endothelial fate and angiogenic properties of human cd34
TRANSCRIPT

Endothelial Fate and Angiogenic Properties of HumanCD34� Progenitor Cells in Zebrafish
Ombretta Pozzoli, Pietro Vella, Grazia Iaffaldano, Valeria Parente, Paolo Devanna, Marta Lacovich,Carla Lora Lamia, Umberto Fascio, Daniela Longoni, Franco Cotelli,
Maurizio C. Capogrossi, Maurizio Pesce
Objective—The vascular competence of human-derived hematopoietic progenitors for postnatal vascularization is stillpoorly characterized. It is unclear whether, in the absence of ischemia, hematopoietic progenitors participate inneovascularization and whether they play a role in new blood vessel formation by incorporating into developing vesselsor by a paracrine action.
Methods and Results—In the present study, human cord blood–derived CD34� (hCD34�) cells were transplanted into pre-and postgastrulation zebrafish embryos and in an adult vascular regeneration model induced by caudal fin amputation.When injected before gastrulation, hCD34� cells cosegregated with the presumptive zebrafish hemangioblasts,characterized by Scl and Gata2 expression, in the anterior and posterior lateral mesoderm and were involved in earlydevelopment of the embryonic vasculature. These morphogenetic events occurred without apparent lineage reprogram-ming, as shown by CD45 expression. When transplanted postgastrulation, hCD34� cells were recruited into developingvessels, where they exhibited a potent paracrine proangiogenic action. Finally, hCD34� cells rescued vascular defectsinduced by Vegf-c in vivo targeting and enhanced vascular repair in the zebrafish fin amputation model.
Conclusion—These results indicate an unexpected developmental ability of human-derived hematopoietic progenitors andsupport the hypothesis of an evolutionary conservation of molecular pathways involved in endothelial progenitordifferentiation in vivo. (Arterioscler Thromb Vasc Biol. 2011;31:1589-1597.)
Key Words: endothelium � vascular biology � angiogenesis � embryology � stem cells
It has long been supposed that cells present in the bonemarrow, peripheral blood (PB), and cord blood (CB),
which copurify with hematopoietic stem cells (HSCs), alsogive rise to endothelial cells. This speculation was supportedby the notion that CD34� and CD133� cells differentiate, inculture and in vivo, into cells that express mature endothelialcell markers. These cells, called endothelial progenitor cells(EPCs),1–3 have been the subject of numerous basic andtranslational studies showing their participation in neovascu-larization. Recent studies have revealed possible separationbetween hematopoietic- and nonhematopoietic-derivedEPCs.4–8 These 2 cell types have been called early and lateEPCs,9 or colony-forming unit–endothelial cells (CFU-ECs)and endothelial colony-forming cells (ECFCs).7 They arefundamentally distinguished on the basis of hematopoieticmarker expression (eg, CD45) and the ability to proliferate or
to differentiate into endothelial cells.10 Separation betweenhematopoietic and the endothelial lineages during earlymouse11 and zebrafish12 development is established during byasymmetrical division of primitive cells located in the dorsalaorta endothelium, through a novel differentiation eventcalled endothelial-hematopoietic transition (EHT).13–16 Moreuncertain is whether cells with a similar potency of generat-ing HSCs or endothelial cells are present at postnatal stages17;recently, however, derivation of endothelial cells fromCD34�/CD38�/CD45�/CD133� CB progenitors was dem-onstrated,18 suggesting the existence of similar progenitors atleast during fetal life.
The zebrafish (Danio rerio) embryo provides severaladvantages for in vivo study of vascular morphogenesis,based on the opportunity to observe in real time the devel-opment of embryonic blood vessels19,20 and also on the
Received on: June 21, 2010; final version accepted on: March 31, 2011.From the Laboratorio di Biologia Vascolare e Medicina Rigenerativa, Centro Cardiologico Monzino, Istituto di Ricovero e Cura a Carattere Scientifico,
Milan, Italy (O.P., P.V., G.I., V.P., P.D., M.L., M.P.); Laboratorio di Patologia Vascolare, Istituto Dermopatico dell’Immacolata, Istituto di Ricovero eCura a Carattere Scientifico, Rome, Italy (M.C.C.); Laboratorio di Biologia dello Sviluppo (C.L.L., F.C.) and Centro Interdipartimentale MicroscopiaAvanzata (U.F.), Universita di Milano, Milan, Italy; Centro Ricerche Tettamanti, Monza, Italy (D.L.).
Drs Capogrossi and Pesce contributed equally to this work.Correspondence to Maurizio Pesce, PhD, Laboratorio di Biologia Vascolare e Medicina Rigenerativa, Centro Cardiologico Monzino, Istituto di
Ricovero e Cura a Carattere Scientifico, Via Parea 4, 20138 Milan, Italy; (E-mail [email protected]) Ombretta Pozzoli, PhD, Laboratorio diBiologia Vascolare e Medicina Rigenerativa, Centro Cardiologico Monzino, Istituto di Ricovero e Cura a Carattere Scientifico, Via Parea 4, 20138 Milan,Italy; (E-mail [email protected]) or Franco Cotelli, Laboratorio di Biologia dello Sviluppo, Universita degli Studi di Milano, Via Celoria 26,20133 Milan, Italy (E-mail [email protected]).
© 2011 American Heart Association, Inc.
Arterioscler Thromb Vasc Biol is available at http://atvb.ahajournals.org DOI: 10.1161/ATVBAHA.111.226969
1589
by guest on March 16, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
arch 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on March 16, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
arch 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on March 16, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
arch 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on March 16, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
arch 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on March 16, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
arch 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on March 16, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
arch 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on March 16, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
arch 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on March 16, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
arch 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on March 16, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
arch 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on March 16, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
arch 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on March 16, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
arch 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on March 16, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
arch 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on March 16, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
arch 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on March 16, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
arch 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on March 16, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
arch 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on March 16, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
arch 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on March 16, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
arch 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on March 16, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
arch 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from

remarkable conservation of molecular pathways regulatingvascular development in mammals, such as those regulatedby endothelial (eg, Flk-1) or hematopoietic (eg, Scl, Runx1)master genes.21–23
The objective of the present work was to explore thedevelopmental potency and the vascular competence of hu-man CB CD34� cells, exploiting the flexibility offered byzebrafish system.
Materials and MethodsAn expanded supplemental version of the Methods section isavailable online at http://atvb.ahajournals.org.
Ethical StatementCollection of CB samples was performed with written consent on avoluntary basis and after approval by institutional review boards.Authorization numbers are provided in the supplemental material.
Fish MaintenanceBreeding zebrafish, wild-type AB strain, transgenic (Tg) strainTg(KDRI:enhanced green fluorescent protein [EGFP])S843 (ZIRC,University of Oregon), and the Tg(fli1:EGFP)y1 line (from thelaboratory of Nathan Lawson, University of Massachusetts MedicalSchool) were maintained at 28°C on a 14-hour light/10-hour darkcycle, according to established procedures.
hCD34� and hCD14� Cell Selectionand CharacterizationIsolation of human CB–derived CD34� (hCD34�) and CD14� cellswas performed by using magnetic selection with a MINI-MACSsystem after separation of CB and PB mononuclear cells onto densitygradients. hCD34� cell purity (87.4�2.9%; n�9) was assessed byflow cytometry in preliminary experiments. Before injection, cellswere labeled using Orange Cell Tracker (Molecular Probes, Invitro-gen) according to the manufacturer’s instruction.
Cell Transplantation Assays and MorpholinoRNA InjectionBlastula stage or tricaine (0.04 mg/mL, Sigma) anesthetized 48 hourspost fertilization (hpf) stage AB or Tg(fli1:EGFP)y1 zebrafish strains
embryos were injected with Orange Cell Tracker–labeled CD34�,CD14� cells (4 nL injection volume, 500 to 1000 cells/embryo), orVegf-c morpholino (MO) RNA.
Embryo Sectioning and Confocal MicroscopyFor tissue sectioning, CD34� cell-injected embryos were fixed with4% paraformaldehyde, embedded in cryostat mounting medium(Killik, Bioptica), and transversally cut (8 �m) at different sectioningplanes in the regions of the head and the dorsal artery/dorsal aortalocations for immunofluorescence.
Whole-Mount Alkaline PhosphataseStaining, Immunodetection Assays, andIn Situ HybridizationZebrafish control and cell-injected embryos were fixed at theindicated time points using 4% paraformaldehyde for 2 hours atroom temperature, after which they were processed for in situhybridization, alkaline phosphatase staining, terminal deoxynucleo-tidyl transferase dUTP nick-end labeling, or phosphohistone H3whole-mount immunofluorescence.
Statistical AnalysisStatistical analysis was performed by unpaired t test or 1-wayANOVA with Newman-Keuls post hoc analysis using Prism GraphPadstatistical software; each experiment was carried out in at least intriplicate. Data are indicated as mean�SE.
ResultsFate of hCD34� Progenitors Transplanted Into theDeveloping Zebrafish BlastulaOrange Cell Tracker–labeled hCD34� cells were transplantedin Tg(fli1:EGFP)y1 embryos before the onset of gastrulation,at midblastula stage. The distribution of hCD34� cells in thedeveloping embryos was analyzed immediately after injec-tion and at tailbud and somite stages. Initially, cells remainedmostly localized at the injection site at the embryo animalpole (Figure 1A). From tailbud stage to somite stage, anumber of human cells were found in the midline, the head
Figure 1. Early segregation of hCD34�
cells after injection into the zebrafishblastula. Tg(fli1:enhanced green fluores-cent protein [EGFP])y1 embryos wereinjected with Orange Cell Tracker–labeledhCD34� cells at the blastula stage. A,Injected gastrula at 50% epiboly (top left),tailbud (top right), 14-somite-stage (bot-tom left) and 18-somite-stage (bottomright) injected embryo. Images result frommerging of bright-field and rhodaminefluorescence pictures indicating injectedhuman cells (arrows). The 18-somiteinjected embryo image results from merg-ing dark-field green and red fluorescencepictures, representing the embryonic vas-culature and human cells, respectively. Inall pictures, a, d, and p indicate the ante-rior, dorsal, and posterior sides of theembryos, respectively; ICM, intermediatecell mass. B and C, Different distributionof blastula-injected hCD34� (B) andhCD14� (C) cells in 3- to 5-somite-stagedeveloping embryos (left) and expressionof Gata2 and Scl transcripts (right). Pic-tures showing human cells were obtained
by merging rhodamine fluorescence with bright-field images of the injected embryos. The same transplanted embryos are shown onthe right after in situ detection of Gata2 and Scl transcripts. In all pictures, embryos are oriented with the anterior (a) on the right sideof each picture. hm indicates head mesenchyme.
1590 Arterioscler Thromb Vasc Biol July 2011
by guest on March 16, 2018
http://atvb.ahajournals.org/D
ownloaded from
mesenchyme, and the intermediate cell mass regions (Figure1A). In situ hybridization using riboprobes to detect Scl andGata2 mRNAs were then performed. The results revealed astrict association of human cells with multipotent Scl� andGata2� mesoderm cells (Figure 1B), suggesting cosegrega-tion of human cells with zebrafish hemangioblasts. Thisbehavior was specific; in fact, PB cells devoid of the CD14�
monocyte cell population, having proangiogenic activity,24
were not associated with Scl� and Gata2� regions (Figure 1Cand Supplemental Figure I).
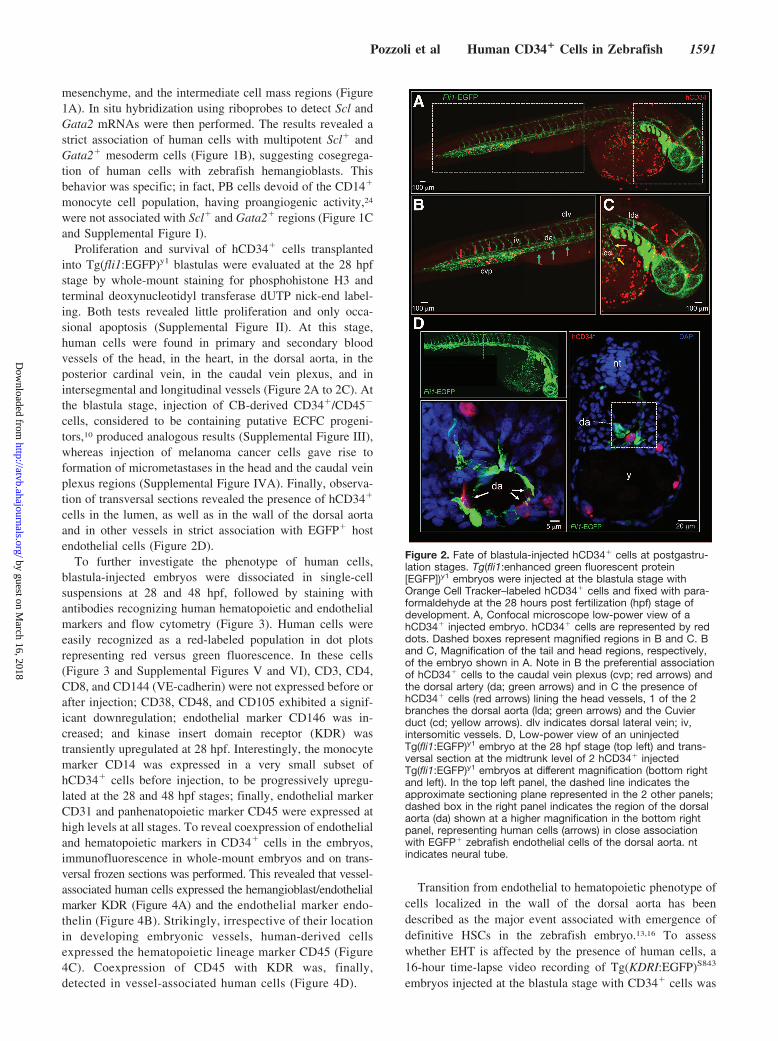
Proliferation and survival of hCD34� cells transplantedinto Tg(fli1:EGFP)y1 blastulas were evaluated at the 28 hpfstage by whole-mount staining for phosphohistone H3 andterminal deoxynucleotidyl transferase dUTP nick-end label-ing. Both tests revealed little proliferation and only occa-sional apoptosis (Supplemental Figure II). At this stage,human cells were found in primary and secondary bloodvessels of the head, in the heart, in the dorsal aorta, in theposterior cardinal vein, in the caudal vein plexus, and inintersegmental and longitudinal vessels (Figure 2A to 2C). Atthe blastula stage, injection of CB-derived CD34�/CD45�
cells, considered to be containing putative ECFC progeni-tors,10 produced analogous results (Supplemental Figure III),whereas injection of melanoma cancer cells gave rise toformation of micrometastases in the head and the caudal veinplexus regions (Supplemental Figure IVA). Finally, observa-tion of transversal sections revealed the presence of hCD34�
cells in the lumen, as well as in the wall of the dorsal aortaand in other vessels in strict association with EGFP� hostendothelial cells (Figure 2D).
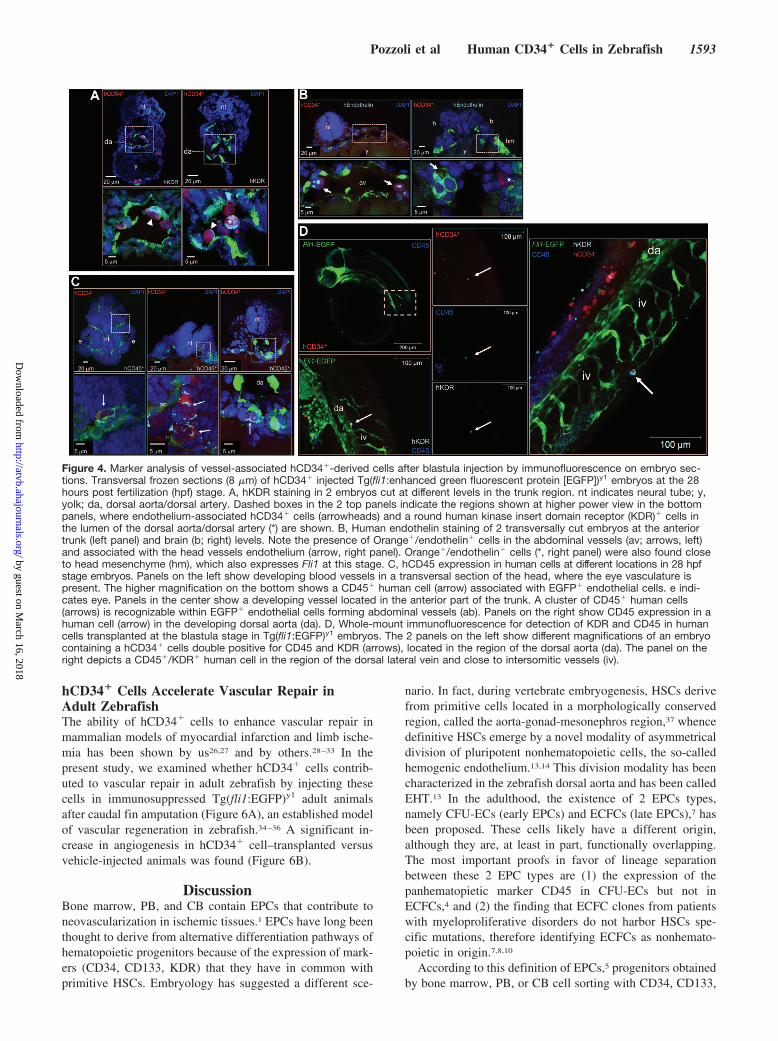
To further investigate the phenotype of human cells,blastula-injected embryos were dissociated in single-cellsuspensions at 28 and 48 hpf, followed by staining withantibodies recognizing human hematopoietic and endothelialmarkers and flow cytometry (Figure 3). Human cells wereeasily recognized as a red-labeled population in dot plotsrepresenting red versus green fluorescence. In these cells(Figure 3 and Supplemental Figures V and VI), CD3, CD4,CD8, and CD144 (VE-cadherin) were not expressed before orafter injection; CD38, CD48, and CD105 exhibited a signif-icant downregulation; endothelial marker CD146 was in-creased; and kinase insert domain receptor (KDR) wastransiently upregulated at 28 hpf. Interestingly, the monocytemarker CD14 was expressed in a very small subset ofhCD34� cells before injection, to be progressively upregu-lated at the 28 and 48 hpf stages; finally, endothelial markerCD31 and panhenatopoietic marker CD45 were expressed athigh levels at all stages. To reveal coexpression of endothelialand hematopoietic markers in CD34� cells in the embryos,immunofluorescence in whole-mount embryos and on trans-versal frozen sections was performed. This revealed that vessel-associated human cells expressed the hemangioblast/endothelialmarker KDR (Figure 4A) and the endothelial marker endo-thelin (Figure 4B). Strikingly, irrespective of their locationin developing embryonic vessels, human-derived cellsexpressed the hematopoietic lineage marker CD45 (Figure4C). Coexpression of CD45 with KDR was, finally,detected in vessel-associated human cells (Figure 4D).
Transition from endothelial to hematopoietic phenotype ofcells localized in the wall of the dorsal aorta has beendescribed as the major event associated with emergence ofdefinitive HSCs in the zebrafish embryo.13,16 To assesswhether EHT is affected by the presence of human cells, a16-hour time-lapse video recording of Tg(KDRI:EGFP)S843
embryos injected at the blastula stage with CD34� cells was
Figure 2. Fate of blastula-injected hCD34� cells at postgastru-lation stages. Tg(fli1:enhanced green fluorescent protein[EGFP])y1 embryos were injected at the blastula stage withOrange Cell Tracker–labeled hCD34� cells and fixed with para-formaldehyde at the 28 hours post fertilization (hpf) stage ofdevelopment. A, Confocal microscope low-power view of ahCD34� injected embryo. hCD34� cells are represented by reddots. Dashed boxes represent magnified regions in B and C. Band C, Magnification of the tail and head regions, respectively,of the embryo shown in A. Note in B the preferential associationof hCD34� cells to the caudal vein plexus (cvp; red arrows) andthe dorsal artery (da; green arrows) and in C the presence ofhCD34� cells (red arrows) lining the head vessels, 1 of the 2branches the dorsal aorta (lda; green arrows) and the Cuvierduct (cd; yellow arrows). dlv indicates dorsal lateral vein; iv,intersomitic vessels. D, Low-power view of an uninjectedTg(fli1:EGFP)y1 embryo at the 28 hpf stage (top left) and trans-versal section at the midtrunk level of 2 hCD34� injectedTg(fli1:EGFP)y1 embryos at different magnification (bottom rightand left). In the top left panel, the dashed line indicates theapproximate sectioning plane represented in the 2 other panels;dashed box in the right panel indicates the region of the dorsalaorta (da) shown at a higher magnification in the bottom rightpanel, representing human cells (arrows) in close associationwith EGFP� zebrafish endothelial cells of the dorsal aorta. ntindicates neural tube.
Pozzoli et al Human CD34� Cells in Zebrafish 1591
by guest on March 16, 2018
http://atvb.ahajournals.org/D
ownloaded from

