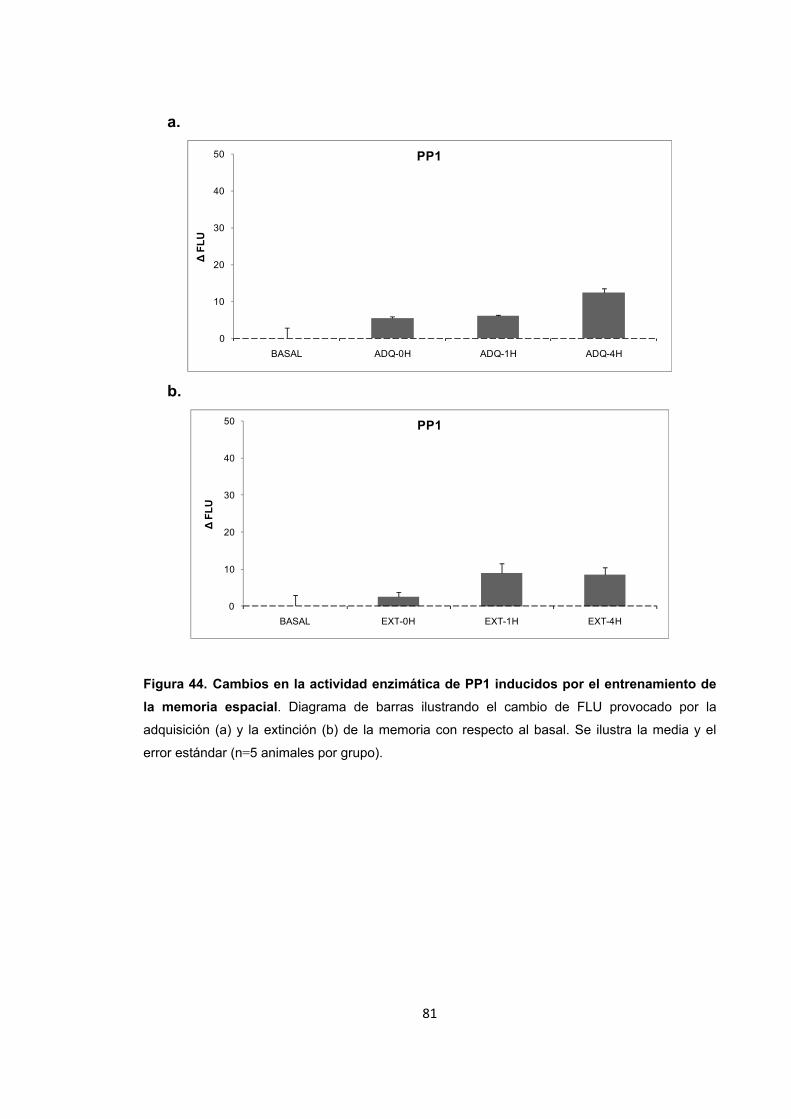
81
a.
0
10
20
30
40
50
BASAL ADQ-0H ADQ-1H ADQ-4H
ΔF
LU
PP1
b.
0
10
20
30
40
50
BASAL EXT-0H EXT-1H EXT-4H
ΔF
LU
PP1
Figura 44. Cambios en la actividad enzimática de PP1 inducidos por el entrenamiento de
la memoria espacial. Diagrama de barras ilustrando el cambio de FLU provocado por la
adquisición (a) y la extinción (b) de la memoria con respecto al basal. Se ilustra la media y el
error estándar (n=5 animales por grupo).
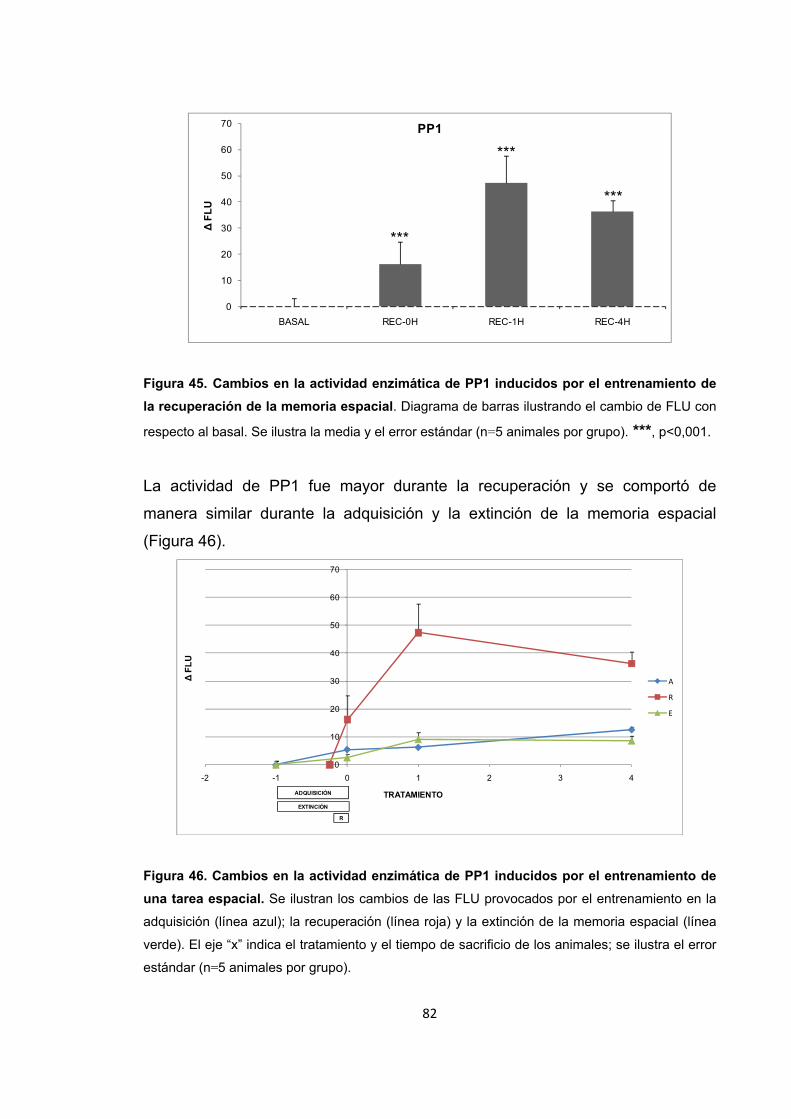
82
0
10
20
30
40
50
60
70
BASAL REC-0H REC-1H REC-4H
ΔF
LU
PP1
Figura 45. Cambios en la actividad enzimática de PP1 inducidos por el entrenamiento de
la recuperación de la memoria espacial. Diagrama de barras ilustrando el cambio de FLU con
respecto al basal. Se ilustra la media y el error estándar (n=5 animales por grupo). ***, p<0,001.
La actividad de PP1 fue mayor durante la recuperación y se comportó de
manera similar durante la adquisición y la extinción de la memoria espacial
(Figura 46).
0
10
20
30
40
50
60
70
-2 -1 0 1 2 3 4
ΔF
LU
TRATAMIENTO
A
R
E
ADQUISICIÓN
EXTINCIÓN
R
Figura 46. Cambios en la actividad enzimática de PP1 inducidos por el entrenamiento de
una tarea espacial. Se ilustran los cambios de las FLU provocados por el entrenamiento en la
adquisición (línea azul); la recuperación (línea roja) y la extinción de la memoria espacial (línea
verde). El eje “x” indica el tratamiento y el tiempo de sacrificio de los animales; se ilustra el error
estándar (n=5 animales por grupo).
***
***
***
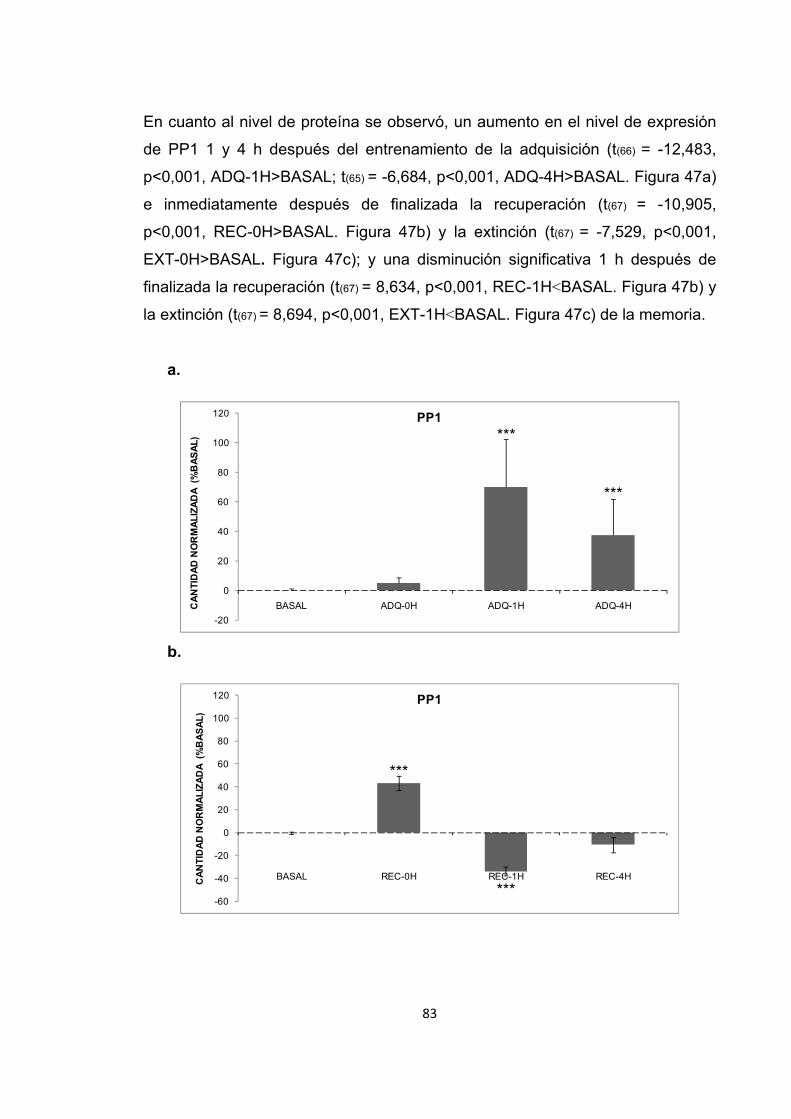
83
En cuanto al nivel de proteína se observó, un aumento en el nivel de expresión
de PP1 1 y 4 h después del entrenamiento de la adquisición (t(66) = -12,483,
p<0,001, ADQ-1H>BASAL; t(65) = -6,684, p<0,001, ADQ-4H>BASAL. Figura 47a)
e inmediatamente después de finalizada la recuperación (t(67) = -10,905,
p<0,001, REC-0H>BASAL. Figura 47b) y la extinción (t(67) = -7,529, p<0,001,
EXT-0H>BASAL. Figura 47c); y una disminución significativa 1 h después de
finalizada la recuperación (t(67) = 8,634, p<0,001, REC-1H<BASAL. Figura 47b) y
la extinción (t(67) = 8,694, p<0,001, EXT-1H<BASAL. Figura 47c) de la memoria.
a.
-20
0
20
40
60
80
100
120
BASAL ADQ-0H ADQ-1H ADQ-4HCA
NT
IDA
D N
OR
MA
LIZ
AD
A (
%B
AS
AL
)
PP1
b.
-60
-40
-20
0
20
40
60
80
100
120
BASAL REC-0H REC-1H REC-4HCA
NT
IDA
D N
OR
MA
LIZ
AD
A (
%B
AS
AL
)
PP1
***
***
***
***

84
c.
-60
-40
-20
0
20
40
60
80
100
120
BASAL EXT-0H EXT-1H EXT-4H
CA
NT
IDA
D N
OR
MA
LIZ
AD
A (
%B
AS
AL
)
PP1
Figura 47. Cambios en el nivel de expresión de PP1 inducidos por el entrenamiento en una
tarea espacial. Diagrama de barras ilustrando los cambios en las FLU en animales entrenados
en la adquisición (a), recuperación (b) y extinción (c) de la memoria espacial con respecto al
basal. Se ilustra la media y el error estándar (n=5 animales por grupo). ***, p<0,001.
6.9. Efecto del estrés sobre la formación de memoria espacial y su
relación con la expresión de PP1
Finalmente al evaluar los efectos del estrés por inmovilización en las tres fases
de la formación de memoria y su relación con la actividad enzimática y el nivel
de expresión de PP1, se observó que con respecto al grupo basal y al grupo de
adquisición, el estrés, tanto corto como prolongado previo a la adquisición, no
modificó la actividad enzimática de PP1 (Figura 48a; 48b y 49).
***
***
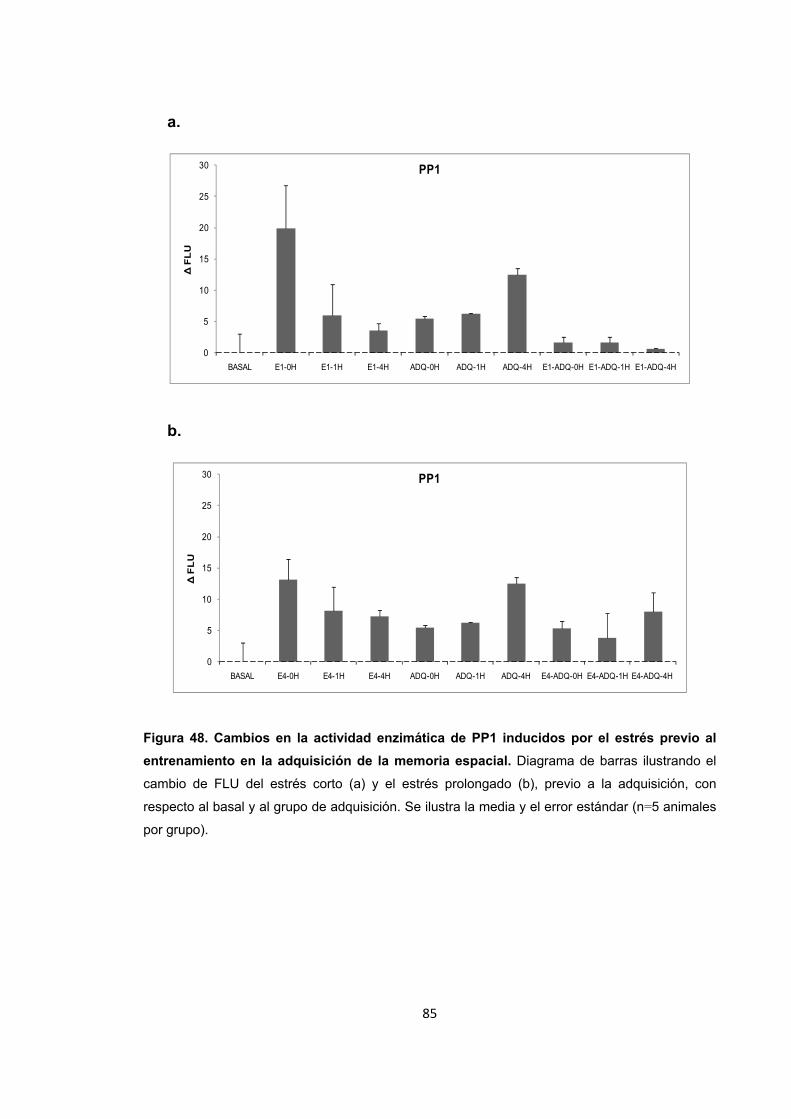
85
a.
0
5
10
15
20
25
30
BASAL E1-0H E1-1H E1-4H ADQ-0H ADQ-1H ADQ-4H E1-ADQ-0H E1-ADQ-1H E1-ADQ-4H
ΔF
LU
PP1
b.
0
5
10
15
20
25
30
BASAL E4-0H E4-1H E4-4H ADQ-0H ADQ-1H ADQ-4H E4-ADQ-0H E4-ADQ-1H E4-ADQ-4H
ΔF
LU
PP1
Figura 48. Cambios en la actividad enzimática de PP1 inducidos por el estrés previo al
entrenamiento en la adquisición de la memoria espacial. Diagrama de barras ilustrando el
cambio de FLU del estrés corto (a) y el estrés prolongado (b), previo a la adquisición, con
respecto al basal y al grupo de adquisición. Se ilustra la media y el error estándar (n=5 animales
por grupo).

86
0
5
10
15
20
25
30
-6 -4 -2 0 2 4 6
ΔF
LU
TRATAMIENTO
A
E4
E4 y A
E1
E1 y A
Figura 49. Cambios en la actividad enzimática de PP1 inducidos por el estrés previo a la
adquisición de la memoria. Se ilustran los cambios de FLU inducidos por el estrés corto (línea
azul tenue) y prolongado (línea verde) previo al entrenamiento de la adquisición de la memoria
espacial; también se ilustra la adquisición (línea azul oscura), el estrés corto (línea purpura) y el
estrés prolongado (línea roja); el eje “x” indica el tratamiento y el tiempo de sacrificio de los
animales y las barras indican el error estándar (n= 5 animales por grupo).
En cuanto al nivel de proteína total, se observó un aumento estadísticamente
significativo en el nivel de expresión de PP1 comparado con el basal,
inmediatamente y 1 h después de finalizado el estrés corto previo a la
adquisición (t(66) = -4,842, p<0,001, E1-ADQ-0H>BASAL; t(65) = -4,066, p<0,001,
E1-ADQ-1H>BASAL;) y 1 h después de finalizado el estrés prolongado previo a
la adquisición (t(67) = -4,562, p<0,001, E4-ADQ-1H>BASAL) (Figura 50a y 50b).

