enhanced tumor cell killing in the presence of ganciclovir...
TRANSCRIPT

[CANCER RESEARCH 58. 5731-5737. December 15. 19981
Enhanced Tumor Cell Killing in the Presence of Ganciclovir by Herpes SimplexVirus Type 1 Vector-directed Coexpression of Human Tumor NecrosisFactor-«and Herpes Simplex Virus Thymidine Kinase1
Shusuke Moriuchi, Thomas Oligino, David Krisky, Peggy Marconi, David Fink, Justus Cohen, andJoseph C. Glorioso2
Departments of Molecular Genetics and Biochemistry IS. M., T. O.. D. K.. P. M., J. C. J. C. C.I and Neurology ¡D.F.¡.University of Pittsburgh School of Medicine, Pittsburgh.Penns\lvania 15261
ABSTRACT
Past studies have documented the promise of herpes simplex virus type 1(HSV-1) IIn mil liiic.'kinase (TK) suicide gene therapy as a potential antitumor
treatment. HSV-TK converts the pro-drug ganciclovir (GCV) into a toxic
nucleotide analogue, the incorporation of which into cellular DNA blocks cellproliferation. In this report, we have examined the hypothesis that the effectiveness of HSV-TK suicide gene therapy can be enhanced by coexpression ofthe antitumor cytokine human tumor necrosis factor-a (TNF-a) from thesame replication-defective HSV-1 vector. In vitro testing demonstrated thatTNF-a expression from this vector potentiated the killing of both TNF-a-sensitive L929 tumor cells and TNF-a-resistant U-87 MG cells in the presence
of GCV. Furthermore, treatment of established intradermal L929 tumors invivo with the TNF-o/TK vector and GCV resulted in prolonged animalsurvival compared with treatment with parental HSV-TK vector in thepresence or absence of GCV. Treatment of intracerebral U-87 MG tumors
showed a clear benefit of TK therapy, but a significant further increase insurvival using the TNF-a vector could not be demonstrated. We found thatpotentiation of cell killing in vitro required intracellular TNF-a becausepurified protein added to the culture medium of cells infected with HSV-TK
vector failed to have the same effect. Accordingly, potentiation in vivo shoulddepend on efficient infection, but immunohistochemical analysis indicatedthat virus administration by U-87 MG ¡niradiinomi injection was inadequate,
resulting in an estimated <1% infection of all tumor cells. Moreover, themajority of infected tumor cells were localized at the tumor margin. Together, these results suggest that TNF-enhanced tk gene therapy shouldprovide a useful treatment for TNF-a-sensitive tumors and perhaps also forTNF-a-resistant tumors if vector delivery can be improved to increase the
percentage of transduced tumor cells.
INTRODUCTION
Gene therapy is a relatively new technique with potential as atreatment for cancer and a wide variety of other diseases. One anti-
cancer gene therapy approach that has already been applied in clinicalsettings is the direct transfer of genes that activate prodrugs locally orinduce programmed tumor cell death (1-10). Although such "suicide"
gene therapy approaches have been successful to varying degrees inboth preclinical and clinical studies, it is possible that "cure" rates can
be improved through the use of multiple therapeutic genes, the products or which destroy tumor cells by a combination of mechanisms.Experiments described in this report attempt to test this hypothesis byvector-mediated codelivery to the tumor mass of the antitumor cytokine TNF3-a and the HSV-1 TK suicide gene followed by systemic
treatment with GCV.
Received 4/29/98: accepted 10/20/98.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby marked advertisement in accordance with18 U.S.C. Section 1734 solely to indicate this fact.
1This work was supported by Grant 108666 from (he NIH (to J. C. G.) and by a grant
from GenVec. Inc.. Rockville, MD.2 To whom requests for reprints should be addressed, al Department of Molecular
Genetics and Biochemistry. E1240 Biomedicai Science Tower. University of PittsburghSchool of Medicine. Pittsburgh, PA 15261. Phone: (412) 648-8106: Fax: (412) 624-8997:E-mail: [email protected].
1The abbreviations used are: TNF. tumor necrosis factor: HSV-1, herpes simplex virus
type 1; TK. thymidine kinase; GCV, ganciclovir; NK. natural killer: ATCC, American
HSV-TK is a commonly used suicide gene in both experimental andclinical settings (1-10). Expression of HSV-TK in tumor cells results
in tumor cell death when the cells are exposed to the antiviral prodrugGCV as the TK protein phosphorylates the prodrug into a toxicnucleotide analogue, which terminates strand elongation during DNAreplication. A powerful feature of the TK/GCV approach is that thetransduction of a small fraction of the targeted cells can result insignificant antitumor activity (1-10). This has been ascribed to the
killing of untransduced neighboring cells and is referred to as thebystander effect (1). It has been demonstrated that the cell-to-celltransfer of phosphorylated GCV via gap junctions between HSV-TK
transduced cells and neighboring uninfected cells is the primarymechanism for the bystander effect (11-15).
In an attempt to enhance the tumoricidal activity of TK/GCVsuicide gene therapy, we have created a replication-defective HSV-1vector expressing the human TNF-a gene in conjunction with HSV-/A-.
TNF-a has been demonstrated to possess an array of antitumor
activities, including potent cytotoxicity exerted directly on tumor cells(16, 17), enhancement of the expression on tumor cell surfaces ofHLA antigens (18) and intercellular adhesion molecule 1(19) and onlymphocytes of interleukin-2 receptors (20), and promotion of theactivation of such effector cells as NK cells, lymphokine-activatedkiller cells and CTLs (21-23). Yet, despite this promising antitumorprofile, the clinical use of TNF-a has been constrained by the toxicityof systemic TNF-a administration (24, 25). The possibility exists,
however, that this limitation can be overcome by the use of genetransfer methods in which vector-mediated local production of TNF-a
at the site of tumor growth may allow for effective use of this cytokineas an antitumor agent without toxicity to other tissues.
In this study, we assessed the effectiveness of treating tumors withour HSV-1 vector expressing both TNF-a and HSV-TK. We present
evidence from cell culture as well as in vivo experiments that theefficacy of HSV-TK suicide gene therapy can be increased by thesimultaneous expression of TNF-a.
MATERIALS AND METHODS
Cell Culture and Virus Production. Vero African green monkey kidneycells (CCL81; ATCC, Rockville. MD), Vero-derived 7B cells (26), and mouse
L929 fibrosarcoma cells (kindly provided by Dr. Leaf Huang. Department ofPharmacology. University of Pittsburgh) were maintained in MEM (LifeTechnologies. Inc.. Gaithersburg. MD) supplemented with glutamine, antibiotics (PSN; Life Technologies. Inc.). and 10% fetal bovine serum (LTI.Gaithersburg. MD). U-87 MG human glioblastoma cells (HTB-14; ATCC)
were maintained in DMEM supplemented with glutamine, antibiotics, and10% fetal bovine serum.
HSV-1 stocks were prepared by infecting 7B cells, which supply the ICP4
and ICP27 proteins in trans, at a multiplicity of 0.03. Infected cells wereharvested when a 100% cytopathic effect was evident and subjected to three
Type Culture Collection: pfu. plaque-forming unit(s): IE. immediate early: MOI. multiplicity of infection: MIT. 3-(4,5-dimethylthiazol-2-yl)-diphenyltetraœlium bromide:
CMV. cytomegalovirus.