performed. As shown in Supplemental Figure VII and Sup-plemental Movie I, budding of KDR� HSCs was not per-turbed by transplanted cells.
Postgastrula Stage Transplantation of hCD34�
Cells Modulates Embryonic Vascular DevelopmentThe behavior of hCD34� cells injected into the circulatorysystem at the 48 hpf stage Tg(fli1:EGFP)y1 embryos wasassessed by confocal time-lapse analysis. Early homingevents in the blood vessels were observed as early as 30minutes after injection (Supplemental Figure VIIA). hCD34�
cell rolling and adhesion to zebrafish intersegmental vesselswere also observed (Supplemental Movie II).
Transgenic embryos injected with hCD34� cells at the 48 hpfstage were analyzed at the 72 hpf stage, ie, 24 hours afterinjection. Altered blood vessel sprouting in the growing tailvasculature (85% of 72 transplanted embryos; SupplementalFigure VIIIB) and at the level of the subintestinal vein (80% of17 injected embryos; Supplemental Figure VIIIC) were ob-served. Angiogenesis abnormalities were never found afterinjection of PB-derived hCD14� cells (Supplemental FigureVIIID) but were detected in embryos transplanted with theintestinal cancer cell line CACO2 (Supplemental Figure IVB).To further examine whether blood vessel development wasperturbed by transplanted hCD34� cells, the vascular develop-ment of 48 hpf stage Tg(fli1:EGFP)y1 injected embryos wasmonitored from 2 to 12 hours postinjection by confocal video-microscopy. A strict association was found between red-labeledcells and the regions of ectopic vascular sprouting (Supplemen-tal Figure IX and Supplemental Movie III).
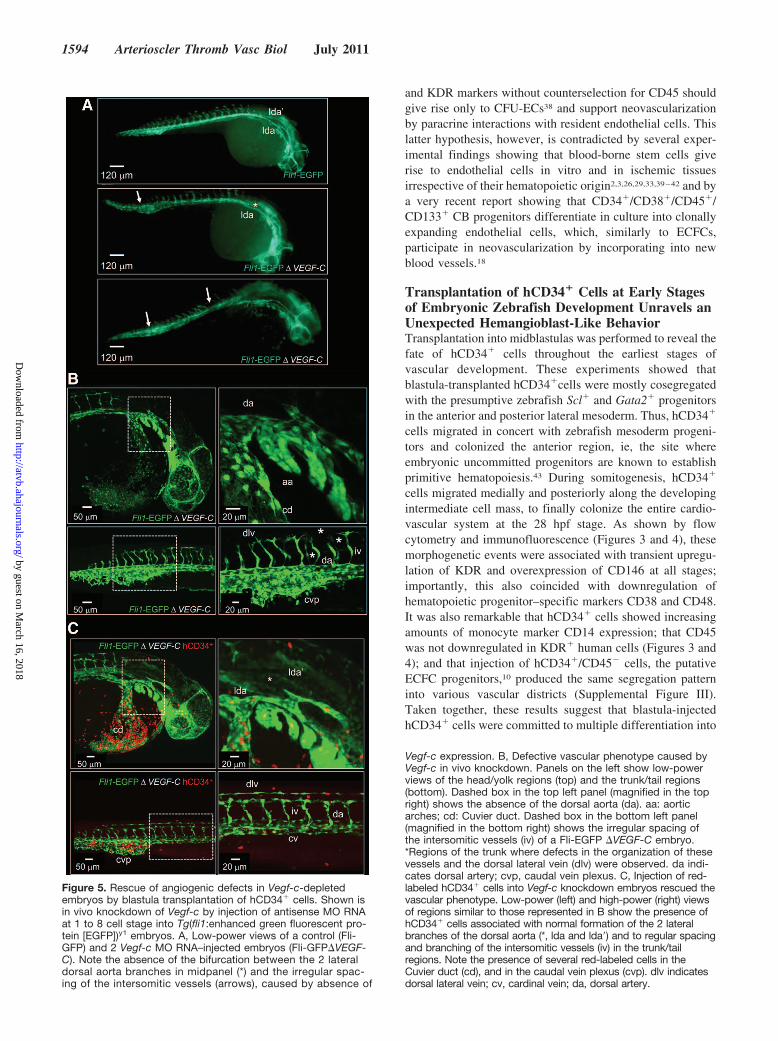
Transplanted hCD34� Cells Rescue DefectiveAngiogenesis in the Zebrafish Embryo byMultifactorial Paracrine EffectThe previous data suggested that transplanted hCD34�
cells have paracrine effects in the developing embryos. Toassess this hypothesis, cells were injected at the blastulastage in Tg(fli1:EGFP)y1 embryos, where Vegf-c wasknocked down by MO RNA injection at the 1- to 4-cellstage. As shown in Figure 5A, embryos depleted of Vegf-cexhibited defects in the development of the vascularsystem (30% of 50 MO RNA-injected embryos). Vegf-cdepletion mostly affected the formation of the dorsal aorta,the posterior cardinal vein, the intersegmental vessels, andspecific head arteries25 (Figure 5B). These defects wereobserved in only 9% (n�32) of Vegf-c-depleted/hCD34�
cell-injected embryos (Figure 5C). Interestingly, pheno-type rescue was complete even in vascular regions whereCD34� cells failed to localize, suggesting complementa-tion of Vegf-c knockdown defects by soluble factorssecreted by human cells. Finally, hCD14� cell injection inVegf-c-depleted embryos did not rescue defective bloodvessel formation (100% of vascular abnormalities, n�5embryos injected with Vegf-c MO RNA and hCD14� cells;data not shown). To identify paracrine factors participatingin the rescue effect by injected cells, the secretion ofinterleukins, chemokines, and angiogenic growth factorswas assessed by Luminex assay (Table). The resultsshowed production of several factors, including chemo-kines, proinflammatory, and proangiogenic cytokines, sug-gesting a potent paracrine effect by hCD34� cells.
Figure 3. Flow cytometry analysis of blastula-injected hCD34� cells phenotype at 28 and 48 hours post fertilization (hpf). Tg(fli1:enhancedgreen fluorescent protein [EGFP])y1 embryos were injected with hCD34� cells and then dissociated and labeled with human anti-bodies recognizing hematopoietic and endothelial cell markers for analysis by flow cytometry. A to C, Human cells were recog-nized in green vs red fluorescence dot plots by comparison with uninjected embryos. These cells were logically gated for their redfluorescence by use of 488 nm (A) and 561 nm (C) lasers and analyzed for the expression of each of the human markers. B,Quantification of results shown in A and C for hematopoietic (top) and endothelial (bottom) markers in hCD34� cells before injec-tion (open bars) and at 28 (gray bars) 48 hpf (black bars). *P�0.05 by 1-way ANOVA with Newman-Keuls post hoc analysis; n�3.Orange Tracker indicates Orange Cell Tracker.
1592 Arterioscler Thromb Vasc Biol July 2011
by guest on March 16, 2018
http://atvb.ahajournals.org/D
ownloaded from
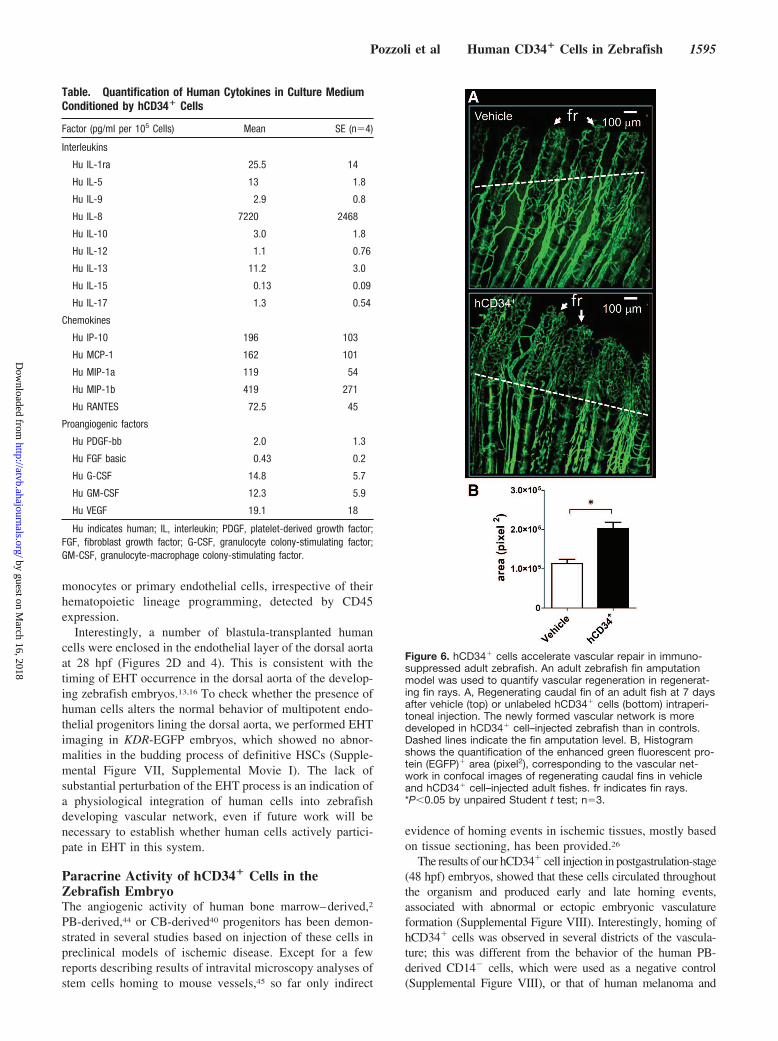
hCD34� Cells Accelerate Vascular Repair inAdult ZebrafishThe ability of hCD34� cells to enhance vascular repair inmammalian models of myocardial infarction and limb ische-mia has been shown by us26,27 and by others.28–33 In thepresent study, we examined whether hCD34� cells contrib-uted to vascular repair in adult zebrafish by injecting thesecells in immunosuppressed Tg(fli1:EGFP)y1 adult animalsafter caudal fin amputation (Figure 6A), an established modelof vascular regeneration in zebrafish.34–36 A significant in-crease in angiogenesis in hCD34� cell–transplanted versusvehicle-injected animals was found (Figure 6B).
DiscussionBone marrow, PB, and CB contain EPCs that contribute toneovascularization in ischemic tissues.1 EPCs have long beenthought to derive from alternative differentiation pathways ofhematopoietic progenitors because of the expression of mark-ers (CD34, CD133, KDR) that they have in common withprimitive HSCs. Embryology has suggested a different sce-
nario. In fact, during vertebrate embryogenesis, HSCs derivefrom primitive cells located in a morphologically conservedregion, called the aorta-gonad-mesonephros region,37 whencedefinitive HSCs emerge by a novel modality of asymmetricaldivision of pluripotent nonhematopoietic cells, the so-calledhemogenic endothelium.13,14 This division modality has beencharacterized in the zebrafish dorsal aorta and has been calledEHT.13 In the adulthood, the existence of 2 EPCs types,namely CFU-ECs (early EPCs) and ECFCs (late EPCs),7 hasbeen proposed. These cells likely have a different origin,although they are, at least in part, functionally overlapping.The most important proofs in favor of lineage separationbetween these 2 EPC types are (1) the expression of thepanhematopietic marker CD45 in CFU-ECs but not inECFCs,4 and (2) the finding that ECFC clones from patientswith myeloproliferative disorders do not harbor HSCs spe-cific mutations, therefore identifying ECFCs as nonhemato-poietic in origin.7,8,10
According to this definition of EPCs,5 progenitors obtainedby bone marrow, PB, or CB cell sorting with CD34, CD133,
Figure 4. Marker analysis of vessel-associated hCD34�-derived cells after blastula injection by immunofluorescence on embryo sec-tions. Transversal frozen sections (8 �m) of hCD34� injected Tg(fli1:enhanced green fluorescent protein [EGFP])y1 embryos at the 28hours post fertilization (hpf) stage. A, hKDR staining in 2 embryos cut at different levels in the trunk region. nt indicates neural tube; y,yolk; da, dorsal aorta/dorsal artery. Dashed boxes in the 2 top panels indicate the regions shown at higher power view in the bottompanels, where endothelium-associated hCD34� cells (arrowheads) and a round human kinase insert domain receptor (KDR)� cells inthe lumen of the dorsal aorta/dorsal artery (*) are shown. B, Human endothelin staining of 2 transversally cut embryos at the anteriortrunk (left panel) and brain (b; right) levels. Note the presence of Orange�/endothelin� cells in the abdominal vessels (av; arrows, left)and associated with the head vessels endothelium (arrow, right panel). Orange�/endothelin� cells (*, right panel) were also found closeto head mesenchyme (hm), which also expresses Fli1 at this stage. C, hCD45 expression in human cells at different locations in 28 hpfstage embryos. Panels on the left show developing blood vessels in a transversal section of the head, where the eye vasculature ispresent. The higher magnification on the bottom shows a CD45� human cell (arrow) associated with EGFP� endothelial cells. e indi-cates eye. Panels in the center show a developing vessel located in the anterior part of the trunk. A cluster of CD45� human cells(arrows) is recognizable within EGFP� endothelial cells forming abdominal vessels (ab). Panels on the right show CD45 expression in ahuman cell (arrow) in the developing dorsal aorta (da). D, Whole-mount immunofluorescence for detection of KDR and CD45 in humancells transplanted at the blastula stage in Tg(fli1:EGFP)y1 embryos. The 2 panels on the left show different magnifications of an embryocontaining a hCD34� cells double positive for CD45 and KDR (arrows), located in the region of the dorsal aorta (da). The panel on theright depicts a CD45�/KDR� human cell in the region of the dorsal lateral vein and close to intersomitic vessels (iv).
Pozzoli et al Human CD34� Cells in Zebrafish 1593
by guest on March 16, 2018
http://atvb.ahajournals.org/D
ownloaded from
and KDR markers without counterselection for CD45 shouldgive rise only to CFU-ECs38 and support neovascularizationby paracrine interactions with resident endothelial cells. Thislatter hypothesis, however, is contradicted by several exper-imental findings showing that blood-borne stem cells giverise to endothelial cells in vitro and in ischemic tissuesirrespective of their hematopoietic origin2,3,26,29,33,39–42 and bya very recent report showing that CD34�/CD38�/CD45�/CD133� CB progenitors differentiate in culture into clonallyexpanding endothelial cells, which, similarly to ECFCs,participate in neovascularization by incorporating into newblood vessels.18
Transplantation of hCD34� Cells at Early Stagesof Embryonic Zebrafish Development Unravels anUnexpected Hemangioblast-Like BehaviorTransplantation into midblastulas was performed to reveal thefate of hCD34� cells throughout the earliest stages ofvascular development. These experiments showed thatblastula-transplanted hCD34�cells were mostly cosegregatedwith the presumptive zebrafish Scl� and Gata2� progenitorsin the anterior and posterior lateral mesoderm. Thus, hCD34�
cells migrated in concert with zebrafish mesoderm progeni-tors and colonized the anterior region, ie, the site whereembryonic uncommitted progenitors are known to establishprimitive hematopoiesis.43 During somitogenesis, hCD34�
cells migrated medially and posteriorly along the developingintermediate cell mass, to finally colonize the entire cardio-vascular system at the 28 hpf stage. As shown by flowcytometry and immunofluorescence (Figures 3 and 4), thesemorphogenetic events were associated with transient upregu-lation of KDR and overexpression of CD146 at all stages;importantly, this also coincided with downregulation ofhematopoietic progenitor–specific markers CD38 and CD48.It was also remarkable that hCD34� cells showed increasingamounts of monocyte marker CD14 expression; that CD45was not downregulated in KDR� human cells (Figures 3 and4); and that injection of hCD34�/CD45� cells, the putativeECFC progenitors,10 produced the same segregation patterninto various vascular districts (Supplemental Figure III).Taken together, these results suggest that blastula-injectedhCD34� cells were committed to multiple differentiation into
Figure 5. Rescue of angiogenic defects in Vegf-c-depletedembryos by blastula transplantation of hCD34� cells. Shown isin vivo knockdown of Vegf-c by injection of antisense MO RNAat 1 to 8 cell stage into Tg(fli1:enhanced green fluorescent pro-tein [EGFP])y1 embryos. A, Low-power views of a control (Fli-GFP) and 2 Vegf-c MO RNA–injected embryos (Fli-GFP�VEGF-C). Note the absence of the bifurcation between the 2 lateraldorsal aorta branches in midpanel (*) and the irregular spac-ing of the intersomitic vessels (arrows), caused by absence of
Vegf-c expression. B, Defective vascular phenotype caused byVegf-c in vivo knockdown. Panels on the left show low-powerviews of the head/yolk regions (top) and the trunk/tail regions(bottom). Dashed box in the top left panel (magnified in the topright) shows the absence of the dorsal aorta (da). aa: aorticarches; cd: Cuvier duct. Dashed box in the bottom left panel(magnified in the bottom right) shows the irregular spacing ofthe intersomitic vessels (iv) of a Fli-EGFP �VEGF-C embryo.*Regions of the trunk where defects in the organization of thesevessels and the dorsal lateral vein (dlv) were observed. da indi-cates dorsal artery; cvp, caudal vein plexus. C, Injection of red-labeled hCD34� cells into Vegf-c knockdown embryos rescued thevascular phenotype. Low-power (left) and high-power (right) viewsof regions similar to those represented in B show the presence ofhCD34� cells associated with normal formation of the 2 lateralbranches of the dorsal aorta (*, lda and lda�) and to regular spacingand branching of the intersomitic vessels (iv) in the trunk/tailregions. Note the presence of several red-labeled cells in theCuvier duct (cd), and in the caudal vein plexus (cvp). dlv indicatesdorsal lateral vein; cv, cardinal vein; da, dorsal artery.
1594 Arterioscler Thromb Vasc Biol July 2011
by guest on March 16, 2018
http://atvb.ahajournals.org/D
ownloaded from
monocytes or primary endothelial cells, irrespective of theirhematopoietic lineage programming, detected by CD45expression.
Interestingly, a number of blastula-transplanted humancells were enclosed in the endothelial layer of the dorsal aortaat 28 hpf (Figures 2D and 4). This is consistent with thetiming of EHT occurrence in the dorsal aorta of the develop-ing zebrafish embryos.13,16 To check whether the presence ofhuman cells alters the normal behavior of multipotent endo-thelial progenitors lining the dorsal aorta, we performed EHTimaging in KDR-EGFP embryos, which showed no abnor-malities in the budding process of definitive HSCs (Supple-mental Figure VII, Supplemental Movie I). The lack ofsubstantial perturbation of the EHT process is an indication ofa physiological integration of human cells into zebrafishdeveloping vascular network, even if future work will benecessary to establish whether human cells actively partici-pate in EHT in this system.
Paracrine Activity of hCD34� Cells in theZebrafish EmbryoThe angiogenic activity of human bone marrow–derived,2
PB-derived,44 or CB-derived40 progenitors has been demon-strated in several studies based on injection of these cells inpreclinical models of ischemic disease. Except for a fewreports describing results of intravital microscopy analyses ofstem cells homing to mouse vessels,45 so far only indirect
evidence of homing events in ischemic tissues, mostly basedon tissue sectioning, has been provided.26
The results of our hCD34� cell injection in postgastrulation-stage(48 hpf) embryos, showed that these cells circulated throughoutthe organism and produced early and late homing events,associated with abnormal or ectopic embryonic vasculatureformation (Supplemental Figure VIII). Interestingly, homing ofhCD34� cells was observed in several districts of the vascula-ture; this was different from the behavior of the human PB-derived CD14� cells, which were used as a negative control(Supplemental Figure VIII), or that of human melanoma and
Table. Quantification of Human Cytokines in Culture MediumConditioned by hCD34� Cells
Factor (pg/ml per 105 Cells) Mean SE (n�4)
Interleukins
Hu IL-1ra 25.5 14
Hu IL-5 13 1.8
Hu IL-9 2.9 0.8
Hu IL-8 7220 2468
Hu IL-10 3.0 1.8
Hu IL-12 1.1 0.76
Hu IL-13 11.2 3.0
Hu IL-15 0.13 0.09
Hu IL-17 1.3 0.54
Chemokines
Hu IP-10 196 103
Hu MCP-1 162 101
Hu MIP-1a 119 54
Hu MIP-1b 419 271
Hu RANTES 72.5 45
Proangiogenic factors
Hu PDGF-bb 2.0 1.3
Hu FGF basic 0.43 0.2
Hu G-CSF 14.8 5.7
Hu GM-CSF 12.3 5.9
Hu VEGF 19.1 18
Hu indicates human; IL, interleukin; PDGF, platelet-derived growth factor;FGF, fibroblast growth factor; G-CSF, granulocyte colony-stimulating factor;GM-CSF, granulocyte-macrophage colony-stimulating factor.
Figure 6. hCD34� cells accelerate vascular repair in immuno-suppressed adult zebrafish. An adult zebrafish fin amputationmodel was used to quantify vascular regeneration in regenerat-ing fin rays. A, Regenerating caudal fin of an adult fish at 7 daysafter vehicle (top) or unlabeled hCD34� cells (bottom) intraperi-toneal injection. The newly formed vascular network is moredeveloped in hCD34� cell–injected zebrafish than in controls.Dashed lines indicate the fin amputation level. B, Histogramshows the quantification of the enhanced green fluorescent pro-tein (EGFP)� area (pixel2), corresponding to the vascular net-work in confocal images of regenerating caudal fins in vehicleand hCD34� cell–injected adult fishes. fr indicates fin rays.*P�0.05 by unpaired Student t test; n�3.
Pozzoli et al Human CD34� Cells in Zebrafish 1595
by guest on March 16, 2018
http://atvb.ahajournals.org/D
ownloaded from