87
a.
-15
-10
-5
0
5
10
15
20
25
30
BASAL E1-ADQ-0H E1-ADQ-1H E1-ADQ-4H E4-ADQ-0H E4-ADQ-1H E4-ADQ-4HCA
NT
IDA
D N
OR
MA
LIZ
AD
A (
%B
AS
AL
)PP1
b.
(1) MW 1 2 3 4 5 6 7 8 MW 9 10 11 12 13 14 15 16
Figura 50. Cambios en el nivel de expresión de PP1 inducidos por el estrés corto y
prolongado previo al entrenamiento de la adquisición de la memoria espacial. (a).
Diagrama de barras ilustrando el cambio en el nivel de expresión de PP1 con respecto al basal
durante el estrés previo a la adquisición. Se ilustra la media y el error estándar (n=5 animales
por grupo). ***, p<0,001. (b). (1). SDS-PAGE; (2). WB. Carriles 1, 2, 9 y 10 muestras en
condiciones basales; carriles 3, 4,11 y 12 condiciones de estrés 1H-adquisición y sacrificio
inmediato; carriles 5, 6, 13 y 14 condiciones de estrés 1H-adquisición y sacrificio 1 H después y
carriles 7, 8, 15 y 16 condiciones de estrés 1H-adquisición y sacrificio 4H después. MW,
marcador de peso molecular (Estándar pre-teñido de amplio rango, BIO-RAD).
198,84 115,70 96,743 53,541 37,134
198,84 115,70 96,743 53,541 37,134
*** ******
KDa KDa
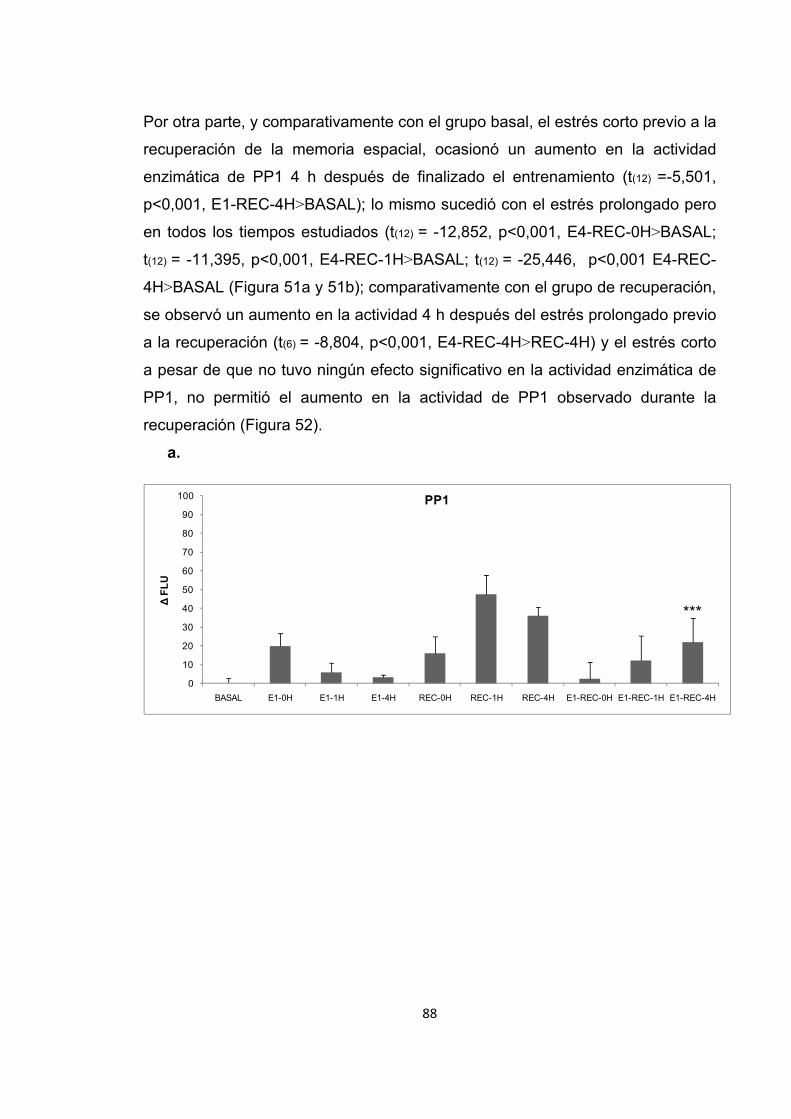
88
Por otra parte, y comparativamente con el grupo basal, el estrés corto previo a la
recuperación de la memoria espacial, ocasionó un aumento en la actividad
enzimática de PP1 4 h después de finalizado el entrenamiento (t(12) =-5,501,
p<0,001, E1-REC-4H>BASAL); lo mismo sucedió con el estrés prolongado pero
en todos los tiempos estudiados (t(12) = -12,852, p<0,001, E4-REC-0H>BASAL;
t(12) = -11,395, p<0,001, E4-REC-1H>BASAL; t(12) = -25,446, p<0,001 E4-REC-
4H>BASAL (Figura 51a y 51b); comparativamente con el grupo de recuperación,
se observó un aumento en la actividad 4 h después del estrés prolongado previo
a la recuperación (t(6) = -8,804, p<0,001, E4-REC-4H>REC-4H) y el estrés corto
a pesar de que no tuvo ningún efecto significativo en la actividad enzimática de
PP1, no permitió el aumento en la actividad de PP1 observado durante la
recuperación (Figura 52).
a.
0
10
20
30
40
50
60
70
80
90
100
BASAL E1-0H E1-1H E1-4H REC-0H REC-1H REC-4H E1-REC-0H E1-REC-1H E1-REC-4H
ΔF
LU
PP1
***
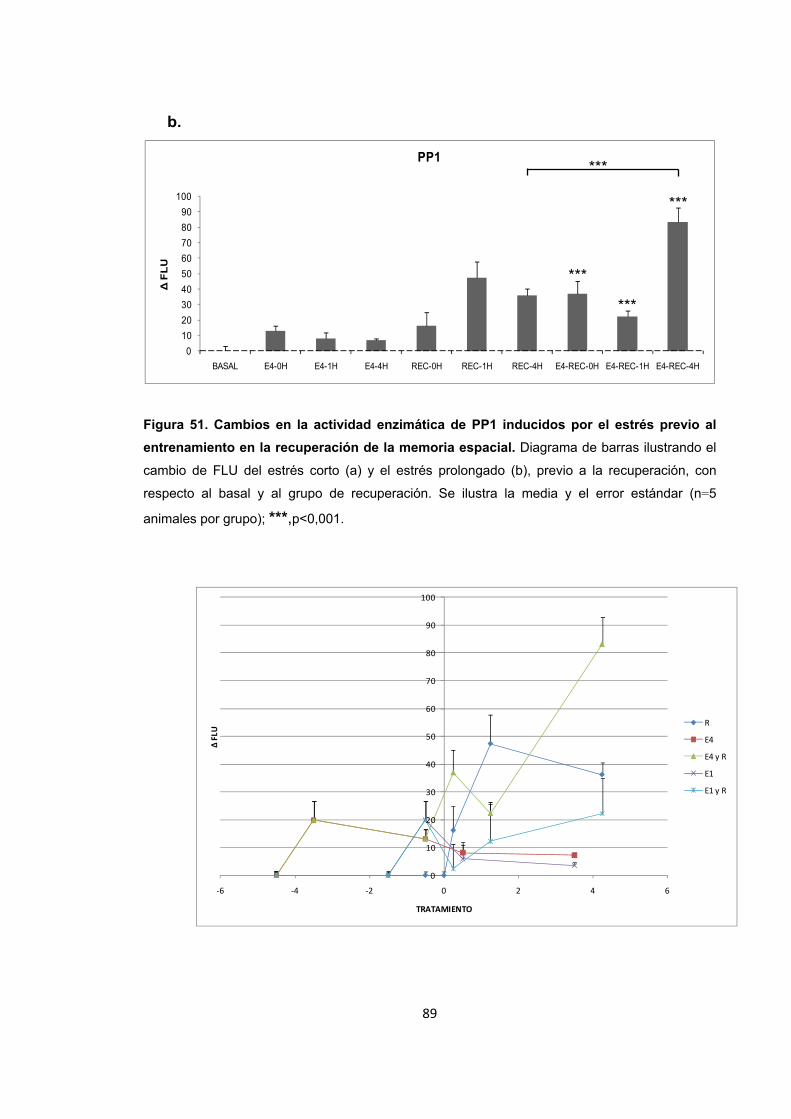
89
b.
0
10
20
30
40
50
60
70
80
90
100
BASAL E4-0H E4-1H E4-4H REC-0H REC-1H REC-4H E4-REC-0H E4-REC-1H E4-REC-4H
ΔF
LU
PP1
Figura 51. Cambios en la actividad enzimática de PP1 inducidos por el estrés previo al
entrenamiento en la recuperación de la memoria espacial. Diagrama de barras ilustrando el
cambio de FLU del estrés corto (a) y el estrés prolongado (b), previo a la recuperación, con
respecto al basal y al grupo de recuperación. Se ilustra la media y el error estándar (n=5
animales por grupo); ***,p<0,001.
0
10
20
30
40
50
60
70
80
90
100
‐6 ‐4 ‐2 0 2 4 6
ΔFLU
TRATAMIENTO
R
E4
E4 y R
E1
E1 y R
***
***
***
***
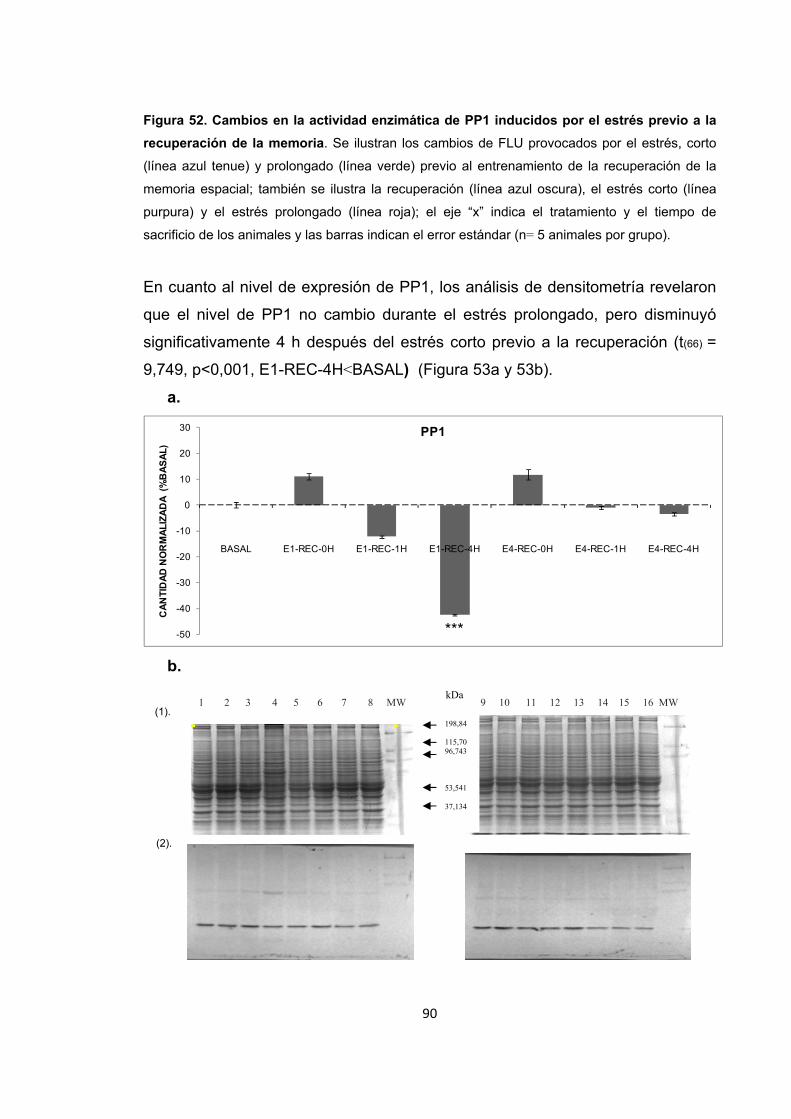
90
Figura 52. Cambios en la actividad enzimática de PP1 inducidos por el estrés previo a la
recuperación de la memoria. Se ilustran los cambios de FLU provocados por el estrés, corto
(línea azul tenue) y prolongado (línea verde) previo al entrenamiento de la recuperación de la
memoria espacial; también se ilustra la recuperación (línea azul oscura), el estrés corto (línea
purpura) y el estrés prolongado (línea roja); el eje “x” indica el tratamiento y el tiempo de
sacrificio de los animales y las barras indican el error estándar (n= 5 animales por grupo).
En cuanto al nivel de expresión de PP1, los análisis de densitometría revelaron
que el nivel de PP1 no cambio durante el estrés prolongado, pero disminuyó
significativamente 4 h después del estrés corto previo a la recuperación (t(66) =
9,749, p<0,001, E1-REC-4H<BASAL) (Figura 53a y 53b).
a.
-50
-40
-30
-20
-10
0
10
20
30
BASAL E1-REC-0H E1-REC-1H E1-REC-4H E4-REC-0H E4-REC-1H E4-REC-4H
CA
NT
IDA
D N
OR
MA
LIZ
AD
A (
%B
AS
AL
)
PP1
b.
1 2 3 4 5 6 7 8 MW 9 10 11 12 13 14 15 16 MW
198,84 115,70 96,743 53,541 37,134
***
(1).
(2).
kDa
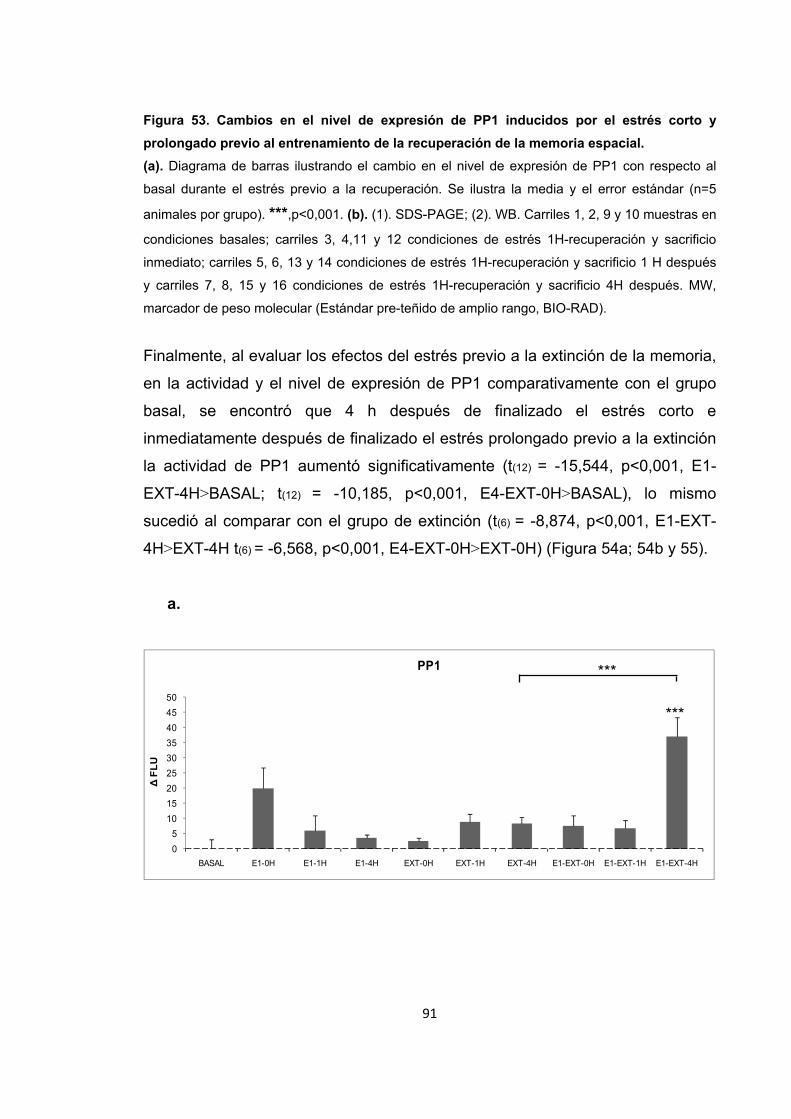
91
Figura 53. Cambios en el nivel de expresión de PP1 inducidos por el estrés corto y
prolongado previo al entrenamiento de la recuperación de la memoria espacial.
(a). Diagrama de barras ilustrando el cambio en el nivel de expresión de PP1 con respecto al
basal durante el estrés previo a la recuperación. Se ilustra la media y el error estándar (n=5
animales por grupo). ***,p<0,001. (b). (1). SDS-PAGE; (2). WB. Carriles 1, 2, 9 y 10 muestras en
condiciones basales; carriles 3, 4,11 y 12 condiciones de estrés 1H-recuperación y sacrificio
inmediato; carriles 5, 6, 13 y 14 condiciones de estrés 1H-recuperación y sacrificio 1 H después
y carriles 7, 8, 15 y 16 condiciones de estrés 1H-recuperación y sacrificio 4H después. MW,
marcador de peso molecular (Estándar pre-teñido de amplio rango, BIO-RAD).
Finalmente, al evaluar los efectos del estrés previo a la extinción de la memoria,
en la actividad y el nivel de expresión de PP1 comparativamente con el grupo
basal, se encontró que 4 h después de finalizado el estrés corto e
inmediatamente después de finalizado el estrés prolongado previo a la extinción
la actividad de PP1 aumentó significativamente (t(12) = -15,544, p<0,001, E1-
EXT-4H>BASAL; t(12) = -10,185, p<0,001, E4-EXT-0H>BASAL), lo mismo
sucedió al comparar con el grupo de extinción (t(6) = -8,874, p<0,001, E1-EXT-
4H>EXT-4H t(6) = -6,568, p<0,001, E4-EXT-0H>EXT-0H) (Figura 54a; 54b y 55).
a.
0
5
10
15
20
25
30
35
40
45
50
BASAL E1-0H E1-1H E1-4H EXT-0H EXT-1H EXT-4H E1-EXT-0H E1-EXT-1H E1-EXT-4H
ΔF
LU
PP1
***
***

92
b.
0
5
10
15
20
25
30
35
40
45
50
BASAL E4-0H E4-1H E4-4H EXT-0H EXT-1H EXT-4H E4-EXT-0H E4-EXT-1H E4-EXT-4H
ΔF
LU
PP1
Figura 54. Cambios en la actividad enzimática de PP1 inducidos por el estrés previo al
entrenamiento en la extinción de la memoria espacial. Diagrama de barras ilustrando el
cambio en la FLU del estrés corto (a) y prolongado (b) previo al entrenamiento en la extinción de
la memoria espacial con respecto al grupo basal y al grupo de entrenamiento en la extinción. Se
ilustra la media y el error estándar (n= 5 animales por grupo). ***,p<0,001.
0
5
10
15
20
25
30
35
40
45
50
-6 -4 -2 0 2 4 6
ΔF
LU
TRATAMIENTO
E
E4
E4 y E
E1
E1 y E
Figura 55. Cambios en la actividad enzimática inducidos por el estrés previo a la extinción
de la memoria. Se ilustran los cambios de FLU provocados por el estrés, corto (línea azul
***
***

93
tenue) y prolongado (línea verde), previo al entrenamiento de la extinción de la memoria
espacial; también se ilustra la extinción (línea azul oscura), el estrés corto (línea purpura) y el
estrés prolongado (línea roja); el eje “x” indica el tratamiento y el tiempo de sacrificio de los
animales, las barras indican el error estándar (n= 5 animales por grupo).
Así mismo, a nivel de la expresión de PP1, se encontró que comparativamente
con el grupo basal, el nivel de PP1 aumentó inmediatamente y 1 h después de
finalizado el estrés corto previo a la extinción (t(66) = -12,130, p<0,001, E1-EXT-
0H>BASAL; t(66) = -9,229, p<0,001, E1-EXT-1H>BASAL), y retornó a los niveles
basales 4 h después de finalizado todo el entrenamiento; el nivel de PP1 se
mantuvo cercano al basal con el estrés prolongado, pero disminuyo
drásticamente 4 h después del estrés prolongado previo a la extinción (t(66) =
6,201, p<0,001, E4-EXT-4H<BASAL) (Figura 56; WB 1 y 2).
-40
-30
-20
-10
0
10
20
30
40
50
60
BASAL E1-EXT-0H E1-EXT-1H E1-EXT-4H E4-EXT-0H E4-EXT-1H E4-EXT-4H
CA
NT
IDA
D N
OR
MA
LIZ
AD
A (
%B
AS
AL
)
PP1
Figura 56. Cambios en el nivel de expresión de PP1 inducidos por el estrés corto y
prolongado previo al entrenamiento en la extinción de la memoria espacial. Diagrama de
barras ilustrando el cambio en el nivel de expresión de PP1 con respecto al basal. Se ilustra la
media y el error estándar (n=5 animales por grupo). ***,p<0,001.
***
***
***
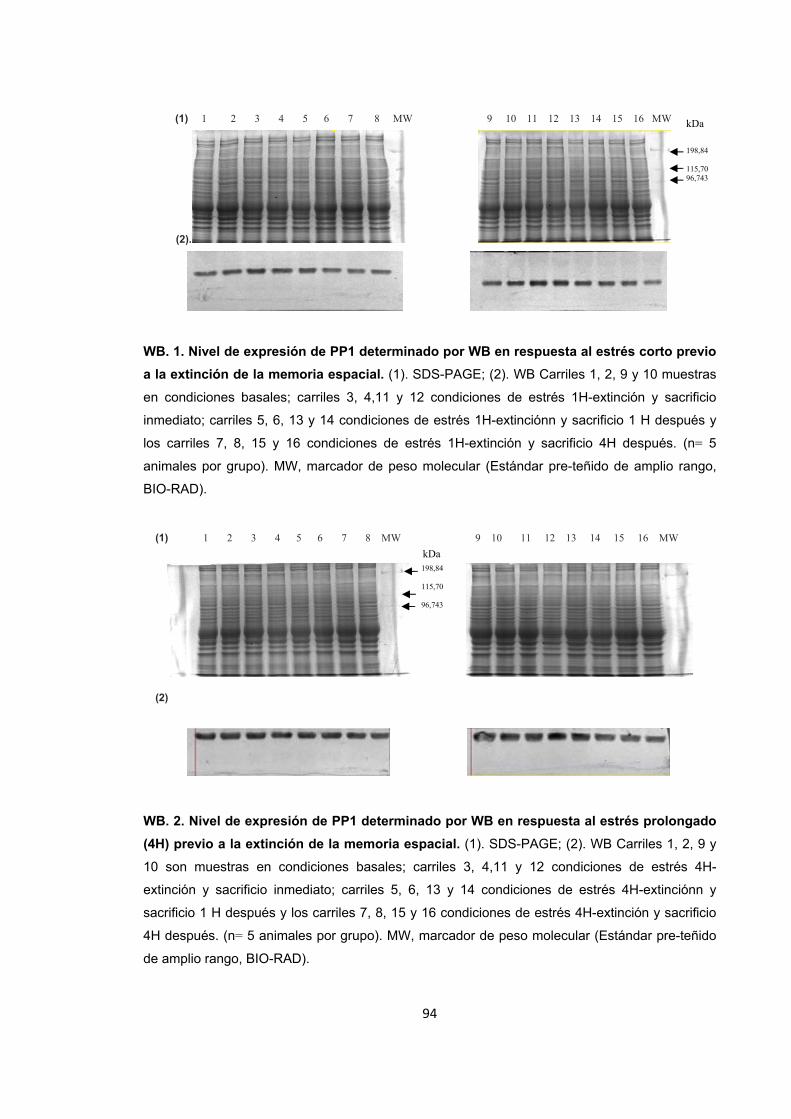
94
(1) 1 2 3 4 5 6 7 8 MW 9 10 11 12 13 14 15 16 MW
(2).
WB. 1. Nivel de expresión de PP1 determinado por WB en respuesta al estrés corto previo
a la extinción de la memoria espacial. (1). SDS-PAGE; (2). WB Carriles 1, 2, 9 y 10 muestras
en condiciones basales; carriles 3, 4,11 y 12 condiciones de estrés 1H-extinción y sacrificio
inmediato; carriles 5, 6, 13 y 14 condiciones de estrés 1H-extinciónn y sacrificio 1 H después y
los carriles 7, 8, 15 y 16 condiciones de estrés 1H-extinción y sacrificio 4H después. (n= 5
animales por grupo). MW, marcador de peso molecular (Estándar pre-teñido de amplio rango,
BIO-RAD).
(1) 1 2 3 4 5 6 7 8 MW 9 10 11 12 13 14 15 16 MW
(2)
WB. 2. Nivel de expresión de PP1 determinado por WB en respuesta al estrés prolongado
(4H) previo a la extinción de la memoria espacial. (1). SDS-PAGE; (2). WB Carriles 1, 2, 9 y
10 son muestras en condiciones basales; carriles 3, 4,11 y 12 condiciones de estrés 4H-
extinción y sacrificio inmediato; carriles 5, 6, 13 y 14 condiciones de estrés 4H-extinciónn y
sacrificio 1 H después y los carriles 7, 8, 15 y 16 condiciones de estrés 4H-extinción y sacrificio
4H después. (n= 5 animales por grupo). MW, marcador de peso molecular (Estándar pre-teñido
de amplio rango, BIO-RAD).
198,84 115,70 96,743
198,84 115,70 96,743
kDa
kDa

95
Se observó un comportamiento muy variable en la actividad enzimática de
ERK1/2, PP1 y PP2B, en respuesta a cada uno de los tratamientos (Figuras en
anexo 10, A10), también se observó, mediante un análisis de varianza,
diferencias significativas entre los grupos experimentales estudiados para cada
una de las enzimas (A11)

96
7. DISCUSIÓN
En este estudio, se encontró que la actividad y el nivel de expresión de las
enzimas estudiadas en el hipocampo tuvieron profundas modificaciones
provocadas por el estrés por inmovilización, el entrenamiento de una tarea de
memoria espacial y por la interacción de los dos anteriores. A continuación se
realiza una descripción detallada del comportamiento de cada una de las
enzimas en respuesta a los diferentes tratamientos.
7.1. Efecto del estrés por inmovilización sobre el nivel de
expresión y la actividad de ERK1/2, PP1 Y PP2B
Al observar en detalle el comportamiento de cada una de las enzimas en
respuesta al estrés se pudo establecer que:
1) El estrés por inmovilización, independientemente de su duración, no
causó modificaciones significativas en el nivel de expresión de ERK1 y
ERK2 en el hipocampo de ratas; sin embargo, el estrés agudo
prolongado, pero no el de corta duración, provocó un aumento
significativo en la actividad de ERK1/2. Este incremento fue mayor
inmediatamente después de finalizado el estrés y se redujo
progresivamente en las siguientes 4 h, pero continuó siendo
significativo.
2) El estrés por inmovilización tuvo un mayor efecto en la actividad
enzimática de PP2B, el estrés agudo, tanto corto como prolongado,
ocasionó un aumento en la actividad enzimática de PP2B, pero
únicamente 4 h después del estrés corto, e inmediatamente después
del estrés prolongado permaneciendo significativamente alto 1 y 4 h
después de finalizado el estrés.
3) Tanto el nivel de expresión como la actividad enzimática de PP1, se
modificaron por el estrés por inmovilización; se observó que el nivel de