5731
on May 29, 2018. © 1998 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
TNF-a AND HSV-ik COMBINATIONGENE THERAPY
cycles of freeze-thawing (-80°C/37°C) and a single burst of sonication. The
virus was (hen separated from cell debris by centrifugation and purified awayfrom free protein and membrane particles by density gradient centrifugation(OptiPrep; Lite Technologies, Inc.). Virus stocks were aliquoted into 2-mlcryogenic tubes (Corning Glass Works. Corning. NY) and stored at —80°C.Final titers averaged 5 x IO7 to 5 X 10s pfu/ml.
Construction of Plasmids and Recombinant Vectors. Plasmids wereisolated and grown in Escherichia coli strain DH5a (Life Technologies, Inc.).Plasmid-transformed bacteria were selected on YT agar plates (YT is 5 g ofyeast extract, 5 g of NaCl, and 8 g of bacto-tryptone per liter distilled water,
pH 7.0) containing 150 ¿¿g/mlampicillin (Sigma Chemical Co., St. Louis. MO)and grown in 2X YT broth containing 150 /j,g/ml ampicillin.
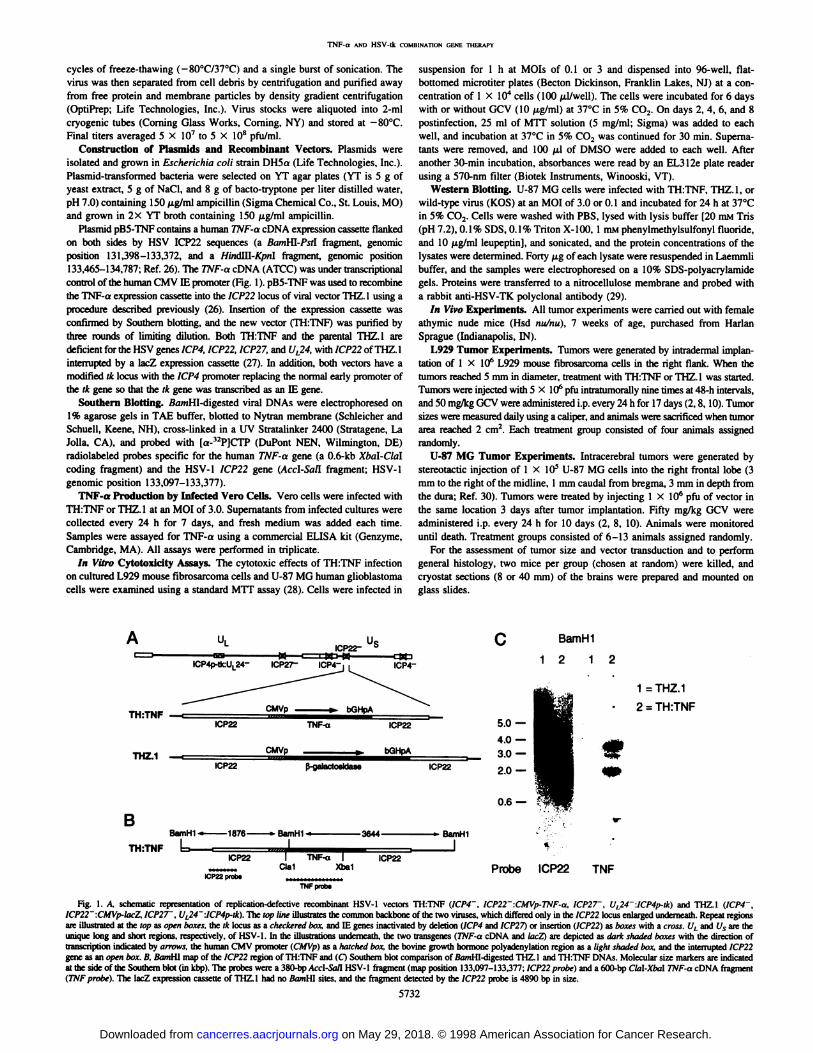
Plasmid pB5-TNF contains a human TNF-a cDNA expression cassette flankedon both sides by HSV ICP22 sequences (a BanA\\-Pst\ fragment, genomicposition 131,398-133,372, and a Hindlll-Kpnl fragment, genomic position133,465-134,787: Ref. 26). The TNF-a cDNA (ATCC) was under transcriptionalcontrol of the human CMV IE promoter (Fig. 1). pB5-TNF was used to recombinethe TNF-a expression cassette into the ICP22 locus of viral vector THZ. 1 using a
procedure described previously (26). Insertion of the expression cassette wasconfirmed by Southern blotting, and the new vector (TH:TNF) was purified bythree rounds of limiting dilution. Both TH:TNF and the parental THZ.l aredeficient for the HSV genes ICP4. ICP22. ICP27, and U,24, with 1CP22 of THZ. Iinterrupted by a lacZ expression cassette (27). In addition, both vectors have amodified tk locus with the ICP4 promoter replacing the normal early promoter ofthe ik gene so that the Ik gene was transcribed as an IE gene.
Southern Blotting. BoniHI-digested viral DNAs were electrophoresed on
\% agarose gels in TAE buffer, blotted to Nytran membrane (Schleicher andSchuell, Keene, NH), cross-linked in a UV Stratalinker 2400 (Stratagene, LaJolla, CA), and probed with [a-'2P]CTP (DuPont NEN, Wilmington, DE)
radiolabeled probes specific for the human TNF-a gene (a 0.6-kb Xbal-Clalcoding fragment) and the HSV-1 ICP22 gene (Accl-Sall fragment: HSV-1genomic position 133,097-133,377).
TNF-a Production by Infected Vero Cells. Vero cells were infected with
TH:TNF or THZ. 1 at an MOI of 3.0. Supernatants from infected cultures werecollected every 24 h for 7 days, and fresh medium was added each time.Samples were assayed for TNF-a using a commercial ELISA kit (Genzyme,
Cambridge, MA). All assays were performed in triplicate.In Vitro Cytotoxicity Assays. The cytotoxic effects of TH:TNF infection
on cultured L929 mouse fibrosarcoma cells and U-87 MG human glioblastoma
cells were examined using a standard MTT assay (28). Cells were infected in
suspension for l h at MOIs of 0.1 or 3 and dispensed into 96-well, flat-
bottomed microtiter plates (Becton Dickinson, Franklin Lakes, NJ) at a concentration of 1 X IO4 cells (100 fil/well). The cells were incubated for 6 dayswith or without GCV (10 ng/ml) at 37°Cin 5% CO2. On days 2, 4, 6, and 8
postinfection, 25 ml of MTT solution (5 mg/ml: Sigma) was added to eachwell, and incubation at 37°Cin 5% CO2 was continued for 30 min. Superna
tants were removed, and 100 /xl of DMSO were added to each well. Afteranother 30-min incubation, absorbances were read by an EL312e plate readerusing a 570-nm filter (Biotek Instruments, Winooski, VT).