CACO-2 stem cells, whose presence was mostly associated withthe head; the caudal vein plexus, where they formed microme-tastases; or the yolk region, where they caused ectopic vesselsprouting (Supplemental Figure IV).46
Direct participation in primary angiogenesis was not theonly effect of hCD34� cell injection into developing ze-brafish embryos. In fact, hCD34� cells rescued vasculardefects caused by Vegf-c knockdown (Figure 6), and whentransplanted into immunosuppressed adults, they acceleratedregeneration of the vascular network in the amputated caudalfins. This activity was due to secretion of several cytokinesand chemokines implicated in (lympho)angiogenesis andinflammatory response (Supplemental Table I). Taken to-gether, and in line with the large existing literature on EPCs,these results suggest that transplanted hCD34� cells exert apotent paracrine effect47 that rescues vascular developmentaldefects due to knockdown of Vegf-C, deregulates formationof normal vascular network in postgastrulation zebrafishembryos, and accelerates wound repair in adults.
Concluding RemarksThe identity of human-derived endothelial progenitors is astill open issue. Although the distinction between the 2 EPCtypes10 has allowed reconciliation of the embryo with theadult stem cell worlds, it is still not clear whether, underpathological conditions or in developmental biology contexts,these cells may cross their lineage barriers and noncanoni-cally differentiate into alternative or mixed phenotypes. Ourfindings suggest that this might be the case and call for furtherinvestigation aimed at clarifying the molecular nature of thesepossible events.
To our knowledge, the present study represents the firstexample of successful human stem cell transplantation intodeveloping zebrafish embryos. In fact, apart from previousinvestigations showing the fate of human cancer cells in thedeveloping zebrafish48 and the recent demonstration of an-giogenic activity of mouse and human tumor cell lines,46 noother evidence exists that human stem cells colonize thedeveloping zebrafish embryos and respond to specific devel-opmental cues. In this context, our results may also haveimportant translational implication for in utero correction ofvascular defects in human embryos by injected stem cells.
It has been discussed that the developing zebrafish is aninteresting microenvironment in which modulatory effects onthe fate of injected cells are likely due to the presence ofdevelopmental mechanisms that are conserved among verte-brates.48 In addition, given the relative ease of zebrafishgenetic manipulation systems, xenotransplantation of humancells in zebrafish has been proposed as a reference techniqueto define the properties of human cancer (stem) cells.49 Ourstudy is in line with these emerging concepts and shows theversatility of this system to also address fundamental humandevelopmental and cell biology questions.
AcknowledgmentsWe thank Dr Mammoliti and the obstetrics group at Melzo Hospitalfor CB collection, and we thank Dr Nathan Lawson for theTg(fli1:EGFP)y1 line. We also thank Drs Serena Balasso and LauraMasala for fish breeding and transgenic line screenings and Drs
Gianluca Deflorian and Federica Pezzimenti of the Zebrafish Facilitycare at Istituto FIRC di Oncologia Molecolare Institute, Milan, Italy.
Sources of FundingThis work was supported by European Union–funded grants UlcerTherapy (contract no: LSHB-CT-2005-512102) and Thercord (contractno: LSHB-CT-2005-018817) and by Fondazione Cariplo NOBELProject “Tumour Stem Cells” 2006 (to F.C.).
DisclosuresNone.
References1. Asahara T, Murohara T, Sullivan A, Silver M, van der Zee R, Li T,
Witzenbichler B, Schatteman G, Isner JM. Isolation of putative progenitorendothelial cells for angiogenesis. Science. 1997;275:964–967.
2. Takahashi T, Kalka C, Masuda H, Chen D, Silver M, Kearney M, MagnerM, Isner JM, Asahara T. Ischemia- and cytokine-induced mobilization ofbone marrow-derived endothelial progenitor cells for neovascularization.Nat Med. 1999;5:434–438.
3. Gehling UM, Ergun S, Schumacher U, Wagener C, Pantel K, Otte M,Schuch G, Schafhausen P, Mende T, Kilic N, Kluge K, Schafer B,Hossfeld DK, Fiedler W. In vitro differentiation of endothelial cells fromAC133-positive progenitor cells. Blood. 2000;95:3106–3112.
4. Prater DN, Case J, Ingram DA, Yoder MC. Working hypothesis toredefine endothelial progenitor cells. Leukemia. 2007;21:1141–1149.
5. Ingram DA, Caplice NM, Yoder MC. Unresolved questions, changingdefinitions, and novel paradigms for defining endothelial progenitor cells.Blood. 2005;106:1525–1531.
6. Ingram DA, Mead LE, Tanaka H, Meade V, Fenoglio A, Mortell K,Pollok K, Ferkowicz MJ, Gilley D, Yoder MC. Identification of a novelhierarchy of endothelial progenitor cells using human peripheral andumbilical cord blood. Blood. 2004;104:2752–2760.
7. Yoder MC, Mead LE, Prater D, Krier TR, Mroueh KN, Li F, Krasich R,Temm CJ, Prchal JT, Ingram DA. Redefining endothelial progenitor cellsvia clonal analysis and hematopoietic stem/progenitor cell principals.Blood. 2007;109:1801–1809.
8. Piaggio G, Rosti V, Corselli M, Bertolotti F, Bergamaschi G, Pozzi S,Imperiale D, Chiavarina B, Bonetti E, Novara F, Sessarego M, Villani L,Garuti A, Massa M, Ghio R, Campanelli R, Bacigalupo A, Pecci A,Viarengo G, Zuffardi O, Frassoni F, Barosi G. Endothelial colony-forming cells from patients with chronic myeloproliferative disorders lackthe disease-specific molecular clonality marker. Blood. 2009;114:3127–3130.
9. Hur J, Yoon CH, Kim HS, Choi JH, Kang HJ, Hwang KK, Oh BH, LeeMM, Park YB. Characterization of two types of endothelial progenitorcells and their different contributions to neovasculogenesis. ArteriosclerThromb Vasc Biol. 2004;24:288–293.
10. Timmermans F, Plum J, Yoder MC, Ingram DA, Vandekerckhove B,Case J. Endothelial progenitor cells: identity defined? J Cell Mol Med.2009;13:87–102.
11. Huber TL, Kouskoff V, Fehling HJ, Palis J, Keller G. Haemangioblastcommitment is initiated in the primitive streak of the mouse embryo.Nature. 2004;432:625–630.
12. Vogeli KM, Jin SW, Martin GR, Stainier DY. A common progenitor forhaematopoietic and endothelial lineages in the zebrafish gastrula. Nature.2006;443:337–339.
13. Kissa K, Herbomel P. Blood stem cells emerge from aortic endotheliumby a novel type of cell transition. Nature. 2010;464:112–115.
14. Boisset JC, van Cappellen W, Andrieu-Soler C, Galjart N, Dzierzak E,Robin C. In vivo imaging of haematopoietic cells emerging from themouse aortic endothelium. Nature. 2010;464:116–120.
15. Swiers G, Speck NA, de Bruijn MF. Visualizing blood cell emergencefrom aortic endothelium. Cell Stem Cell. 2010;6:289–290.
16. Bertrand JY, Chi NC, Santoso B, Teng S, Stainier DY, Traver D. Haema-topoietic stem cells derive directly from aortic endothelium during devel-opment. Nature. 2010;464:108–111.
17. Pelosi E, Valtieri M, Coppola S, Botta R, Gabbianelli M, Lulli V,Marziali G, Masella B, Muller R, Sgadari C, Testa U, Bonanno G,Peschle C. Identification of the hemangioblast in postnatal life. Blood.2002;100:3203–3208.
18. Ramos AL, Darabi R, Akbarloo N, Borges L, Catanese J, Dineen SP,Brekken RA, Perlingeiro RC. Clonal analysis reveals a common pro-
1596 Arterioscler Thromb Vasc Biol July 2011
by guest on March 16, 2018
http://atvb.ahajournals.org/D
ownloaded from

genitor for endothelial, myeloid, and lymphoid precursors in umbilicalcord blood. Circ Res. 2010;107:1460–1469.
19. Lawson ND, Weinstein BM. In vivo imaging of embryonic vasculardevelopment using transgenic zebrafish. Dev Biol. 2002;248:307–318.
20. Zhang XY, Rodaway AR. SCL-GFP transgenic zebrafish: in vivoimaging of blood and endothelial development and identification of theinitial site of definitive hematopoiesis. Dev Biol. 2007;307:179–194.
21. Chen MJ, Yokomizo T, Zeigler BM, Dzierzak E, Speck NA. Runx1 isrequired for the endothelial to haematopoietic cell transition but notthereafter. Nature. 2009;457:887–891.
22. North TE, Goessling W, Peeters M, Li P, Ceol C, Lord AM, Weber GJ,Harris J, Cutting CC, Huang P, Dzierzak E, Zon LI. Hematopoietic stemcell development is dependent on blood flow. Cell. 2009;137:736–748.
23. Lancrin C, Sroczynska P, Serrano AG, Gandillet A, Ferreras C, KouskoffV, Lacaud G. Blood cell generation from the hemangioblast. J Mol Med.2010;88:167–172.
24. Romagnani P, Annunziato F, Liotta F, Lazzeri E, Mazzinghi B, Frosali F,Cosmi L, Maggi L, Lasagni L, Scheffold A, Kruger M, Dimmeler S,Marra F, Gensini G, Maggi E, Romagnani S. CD14�CD34low cells withstem cell phenotypic and functional features are the major source ofcirculating endothelial progenitors. Circ Res. 2005;97:314–322.
25. Ober EA, Olofsson B, Makinen T, Jin SW, Shoji W, Koh GY, Alitalo K,Stainier DY. Vegfc is required for vascular development and endodermmorphogenesis in zebrafish. EMBO Rep. 2004;5:78–84.
26. Pesce M, Orlandi A, Iachininoto MG, Straino S, Torella AR, Rizzuti V,Pompilio G, Bonanno G, Scambia G, Capogrossi MC. Myoendothelialdifferentiation of human umbilical cord blood-derived stem cells in is-chemic limb tissues. Circ Res. 2003;93:e51–e62.
27. Gaipa G, Tilenni M, Straino S, Burba I, Zaccagnini G, Belotti D, Biagi E,Valentini M, Perseghin P, Parma M, Di Campli C, Biondi A, CapogrossiMC, Pompilio G, Pesce M. GMP-based CD133(�) cells isolationmaintains progenitor angiogenic properties and enhances standardizationin cardiovascular cell therapy. J Cell Mol Med. 2010;14:1619–1634.
28. Kocher AA, Schuster MD, Szabolcs MJ, Takuma S, Burkhoff D, WangJ, Homma S, Edwards NM, Itescu S. Neovascularization of ischemicmyocardium by human bone-marrow-derived angioblasts prevents car-diomyocyte apoptosis, reduces remodeling and improves cardiacfunction. Nat Med. 2001;7:430–436.
29. Botta R, Gao E, Stassi G, Bonci D, Pelosi E, Zwas D, Patti M, ColonnaL, Baiocchi M, Coppola S, Ma X, Condorelli G, Peschle C. Heart infarctin NOD-SCID mice: therapeutic vasculogenesis by transplantation ofhuman CD34� cells and low dose CD34�KDR� cells. FASEB J. 2004;18:1392–1394.
30. Kawamoto A, Iwasaki H, Kusano K, Murayama T, Oyamada A, Silver M,Hulbert C, Gavin M, Hanley A, Ma H, Kearney M, Zak V, Asahara T,Losordo DW. CD34-positive cells exhibit increased potency and safetyfor therapeutic neovascularization after myocardial infarction comparedwith total mononuclear cells. Circulation. 2006;114:2163–2169.
31. Ott I, Keller U, Knoedler M, Gotze KS, Doss K, Fischer P, Urlbauer K,Debus G, von Bubnoff N, Rudelius M, Schomig A, Peschel C, Oost-endorp RA. Endothelial-like cells expanded from CD34� blood cellsimprove left ventricular function after experimental myocardialinfarction. FASEB J. 2005;19:992–994.
32. Schatteman GC, Hanlon HD, Jiao C, Dodds SG, Christy BA. Blood-derived angioblasts accelerate blood-flow restoration in diabetic mice.J Clin Invest. 2000;106:571–578.
33. Madeddu P, Emanueli C, Pelosi E, Salis MB, Cerio AM, Bonanno G,Patti M, Stassi G, Condorelli G, Peschle C. Transplantation of low doseCD34�KDR� cells promotes vascular and muscular regeneration inischemic limbs. FASEB J. 2004;18:1737–1739.
34. Bayliss PE, Bellavance KL, Whitehead GG, Abrams JM, Aegerter S,Robbins HS, Cowan DB, Keating MT, O’Reilly T, Wood JM, RobertsTM, Chan J. Chemical modulation of receptor signaling inhibits regen-erative angiogenesis in adult zebrafish. Nat Chem Biol. 2006;2:265–273.
35. Huang CC, Lawson ND, Weinstein BM, Johnson SL. reg6 is required forbranching morphogenesis during blood vessel regeneration in zebrafishcaudal fins. Dev Biol. 2003;264:263–274.
36. Iovine MK. Conserved mechanisms regulate outgrowth in zebrafish fins.Nat Chem Biol. 2007;3:613–618.
37. Medvinsky A, Dzierzak E. Definitive hematopoiesis is autonomouslyinitiated by the AGM region. Cell. 1996;86:897–906.
38. Case J, Mead LE, Bessler WK, Prater D, White HA, Saadatzadeh MR,Bhavsar JR, Yoder MC, Haneline LS, Ingram DA. HumanCD34�AC133�VEGFR-2� cells are not endothelial progenitor cellsbut distinct, primitive hematopoietic progenitors. Exp Hematol. 2007;35:1109–1118.
39. Asahara T, Masuda H, Takahashi T, Kalka C, Pastore C, Silver M,Kearne M, Magner M, Isner JM. Bone marrow origin of endothelialprogenitor cells responsible for postnatal vasculogenesis in physiologicaland pathological neovascularization. Circ Res. 1999;85:221–228.
40. Murohara T, Ikeda H, Duan J, Shintani S, Sasaki K, Eguchi H, OnitsukaI, Matsui K, Imaizumi T. Transplanted cord blood-derived endothelialprecursor cells augment postnatal neovascularization. J Clin Invest. 2000;105:1527–1536.
41. De Falco E, Porcelli D, Torella AR, Straino S, Iachininoto MG, OrlandiA, Truffa S, Biglioli P, Napolitano M, Capogrossi MC, Pesce M. SDF-1involvement in endothelial phenotype and ischemia-induced recruitmentof bone marrow progenitor cells. Blood. 2004;104:3472–3482.
42. Melero-Martin JM, Khan ZA, Picard A, Wu X, Paruchuri S, Bischoff J. Invivo vasculogenic potential of human blood-derived endothelial pro-genitor cells. Blood. 2007;109:4761–4768.
43. Zon LI. Developmental biology of hematopoiesis. Blood. 1995;86:2876–2891.
44. Kalka C, Masuda H, Takahashi T, Kalka-Moll WM, Silver M, KearneyM, Li T, Isner JM, Asahara T. Transplantation of ex vivo expandedendothelial progenitor cells for therapeutic neovascularization. Proc NatlAcad Sci U S A. 2000;97:3422–3427.
45. Mazo IB, Gutierrez-Ramos JC, Frenette PS, Hynes RO, Wagner DD, vonAndrian UH. Hematopoietic progenitor cell rolling in bone marrowmicrovessels: parallel contributions by endothelial selectins and vascularcell adhesion molecule 1. J Exp Med. 1998;188:465–474.
46. Nicoli S, Ribatti D, Cotelli F, Presta M. Mammalian tumor xenograftsinduce neovascularization in zebrafish embryos. Cancer Res. 2007;67:2927–2931.
47. Gnecchi M, Zhang Z, Ni A, Dzau VJ. Paracrine mechanisms in adult stemcell signaling and therapy. Circ Res. 2008;103:1204–1219.
48. Diez-Torre A, Andrade R, Eguizabal C, Lopez E, Arluzea J, Silio M,Arechaga J. Reprogramming of melanoma cells by embryonic microen-vironments. Int J Dev Biol. 2009;53:1563–1568.
49. Dovey MC, Zon LI. Defining cancer stem cells by xenotransplantation inzebrafish. Methods Mol Biol. 2009;568:1–5.
Pozzoli et al Human CD34� Cells in Zebrafish 1597
by guest on March 16, 2018
http://atvb.ahajournals.org/D
ownloaded from

