epithelial transport of drugs in cell culture. ii: effect of extracellular calcium concentration on...
TRANSCRIPT

Epithelial Transport of Drugs in Cell Culture. II: Effect of Extracel I u lar Calcium Concent rat ion on the Paracel I u lar Transport of Drugs of Different Lipophilicities across Monolayers of Intestinal Epithelial (Caco-2) Cells
PER ARTURSSON~ AND CHRISTINE MAGNUSSON Received June 5,. 1989, from the Department of Pharmaceutics, Uppsala University, BMC, Box 580, S-751 23 Uppsala, Sweden. Accepted for publication September 18, 1989.
Abstract 0 A human intestinal cell line, Caco-2, was used as a model to study the passive diffusion of a homologous series of drugs (p-blocking agents) of different lipophilicity across intestinal epithelium. The perme- ability of the Cam-2 monolayers was modulated by the use of a calcium switch assay. The transmembrane resistance could be reversibly de- creased from -280 ohms - cm2 (a resistance similar to that of colon epithelium) to -60 ohms.cm2 (a resistance similar to that of small intestine epithelium). Transmission electron microscopy showed that the increased electrical permeability was caused by a reversible separation of the components of the junctional complex and not by cell detachment. In general, the increased paracellular permeability resulted in a 2- to 9-fold increase in the apparent permeability coefficients for the more hydrophilicdrugs(e.g.,from0.20 f 0.010 x lO-’to 1.43 k 0.185 x cm/s for atenolol), while the transport parameters for the more lipophilic drugs remained unchanged (e.g., 43.03 2 3.64 x lO-’and 46.10 f 3.25 x cm/s for propranolol). These findings indicate that it is possible to study the contribution of the paracellular pathway to the transport of drugs in the Caco-2 model.
The epithelial cells lining the gastrointestinal tract are a major barrier to absorption of orally administered drugs.1 Drugs can pass this barrier by two parallel routes: the transcellular route (across the cell membranes) and the paracellular route (through the intercellular spaces). Most drugs are relatively lipophilic and partition rapidly from the luminal fluid into the cell membranes. These drugs are normally completely absorbed from the gastrointestinal tract.2 However, many hydrophilic drugs will not partition into the cell membranes and are consequently excluded fmm this pathway.’
The alternative paracellular pathway occupies a very small surface area as compared with the cell membranes and is sealed by the junctional complex.3 It has been shown that the aqueous pores in the junctional complex have a mean diam- eter of only a few Angstrom units, which theoretically limits the intercellular transport to molecules that are much smaller than drugs.4 However, accumulating findings indi- cate that the intercellular junctions of different epithelia are dynamic structures that can modulate their permeability in response to various stimuli such as hormones and nutri- ents.3.5.6 In addition, the integrity of the junctional complex requires the presence of calcium ions.7-8 The reversible open- ing of intercellular spaces is of considerable interest in pharmaceutics since it shows a way by which the absorption of more hydrophilic drugs (that do not partition significantly into the cell membranes) can be increased.
An experimental separation of the two parallel pathways would allow systematic studies on the influence of different physicochemical properties, such as lipophilicity, on the drug transport across each of the pathways. However, studies on the relative importance of the transcellular and paracellular
pathways are limited due to the lack of suitable experimental models.9 In this paper, we use a new cell culture model10-12 to study the transepithelial transport of a series of drugs under conditions where the junctional complex is reversibly opened and closed. P-Adrenoceptor antagonists were chosen as model drugs since they represent a homologous series of clinically relevant drugs that covers a broad range of different li- pophilicities. In addition, these drugs are absorbed across Caco-2 monolayers by passive diffusion.10 In general, the results indicate that the permeability of the paracellular pathway has an influence on the transport of hydrophilic drugs, while this pathway is of limited or no importance for the transport of more lipophilic drugs that are taken up rapidly by the transcellular route.