Western Blotting. U-87 MG cells were infected with TH:TNF, THZ.l, orwild-type virus (KOS) at an MOI of 3.0 or 0.1 and incubated for 24 h at 37°C
in 5% CO2. Cells were washed with PBS, lysed with lysis buffer [20 mM Tris(pH 7.2), 0.1% SDS, 0.1% Triton X-100, 1 mM phenylmethylsulfonyl fluoride,
and 10 fig/ml leupeptin], and sonicated, and the protein concentrations of thelysates were determined. Forty /j.g of each lysate were resuspended in Laemmlibuffer, and the samples were electrophoresed on a 10% SDS-polyacrylamide
gels. Proteins were transferred to a nitrocellulose membrane and probed witha rabbit ami-HSV-TK polyclonal antibody (29).
In Vivo Experiments. All tumor experiments were carried out with femaleathymic nude mice (Hsd nu/nu), 1 weeks of age, purchased from HarÃanSprague (Indianapolis, IN).
L929 Tumor Experiments. Tumors were generated by intradermal implantation of 1 X IO6 L929 mouse fibrosarcoma cells in the right flank. When the
tumors reached 5 mm in diameter, treatment with TH:TNF or THZ. 1 was started.Tumors were injected with 5 x IO6pfu intratumorally nine times at 48-h intervals,
and 50 mg/kg GCV were administered i.p. every 24 h for 17 days (2,8,10). Tumorsizes were measured daily using a caliper, and animals were sacrificed when tumorarea reached 2 cm2. Each treatment group consisted of four animals assigned
randomly.U-87 MG Tumor Experiments. Intracerebral tumors were generated by
stereotactic injection of 1 x 1()5U-87 MG cells into the right frontal lobe (3
mm to the right of the midline, 1 mm caudal from bregma, 3 mm in depth fromthe dura: Ref. 30). Tumors were treated by injecting 1 x IO6 pfu of vector in
the same location 3 days after tumor implantation. Fifty mg/kg GCV wereadministered i.p. every 24 h for 10 days (2, 8, 10). Animals were monitoreduntil death. Treatment groups consisted of 6-13 animals assigned randomly.
For the assessment of tumor size and vector transduction and to performgeneral histology, two mice per group (chosen at random) were killed, andcryostat sections (8 or 40 mm) of the brains were prepared and mounted onglass slides.
ICP22-
ICP4p-tk:U,24- ICP27- ICP4~ ICP4-
TH:TNF
THZ.1
CMVp bGHpA
ICP22CICP22TNF-aß-galactosidaseICP22bGHpAICP225.04.02.0
B
TH:TNF |ICP22ic'p22p*rôb«l 11dalTNF-a
1
Xba1ICP22
TNF probe
BamH1
12 12
1 = THZ.1
2 = TH:TNF
0.6 —
Probe ICP22 TNF
Fig. I. A. schematic representation of replication-defectiverecombinant HSV-1 vectors TH:TNF (ICP4~. iCP22~\CMVp-TNF-a, 1CP27~. U,24~ \ICP4p-ik) and THZ.1 (KP4~,ICP22~:CMVp-lacZ,ICP27~. UL24~:ICP4p-tk).The ¡opline illustratesthe common backboneof the two viruses,which differedonly in the ICP22 locusenlargedunderneath.Repeatregions
are illustratedat the lop as open boxes, the tk locus as a checkeredbox, and IE genes inactivatedby deletion(ICP4 and ICP27) or insertion(ICP22) as boxes with a cross. if¡and i/s-are theunique long and short regions, respectively,of HSV-1. In the illustrationsunderneath,the two transgenes(TNF-a cDNA and lacZ) are depicted as dark shaded boxes with the directionoftranscriptionindicatedby arrows, the human CMV promoter(CMVp)as a hatched box, the bovine growth hormone polyadenylationregion as a light sluuledbox, and the interruptedICP22gene as an open box.B. BanMl map of the ICP22 region of TH:TNF and (O Southernblot comparisonof SumHI-digestedTHZ.1and TH:TNF DNAs. Molecularsize markersare indicatedat the side of the Southernblot (in kbp).The probeswere a 380-bpAcc\-Sall HSV-] fragment(map position 133,097-133,377:ICP22 probe) and a 600-bp Cla\-Xbal TNF-a cDNA fragment(TNF probe}. The lacZ expressioncassetteof THZ. I had no BamHl sites, and the fragmentdetected by the ICP22 probe is 4890 bp in size.
5732
on May 29, 2018. © 1998 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

TNF-tr AND HSV-Ik COMBINATION CENE TOHIAPY
Immunohistochemistry. Brains were fixed by perfusion with 4%paraformaldehyde and resected. Cryostat sections were prepared on poly-L-
lysine slides, rinsed with 1% H2O, in methanol and with PBS. and stained witha rabbit anti-HSV-1-antigen antibody (Accurate. Westbury, NY; 1:2000 dilution) in 5% normal goat or rabbit serum overnight at 4°C.The sections were
washed three times with 0.3% Triton-X in PBS (PBS-T). incubated withbiotin-conjugated goat anti-rabbit for 2 h at room temperature, and againwashed three times with PBS-T. The sections were then incubated withavidin-biotin complex (ELITE kit; Vector. Burlingame, CA; 1:400 dilution)for l h at room temperature, washed with PBS-T three times, stained with 0.1%
diaminobenzidine tetrahydrochloride (DAB substrate kit; Vector), washed withtap water, and counterstained with hematoxylin.
RESULTS
TH:TNF Vector Construction. A replication-defective genomicHSV-1 vector for coexpression of HSV-TK and the tumoricidal cytokineTNF-a was constructed by recombining a TNF-a expression cassette into
the ICP22 locus of a previously described lacZ expression vector, THZ. 1(Fig. \A, TH:TNF). The parental THZ.l had the following features: (a) itwas deleted for the two essential IE genes ICP4 and ICP27 and wastherefore highly defective for early (E) and late (L) gene expression andincapable of productive infection; (b) it contained a lacZ expressioncassette interrupting and thereby inactivating a third IE gene, ICP22; and(c) it had a modified tk locus in which the native E promoter (as well asthe U'¡24gene encoded on the opposite DNA strand) had been replaced
by the ICP4 IE promoter (ICP4p) to express TK as an IE gene product(27). It has been reported previously that combined IE gene deficienciessuch as those of THZ. 1 substantially reduce the undesirable cytotoxicityof wild-type HSV-1 and mutant vectors deleted for single IE genes (26),which results in extended transgene expression (26). The TNF-o expression cassette used for recombination consisted of a human TNF-a cDNA
between the human CMV IE promoter (CMVp) and a polyadenylationregion from the bovine growth hormone gene and was surrounded byICP22 sequences in a plasmid background. The TNF-a cassette was
introduced in place of the resident lacZ cassette of THZ. 1 by homologousrecombination using an efficient protocol described previously (26), andthe structure of the final TH:TNF isolate was confirmed by Southern blotanalysis (Fig. 15).