Capogrossi and Maurizio PesceLacovich, Carla Lora Lamia, Umberto Fascio, Daniela Longoni, Franco Cotelli, Maurizio C.
Ombretta Pozzoli, Pietro Vella, Grazia Iaffaldano, Valeria Parente, Paolo Devanna, MartaZebrafish
Progenitor Cells in+Endothelial Fate and Angiogenic Properties of Human CD34
Print ISSN: 1079-5642. Online ISSN: 1524-4636 Copyright © 2011 American Heart Association, Inc. All rights reserved.
Greenville Avenue, Dallas, TX 75231is published by the American Heart Association, 7272Arteriosclerosis, Thrombosis, and Vascular Biology
doi: 10.1161/ATVBAHA.111.2269692011;31:1589-1597; originally published online April 28, 2011;Arterioscler Thromb Vasc Biol.
http://atvb.ahajournals.org/content/31/7/1589World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://atvb.ahajournals.org/content/suppl/2013/10/17/ATVBAHA.111.226969.DC2 http://atvb.ahajournals.org/content/suppl/2011/04/28/ATVBAHA.111.226969.DC1
Data Supplement (unedited) at:
http://atvb.ahajournals.org//subscriptions/
at: is onlineArteriosclerosis, Thrombosis, and Vascular Biology Information about subscribing to Subscriptions:
http://www.lww.com/reprints
Information about reprints can be found online at: Reprints:
document. Question and AnswerPermissions and Rightspage under Services. Further information about this process is available in the
which permission is being requested is located, click Request Permissions in the middle column of the WebCopyright Clearance Center, not the Editorial Office. Once the online version of the published article for
can be obtained via RightsLink, a service of theArteriosclerosis, Thrombosis, and Vascular Biologyin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on March 16, 2018
http://atvb.ahajournals.org/D
ownloaded from

O. Pozzoli et al. ATVB/2011/226969 R1 Human CD34+ endothelial fate in zebrafish
1
SUPPLEMENT MATERIAL
Expanded ethical statement
An Institutional Review Board formal approval for cord blood collection at Melzo Hospital was
obtained to this aim (December 12, 2008; authorization no 843). Collection of peripheral blood was
performed at Centro Cardiologico Monzino on a voluntary basis from healthy subjects. The
experimentation involving the use of animals conforms with the Guide for the Care and Use of
Laboratory Animals published by the US National Institutes of Health (NIH Publication No. 85-23,
revised 1996) and the Helsinki Declaration. Specific authorization to proceed with animal
experimentation in the present study was obtained by Italian Ministry of Health (authorization no
08/2009) from the zebrafish Facility at IEO/IFOM campus.
Cells isolation and culture; flow cytometry
CD34+ cells were isolated using a magnetic isolation method (MINI-MACS, Miltenyi Biotech)
already described by us 1. After diluting 1:3 freshly collected cord blood with PBS, whole blood
was layered over Lymphoprep (1,077g/mL density) followed by centrifugation at 800xg for 30min
at room temperature. Cord blood mononuclear cells (CBMNCs) were then collected, washed twice
with PBS containing 2mM EDTA and 5% FBS and finally processed for CD34+ cells isolation and
culture. MACS-isolated hCD34+ cells were maintained in culture for one day before injection into a
serum-free medium (Stem Span, Stem Cell Technologies) supplemented with IL-3/6 (20 ng/ml),
SCF and Flt3-L (both at 100 ng/ml).
For the isolation of CD14- cells, peripheral blood samples from healthy volunteers were collected.
After dilution 1:3 with PBS, peripheral blood was loaded onto Lymphoprep for the peripheral blood
mononuclear cell (PBMNCs) separation. CD14- cells were isolated by negative selection using the
CD14 Isolation MINI-MACS Kit (Miltenyi Biotech). After isolation and before culture, CD14- cells

O. Pozzoli et al. ATVB/2011/226969 R1 Human CD34+ endothelial fate in zebrafish
2
were maintained in DMEM supplemented with 10% FCS for one day before injection. The
immunophenotype of CD34+ and CD14- cells was assessed by flow cytometry as follows.
Immunophenotype of CD34+ cells at T0
Immunophenotype of CD34+ cells prior injection was assessed by incubating cells with 1-10µg
specific Pacific Blue-CD3, FITC-CD34, APC-Cy7-CD45, Alexa-700-CD48, APC-KDR, APC-
CD144, APC-CD14 and APC-CD31 antibodies. All antibodies were purchased from BD-
Pharmingen except for CD48 and KDR that were purchased from Ex-Bio and R&D Systems,
respectively. Mouse IgG conjugated with the same fluorochromes were used as isotype controls.
Cells (1-5x105) were incubated at RT for 15’ after, which they were washed and immediately
analyzed by FACSAria high throughput cell sorter system (Beckton-Dickinson). 5x104 events were
scored for each sample.
Immunophenotype of CD34+-derived cells at 28 and 48 hpf
To assess the phenotype of human cells after blastula injection, Tg(fli1:EGFP)y1 embryos injected
with Orange+hCD34+ cells were dissociated at 28 and 48 hpf by using a dissociation solution
containing 0.1 mg/ml Collagenase IV and 1mM EDTA in Ca2+-free Ringer’s Solution, until
reaching a homogeneous unicellular suspension. Cells were then filtered through a 70µm mesh
filter and then labeled with the indicated antibodies under conditions as described above. The
following gating strategy was adopted to recognize human-derived cells. In a first dot plot, the
Orange tracker fluorescence was plotted against the EGFP fluorescence, to clearly discriminate
human cells from embryos’ endothelial cells (Figure 3). The Orange+ cells were recognized by
comparing the plots obtained from cells uninjected and cells-injected embryos (Figure 3). These
cells were finally gated to analyze the expression of hematopoietic and endothelial markers in
multicolor analysis. Given the small percentage (0.2%-0.5%) of human cells compared to cells of
the host embryos, this analysis was performed by analyzing a minimum of 105 total events.

O. Pozzoli et al. ATVB/2011/226969 R1 Human CD34+ endothelial fate in zebrafish
3
Purity checking of CD14- cells
Flow cytometry was used to assess negative depletion of CD14+ cells from PBMNCs before
labeling with Orange Tracker and injection into developing zebrafish embryos. To this purpose, a
PE-labeled anti-CD14 antibody (Beckton Dickinson) was used (Figure I). hCD14- cells were used
as negative control as they are similar in morphology to hCD34+ cells but, unlike CD34+ cells and
CD14+ monocytes, they do not behave as endothelial progenitor cells and exhibit no angiogenic
activity2.
Isolation of CD34+/CD45- cells
Isolation of CD34+/CD45- cells was performed by a high throughput cell sorting method using
FACSAria system. Briefly, hCD34+ cells were first isolated using MINI-MACS (see above) and
then cultured for 2 days before sorting. On the day of sorting, CD45- cells were separated from
CD45+ by flow cytometry, after staining with an anti CD45-APC antibody. After sorting, cells were
cultured for 5 additional days to let them grow in sufficient numbers to perform embryo injection.
Before injection, these cells were labeled using Orange Tracker dye as described below.
Human cells transplantation; Vegf-C morpholino RNA injection
Embryos were collected by natural spawning and staged. Tg(fli1:EGFP)y1 blastula-stage embryos or
tricaine (0.04 mg/mL, Sigma)-anesthetized 48 hpf stage embryos were injected with 4 nL PBS
containing 100-500 Orange Tracker (Invitrogen) labeled CD34+ or CD14- cells per embryo by using
a FemtoJet Express Microinjector (Eppendorf), as described by Traver et al. (2003)3. Blastula-stage
embryos were also injected with a 4 nL drop/embryo of PBS and 100-500 Orange Tracker labeled
CD34+/CD45- cells, sorted from a freshly isolated CD34+ cell population as described above.
Tg(fli1:EGFP)y1 CD34+/CD45- injected embryos were imaged by confocal microscopy in order to
visualize human cells homing and embryonic vascular phenotype.

O. Pozzoli et al. ATVB/2011/226969 R1 Human CD34+ endothelial fate in zebrafish
4
Tg(fli1:EGFP)y1 blastula-stage embryos or tricaine (0.04 mg/mL, Sigma)-anesthetized 28 hpf stage
embryos were injected with about 100 Orange Tracker labeled melanoma cancer cells (from
primary culture) and CACO-2 cells respectively, then imaged by confocal microscopy (LSM710,
Zeiss) 24h after injection. Furthermore, CACO-2 injected embryos were cultured until 7dpf and
imaged in order to visualize ectopic vessels induced by cancer cells.
Zebrafish Vegf-c antisense morpholino (MO) RNA was injected into 1-4 cell stage transgenic
Tg(fli1:EGFP)y1 embryos, as described4.
Embryo sectioning, immunofluorescence and confocal microscopy
Embryo sectioning
Zebrafish control and injected embryos at 72 hpf stage were fixed using 4% PFA for 2h at room
temperature. 8 µm transversal frozen sections of OCT-embedded 4% PFA fixed embryos, were cut,
immunostained and mounted. Images were acquired by confocal microscopy.
Immunofluorescence
Immunofluorescence on whole-mount embryos and frozen sections was performed by using
polyclonal mouse and rabbit anti-human antibodies, diluted from 1:50 to 1:1000: CD45 (Abcam),
KDR (Abcam), Endothelin (Abcam) and Ph-H3 (Bethyl Antibodies, TEMA). Goat anti-mouse and
anti-rabbit Alexa Fluor 633 and 350 (Molecular Probes, Invitrogen), diluted 1:200, were used for
the secondary staining. DAPI staining (Sigma) was performed for nuclear labeling where present.
TUNEL assay was performed according to manufacturer’s instructions (Roche Applied Science).
Confocal imaging
Live injected Tg(fli1:EGFP)y1 embryos were imaged either by a Leica stereo-microscope equipped
with a digital camera and the Leica IM100™ software, or by a Leica TCSSP2AOBS confocal
microscopy and a Leica software for time-lapse in vivo imaging, equipped with a Ar/Kr and a UV

O. Pozzoli et al. ATVB/2011/226969 R1 Human CD34+ endothelial fate in zebrafish
5
laser (GFP: 488 nm filter for excitation and 500-600 nm AOBS filter for emission; Orange Cell
Tracker dye: 594 nm filter for excitation and 600-700 nm AOBS filter for emission).
Injected and/or immunostained whole-mount embryos and embryo sections were imaged by the
LSM710 Zeiss confocal inverted microscope and the Zeiss software Zen 2008, equipped with a
Ar/Kr and a UV laser (GFP: 488 nm filter for excitation and 500-600 nm filter for emission; Orange
Cell Tracker dye: 594 nm filter for excitation and 600-700 nm filter for emission; Alexa 633: 612
nm filter for excitation and 700 nm filter for emission).
To document EHT events during vascular development of hCD34+ injected embryos, we performed
a 16h time-lapse imaging of live injected Tg(KDRI:EGFP)S843, 24h after blastula injection with a
high magnification (20X) of the dorsal aorta region (1-12 somites), by using the LSM710 Zeiss
confocal inverted microscope and the Zeiss software Zen 2008, equipped with the laser as
described above.
Whole-mount alkaline phosphatase staining and in situ hybridization methods
Alkaline phosphatase staining
After fixation and dehydratation using a methanol series, embryos were stained with a solution
containing NBT/BCIP (Roche Applied Science).
In situ hybridization
In situ hybridization to detect Scl and Gata2 genes was performed as described in 5. Riboprobes
were synthesized by using the T3/T7 MaxiScript kit (Ambion). Hybridization was performed in
50% formamide at 65°C for an overnight. After washing, color reaction was developed by BM
purple (Roche Applied Science).
TUNEL

O. Pozzoli et al. ATVB/2011/226969 R1 Human CD34+ endothelial fate in zebrafish
6
TUNEL staining was performed using the Fluorescein In Situ Cell Death Detection kit (TUNEL,
Roche Applied Science).
Fin regeneration model
Regenerative angiogenesis experiments were performed on caudal fins of adult Tg(fli1:EGFP)y1
zebrafish that were amputated at approximately 50% proximal/distal level. Amputated fish were
kept individually, deprived of food, and were immunosuppressed using 0.02 µg/µl Cyclosporine A
in fish water starting 1 day before the amputation throughout the experiment. The third day post-
amputation, adults were injected intraperitoneally with vehicle or hCD34+ cells, and kept at 25°C
for 7 additional days. Ten days post-amputation injected adults were sacrificed and whole
regenerating caudal fins were cut and fixed in 4% PFA. Images of the regenerating fin rays were
obtained by confocal microscopy, and angiogenesis was quantified by using AxioMeasure software
(Zeiss). Quantification of the blood vessels regeneration in the fin rays of amputated fins was
performed using the Axio Measure plugin in the AxioVision software (Zeiss). The areas of three fin
rays occupied by green fluorescence was measured (in pixel2) over a fixed selected area in the
LSM710 Zeiss confocal microscope field showing the amputation (see also figure 6A).

O. Pozzoli et al. ATVB/2011/226969 R1 Human CD34+ endothelial fate in zebrafish
7
SUPPLEMENTARY REFERENCES
1. Pesce M, Orlandi A, Iachininoto MG, Straino S, Torella AR, Rizzuti V, Pompilio G,
Bonanno G, Scambia G, Capogrossi MC. Myoendothelial differentiation of human
umbilical cord blood-derived stem cells in ischemic limb tissues. Circ Res. 2003;93:e51-62.
2. Urbich C, Heeschen C, Aicher A, Dernbach E, Zeiher AM, Dimmeler S. Relevance of
monocytic features for neovascularization capacity of circulating endothelial progenitor
cells. Circulation. 2003;108:2511-2516. Epub 2003 Oct 2527.
3. Traver D, Paw BH, Poss KD, Penberthy WT, Lin S, Zon LI. Transplantation and in vivo
imaging of multilineage engraftment in zebrafish bloodless mutants. Nat Immunol.
2003;4:1238-1246.
4. Ober EA, Olofsson B, Makinen T, Jin SW, Shoji W, Koh GY, Alitalo K, Stainier DY.
Vegfc is required for vascular development and endoderm morphogenesis in zebrafish.
EMBO Rep. 2004;5:78-84.
5. Thisse C, Thisse B, Postlethwait JH. Expression of snail2, a second member of the zebrafish
snail family, in cephalic mesendoderm and presumptive neural crest of wild-type and
spadetail mutant embryos. Dev Biol. 1995;172:86-99.

O. Pozzoli et al. ATVB/2011/226969 R1 Human CD34+ endothelial fate in zebrafish
8
SUPPLEMENTARY FIGURES LEGENDS
Figure I. Purity checking of negatively selected human peripheral blood-derived CD14- cells.
After negative selection by MINI-MACS, CD14- and CD14+ cells were cultured for one day. They
then were labelled with anti CD14-PE antibody and analyzed by flow cytometry. Panel on the top
shows that the majority of CD14+ cells (blue gate in physical dot plot) were retained in the MINI-
MACS column, while CD14- cells (red gate in physical dot plots in panels on the bottom) were
fairly pure and were not contaminated by cells expressing CD14.
Figure II. Proliferation and cell death of hCD34+ labeled cells injected into the zebrafish
blastula. Tg(fli1:EGFP)y1 and wild-type (WT) embryos were injected with Orange Cell-Tracker
labeled hCD34+ cells, 24 hours after isolation from human cord blood. Transplanted embryos were
cultured and PFA-fixed at 28 hpf stage of development. Whole-mount embryos are oriented with
anterior to the right and dorsal to the top.
A) Whole-mount Ph-H3 immunofluorescence detected a low hCD34+ cells proliferation rate
(1.25±1.01% of injected cells). Pictures in the head and the trunk regions were taken at low (left
panels) and high (right panels) magnification. Dashed boxes in the left panels indicate magnified
regions in right panels. Green fluorescence represents the embryonic vessels in the Tg(fli1:EGFP)y1
embryos; h: heart; e: eye; cvp: caudal vein plexus; da: dorsal artery; iv: intersegmental vessels.
Arrows indicate Ph-H3+ human cells.
B) TUNEL staining revealed only occasional hCD34+ cells apoptosis (0.54±0.60% of injected
cells). Pictures in the yolk sac and trunk regions were taken at low (left panels) and high (right
panels) magnification. Dashed boxes in the left panels indicate magnified regions in right panels.
Arrows indicate TUNEL+ human cells in the magnified regions.

O. Pozzoli et al. ATVB/2011/226969 R1 Human CD34+ endothelial fate in zebrafish
9
Figure III. Localization of blastulae-injected CD34+/CD45- cells in the developing embryos at
28 hpf. High throughput sorted CD34+/CD45- cells were injected at blastula stage. At this stage,
Orange Tracker+ cells were observed in several vascular districs such as the head, the dorsal aorta
(da), the dorsal lateral vein (dlv) and the caudal vein plexus (cvp) in proximity of intersegmental
vessels (iv).
Figure IV. Injection of human cancer stem cells (CSCs) from melanoma primary culture and
CACO-2 cell line.
(A) Cancer stem cells were injected at the blastula stage (top panel) or in the Cuvier’s duct after
gastrulation (bottom panel). Note the presence in both cases of red-labelled CSCs in the yolk and
the caudal vein plexus regions, where these cells formed micro-metastases.
(B). Angiogenic sprouting in the same embryo depicted in bottom panel in A at 7 days post
fertilization (7 dpf). The CSCs were not present at this stage likely due to rejection by the host
immune system. However, the presence of CACO-2 cells in the yolk region caused an evident
ectopic vessel sprouting (arrows), as evidenced by the presence of an expanded vascular network
localized ventrally to the yolk in this embryo (right panels). Left panels indicate the vascularisation
of the yolk in an uninjected embryo at a comparable developmental stage.
Figure V. Immunophenotype of hCD34+ cells before injection into developing blastulae. Cells
were analyzed for the expression of the indicated hematopoietic or endothelial markers by flow
cytometry one day after isolation. The dot plots at the top of each panel indicate the physical
parameters of cells that were gated to analyze the expression of hematopoietic and endothelial cells
markers. The histogram plots show the marker positive and negative for each marker. Gate was
established by staining with isotype control antibodies.