97
expresión de PP1 aumentó inmediatamente finalizado el estrés corto,
se redujo progresivamente a la hora, y alcanzó el nivel de expresión
inicial a las 4 h; con el estrés agudo prolongado, se observó un pico
máximo de expresión de PP1 inmediatamente después de finalizado
el estrés y una reducción estadísticamente significativa a la hora y a
las 4 h; en relación a la actividad enzimática de PP1, esta tuvo dos
picos máximos de actividad, tanto con el estrés agudo de corta
duración como con el prolongado, pero únicamente inmediatamente
después de finalizado el estrés.
Trabajos previos han documentado la activación de estas enzimas en respuesta
al estrés; Meller y col. (2003) demostraron que un estrés agudo por 30 min
incrementaba el nivel de ERK1 y ERK2 en su forma fosforilada, inmediatamente
terminaba el estrés, pero un estrés crónico repetitivo no causaba ningún efecto;
así mismo Shimizu y col. (2004) mostraron que después de 2 h de restricción, el
nivel de ERK1 y ERK2 fosforilado incrementaba drásticamente y permanecía
alto 24 h después; de la misma manera, el estrés por inmovilización por 2 h
durante días consecutivos causaba un incremento en la fosforilación de ERK1 y
ERK2 en el hipocampo de ratas 1 h después de finalizado el estrés (Lee y col.,
2006); consistente con nuestros resultados, no observaron un aumento en el
nivel de expresión de ERK1 y ERK2 total en respuesta a la restricción.
Por otra parte, Takahashi y col. (2001), al estudiar la influencia del estrés por
restricción sobre la expresión y actividad de PP2B en el cerebro de ratas,
encontraron que el nivel de expresión de PP2B no se alteraba, pero su actividad
enzimática aumentaba en respuesta a un estrés por restricción de 45 ó 90 min;
resultados similares obtuvo el grupo de Morinobu y col. (2003). Sin embargo, no
se ha logrado definir con claridad y precisión los mecanismos que expliquen el
comportamiento de esta enzima en respuesta al estrés, pero es posible que el
aumento en la actividad de PP2B se deba a un aumento en los niveles de calcio
provocados por este tipo de estrés; varios estudios en ratones han demostrado
que los niveles de calcio aumentan en respuesta a un estrés por restricción

98
(Satoh y Shimeki, 2010; Wen-feng y col., 2008). El comportamiento de PP1 en
respuesta al estrés, y específicamente a un estrés por restricción no ha sido
definido, pero no se descarta que esta enzima pueda tener un papel importante
en los eventos moleculares relacionados con el estrés, debido a que se ha
descrito una función neuroprotectora de PP1 en un modelo de isquemia cerebral
de ratón adulto (Hédou y col., 2008).
De esta manera el estrés por inmovilización ocasionó principalmente un
aumento en la actividad enzimática de las enzimas estudiadas; siendo ERK1/2 y
PP2B principalmente modificadas con el estrés prolongado.
De acuerdo con los resultados obtenidos en este trabajo y teniendo en cuenta
los resultados de otros grupos de investigación, se puede concluir que el estrés,
dependiendo de su duración y repetición, desencadena la activación diferencial
de estas enzimas en el hipocampo de ratas. Así mismo el protocolo de estrés
empleado, la duración y el tiempo transcurrido desde la finalización del estrés,
son determinantes claves en su activación.
Para tratar de discernir el mecanismo por el cual estas enzimas están siendo
activadas en respuesta al estrés, es necesario hacer un análisis detallado de las
hormonas y neurotransmisores liberados en respuesta al estrés y establecer su
relación con la activación de estas enzimas. Trabajos de investigación dirigidos
a evaluar el papel de estas hormonas y neurotransmisores en la expresión de
ciertas proteínas en el cerebro, han dado a conocer diferentes aproximaciones
que nos permiten entender como es la cascada de señalización de estas
enzimas en respuesta al estrés (Figura 57). Qiu y col. (2001), reportaron que la
corticosterona puede inducir rápidamente la fosforilación y activación de ERK1 y
ERK2 e inducir su traslocación al núcleo sin cambiar el nivel de ERK1 y ERK2
total. Estos autores postularon también que PKC está corriente arriba de ERK1 y
ERK2 y que su fosforilación, inducida también por corticosterona, conducía a un
aumento en su actividad a través de c-Raf-1 (Qiu y col., 1998). En ratones
sometidos a estrés por restricción se observó que ERK1 y ERK2 se activaron en

99
extractos de hipocampo, pero únicamente 60-120 min después del inicio del
estrés, mientras que Raf-1 que esta corriente arriba de ERK aumento su
actividad 30 min después del inicio del estrés (Revest y col., 2005). Un
comportamiento similar se describió en líneas celulares, células AtT20,
observándose un aumento de ERK1 y ERK2 fosforilada 1 y 3 h después del
tratamiento con corticosterona (Tasker, 2006).
Por otra parte, el factor liberador de corticotropina (CRF, de sus siglas en
inglés), juega un papel importante en el control de la función del eje hipotálamo-
pituitaria-adrenal, y controla la respuesta endocrina frente a eventos estresantes
a través de receptores acoplados a proteína G (Gs, Gq/G11). Se ha
documentado que ERK1 y ERK2 es activada por CRF a través del receptor 1
(CRF-1) en el hipocampo, esta activación es dependiente de la actividad de
PKA, la cual media la fosforilación de ERK1 y ERK2 mediante la activación
secuencial de Rap1 y de B-Raf, moléculas claves en la activación de ERK1/2
por señales dependientes de AMPc (Arzt y Holsboer, 2006 y Elliott-Hunt y col.,
2002).
La norepinefrina, también induce la fosforilación y activación de ERK1 y ERK2.
Esto se demostró en un estudio en cultivos de neuronas primarias de la corteza
cerebral. La norepinefrina, se une a receptores α y β adrenérgicos acoplados a
proteína G (Tolbert, 2003).
La vasopresina, es un regulador importante de la secreción de ACTH. Su
expresión y secreción dentro de la circulación portal hipofisaria, así como el
número de receptores, se incrementan durante la fase tardía de un estrés agudo
y con el estrés crónico (Aguilera y Rabadan-Diehl 2000). Una de las posibles
rutas de señalización de este neuropéptido en respuesta al estrés, se describió
en una línea celular hipotalámica (H32), en esta línea celular se demostró que
este neuropéptido ejerce su efecto a través de la activación del receptor para la
vasopresina tipo 1 (V1bR) (Volpi y col., 2006); estos autores plantearon el
siguiente modelo: la liberación de vasopresina a la circulación portal hipofisaria

100
inducida por el estrés, conlleva a la activación del la señalización a través de
V1Br, este es un receptor acoplado a proteína G (Gαq/11), que ejerce su acción a
través de la activación de fosfolipasa C, intermediarios de esta ruta así como
inositol trifosfato (IP3) y el aumento en los niveles de calcio citosólicos inducen
la activación del receptor del factor de crecimiento epidermal (EGFR), el cual a
su vez induce la activación de la ruta MAPK (ERK1/2) a través de Ras (Fuentes
y col., 2008; Chen & Aguilera, 2010). En el hipocampo, el receptor para la
vasopresina es el AVPR1B, el cual pertenece al grupo de los receptores
acoplados a proteína G (Gs, Gq/11) y ejerce su señalización a través de
fosfolipasa C (Roper y col., 2010).
Finalmente, Los receptores de mineralocorticoides (MR), que se encuentran
predominantemente en el hipocampo, presentan de 4 a 10 veces más afinidad
por la corticosterona, que los receptores de glucocorticoides (GR) y por lo tanto
son ocupados en su mayoría cuando hay niveles bajos de corticosterona. En un
trabajo realizado por Olijslagers y col. (2008), se llego a la conclusión que la
corticosterona, se une a MR localizados en membranas postsinápticas de
neuronas piramidales en el área CA1 del hipocampo y activa la ruta ERK1/2 a
través de la activación de receptores acoplados a proteínas G.
Figura 57. Rutas de señalización activadas en respuesta al estrés. Se ilustran las posibles
rutas de señalización que se activan en respuesta al estrés y conducen a la activación de
ERK1/2. CRF: factor liberador de corticotropina; AC: adenilato ciclasa; cAMP: adenosina

101
monofosfato ciclico; PKA: proteína quinasa A; AVP: arginina vasopresina;V1Br: receptor 1B de
la vasopresina; PLC: fosfolipasa C; DAG: diacilglicerol; IP3:inositol trifosfato; PKC: proteína
quinasa C; MR: receptor de mineralocorticoides; Rap-1:proteína 1 relacionada a Ras;BRaf y
cRAS: proteínas serina treonina quinasas; ERK1/2: quinasas 1 y 2 reguladas por señales
extracelulares. (Tomado y modificado de Michael J. Shipstm (1995). TEM.6; 8).
En cuanto a la activación de fosfatasas en respuesta al estrés y los mecanismos
moleculares que subyacen a este evento son muy poco conocidos y la literatura
al respecto es escasa, razón por la cual nuestro trabajo sienta las bases para
futuras investigaciones al respecto.
7.2. Efecto del aprendizaje de una tarea de memoria espacial en el
nivel de expresión y la actividad de ERK1/2, PP1 Y PP2B
Durante el proceso de formación de la memoria espacial, también se observaron
modificaciones en la actividad enzimática de ERK1/2, PP1 y PP2B, en el
hipocampo de las ratas. En términos generales, la actividad de estas enzimas
aumentó durante las tres fases de la formación de memoria.
1) La actividad enzimática de ERK1/2 aumentó de manera significativa
comparativamente con el grupo basal, con el entrenamiento en la
adquisición, la recuperación y la extinción de la memoria espacial en los
tres tiempos estudiados, siendo muy similar en la adquisición y la
recuperación, y menor en la extinción de la memoria espacial. En cuanto
al nivel de expresión, se observó un aumento significativo en la
recuperación y la extinción de la memoria espacial, principalmente 4 h
después del entrenamiento.
2) La actividad enzimática de PP2B aumentó considerablemente durante las
tres fases de la formación de la memoria; esta actividad, fue equivalente
inmediatamente y 4 h después de haber finalizado el entrenamiento en la
adquisición y la extinción de la memoria, y similar en las tres fases 4 h
después de finalizado el entrenamiento. También se observó un pico
máximo de actividad inmediatamente después de haberse finalizado el

102
entrenamiento en la recuperación de la memoria espacial. Se observó
principalmente un aumento en el nivel de expresión de PP2B con el
entrenamiento en la recuperación de la memoria espacial 1 y 4 h después
de finalizado el entrenamiento.
3) La actividad de PP1 no cambio durante el entrenamiento en la adquisición
y la extinción de la memoria espacial, incluso su actividad fue muy similar
en estas dos fases, pero si lo hizo con el entrenamiento en la
recuperación de la memoria en todos los tiempos estudiados,
observándose una mayor actividad 1 y 4 h después del entrenamiento.
Sin embargo, los niveles de proteína total si cambiaron con el
entrenamiento en la adquisición, la recuperación y la extinción,
observándose un aumento en el nivel de PP1 1 y 4 h después de
finalizada la adquisición e inmediatamente después de finalizada la
recuperación y la extinción de la memoria espacial, y una disminución
significativa 1 h después de finalizadas estas dos fases.
Estas enzimas son expresadas en abundancia en neuronas del sistema
nervioso central maduro y en diferentes aéreas en el cerebro, específicamente
en neocorteza, hipocampo, amígdala y cerebelo, que son áreas involucradas en
procesos de aprendizaje y memoria (Atkins y col., 1998; Fiore y col., 1993; Flood
y col., 1998). El papel de ERK1/2 en la formación de memoria se ha
documentado en diferentes trabajos. La administración de un inhibidor
especifico de MEK (SL327), que impide la fosforilación de ERK1/2, deteriora el
aprendizaje contextual y el aprendizaje espacial (Selcher y col., 1999).
Adicionalmente, otro inhibidor de MEK, PD098059, bloquea la memoria espacial
a largo plazo en ratas entrenadas en el laberinto acuático de Morris (Blum y col.,
1999). ERK1 y ERK2 presenta un 84% de homología y comparten muchas
funciones diversas, como la regulación de la formación de memoria; estas son
activadas en respuesta a muchas señales, incluyendo receptores de factores de
crecimiento, integrinas, Src, Fyn y receptores acoplados a proteína G, así como
también, mediante la activación de receptores NMDA de glutamato (NMDAR) y
la estimulación eléctrica, los cuales rápidamente inducen la activación de ERK

103
(Ramos, 2008). Los NMDAR contribuyen a la adquisición de nuevas tareas de
memoria a través de la activación de ERK1/2 (Haddad, 2005). De esta manera,
ERK1/2 puede estar siendo activada por diferentes rutas de señalización en el
hipocampo, uno de los posibles modelos es, el aumento en los niveles de calcio
citosólico inducidos por la activación de los NMDAR en respuesta al
entrenamiento en la memoria espacial y a la activación de receptores acoplados
a proteína G (GPCR) (Figura 58); la activación de GPCR conduce a un aumento
a la actividad enzimática de PKA a través de un aumento de AMPc, PKA
fosforila los NMDAR y directa e indirectamente activa MEK1/2 promoviendo la
activación de ERK1/2. Otra posible ruta es a través de la activación de PKC, la
cual también es activada durante los procesos de formación de memoria y
puede inducir la fosforilación de c-Raf y activar de esta manera MEK1/2 y
ERK1/2; PKA también lo hace directamente a través de la fosforilación directa
de Raf-1 (Haddad, 2005). En cuanto al nivel de expresión, solo se observó un
aumento 4 h después del entrenamiento, esto es consistente con el tiempo en el
que transcurre la síntesis proteína.
Figura 58. Activación de ERK1/2 en el hipocampo durante los procesos de aprendizaje y
formación de memoria. Se ilustra la ruta de señalización activada durante los procesos de
aprendizaje con la consecuente activación de ERK1/2. Ca2+: calcio; GPCR: receptores
acoplados a proteína G; Gsα: subunidad alfa de la proteína G; PD-98059: Inhibidor de MEK
(Tomado y modificado de: John J. Haddad; Progress in Neurobiology 77 (2005) 252–282).

104
Un aspecto muy importante que queda por investigar son los efectores corriente
abajo de ERK1/2 que pueden contribuir con los procesos de aprendizaje y
memoria, tales como proteínas de vesículas sinápticas y de citoesqueleto,
canales iónicos y factores de transcripción como Elk-1.
Por otra parte, la PP2B a diferencia de otras fosfatasas, tiene una estrecha
especificidad de sustrato, siendo la fosfoproteína neuronal regulada por AMPc y
dompamina, inhibidor 1, factor nuclear de celulas T activadas (NFAT), NMDAR y
el inositol trifosfato, de sus siglas en ingles DARP32, I-1, NFAT, NMDAR e IP3
respectivamente, los principales blancos de esta enzima. Tiene una distribución
principalmente citoplasmática y en ausencia de algún estímulo se encuentra
principalmente inactiva (Rusnak y Mertz, 2000). Mediante el uso de FK506, un
inhibidor especifico de PP2B, se demostró que está involucrada en los procesos
de potenciación a largo plazo (LTP, modelo sináptico del aprendizaje y la
memoria) en el hipocampo y en la regulación de la actividad de los NMDAR (Lu,
1996). Pequeños aumentos en la concentración de calcio intracelular, inducidos
por estimulaciones de baja frecuencia, son suficientes para activar PP2B a
través de los NMDAR (Munton y col., 2004). Por lo tanto, existe una relación
estrecha entre los cambios en la plasticidad sináptica asociados con los
procesos de formación de memoria y la actividad enzimática de PP2B, que se
reflejan en el aumento en la actividad enzimática observado durante la
adquisición, la recuperación y la extinción de la memoria espacial. Por otra
parte, Mansuy y col. (1998), encontraron que ratones que expresaban una forma
truncada de la PP2B presentaban un defecto específico en la memoria espacial
en el laberinto de Barnes; sin embargo, la sobreexpresión de una forma normal
de PP2B, revertía dicho defecto en la memoria. Así mismo, Zeng y col. (2001)
mostraron que ratones Knock-out para PP2B tenían dificultades de aprendizaje
en el laberinto acuático de Morris y en el reaprendizaje de una nueva
localización de la plataforma. También se ha reportado que PP2B es una
enzima importante en la adaptación a cambios en un contexto conocido, en
línea con esta hipótesis, Runyan y col. (2005), mostraron que la inhibición
farmacológica de PP2B en la corteza pre-frontal de ratas, impedía el aprendizaje

105
de una nueva localización de la plataforma en el laberinto acuático de Morris. Lin
y col. (2003), mostraron que el bloqueo de la actividad de PP2B en la amígdala
basolateral durante la extinción del entrenamiento en un paradigma de
condicionamiento al miedo, no permitía la extinción de memoria al miedo
previamente formada. Por último, Havekes y col. (2006), mostraron que el nivel
de PP2B disminuía durante el entrenamiento en el laberinto en Y, pero
retornaba a niveles basales cuando se revertía la tarea, indicando que está
fosfatasa es esencial para adaptarse a cambios en un mismo contexto. Sin
embargo, otras publicaciones reportan un efecto negativo de PP2B en la
regulación de los procesos de formación de memoria (Malleret y col., 2001;
Dineley y col., 2007; Mansuy, 2003). Teniendo en cuenta estos resultados
encontrados y los obtenidos en este estudio, es importante recalcar el aumento
en la actividad enzimática de PP2B observado durante la extinción de la
memoria, se sabe que en esta fase, se establece una nueva memoria y un
aumento en la variabilidad en el repertorio comportamental de un organismo,
que le permite adaptarse a una nueva tarea en un mismo contexto.
En cuanto a PP1, esta puede unirse a más de 60 proteínas reguladoras y de
anclaje a través de su subunidad catalítica y se localiza en diferentes
compartimentos sub-celulares específicos (Mansuy y Shenolikar, 2006), esto
hace de PP1 una proteína muy compleja de estudiar con un amplio espectro de
funciones, ejerce un papel importante en la regulación del ciclo celular, la
transducción de señales, el metabolismo del glicógeno, la contracción muscular,
la apoptosis y la función neural, así como la plasticidad sináptica, la liberación
de neurotransmisores y el crecimiento de neuritas (Munton y col., 2004) y
además, tiene efecto neuroprotector (Hédou y col, 2008). La PP1 citoplasmática
es conocida como un regulador negativo universal de la memoria y la plasticidad
sináptica. En el cerebro, el pool de PP1 presente en las terminales sinápticas,
regula negativamente la señalización neuronal y la eficiencia sináptica, actuando
como un limitante molecular del aprendizaje y como un supresor de la memoria;
por otra parte, la PP1 nuclear es un regulador crítico de la remodelación de la
cromatina en el cerebro de mamíferos, que controla las modificaciones post-

106
traduccionales de las histonas y la transcripción de genes asociados con la
memoria a largo plazo, tales como elementos de unión de respuesta al AMPc
(CREB) y el factor nuclear kappa-B (NFκB) (Koshibu y col., 2009). Esto es
fundamentado en trabajos que concluyen que la inhibición de la PP1 presente
en el núcleo de neuronas en la amígdala altera la fosforilación, acetilación y
metilación de residuos específicos sobre varias proteínas histonas e incrementa
la memoria al miedo a largo plazo y la LTP (Morishita y col., 2001; Jouvenceau y
col., 2006). Genoux y col., 2002 y Graff y col., 2010 reportaron que la PP1
nuclear tiene un papel importante en el control de los eventos transcripcionales
críticos para la formación de memoria y la plasticidad sináptica a largo plazo. En
efecto, este grupo encontró que la inhibición de la PP1 nuclear estaba asociada
con una regulación positiva o negativa de genes involucrados en diferentes
funciones celulares. Más aún, la inhibición de la PP1 nuclear facilitó la memoria
espacial dependiente de hipocampo. Esto tiene un papel importante en la
regulación epigenética de la memoria al miedo en la amígdala (Koshibu y col.,
2011).
La administración de inhibidores de fosfatasas (ácido ocadáico, caliculina A,
ciclosporina A) tiene efectos deletéreos sobre varias formas de memoria, lo que
sugiere que PP1, PP2A y PP2B podrían actuar como facilitadores del
aprendizaje (Vianna y col., 2010; Zhao y col., 1995, Bennett y col., 1996, 2001,
2003; He y col., 2001).
De acuerdo con lo mencionado anteriormente, es posible que el aumento en la
actividad de PP1 y PP2B observado se deba a: durante el aprendizaje se
activarían los NMDAR, la entrada de calcio a través de estos receptores
provocaría un aumento en la actividad de PP2B, lo que llevaría a la des-
fosforilación de DARPP32 (G-substrate and dopamine and cAMP-regulated
phosphoprotein Mr 32000) (Halpain y col, 1990), esto conllevaría a una
desinhibición de la PP1, asimismo, la PP2B también puede defosforilar el
inhibidor 1 de PP1 (I1) dejando a PP1 activa.