The TH:TNF vector was tested for expression of TNF-a by infec
tion of noncomplementing Vero cells and determination by ELISA ofthe amounts of TNF-a produced during successive 24-h periods over
7 days. As illustrated in Fig. 2A, medium (1 ml) collected after thefirst day contained 13.6 ng of TNF-a, but this was followed by aprecipitous decline in production on day 2 and subsequent days (1-2
ng/24 h), and the protein was no longer detectable by day 7.The bioactivity of TNF-a produced in this experiment was tested
by exposure of cultured TNF-a-sensitive L929 fibrosarcoma cells to
medium collected after the first day of infection. As measured bystandard MTT viability assay, L929 cells treated with medium fromeither mock-infected or THZ. 1 control-infected Vero cells showed no
significant reduction in viability relative to untreated L929 cells (Fig.2B). In contrast, a dramatic reduction was observed for cells treatedwith medium from TH:TNF-infected cells, and this reduction was
essentially identical over time to that seen with unconditioned medium supplemented with 10 ng of recombinant TNF-a protein (Fig.2ß).These results demonstrated that TNF-a produced by the TH:TNF
vector was biologically active and comparable in specific activity torecombinant TNF-a.
Enhanced Killing of TNF-a-sensitive Cells in Vitro by TH:TNFand GCV. To determine whether vector-directed production ofTNF-a could enhance HSV-TK/GCV-mediated cell killing of a TNF-a-sensitive cell line. L929 cells were infected at low multiplicity
(MOI, 0.1 ) with TH:TNF or parental vector THZ. 1, and the cells were
A.Vero/TH:TNF mediumVero/THZ.1 medium1 ng/ml TNF-a10 ng/ml INI- •
1234567Days post-infection
Fig. 2. A, production of TNF-a by TH:TNF-infected Vero cells. Vero cells (1 X IO6)
were infected with TH:TNF or control virus THZ.l al an MOI of 3.0. Media werecollected every 24 h and assayed hy TNF-a ELISA. The results are shown as the amountof TNF-a in 1 ml medium at each time point. B, biological activity of TNF-a producedby infected Vero cells. Vero cell media collected 24 h after infection (see A] wereincubated with cultures of TNF-a-scnsitive L929 cells, and survival over a 4-day periodwas monitored by MTT assay. The results are shown as the percentage of survival relativeto cultures treated with medium from mock-infected Vero cells, and the values represent
averages of triplicate determinations; bars, SD. For comparison, survival of L929 cells inI ml of medium supplemented with I ng/ml or 10 ng/ml purified recomhinant TNF-aprotein (Genzyme. Cambridge, MA) was also determined.
maintained in the presence or absence of GCV. At 24-h intervals, cell
viability was determined by MTT assay, and the results were plottedas the percentage of survival relative to mock-infected cells (Fig. 3/4).THZ. 1-infected cultures exposed to GCV displayed a 40% reduction
in viability 3 days after infection, compared with <20% in theabsence of GCV. Survival of TH:TNF-infected cultures was ~50%
but was reduced to 35% in the presence of GCV. Hence, the combination of HSV-TK/GCV and TNF-a augmented the cytocidal effi
ciency of each treatment alone.The common observation that GCV treatment kills more cells than are
actually infected with virus, producing activating TK protein, is referredto as the bystander effect. The bystander effect is thought to result fromintercellular spread of activated GCV from infected cells to uninfectedneighboring cells via gap junctions (11-15) and is therefore best observedat low MOI. For example, exposure of THZ. 1-infected cells to GCV (Fig.
3A) reduced cell survival by nearly 25%, although <10% of the cellswere infected with the nonreplicating vector. Likewise, the survival ofTH:TNF-infected cultures was reduced by >15% in the presence of
GCV, suggesting that the bystander effect was operational in thesecultures as well and that cell killing by activated GCV was additive withthe cytotoxic effect of TNF-a expression.
Fig. 3B shows the results of a second experiment performed at a10-fold higher MOI, where ~70% of the cells are expected to be infected.A bystander effect was not obvious here for either THZ.l- or TH:TNF-infected cells. GCV reduced the viability of THZ. 1-infected cells by 30%,
an amount similar to that observed for the earlier MOI of 0.1. TheTH:TNF-infected culture was obliterated in 2 days, but there was no
evidence that GCV accelerated this process because the survival curveswith and without GCV were essentially the same. These results suggestedthat combination treatment could be advantageous over single genetreatment at lower MOIs, although not at higher MOIs, which wasencouraging because the level of infection attainable with nonreplicatingvectors in vivo is generally low. Moreover, limited infection without viralspread should avoid the majority of undesirable side effects associatedwith sustained TNF-a expression at higher levels.
Enhanced Killing in Vitro by TH:TNF and GCV of a TNF-a-resistant Glioblastoma Cell Line. Unlike L929 cells, most humantumors are resistant to TNF-a-receptor-dependent cell killing, includ
ing those that arise in the central nervous system (31, 32). It was of
5733
on May 29, 2018. © 1998 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

TNF-a AND HSV-tk COMBINATIONÃœENETHERAPY
150-, L929
MOI=0.1
THiTNF B.TH:TNF+GCVTHZ.1THZ.1+GCV 100-1L929
MObl.O
23 01
Days post-infection
Fig. 3. Survival of TH:TNF- and THZ.l-infected L929 cells in the presence or absence
of GCV. L929 cells were infected with TH:TNF or THZ. I at MOIs of 0.1 (A) or 1.0 (B),and cell survival in the presence or absence of GCV (10 ¿¿g/ml)was measured by MTTassay. The results are plotted as survival relative to untreated mock-infected cells, and
each value represents the average of triplicate determinations; bars, SD.
interest, therefore, to determine whether vector-directed TNF-a expression could enhance the killing by HSV-TK/GCV of U-87 MGcells, a well-defined human glioblastoma cell line which in our handsis resistant to TNF-a concentrations of as high as 100 ng/ml.4 Ac
cordingly, U-87 MG cells were infected at low MOI, and viability
with or without exposure to GCV was determined by MTT assay.In the absence of GCV, cultures infected with TH:TNF at an MOI
of 0.1 showed no significant reduction in viability compared withmock- or THZ. 1-infected cells (Fig. 44), which was consistent withthe established TNF-a resistance of these cells. In the presence ofGCV, the viability of THZ.l-infected cultures was reduced by 50% 6
days after infection, suggesting a robust bystander effect. Surprisingly, cultures infected with TH:TNF and exposed to GCV showed asubstantially larger decrease in viability (90% by day 6; Fig. 44;P < 0.01, Student's t test). Although the mechanism of this enhance
ment is unclear, this observation indicated that the combination ofvector-directed TNF-a expression and HSV-TK expression with GCVtreatment could be beneficial not only against TNF-a-sensitive tumorcells like L929 but also against TNF-a-resistant tumors.
The preceding result could be a consequence of TK- and/or GCV-mediated sensitization of the cells to TNF-a or vice versa, TNF-a-
mediated potentiation of the bystander effect. To distinguish betweenthese alternatives, we tested whether TNF-a protein added to the
growth medium would reduce the viability in the presence of GCV ofU-87 MG cells infected with THZ.l at an MOI of 0.1. The resultsshowed that TNF-a added in this manner (10 ng/ml) had no signifi
cant effect (Fig. 45), indicating that the increased cell killing byTH:TNF/GCV compared with THZ.l/GCV was dependent on intra-cellular production of TNF-a by the vector and was not mediated bysecreted TNF-a or through sensitization of the cells to extracellularTNF-a by HSV-TK/GCV. The immunoblot of Fig. 4C demonstratesthat there were no significant differences in the amounts of HSV-TKproduced by U-87 MG cells infected with TH:TNF or THZ.l at highor low MOI, which excluded the possibility that vector-producedTNF-a increased cell killing by raising the level of TK protein.