Experimental Section Drugs and Radiolabeled Markerp3H-Labe1ed and unlabeled
atenolol (specific radioactivity 0.627 nCi/nmol); H216/44 [(S)- 4-hydroxy-N-~2-(~2-hydroxy-3-~4-~2-~2-~cyclopropylmethoxy)eth- oxy)ethyl)phenoxy)-propyl)amino)ethyl)-l-piperidinecarboxamide; specific radioactivity 16.7 pCilnmol1, alprenolol (specific radioactiv- ity 0.252 nCi/nmol), and metoprolol (specific radioactivity 0.475 pcilnmol) were generous gifts from Dr. Kurt-Jorgen Hoffman, Hassle AB, Giiteborg, Sweden, as was 14C-labeled practolol (2.1 nCi/nmol). [3H]Propranolol (26.6 Ci/mmol) and [14C]polyethyleneglycol (MW 4000; specific radioactivity 15.0 Ci/mg) were obtained from New England Nuclear, Boston, MA. The radiolabeled compounds had a radiochemical purity of 9699%. Propranolol and alprenolol were purchased from Sigma Chemical, St. Louis, MO.
Octanol-water distribution coefficients (D) were kindly provided by Dr. Kurt-Jorgen Hoffman, Hassle AB, Goteborg, Sweden, and have also been published elsewhere.1”15
Cells-Caco-2 cells, originating from a human colorectal carcinoma16 were obtained from American Tissue Culture Collection, Rockville, MD. Cells of passage numbers 85-95 were used. The cells were maintained in Dulbecco’s modified Eagle medium (DMEM) containing 10% fetal calf serum, 1% nonessential amino acids, benzylpenicillin (100 U/mL), and streptomycin (10 pg/mL). All tissue culture media were obtained from Gibco through Laboratorie Design AB, Lidingo, Sweden. The medium was changed every second day. The cells were mycoplasma negative as determined with Hoechst 33258.17
The cells were added to polycarbonate membrane culture plate inserts (Nucell; pore size 0.4 pm; diameter 24 mm; Nuclepore, Pleasanton, CA) as previously described.9 The inserts were placed in tissue culture wells (Costar, Badhoevedorp, The Netherlands; cat. no. 3506) and 2 x lo6 cells were added to each insert. The cells were allowed to grow and differentiate for up to 30 days. After this time, Caco-2 monolayers may start to detach from their support.18
Integrity of the Monolayera-The integrity of the monolayers was determined by measurement of the potential difference over the cells, by light microscopy, and by following the transepithelial transport of a macromolecular marker, polyethyleneglycol (MW 4000), as de- scribed previously.1o-12 The potential difference was expressed as transmembrane resistance (ohmsicm? after subtraction of the in-
0022-3549/90/0700-0595$0 1.00/0 0 1990, American Pharmaceutical Association
Journal of Pharmaceutical Sciences / 595 Vol. 79, No. 7, July 1990

trinsic resistance of the model (i.e., the resistance obtained over cell free inserts).l0.19
Electron Microscopy-Transmission electron microscopy was per- formed essentially as previously described.20 The filters were washed in phosphate-buffered saline (pH 7.2) and fixed in a solution contain- ing 2.0% glutaraldehyde and 0.1 M sucrose in 0.1 M cacodylate buffer (pH 7.3) for 1 h a t 25 "C. The cells were rinsed in 0.1 M phosphate- buffered saline and fixed with 1% osmium tetroxide in 0.1 M sodium cacodylate * HCI buffer for 1 h a t 25 "C. The filters were rinsed in 0.1 M phosphate-buffered saline and dehydrated in ethanol, infiltrated, embedded in Epon resin, and cured in an oven at 60 "C for 48 h. The preparations were sectioned in a microtome, stained with uranyl acetate and lead citrate, and examined and photographed on a transmission electron microscope (Philips 201).
Scanning electron microscopy was performed as previously described.21 The cells were examined and photographed in a trans- mission electron microscope (Jeol 100-0 equipped with a side entry goniometer and a scanning attachment.