O. Pozzoli et al. ATVB/2011/226969 R1 Human CD34+ endothelial fate in zebrafish
10
Figure VI. Flow cytometry analysis of CD4 and CD8 markers expression in hCD34+ cells
before (T0 CD34+) and after injection into developing blastulae. None of these markers was
expressed at all time points. Logical gating strategy applied to recognize human cells in suspensions
obtained from enzymatically dissociating the embryos is the same as that used in Figure 3.
Figure VII. EHT imaging in a Tg(KDRI:EGFP)S843 transgenic embryo transplanted with
hCD34+ cells at blastula stage. The four panels are four subsequent time frames of the same 16 hrs
time lapse experiment, started at 24hpf (see also Movie 1). The dashed square indicates a region of
the dorsal aorta where budding of a KDR+ HSCs (*) appears to emerge from the hemogenic
endothelium (arrow).
Figure VIII. Homing of hCD34+ cells in the vasculature of post-gastrulation stages zebrafish
embryos. Tg(fli1:EGFP)y1 and wild-type (WT) embryos received hCD34+ cells into the Cuvier duct
at 48 hpf stage. Transplanted embryos were cultured until 72 hpf stage.
A) Two-hour confocal microscopy tracking of circulating red-labeled hCD34+ cells, immediately
after transplantation into a Tg(fli1:EGFP)y1 embryo. The four images represent the 12, 24, 48 and
110 minute time frames, respectively. hCD34+ cells produced early and late homing events as
shown by the presence of red cells associated to the vasculature as early as 12 minutes after
injection (dashed squares) and at later time points (dashed circles). For further details about
circulation, rolling and homing of hCD34+ cells in the vasculature at post gastrulation stages see
also Movie 1 in the supplementary material. Embryos are oriented with the anterior on the top left
and posterior to the lower right. Da: dorsal aorta/dorsal artery; iv: intersegmental vessels.
B) Injection of unlabeled hCD34+ cells into the zebrafish embryo peripheral circulation caused
vascular abnormalities in the tail vasculature. These defects were never observed in vehicle-injected
embryos. Upper panels show the tail region of an embryo injected with vehicle only (low
magnification in the left and high magnification in the right upper panels). The lower panel shows

O. Pozzoli et al. ATVB/2011/226969 R1 Human CD34+ endothelial fate in zebrafish
11
the tail region of a hCD34+ cells injected embryo; ectopic vessels sprouting is apparent between the
dorsal artery and the insersegmental vessels. Embryos are oriented with the anterior to the right and
posterior to the left. cvp: caudal vein plexus; da: dorsal artery; dlv: dorsal longitudinal vessels; iv:
intersegmental vessels; pcv: posterior cardinal vein.
C) Stereoscope imaging of whole-mount alkaline phosphatase staining at the trunk-yolk region in
one vehicle-injected post-gastrulation WT embryo (upper panel) and one injected with unlabeled
hCD34+ cells (lower panel). Note the abnormal development of the subintestinal vein (siv) in the
embryo transplanted with human cells, compared to that injected with vehicle only. Embryos are
oriented with the anterior to the right and posterior to the left.
D) Confocal microscope imaging of the tail region in a post-gastrulation embryo injected with
vehicle only (upper panel) or orange tracker labeled CD14- cells (lower panel). hCD14- cells did not
preferentially home into the vasculature and were never associated to ectopic vessels sprouting.
Embryos are oriented with the anterior to the right. iv: intersomitic vessels; da: dorsal aorta.
Figure IX. Imaging of ectopic vessel sprouting in embryos injected at post-gastrulation stages.
High magnification fields from (2-12h) confocal in vivo videomicroscopy (see also Movie 2 in
supplementary material) of tail region in a Tg(fli1:EGFP)y1 embryo, injected with hCD34+ cells at
48 hpf, showing an altered secondary blood vessel associated to human cells localization. Arrows
indicate a hCD34+ cells cluster located in the forming dorsal longitudinal vessel (dlv); dashed
squares indicate a single hCD34+ cell in an intersegmental vessel (iv). Note the presence of an
abnormally developed branching between the dorsal lateral vein (dlv) and two intersegmental
vessels (iv). The embryo is oriented with anterior to the right and dorsal to the top.
Movie 1. Time lapse experiment in a hCD34+ cells blastula-injected Tg(KDRI:EGFP)S843
started at 24hpf. A budding of a KDR+ HSCs appears to emerge from the hemogenic endothelium
located in the wall of the dorsal aorta.

O. Pozzoli et al. ATVB/2011/226969 R1 Human CD34+ endothelial fate in zebrafish
12
Movie 2. Short time (0-2 hrs) confocal videomicroscopy recording showing in real time circulation,
rolling and homing events of red-labeled hCD34+ cells into the embryonic vasculature, recognized
by green fluorescence. Representative time frames from this video are also shown in Figure 5A.
Movie 3. Long time (2-12 hrs) confocal videomicroscopy recording showing abnormal growth of a
branch between dorsal lateral vein and intersegmental vessels in proximity with a hCD34+ cells
cluster. Representative time frames from this video are also shown in Figure V.

O. Pozzoli et al. ATVB/2011/226969 R1 Human CD34+ endothelial fate in zebrafish
13
FIGURE I

O. Pozzoli et al. ATVB/2011/226969 R1 Human CD34+ endothelial fate in zebrafish
14
FIGURE II

O. Pozzoli et al. ATVB/2011/226969 R1 Human CD34+ endothelial fate in zebrafish
15
FIGURE III

O. Pozzoli et al. ATVB/2011/226969 R1 Human CD34+ endothelial fate in zebrafish
16
FIGURE IV

O. Pozzoli et al. ATVB/2011/226969 R1 Human CD34+ endothelial fate in zebrafish
17
FIGURE V

O. Pozzoli et al. ATVB/2011/226969 R1 Human CD34+ endothelial fate in zebrafish
18
FIGURE VI

O. Pozzoli et al. ATVB/2011/226969 R1 Human CD34+ endothelial fate in zebrafish
19
FIGURE VII

O. Pozzoli et al. ATVB/2011/226969 R1 Human CD34+ endothelial fate in zebrafish
20
FIGURE VIII

O. Pozzoli et al. ATVB/2011/226969 R1 Human CD34+ endothelial fate in zebrafish
21
FIGURE IX

76
Summary
그 동안의 내피전구세포 등을 비롯한 심혈관계
전구세포에 대한 집중적인 연구에도 불구하고
인간 유래 조혈전구세포(hematopo ie t i c
progenitor)가 태생기후 혈관조성(postnatal
vascularization)에 어떠한 역할을 갖는지는 아직
도 불분명한 상태이다. 더구나, 허혈 손상이 없는
태생기의 정상 심혈관계 발생에 조혈전구세포가
신생혈관조성(neovascularization)에 참여하는지
그리고 태아기에 발생하는 혈관망에 조혈전구세
포가 직접적으로 포함되는지 아니면 측분비 효
과(paracrine action)를 나타내는지도 불분명하
다. 이번 연구에서는 인간 제대혈 유래 CD34+
(hCD34+) 세포를 zebraf ish의 장배형성기
(gastrulation) 전과 후, 그리고, 성체 혈관재생의
형태인 꼬리지느러미 절단(caudal fin amputation)
모델에 주입하여 hCD34+ 세포가 갖는 혈관신생
효과를 확인하였다.
장배형성기 전에 주입된 hCD34+세포는 Scl,
Gata2로 대변되는 zebrafish hemangioblast
와 함께 동행하여 전, 후측 외중배엽(lateral
mesoderm)에서 배아 심혈관계 발생에 직접적
으로 관계하고 이는 CD45로 표지되는 lineage
reprogramming과 무관하게 작동된다. 또한, 장
배형성기 이후에 주입된 hCD34+ 세포는 형성중
인 배아 심혈관계에 넓게 분포되어 심혈관계 발생
에 강력한 측분비 효과를 보이며 이러한 효과를
통하여 꼬리지느러미 절단 모델에서도 혈관 재생
효과를 갖는 것을 밝혔다. 이 결과들은 인간 유래
조혈전구세포가 심혈관계 발생에 상당한 역할을
하며 내피전구세포로의 분화에 분자경로의 진화
적 보존을 뒷받침하는 단서가 된다.
Zebrafish의 장배형성기 전, 후에 주입된 인간 제대 유래 CD34+ 세포는 다양한 방법으로 배아 심혈관계 발생에 기여한다.
배 장 환 교수충북대학교병원 순환기내과

77
Commentary
골수, 말초혈액, 그리고 제대혈에 존재하는 일
부의 세포가 내피세포로 분화되며, 이는 HSC
(hematopoietic stem cell)와 발생학적 연관이 있
음이 밝혀져 있다. 그리고, CD34+와 CD133+를 보
이는 세포들이 체내, 외에서 성숙한 내피세포 표
지자를 갖는 세포로 분화되어 이러한 세포들을
EPC (endothelial progenitor cell)로 부르게 되었
다. 하지만, 최근의 결과에서 EPC가 조혈전구세포
유래와 그렇지 않은 세포로 나뉠 수 있다는 것이
밝혀졌고, 이는 전, 후기 EPC로 불리게 되었으며,
또한, CFU-EC (colony-forming unit-endothelial
cell)와 ECFC (endothelial colony-forming cell)
로 불려지기도 한다. 이들은 기본적으로 조혈세
포 표지자인 CD45의 발현여부와 내피세포로의
분화력에 의하여 분류된다. 조혈세포계와 내피세
포계로의 분리는 생쥐와 zebrafish의 발생에서
는 배측 대동맥 내피(dorsal aorta endothelium)
에 위치한 원시세포가 비대칭 분리를 보이는 EHT
(endothelial-hematopoietic transition) 동안
에 나타나게 된다. 더구나, 조혈줄기세포에서 유
래된 세포와 내피전구세포가 같은 혈관조성능력
을 배아기와 출생 후에 갖는지 그리고 제대유래
CD34+/CD38+/CD45+/CD133+ 전구세포에서 얻어
진 내피세포가 배아기에 어떠한 역할을 하는지도
불분명한 상태이다.
이번 연구자들은 hCD34+ 세포를 zebrafish의 장
배형성기 전, 후와 성체 신생혈관조성 모델인 꼬
리지느러미 절단 모델에 주입하여 신생혈관에 미
치는 영향을 검증하였다. Zebrafish의 중기포배
(midblastula stage)에 주입된 hCD34+ 전구세포
는 미아기(tailbud stage)와 체절기(somite stage)
에 이동하여 상당수의 세포가 배아의 중앙부와
두부 간엽, 중간세포괴(intermediate cell mass)
부위에서 관찰되며, 이는 zebrafish의 혈관모세포
(hemangioblast) 표지자인 Scl+, Gata2+를 발현하
는 중배엽세포의 분포와 연관을 갖게 된다. 주입
후 24, 28hpf (hours post fertilization)에 hCD34+
세포를 보면 CD3/4/8/144는 발현이 없고,
CD38/48/105는 발현이 감소되며, 내피표지자인
CD146과 KDR (kinase insert domain receptor)
은 28 hpf에 상승하고 내피표지자인 CD31과 전
조혈(panhenatopoietic) 표지자인 CD45는 전기
간에 걸쳐 상승하는 것을 알 수 있다. 배측 대동맥
에 분포한 세포들의 내피세포에서 조혈세포로의
표지자 변화는 zebrafish 배아에서 HSC의 발생
기와 맞물려 있어 주입된 hCD34+ 세포와 배아의
HSC간에는 발생학적 연관을 가짐을 보여준다. 즉,
포배기에 주입된 hCD34+ 세포는 CD45의 발현으
로 대변되는 조혈세포로서의 프로그래밍과는 무
관하게 단핵구나 내피세포로의 다양한 분화를 하
게 되는 것이다.
또한, zebrafish의 배아의 1-4 세포기에 vegf-c
를 유전적으로 결손시키면 30% 정도에서 심혈관
계 발생의 이상이 생기지만 hCD34+ 세포를 포배
기에 주입하면 IL-8, IL-1ra, MIP-1, G-CSF, GM-
CSF, VEGF 등의 다양한 interleukin, cytokine,
chemokine을 생성하여 강력한 혈관신생 효과를
측분비 형식으로 보여 심혈관계 발생 이상을 교
정하게 된다(Figure. 1). 이러한 다양한 혈관신생
효과를 바탕으로 생후 7일령의 zebrafish 성체에
꼬리지느러미 절단 모델을 만든 이후에 hCD34+
세포를 주입하면 신생혈관망이 대조군에 비하
여 현격하게 증가하는 것을 알 수 있다. 결론적으
로, 인간 제대 유래 세포 중 조혈전구세포 표지자
인 CD34+ 세포는 내피세포로의 분화력을 보이며
이는 zebrafish의 HSC과의 연관된 직접적인 분