107
Por otra parte, también es posible una interacción entre las quinasas y las
fosfatasas en términos de la regulación de su actividad. En cardimiocitos, la
activación de los receptores opioides-κ (receptores acoplados a proteína G, que
también se encuentran en cerebro) activaría PKC y PKA. La activación de PKC
incrementaría la actividad de PP1 a través de una interacción directa o a través
de la inhibición de PKA. La activación de PKA fosforilaría la neurabina I (la cual
es altamente expresada en las neuronas y regula el crecimiento de las neuritas).
Como la neurabina I sin fosforilar, se une directamente a la PP1 inhibiéndola, su
fosforilación por PKA disminuiría su interacción con PP1 y por lo tanto la dejaría
activa (Figura 59) (Mc AVOY y col., 1999).
Figura 59. Rutas de señalización que conduce a un cambio en la actividad de PP1 en el
hipocampo durante los procesos de aprendizaje y formación de memoria. Ilustración de las
rutas de señalización desencadenas por la activación de los receptores κ-opioides y la
subsecuente activación de PP1. (tomado y modificado de British Journal of Pharmacology, 1998,
124; 3: 600-6)
Los mecanismos de acción y de señalización de cada una de estas enzimas son
muy diversos y varían dependiendo de la fase y de la forma de aprendizaje y
memoria. De esta manera, cada fase de la formación de memoria recluta
diferentes estructuras anatómicas del cerebro y diferentes rutas de señalización
intracelular, las cuales son activadas diferencialmente durante las diferentes
fases temporales de la formación de memoria y en las distintas formas de

108
aprendizaje. Es importante tener en cuenta que no solo el tipo de memoria
influye en el patrón de expresión de proteínas en el cerebro, específicamente en
el hipocampo, sino que también influye el tipo de paradigma de aprendizaje
utilizado. En efecto, Zheng y col. (2009), reportaron que en el hipocampo los
cambios en la expresión de proteínas relacionadas con la memoria espacial
eran significativamente distintos en función de que el entrenamiento se hubiera
hecho en el laberinto de Barnes o en el laberinto en T múltiple.
7.3. Efecto de la interacción del estrés agudo y los procesos de
aprendizaje en el nivel de expresión y la actividad de ERK1/2,
PP1 Y PP2B
Es necesario hacer una descripción de los resultados obtenidos a nivel
comportamental (Grupo de aprendizaje y comportamiento animal) para poder
establecer una relación directa de la interacción del estrés agudo previo a las
tres fases de la formación de la memoria.
7.4. Efectos comportamentales del estrés agudo inducido antes de
la adquisición de la memoria espacial.
Los animales estresados, independientemente de la duración de la restricción
de movimientos, adquirieron satisfactoriamente la tarea espacial. No obstante, la
adquisición de la tarea espacial fue más lenta en los animales estresados que
en los no estresados, comportamiento que se refleja en los siguientes
indicadores: latencia de escape [estrés 4 horas (E4h) > no estrés (NE), p <
0,05], errores de agujero [estrés 1 hora (E1h) > NE, p < 0,01; E4h > NE, p <
0,01], errores ponderados [E1h > NE, p < 0,001; E4h > NE, p < 0,001], distancia
media a meta [E4h > NE, p < 0,05] y número total de exploraciones [E1h > NE, p
< 0,001; E4h > NE, p < 0,001]. Como se infiere de los datos anteriores, el
enlentecimiento del cambio comportamental durante el entrenamiento fue mayor
en los sujetos sometidos a estrés durante 4 horas.

109
7.5. Efectos comportamentales del estrés agudo inducido antes de
la recuperación de la memoria espacial.
los sujetos estresados mostraron un comportamiento significativamente distinto
que el de los animales no estresados en los siguientes indicadores: latencia a
meta [E1h > NE, p < 0,001; E4h > NE, p < 0,001], exploración de agujeros
diferentes al agujero meta [E1h > NE, p < 0,001; E4h > NE, p < 0,01],
exploración ponderada de agujeros diferentes al agujero meta [E1h > NE, p <
0,001; E4h > NE, p < 0,001], distancia media a meta [E1h > NE, p < 0,01; E4h >
NE, p < 0,01] y exploraciones porcentuales del agujero meta [E1h < NE, p <
0,001; E4h < NE, p < 0,001], lo que indica que el estrés interfiere en la
recuperación de la memoria.
7.6. Efectos comportamentales del estrés agudo inducido antes de
la extinción de la memoria espacial.
Durante la extinción, los animales no estresados persistieron en explorar
preferencialmente el agujero meta hasta el sexto ensayo de extinción, esta
preferencia desapareció durante el séptimo ensayo de extinción y se recuperó,
si bien marginalmente, durante el octavo ensayo de extinción. En contraste, en
los animales estresados, la preferencia por el agujero meta desapareció a partir
del quinto ensayo de extinción y no hubo recuperación en ensayos sucesivos.
Los resultados indican que el estrés agudo deteriora la adquisición y
recuperación de la memoria espacial, mientras que facilita la extinción de la
misma. A mayor intensidad y duración del estrés, mayor es la magnitud de estos
efectos.
Finalmente, el estrés agudo inducido antes de fases específicas del
entrenamiento en una tarea espacial también ocasionó modificaciones en la
actividad y el nivel de expresión de las enzimas estudiadas, observándose un
comportamiento más variable tanto en la actividad como en el nivel de
expresión. Los resultados obtenidos indican:

110
1) El estrés corto inducido antes de la adquisición de la memoria espacial
sin importar su duración, no permitió el aumento en la actividad
enzimática de ERK1/2 observado durante esta fase de la memoria
espacial; comparativamente con el grupo basal, el estrés, tanto corto
como prolongado, inducido antes de la adquisición, ocasionó un
aumento en la actividad enzimática de ERK1/2; en cuanto al nivel de
expresión de ERK1/2 esté aumentó con el estrés corto inducido antes
de la adquisición de la memoria espacial.
2) Solo el estrés corto inducido antes de la adquisición de la memoria
espacial no permitió el aumento en la actividad enzimática de PP2B y
PP1 observado durante la adquisición, aunque esté solo fue
significativo para PP2B; comparativamente con el grupo basal, la
actividad de PP2B, aumentó solo en condiciones de estrés prolongado
inducido antes de la adquisición; en términos generales, la actividad
de PP1 no se modificó con el estrés inducido antes de la adquisición,
sin embargo, el nivel de expresión de PP1 aumentó inmediatamente y
1 h después de finalizado el entrenamiento con el estrés corto
inducido antes de la adquisición de la memoria y 1 h después de
finalizado el entrenamiento con el estrés prolongado inducido antes de
la adquisición.
3) El estrés agudo inducido antes de la recuperación de la memoria tuvo
un efecto más dramático en la actividad de ERK1/2. Sin importar su
duración, la inducción del estrés agudo antes de la recuperación, no
permitió el aumento de actividad de ERK1/2 observado durante la
recuperación (sin estrés). El estrés agudo de corta duración inducido
antes de la recuperación no permitió el aumento en PP2B y PP1; si
bien este efecto no fue estadísticamente significativo para PP1, debido
a que las desviaciones estándar fueron muy grandes. Por otra parte,
el estrés agudo prolongado inducido antes de la recuperación de la
memoria no alteró el aumento en la actividad enzimática de PP1 y
PP2B observado durante la recuperación. El nivel de expresión de

111
ERK1/2 aumentó con el estrés prolongado inducido antes de la
recuperación 1 y 4 h después de finalizado todo el entrenamiento,
mientras que el nivel de PP2B y PP1 disminuyó.
4) El estrés agudo inducido antes de la extinción de la memoria espacial,
no tuvo un efecto considerable en la actividad de ERK1/2 y PP2B
cuando se comparó con el grupo de extinción. Sin embargo, cuando
se comparó el grupo basal, la actividad enzimática de ERK1/2
aumentó con el estrés corto inducido antes de la extinción
inmediatamente y 4 h después de finalizado todo el entrenamiento, e
inmediatamente finalizó el estrés prolongado inducido antes de la
extinción. Por otra parte el estrés previo a la extinción tuvo un efecto
más notorio en el nivel de expresión de PP2B, observándose
principalmente un aumento.
5) Por ultimo, el estrés inducido antes de la extinción de la memoria, tuvo
más impacto sobre el nivel de expresión y la actividad enzimática de
PP1. Con respecto al grupo basal, se observó un incremento en el
nivel de la expresión de PP1 inmediatamente y 1 h después de
finalizado el estrés corto inducido antes de la extinción; con respecto
al grupo de extinción (sin estrés), se observó un aumento en la
actividad de PP1 4 h después de finalizado el estrés corto inducido
antes de la extinción e inmediatamente finalizo el estrés prolongado
inducido antes de la extinción; por lo tanto el estrés inducido antes de
la extinción de la memoria incremento tanto el nivel de expresión
como la actividad enzimática de PP1.
Existe importante evidencia que involucra a ERK1/2 como uno de los posibles
candidatos que puede estar mediando los efectos en el aprendizaje inducidos
por el estrés. Revest y col. (2005, 2010), describieron la primera cascada
molecular que soporta el incremento de la memoria relacionado con el estrés, la
cual involucra la activación del receptor de glucocorticoides (GR), el factor de

112
transcripción Egr-1 y finalmente ERK1/2 (GR/Egr-1/MAPK). Además reportaron
que la activación de GR inducida por estrés se asociaba con un incremento en
la expresión de Ras y Raf-1 30 min después del estrés, con un incremento
nuclear de ERK1/2 60 min después del estrés, y con un incremento de la forma
fosforilada de ERK1/2 120 min después del comienzo del estrés. Así mismo,
reportaron que en el hipocampo y corriente abajo de esta cascada, la sinapsina-
Ia/Ib (proteínas de las vesículas presinápticas que controlan la liberación de
neurotransmisores), es una de las principales proteínas efectoras que esta
siendo activada por esta ruta. Otro trabajo reportó, que el incremento de
corticosterona durante el estrés agudo, conducía a la activación de GR y a la
síntesis y liberación del factor neurotrófico derivado del cerebro, de sus siglas en
ingles BDNF, el cual actúa sobre el receptor TrkB, lo que conduce a la
activación secuencial de una serie de quinasas que incluye PKC, RAS/RAF-1,
MEK1/2 y finalmente ERK1/2, la cual en este trabajo perjudicaría la LTP y
promovería la inducción de la LTD (Yang Chih-Hao y col., 2004).
La modulación del aprendizaje y la memoria parece ser uno de los principales
papeles de CRF en roedores y humanos. Así, la inyección intra-cerebro-
ventricular de CRF antes o inmediatamente después del entrenamiento
incrementa la memoria en tareas de aprendizaje (pruebas de discriminación
visual en el laberinto en Y; tareas de evasión pasiva, y tareas de evasión activa,
aprendizaje de evasión inhibitorio (Radulovic y col., 1999). De otra parte, en un
estudio realizado con dos cepas de ratones BALBc y C57BL/6N (Blank y col.,
2003) para tratar de establecer las rutas de transducción de señales del CRF en
el hipocampo, se encontró que CRF incrementa la actividad neuronal en el área
CA1 del hipocampo a través de PKC en ratones BALBc y a través de PKA en
ratones C57BL/6N; no obstante, la administración de CRF solo mejora el
aprendizaje en ratones BALBc. Otro estudio, realizado por Sananbenesi y col.
(2003), demostró que la inmovilización por 1 h previo a un aprendizaje al miedo
dependiente del contexto incrementa la fosforilación de ERK1/2 30, 60 y 120
minutos después del entrenamiento, y que este efecto depende de Mek1/2 y lo
que conduce a un incrementó la persistencia de esta memoria. De esta manera,

113
como lo indican nuestros resultados, el estrés previo a la adquisición de la
memoria espacial no interfiere negativamente con el nivel de expresión y la
actividad de esta enzima. Además, en respuesta al estrés, hay varias rutas de
señalización que pueden estar manteniendo ERK1/2 activa, como las mediadas
por PKC, PKA, BDNF, Egr-1, o el AMPc. El incremento de AMPc asociado a la
activación de CRF1 y CRF2, de una manera independiente de PKA, regula el
factor intercambiador de nucleótidos de guanina y activa ERK1/2.
Por otra parte, el bloqueo del aumento de la actividad enzimática de ERK1/2 tras
la recuperación de la memoria espacial debido al estrés agudo corto, se ajusta a
lo obtenido por otros grupos de investigación, que concluyen que esta enzima
está relacionada con los procesos de memoria y que el estrés afecta
específicamente la recuperación. Han sido pocos los estudios que se han
enfocado en los efectos del estrés o de los glucocorticoides en la recuperación
de la memoria. Cai y col. (2006), reportaron que cuando los glucocorticoides son
administrados inmediatamente después de la recuperación de la memoria al
miedo contextual, esta era significativamente disminuida. Sin embargo, es difícil
tratar de establecer una cascada de señalización intracelular a través de la cual
se esté generando la disminución en la activación de ERK1/2 mediada por
glucocorticoides, dada la complejidad, el numero y la variabilidad de moléculas
que se activan en el cerebro en respuesta al estrés. Es posible que mediante la
activación de la vía dopaminérgica en respuesta al estrés, y específicamente la
activación del receptor D2, se disminuya la vía de señalización del AMPc y de la
proteína quinasa A (PKA) (cAMP/PKA), la cual es una de las vías de activación
de ERK1/2. También se reportó, aunque no es muy clara la vía de señalización
corriente arriba, que PKA puede inhibir la actividad de ERK1/2 a través de la
fosforilación del residuo ser43 en Raf-1, esto perjudicaría su interacción con Ras
y, corriente abajo detiene la cascada de señalización que finaliza con la
fosforilación de ERK1/2 (Roberson y col., 1999). Por otra parte, como vimos en
nuestros resultados, la actividad enzimática de ERK1/2 disminuyó tanto con el
estrés agudo de corta duración como con el estrés prolongado, pero la actividad
de PP2B solo disminuyó con el estrés de corta duración. Dado este resultado,

114
es probable que PP2B sea responsable de que no aumentara la actividad de
ERK1/2 cuando se indujo estrés agundo prolongado antes de la recuperación.
Como vimos, la actividad de PP2B aumentó con el estrés prolongado, con la
recuperación y con el estrés prolongado inducido antes de la recuperación.
Según la literatura, en respuesta al estrés hay una regulación dependiente de
glutamato de PP2B, el aumento en la activación de esta enzima conllevaría a la
des-fosforilación de Rap1GAP en los residuos ser-441 y ser-499, a la
inactivación de Rap1 y por lo tanto de ERK1/2 (McAvoya y col., 2009).
La extinción de la memoria es considerada una nueva forma de aprendizaje, y al
igual que otros tipos de aprendizaje, es regulada por: el aumento en la expresión
génica, la síntesis de proteínas, la activación de las quinasas PKA y ERK1/2 y la
activación de los NMDAR (Cammarota y col., 2004). La extinción también es
susceptible a los efectos del estrés, pero, a diferencia de las otras fases de la
memoria espacial, no se sabe con certeza cuales son los mecanismos
moleculares que subyacen a los efectos del estrés sobre esta fase. Sin
embargo, previas investigaciones le confieren un papel facilitador a los
glucocorticoides en la extinción de la memoria (Yang y col., 2006), así como
también un papel importante a los NMDAR, y a numerosas cascadas de
señalización, como las MAPK, fosfatidilinositol 3-quinasa (PI3K) y PP2B y genes
tempranos inmediatos como zif268 y c-fos (Yang y Lu, 2005). Los
glucocorticoides alteran la conductancia y la expresión de los canales de calcio,
y a través de este mecanismo los glucocorticoides también podrian modificar la
extinción de la memoria (Yang, 2007). Por otra parte, el estrés, a través de la
activación de los GR, perjudica la absorción de glutamato en la región CA1 del
hipocampo, lo que prolonga la activación de los NMDAR y conlleva a la
activación diferencial de ciertas cascadas de señalización, dependiendo de la
estructura cerebral en cuestión y del tipo de aprendizaje (Yang y col., 2007).
A groso modo, la actividad enzimática de ERK1/2 y PP2B no cambio en
condiciones de estrés agudo previo a la extinción de la memoria espacial, sin
embargo la actividad de ERK1/2 aumento con respecto al grupo basal en

115
condiciones de estrés agudo corto y prolongado previo a la extinción, pero esta
no fue mayor en ninguno de los casos a la obtenida durante el entrenamiento de
la extinción, lo que indica que ERK1/2 y PP2B no se ven modificadas con el
estrés previo a la extinción y de acuerdo con los resultados comportamentales,
en donde se concluye que el estrés mejora la extinción de la memoria, es
posible que otras proteínas se estén activando específicamente en esta fase del
entrenamiento. De acuerdo con nuestros resultados PP1 tiene un papel
importante durante este proceso, sin embargo, no se sabe con certeza los
mecanismos por los cuales el estrés puede estar activando esta enzima y cual
es su papel durante la extinción de la memoria espacial.
Por otra parte, se sabe que los GR dentro del núcleo, dependiendo de la
concentración de corticosterona, pueden inducir tanto la transcripción génica, a
través de su unión secuencias especificas de DNA en las regiones enhancer del
promotor (GRE) de ciertos genes sensibles a esteroides y la acetilación de
histonas (H4) a través de la activación de proteínas co-activadoras como CREB-
binding protein (CBP), p300/CBP activating factor (PCAF) y steroid-receptor
coactivator (SRC-1) que tiene actividad de histona acetiltransferasa (HTA) ó
pueden reducir la transcripción génica, interfiriendo con la transactivación
mediada por co-activadores (CBP) a través de una interacción directa o a través
de alteraciones a nivel de la cromatina, esto se demostró en pacientes con asma
en donde bajas concentraciones de corticosterona puede inducir la activación de
histonas deacetilasas (HDAC2) (Figura 60) (Barnes y col., 2005; Ito y col.,2000);
este hallazgo explicaría el hecho de que encontremos una expresión diferencial
de las proteínas estudiadas, que sustenta los aumentos y disminuciones
observados.

116
Figura 60. Efecto de los glucocorticoides en la transcripción génica. Se ilustra el papel de la
corticosterona en los procesos de transcripción génica (Tomado de Barnes P.J, Adcock I.M, Ito
K; Eur Respir J. 25: 552-63).
Estos hallazgos indican que el estrés previo a un aprendizaje, interfiere con el
proceso de formación de nuevas memorias, a través de la activación diferencial
de rutas de señalización intracelular, quinasas y fosfatasas, que permite explicar
la relación entre los procesos celulares desencadenados por el estrés y los
procesos neurobiológicos en los que se basa la memoria.
No es posible formular un modelo universal que explique la relación entre los
procesos de aprendizaje y los mecanismos moleculares subyacentes. Sin
embargo lo comúnmente aceptado y más conocido es el balance entre las
proteínas quinasas y fosfatasas, que operan en el cerebro para controlar la
eficacia sináptica, el aprendizaje y la memoria, regulando positiva o
negativamente las rutas de transducción de señales (Lisman, 1989). Así mismo,
no podemos hablar de un modelo “lineal” en donde solo intervienen 2 moléculas
protagonistas, ya que existen otras moléculas y otros factores que ejercen un
papel determinante en estos procesos, además es importante tener en cuenta,
que durante los procesos de aprendizaje y formación de memoria la síntesis de
proteínas y la expresión de genes es inespecífica en cada fase del entrenamie

117
8. CONCLUSIONES
1. El estrés agudo provocó aumento de la actividad enzimática de todas
las enzimas evaluadas. La actividad de ERK1/2 presentó el aumento
más evidente y persistente, independientemente de la duración del
estrés agudo. La actividad de PP2B aumentó después del estrés,
siendo este fenómeno más temprano e intenso cuando el estrés
agudo se prolongo. La actividad de PP1 aumentó levemente
inmediatamente después de finalizado el estrés agudo,
independientemente de su duración.
2. La adquisición del aprendizaje espacial indujo un aumento de la
actividad enzimática de ERK1/2 y de PP2B, la cual se mantuvo
elevada hasta 4 h después de finalizado el entrenamiento. En
contraste, la adquisición solo provocó un aumento tardío del nivel de
expresión de PP1, pero no de su actividad enzimática.
3. La recuperación de la memoria espacial provocó un aumento de la
actividad enzimática de todas las enzimas evaluadas, este aumento
persistió hasta por 4 h después de finalizado el entrenamiento. La
recuperación también ocasionó cambios en el nivel de expresión de
estas enzimas: ERK1/2 y PP1 aumentaron tempranamente, mientras
que PP2B lo hizo tardíamente.
4. El entrenamiento de la extinción de la memoria espacial indujo un
aumento persistente de la actividad enzimática de ERK1/2 y PP2B. El
nivel de expresión de PP1 aumentó inmediatamente después de
finalizada la extinción, pero disminuyó 1 h después de finalizado el
entrenamiento, sin presentar cambios significativos en su actividad
enzimática.
5. El estrés agudo inducido antes de la adquisición de la memoria
espacial indujo cambios en la actividad de ERK1/2 y PP2B. La
actividad de ERK1/2, con respecto a la actividad basal, presentó el

118
aumento más evidente y persistente independientemente de la
duración del estrés agudo. No obstante, cuando el estrés agudo fue
de corta duración, el aumento de la actividad de ERK1/2 fue
significativamente menor que el observado durante la adquisición. El
estrés agudo de corta duración, pero no el prolongado, bloqueó el
incremento en la actividad enzimática de PP2B inducido por la
adquisición. En contraste, el estrés agudo antes de la adquisición solo
provocó aumento del nivel de expresión de PP1.
6. El estrés agudo inducido, antes de la recuperación de la memoria
espacial, provocó cambios en todas las enzimas evaluadas, los cuales
fueron dependientes de la duración del estrés agudo para PP2B y
PP1. El estrés agudo, independientemente de su duración, impidió el
aumento de la actividad de ERK1/2 observado tras la recuperación. La
actividad de PP1 y PP2B aumentó persistentemente con el estrés
agudo prolongado previo a la recuperación, siendo este aumento más
notorio para PP1. No obstante, cuando el estrés agudo fue de corta
duración el aumento en la actividad de PP2B fue significativamente
menor que el observado tras la recuperación.
7. El estrés agudo inducido antes de la extinción de la memoria espacial
provocó un aumento en la actividad de ERK1/2 y PP1. La actividad de
ERK1/2 aumentó de forma persistente cuando el estrés agudo fue de
corta duración y solo ocurrió tempranamente cuando el estrés fue
prolongado. Estos cambios fueron significativamente menores de lo
observado tras la extinción sin estrés previo. La actividad de PP1
aumentó de forma tardía cuando el estrés agudo fue de corta duración
y de forma temprana cuando el estrés fue prolongado. La actividad de
PP2B no se modificó, pero sí su nivel de expresión, el cual aumentó
principalmente con el estrés agudo de corta duración inducido antes
de la extinción.

119
9. ANEXOS
A1. Inducción de estrés agudo por restricción de movimientos y
entrenamiento de la memoria espacial.
1. Inducción de estrés agudo por restricción de movimientos
Los animales se introdujeron en un cilindro de PVC con un perímetro
ligeramente superior al perímetro abdominal, con tapas removibles en los
extremos, durante un periodo corto (1 hora) y un periodo prolongado (4
horas).
2. Entrenamiento del aprendizaje de una tarea de memoria espacial
Se indujo mediante un protocolo de entrenamiento abreviado en el
laberinto de Barnes. El laberinto es una plataforma circular elevada, con
18 agujeros en la periferia, uno de los cuales conduce a una caja, ubicada
bajo la superficie, en la cual el sujeto puede ocultarse. En las paredes
blancas que rodean el laberinto hay figuras geométricas negras que son
las únicas claves disponibles para que el sujeto pueda orientarse en el
espacio. Una intensa iluminación y un ruido blanco motivan al sujeto a
buscar refugio. Se evaluaron tres fases de la memoria espacial:
2.1. Adquisición
Durante esta sesión se hacen 8 ensayos, separados entre sí por
intervalos de 4 min. En la primera sesión, se introduce el sujeto en
el centro del laberinto y se le permite explorarlo en ensayos de
hasta 4 min de duración. Si el animal encuentra la caja de escape
en un tiempo menor, se apagan las luces y la fuente sonora y se
da por terminado el ensayo.