Meaeurements of Drug Transport-Drug solutions were pre- pared from the radiolabeled isotopes and the corresponding unlabeled compounds in ethanol to give final concentrations of 1 x 10 M. The ethanol concentration in the cell culture medium was usually -0.05% and never exceeded 0.25%. Control experiments, including measure- ments of transmembrane resistance, [l4C1PEG transport, light mi- croscopy, and transmission electron microsropy showed that the monolayers were unaffected by this concentration of ethanol.10
All transport experiments were performed in air at 95% relative humidity and 37 "C in serum-free medium (DMEM; pH 7.3) contain- ing 1% nonessential amino acids, 10 mM HEPES buffer, and 0.1% human serum albumin. The monolayers were 14-30 days old. Under these conditions, the integrity of the monolayers was intact for a t least 6 h. The monolayers were agitated on a microscope slide mixer (Relax 3, Kebo Lab, Stockholm, Sweden) a t 10 rpm and a 2.5" angle. The radiolabeled drug solutions were added to the apical side of the cells and the samples were mixed for 30 s. At this time, the initial concentration (C,) in the donor chamber was determined from a 50- or 100-pL sample. (No radioactivity was found in the receiving chamber after 30 s.) Subsequently, a maximum of five samples (50-100 pL) were taken from each chamber at regular time intervals.10 Some experiments were performed in low calcium me- dium, that is, after treatment with medium containing 2.5 mM EGTA [ethylene glycol-bis-(B-aminoethyl etherUV,N,iV' ,"-tetraacetic acid; Sigma, St. Louis, MO; purity: -97%1 for 45 min.
The integrity of the monolayers was checked a t the end of each experiment by measurement of the transmembrane resistance. The monolayers were then washed five times with phosphate-buffered saline and the radioactivity of the polycarbonate filters was deter- mined. The results were corrected for dilution and expressed as the concentration C a t time t .
All rate constants were obtained under "sink" conditions (i.e., before > 10% of the drug had been transported) from the linear drug appearance curves in the receiving chambers. (No disappearance slopes could be obtained for the hydrophilic drugs that were trans- ported slowly across the monolayers.) By the use of "s ink conditions, the influence of drug diffusing back from the receiving to the donor chamber could be minimized.
The apparent permeability coefficient Papp) was determined ac- cording to eq 1:1:3
mo
100
(cm x s - l ) - AQ - At x 60 x A x C,
-
-
(1)
where AQlAt is the permeability rate (pg X min '), C, the initial concentration in the donor chamber (fig x mL '1, and A is the surface area of the membrane (cm2).10
Treatment with a Selective Calcium Chelator-The selective calcium chelator EGTA was used to bind free extracellular Ca2' ions. All EGTA experiments were performed under the same conditions as the drug transport studies. The serum-free medium was supple- mented with 2.5 mM EGTA. The EGTA-containing medium was added to the two filter chambers for up to 120 min (usually 45 mid. When appropriate, the EGTA medium was exchanged to serum-free medium (containing 1.8 mM Ca") and the cells were incubated for another 2 4 h. The integrity ofthe monolayers was checked at the end of each experiment by measurement of transmembrane resistance.
Results Effect of Low Extracellular Ca2+ on Monolayer Integri-
ty-Initial experiments showed that the minimal effective concentration of EGTA on the transmembrane resistance was 2.5 mM. This corres onds to an extracellular Ca2+ concen- tration of 2-4 x 10- M.22 (A more precise calculation of free Ca2 could not be made since the binding constant of Ca2+ to EGTA in the complex cell culture medium is different than the published association constants.) This Concentration is at least 10 times larger than the normal intracellular calcium concentration which is in the 100 nM range.23
A gradual decrease in transmembrane resistance with time was observed when the monolayers were exposed to low Ca2 +
medium (containing 2.5 mM EGTA), as shown in Figure 1. The decrease plateaued after -45 min (i.e., a t a transmem- brane resistance of -50 ohms * cm').
The reversibility of the decrease in resistance was investi- gated after replacement of the low Ca'' medium with normal (high Ca2+) medium containing 1.8 mM Ca2+ (Table I). After 45 min of EGTA treatment, the transmembrane resistance returned to normal values within the 2-h recovery period. However, exposure of the cells to low Ca2+ medium for a longer time (90 min) resulted in an irreversible decrease in transmembrane resistance. Investigation of the monolayers in the light microscope showed that some cells had been detached from the filters. In contrast, monolayers that had been exposed to EGTA for 45 rnin were continuous and indistinguishable from control monolayers. These data indi- cate that the intercellular junctions are more sensitive to decreases in extracellular Ca2+ than the sites of focal contact (which anchor the cells to the substrate).