78
화력과 다양한 조혈인자자의 측분비능으로 배아
의 심혈관계 신생에 영향을 미치고 성체에서도 신
생혈관조성능을 보여주며 이는 조혈세포로서의
reprogramming과는 무관하게 일어난다. 이는 인
간 태아에서 발생하는 심혈관계의 발생 이상을 줄
기세포의 주입으로 교정할 수 있다는 가능성을 보
여준다.
REFERENCESAsahara T, Murohara T, Sullivan A, Silver M, van der Zee R, 1. Li T, Witzenbichler B, Schatteman G, Isner JM. Isolation of putative progenitor endothelial cells for angiogenesis. Science. 1997;275:964-967.Yoder MC, Mead LE, Prater D, Krier TR, Mroueh KN, Li 2. F, Krasich R, Temm CJ, Prchal JT, Ingram DA. Redefining endothelial progenitor cells via clonal analysis and hematopoietic stem/progenitor cell principals. Blood. 2007;109:1801-1809.Hur J, Yoon CH, Kim HS, Choi JH, Kang HJ, Hwang KK, 3. Oh BH, Lee MM, Park YB. Characterization of two types of endothelial progenitor cells and their different contributions to neovasculogenesis. Arterioscler Thromb Vasc Biol. 2004;24:288-293.Huber TL , Kouskof f V, Feh l i ng HJ, Pa l i s J, Kel le r G. 4. Haemangioblast commitment is initiated in the primitive streak of the mouse embryo. Nature. 2004;432:625-630.Murohara T, Ikeda H, Duan J, Shintani S, Sasaki K, Eguchi 5. H, Onitsuka I, Matsui K, Imaizumi T. Transplanted cord blood-derived endothelial precursor cells augment postnatal neovascularization. J Clin Invest. 2000;105:1527-1536.
Endothelial Fate and Angiogenic Properties of HumanCD34� Progenitor Cells in Zebrafish
Ombretta Pozzoli, Pietro Vella, Grazia Iaffaldano, Valeria Parente, Paolo Devanna, Marta Lacovich,Carla Lora Lamia, Umberto Fascio, Daniela Longoni, Franco Cotelli,
Maurizio C. Capogrossi, Maurizio Pesce
Objective—The vascular competence of human-derived hematopoietic progenitors for postnatal vascularization is stillpoorly characterized. It is unclear whether, in the absence of ischemia, hematopoietic progenitors participate inneovascularization and whether they play a role in new blood vessel formation by incorporating into developing vesselsor by a paracrine action.
Methods and Results—In the present study, human cord blood–derived CD34� (hCD34�) cells were transplanted into pre-and postgastrulation zebrafish embryos and in an adult vascular regeneration model induced by caudal fin amputation.When injected before gastrulation, hCD34� cells cosegregated with the presumptive zebrafish hemangioblasts,characterized by Scl and Gata2 expression, in the anterior and posterior lateral mesoderm and were involved in earlydevelopment of the embryonic vasculature. These morphogenetic events occurred without apparent lineage reprogram-ming, as shown by CD45 expression. When transplanted postgastrulation, hCD34� cells were recruited into developingvessels, where they exhibited a potent paracrine proangiogenic action. Finally, hCD34� cells rescued vascular defectsinduced by Vegf-c in vivo targeting and enhanced vascular repair in the zebrafish fin amputation model.
Conclusion—These results indicate an unexpected developmental ability of human-derived hematopoietic progenitors andsupport the hypothesis of an evolutionary conservation of molecular pathways involved in endothelial progenitordifferentiation in vivo. (Arterioscler Thromb Vasc Biol. 2011;31:1589-1597.)
Key Words: endothelium � vascular biology � angiogenesis � embryology � stem cells
It has long been supposed that cells present in the bonemarrow, peripheral blood (PB), and cord blood (CB),
which copurify with hematopoietic stem cells (HSCs), alsogive rise to endothelial cells. This speculation was supportedby the notion that CD34� and CD133� cells differentiate, inculture and in vivo, into cells that express mature endothelialcell markers. These cells, called endothelial progenitor cells(EPCs),1–3 have been the subject of numerous basic andtranslational studies showing their participation in neovascu-larization. Recent studies have revealed possible separationbetween hematopoietic- and nonhematopoietic-derivedEPCs.4–8 These 2 cell types have been called early and lateEPCs,9 or colony-forming unit–endothelial cells (CFU-ECs)and endothelial colony-forming cells (ECFCs).7 They arefundamentally distinguished on the basis of hematopoieticmarker expression (eg, CD45) and the ability to proliferate or
to differentiate into endothelial cells.10 Separation betweenhematopoietic and the endothelial lineages during earlymouse11 and zebrafish12 development is established during byasymmetrical division of primitive cells located in the dorsalaorta endothelium, through a novel differentiation eventcalled endothelial-hematopoietic transition (EHT).13–16 Moreuncertain is whether cells with a similar potency of generat-ing HSCs or endothelial cells are present at postnatal stages17;recently, however, derivation of endothelial cells fromCD34�/CD38�/CD45�/CD133� CB progenitors was dem-onstrated,18 suggesting the existence of similar progenitors atleast during fetal life.
The zebrafish (Danio rerio) embryo provides severaladvantages for in vivo study of vascular morphogenesis,based on the opportunity to observe in real time the devel-opment of embryonic blood vessels19,20 and also on the
Received on: June 21, 2010; final version accepted on: March 31, 2011.From the Laboratorio di Biologia Vascolare e Medicina Rigenerativa, Centro Cardiologico Monzino, Istituto di Ricovero e Cura a Carattere Scientifico,
Milan, Italy (O.P., P.V., G.I., V.P., P.D., M.L., M.P.); Laboratorio di Patologia Vascolare, Istituto Dermopatico dell’Immacolata, Istituto di Ricovero eCura a Carattere Scientifico, Rome, Italy (M.C.C.); Laboratorio di Biologia dello Sviluppo (C.L.L., F.C.) and Centro Interdipartimentale MicroscopiaAvanzata (U.F.), Universita di Milano, Milan, Italy; Centro Ricerche Tettamanti, Monza, Italy (D.L.).
Drs Capogrossi and Pesce contributed equally to this work.Correspondence to Maurizio Pesce, PhD, Laboratorio di Biologia Vascolare e Medicina Rigenerativa, Centro Cardiologico Monzino, Istituto di
Ricovero e Cura a Carattere Scientifico, Via Parea 4, 20138 Milan, Italy; (E-mail [email protected]) Ombretta Pozzoli, PhD, Laboratorio diBiologia Vascolare e Medicina Rigenerativa, Centro Cardiologico Monzino, Istituto di Ricovero e Cura a Carattere Scientifico, Via Parea 4, 20138 Milan,Italy; (E-mail [email protected]) or Franco Cotelli, Laboratorio di Biologia dello Sviluppo, Universita degli Studi di Milano, Via Celoria 26,20133 Milan, Italy (E-mail [email protected]).
© 2011 American Heart Association, Inc.
Arterioscler Thromb Vasc Biol is available at http://atvb.ahajournals.org DOI: 10.1161/ATVBAHA.111.226969
1589 by IMED Korea on October 10, 2011http://atvb.ahajournals.org/Downloaded from
Figure 1. hCD34+ 세포의 주입은 antisense MO RNA를 zebrafish 배아의 1-8 세포기에 주입하여 Vegf-c를 발현이 결핍되어 발생하는 심혈관계 발생 이상을 정상화한다. A. 위) GFP 발현 정상 zebrafish의 배아. 중간, 아래) Vegf-c 발현이 없어진 배아, 측배측 대동맥(lda) 사이에 분지가 없으며 체절간 혈관 사이에 불규칙적인 간극이 보인다. B. vegf 결손 배아. 좌측 위) 머리와 난황부위, 우측 위) 배측 대동맥이 관찰되지 않는다. 좌측 아래) 체부와 꼬리 부위, 좌측 아래) 체절간 혈관 사이의 간극이 불규칙적이다. C. hCD34+ 세포가 주입된 Vegf-c 결손 배아. 좌측 위) 머리와 난황부위, 우측 위) 두 개의 배측 대동맥사이에 정상적인 가지 연결이 보인다. 좌측 아래) 체부와 꼬리 부위, 좌측 아래) 체절 간 혈관에 간극이 정상적으로 규칙적이다.
and KDR markers without counterselection for CD45 shouldgive rise only to CFU-ECs38 and support neovascularizationby paracrine interactions with resident endothelial cells. Thislatter hypothesis, however, is contradicted by several exper-imental findings showing that blood-borne stem cells giverise to endothelial cells in vitro and in ischemic tissuesirrespective of their hematopoietic origin2,3,26,29,33,39–42 and bya very recent report showing that CD34�/CD38�/CD45�/CD133� CB progenitors differentiate in culture into clonallyexpanding endothelial cells, which, similarly to ECFCs,participate in neovascularization by incorporating into newblood vessels.18
Transplantation of hCD34� Cells at Early Stagesof Embryonic Zebrafish Development Unravels anUnexpected Hemangioblast-Like BehaviorTransplantation into midblastulas was performed to reveal thefate of hCD34� cells throughout the earliest stages ofvascular development. These experiments showed thatblastula-transplanted hCD34�cells were mostly cosegregatedwith the presumptive zebrafish Scl� and Gata2� progenitorsin the anterior and posterior lateral mesoderm. Thus, hCD34�
cells migrated in concert with zebrafish mesoderm progeni-tors and colonized the anterior region, ie, the site whereembryonic uncommitted progenitors are known to establishprimitive hematopoiesis.43 During somitogenesis, hCD34�
cells migrated medially and posteriorly along the developingintermediate cell mass, to finally colonize the entire cardio-vascular system at the 28 hpf stage. As shown by flowcytometry and immunofluorescence (Figures 3 and 4), thesemorphogenetic events were associated with transient upregu-lation of KDR and overexpression of CD146 at all stages;importantly, this also coincided with downregulation ofhematopoietic progenitor–specific markers CD38 and CD48.It was also remarkable that hCD34� cells showed increasingamounts of monocyte marker CD14 expression; that CD45was not downregulated in KDR� human cells (Figures 3 and4); and that injection of hCD34�/CD45� cells, the putativeECFC progenitors,10 produced the same segregation patterninto various vascular districts (Supplemental Figure III).Taken together, these results suggest that blastula-injectedhCD34� cells were committed to multiple differentiation into
Figure 5. Rescue of angiogenic defects in Vegf-c-depletedembryos by blastula transplantation of hCD34� cells. Shown isin vivo knockdown of Vegf-c by injection of antisense MO RNAat 1 to 8 cell stage into Tg(fli1:enhanced green fluorescent pro-tein [EGFP])y1 embryos. A, Low-power views of a control (Fli-GFP) and 2 Vegf-c MO RNA–injected embryos (Fli-GFP�VEGF-C). Note the absence of the bifurcation between the 2 lateraldorsal aorta branches in midpanel (*) and the irregular spac-ing of the intersomitic vessels (arrows), caused by absence of
Vegf-c expression. B, Defective vascular phenotype caused byVegf-c in vivo knockdown. Panels on the left show low-powerviews of the head/yolk regions (top) and the trunk/tail regions(bottom). Dashed box in the top left panel (magnified in the topright) shows the absence of the dorsal aorta (da). aa: aorticarches; cd: Cuvier duct. Dashed box in the bottom left panel(magnified in the bottom right) shows the irregular spacing ofthe intersomitic vessels (iv) of a Fli-EGFP �VEGF-C embryo.*Regions of the trunk where defects in the organization of thesevessels and the dorsal lateral vein (dlv) were observed. da indi-cates dorsal artery; cvp, caudal vein plexus. C, Injection of red-labeled hCD34� cells into Vegf-c knockdown embryos rescued thevascular phenotype. Low-power (left) and high-power (right) viewsof regions similar to those represented in B show the presence ofhCD34� cells associated with normal formation of the 2 lateralbranches of the dorsal aorta (*, lda and lda�) and to regular spacingand branching of the intersomitic vessels (iv) in the trunk/tailregions. Note the presence of several red-labeled cells in theCuvier duct (cd), and in the caudal vein plexus (cvp). dlv indicatesdorsal lateral vein; cv, cardinal vein; da, dorsal artery.
1594 Arterioscler Thromb Vasc Biol July 2011
by IMED Korea on October 10, 2011http://atvb.ahajournals.org/Downloaded from
and KDR markers without counterselection for CD45 shouldgive rise only to CFU-ECs38 and support neovascularizationby paracrine interactions with resident endothelial cells. Thislatter hypothesis, however, is contradicted by several exper-imental findings showing that blood-borne stem cells giverise to endothelial cells in vitro and in ischemic tissuesirrespective of their hematopoietic origin2,3,26,29,33,39–42 and bya very recent report showing that CD34�/CD38�/CD45�/CD133� CB progenitors differentiate in culture into clonallyexpanding endothelial cells, which, similarly to ECFCs,participate in neovascularization by incorporating into newblood vessels.18
Transplantation of hCD34� Cells at Early Stagesof Embryonic Zebrafish Development Unravels anUnexpected Hemangioblast-Like BehaviorTransplantation into midblastulas was performed to reveal thefate of hCD34� cells throughout the earliest stages ofvascular development. These experiments showed thatblastula-transplanted hCD34�cells were mostly cosegregatedwith the presumptive zebrafish Scl� and Gata2� progenitorsin the anterior and posterior lateral mesoderm. Thus, hCD34�
cells migrated in concert with zebrafish mesoderm progeni-tors and colonized the anterior region, ie, the site whereembryonic uncommitted progenitors are known to establishprimitive hematopoiesis.43 During somitogenesis, hCD34�
cells migrated medially and posteriorly along the developingintermediate cell mass, to finally colonize the entire cardio-vascular system at the 28 hpf stage. As shown by flowcytometry and immunofluorescence (Figures 3 and 4), thesemorphogenetic events were associated with transient upregu-lation of KDR and overexpression of CD146 at all stages;importantly, this also coincided with downregulation ofhematopoietic progenitor–specific markers CD38 and CD48.It was also remarkable that hCD34� cells showed increasingamounts of monocyte marker CD14 expression; that CD45was not downregulated in KDR� human cells (Figures 3 and4); and that injection of hCD34�/CD45� cells, the putativeECFC progenitors,10 produced the same segregation patterninto various vascular districts (Supplemental Figure III).Taken together, these results suggest that blastula-injectedhCD34� cells were committed to multiple differentiation into
Figure 5. Rescue of angiogenic defects in Vegf-c-depletedembryos by blastula transplantation of hCD34� cells. Shown isin vivo knockdown of Vegf-c by injection of antisense MO RNAat 1 to 8 cell stage into Tg(fli1:enhanced green fluorescent pro-tein [EGFP])y1 embryos. A, Low-power views of a control (Fli-GFP) and 2 Vegf-c MO RNA–injected embryos (Fli-GFP�VEGF-C). Note the absence of the bifurcation between the 2 lateraldorsal aorta branches in midpanel (*) and the irregular spac-ing of the intersomitic vessels (arrows), caused by absence of
Vegf-c expression. B, Defective vascular phenotype caused byVegf-c in vivo knockdown. Panels on the left show low-powerviews of the head/yolk regions (top) and the trunk/tail regions(bottom). Dashed box in the top left panel (magnified in the topright) shows the absence of the dorsal aorta (da). aa: aorticarches; cd: Cuvier duct. Dashed box in the bottom left panel(magnified in the bottom right) shows the irregular spacing ofthe intersomitic vessels (iv) of a Fli-EGFP �VEGF-C embryo.*Regions of the trunk where defects in the organization of thesevessels and the dorsal lateral vein (dlv) were observed. da indi-cates dorsal artery; cvp, caudal vein plexus. C, Injection of red-labeled hCD34� cells into Vegf-c knockdown embryos rescued thevascular phenotype. Low-power (left) and high-power (right) viewsof regions similar to those represented in B show the presence ofhCD34� cells associated with normal formation of the 2 lateralbranches of the dorsal aorta (*, lda and lda�) and to regular spacingand branching of the intersomitic vessels (iv) in the trunk/tailregions. Note the presence of several red-labeled cells in theCuvier duct (cd), and in the caudal vein plexus (cvp). dlv indicatesdorsal lateral vein; cv, cardinal vein; da, dorsal artery.
1594 Arterioscler Thromb Vasc Biol July 2011
by IMED Korea on October 10, 2011http://atvb.ahajournals.org/Downloaded from
and KDR markers without counterselection for CD45 shouldgive rise only to CFU-ECs38 and support neovascularizationby paracrine interactions with resident endothelial cells. Thislatter hypothesis, however, is contradicted by several exper-imental findings showing that blood-borne stem cells giverise to endothelial cells in vitro and in ischemic tissuesirrespective of their hematopoietic origin2,3,26,29,33,39–42 and bya very recent report showing that CD34�/CD38�/CD45�/CD133� CB progenitors differentiate in culture into clonallyexpanding endothelial cells, which, similarly to ECFCs,participate in neovascularization by incorporating into newblood vessels.18
Transplantation of hCD34� Cells at Early Stagesof Embryonic Zebrafish Development Unravels anUnexpected Hemangioblast-Like BehaviorTransplantation into midblastulas was performed to reveal thefate of hCD34� cells throughout the earliest stages ofvascular development. These experiments showed thatblastula-transplanted hCD34�cells were mostly cosegregatedwith the presumptive zebrafish Scl� and Gata2� progenitorsin the anterior and posterior lateral mesoderm. Thus, hCD34�
cells migrated in concert with zebrafish mesoderm progeni-tors and colonized the anterior region, ie, the site whereembryonic uncommitted progenitors are known to establishprimitive hematopoiesis.43 During somitogenesis, hCD34�
cells migrated medially and posteriorly along the developingintermediate cell mass, to finally colonize the entire cardio-vascular system at the 28 hpf stage. As shown by flowcytometry and immunofluorescence (Figures 3 and 4), thesemorphogenetic events were associated with transient upregu-lation of KDR and overexpression of CD146 at all stages;importantly, this also coincided with downregulation ofhematopoietic progenitor–specific markers CD38 and CD48.It was also remarkable that hCD34� cells showed increasingamounts of monocyte marker CD14 expression; that CD45was not downregulated in KDR� human cells (Figures 3 and4); and that injection of hCD34�/CD45� cells, the putativeECFC progenitors,10 produced the same segregation patterninto various vascular districts (Supplemental Figure III).Taken together, these results suggest that blastula-injectedhCD34� cells were committed to multiple differentiation into
Figure 5. Rescue of angiogenic defects in Vegf-c-depletedembryos by blastula transplantation of hCD34� cells. Shown isin vivo knockdown of Vegf-c by injection of antisense MO RNAat 1 to 8 cell stage into Tg(fli1:enhanced green fluorescent pro-tein [EGFP])y1 embryos. A, Low-power views of a control (Fli-GFP) and 2 Vegf-c MO RNA–injected embryos (Fli-GFP�VEGF-C). Note the absence of the bifurcation between the 2 lateraldorsal aorta branches in midpanel (*) and the irregular spac-ing of the intersomitic vessels (arrows), caused by absence of
Vegf-c expression. B, Defective vascular phenotype caused byVegf-c in vivo knockdown. Panels on the left show low-powerviews of the head/yolk regions (top) and the trunk/tail regions(bottom). Dashed box in the top left panel (magnified in the topright) shows the absence of the dorsal aorta (da). aa: aorticarches; cd: Cuvier duct. Dashed box in the bottom left panel(magnified in the bottom right) shows the irregular spacing ofthe intersomitic vessels (iv) of a Fli-EGFP �VEGF-C embryo.*Regions of the trunk where defects in the organization of thesevessels and the dorsal lateral vein (dlv) were observed. da indi-cates dorsal artery; cvp, caudal vein plexus. C, Injection of red-labeled hCD34� cells into Vegf-c knockdown embryos rescued thevascular phenotype. Low-power (left) and high-power (right) viewsof regions similar to those represented in B show the presence ofhCD34� cells associated with normal formation of the 2 lateralbranches of the dorsal aorta (*, lda and lda�) and to regular spacingand branching of the intersomitic vessels (iv) in the trunk/tailregions. Note the presence of several red-labeled cells in theCuvier duct (cd), and in the caudal vein plexus (cvp). dlv indicatesdorsal lateral vein; cv, cardinal vein; da, dorsal artery.
1594 Arterioscler Thromb Vasc Biol July 2011
by IMED Korea on October 10, 2011http://atvb.ahajournals.org/Downloaded from