Interestingly, we observed that TH:TNF infection at high MOIcaused a dramatic reduction in U-87 MG cell viability in the absence
of GCV (Fig. 4£>).Because these cells are resistant to high levels ofextracellular TNF-a,5 this must also be due to intracellular TNF-a
produced by the vector, suggesting that the two effects, increasedkilling at low MOI in the presence of GCV and at high MOI in theabsence of GCV, may be manifestations of the same mechanism.
4 S. M . unpublished results.* S. M., unpublished observations.
Treatment of IM-«-sensitive L929 Tumors by Combination
Suicide Gene Therapy. To determine whether the increased effectiveness of combination gene treatment evident in vitro could also be observed in vivo, we first tested the effect of intratumoral vector injectionfollowed by GCV treatment on established L929 tumors. L929 cells(1 X IO6) were implanted s.c. into the right flank of athymic nude mice,
and tumors were left to grow to a diameter of 2 mm. At this point and onalternate days thereafter, the tumors were injected with medium, THZ. 1virus, or TH:TNF (5 x IO6 pfu) for a total of nine treatments until 20
days after implantation. During this period, selected groups receivedGCV (1 mg) by i.p. injection every day from the start of vector treatment.Tumor size was monitored daily, and animals were sacrificed whentumor area exceeded 2 cm2 or if ulcérationof the injected area occurred.
Tumors in animals treated with TH:TNF plus GCV were significantly retarded in growth compared with all other groups, includingTHZ. 1 with or without GCV and TH:TNF alone (Fig. 5A). A Kaplan-
Meier survival plot (Fig. 5ß)shows the end points for the animals inthis experiment, which fell near the end of the treatment period for allgroups (median, 17-20 days), except for the group treated with TNF-a
vector plus GCV (median, 29 days). As suggested by the plot, tumorgrowth in the latter group resumed after the discontinuation of treatment (Fig. 55 and data not shown). These results demonstrated thepromise of combination TNF-a-TK gene therapy for the treatment ofTNF-a-sensitive tumors and added an incentive to test the sametreatment against TNF-a-resistant tumors.
Treatment of U-87 MG TNF-a-resistant Tumors. The primaryincentive to test our combination gene therapy against U-87 MG tumorswas the encouraging, albeit unexplained, observation earlier that vector-directed TNF-a expression could augment the killing by TK-activatedGCV of these TNF-a-resistant cells in vitro. U-87 MG cells (1 X IO5)
were stereotactically implanted into athymic nude mice at a site whereglioblastomas may develop naturally, the right frontal lobe of the brain.Three days later, the mice were randomly divided into six groups andinjected with medium or virus (1 X IO6 pfu) using the same stereotactic
coordinates as used earlier for tumor implantation. For practical reasons,vector application in this model was initially limited to a single injection.GCV was administered to selected groups by daily i.p. injection (1 mg)for a period of 10 days, starting at the day of vector application, and theanimals were observed until death.
The results in Fig. 6 readily suggest an increase in survival times foranimals injected with TH:TNF or THZ.l and treated with GCV.Median survival times for these groups were 33 and 29 days, respectively, which was significantly longer (P < 0.01, Wilcoxon test) thanfor the control groups (THZ.l only, PBS or PBS+GCV), the meansurvival times of which ranged from 22 to 23 days. However, onlytreatments involving TH:TNF+GCV were significantly differentfrom the treatments using TH:TNF (P < 0.01). Complete cures withsurvival past 80 days were observed for 1 of 12 animals in theTHZ.l/GCV group and 2 of 14 animals in the TH:TNF/GCV group.At the same time, we found that the difference in survival timesbetween these two groups was not statistically significant (P = 0.18).
A second experiment with three viral injections on alternate daysshowed essentially identical results (data not shown).
We examined the spread of viral infection after U-87 MG tumorinjection because our earlier results in vitro had indicated that TNF-a
could enhance the cytocidal activity of TK expression with GCVtreatment toward U-87 MG cells only when it was expressed intra-
cellularly. Tumor sections were prepared from animals sacrificed 3days after viral injection and were stained with antibodies directedagainst wild-type HSV-1 antigens. The results for TH:TNF-injected
animals demonstrated that only a small portion of the tumor wasinfected, with staining limited mainly to peripheral areas and fewantigen-positive cells detectable inside the tumor mass (Fig. 7,4); no
5734
on May 29, 2018. © 1998 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

TNF-a AND HSV-lk COMBINATION GENE THERAPY
Fig. 4. A, survival of TH:TNF- and THZ. 1-infected U-87 MG cells in
the presence or absence of GCV. Cells were infected at an MOI of 0.1.and survival was measured relative to untreated mock-infected cells, asbefore: bars, SD. B, TNF-a protein added to the medium of THZ. I-infected cells failed to reproduce the effect of TH:TNF infection on cellsurvival in the presence of GCV. U-87 MG cells were infected at an MOIof 0.1, and TNF-a was added at 10 ng/ml where indicated, with or
without GCV. The graph shows cell survival 5 days after infection(averages of three determinations); bars. SD. C immunoblot showingHSV-TK expression in infected U-87 MG cells. Cells were infected withTH:TNF (Lanes 1 and 4), THZ.1 (Lanes 2 and 5), or wild-type virus(KOS: Lanes 3 and 6) at MOIs of 3.0 or 0.1, and lysates were prepared24 h later for electrophoresis of equal amounts of protein (40 fig/lane) ona 10% SDS-polyacrylamide gel. electroblotting. and reaction with apolyclonal HSV-TK-specific antibody (38). The lower level of HSV-TK
expression in Lane 3 compared with Lanes 1 and 2 can be attributed toseveral causes, including the early HSV-rt promoter used for expressionby KOS virus versus the immediate-early ICP4 promoter used by the twoother viruses. D, TH:TNF infection of TNF-a-resistant U-87 MG cells athigh MOI was cytotoxic in the absence of GCV. Cells were infected at anMOI of 3.0, and relative survival with or without exposure to GCV wasdetermined as before. Unlike TH:TNF infection at low MOI (see A),infection at high MOI reduced cell survival dramatically; bars, SD.
U-87MGMOI=0.1
TH:TNF
TH:TNF+GCV B.THZ.1THZ.1+GCV
100-
S? 50-
100-
I/5J
W 50-
äS
25-
MOI=0.1
246Days post-infection
^ ? >mm
c.
HSV-TK*-
D.123456 U-87MG
MOI=3.0
MOI=3.0 MOI=0.1
o
TH:TNFTH:TNF+GCVTHZ.1THZ.1+GCV
0246Days post-infection
HSV-positive cells were detected in tumors injected with medium (Fig.
IB). These observations would suggest that enhanced tumor control byTNF-a expression in the context of HSV-TK/GCV therapy may bedemonstrable in this TNF-a-resistant brain tumor model only if a higher
level of infection can be achieved. At the same time, it is noteworthy thatHSV-TK/GCV therapy alone was beneficial in this model despite the
limited spread of the virus, which can be attributed to the bystander effect.