The kinetics of the restoration of the transmembrane resistance is shown in Figure 2. A gradual increase of the transmembrane resistance was observed over the 2-h period at 37 "C. However, the resistance was not completely restored when the incubation was performed at room temperature (22 "C). This indicates that the reassembly of the junctional complex is an energy-dependent process.
Scanning electron microscopy verified the finding in the light microscope that the monolayers were continuous (Fig- ures 3 and 4). However, the normal polygonal shape of the untreated Caco-2 cells was lost after 45 min of exposure to the low Ca2+ medium (Figure 4). These cells appeared to be contracted and rounded as compared with the untreated cells. Both untreated and EGTA-treated monolayers were heterog- enous in that they contained cells of different sizes and shapes.
B
"E 0
cn c 0
E
0 1 I I I - 2 0 30 8 0 1 3 0
Time, min Flgure 1-Effect of 2.5 mM EGTA on transmembrane resistance (n = 3; 2 SO).
596 ,' Journal of Pharmaceutical Sciences Vol. 79, No. 7, July 1990

Table &Effect of EGTA Treatment on Caco-2 Monolayers' Resistance, ohms * cm2
Treatment Intact Monolayer D ~ ~ ~ ~ I ~ ~ f t ~ ~ 2 h ~ f t ~ ~ Treatment Treatment
,,, 300
5 2 200
$ u z 100 t
cn E
C
K
None Yes 279 -+ 7 287 t 11 EGTA, 45 min Yes 63 f 7 270 t 11 EGTA, 90 min no <30 <30
investigated in the light microscope at the end of the experiment. 'The cells were exposed to 2.5 mM EGTA; the monolayers were
-
-
-
EGTA
n
0 I -50 5 0 1 5 0 2 5 0
Time, min Figure 2-The effect of a 45-min exposure of 2.5 mM EGTA on transmembrane resistance is reversible at 37 "C (closed symbols), but not at 22 "C (open symbols; n = 6; S D ) .
Figure +Scanning electron micrograph of untreated Caco-2 monolay- ers. The bar indicates 10 WM.
The transmission electron micrographs verified previous findings that the Caco-2 cells have a morphological appear- ance similar to that of well-differentiated small intestine absorbtive ~ells.11.12~18~24 The three components of the inter- cellular junctional complex (tight junctions or zonula occlu-
Figure &Scanning electron micrograph of EGTA-treated Cam2 monolayers (2.5 mM; 45 min). The bar indicates 10 pM. The cracks in the upper right and lower left corners are artifacts from the preparation of the specimens. Similar cracks were also present in the control specimens.
dens, intermediate junctions or zonula adherens, and desmo- somes or macula adherens) were present (see Figure 5). The polarity of the cells was clearly maintained after EGTA treatment for 45 min (Figure 6). However, in agreement with the findings in the scanning electron microscope, the cells showed brush border convexities. Moreover, the components of the junctional complex were often separated or dissolved. Thus, tight junctions and intermediate junctions were sparse and the outer leaflets of the plasma membranes appeared to be separated by a 40-80-nm zone a t the sites of the desmo- somes as compared with the normal 30-50 nm4 (insert in Figure 6). However, the visible junctional structures of adjacent cells appeared in parallel, which indicates that they were still in contact. (Figure 6).
Effect of Low Extracellular Ca2+ on Drug TransporGA clear effect of the reduction of the extracellular Ca2+ concen- tration was observed for the hydrophilic drug atenolol (see Figure 7). The transport of atenolol was increased about sixfold after the 45-min exposure to the low Ca2+ medium. The increase in permeability was reversible since the trans- port rate returned to normal levels after 2 h of recovery in high Ca2+ medium. Thus, the EGTA-induced decrease in transmembrane resistance was paralleled by an increase in atenolol transport (Figures 2 and 7).