79
Endothelial Fate and Angiogenic Properties of HumanCD34� Progenitor Cells in Zebrafish
Ombretta Pozzoli, Pietro Vella, Grazia Iaffaldano, Valeria Parente, Paolo Devanna, Marta Lacovich,Carla Lora Lamia, Umberto Fascio, Daniela Longoni, Franco Cotelli,
Maurizio C. Capogrossi, Maurizio Pesce
Objective—The vascular competence of human-derived hematopoietic progenitors for postnatal vascularization is stillpoorly characterized. It is unclear whether, in the absence of ischemia, hematopoietic progenitors participate inneovascularization and whether they play a role in new blood vessel formation by incorporating into developing vesselsor by a paracrine action.
Methods and Results—In the present study, human cord blood–derived CD34� (hCD34�) cells were transplanted into pre-and postgastrulation zebrafish embryos and in an adult vascular regeneration model induced by caudal fin amputation.When injected before gastrulation, hCD34� cells cosegregated with the presumptive zebrafish hemangioblasts,characterized by Scl and Gata2 expression, in the anterior and posterior lateral mesoderm and were involved in earlydevelopment of the embryonic vasculature. These morphogenetic events occurred without apparent lineage reprogram-ming, as shown by CD45 expression. When transplanted postgastrulation, hCD34� cells were recruited into developingvessels, where they exhibited a potent paracrine proangiogenic action. Finally, hCD34� cells rescued vascular defectsinduced by Vegf-c in vivo targeting and enhanced vascular repair in the zebrafish fin amputation model.
Conclusion—These results indicate an unexpected developmental ability of human-derived hematopoietic progenitors andsupport the hypothesis of an evolutionary conservation of molecular pathways involved in endothelial progenitordifferentiation in vivo. (Arterioscler Thromb Vasc Biol. 2011;31:1589-1597.)
Key Words: endothelium � vascular biology � angiogenesis � embryology � stem cells
It has long been supposed that cells present in the bonemarrow, peripheral blood (PB), and cord blood (CB),
which copurify with hematopoietic stem cells (HSCs), alsogive rise to endothelial cells. This speculation was supportedby the notion that CD34� and CD133� cells differentiate, inculture and in vivo, into cells that express mature endothelialcell markers. These cells, called endothelial progenitor cells(EPCs),1–3 have been the subject of numerous basic andtranslational studies showing their participation in neovascu-larization. Recent studies have revealed possible separationbetween hematopoietic- and nonhematopoietic-derivedEPCs.4–8 These 2 cell types have been called early and lateEPCs,9 or colony-forming unit–endothelial cells (CFU-ECs)and endothelial colony-forming cells (ECFCs).7 They arefundamentally distinguished on the basis of hematopoieticmarker expression (eg, CD45) and the ability to proliferate or
to differentiate into endothelial cells.10 Separation betweenhematopoietic and the endothelial lineages during earlymouse11 and zebrafish12 development is established during byasymmetrical division of primitive cells located in the dorsalaorta endothelium, through a novel differentiation eventcalled endothelial-hematopoietic transition (EHT).13–16 Moreuncertain is whether cells with a similar potency of generat-ing HSCs or endothelial cells are present at postnatal stages17;recently, however, derivation of endothelial cells fromCD34�/CD38�/CD45�/CD133� CB progenitors was dem-onstrated,18 suggesting the existence of similar progenitors atleast during fetal life.
The zebrafish (Danio rerio) embryo provides severaladvantages for in vivo study of vascular morphogenesis,based on the opportunity to observe in real time the devel-opment of embryonic blood vessels19,20 and also on the
Received on: June 21, 2010; final version accepted on: March 31, 2011.From the Laboratorio di Biologia Vascolare e Medicina Rigenerativa, Centro Cardiologico Monzino, Istituto di Ricovero e Cura a Carattere Scientifico,
Milan, Italy (O.P., P.V., G.I., V.P., P.D., M.L., M.P.); Laboratorio di Patologia Vascolare, Istituto Dermopatico dell’Immacolata, Istituto di Ricovero eCura a Carattere Scientifico, Rome, Italy (M.C.C.); Laboratorio di Biologia dello Sviluppo (C.L.L., F.C.) and Centro Interdipartimentale MicroscopiaAvanzata (U.F.), Universita di Milano, Milan, Italy; Centro Ricerche Tettamanti, Monza, Italy (D.L.).
Drs Capogrossi and Pesce contributed equally to this work.Correspondence to Maurizio Pesce, PhD, Laboratorio di Biologia Vascolare e Medicina Rigenerativa, Centro Cardiologico Monzino, Istituto di
Ricovero e Cura a Carattere Scientifico, Via Parea 4, 20138 Milan, Italy; (E-mail [email protected]) Ombretta Pozzoli, PhD, Laboratorio diBiologia Vascolare e Medicina Rigenerativa, Centro Cardiologico Monzino, Istituto di Ricovero e Cura a Carattere Scientifico, Via Parea 4, 20138 Milan,Italy; (E-mail [email protected]) or Franco Cotelli, Laboratorio di Biologia dello Sviluppo, Universita degli Studi di Milano, Via Celoria 26,20133 Milan, Italy (E-mail [email protected]).
© 2011 American Heart Association, Inc.
Arterioscler Thromb Vasc Biol is available at http://atvb.ahajournals.org DOI: 10.1161/ATVBAHA.111.226969
1589 by IMED Korea on October 10, 2011http://atvb.ahajournals.org/Downloaded from
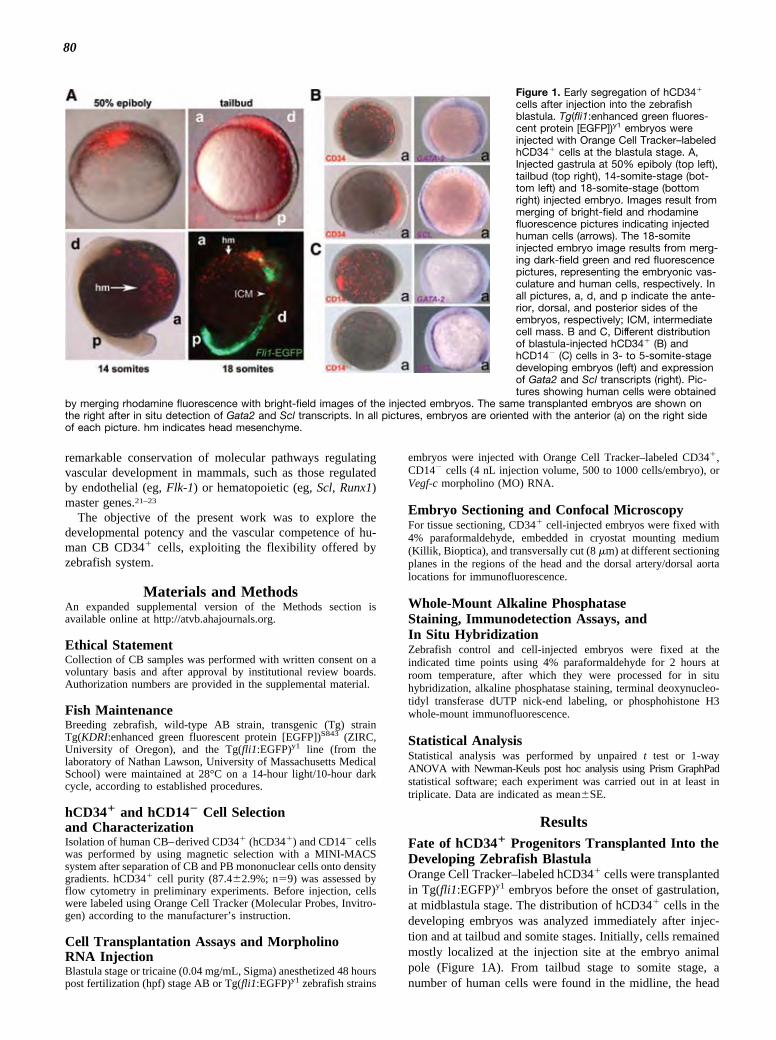
80
mesenchyme, and the intermediate cell mass regions (Figure1A). In situ hybridization using riboprobes to detect Scl andGata2 mRNAs were then performed. The results revealed astrict association of human cells with multipotent Scl� andGata2� mesoderm cells (Figure 1B), suggesting cosegrega-tion of human cells with zebrafish hemangioblasts. Thisbehavior was specific; in fact, PB cells devoid of the CD14�
monocyte cell population, having proangiogenic activity,24
were not associated with Scl� and Gata2� regions (Figure 1Cand Supplemental Figure I).
Proliferation and survival of hCD34� cells transplantedinto Tg(fli1:EGFP)y1 blastulas were evaluated at the 28 hpfstage by whole-mount staining for phosphohistone H3 andterminal deoxynucleotidyl transferase dUTP nick-end label-ing. Both tests revealed little proliferation and only occa-sional apoptosis (Supplemental Figure II). At this stage,human cells were found in primary and secondary bloodvessels of the head, in the heart, in the dorsal aorta, in theposterior cardinal vein, in the caudal vein plexus, and inintersegmental and longitudinal vessels (Figure 2A to 2C). Atthe blastula stage, injection of CB-derived CD34�/CD45�
cells, considered to be containing putative ECFC progeni-tors,10 produced analogous results (Supplemental Figure III),whereas injection of melanoma cancer cells gave rise toformation of micrometastases in the head and the caudal veinplexus regions (Supplemental Figure IVA). Finally, observa-tion of transversal sections revealed the presence of hCD34�
cells in the lumen, as well as in the wall of the dorsal aortaand in other vessels in strict association with EGFP� hostendothelial cells (Figure 2D).
To further investigate the phenotype of human cells,blastula-injected embryos were dissociated in single-cellsuspensions at 28 and 48 hpf, followed by staining withantibodies recognizing human hematopoietic and endothelialmarkers and flow cytometry (Figure 3). Human cells wereeasily recognized as a red-labeled population in dot plotsrepresenting red versus green fluorescence. In these cells(Figure 3 and Supplemental Figures V and VI), CD3, CD4,CD8, and CD144 (VE-cadherin) were not expressed before orafter injection; CD38, CD48, and CD105 exhibited a signif-icant downregulation; endothelial marker CD146 was in-creased; and kinase insert domain receptor (KDR) wastransiently upregulated at 28 hpf. Interestingly, the monocytemarker CD14 was expressed in a very small subset ofhCD34� cells before injection, to be progressively upregu-lated at the 28 and 48 hpf stages; finally, endothelial markerCD31 and panhenatopoietic marker CD45 were expressed athigh levels at all stages. To reveal coexpression of endothelialand hematopoietic markers in CD34� cells in the embryos,immunofluorescence in whole-mount embryos and on trans-versal frozen sections was performed. This revealed that vessel-associated human cells expressed the hemangioblast/endothelialmarker KDR (Figure 4A) and the endothelial marker endo-thelin (Figure 4B). Strikingly, irrespective of their locationin developing embryonic vessels, human-derived cellsexpressed the hematopoietic lineage marker CD45 (Figure4C). Coexpression of CD45 with KDR was, finally,detected in vessel-associated human cells (Figure 4D).
Transition from endothelial to hematopoietic phenotype ofcells localized in the wall of the dorsal aorta has beendescribed as the major event associated with emergence ofdefinitive HSCs in the zebrafish embryo.13,16 To assesswhether EHT is affected by the presence of human cells, a16-hour time-lapse video recording of Tg(KDRI:EGFP)S843
embryos injected at the blastula stage with CD34� cells was
Figure 2. Fate of blastula-injected hCD34� cells at postgastru-lation stages. Tg(fli1:enhanced green fluorescent protein[EGFP])y1 embryos were injected at the blastula stage withOrange Cell Tracker–labeled hCD34� cells and fixed with para-formaldehyde at the 28 hours post fertilization (hpf) stage ofdevelopment. A, Confocal microscope low-power view of ahCD34� injected embryo. hCD34� cells are represented by reddots. Dashed boxes represent magnified regions in B and C. Band C, Magnification of the tail and head regions, respectively,of the embryo shown in A. Note in B the preferential associationof hCD34� cells to the caudal vein plexus (cvp; red arrows) andthe dorsal artery (da; green arrows) and in C the presence ofhCD34� cells (red arrows) lining the head vessels, 1 of the 2branches the dorsal aorta (lda; green arrows) and the Cuvierduct (cd; yellow arrows). dlv indicates dorsal lateral vein; iv,intersomitic vessels. D, Low-power view of an uninjectedTg(fli1:EGFP)y1 embryo at the 28 hpf stage (top left) and trans-versal section at the midtrunk level of 2 hCD34� injectedTg(fli1:EGFP)y1 embryos at different magnification (bottom rightand left). In the top left panel, the dashed line indicates theapproximate sectioning plane represented in the 2 other panels;dashed box in the right panel indicates the region of the dorsalaorta (da) shown at a higher magnification in the bottom rightpanel, representing human cells (arrows) in close associationwith EGFP� zebrafish endothelial cells of the dorsal aorta. ntindicates neural tube.
Pozzoli et al Human CD34� Cells in Zebrafish 1591
by IMED Korea on October 10, 2011http://atvb.ahajournals.org/Downloaded from
remarkable conservation of molecular pathways regulatingvascular development in mammals, such as those regulatedby endothelial (eg, Flk-1) or hematopoietic (eg, Scl, Runx1)master genes.21–23
The objective of the present work was to explore thedevelopmental potency and the vascular competence of hu-man CB CD34� cells, exploiting the flexibility offered byzebrafish system.
Materials and MethodsAn expanded supplemental version of the Methods section isavailable online at http://atvb.ahajournals.org.
Ethical StatementCollection of CB samples was performed with written consent on avoluntary basis and after approval by institutional review boards.Authorization numbers are provided in the supplemental material.
Fish MaintenanceBreeding zebrafish, wild-type AB strain, transgenic (Tg) strainTg(KDRI:enhanced green fluorescent protein [EGFP])S843 (ZIRC,University of Oregon), and the Tg(fli1:EGFP)y1 line (from thelaboratory of Nathan Lawson, University of Massachusetts MedicalSchool) were maintained at 28°C on a 14-hour light/10-hour darkcycle, according to established procedures.
hCD34� and hCD14� Cell Selectionand CharacterizationIsolation of human CB–derived CD34� (hCD34�) and CD14� cellswas performed by using magnetic selection with a MINI-MACSsystem after separation of CB and PB mononuclear cells onto densitygradients. hCD34� cell purity (87.4�2.9%; n�9) was assessed byflow cytometry in preliminary experiments. Before injection, cellswere labeled using Orange Cell Tracker (Molecular Probes, Invitro-gen) according to the manufacturer’s instruction.
Cell Transplantation Assays and MorpholinoRNA InjectionBlastula stage or tricaine (0.04 mg/mL, Sigma) anesthetized 48 hourspost fertilization (hpf) stage AB or Tg(fli1:EGFP)y1 zebrafish strains
embryos were injected with Orange Cell Tracker–labeled CD34�,CD14� cells (4 nL injection volume, 500 to 1000 cells/embryo), orVegf-c morpholino (MO) RNA.
Embryo Sectioning and Confocal MicroscopyFor tissue sectioning, CD34� cell-injected embryos were fixed with4% paraformaldehyde, embedded in cryostat mounting medium(Killik, Bioptica), and transversally cut (8 �m) at different sectioningplanes in the regions of the head and the dorsal artery/dorsal aortalocations for immunofluorescence.
Whole-Mount Alkaline PhosphataseStaining, Immunodetection Assays, andIn Situ HybridizationZebrafish control and cell-injected embryos were fixed at theindicated time points using 4% paraformaldehyde for 2 hours atroom temperature, after which they were processed for in situhybridization, alkaline phosphatase staining, terminal deoxynucleo-tidyl transferase dUTP nick-end labeling, or phosphohistone H3whole-mount immunofluorescence.
Statistical AnalysisStatistical analysis was performed by unpaired t test or 1-wayANOVA with Newman-Keuls post hoc analysis using Prism GraphPadstatistical software; each experiment was carried out in at least intriplicate. Data are indicated as mean�SE.
ResultsFate of hCD34� Progenitors Transplanted Into theDeveloping Zebrafish BlastulaOrange Cell Tracker–labeled hCD34� cells were transplantedin Tg(fli1:EGFP)y1 embryos before the onset of gastrulation,at midblastula stage. The distribution of hCD34� cells in thedeveloping embryos was analyzed immediately after injec-tion and at tailbud and somite stages. Initially, cells remainedmostly localized at the injection site at the embryo animalpole (Figure 1A). From tailbud stage to somite stage, anumber of human cells were found in the midline, the head
Figure 1. Early segregation of hCD34�
cells after injection into the zebrafishblastula. Tg(fli1:enhanced green fluores-cent protein [EGFP])y1 embryos wereinjected with Orange Cell Tracker–labeledhCD34� cells at the blastula stage. A,Injected gastrula at 50% epiboly (top left),tailbud (top right), 14-somite-stage (bot-tom left) and 18-somite-stage (bottomright) injected embryo. Images result frommerging of bright-field and rhodaminefluorescence pictures indicating injectedhuman cells (arrows). The 18-somiteinjected embryo image results from merg-ing dark-field green and red fluorescencepictures, representing the embryonic vas-culature and human cells, respectively. Inall pictures, a, d, and p indicate the ante-rior, dorsal, and posterior sides of theembryos, respectively; ICM, intermediatecell mass. B and C, Different distributionof blastula-injected hCD34� (B) andhCD14� (C) cells in 3- to 5-somite-stagedeveloping embryos (left) and expressionof Gata2 and Scl transcripts (right). Pic-tures showing human cells were obtained
by merging rhodamine fluorescence with bright-field images of the injected embryos. The same transplanted embryos are shown onthe right after in situ detection of Gata2 and Scl transcripts. In all pictures, embryos are oriented with the anterior (a) on the right sideof each picture. hm indicates head mesenchyme.
1590 Arterioscler Thromb Vasc Biol July 2011
by IMED Korea on October 10, 2011http://atvb.ahajournals.org/Downloaded from

81
mesenchyme, and the intermediate cell mass regions (Figure1A). In situ hybridization using riboprobes to detect Scl andGata2 mRNAs were then performed. The results revealed astrict association of human cells with multipotent Scl� andGata2� mesoderm cells (Figure 1B), suggesting cosegrega-tion of human cells with zebrafish hemangioblasts. Thisbehavior was specific; in fact, PB cells devoid of the CD14�
monocyte cell population, having proangiogenic activity,24
were not associated with Scl� and Gata2� regions (Figure 1Cand Supplemental Figure I).
Proliferation and survival of hCD34� cells transplantedinto Tg(fli1:EGFP)y1 blastulas were evaluated at the 28 hpfstage by whole-mount staining for phosphohistone H3 andterminal deoxynucleotidyl transferase dUTP nick-end label-ing. Both tests revealed little proliferation and only occa-sional apoptosis (Supplemental Figure II). At this stage,human cells were found in primary and secondary bloodvessels of the head, in the heart, in the dorsal aorta, in theposterior cardinal vein, in the caudal vein plexus, and inintersegmental and longitudinal vessels (Figure 2A to 2C). Atthe blastula stage, injection of CB-derived CD34�/CD45�
cells, considered to be containing putative ECFC progeni-tors,10 produced analogous results (Supplemental Figure III),whereas injection of melanoma cancer cells gave rise toformation of micrometastases in the head and the caudal veinplexus regions (Supplemental Figure IVA). Finally, observa-tion of transversal sections revealed the presence of hCD34�
cells in the lumen, as well as in the wall of the dorsal aortaand in other vessels in strict association with EGFP� hostendothelial cells (Figure 2D).
To further investigate the phenotype of human cells,blastula-injected embryos were dissociated in single-cellsuspensions at 28 and 48 hpf, followed by staining withantibodies recognizing human hematopoietic and endothelialmarkers and flow cytometry (Figure 3). Human cells wereeasily recognized as a red-labeled population in dot plotsrepresenting red versus green fluorescence. In these cells(Figure 3 and Supplemental Figures V and VI), CD3, CD4,CD8, and CD144 (VE-cadherin) were not expressed before orafter injection; CD38, CD48, and CD105 exhibited a signif-icant downregulation; endothelial marker CD146 was in-creased; and kinase insert domain receptor (KDR) wastransiently upregulated at 28 hpf. Interestingly, the monocytemarker CD14 was expressed in a very small subset ofhCD34� cells before injection, to be progressively upregu-lated at the 28 and 48 hpf stages; finally, endothelial markerCD31 and panhenatopoietic marker CD45 were expressed athigh levels at all stages. To reveal coexpression of endothelialand hematopoietic markers in CD34� cells in the embryos,immunofluorescence in whole-mount embryos and on trans-versal frozen sections was performed. This revealed that vessel-associated human cells expressed the hemangioblast/endothelialmarker KDR (Figure 4A) and the endothelial marker endo-thelin (Figure 4B). Strikingly, irrespective of their locationin developing embryonic vessels, human-derived cellsexpressed the hematopoietic lineage marker CD45 (Figure4C). Coexpression of CD45 with KDR was, finally,detected in vessel-associated human cells (Figure 4D).
Transition from endothelial to hematopoietic phenotype ofcells localized in the wall of the dorsal aorta has beendescribed as the major event associated with emergence ofdefinitive HSCs in the zebrafish embryo.13,16 To assesswhether EHT is affected by the presence of human cells, a16-hour time-lapse video recording of Tg(KDRI:EGFP)S843
embryos injected at the blastula stage with CD34� cells was
Figure 2. Fate of blastula-injected hCD34� cells at postgastru-lation stages. Tg(fli1:enhanced green fluorescent protein[EGFP])y1 embryos were injected at the blastula stage withOrange Cell Tracker–labeled hCD34� cells and fixed with para-formaldehyde at the 28 hours post fertilization (hpf) stage ofdevelopment. A, Confocal microscope low-power view of ahCD34� injected embryo. hCD34� cells are represented by reddots. Dashed boxes represent magnified regions in B and C. Band C, Magnification of the tail and head regions, respectively,of the embryo shown in A. Note in B the preferential associationof hCD34� cells to the caudal vein plexus (cvp; red arrows) andthe dorsal artery (da; green arrows) and in C the presence ofhCD34� cells (red arrows) lining the head vessels, 1 of the 2branches the dorsal aorta (lda; green arrows) and the Cuvierduct (cd; yellow arrows). dlv indicates dorsal lateral vein; iv,intersomitic vessels. D, Low-power view of an uninjectedTg(fli1:EGFP)y1 embryo at the 28 hpf stage (top left) and trans-versal section at the midtrunk level of 2 hCD34� injectedTg(fli1:EGFP)y1 embryos at different magnification (bottom rightand left). In the top left panel, the dashed line indicates theapproximate sectioning plane represented in the 2 other panels;dashed box in the right panel indicates the region of the dorsalaorta (da) shown at a higher magnification in the bottom rightpanel, representing human cells (arrows) in close associationwith EGFP� zebrafish endothelial cells of the dorsal aorta. ntindicates neural tube.
Pozzoli et al Human CD34� Cells in Zebrafish 1591
by IMED Korea on October 10, 2011http://atvb.ahajournals.org/Downloaded from
remarkable conservation of molecular pathways regulatingvascular development in mammals, such as those regulatedby endothelial (eg, Flk-1) or hematopoietic (eg, Scl, Runx1)master genes.21–23
The objective of the present work was to explore thedevelopmental potency and the vascular competence of hu-man CB CD34� cells, exploiting the flexibility offered byzebrafish system.
Materials and MethodsAn expanded supplemental version of the Methods section isavailable online at http://atvb.ahajournals.org.
Ethical StatementCollection of CB samples was performed with written consent on avoluntary basis and after approval by institutional review boards.Authorization numbers are provided in the supplemental material.
Fish MaintenanceBreeding zebrafish, wild-type AB strain, transgenic (Tg) strainTg(KDRI:enhanced green fluorescent protein [EGFP])S843 (ZIRC,University of Oregon), and the Tg(fli1:EGFP)y1 line (from thelaboratory of Nathan Lawson, University of Massachusetts MedicalSchool) were maintained at 28°C on a 14-hour light/10-hour darkcycle, according to established procedures.
hCD34� and hCD14� Cell Selectionand CharacterizationIsolation of human CB–derived CD34� (hCD34�) and CD14� cellswas performed by using magnetic selection with a MINI-MACSsystem after separation of CB and PB mononuclear cells onto densitygradients. hCD34� cell purity (87.4�2.9%; n�9) was assessed byflow cytometry in preliminary experiments. Before injection, cellswere labeled using Orange Cell Tracker (Molecular Probes, Invitro-gen) according to the manufacturer’s instruction.
Cell Transplantation Assays and MorpholinoRNA InjectionBlastula stage or tricaine (0.04 mg/mL, Sigma) anesthetized 48 hourspost fertilization (hpf) stage AB or Tg(fli1:EGFP)y1 zebrafish strains
embryos were injected with Orange Cell Tracker–labeled CD34�,CD14� cells (4 nL injection volume, 500 to 1000 cells/embryo), orVegf-c morpholino (MO) RNA.
Embryo Sectioning and Confocal MicroscopyFor tissue sectioning, CD34� cell-injected embryos were fixed with4% paraformaldehyde, embedded in cryostat mounting medium(Killik, Bioptica), and transversally cut (8 �m) at different sectioningplanes in the regions of the head and the dorsal artery/dorsal aortalocations for immunofluorescence.
Whole-Mount Alkaline PhosphataseStaining, Immunodetection Assays, andIn Situ HybridizationZebrafish control and cell-injected embryos were fixed at theindicated time points using 4% paraformaldehyde for 2 hours atroom temperature, after which they were processed for in situhybridization, alkaline phosphatase staining, terminal deoxynucleo-tidyl transferase dUTP nick-end labeling, or phosphohistone H3whole-mount immunofluorescence.
Statistical AnalysisStatistical analysis was performed by unpaired t test or 1-wayANOVA with Newman-Keuls post hoc analysis using Prism GraphPadstatistical software; each experiment was carried out in at least intriplicate. Data are indicated as mean�SE.
ResultsFate of hCD34� Progenitors Transplanted Into theDeveloping Zebrafish BlastulaOrange Cell Tracker–labeled hCD34� cells were transplantedin Tg(fli1:EGFP)y1 embryos before the onset of gastrulation,at midblastula stage. The distribution of hCD34� cells in thedeveloping embryos was analyzed immediately after injec-tion and at tailbud and somite stages. Initially, cells remainedmostly localized at the injection site at the embryo animalpole (Figure 1A). From tailbud stage to somite stage, anumber of human cells were found in the midline, the head
Figure 1. Early segregation of hCD34�
cells after injection into the zebrafishblastula. Tg(fli1:enhanced green fluores-cent protein [EGFP])y1 embryos wereinjected with Orange Cell Tracker–labeledhCD34� cells at the blastula stage. A,Injected gastrula at 50% epiboly (top left),tailbud (top right), 14-somite-stage (bot-tom left) and 18-somite-stage (bottomright) injected embryo. Images result frommerging of bright-field and rhodaminefluorescence pictures indicating injectedhuman cells (arrows). The 18-somiteinjected embryo image results from merg-ing dark-field green and red fluorescencepictures, representing the embryonic vas-culature and human cells, respectively. Inall pictures, a, d, and p indicate the ante-rior, dorsal, and posterior sides of theembryos, respectively; ICM, intermediatecell mass. B and C, Different distributionof blastula-injected hCD34� (B) andhCD14� (C) cells in 3- to 5-somite-stagedeveloping embryos (left) and expressionof Gata2 and Scl transcripts (right). Pic-tures showing human cells were obtained
by merging rhodamine fluorescence with bright-field images of the injected embryos. The same transplanted embryos are shown onthe right after in situ detection of Gata2 and Scl transcripts. In all pictures, embryos are oriented with the anterior (a) on the right sideof each picture. hm indicates head mesenchyme.
1590 Arterioscler Thromb Vasc Biol July 2011
by IMED Korea on October 10, 2011http://atvb.ahajournals.org/Downloaded from