120
2.2. Recuperación
Un día después de la sesión de adquisición se hace la sesión de
evaluación, la cual consta de dos ensayos: En el primer ensayo, se
evalúa la ejecución del animal en condiciones idénticas a las de los
ensayos de adquisición (prueba con caja). En el segundo ensayo,
se retira la caja de escape para evaluar si el animal explora
preferentemente el agujero que conducía a la caja de escape
(prueba sin caja).
2.3. Extinción
Inmediatamente después de la recuperación, con el fin de extinguir
la preferencia espacial adquirida el día anterior, el sujeto es
sometido a siete ensayos adicionales idénticos a la prueba sin
caja. Para caracterizar la eficiencia de la ejecución de la tarea a lo
largo del proceso completo de entrenamiento el comportamiento
del animal en cada ensayo se graba en video y posteriormente se
registra una serie de indicadores comportamentales.
3 Inducción del estrés previo al aprendizaje de la tarea de memoria
espacial.
3.1. Estrés agudo inducido antes de la sesión de adquisición
Los animales se sometieron a restricción de movimientos por
períodos de 1 ó 4 h antes de la sesión de adquisición (en ambos
casos, la restricción de movimientos terminó 30 min antes de
iniciar el entrenamiento). Se evalúo la ejecución durante los
ensayos de adquisición y los ensayos de evaluación y se comparó
con la ejecución de sujetos que recibieron el entrenamiento
completo pero no se sometieron a restricción de movimientos.

121
3.2. Estrés agudo inducido antes de la sesión de recuperación y
extinción
Un día después, a los animales que habían adquirido previamente
el aprendizaje de la memoria espacial, se les sometió a restricción
de movimientos por períodos de 1 ó 4 h antes de la sesión de
evaluación y extinción (en ambos casos, la restricción de
movimientos terminó 30 min antes de iniciar el entrenamiento). Se
evalúo la ejecución durante los ensayos de evaluación y los
ensayos de extinción y se comparó con la ejecución de sujetos que
recibieron el entrenamiento completo pero no se sometieron a
restricción de movimientos.
-20
-15
-10
-5
0
5
10
15
20
BASAL E1-0H E1-1H E1-4H E4-0H E4-1H E4-4H
CA
NT
IDA
D N
OR
MA
LIZ
AD
A (
%B
AS
AL
)
ERK1/2
ERK 1
ERK 2
Figura A2. Cambios en el nivel de expresión de ERK1/2 provocados por el estrés por
inmovilización. Diagrama de barras ilustrando el cambio en el nivel de expresión de ERK1/2.
Se ilustra la media y el error estándar (n=5 animales por grupo). Los animales se sometieron a
estrés corto 1 h (E1) y prolongado 4 h (E4) y sacrificio inmediato (0H), a la hora (1H) y a las 4
horas (4H). (n= 5 animales por grupo). MW, marcador de peso molecular (Estándar pre-teñido de
amplio rango, BIO-RAD).
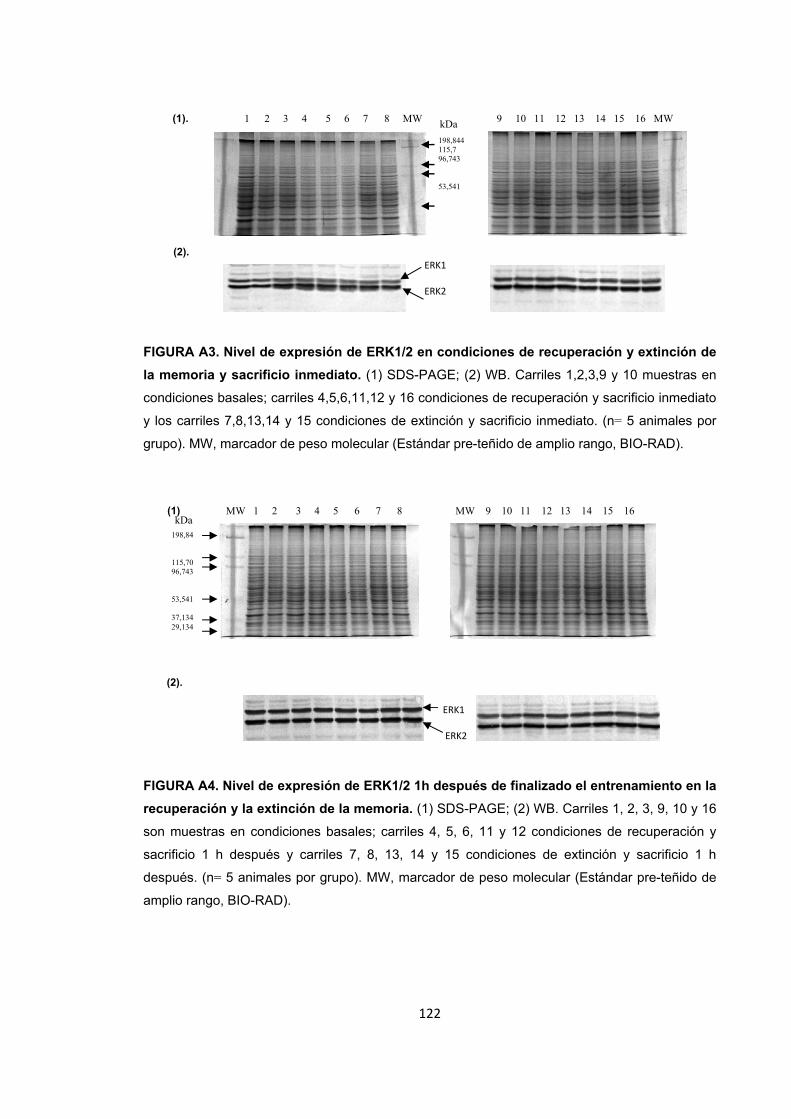
122
(1). 1 2 3 4 5 6 7 8 MW 9 10 11 12 13 14 15 16 MW
(2).
FIGURA A3. Nivel de expresión de ERK1/2 en condiciones de recuperación y extinción de
la memoria y sacrificio inmediato. (1) SDS-PAGE; (2) WB. Carriles 1,2,3,9 y 10 muestras en
condiciones basales; carriles 4,5,6,11,12 y 16 condiciones de recuperación y sacrificio inmediato
y los carriles 7,8,13,14 y 15 condiciones de extinción y sacrificio inmediato. (n= 5 animales por
grupo). MW, marcador de peso molecular (Estándar pre-teñido de amplio rango, BIO-RAD).
(1) MW 1 2 3 4 5 6 7 8 MW 9 10 11 12 13 14 15 16
(2).
FIGURA A4. Nivel de expresión de ERK1/2 1h después de finalizado el entrenamiento en la
recuperación y la extinción de la memoria. (1) SDS-PAGE; (2) WB. Carriles 1, 2, 3, 9, 10 y 16
son muestras en condiciones basales; carriles 4, 5, 6, 11 y 12 condiciones de recuperación y
sacrificio 1 h después y carriles 7, 8, 13, 14 y 15 condiciones de extinción y sacrificio 1 h
después. (n= 5 animales por grupo). MW, marcador de peso molecular (Estándar pre-teñido de
amplio rango, BIO-RAD).
ERK1 ERK2
ERK1 ERK2
198,844 115,7 96,743 53,541
198,84 115,70 96,743 53,541 37,134 29,134
kDa
kDa
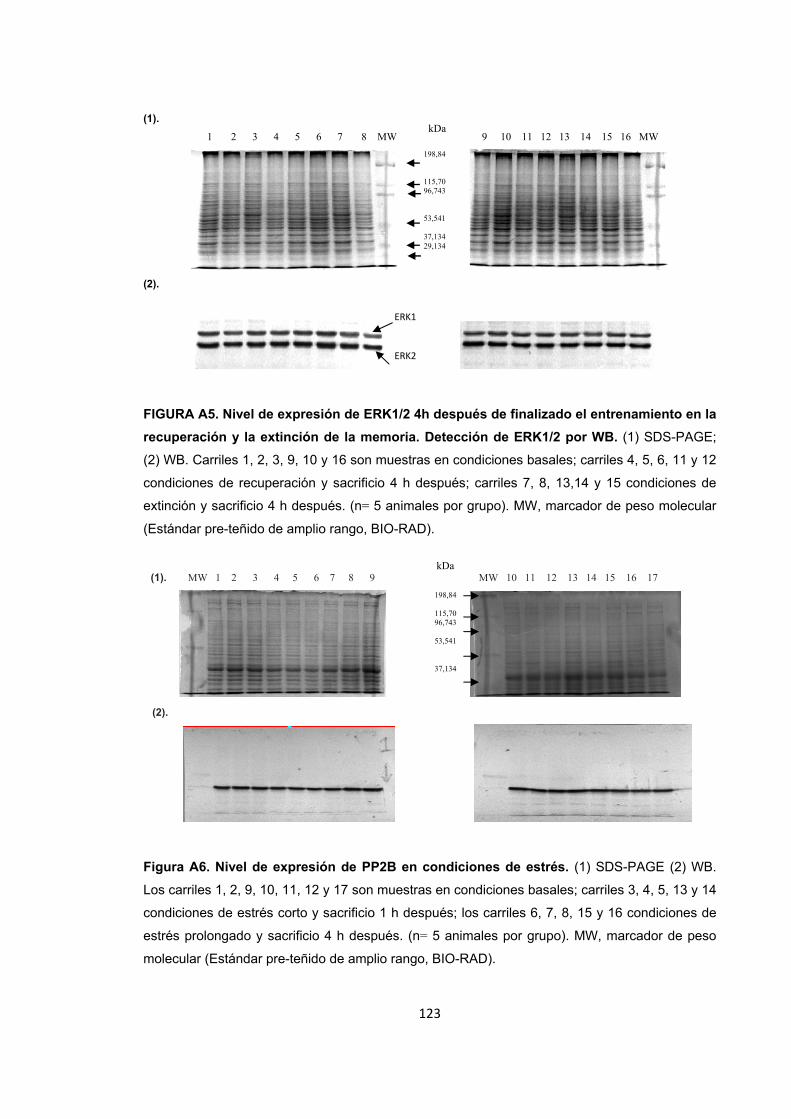
123
(1).
1 2 3 4 5 6 7 8 MW 9 10 11 12 13 14 15 16 MW
(2).
FIGURA A5. Nivel de expresión de ERK1/2 4h después de finalizado el entrenamiento en la
recuperación y la extinción de la memoria. Detección de ERK1/2 por WB. (1) SDS-PAGE;
(2) WB. Carriles 1, 2, 3, 9, 10 y 16 son muestras en condiciones basales; carriles 4, 5, 6, 11 y 12
condiciones de recuperación y sacrificio 4 h después; carriles 7, 8, 13,14 y 15 condiciones de
extinción y sacrificio 4 h después. (n= 5 animales por grupo). MW, marcador de peso molecular
(Estándar pre-teñido de amplio rango, BIO-RAD).
(1). MW 1 2 3 4 5 6 7 8 9 MW 10 11 12 13 14 15 16 17
(2).
Figura A6. Nivel de expresión de PP2B en condiciones de estrés. (1) SDS-PAGE (2) WB.
Los carriles 1, 2, 9, 10, 11, 12 y 17 son muestras en condiciones basales; carriles 3, 4, 5, 13 y 14
condiciones de estrés corto y sacrificio 1 h después; los carriles 6, 7, 8, 15 y 16 condiciones de
estrés prolongado y sacrificio 4 h después. (n= 5 animales por grupo). MW, marcador de peso
molecular (Estándar pre-teñido de amplio rango, BIO-RAD).
ERK1 ERK2
198,84 115,70 96,743 53,541 37,134 29,134
198,84 115,70 96,743 53,541 37,134
kDa
kDa
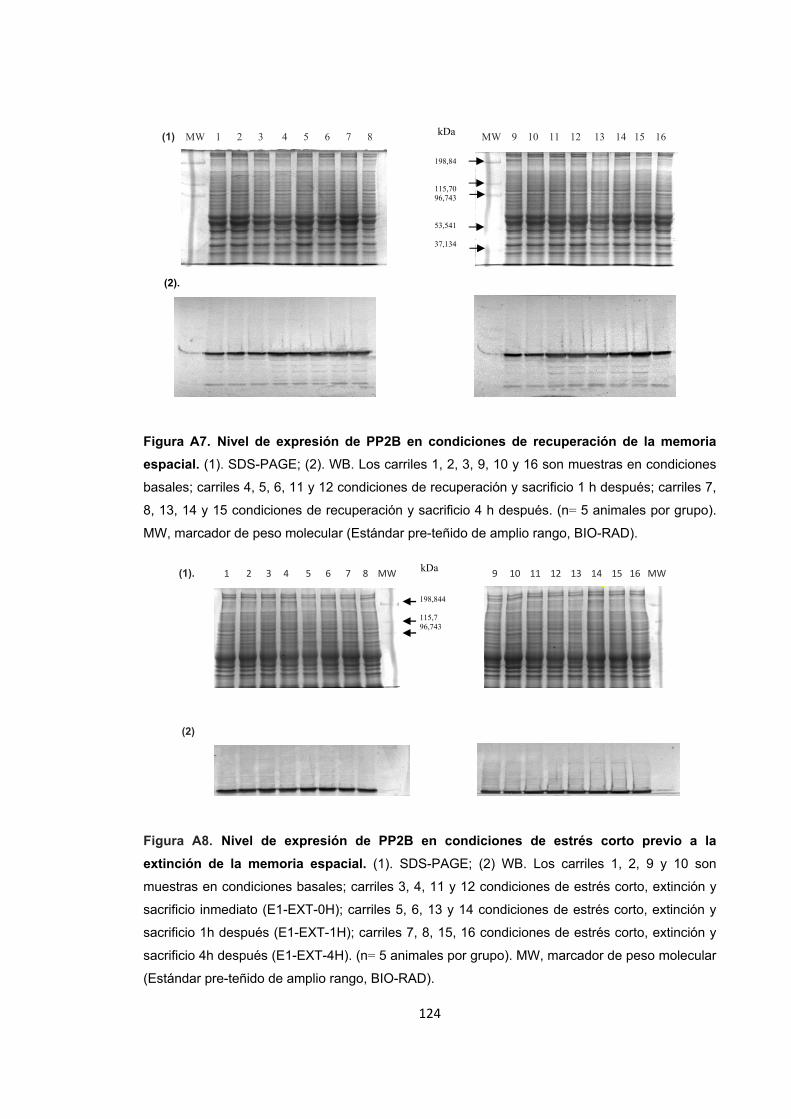
124
(1) MW 1 2 3 4 5 6 7 8 MW 9 10 11 12 13 14 15 16
(2).
Figura A7. Nivel de expresión de PP2B en condiciones de recuperación de la memoria
espacial. (1). SDS-PAGE; (2). WB. Los carriles 1, 2, 3, 9, 10 y 16 son muestras en condiciones
basales; carriles 4, 5, 6, 11 y 12 condiciones de recuperación y sacrificio 1 h después; carriles 7,
8, 13, 14 y 15 condiciones de recuperación y sacrificio 4 h después. (n= 5 animales por grupo).
MW, marcador de peso molecular (Estándar pre-teñido de amplio rango, BIO-RAD).
(1). 1 2 3 4 5 6 7 8 MW 9 10 11 12 13 14 15 16 MW
(2)
Figura A8. Nivel de expresión de PP2B en condiciones de estrés corto previo a la
extinción de la memoria espacial. (1). SDS-PAGE; (2) WB. Los carriles 1, 2, 9 y 10 son
muestras en condiciones basales; carriles 3, 4, 11 y 12 condiciones de estrés corto, extinción y
sacrificio inmediato (E1-EXT-0H); carriles 5, 6, 13 y 14 condiciones de estrés corto, extinción y
sacrificio 1h después (E1-EXT-1H); carriles 7, 8, 15, 16 condiciones de estrés corto, extinción y
sacrificio 4h después (E1-EXT-4H). (n= 5 animales por grupo). MW, marcador de peso molecular
(Estándar pre-teñido de amplio rango, BIO-RAD).
198,84 115,70 96,743 53,541 37,134
198,844 115,7 96,743
kDa
kDa