Similar experiments with propranolol, a drug that is -2000 times more lipophilic than atenolol, gave different results (Figure 8). The reversible opening of the junctional com- plex had no effect on the transport rate of propranolol. Obviously, the transport of this drug is so fast that the contribution of the widened intercellular spaces to the overall transport is insignificant. Thus, the paracellular pathway is of limited importance for highly lipophilic drugs such as propranolol.
The experiments shown in Figures 7 and 8 were repeated with several other @blocking agents; the results are summa- rized in Figures 9 and 10 and Table 11. The reversible opening
Journal of Pharmaceutical Sciences I 597 Vol. 79, No. 7, July 1990

Flgure &Transmission electron micrographs of untreated cells. The plasma membranes of adjacent cells are brought into close contact by the components of the junctional complex. Tight junctions are visible (arrowheads; upper micrograph), as are desmosomes (arrowheads; lower micrograph). The bars indicate 1 pM.
of the junctional complex had a significant effect on the transport of the two most hydrophilic drugs and H216/44. Thus, the permeability coefficients of H216144, practolol, and atenolol increased 2-fold, 2-6-fold, and 4-9-fold, respectively (three different experiments), while the permeability of me- toprolol, alprenolol, and propranolol remained unchanged (Figure 10).
Discussion Epithelial cells are joined by a junctional complex which
is comprised of three separate structures: tight junctions, intermediate junctions, and desmosomes.4 The integrity of these structures is dependent on Ca2' .4*7.8-22 In this study, the Ca2+ dependence of the junctional complex was used to investigate the contribution of the paracellular pathway to the transepithelial transport of a homologous series of drugs. These studies were facilitated by the use of well- characterized and homogenous monolayers of human intes- tinal absorbtive (Caco-2) cells.10-12J8.24 Thus, the influence of several complicating factors such as mucus and other cell populations could be excluded. Another advantage was that the Caco-2 monolayers have a transmembrane resist-
ance of -250 ohms - cm2, which is similar to that of colonic mucosa. This resistance is so high that the transport of hydrophilic compounds is reduced to very low levels. Even snail changes in part :ellular permeability are therefore readily detectable.
The results indicate that the effects on transmembrane resistance were reversible. No cell detachment could be observed under the selected experimental conditions. How- ever, the detachment of a very small number of cells cannot be completely ruled out. Only a limited number of cells from relatively small areas of the monolayers can be studied in the transmission electron microscope and it is possible that losses of single cells occurred on other parts of the filter. (The detachment of more than one cell can easily be observed in the light microscope.) However, the observa- tions in the light microscope, the reversibility of the increases in permeability, and the finding in this study as well as by others7.22 that cell detachment after prolonged exposure to EGTA appears to be secondary to the retraction of the components of the junctional complex, supports the observations in the transmission electron microscope that no cell detachment occurred.
This is important since the detachment of a small number of cells will give erroneous permeability constants. Thus, a 20-times increase in the transport rate of atenolol was observed when the cells were exposed to EGTA for 90 min-an exposure time that induced irreversible damage to the mono- layers. This value is 2-5 times larger than those obtained under the reversible conditions used in this study. It can be concluded that studies on the effects of the various additives on the absorption of drugs are of limited value without the appropriate examination of possible injuries on the epithelial barrier.
In general, the opening of the junctional complex had no effect on the transport of the more lipophilic drugs. These drugs partition rapidly into the cell membranes. The distri- bution to the intercellular spaces will therefore be limited even after EGTA treatment since the cell membranes cover a much larger surface area than the intercellular spaces. However, the more hydrophilic drugs will have a low solu- bility in the cell membranes. The transport through the paracellular pathway is therefore more important for these drugs. Manipulation of the paracellular permeability in- creased the absorption of the otherwise slowly absorbed hydrophilic drugs significantly. The largest increase was observed for atenolol, a drug that is incompletely absorbed in humans.25 It is likely that a selective opening of the junctional complex can be used to increase the absorption of other hydrophilic drugs as well. The transport of H216/44 was also increased. This lipophilic compound is transported at an unexpectedly slow rate across intact Caco-2 monolayers.10 The enhanced transport of H216/44 indicates that the para- cellular pathway is of importance not only for hydrophilic, but also for hydrophobic drugs that are transported slowly across the intestinal epithelium.