DISCUSSION
It has been shown previously that suicide gene therapy can be aneffective methodology for killing tumor cells in culture and in vivo(1-10). For example, the survival of rats implanted with tumor-
generating 9 L gliosarcoma cells was prolonged 1.5 times or more byHSV-TK/GCV therapy, and some rats were cured completely (4,7-10). However, the results of clinical trials indicate a need for greater
efficacy. In a recent trial, 10 glioblastoma patients were treated byintratumoral implantation of cells producing a retroviral vector expressing HSV-TK, but just one patient showed a complete responsewith long-term survival (4). We investigated the hypothesis thatcoexpression in tumor cells of the antitumor cytokine TNF-a with theHSV-rfc suicide gene could increase the efficacy of GCV-mediatedtumor cell killing. To test this hypothesis, we created a double-recombinant, replication-defective HSV-1 vector for simultaneousintratumoral delivery of the TNF-a and tk genes. We evaluated theability of this vector to kill TNF-a-sensitive and TNF-a-resistant
tumor cell lines in vitro and corresponding solid tumors in vivo incomparison with a vector that expressed only TK.
In experiments with the TNF-a-sensitive tumor cell line L929, wefound that coexpression of HSV-TK with TNF-a resulted in greater
cell killing in the presence of GCV than expression of TK alone. Thistrend was also observed in allograft experiments with athymic nudemice where established intradermal L929 tumors were treated bydirect vector injections into the tumor mass. These experiments demonstrated that animals treated with the TH:TNF vector plus GCV hadsignificantly longer survival times than animals in any of the controlgroups, including animals treated with parental HSV-iA: vector plusGCV, and the results indicated that the combination of two tumor-,
killing mechanisms provided an additive effect. In this experimentaltumor model, TNF-a could be released locally from transduced tumor
cells and be taken up by neighboring tumor cells, providing anadditional bystander effect to that of TK-activated GCV.
Because tumors are often resistant to exogenous TNF-a, it was of'
practical interest to examine the response of a representative TNF-a-resistant tumor cell line, U-87 MG, to infection with TH:TNF. In
addition, these experiments would test the assumption that the effectof vector-derived TNF-a in the L929 experiments was mediated byinteraction of secreted TNF-a with cell surface receptors. Surprisingly,
we found that TH:TNF infection with exposure to GCV showed asignificant increase in cell killing compared with THZ. 1/GCV (90%versus 50% cell killing). We confirmed that this effect was independent
5735
on May 29, 2018. © 1998 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

TNF-a AND HSV-tk COMBINATIONGENETHERAPY
30»
100-
•TH:TNF° TH:TNF+GCV•THZ.10 THZ.1-fGCV•No treatmentA GCV
5 10 15 20GCVAAAAAAAAAAAAAAAAAvimsA i A A i i i i i
Days post-Implantation
B.
50 -
•TH:TNF only (20 d.)O TH:TNF+GCV (29 d.)•THZ.1 only (20 d.)O THZ.1+GCV(21 d.)•No treatment (19 d.)A GCV(17d.)
5 10 15 20 25 30 35
GCV AAAAAAAAAAAAAAAAAvirus A A A A A A A A A
Days post-implantation
Fig. 5. Treatment of intradermal L929 tumors in alhymic nude mice. Tumors weretreated by virus injection on alternate days with or without daily i.p. administration ofGCV over a period of 17 days. A, tumor size was measured regularly, and the valuesshown represent averages for the animals in each group (n = 4); bars, SD. B. Kaplan-Meier survival plot. Mice were sacrificed when tumor area reached 2 cm2. Median
survival times for all groups are listed with the symbol key.
of cell surface receptor-mediated signaling by demonstrating that theaddition of purified recombinant TNF-a to the culture medium of THZ. 1-
infected cells failed to increase cell killing. These results indicated thatthe intracellular production of TNF-a increased cell death due to acti
vated GCV. We are presently exploring the underlying mechanism,which could involve, for example, increased formation of gap junctionsbetween tumor cells, allowing for increased transmission of activatedGCV, or inhibition of the activation of NF-«B, resulting in down-regulation of the antiapoptotic pathway of the cell (33-35).
Unlike the earlier results with L929 cells, the additive effect ofcombination treatment observed with U-87 MG cells in vitro could
not be duplicated in vivo. Independent inoculation of intracerebralU-87 MG glioblastomas with the TH:TNF and THZ.1 vectors accom
panied by regular injections of GCV clearly prolonged the survival ofthe host animals significantly, but a meaningful difference betweenthe two vectors was not observed. Because we had found that vector-directed TNF-a expression was short-lived in Vero cells, we deter
mined whether repeated vector injections would differentiate betweenthe TH:TNF and THZ.1 treatments. Regardless of whether the vectorswere injected once or three times on alternate days, the differences in theresults for the two vectors remained statistically insignificant. WhereasGCV treatment is able to reach uninfected cells that do not express theviral tk gene (bystander effect), the observation that TNF-a must be
expressed intracellularly to enhance the killing of U-87 MG cells byactivated GCV in vitro suggested that the benefits of TNF-a expression
may be limited to infected tumor cells. We reasoned, therefore, thatinefficient infection could explain the absence of a clear difference in vivobetween TH:TNF and THZ.1. Indeed, we found by immunochemicalstaining of treated U-87 MG tumor sections that < 1% of the tumor cellsexpressed HSV-1 antigens, which could be due in part to limited viral
diffusion inside tumors. Hence, a more efficient procedure for vectordelivery is required to establish whether TNF-a coexpression withHSV-TK is beneficial against TNF-a-resistant tumors. At the same time,eradication of solid tumors by any gene-therapy protocol alone is not a
realistic goal at present, and the greater promise in this area may lie inpostsurgical treatment to eliminate malignant cells left behind in theperiphery; following surgery, the target cell population is much reducedin size and presumably more accessible to vector delivery. Accordingly,experiments are now in progress to compare our vectors under theseconditions.
Local production of TNF-a is likely to have additional benefits for
antitumor therapy by mechanisms other than direct tumor cell killing:(a) Extracellular TNF-a damages endothelial cells and will cause
thrombosis in the tumor vasculature when applied locally (36). Thissensitizes the vasculature to radiation-induced collapse, and localproduction of TNF-a by virus-infected tumor cells therefore has the
potential to enhance the efficiency of radiation treatment (37, 38). Wehave stained TH:TNF-injected U-87 MG tumor sections with human-specific anti-TNF-a antibodies and observed a more widespread distribution of positive signals than with anti-HSV-1 antibodies; thespecificity for human TNF-a was confirmed by the observation thatthe TNF-a antibodies failed to stain brain tumors injected with THZ. 1(data not shown). By indicating secretion and diffusion of vector-derived TNF-a, these results suggest that infection of even a fraction
of the tumor could benefit the efficiency of radiation therapy. Moreover, vector-directed local TNF-a production should offer a clear
advantage over systemic administration because it would prevent thesevere side effects (e.g., hypotension, fever, diarrhea, and others)known to accompany systemic TNF-a delivery (24, 25).