125
1.
1 2 3 4 5 6 7 8 MW 9 10 11 12 13 14 15 16 MW
2.
Figura A9. Nivel de expresión de PP2B en condiciones de estrés prolongado previo a la
extinción. (1) SDS-PAGE; (2) WB. Los carriles 1, 2, 9 y 10 son muestras en condiciones
basales; carriles 3, 4, 11 y 12 condiciones de estrés prolongado, extinción y sacrificio inmediato
(E4-EXT-0H); carriles 5, 6, 13 y 14 condiciones de estrés prolongado, extinción y sacrificio 1h
después (E4-EXT-1H); carriles 7, 8, 15, 16 condiciones de estrés prolongado, extinción y
sacrificio 4h después (E4-EXT-4H). (n= 5 animales por grupo). MW, marcador de peso molecular
(Estándar pre-teñido de amplio rango, BIO-RAD).
a).
0
50
100
150
200
250
‐5 ‐4 ‐3 ‐2 ‐1 0 1 2 3 4 5
ΔEN
LA ACTIVIDAD ENZIMATICA
TRATAMIENTO
E1 ERK1/2
E1 PP1
E1 PP2B
b).
198,844 115,7 96,743
kDa
126
0
50
100
150
200
250
300
‐5 ‐4 ‐3 ‐2 ‐1 0 1 2 3 4 5
ΔEN
LA ACTIVIDAD ENZIMATICA
TRATAMIENTO
E4 ERK1/2
E4 PP1
E4 PP2B
c).
0
50
100
150
200
250
300
350
400
‐5 ‐4 ‐3 ‐2 ‐1 0 1 2 3 4 5
ΔE
N L
A A
CT
IVID
AD
EN
ZIM
AT
ICA
TRATAMIENTO
ADQ ERK1/2
ADQ PP1
ADQ PP2B
d).
0
50
100
150
200
250
300
350
400
‐5 ‐4 ‐3 ‐2 ‐1 0 1 2 3 4 5
ΔE
N L
A A
CT
IVID
AD
EN
ZIM
AT
ICA
TRATAMIENTO
REC ERK1/2
REC PP1
REC PP2B
e).
127
0
20
40
60
80
100
120
140
160
180
200
‐5 ‐4 ‐3 ‐2 ‐1 0 1 2 3 4 5
ΔE
N L
A A
CT
IVID
AD
EN
ZIM
AT
ICA
TRATAMIENTO
EXT ERK1/2
EXT PP1
EXT PP2B
f).
0
50
100
150
200
250
300
-6 -4 -2 0 2 4 6
ΔE
N L
A A
CT
IVID
AD
EN
ZIM
AT
ICA
TRATAMIENTO
E1 y ADQ ERK1/2
E1 y ADQ PP1
E1 y ADQ PP2B
g).
128
0
50
100
150
200
250
300
-6 -4 -2 0 2 4 6
ΔE
N L
A A
CT
IVID
AD
EN
ZIM
AT
ICA
TRATAMIENTO
E4 y ADQ ERK1/2
E4 y ADQ PP1
E4 y ADQ PP2B
h).
-50
0
50
100
150
200
250
300
-5 -4 -3 -2 -1 0 1 2 3 4 5
ΔE
N L
A A
CT
IVID
AD
EN
ZIM
AT
ICA
TRATAMIENTO
E1 y REC ERK1/2
E1 y REC PP1
E1 y REC PP2B
i).
-50
0
50
100
150
200
250
300
-5 -4 -3 -2 -1 0 1 2 3 4 5
ΔE
N L
A A
CT
IVID
AD
EN
ZIM
AT
ICA
TRATAMIENTO
E4 y REC ERK1/2
E4 y REC PP1
E4 y REC PP2B
j).
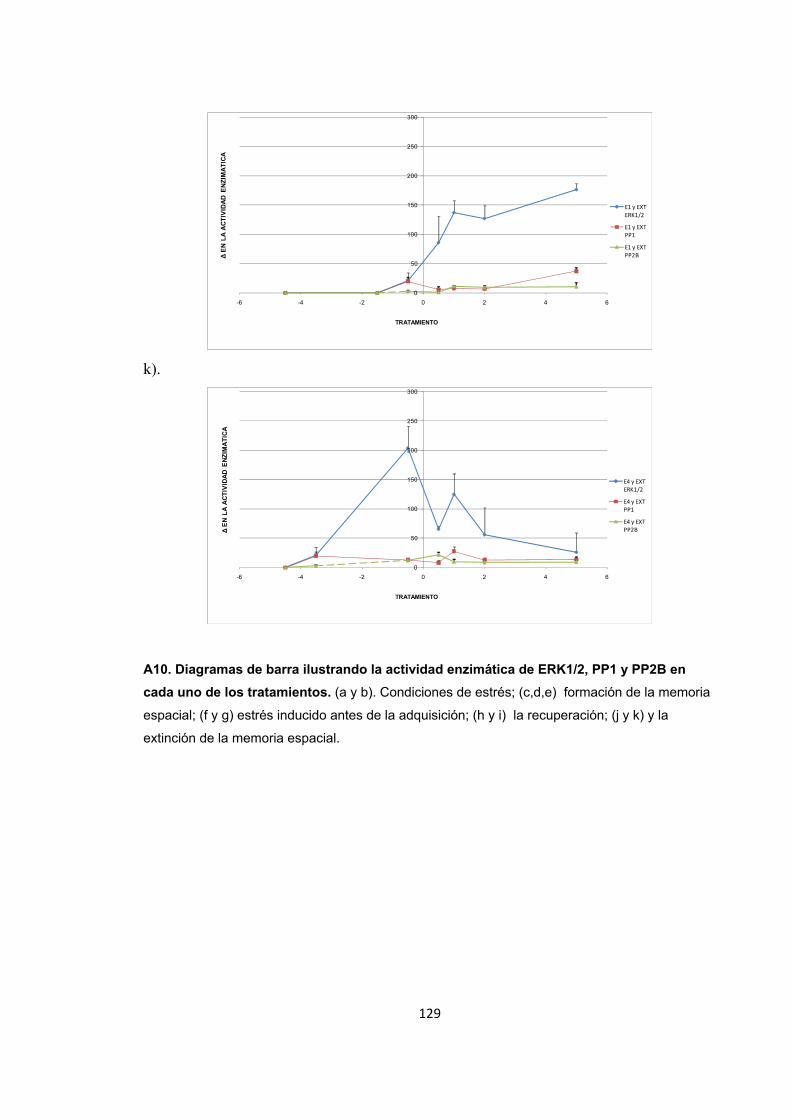
129
0
50
100
150
200
250
300
-6 -4 -2 0 2 4 6
ΔE
N L
A A
CT
IVID
AD
EN
ZIM
AT
ICA
TRATAMIENTO
E1 y EXT ERK1/2
E1 y EXT PP1
E1 y EXT PP2B
k).
0
50
100
150
200
250
300
-6 -4 -2 0 2 4 6
ΔE
N L
A A
CT
IVID
AD
EN
ZIM
AT
ICA
TRATAMIENTO
E4 y EXT ERK1/2
E4 y EXT PP1
E4 y EXT PP2B
A10. Diagramas de barra ilustrando la actividad enzimática de ERK1/2, PP1 y PP2B en
cada uno de los tratamientos. (a y b). Condiciones de estrés; (c,d,e) formación de la memoria
espacial; (f y g) estrés inducido antes de la adquisición; (h y i) la recuperación; (j y k) y la
extinción de la memoria espacial.
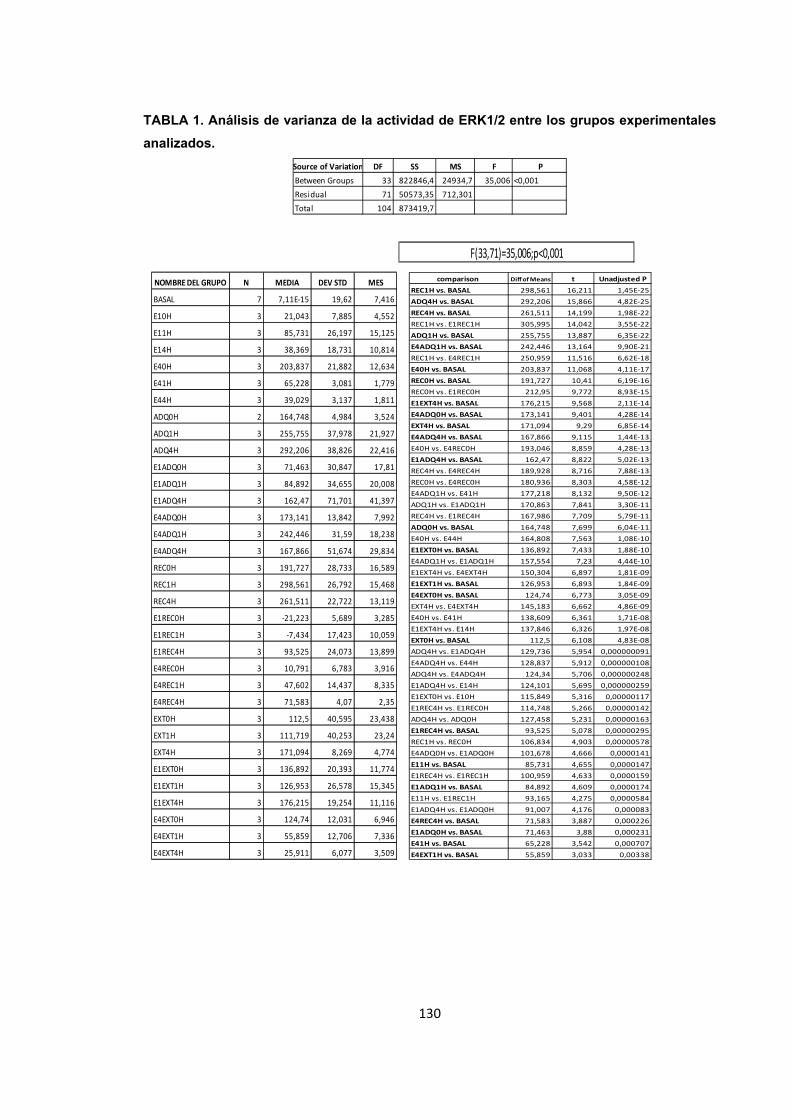
130
TABLA 1. Análisis de varianza de la actividad de ERK1/2 entre los grupos experimentales
analizados.
Source of Variation DF SS MS F P
Between Groups 33 822846,4 24934,7 35,006 <0,001
Residual 71 50573,35 712,301
Total 104 873419,7
F(33,71)=35,006;p<0,001
NOMBRE DEL GRUPO N MEDIA DEV STD MES
BASAL 7 7,11E‐15 19,62 7,416
E10H 3 21,043 7,885 4,552
E11H 3 85,731 26,197 15,125
E14H 3 38,369 18,731 10,814
E40H 3 203,837 21,882 12,634
E41H 3 65,228 3,081 1,779
E44H 3 39,029 3,137 1,811
ADQ0H 2 164,748 4,984 3,524
ADQ1H 3 255,755 37,978 21,927
ADQ4H 3 292,206 38,826 22,416
E1ADQ0H 3 71,463 30,847 17,81
E1ADQ1H 3 84,892 34,655 20,008
E1ADQ4H 3 162,47 71,701 41,397
E4ADQ0H 3 173,141 13,842 7,992
E4ADQ1H 3 242,446 31,59 18,238
E4ADQ4H 3 167,866 51,674 29,834
REC0H 3 191,727 28,733 16,589
REC1H 3 298,561 26,792 15,468
REC4H 3 261,511 22,722 13,119
E1REC0H 3 ‐21,223 5,689 3,285
E1REC1H 3 ‐7,434 17,423 10,059
E1REC4H 3 93,525 24,073 13,899
E4REC0H 3 10,791 6,783 3,916
E4REC1H 3 47,602 14,437 8,335
E4REC4H 3 71,583 4,07 2,35
EXT0H 3 112,5 40,595 23,438
EXT1H 3 111,719 40,253 23,24
EXT4H 3 171,094 8,269 4,774
E1EXT0H 3 136,892 20,393 11,774
E1EXT1H 3 126,953 26,578 15,345
E1EXT4H 3 176,215 19,254 11,116
E4EXT0H 3 124,74 12,031 6,946
E4EXT1H 3 55,859 12,706 7,336
E4EXT4H 3 25,911 6,077 3,509
comparison Diff of Means t Unadjusted P
REC1H vs. BASAL 298,561 16,211 1,45E‐25
ADQ4H vs. BASAL 292,206 15,866 4,82E‐25
REC4H vs. BASAL 261,511 14,199 1,98E‐22
REC1H vs. E1REC1H 305,995 14,042 3,55E‐22
ADQ1H vs. BASAL 255,755 13,887 6,35E‐22
E4ADQ1H vs. BASAL 242,446 13,164 9,90E‐21
REC1H vs. E4REC1H 250,959 11,516 6,62E‐18
E40H vs. BASAL 203,837 11,068 4,11E‐17
REC0H vs. BASAL 191,727 10,41 6,19E‐16
REC0H vs. E1REC0H 212,95 9,772 8,93E‐15
E1EXT4H vs. BASAL 176,215 9,568 2,11E‐14
E4ADQ0H vs. BASAL 173,141 9,401 4,28E‐14
EXT4H vs. BASAL 171,094 9,29 6,85E‐14
E4ADQ4H vs. BASAL 167,866 9,115 1,44E‐13
E40H vs. E4REC0H 193,046 8,859 4,28E‐13
E1ADQ4H vs. BASAL 162,47 8,822 5,02E‐13
REC4H vs. E4REC4H 189,928 8,716 7,88E‐13
REC0H vs. E4REC0H 180,936 8,303 4,58E‐12
E4ADQ1H vs. E41H 177,218 8,132 9,50E‐12
ADQ1H vs. E1ADQ1H 170,863 7,841 3,30E‐11
REC4H vs. E1REC4H 167,986 7,709 5,79E‐11
ADQ0H vs. BASAL 164,748 7,699 6,04E‐11
E40H vs. E44H 164,808 7,563 1,08E‐10
E1EXT0H vs. BASAL 136,892 7,433 1,88E‐10
E4ADQ1H vs. E1ADQ1H 157,554 7,23 4,44E‐10
E1EXT4H vs. E4EXT4H 150,304 6,897 1,81E‐09
E1EXT1H vs. BASAL 126,953 6,893 1,84E‐09
E4EXT0H vs. BASAL 124,74 6,773 3,05E‐09
EXT4H vs. E4EXT4H 145,183 6,662 4,86E‐09
E40H vs. E41H 138,609 6,361 1,71E‐08
E1EXT4H vs. E14H 137,846 6,326 1,97E‐08
EXT0H vs. BASAL 112,5 6,108 4,83E‐08
ADQ4H vs. E1ADQ4H 129,736 5,954 0,000000091
E4ADQ4H vs. E44H 128,837 5,912 0,000000108
ADQ4H vs. E4ADQ4H 124,34 5,706 0,000000248
E1ADQ4H vs. E14H 124,101 5,695 0,000000259
E1EXT0H vs. E10H 115,849 5,316 0,00000117
E1REC4H vs. E1REC0H 114,748 5,266 0,00000142
ADQ4H vs. ADQ0H 127,458 5,231 0,00000163
E1REC4H vs. BASAL 93,525 5,078 0,00000295
REC1H vs. REC0H 106,834 4,903 0,00000578
E4ADQ0H vs. E1ADQ0H 101,678 4,666 0,0000141
E11H vs. BASAL 85,731 4,655 0,0000147
E1REC4H vs. E1REC1H 100,959 4,633 0,0000159
E1ADQ1H vs. BASAL 84,892 4,609 0,0000174
E11H vs. E1REC1H 93,165 4,275 0,0000584
E1ADQ4H vs. E1ADQ0H 91,007 4,176 0,000083
E4REC4H vs. BASAL 71,583 3,887 0,000226
E1ADQ0H vs. BASAL 71,463 3,88 0,000231
E41H vs. BASAL 65,228 3,542 0,000707
E4EXT1H vs. BASAL 55,859 3,033 0,00338
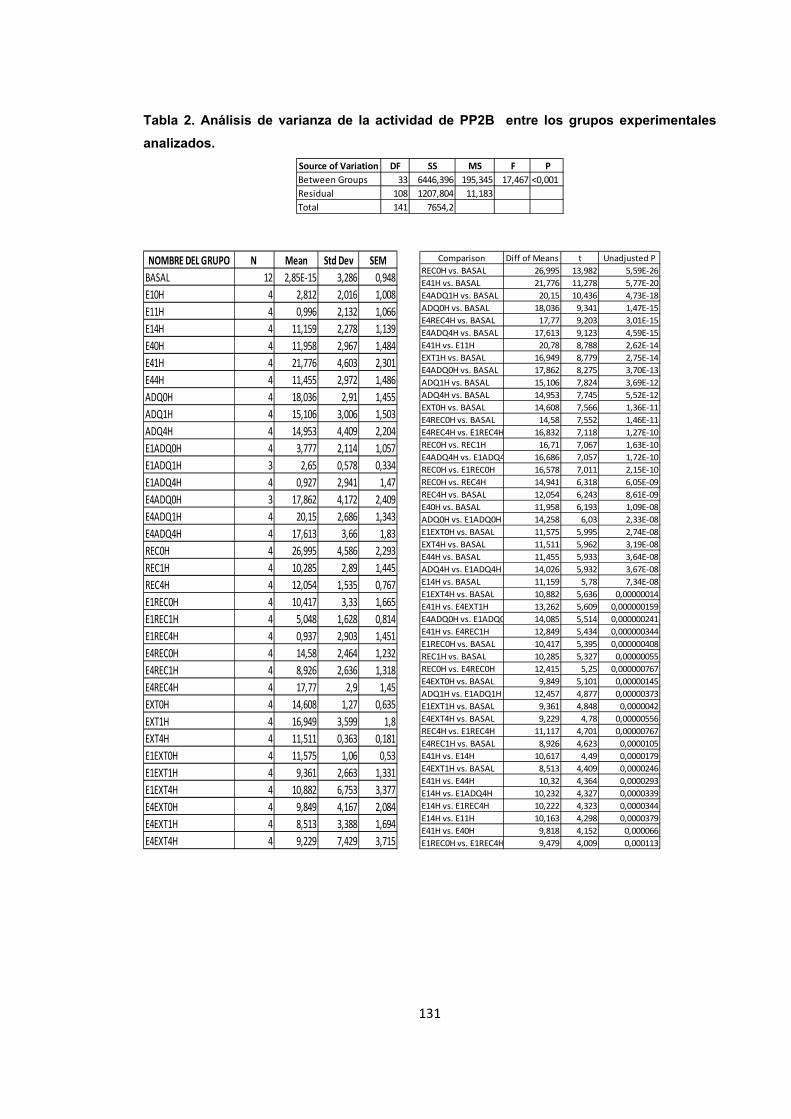
131
Tabla 2. Análisis de varianza de la actividad de PP2B entre los grupos experimentales
analizados.
Source of Variation DF SS MS F P
Between Groups 33 6446,396 195,345 17,467 <0,001
Residual 108 1207,804 11,183
Total 141 7654,2
NOMBRE DEL GRUPO N Mean Std Dev SEM
BASAL 12 2,85E‐15 3,286 0,948
E10H 4 2,812 2,016 1,008
E11H 4 0,996 2,132 1,066
E14H 4 11,159 2,278 1,139
E40H 4 11,958 2,967 1,484
E41H 4 21,776 4,603 2,301
E44H 4 11,455 2,972 1,486
ADQ0H 4 18,036 2,91 1,455
ADQ1H 4 15,106 3,006 1,503
ADQ4H 4 14,953 4,409 2,204
E1ADQ0H 4 3,777 2,114 1,057
E1ADQ1H 3 2,65 0,578 0,334
E1ADQ4H 4 0,927 2,941 1,47
E4ADQ0H 3 17,862 4,172 2,409
E4ADQ1H 4 20,15 2,686 1,343
E4ADQ4H 4 17,613 3,66 1,83
REC0H 4 26,995 4,586 2,293
REC1H 4 10,285 2,89 1,445
REC4H 4 12,054 1,535 0,767
E1REC0H 4 10,417 3,33 1,665
E1REC1H 4 5,048 1,628 0,814
E1REC4H 4 0,937 2,903 1,451
E4REC0H 4 14,58 2,464 1,232
E4REC1H 4 8,926 2,636 1,318
E4REC4H 4 17,77 2,9 1,45
EXT0H 4 14,608 1,27 0,635
EXT1H 4 16,949 3,599 1,8
EXT4H 4 11,511 0,363 0,181
E1EXT0H 4 11,575 1,06 0,53
E1EXT1H 4 9,361 2,663 1,331
E1EXT4H 4 10,882 6,753 3,377
E4EXT0H 4 9,849 4,167 2,084
E4EXT1H 4 8,513 3,388 1,694
E4EXT4H 4 9,229 7,429 3,715
Comparison Diff of Means t Unadjusted P
REC0H vs. BASAL 26,995 13,982 5,59E‐26
E41H vs. BASAL 21,776 11,278 5,77E‐20
E4ADQ1H vs. BASAL 20,15 10,436 4,73E‐18
ADQ0H vs. BASAL 18,036 9,341 1,47E‐15
E4REC4H vs. BASAL 17,77 9,203 3,01E‐15
E4ADQ4H vs. BASAL 17,613 9,123 4,59E‐15
E41H vs. E11H 20,78 8,788 2,62E‐14
EXT1H vs. BASAL 16,949 8,779 2,75E‐14
E4ADQ0H vs. BASAL 17,862 8,275 3,70E‐13
ADQ1H vs. BASAL 15,106 7,824 3,69E‐12
ADQ4H vs. BASAL 14,953 7,745 5,52E‐12
EXT0H vs. BASAL 14,608 7,566 1,36E‐11
E4REC0H vs. BASAL 14,58 7,552 1,46E‐11
E4REC4H vs. E1REC4H 16,832 7,118 1,27E‐10
REC0H vs. REC1H 16,71 7,067 1,63E‐10
E4ADQ4H vs. E1ADQ4 16,686 7,057 1,72E‐10
REC0H vs. E1REC0H 16,578 7,011 2,15E‐10
REC0H vs. REC4H 14,941 6,318 6,05E‐09
REC4H vs. BASAL 12,054 6,243 8,61E‐09
E40H vs. BASAL 11,958 6,193 1,09E‐08
ADQ0H vs. E1ADQ0H 14,258 6,03 2,33E‐08
E1EXT0H vs. BASAL 11,575 5,995 2,74E‐08
EXT4H vs. BASAL 11,511 5,962 3,19E‐08
E44H vs. BASAL 11,455 5,933 3,64E‐08
ADQ4H vs. E1ADQ4H 14,026 5,932 3,67E‐08
E14H vs. BASAL 11,159 5,78 7,34E‐08
E1EXT4H vs. BASAL 10,882 5,636 0,00000014
E41H vs. E4EXT1H 13,262 5,609 0,000000159
E4ADQ0H vs. E1ADQ0 14,085 5,514 0,000000241
E41H vs. E4REC1H 12,849 5,434 0,000000344
E1REC0H vs. BASAL 10,417 5,395 0,000000408
REC1H vs. BASAL 10,285 5,327 0,00000055
REC0H vs. E4REC0H 12,415 5,25 0,000000767
E4EXT0H vs. BASAL 9,849 5,101 0,00000145
ADQ1H vs. E1ADQ1H 12,457 4,877 0,00000373
E1EXT1H vs. BASAL 9,361 4,848 0,0000042
E4EXT4H vs. BASAL 9,229 4,78 0,00000556
REC4H vs. E1REC4H 11,117 4,701 0,00000767
E4REC1H vs. BASAL 8,926 4,623 0,0000105
E41H vs. E14H 10,617 4,49 0,0000179
E4EXT1H vs. BASAL 8,513 4,409 0,0000246
E41H vs. E44H 10,32 4,364 0,0000293
E14H vs. E1ADQ4H 10,232 4,327 0,0000339
E14H vs. E1REC4H 10,222 4,323 0,0000344
E14H vs. E11H 10,163 4,298 0,0000379
E41H vs. E40H 9,818 4,152 0,000066
E1REC0H vs. E1REC4H 9,479 4,009 0,000113
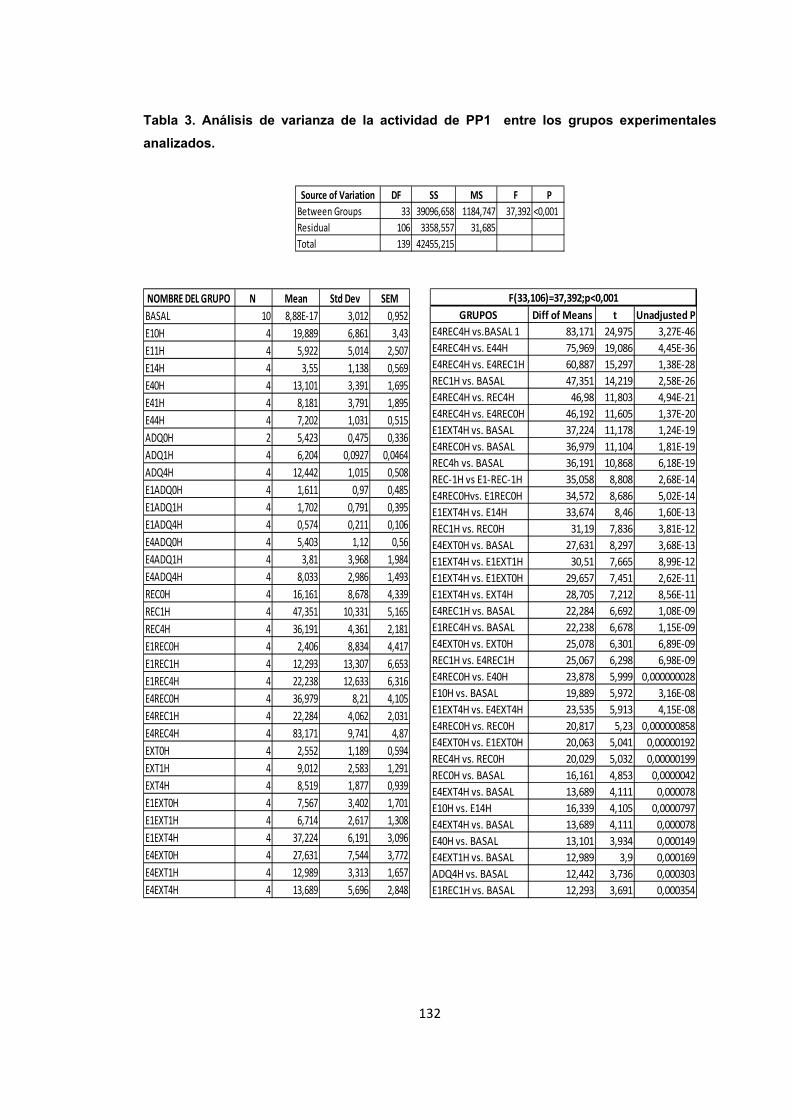
132
Tabla 3. Análisis de varianza de la actividad de PP1 entre los grupos experimentales
analizados.
Source of Variation DF SS MS F P
Between Groups 33 39096,658 1184,747 37,392 <0,001
Residual 106 3358,557 31,685
Total 139 42455,215
NOMBRE DEL GRUPO N Mean Std Dev SEM
BASAL 10 8,88E‐17 3,012 0,952
E10H 4 19,889 6,861 3,43
E11H 4 5,922 5,014 2,507
E14H 4 3,55 1,138 0,569
E40H 4 13,101 3,391 1,695
E41H 4 8,181 3,791 1,895
E44H 4 7,202 1,031 0,515
ADQ0H 2 5,423 0,475 0,336
ADQ1H 4 6,204 0,0927 0,0464
ADQ4H 4 12,442 1,015 0,508
E1ADQ0H 4 1,611 0,97 0,485
E1ADQ1H 4 1,702 0,791 0,395
E1ADQ4H 4 0,574 0,211 0,106
E4ADQ0H 4 5,403 1,12 0,56
E4ADQ1H 4 3,81 3,968 1,984
E4ADQ4H 4 8,033 2,986 1,493
REC0H 4 16,161 8,678 4,339
REC1H 4 47,351 10,331 5,165
REC4H 4 36,191 4,361 2,181
E1REC0H 4 2,406 8,834 4,417
E1REC1H 4 12,293 13,307 6,653
E1REC4H 4 22,238 12,633 6,316
E4REC0H 4 36,979 8,21 4,105
E4REC1H 4 22,284 4,062 2,031
E4REC4H 4 83,171 9,741 4,87
EXT0H 4 2,552 1,189 0,594
EXT1H 4 9,012 2,583 1,291
EXT4H 4 8,519 1,877 0,939
E1EXT0H 4 7,567 3,402 1,701
E1EXT1H 4 6,714 2,617 1,308
E1EXT4H 4 37,224 6,191 3,096
E4EXT0H 4 27,631 7,544 3,772
E4EXT1H 4 12,989 3,313 1,657
E4EXT4H 4 13,689 5,696 2,848
GRUPOS Diff of Means t Unadjusted P
E4REC4H vs.BASAL 1 83,171 24,975 3,27E‐46
E4REC4H vs. E44H 75,969 19,086 4,45E‐36
E4REC4H vs. E4REC1H 60,887 15,297 1,38E‐28
REC1H vs. BASAL 47,351 14,219 2,58E‐26
E4REC4H vs. REC4H 46,98 11,803 4,94E‐21
E4REC4H vs. E4REC0H 46,192 11,605 1,37E‐20
E1EXT4H vs. BASAL 37,224 11,178 1,24E‐19
E4REC0H vs. BASAL 36,979 11,104 1,81E‐19
REC4h vs. BASAL 36,191 10,868 6,18E‐19
REC‐1H vs E1‐REC‐1H 35,058 8,808 2,68E‐14
E4REC0Hvs. E1REC0H 34,572 8,686 5,02E‐14
E1EXT4H vs. E14H 33,674 8,46 1,60E‐13
REC1H vs. REC0H 31,19 7,836 3,81E‐12
E4EXT0H vs. BASAL 27,631 8,297 3,68E‐13
E1EXT4H vs. E1EXT1H 30,51 7,665 8,99E‐12
E1EXT4H vs. E1EXT0H 29,657 7,451 2,62E‐11
E1EXT4H vs. EXT4H 28,705 7,212 8,56E‐11
E4REC1H vs. BASAL 22,284 6,692 1,08E‐09
E1REC4H vs. BASAL 22,238 6,678 1,15E‐09
E4EXT0H vs. EXT0H 25,078 6,301 6,89E‐09
REC1H vs. E4REC1H 25,067 6,298 6,98E‐09
E4REC0H vs. E40H 23,878 5,999 0,000000028
E10H vs. BASAL 19,889 5,972 3,16E‐08
E1EXT4H vs. E4EXT4H 23,535 5,913 4,15E‐08
E4REC0H vs. REC0H 20,817 5,23 0,000000858
E4EXT0H vs. E1EXT0H 20,063 5,041 0,00000192
REC4H vs. REC0H 20,029 5,032 0,00000199
REC0H vs. BASAL 16,161 4,853 0,0000042
E4EXT4H vs. BASAL 13,689 4,111 0,000078
E10H vs. E14H 16,339 4,105 0,0000797
E4EXT4H vs. BASAL 13,689 4,111 0,000078
E40H vs. BASAL 13,101 3,934 0,000149
E4EXT1H vs. BASAL 12,989 3,9 0,000169
ADQ4H vs. BASAL 12,442 3,736 0,000303
E1REC1H vs. BASAL 12,293 3,691 0,000354
F(33,106)=37,392;p<0,001