It has been shown that the transport of the more hydro- phobic P-blocking agents proceeds at comparable rates in the Caco-2 model and in rat ileum in situ.10.26 However, the most hydrophilic drugs, practolol and atenolol, were transported two and seven times slower, respectively, than could be expected from the in situ data. It was suggested that these differences could be related to the different electrical param- eters of the two models. Thus, rat ileum has a transmembrane resistance of 60-90 ohms * cm2,27,28 while the corresponding value for the Caco-2 monolayers is 170-400 ohms * cm2.10-12 The results of this study support this hypothesis, since the transport of practolol and atenolol increased %&fold and 4-9-fold, respectively, when the transmembrane resistance was decreased to levels comparable to that of rat ileum. After
598 I Journal of Pharmaceutical Sciences Vol. 79, No. 7, July 1990

Figure &Transmission electron micrographs of EGTA-treated Caco-2 cells (2.5 mM; 45 min). The cells show brush border convexities. The intercellular spaces are widened as compared with the control cells (stars), but the plasma membranes of adjacent cells are still held in contact by the components of the junctional complex (arrowheads). However, the gaps between the junctional structures on adjacent cells are widened (insert). The bars indicate 1 pM. (The insert is an enlargement of the central part in Figure 6).
A
100 200 300 0 Time, min
Figure 7-Effect of EGTA treatment on the transport of 1 x M atenolol across Caco-2 monolayers The transport of atenolol was measured before, immediately after, and 2 h after the EGTA treatment. The same monolayers were used in all three experiments (n = 3; tSD).
exposure to low Ca2' medium, the transport rates of the hydrophilic, as well as the lipophilic p-blocking agents, were comparable to those published for rat ileum.10.26 These results indicate that the paracellular drug permeability of the Caco-2 model is significantly lower than that of rat ileum and more similar to that of colonic epithelium.29 It follows that very hydrophilic drugs such as atenolol are absorbed at compara- bly slow rates across the Caco-2 cells. More leaky monolayers with transmembrane resistances similar to that found in the small intestine are therefore better models of ileal absorption of very hydrophilic drugs.
It is not likely that Ca2+ chelators such as EGTA will be used in vivo to enhance the uptake of drugs that are incom- pletely absorbed. The Ca2+ concentration is dependent on too many endogenous factors to allow a reproducible reduction of
0 20 4 0 60 80 100 Time, min
Flgure &Effect of EGTA treatment on the transport of 1 x lo-' M propranolol across the Caco-2 monolayers. The experimental conditions were the same as in Figure 7 (n = 3; +SO).
its concentration in the gastrointestinal tract. The EGTA treatment gave variable increases in permeability even under the controlled conditions used in this study. In addition, there will always be a risk for overexposure with cell detachment as a result. More selective and less toxic ways to regulate the paracellular permeability have to be found before this strat- egy can be applied in pharmaceutical formulations.3.6~6
In summary, the results indicate that the Caco-2 model can be used to study the relative importance of the paracel- lular and transcellular routes for the transport of a certain drug. Similar experiments can be performed to study the effects of other compounds, such as various pharmaceutical additives. Thus, the Caco-2 model will be a valuable tool in the preformulation and optimization of oral drug delivery systems.