(b) TA'F-a-transduced cells can support the induction of antitu-
mor immunity (39). For example, dendritic cells collected byleukapheresis from peripheral blood have been shown to efficientlymature in the presence of TNF-a and granulocyte/macrophagecolony-stimulating factor (40), and TNF-a expression by autolo-gous tumor cells can therefore increase the abundance of antigen-pre
senting cells required for sensitization of CTLs. Nonimmune antitumorresponses involving activated NK and lymphokine-activated killer cellscan also be induced by locally produced cytokines such as TNF-a (41),
•TH:TNF only (25 d.; n=8|D TH:TNF+GCV(33d.;n=14)•THZ.1only (22 d.; n=9)O THZ.1*GCV(29d.;n*12)A PBS (22 d.; n=6)A PBStGCV (23 d.; n=6)
» 10
GcvAAAAAAAAAAVlrusA
Days post-implantation
Fig. 6. Kaplan-Meier survival plot of athymic nude mice with intracerebral U-87 MGtumors treated by a single viral injection and daily GCV administration for 10 days. Thekey includes the number of animals per treatment (n) and median survival limes.
5736
on May 29, 2018. © 1998 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

TNF-a ANDHSV-ik COMBINATIONGENETHERAPY
12.
Fig. 7. Examination of viral transduction of U-87 MG intracerchral tumor mass.BALB/c (nii/nn) mice with implanted tumors were sacrificed and perfused with fixative.Brain sections were prepared and stained with polyclonal rabbit anti-HSV-1 antigen
antibody. A, intracerebral tumor treated with TH:TNF and GCV (2 days after infection).HSV-I antigen is detected around the tumor margin and in some cells in the tumor core
(arrowheads). B, intracerebral tumor treated with medium. No staining is observed.
and these activated cells have been shown to be effective in destructionof a variety of tumors in animal model systems (41). Whether NK cellsplayed a role in our tumor models has not been determined, but inflammatory cells were observed at the site of virus injection using immuno-histology.5 Thus, vector-directed TNF-a expression may be able to both
increase local inflammation and to recruit and activate antigen-presenting
cells that may induce antitumor immunity. Further preclinical studiesusing syngeneic tumor models where both direct tumor cell killing andthe induction of specific immunity can be evaluated should be useful indetermining whether the combination of TNF-a and tk gene therapy willprovide enhanced effectiveness for the treatment of TNF-or-resistant
tumors and by what mechanisms.
ACKNOWLEDGMENTS
We thank Richard Rouse and Kate Sullivan of the Department of MolecularGenetics and Biochemistry and Makiko Hartman of the Department of Neurology for technical assistance.
REFERENCES
1. Culver, K. M., Ram, Z.. Walbridge, S., Ishii, H., Oldfield. E. H., and Blaese, R. M.In vivo gene transfer with retroviral vector-producing cells for treatment of experimental brain tumors. Science (Washington DC), 256: 1550-1552, 1992.
2. Barba, D., Hardin. J., Ray, J.. and Gage. F. H. Thymidine kinase-mediated killing ofrat brain tumors. J. Neurosurg.. 79: 729-735, 1993.
3. Barba. D., Hardin. J., Sadelain. M.. and Gage. F. H. Development of antitumorimmunity for following thymidine kinase-mediated killing of experimental braintumors. Proc. Nail. Acad. Sci. USA, 91: 4348-4352. 1994.
4. Ram, Z., Culver, K. W.. Oshiro, E. M., Viola, J. J.. DeVroom, H. L., Otto, E., Long.Z., Chiang, Y., McGarrity, G. J.. Muul, L. M.. Katz. D.. Blaese. R. M.. and Oldfield.E. H. Therapy of malignant brain tumors by intratumoral implantation of retroviralvector-producing cells. Nat. Med.. 12: 1354-1361. 1997.
5. Okada, H.. Miyamura. K., Itoh, T.. Hagiwara. M., Wakabayashi, T.. Mizuno, M.,Colosi, P.. Kurtzman. G., and Yoshida, J. Gene therapy against an experimentalglioma using adeno-associated virus vectors. Gene Ther.. 3: 957-964, 1996.
6. Chen. S. H., Shine. H. D., Goodman, J. C. Grossman. R. G.. and Woo, S. L. Genetherapy for brain tumors: regression of experimental gliomas by adenovirus-mediatedgene transfer in vivo. Proc. Nati. Acad. Sci. USA. 91: 3054-3057. 1994.
7. Perez-Cruet, M. J., Trask, T. W.. Chen. S. H., Goodman. J. C.. Woo. S. L., Grossman,R. G., and Shine, H. D. Adenovirus-mediated gene therapy of experimental gliomas.J. Neurosci. Res., 39: 506-511. 1994.
8. Vincent. A. J.. Vogels. R.. Someren, G. V.. Esandi, M. C., Noteboom, J. L., Avezaat,C. J.. Vecht. C.. Bekkum, D. W., Valerio, D., Bout, A., and Hoogerbrugge, P. M.Herpes simplex virus thymidine kinase gene therapy for rat malignant brain tumors.Hum. Gene Ther., 7: 197-205. 1996.
9. Vincent, A. J., Esandi. M. C.. Avezaat, C. J., Vechl, C. J., Sillevis S. P.. van Bekkum.D. W.. Valerio, D., Hoogerbrugge. P. M.. and Bout, A. Preclinical testing ofrecombinant adenoviral herpes simplex virus-thymidine kinase gene therapy forcentral nervous system malignancies. Neurosurgery. 41: 442-452. 1997.
10. Boviatsis, E. J., Park, J. S., Sena-Esteves, M.. Kramm, C. M.. Chase. M., Efird, J. T.,Wei, M. X., Breakfield. X. O.. and Chiocca. E. A. Long-term survival of ratsharboring brain neoplasms treated with ganciclovir and a herpes simplex virus vectorthat retains an intact thymidine kinase gene. Cancer Res., 54: 5745-5751. 1994.
11. Yamasaki, H.. and Katoh. F. Further evidence for the involvement of gap-junctionalintercellular communication in induction and maintenance of transformed foci inBALB/c 3T3 cells. Cancer Res.. 48: 3490-3495, 1988.
16.
18.
19.
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
40.
41.
Freeman. S. M., Abbound, C. N., Whartenby. K. A.. Packman, C. H.. Koeplin. D. S..Moolten. F. L.. and Abranham. G. N. The bystander effect: tumor regression when afraction of the tumor mass is genetically modified. Cancer Res.. 53: 5274-5283, 1993.Wu. J., Cano. W. G.. Meylaerts. S. A.. Qi. P.. Vrionis, F., and Cherington. V.Bystander lumoricidal effect in the treatment of experimental brain tumors. Neurosurgery. 35: 1094-1102. 1994.Bi. W. L.. Parysek, L. M., Wamick. R.. and Stambrook, P. J. In vitro evidence thatmetabolic cooperation is responsible for the bystander effect observed with HCV TKretroviral gene therapy. Hum. Gene Ther., 4: 725-731. 1993.Mesnil. M.. Piccoli. C., Tiraby. G.. Willecke, K., and Yamasaki, H. Bystander killingof cancer cells by herpes simplex virus thymidine kinase gene is mediated byconnexins. Proc. Nati. Acad. Sci. USA. 93: 1831-1835, 1996.Han. S. K.. Brody. S. L.. and Crystal, R. G. Suppression of in vivo lumorigenicity ofhuman lung cancer cells by retrovirus-mediated transfer of the human tumor necrosisfactor-a cDNA. Am. J. Resp. Cell. Mol. Biol.. //: 270-278. 1994.Cao, G., Kuriyama, S.. Du, P., Sakamoto. T., Kong, X.. Masui, K.. and Qi, Z.Complete regression of established murine hepatocellular carcinoma by in vivo tumornecrosis factor a gene transfer. Gastroenterology, 112: 501-510, 1997.Pfizenmaier. K.. Pfizenmaier. K.. Scheurich. P.. Schluter. C., and Kronke. M. Tumornecrosis factor enhances HLA-A. B. C and HLA-DR gene expression in human tumorcells. J. Immunol., 138: 975-980, 1987.Rothlein, R., Czajkowski, M., O'Neill, M. M.. Marlin, S. D., and Merluzzi, V. J.