133

134
10. BIBLIOGRAFÍA
Aguilera G, Rabadan-Diehl C (2000). Vasopressinergic regulation of the hypothalamic–pituitary–adrenal axis: Implications for stress adaptation. Regul. Pept. 96:23–29. Aine Kelly, Serge Laroche, Sabrina Davis (2003). Activation of Mitogen-Activated Protein Kinase/Extracellular Signal-Regulated Kinase in Hippocampal Circuitry Is Required for Consolidation and Reconsolidation of Recognition Memory. J Neurosci. 12; 23: 5354-60. Akirav I, Richter-Levin G (1999). Biphasic Modulation of Hippocampal Plasticity by Behavioral Stress and Basolateral Amygdala Stimulation in the Rat. J Neurosci.19; 23:10530-35. Akirav Irit, Kozenicky Maya,Tal Dadi, Sandi Carmen,Venero Cesar y Gal Richter-Levin (2004). A Facilitative Role for Corticosterone in the Acquisition of a Spatial Task Under Moderate Stress. Learn Mem.11: 188-95. Alfonso J, Frick LR, Silberman DM, Palumbo ML, Genaro AM, Frasch AC (2006). Regulation of hippocampal gene expression is conserved in two species subjected to different stressors and antidepressant treatments. Biol Psychiatry. 59; 3: 244-51. Alistair T. R. Sim (1992). The Regulation and Function of Protein Phosphatases in the Brain. Mol Neurobiol. 5: 229-46. Alkon DL, Epstein H, Kuzirian A, Bennett MC, Nelson TJ (2005). Protein synthesis required for long-term memory is induced by PKC activation on days before associative learning. Proc Natl Acad Sci U S A. 102; 45: 16432-7. Andoh T, Chock PB, Chiueh CC (2002). Preconditioning-mediated neuroprotection: role of nitric oxide, cGMP, and new protein expression. Ann N Y Acad Sci. 962: 1-7. Anisman H, Hayley S, Kelly O, Borowski T, Merali Z. (2001). Psychogenic, neurogenic, and systemic stressor effects on plasma corticosterone and behavior: mouse strain-dependent outcomes. Behav Neurosci..115; 2: 443-54.

135
Araujo AP, DeLucia R, Scavone C, Planeta CS (2003). Repeated predictable or unpredictable stress: effects on cocaine-induced locomotion and cyclic AMP-dependent protein kinase activity. Behav Brain Res. 139; 1-2: 75-81. Atkins, C. M, Selcher, J. C, Petraitis, J. J, Trzaskos, J. M, Sweatt, J. D (1998). The MAPK cascade is required for mammalian associative learning. Nat Neurosci. 1: 602–09. Arzt Eduardo y Holsboer Florian (2006). CRF signaling: molecular specificity for drug targeting in the CNS. Trends Pharmacol Sci. 27; 10: 531-38. Bach ME, Hawkins RD, Osman M, Kandel ER, Mayford M (1995). Impairment of spatial but not contextual memory in CaMKII mutant mice with a selective loss of hippocampal LTP in the range of the theta frequency. Cell. 81; 6: 905-15. Barnes C.A (1979). Memory deficits associated with senescence a neurophysiological and behavioral study in the rat. J Comp Physiol Psychol.93; 3: 74-104. Barnes P.J, Adcock I.M, Ito K (2005). Histone acetylation and deacetylation: importance in inflammatory lung diseases. Eur Respir J. 25: 552-63. Bernabeu Ramón, Schroder Nadja, Quevedo Joao, Cammarota Martín, Izquierdo Iván and MedinaCA Jorge H (1997). Further evidence for the involvement of a hippocampal cGMP/cGMP dependent protein kinase cascade in memory consolidation. Neuroreport. 8; 9–10: 2221-24. Bennett PC, Zhao W, Lawan A, Ng KT (1996). Cyclosporin A, an inhibitor of calcineurin, impairs memory formation in dayold chicks. Brain Res. 730:107-17. Bennett PC, Zhao W, Ng KT (2001). Concentration-dependent effects of protein phosphatases (PP) inhibitors implicate PP1 and PP2A in different stages of memory formation. Neurobiol Learn. 75: 91-110. Bennett PC, Moutsoulas P, Lawen A, Perini E, Ng KT (2003). Novel effects on memory observed following unilateral intracranial administration of okadaic acid, cyclosporine A, FK506 and [VeVal4]CyA. Brain Res. 988:56-6. Blank Thomas, Nijholt Ingrid, Grammatopoulos Dimitris K, Randeva Harpal S Hillhouse Edward W and Spiess Joachim (2003). Corticotropin-Releasing Factor Receptors Couple to Multiple G-Proteins to Activate Diverse Intracellular

136
Signaling Pathways in Mouse Hippocampus: Role in Neuronal Excitability and associative Learning. J Neurosci. 23; 2: 700-07. Blank T, Nijholt I, Vollstaedt S, Spiess J (2003). The corticotropin-releasing factor receptor 1 antagonist CP-154,526 reverses stress-induced learning deficits in mice. Behav Brain Res. 138; 2: 207-13. Blum S, Moore AN, Adams F, Dash PK (1999). A mitogen-activated protein kinase cascade in the CA1/CA2 subfield of the dorsal hippocampus is essential for long-term spatial memory. J Neurosci. 19; 9: 3535-44.
Bonini JS, Da Silva WC, Bevilaqua LR, Medina JH, Izquierdo I, Cammarota M (2007). On the participation of hippocampal PKC in acquisition, consolidation and reconsolidation of spatial memory. Neuroscience. 147; 1: 37-45.
Broadbent NJ, Squire LR, Clark RE (2004). Spatial memory, recognition memory, and the hippocampus. Proc Natl Acad Sci U S A.101(40):14515-20. Burnette W. Neal (1981). "'Western blotting': electrophoretic transfer of proteins from sodium dodecyl sulfate — polyacrylamide gels to unmodified nitrocellulose and radiographic detection with antibody and radioiodinated protein A". Anal Biochem.112 (2): 195–203
Burque TR, Zhang ZY (1998). Protein-tyrosine phosphatases: structure, mechanism, and inhibitor discovery. Biopolymers. 47: 225-41.
Cai WH, Blundell J, Han J, Greene RW, Powell CM (2006). Postreactivation glucocorticoids impair recall of established fear memory. J Neurosci. 26: 9560–66. Cammarota Martín, Barros Daniela M, Vianna Mónica R.M, Bevilaqua Lia R.M, Coitinho Adriana, Szapiro Germánn, Izquierdo Luciana A, Medina Jorge H And Izquierdo Iván (2004). The transition from memory retrieval to extinction. An Acad Bras Cienc. 76; 3: 573-82.
Cano E, Mahadevan LC (1995). Parallel signal processing among mammalian MAPKs. Trends Biochem Sci. 20; 3:117-22.
Cannon W.B (1929a). Organization for physiological homeostasis. Physiol Rev IX. pp. 399–431.
Cannon W.B (1929b). The sympathetic division of the autonomic system in relation to homeostasis. Arch Neurol Psychiatry. 22: 282-94.

137
Carvajal, A (2006). Efectos de la inyección sistémica de corticosterona, como Modelo de estrés agudo, sobre la recuperación de la Memoria espacial en ratas. [Tesis de grado, Maestría en Fisiología]. Bogotá: Facultad de Medicina, Universidad Nacional de Colombia.
Ceulemans H y Bollen M (2004). Functional Diversity of Protein Phosphatase-1, a Cellular Economizer and Reset Button. Physiol Rev. 84: 1–39.
Chao CC, Ma YL, Lee EH (2007). Protein kinase CK2 impairs spatial memory formation through differential cross talk with PI-3 kinase signaling: activation of Akt and inactivation of SGK1. J Neurosci. 6; 27(23): 6243-8.
Chen, R.-H, C. Sarnecki, J. Blenis (1992). Nuclear localization and regulation of the erk- and rsk-encoded protein kinases. Mol. Cell. Biol. 12: 915-27.
Chen, R.-H, C. Abate y J. Blenis (1993). Phosphorylation of the c-Fos transrepression domain by mitogen-activated protein kinase and 90-kDa ribosomal S6 kinase. Proc. Natl. Acad. Sci. USA 90:10952-56.
Chen J, Aguilera G (2010). Vasopressin Protects Hippocampal Neurones in Culture Against Nutrient Deprivation or Glutamate-Induced Apoptosis. J Neuroendocrinol. 22: 1072-81.
Chien WL, Liang KC, Teng CM, Kuo SC, Lee FY, Fu WM (2005). Enhancement of learning behaviour by a potent nitric oxide-guanylate cyclase activator YC-1. Eur J Neurosci. 21; 6: 1679-88.
Cohen P (1989).The structure and regulation of protein phosphatases. Annu Rev Biochem; 58: 453-508.
Cohen P (1991). Targeting subunits of protein phosphatase 1. Methods Enzymol. 99:389-427.
Cohen PTW (1997). Novel protein serine/threonine phosphatases: variety is the spice of life. Trends Biochen Sci. 22: 245-51.
Cordero MI, Sandi C (1998). A role for brain glucocorticoid receptors in contextual fear conditioning: dependence upon training intensity. Brain Res. 9; 786(1-2): 11-7.
Davis S, Laroche S (2006). Mitogen-activated protein kinase/extracellular regulated kinase signalling and memory stabilization: a review. Genes Brain Behav 5 [Suppl 2]:61–72.

138
Denton, R.M, and Tavaré, J.M (1995). Does MAP kinase have a role in insulin action? The case for and against. Eur. J. Biochem. 227: 597-611. de Kloet ER, Joels M, Holsboer F (2005). Stress and the brain: from adaptation to disease. Nat Rev Neurosci. 6: 463-4 de Quervain D, Roozendaal B, McGaugh JL (1998). Stress and glucocorticoids impair retrieval of long-term spatial memory. Nature. 394: 787-90. Diamond DM, Fleshner M, Ingersoll N, Rose GM (1996). Psychological stress impairs spatial working memory: relevance to electrophysiological studies of hippocampal function. Behav Neurosci. 110; 4: 661-72. Diamond DM, Park CR, Heman KL, Rose GM (1999). Exposing rats to a predator impairs spatial working memory in the radial arm water maze. Hippocampus. 9; 5: 542-52. Diamond DM, Kim JJ (2002).The stressed hippocampus, synaptic plasticity and lost memories. Nature. 3:453-62. Dineley, K.T, Hogan, D, Ru, W, Taglialatela, G (2007). Acute inhibition of calcineurin restores associative learning and memory in Tg2576 APP transgenic mice. Neurobiol. Learn. Mem. 88; 2: 217-24. Dwivedi Y, Mondal AC, Shukla PK, Rizavi HS, Lyons J (2004). Altered protein kinase a in brain of learned helpless rats: effects of acute and repeated stress. Biol Psychiatry. 56; 1: 30-40. Dudai Y (2002). The neurobiology of consolidations or, howstable is the engram? Annu Rev Psychol. 55:51–86. Elliott-Hunt Caroline R., Kazlauskaite Jurate, Wilde Geraint J. C., Grammatopoulos Dimitris K. and Hillhouse Edward W (2002). Potential signalling pathways underlying corticotrophin-releasing hormone-mediated neuroprotection from excitotoxicity in rat hippocampus. J Neurochem. 80: 416-25. Eto, M, Kitazawa T, Brautigan D. (2004). Phosphoprotein inhibitor CPI-17 specificity depends on allosteric regulation of protein phosphatase-1 by regulatory subunits. Proc. Natl. Acad. Sci. U. S. A. 101; 8888–93.

139
Fiore, R. S, Bayer, V. E, Pelech, S. L, Posada, J, Cooper, J. A, Baraban, J. M (1993). p42 mitogen-activated protein kinase in brain: prominent localization in neuronal cell bodies and dendrites. Neuroscience, 55, 463–72. Flood, D. G, Finn, J. P, Walton, K. M, Dionne, C. A, Contreras, P. C, Miller, M. (1998). Immunolocalization of the mitogenactivated protein kinases p42MAPK and JNK1, and their regulatory kinases MEK1 and MEK4, in adult rat central nervous system. J comp Neurol. 398: 373–92. Fuentes Lida Q, Reyes Carlos E, Sarmiento José M, Villanueva Carolina I, Figueroa Carlos D, Navarro Javier, and González Carlos B (2008). Vasopressin up-regulates the expression of growth-related immediate-early genes via two distinct EGF receptor transactivation pathways. Cell Signal. 20; 9: 1642–50. García Pérez Hilda Marilín (2000). Electroforesis en geles de poliacrilamida: fundamentos, actualidad e importancia. Univ Diag.1; 2: 31-41. Genoux D, Haditsch U, Knobloch M, Michalon A, Storm D, Mansuy IM (2002). Protein phosphatase 1 is a molecular constraint on learning and memory. Nature. 418; 6901: 970-5. Gerges NZ, Aleisa AM, Schwarz LA, Alkadhi KA (2004). Reduced basal CaMKII levels in hippocampal CA1 region: possible cause of stress-induced impairment of LTP in chronically stressed rats. Hippocampus. 14; 3: 402-10. Giovannini MG (2006). The role of the extracellular signal-regulated kinase pathway in memory encoding. Rev Neurosci. 17:619-34. Gooney M, Shaw K, Kelly A, O'Mara SM, Lynch MA (2002). Long-term potentiation and spatial learning are associated with increased phosphorylation of TrkB and extracellular signal-regulated kinase (ERK) in the dentate gyrus: evidence for a role for brain-derived neurotrophic factor. Behav Neurosci. 116; 3: 455-63. Gräff Johannes, Kyoko Koshibu, Anne Jouvenceau, Patrick Dutar and Isabelle M. Mansuy (2010). Protein phosphatase 1-dependent transcriptional programs for long-term memory and plasticity. Learn Mem. 17: 355-63. Haddad JJ (2005). N-methyl-D-aspartate (NMDA) and the regulation of mitogen-activated protein kinase (MAPK) signaling pathways: a revolving neurochemical axis for therapeutic intervention? Prog Neurobiol. 77: 252-82.

140
Halpain, S. and Greengard, P. (1990). Activation of NMDA receptors induces rapid dephosphorylation of the cytoskeletal protein MAP2. Neuron. 5: 237-46. Hanks SK, Hunter T (1995). Protein kinases 6. The eukaryotic protein kinase superfamily: kinase (catalytic) domain structure and classification. FASEB J. 9: 576-96. Havekes R, Nijholt IM, Luiten PG, Van der Zee EA (2006). Differential involvement of hippocampal calcineurin during learning and reversal learning in a Y-maze task. Learn Mem. 13; 6: 753-9. He J, K. Yamada, L.-B. Zou, and T. Nabeshima (2001). Spatial memory deficit and neurodegeneration induced by the direct injection of okadaic acid into the hippocampus in rats. J Neural Transm.108: 1435-43. Hebert AE, Dash PK (2004). Nonredundant roles for hippocampal and entorhinal cortical plasticity in spatial memory storage. Pharmacol Biochem Behav. 79; 1: 143-53. Hedou GF, Koshibu K, Farinelli M, Kilic E, Gee CE, Kilic U, Baumgartel K, Hermann DM, Mansuy IM (2008). Protein phosphatase 1-dependent bidirectional synaptic plasticity controls ischemic recovery in the adult brain. J Neurosci. 28: 154–62. Herman JP, Cullinan WE. (1997). Neurocircuitry of stress: central control of the hypothalamo-pituitary-adrenocortical axis. Trends Neurosci. 20; 2: 78-84.
Hunter T (1998). The phosphorylation of protein on tyrosine: its role in cell growth and disease. Harvey Lect. 94: 81-119.
Impey S, Obrietan K, Wong ST, Poser S, Yano S, Wayman G, Deloulme JC, Chan G, Storm DR (1998a). Cross talk between ERK and PKA is required for Ca 21 stimulation of CREB-dependent transcription and ERK nuclear translocation. Neuron 21:869-83. Ito k. Barnes PJ, Adcock IM (2000). Glucocorticoid receptor recruitment of histone deacetylase 2 inhibits IL-1β-induced histone H4 acetylation on lysines 8 and 12. Mol Cell Biol. 20: 6891-6903. Izquierdo I, Medina JH (1997). Memory formation: the sequence of biochemical events in the hippocampus and its connection to activity in other brain structures. Neurobiol Learn Mem. 68: 285–316.

141
Izquierdo LA, Vianna M, Barros DM, Mello e Souza T, Ardenghi P, Sant' Anna MK, Rodrigues C, Medinam JH, Izquierdo I (2000). Short- and long-term memory are differentially affected by metabolic inhibitors given into hippocampus and entorhinal cortex. Neurobiol Learn Mem. 73; 2: 141-9.
Izquierdo I, BevilaquaLRM,Rossato JI, Bonini J, Medina JH, Cammarota M (2006). Different molecular cascades in different sites of the brain control consolidation. Trends Neurosci. 29: 496-505.
Jacobson L, Sapolsky R (1991). The Role of the Hippocampus in Feedback Regulation of the Hypothalamic-Pituitary-Adrenocortical Axis. Endocr Rev.12; 2: 118-34.
Jeansok J. Kim and Kenneth S. Yoon (1998). Stress: metaplastic effects in the hippocampus. Trends Neurosci. 21; 505-09. Joëls Marian, Henk Karst, Roel DeRijk, y de Kloet ER. (2008). The coming out of the brain mineralocorticoid receptor. Trends Neurosci.31; 1: 1-7.
Jouvenceau A, Hédou G, Potier B, Kollen M, Dutar P, Mansuy IM (2006). Partial inhibition of PP1 alters bidirectional synaptic plasticity in the hippocampus. Eur J Neurosci. 24; 2: 564-72.
Kim J, Lee H, Han J Packard M. G (2001). Amygdala Is Critical for Stress-Induced Modulation of Hippocampal Long-Term Potentiation and Learning. J NeuroscI. 21; 14: 5222-28.
Kim J.J, Song E.Y and Kosten T.A (2006). Stress effects in the hippocampus: synaptic plasticity and memory, Stress. 9: 1–11.
Koshibu Kyoko, Gräff Johannes, Beullens Monique, Heitz Fabrice D, Berchtold Dominik, Russig Holger, Farinelli Mélissa, Bollen Mathieu, Mansuy Isabelle M (2009). Protein phosphatase 1 regulates the histone code for long-term memory. J Neurosci. 29; 41:13079-89.
Koshibu K, Gräff j, Mansuy IM (2011). Nuclear protein phosphatase-1: an epigenetic regulator of fear memory and amygdala long-term potentiation. Neuroscience. 173():30-6.
Koya E, Spijker S, Homberg JR, Voorn P, Schoffelmeer AN, De Vries TJ, Smit AB (2005). Molecular reactivity of mesocorticolimbic brain areas of high and low

142
grooming rats after elevated plus maze exposure. Brain Res Mol Brain Res. 137; 1-2: 184-92.
Krugers Harmen J, Hoogenraad Casper C y Laurent Groc. (2010). Stress hormones and AMPA receptor trafficking in synaptic plasticity and memory. Nat Rev Neurosci. 11: 675-81.
Lee CT, Tyan SW, Ma YL, Tsai MC, Yang YC, Lee EH (2006). Serum- and glucocorticoid-inducible kinase (SGK) is a target of the MAPK/ERK signaling pathway that mediates memory formation in rats. Eur J Neurosci. 23; 5: 1311-20. Lee CT, Ma YL, Lee EH (2007). Serum- and glucocorticoid-inducible kinase1 enhances contextual fear memory formation through down-regulation of the expression of Hes5. J Neurochem. 100; 6: 1531-42. Li FC, Chan JY, Chan SH, Chang AY (2005). In the rostral ventrolateral medulla, the 70-kDa heat shock protein (HSP70), but not HSP90, confers neuroprotection against fatal endotoxemia via augmentation of nitric-oxide synthase I (NOS I)/protein kinase G signaling pathway and inhibition of NOS II/peroxynitrite cascade. Mol Pharmacol. 68; 1: 179-92. Lin CH, Yeh SH, Lu HY, Gean PW (2003). The similarities and diversities of signal pathways leading to consolidation of conditioning and consolidation of extinction of fear memory. J Neurosci. 23; 23: 8310-7. Lisman, J.E (1989). A mechanism for the Hebb and anti-Hebb Processes Underlying Learning and Memory. Proc. Natl. Acad. Sci. 86:9574-78. López-Calderón A. (1999). Estrés. En: Fernandez-Tresguerres JA (Ed.). Fisiología Humana. Segunda Edición. Madrid: McGraw-Hill Interamericana de España. pp: 1127-1136. Luine VN, Spencer RL, McEwen BS (1993). Effects of chronic corticosterone ingestion on spatial memory performance and hippocampal serotonergic function. Brain Res. 616; 1-2: 65-70. Lu Y.-F, Tomizawa K, Moriwaki A, Hayashi Y, l M. Tokuda, T. Itano, O. Hatase, H. Matsui (1996). Calcineurin inhibitors, FK506 and cyclosporin A, suppress the NMDA receptor-mediated potentials and LTP, but not depotentiation in the rat hippocampus. Brain Res. 729: 142-46.