Journal of Pharmaceutical Sciences I 599 Vol. 79, No. 7, July 1990
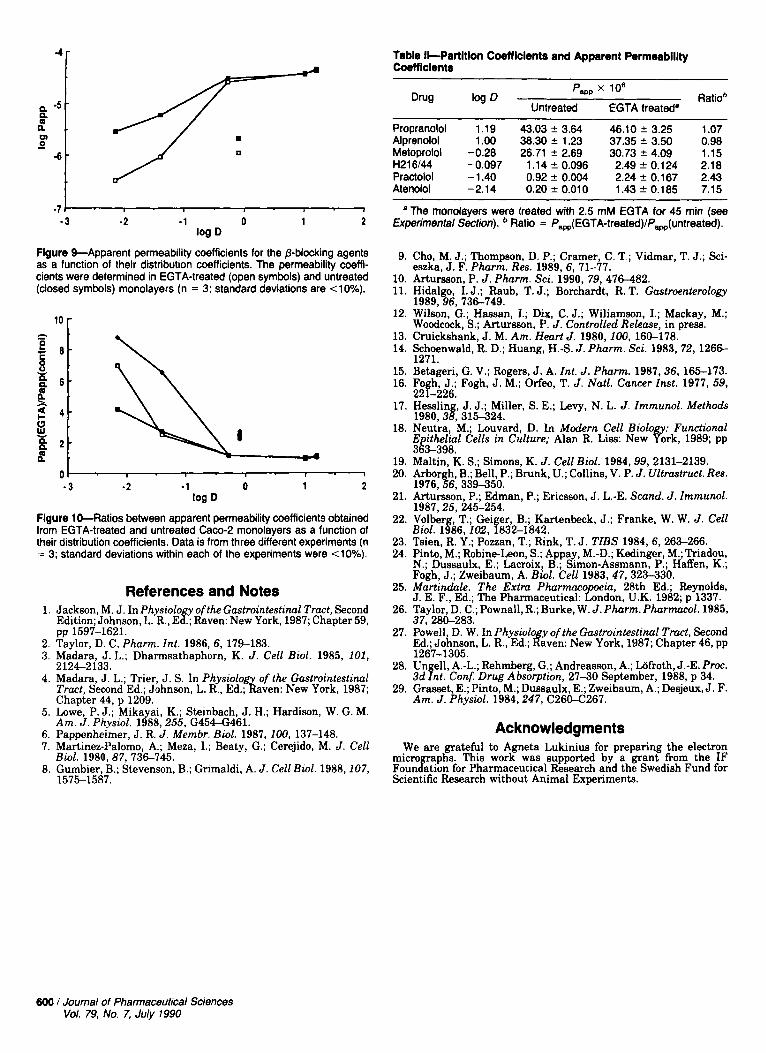
4r Table Il-Partltlon Coeff lclents and Apparent Permeablllty Coemclents
-7 I I 1
- 3 -2 -1 0 1 2
Flgure %Apparent permeability coefficients for the @-blocking agents as a function of their distribution coefficients. The permeability coeffi- cients were determined in EGTA-treated (open symbols) and untreated (closed symbols) monolayers (n = 3; standard deviations are <lo%).
log D
" 0
- 3 - 2 -1 0 1 2 log D
Flgure 10-Ratios between apparent permeability coefficients obtained from EGTA-treated and untreated Cam-2 monolayers as a function of their distribution coefficients. Data is from three different experiments (n = 3; standard deviations within each of the experiments were <lo%).
1.
2. 3.
4.
5.
6. 7.
8.
References and Notes Jackson, M. J. InPhysiolo y of the Gastrointestinal Tract, Second Edition; Johnson, L. R., E f ; Raven: New York, 1987; Chapter 59,
Taylor, D. C. Pharm. Znt. 1986, 6, 179-183. pp 1597-1621.
Madara, J. L.; Dharmsathaphorn, K. J. Cell Biol. 1985, 101, 2 124-2 133. Madara, J. L.; Trier, J . S. In Physwlog of the Gastrointestinal Tract, Second Ed.; Johnson, L. R., Ed.; kiven: New York, 1987; Chapter 44, p 1209. Lowe, P. 5.; Mikayai, K.; Steinbach, J. H.; Hardison, W. G. M. Am. J. Physiol. 1988,255, G45-461. Pamenheimer. J. R. J. Membr. Bwl. 1987.100.137-148. . . M&tinez-Palomo, A.; Meza, I.; Beaty, G.; Cerejido, M. J. Cell Bwl. 1980,87, 736-745. Gumbier, B.; Stevenson, B.; Grimaldi, A. J. Cell Biol. 1988,107, 1575-1587.