Induction of intercellular adhesion molecule 1 on primary and continuous cell lines bypro-inflammatory cytokines. Regulation by pharmacologie agents and neutralizingantibodies. J. Immunol.. 141: 1665-1669, 1988.Plaetinck. G.. Declercq. W., Tavemier. J., Jabholz. M., and Fiers. W. Recombinanttumor necrosis factor can induce interleukin 2 receptor expression and cytolyticactivity in a rat x mouse T cell hybrid. Eur. J. Immunol., 17: 1835-1838, 1987.Ostensen. M. E., Thiele. D. L., and Lipsky, P. E. Enhancement of human natural killercell function by the combined effects of tumor necrosis factor a or interleukin-1 andinterferon-a or interleukin-2. J. Biol. Res. Mod., 8: 53-61, 1987.Ranges. G. E., Figari, 1. S.. Espevik. T., Palladino. M. A., Jr. Inhibition of cytotoxicT cell development by transforming growth factor beta and reversal by recombinanttumor necrosis factor a. i. Exp. Med., 166: 991-998, 1987.Owen-Schaub. L. B.. Gutterman, J. U.. and Grimm, E. A. Synergy of tumor necrosis
factor and interleukin 2 in the activation of human cytotoxic lymphocytes: effect oftumor necrosis factor a and interleukin 2 in the generation of human lymphokine-aclivated killer cell cytotoxicity. Cancer Res.. 48: 788-792, 1988.Moritz, T.. Niederle, N., Baumann. J., May. D., Kurchel. E.. Osieka, R.. Kempeni, J., Schlick,E., and Schmidt. C. G. Phase 1 study of recombinant human tumor necrosis factor A inadvanced malignant disease. Cancer Immune). Immunother. 29: 144-150, 1989.Sidhu. R. S.. and Bollón.A. P. Tumor necrosis factor activities and cancer therapy-aperspective. Pharmacol. Ther.. 57: 79-128. 1993.Krisky. D. M.. Marconi, P. C.. Oligino, T., Rouse, R. J. D., Fink, D. J.. and Glorioso.J. C. Rapid method for construction of recombinant HSV gene transfer vectors. GeneTher., 4: 1120-1125. 1997.Marconi, P., Krisky. D.. Oligino. T.. Poliani. P. L.. Ramakrishnan. R.. Coins, W. F..Fink. D. J., and Glorioso. J. Replication-defective herpes simplex virus vectors forgene transfer in vivo. Proc. Nati. Acad. Sci. USA, 93: 11319-11320. 19%.Mosmann, T. Rapid calorimetrie assay for cellular growth and survival: application toproliferation and cytotoxicity assays. J. Immunol. Methods, 65: 55-63. 1983.Summers, W. P.. Wagner. M., and Summers, W. C. Possible peptide chain termination mutants in thymidine kinase gene of a mammalian virus, herpes simplex virus.Proc. Nati. Acad. Sci. USA, 10: 4081-4084. 1975.Tamura, M.. Shimizu. K.. Yamada. M.. Miyao. Y.. Hayakawa. T.. and Ikenaka. K.Targeted killing of migrating glioma cells by injection of HTK-modified glioma cells.Hum. Gene Ther., 8: 381-391. 1997.Sakuma. S.. Sawamura. Y., Tada. M., Aida. T.. Abe. H.. Suzuki. K.. and Taniguchi.N. Responses of human glioblastoma cells to human natural tumor necrosis factor-a:susceptibility, mechanism of resistance and cytokine production studies. J. Neuro-Oncol., 15: 197-208, 1993.Beyaert, R.. and Fiers. W. Molecular mechanism of tumor necrosis factor-inducedcytoloxicity. What we do understand and what we do not. FEBS Lett.. 340: 9-16. 1994.Beg, A. A., and Baltimore. D. An essential role for NF-KB in preventing TNF-or-induced cell death. Science (Washington DC). 274: 782-784, 1996.Wang, C. Y.. Mayo, M. W., and Baldwin. A. S., Jr. TNF- and cancer therapy-inducedapoptosis: potentiation by inhibition of NF-KB. Science (Washington DC), 274:784-787, 1996.Van Antwerp, D. J., Martin, S. J.. K.iln. T., Green, D. R.. and Verma. I. M. Suppressionof TNF-a-induced apoptosis by NF-xB. Science (Washington DC). 274: 787-789. 1996.Walanabe, N., Niitsu. Y., Umeno, H.. Kuriyama. H.. Neda. H.. Yamauchi. N.. Maeda.M., and Urushizaki, I. Toxic effect of tumor necrosis factor on tumor vasculature inmice. Cancer Res,. 48: 2179-2183, 1988.Gridley. D. S.. Archambeau, J. O., Andres. M. A.. Mao, X. W., Wright. K.. and Slater.J. M. Tumor necrosis factor-a enhances antitumor effects of radiation against gliomaxenografts. Oncol. Res.. 9: 217-227. 1997.Mauceri, H.. Hanna, N. H.. Wayne. J. D.. Hallahan. D. E.. Hellman. S., andWeichselbaum, R. R. Tumor necrosis factor a gene therapy targeted by ionizingradiation selectively damages tumor vasculature. Cancer Res., 56: 4311-4314. 1996.Sato, Y., Koshita, Y., Hirayama, M.. Matuyama, T.. Wakimolo, H., Hamada. H.. andNiitsu, Y. Augmented antitumor effects of killer cells induced by tumor necrosisfactor gene-transduced autologous tumor cells from gastrointestinal cancer patients.Hum. Gene Ther.. 7: 1895-1905. 1996.Caux, C., Dezutter-Dambuyant. C.. Schmitt. D.. and Banchereau. J. GM-CSF andTNF-a cooperate in the generation of dendritic Langerhans cells. Nature (Lond.).360: 258-261. 1992.Shultz. L. D.. and Sidman. C. L. Genetically determined murine models of immunodeficiency. Annu. Rev. Immunol.. 5: 367-403, 1987.
5737
on May 29, 2018. © 1998 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

1998;58:5731-5737. Cancer Res Shusuke Moriuchi, Thomas Oligino, David Krisky, et al. Thymidine Kinase
and Herpes Simplex VirusαHuman Tumor Necrosis Factor-Herpes Simplex Virus Type 1 Vector-directed Coexpression of Enhanced Tumor Cell Killing in the Presence of Ganciclovir by
Updated version
http://cancerres.aacrjournals.org/content/58/24/5731
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/58/24/5731To request permission to re-use all or part of this article, use this link
on May 29, 2018. © 1998 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from