143
Maeda T, Yoshimatsu T, Hamabe W, Fukazawa Y, Kumamoto K, Ozaki M, Kishioka S (2006). Involvement of serine/threonine protein phosphatases sensitive to okadaic acid in restraint stress-induced hyperlocomotion in cocaine-sensitized mice. Br J Pharmacol. 148; 4: 405-12. Malenka R.C (2003). Synaptic plasticity and AMPA receptor trafficking. Ann N Y Acad Sci. 1003: 1-11. Malleret G, Haditsch U, Genoux D, Jones MW, Bliss TV, Vanhoose AM, Weitlauf C, Kandel ER, Winder DG, Mansuy IM (2001). Inducible and reversible enhancement of learning, memory, and long-term potentiation by genetic inhibition of calcineurin. Cell 104:675–86. Mansuy, I.M, Winder DG, Moallem TM, Osman M, Mayford M, Hawkins RD, Kandel ER (1998). Inducible and reversible gene expression with the rtTA system for the study of memory. Neuron. 21:257-65. Mansuy Isabelle (2003). Calcineurin in memory and bidirectional plasticity. Biochem biophys Res Commun. 311; 4:1195-208. Mansuy Isabelle M, Shenolikar Shirish (2006). Protein serine/threonine phosphatases in neuronal plasticity and disorders of learning and memory. Trends Neurosci. 29; 12: 679-86. McAvoy T, Allen P.B, Obaishi H, Nakanishi H, Takai Y, Greengard P, Nairn A.C, Hemmings H.C (1999). Regulation of neurabinI interaction with protein phosphatase 1 by phosphorylation. Biochemistry. 38: 12943–49. McAvoya Thomas, Zhoua Ming-ming, Paul Greengarda, Angus C. Nairna (2009). Phosphorylation of Rap1GAP, a striatally enriched protein, by protein kinase A controls Rap1 activity and dendritic spine morphology. Proc Natl Acad Sci U S A. 106; 9: 3531–36. McEwen BS, Albeck D, Cameron H, Chao HM, Gould E, Hastings N, Kuroda Y, Luine V, Magarinos AM, McKittrick CR (1995). Stress and the brain: a paradoxical role for adrenal steroids. Vitam Horm. 51: 371-402. McLay R N, Reeman S M, Zadina J E (1998). Chronic Corticosterone Impairs Memory Performance in the Barnes Maze. Physiol Behav. 5: 933-37.

144
McNamara RK, Lenox RH (2004). Acute restraint stress reduces protein kinase C gamma in the hippocampus of C57BL/6 but not DBA/2 mice. Neurosci Lett. 368; 3: 293-6.
Medema RH, Bos JL (1993). Role of P21ras in receptor tyrosine kinase signaling. Crit Rev Oncology. 4: 615-61.
Meller E, Shen C, Nikolao TA, Jensen C, Tsimberg Y, Chen J, Gruen RJ (2003) Region-specific effects of acute and repeated restraint stress on the phosphorylation of mitogen-activated protein kinases. Brain Res. 979: 57–64
Moorhead GB, Trinkle-Mulcahy L, Ulke-Lemee A. (2007). Emerging roles of nuclear protein phosphatases. Nat Rev Mol Cell Biol. 8: 234–44.
Mishkin M (1978). Memory in monkeys severely impaired by combined but not by separate removal of amygdala and hippocampus. Nature. 273; 5660: 297-8.
Monti B, Berteotti C, Contestabile A (2005). Dysregulation of memory-related proteins in the hippocampus of aged rats and their relation with cognitive impairment. Hippocampus. 15; 8: 1041-9.
Morinobu S, Fujimaki K, Kawano K, Tanaka K, Takahashi J, Ohkawa M, Yamawaki S, Kato N. (2003). Influence of immobilization stress on the expression and phosphatase activity of protein phosphatase 2A in the rat brain. Biol Psychiatry. 54; 10: 1060-6.
Morris R (1984). Developments of a water-maze procedure for studying spatial learning in the rat. J Neurosci Methods. 11; 1: 47-60.
Morishita W, Connor JH, Xia H, Quinlan EM, Shenolikar S, Malenka RC (2001). Regulation of synaptic strength by protein phosphatase 1. Neuron 32: 1133–48. Munton RP,Vizi S,Mansuy IM (2004). The role of protein phosphatase-1 in a modulation of synaptic and structural plasticity. FEBS Letters 567: 121–28. Nishida Y, Nagata T, Takahashi Y, Sugahara-Kobayashi M, Murata A, Asai S (2004). Alteration of serum/glucocorticoid regulated kinase-1 (sgk-1) gene expression in rat hippocampus after transient global ischemia. Brain Res Mol Brain Res. 123; 1-2: 121-5. Olijslagers J. E, de Kloet E. R, Elgersma Y, van Woerden G. M, Joëls M, Karst H (2008). Rapid changes in hippocampal CA1 pyramidal cell function via pre- as

145
well as postsynaptic membrane mineralocorticoid receptors. European Journal of Neuroscience. 27: 2542–50.
Olton DS (1977). Spatial memory. Sci Am. 236; 6: 89-94.
Olton DS (1979). Mazes, maps, and memory. Am Psychol. 34; 7: 583-96.
Pacák K, Palkovits M (2001). Stressor Specificity of Central Neuroendocrine Responses: Implications for Stress- Related Disorders. Endocr Rev. 22; 4: 502- 54. Pedreira, M.E, Maldonado, H (2003). Protein synthesis subserves reconsolidation or extinction depending on reminder duration. Neuron 38: 863-69. Qi X, Lin W, Li J, Pan Y, Wang W (2006). The depressive-like behaviors are correlated with decreased phosphorylation of mitogen-activated protein kinases in rat brain following chronic forced swim stress. Behav Brain Res. 175; 2: 233-40. Qiu, J, Lou, L. G, Huang, X. Y, Lou, S. J, Pei, G, Chen, Y. Z (1998). Nongenomic mechanisms of glucocorticoid inhibition of nicotine-induced calcium influx in PC12 cells: Involvement of protein kinase C. Endocrinology. 139: 5103–08. Qiu Jian, Wang Ping, Jing Qing, Zhang Wenbo, Li Xiaoyu, Zhong Yongping, Sun Gang, Pei Gang, and Chen Yizhang (2001). Rapid Activation of ERK1/2 Mitogen-Activated Protein Kinase by Corticosterone in PC12 Cells. Biochem Biophys Res Commun. 287: 1017-24. Radulovic J, Ruhmann A, Liepold T, Spiess J (1999). Modulation of learning and anxiety by corticotropin-releasing factor (CRF) and stress: differential roles of CRF receptors 1 and 2. J Neurosci. 19: 5016-25. Ramos JW (2008) The regulation of extracellular signal-regulated kinase (ERK) in mammalian cells. Int J Biochem Cell Biol. 40:2707–19 Revest Jean-Michel, Di Blasi Francesco, Kitchener Pierre, Rouge´-Pont Françoise, Desmedt Aline, Turiault Marc, Tronche François, Pier Vincenzo Piazza (2005). The MAPK pathway and Egr-1 mediate stress-related behavioral effects of glucocorticoids. Nat Neurosci. 8; 5: 664-72.

146
Revest J-M, Kaouane N, Mondin M, Le Roux A, Rougé F -Pont, Vallée M, Barik J, Tronche F, A Desmedtu and Piazza PV (2010). The enhancement of stress-related memory by glucocorticoids depends on synapsin-Ia/Ib. Mol Psychiatry. 15(12):1125, 1140-5. Roberson Erik D, English Joey D, Adams J. Paige, Selcher Joel C, Kondratick Christine and Sweatt J. David. (1999). The Mitogen-Activated Protein Kinase Cascade Couples PKA and PKC to cAMP Response Element Binding Protein Phosphorylation in Area CA1 of Hippocampus. J Neurosci. 19; 11: 4337-48. Roozendaal B, McGaugh JL (1996). Amygdaloid nuclei lesions differentially affect glucocorticoid-induced memory enhancement in an inhibitory avoidance task. Neurobiol Learn Mem. 65: 1- 8. Roozendaal, B., & McGaugh, J. L (1997a). Glucocorticoid receptor agonist and antagonist administration into the basolateral but not central amygdala modulates memory storage. Neurobiol Learn Mem. 67: 176-79. Roozendaal, B., & McGaugh J. L (1997b). Basolateral amygdala lesions block the memory-enhancing effect of glucocorticoid administration in the dorsal hippocampus of rats. Eur J Neurosci. 9: 76-83. Roozendaal B, de Quervain D J, Ferry B, Setlow B, McGaugh JL (2001). Basolateral Amygdala-Nucleus Accumbens Interactions in Mediating Glucocorticoid Enhancement of Memory Consolidation. J Neurosci. 21; 7: 2518-25. Roozendaal Williams CL, McGaugh JL (1999). Glucocorticoid receptor activation in the rat nucleus of the solitary tract facilitates memory consolidation: Involvement of the basolateral amygdala. Eur J Neurosci. 11:1317-23. Roozendaal B. (2002). Stress and Memory: Opposing Effects of Glucocorticoids on Memory Consolidation and Memory Retrieval. Neurobiol Learn Mem. 78; 3: 578-95. Roozendaal B (2003). Systems mediating acute glucocorticoid effects on memory consolidation and retrieval. Prog Neuropsychopharmacology Biol Psychiatry 27; 8: 1213-23. Roper JA, O’Carroll A-M, Young WS and Lolait SJ (2010). The vasopressin Avpr1b receptor: Molecular and pharmacological studies. Stress. 1-18.

147
Runyan Jason D, Moore Anthony N y Dash Pramod K. (2005). A role for prefrontal calcium-sensitive protein phosphatase and kinase activities in working memory. Learn Mem.12: 103-10. Sananbenesi Farahnaz, Fischer André, Schrick Christina, Spiess Joachim, Radulovic Jelena (2003). Behavioral/Systems/Cognitive Mitogen-Activated Protein Kinase Signaling in the Hippocampus and Its Modulation by Corticotropin-Releasing Factor Receptor 2: A Possible Link between Stress and Fear Memory. J Neurosci. 23; 36: 11436 –43. Sandi, C, Woodson, J.C, Haynes, V.F, Park, C.R, Touyarot, K, Lopez-Fernandez, M.A, Venero, C, Diamond, D.M (2005). Acute stress-induced impairment of spatial memory is associated with decreased expression of neural cell adhesion molecule in the hippocampus and prefrontal cortex. Biol. Psychiatry. 57; 8: 856-64. Sasaguri K, Kikuchi M, Hori N, Yuyama N, Onozuka M, Sato S (2005). Suppression of stress immobilization-induced phosphorylation of ERK 1/2 by biting in the rat hypothalamic paraventricular nucleus. Neurosci Lett. 383; 1-2: 160-4. Satoh Eiki, Shimeki Shusuke (2010). Acute Restraint Stress Enhances Calcium Mobilization and Glutamate Exocytosis in Cerebrocortical Synaptosomes from Mice. Neurochem Res. 35:693–701 Schoenebeck B, Bader V, Zhu XR, Schmitz B, Lübbert H, Stichel CC (2005). Sgk1, a cell survival response in neurodegenerative diseases. Mol Cell Neurosci. 30; 2: 249-64. Selye, H. A (1936). Syndrome produced by diverse nocuous agents. Nature. 138: 32. Selcher, J. C, Atkins, C. M, Trzaskos, J. M, Paylor, R, Sweatt, J.D (1999). A necessity for MAP kinase activation in mammalian spatial learning. Learning and Memory. 6: 478–90. Sharifzadeh M, Sharifzadeh K, Naghdi N, Ghahremani MH, Roghani A (2006). Posttraining intrahippocampal infusion of a protein kinase AII inhibitor impairs spatial memory retention in rats. Pharmacol Biochem Behav. 85; 4: 705-12.

148
Sharifzadeh M, Zamanian AR, Gholizadeh S, Tabrizian K, Etminani M, Khalaj S, Zarrindast MR, Roghani A (2007). Post-training intrahippocampal infusion of nicotine-bucladesine combination causes a synergistic enhancement effect on spatial memory retention in rats. Eur J Pharmacol. 562; 3: 212-20. Shen CP, Tsimberg Y, Salvadore C, Meller E (2004). Activation of Erk and JNK MAPK pathways by acute swim stress in rat brain regions. BMC Neurosci. 5; 1: 36. Shimizu Y, Sugama S, Degiorgio L.A, CHO B. P and Joh T.H. (2004). Cell-type specific signal transduction and gene regulation via mitogen activated protein kinase pathway in catecholaminergic neurons by restraint stress. Neuroscience 129: 831–39 Shimizu K, Phan T, Mansuy IM, Storm DR (2007). Proteolytic degradation of SCOP in the hippocampus contributes to activation of MAP kinase and memory. Cell. 128; 6: 1219-29. Shors TJ, Weiss C, Thompson RF (1992). Stress-induced facilitation of classical conditioning. Science 24: 537-39. Shors TJ, Elkabes S, Selcher JC, Black IB (1997). Stress persistently increases NMDA receptor-mediated binding of [3H]PDBu (a marker for protein kinase C) in the amygdala, and re-exposure to the stressful context reactivates the increase. Brain Res. 750; 1-2: 293-300. Suenaga T, Morinobu S, Kawano K, Sawada T, Yamawaki S (2004). Influence of immobilization stress on the levels of CaMKII and phospho-CaMKII in the rat hippocampus. Int J Neuropsychopharmacol. 7; 3: 299-309. Sweatt JD (2001).The neuronal MAP kinase cascade: a biochemical signal integration system subserving synaptic plasticity and memory. J Neurochem. 76: 1-10. Sweatt JD (2004). Mitogen-activated protein kinases in synaptic plasticity and memory. Curr Opin Neurobiol; 14:311-17. Takahashi J, Tanaka K, Morinobu S, Fujimaki K, Li ST, Kato K, Ohkawa M, Yamawaki S, Kato N (2001). Influence of restraint stress on the expression and the serine/threonine phosphatase activity of calcineurin in the rat brain. Synapse. 40; 2: 130-6.

149
Tasker Jeffrey G, Shi Di, Renato Malcher-Lopes (2006). Rapid Glucocorticoid Signaling via Membrane-Associated Receptors. Endocrinology. 147; 12: 5549-56.
Tian Q, Lin ZQ, Wang XC, Chen J, Wang Q, Gong CX, Wang JZ (2004). Injection of okadaic acid into the Meynert nucleus basalis of rat brain induces decreased acetylcholine level and spatial memory deficit. Neuroscience. 126; 2: 277-84. Thorpe WH (1963). Learning and instinct in animals. London: Methuen. Tolbert Lara M., Russell David S., and Duman Ronald S (2003). Norepinephrine Activates Extracellular-Regulated Kinase in Cortical Neurons. Biol. Psychiatry. 54:983–993. Torres A (2006). Efectos del estrés agudo por restricción motora sobre la recuperación y la extinción de una tarea espacial en ratas. [Trabajo de grado]. Bogotá: Facultad de Ciencias Humanas, Universidad Nacional de Colombia. Tsai KJ, Chen SK, Ma YL, Hsu WL, Lee EH (2002). Sgk, a primary glucocorticoid- induced gene, facilitates memory consolidation of spatial learning in rats. Proc Natl Acad Sci USA 99:3990 –3995. Utto, M, J.J. Vasterling, K. Brailey, P.B. Sutker (1993). Memory and attention in combat-related posttraumatic-stress-disorder (PTSD). J. Psychopath. Behav. Assess. 15: 43-52. Van der Geer, P., Hunter, T., and Lindberg, R.A (1994). Receptor protein-tyrosine kinases and their signal transduction pathways. Annu Rev Cell Biol. 10: 251-337. Vianna Monica R.M, Coitinho Adriana, Izquierdo Luciana, Izquierdo Ivan (2010). Effects of intrahippocampal administration of the phosphatase inhibitor okadaic acid Dual effects on memory formation. Dement Neuropsychol. 4; 1: 23-7. Volpi Simona, Liu Ying and Aguilera Greti (2006). Vasopressin increases GAGA binding activity to the V1b receptor promoter through transactivation of the MAP kinase pathway. J Mol Endocrinol. 36: 581–90. Waddell S (2003). Protein phosphatase 1 and memory: practice makes PP1 imperfect? Trends Neurosci. 26; 3: 117-9.

150
Wang Z, Hu SY, Lei DL, Song WX (2006). Effect of chronic stress on PKA and P-CREB expression in hippocampus of rats and the antagonism of antidepressors. Zhong Nan Da Xue Xue Bao Yi Xue Ban. 31: 767-71. Wei L, Hubbard SR, Smith RF, Ellis L (1994). Protein kinase superfamily comparisons of sequence data with three-dimensional structure. Curr Op Struct Biol. 4: 450-5. Wen-feng Teng; Wei-min Sun; Li-fei Shi; Dian-dong Hou; Hui Liu (2008). Effects of Restraint Stress on Iron, Zinc, Calcium, and Magnesium Whole Blood Levels in Mice. Biol Trace Elem Res. 121:243–48. Wu J, Song TB, Li YJ, He KS, Ge L, Wang LR (2007). Prenatal restraint stress impairs learning and memory and hippocampal PKCbeta1 expression and translocation in offspring rats. Brain Res. 1141: 205-13. Yang YL, Lu KT (2005). Facilitation of conditioned fear extinction by D-cycloserine is mediated by mitogen activated protein kinase and PI-3 kinase cascades and requires de novo protein synthesis in basolateral nucleus of amygdale. Neuroscience.134: 247–60. Yang YL, Chao PK, Lu KT (2006). Systemic and intra-amygdala administration of glucocorticoid agonist and antagonist modulate extinction of conditioned fear. Neuropsychopharmacology 31: 912–24. Yang Yi-Ling, Chao Po-Kuan, Ro Long-Sun, Yu-Yuan P Wo and Kwok-Tung Lu (2007). Glutamate NMDA Receptors within the Amygdala Participate in the Modulatory Effect of Glucocorticoids on Extinction of Conditioned Fear in Rats. Neuropsychopharmacology. 32: 1042-51. Yamashita T, Inui S, Maeda K, Hua DR, Takagi K, Fukunaga K, Sakaguchi N (2006). Regulation of CaMKII by alpha4/PP2Ac contributes to learning and memory. Brain Res. 1082; 1: 1-10. Yang Chih-Hao, Huang Chiung-Chun, and Hsu Kuei-Sen (2004). Behavioral Stress Modifies Hippocampal Synaptic Plasticity through Corticosterone-Induced Sustained Extracellular Signal-Regulated Kinase/Mitogen-Activated Protein Kinase Activation. J Neurosci. 24; 49:11029-34.
Yasuda Masahiro y Mayford Mark R (2006). CaMKII Activation in the Entorhinal Cortex Disrupts Previously Encoded Spatial Memory. Neuron. 50; 2: 309-18.

151
Zeng H, Chattarji S, Barbarosie M, Rondi-Reig L, Philpot B. D, Miyakawa T, Bear M. F, Tonegawa S (2001). Forebrainspecific calcineurin knockout selectively impairs bidirectional synaptic plasticity and working/episodic–like memory. Cell. 107: 617–29. Zhao W, Bennett P, Sedman GL, Ng KT (2005). The impairment of long-term memory formation by the phosphatase inhibitor okadaic acid. Brain Res Bull. 36: 557-56. Zheng Jun-Fang, Patil Sudarshan S, Wei-Qiang Chen,Wei An, Jun-Qi He, Harald Höger, and Gert Lubec (2009). Hippocampal Protein Levels Related to Spatial Memory Are Different in the Barnes Maze and in the Multiple T-Maze. J Proteome Res. 8: 4479–86. Ziemiecki A, Harpur AG, Wilks AF (1994). JAK protein tyrosine kinases: their role in cytokine signalling. Trends Cell Biol. 4: 207-12. Zorrilla EP, Schulteis G, Ormsby A, Klaassen A, Ling N, McCarthy JR, Koob GF, De Souza EB (2002). Urocortin shares the memory modulating effects of corticotropin-releasing factor (CRF): mediation by CRF1 receptors. Brain Res. 952: 200-10.