~~~
Propranolol 1.19 43.03 f 3.64 46.10 f 3.25 1.07 Alprenolol 1.00 38.30 f 1.23 37.35 * 3.50 0.98 Metoprolol -0.28 26.71 f 2.69 30.73 f 4.09 1.15 H216M -0.097 1.14 f 0.096 2.49 f 0.124 2.18 Practolol -1.40 0.92 f 0.004 2.24 f 0.167 2.43 Atenolol -2.14 0.20 * 0.010 1 . 4 3 f 0.185 7.15
The monolayers were treated with 2.5 mM EGTA for 45 min (see Experimental Section). Ratio = fw(EGTA-treated)/f,(untreated).
9. Cho, M. J.; Thompson, D. P.; Cramer, C. T.; Vidmar, T. J.; Sci- eszka, J. F. Pharm. Res. 1989, 6, 71-77.
10. Artursson. P. J. Pharm. Sci. 1990. 79. 476482. 11. Hidalgo, I. J.; Raub, T. J.; Borchardt, R. T. Gastroenterology
1989.96. 736-749. 12. Wilson, G.; Hassan, I.; Dix, C. J.; Wiliamson, I.; Mackay, M.;
Woodcock, S.; Artursson, P. J. Controlled Release, in press. 13. Cruickshank, J. M. Am. Heart J . 1980,100, 160-178. 14. Schoenwald, R. D.; Huang, H A . J. Phurm. Sci. 1983, 72, 1 2 6 6
15. Betageri, G. V.; Rogers, J. A. Znt. J. Pharm. 1987,36, 165-173. 16. Fo h, J.; Fogh, J. M.; Orfeo, T. J. Natl. Cancer Inst. 1977, 59,
1271.
22f-226. 17. Hesslin , J. J.; Miller, S. E.; Levy, N. L. J. Zmmunol. Methods
1980,3j, 315-324. 18. Neutra, M.; huvard , D. In Modern Cell Biolo : Functional
E itheliul Cells in Culture; Alan R. Liss: New E r k , 1989; pp 3l3-398
19. Maltin, K. S.; Simons, K. J. Cell Biol. 1984,99, 2131-2139. 20. Arbor h, B.; Bell, P.; Brunk, U.; Collins, V. P. J. Ultmtruct . Res.
1976,%6,339-350. 21. Arturseon, P.; Edman, P.; Ericsson, J. L.-E. Scand. J. Zmmunol.
1987,25,245-254. 22. Volber , T.; Gei er, B ; Kartenbeck, J.; Franke, W. W. J. Cell
Bwl. 1886,102, f832-1842. 23. Tsien, R. Y.; Pozzan, T.; Rink, T. J. TZBS 1984, 6, 263-266. 24. Pinto, M.; Robine-Leon, S . ; Appa , M. D Kedinger, M.; Triadou,
N.; Dussaulx, E.; Lacroix? B.; dmoi-A&mann, P.; Haffen, K.; Foah. J.: Zweibaum. A. Bwl. Cell 1983, 47, 323-330.
25. M&&& The Extra Phnrmaco iu, 28th Ed.; Reynolds,
26. Taylor, D. C.; Pownall, R.; Burke, W. J. Pharm.Pharmacol.1985, J. E. F., Ed.; The Pharmaceutical: E d o n , U.K. 1982; p 1337.
37. 28n-283. - . , -- - - - - . 27. Powell, D. W. In Physwlo y of the Gastrointestinal T m t , Second
Ed.; Johnson, L. R., Ed.; Ifwen: New York, 1987; Chapter 46, pp
28. Un ell, A.-L.; Rehmberg, G.; Andreasson, A.; Lijfroth, J.-E. P m . 3d ht. Conf. Drug Absorption, 2 7 3 0 September, 1988, p 34.
29. Grasset, E.; Pinto, M.; Dussaulx, E.; Zweibaum, A.; Desjeux, J. F. Am. J . Physwl. 1984,247, C260.4267.
1267-1305.
Acknowledgments We are grateful to Agneta Lukinius for preparing the electron
micro aphs. This work was supported by a grant from the IF Foungtion for Pharmaceutical Research and the Swedish Fund for Scientific Research without Animal Experiments.
600 I Journal of Pharmaceutical Sciences Vol. 79, No. 7, July 1990