82
hCD34� Cells Accelerate Vascular Repair inAdult ZebrafishThe ability of hCD34� cells to enhance vascular repair inmammalian models of myocardial infarction and limb ische-mia has been shown by us26,27 and by others.28–33 In thepresent study, we examined whether hCD34� cells contrib-uted to vascular repair in adult zebrafish by injecting thesecells in immunosuppressed Tg(fli1:EGFP)y1 adult animalsafter caudal fin amputation (Figure 6A), an established modelof vascular regeneration in zebrafish.34–36 A significant in-crease in angiogenesis in hCD34� cell–transplanted versusvehicle-injected animals was found (Figure 6B).
DiscussionBone marrow, PB, and CB contain EPCs that contribute toneovascularization in ischemic tissues.1 EPCs have long beenthought to derive from alternative differentiation pathways ofhematopoietic progenitors because of the expression of mark-ers (CD34, CD133, KDR) that they have in common withprimitive HSCs. Embryology has suggested a different sce-
nario. In fact, during vertebrate embryogenesis, HSCs derivefrom primitive cells located in a morphologically conservedregion, called the aorta-gonad-mesonephros region,37 whencedefinitive HSCs emerge by a novel modality of asymmetricaldivision of pluripotent nonhematopoietic cells, the so-calledhemogenic endothelium.13,14 This division modality has beencharacterized in the zebrafish dorsal aorta and has been calledEHT.13 In the adulthood, the existence of 2 EPCs types,namely CFU-ECs (early EPCs) and ECFCs (late EPCs),7 hasbeen proposed. These cells likely have a different origin,although they are, at least in part, functionally overlapping.The most important proofs in favor of lineage separationbetween these 2 EPC types are (1) the expression of thepanhematopietic marker CD45 in CFU-ECs but not inECFCs,4 and (2) the finding that ECFC clones from patientswith myeloproliferative disorders do not harbor HSCs spe-cific mutations, therefore identifying ECFCs as nonhemato-poietic in origin.7,8,10
According to this definition of EPCs,5 progenitors obtainedby bone marrow, PB, or CB cell sorting with CD34, CD133,
Figure 4. Marker analysis of vessel-associated hCD34�-derived cells after blastula injection by immunofluorescence on embryo sec-tions. Transversal frozen sections (8 �m) of hCD34� injected Tg(fli1:enhanced green fluorescent protein [EGFP])y1 embryos at the 28hours post fertilization (hpf) stage. A, hKDR staining in 2 embryos cut at different levels in the trunk region. nt indicates neural tube; y,yolk; da, dorsal aorta/dorsal artery. Dashed boxes in the 2 top panels indicate the regions shown at higher power view in the bottompanels, where endothelium-associated hCD34� cells (arrowheads) and a round human kinase insert domain receptor (KDR)� cells inthe lumen of the dorsal aorta/dorsal artery (*) are shown. B, Human endothelin staining of 2 transversally cut embryos at the anteriortrunk (left panel) and brain (b; right) levels. Note the presence of Orange�/endothelin� cells in the abdominal vessels (av; arrows, left)and associated with the head vessels endothelium (arrow, right panel). Orange�/endothelin� cells (*, right panel) were also found closeto head mesenchyme (hm), which also expresses Fli1 at this stage. C, hCD45 expression in human cells at different locations in 28 hpfstage embryos. Panels on the left show developing blood vessels in a transversal section of the head, where the eye vasculature ispresent. The higher magnification on the bottom shows a CD45� human cell (arrow) associated with EGFP� endothelial cells. e indi-cates eye. Panels in the center show a developing vessel located in the anterior part of the trunk. A cluster of CD45� human cells(arrows) is recognizable within EGFP� endothelial cells forming abdominal vessels (ab). Panels on the right show CD45 expression in ahuman cell (arrow) in the developing dorsal aorta (da). D, Whole-mount immunofluorescence for detection of KDR and CD45 in humancells transplanted at the blastula stage in Tg(fli1:EGFP)y1 embryos. The 2 panels on the left show different magnifications of an embryocontaining a hCD34� cells double positive for CD45 and KDR (arrows), located in the region of the dorsal aorta (da). The panel on theright depicts a CD45�/KDR� human cell in the region of the dorsal lateral vein and close to intersomitic vessels (iv).
Pozzoli et al Human CD34� Cells in Zebrafish 1593
by IMED Korea on October 10, 2011http://atvb.ahajournals.org/Downloaded from
performed. As shown in Supplemental Figure VII and Sup-plemental Movie I, budding of KDR� HSCs was not per-turbed by transplanted cells.
Postgastrula Stage Transplantation of hCD34�
Cells Modulates Embryonic Vascular DevelopmentThe behavior of hCD34� cells injected into the circulatorysystem at the 48 hpf stage Tg(fli1:EGFP)y1 embryos wasassessed by confocal time-lapse analysis. Early homingevents in the blood vessels were observed as early as 30minutes after injection (Supplemental Figure VIIA). hCD34�
cell rolling and adhesion to zebrafish intersegmental vesselswere also observed (Supplemental Movie II).
Transgenic embryos injected with hCD34� cells at the 48 hpfstage were analyzed at the 72 hpf stage, ie, 24 hours afterinjection. Altered blood vessel sprouting in the growing tailvasculature (85% of 72 transplanted embryos; SupplementalFigure VIIIB) and at the level of the subintestinal vein (80% of17 injected embryos; Supplemental Figure VIIIC) were ob-served. Angiogenesis abnormalities were never found afterinjection of PB-derived hCD14� cells (Supplemental FigureVIIID) but were detected in embryos transplanted with theintestinal cancer cell line CACO2 (Supplemental Figure IVB).To further examine whether blood vessel development wasperturbed by transplanted hCD34� cells, the vascular develop-ment of 48 hpf stage Tg(fli1:EGFP)y1 injected embryos wasmonitored from 2 to 12 hours postinjection by confocal video-microscopy. A strict association was found between red-labeledcells and the regions of ectopic vascular sprouting (Supplemen-tal Figure IX and Supplemental Movie III).
Transplanted hCD34� Cells Rescue DefectiveAngiogenesis in the Zebrafish Embryo byMultifactorial Paracrine EffectThe previous data suggested that transplanted hCD34�
cells have paracrine effects in the developing embryos. Toassess this hypothesis, cells were injected at the blastulastage in Tg(fli1:EGFP)y1 embryos, where Vegf-c wasknocked down by MO RNA injection at the 1- to 4-cellstage. As shown in Figure 5A, embryos depleted of Vegf-cexhibited defects in the development of the vascularsystem (30% of 50 MO RNA-injected embryos). Vegf-cdepletion mostly affected the formation of the dorsal aorta,the posterior cardinal vein, the intersegmental vessels, andspecific head arteries25 (Figure 5B). These defects wereobserved in only 9% (n�32) of Vegf-c-depleted/hCD34�
cell-injected embryos (Figure 5C). Interestingly, pheno-type rescue was complete even in vascular regions whereCD34� cells failed to localize, suggesting complementa-tion of Vegf-c knockdown defects by soluble factorssecreted by human cells. Finally, hCD14� cell injection inVegf-c-depleted embryos did not rescue defective bloodvessel formation (100% of vascular abnormalities, n�5embryos injected with Vegf-c MO RNA and hCD14� cells;data not shown). To identify paracrine factors participatingin the rescue effect by injected cells, the secretion ofinterleukins, chemokines, and angiogenic growth factorswas assessed by Luminex assay (Table). The resultsshowed production of several factors, including chemo-kines, proinflammatory, and proangiogenic cytokines, sug-gesting a potent paracrine effect by hCD34� cells.
Figure 3. Flow cytometry analysis of blastula-injected hCD34� cells phenotype at 28 and 48 hours post fertilization (hpf). Tg(fli1:enhancedgreen fluorescent protein [EGFP])y1 embryos were injected with hCD34� cells and then dissociated and labeled with human anti-bodies recognizing hematopoietic and endothelial cell markers for analysis by flow cytometry. A to C, Human cells were recog-nized in green vs red fluorescence dot plots by comparison with uninjected embryos. These cells were logically gated for their redfluorescence by use of 488 nm (A) and 561 nm (C) lasers and analyzed for the expression of each of the human markers. B,Quantification of results shown in A and C for hematopoietic (top) and endothelial (bottom) markers in hCD34� cells before injec-tion (open bars) and at 28 (gray bars) 48 hpf (black bars). *P�0.05 by 1-way ANOVA with Newman-Keuls post hoc analysis; n�3.Orange Tracker indicates Orange Cell Tracker.
1592 Arterioscler Thromb Vasc Biol July 2011
by IMED Korea on October 10, 2011http://atvb.ahajournals.org/Downloaded from
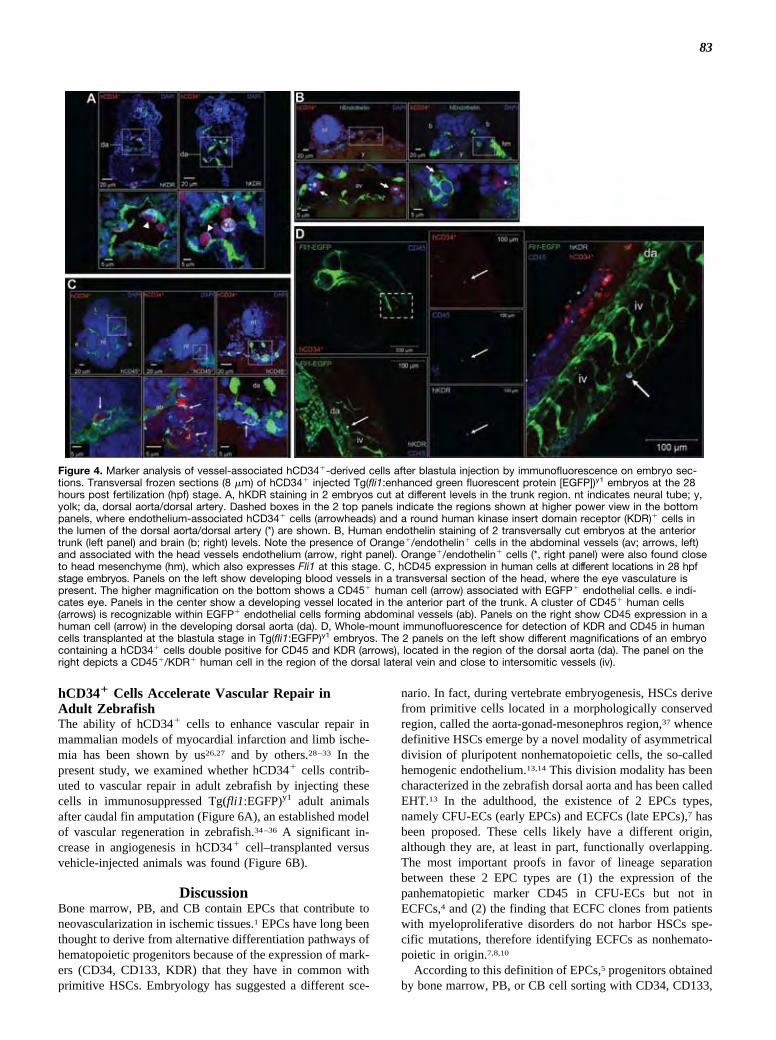
83
hCD34� Cells Accelerate Vascular Repair inAdult ZebrafishThe ability of hCD34� cells to enhance vascular repair inmammalian models of myocardial infarction and limb ische-mia has been shown by us26,27 and by others.28–33 In thepresent study, we examined whether hCD34� cells contrib-uted to vascular repair in adult zebrafish by injecting thesecells in immunosuppressed Tg(fli1:EGFP)y1 adult animalsafter caudal fin amputation (Figure 6A), an established modelof vascular regeneration in zebrafish.34–36 A significant in-crease in angiogenesis in hCD34� cell–transplanted versusvehicle-injected animals was found (Figure 6B).
DiscussionBone marrow, PB, and CB contain EPCs that contribute toneovascularization in ischemic tissues.1 EPCs have long beenthought to derive from alternative differentiation pathways ofhematopoietic progenitors because of the expression of mark-ers (CD34, CD133, KDR) that they have in common withprimitive HSCs. Embryology has suggested a different sce-
nario. In fact, during vertebrate embryogenesis, HSCs derivefrom primitive cells located in a morphologically conservedregion, called the aorta-gonad-mesonephros region,37 whencedefinitive HSCs emerge by a novel modality of asymmetricaldivision of pluripotent nonhematopoietic cells, the so-calledhemogenic endothelium.13,14 This division modality has beencharacterized in the zebrafish dorsal aorta and has been calledEHT.13 In the adulthood, the existence of 2 EPCs types,namely CFU-ECs (early EPCs) and ECFCs (late EPCs),7 hasbeen proposed. These cells likely have a different origin,although they are, at least in part, functionally overlapping.The most important proofs in favor of lineage separationbetween these 2 EPC types are (1) the expression of thepanhematopietic marker CD45 in CFU-ECs but not inECFCs,4 and (2) the finding that ECFC clones from patientswith myeloproliferative disorders do not harbor HSCs spe-cific mutations, therefore identifying ECFCs as nonhemato-poietic in origin.7,8,10
According to this definition of EPCs,5 progenitors obtainedby bone marrow, PB, or CB cell sorting with CD34, CD133,
Figure 4. Marker analysis of vessel-associated hCD34�-derived cells after blastula injection by immunofluorescence on embryo sec-tions. Transversal frozen sections (8 �m) of hCD34� injected Tg(fli1:enhanced green fluorescent protein [EGFP])y1 embryos at the 28hours post fertilization (hpf) stage. A, hKDR staining in 2 embryos cut at different levels in the trunk region. nt indicates neural tube; y,yolk; da, dorsal aorta/dorsal artery. Dashed boxes in the 2 top panels indicate the regions shown at higher power view in the bottompanels, where endothelium-associated hCD34� cells (arrowheads) and a round human kinase insert domain receptor (KDR)� cells inthe lumen of the dorsal aorta/dorsal artery (*) are shown. B, Human endothelin staining of 2 transversally cut embryos at the anteriortrunk (left panel) and brain (b; right) levels. Note the presence of Orange�/endothelin� cells in the abdominal vessels (av; arrows, left)and associated with the head vessels endothelium (arrow, right panel). Orange�/endothelin� cells (*, right panel) were also found closeto head mesenchyme (hm), which also expresses Fli1 at this stage. C, hCD45 expression in human cells at different locations in 28 hpfstage embryos. Panels on the left show developing blood vessels in a transversal section of the head, where the eye vasculature ispresent. The higher magnification on the bottom shows a CD45� human cell (arrow) associated with EGFP� endothelial cells. e indi-cates eye. Panels in the center show a developing vessel located in the anterior part of the trunk. A cluster of CD45� human cells(arrows) is recognizable within EGFP� endothelial cells forming abdominal vessels (ab). Panels on the right show CD45 expression in ahuman cell (arrow) in the developing dorsal aorta (da). D, Whole-mount immunofluorescence for detection of KDR and CD45 in humancells transplanted at the blastula stage in Tg(fli1:EGFP)y1 embryos. The 2 panels on the left show different magnifications of an embryocontaining a hCD34� cells double positive for CD45 and KDR (arrows), located in the region of the dorsal aorta (da). The panel on theright depicts a CD45�/KDR� human cell in the region of the dorsal lateral vein and close to intersomitic vessels (iv).
Pozzoli et al Human CD34� Cells in Zebrafish 1593
by IMED Korea on October 10, 2011http://atvb.ahajournals.org/Downloaded from
performed. As shown in Supplemental Figure VII and Sup-plemental Movie I, budding of KDR� HSCs was not per-turbed by transplanted cells.
Postgastrula Stage Transplantation of hCD34�
Cells Modulates Embryonic Vascular DevelopmentThe behavior of hCD34� cells injected into the circulatorysystem at the 48 hpf stage Tg(fli1:EGFP)y1 embryos wasassessed by confocal time-lapse analysis. Early homingevents in the blood vessels were observed as early as 30minutes after injection (Supplemental Figure VIIA). hCD34�
cell rolling and adhesion to zebrafish intersegmental vesselswere also observed (Supplemental Movie II).
Transgenic embryos injected with hCD34� cells at the 48 hpfstage were analyzed at the 72 hpf stage, ie, 24 hours afterinjection. Altered blood vessel sprouting in the growing tailvasculature (85% of 72 transplanted embryos; SupplementalFigure VIIIB) and at the level of the subintestinal vein (80% of17 injected embryos; Supplemental Figure VIIIC) were ob-served. Angiogenesis abnormalities were never found afterinjection of PB-derived hCD14� cells (Supplemental FigureVIIID) but were detected in embryos transplanted with theintestinal cancer cell line CACO2 (Supplemental Figure IVB).To further examine whether blood vessel development wasperturbed by transplanted hCD34� cells, the vascular develop-ment of 48 hpf stage Tg(fli1:EGFP)y1 injected embryos wasmonitored from 2 to 12 hours postinjection by confocal video-microscopy. A strict association was found between red-labeledcells and the regions of ectopic vascular sprouting (Supplemen-tal Figure IX and Supplemental Movie III).
Transplanted hCD34� Cells Rescue DefectiveAngiogenesis in the Zebrafish Embryo byMultifactorial Paracrine EffectThe previous data suggested that transplanted hCD34�
cells have paracrine effects in the developing embryos. Toassess this hypothesis, cells were injected at the blastulastage in Tg(fli1:EGFP)y1 embryos, where Vegf-c wasknocked down by MO RNA injection at the 1- to 4-cellstage. As shown in Figure 5A, embryos depleted of Vegf-cexhibited defects in the development of the vascularsystem (30% of 50 MO RNA-injected embryos). Vegf-cdepletion mostly affected the formation of the dorsal aorta,the posterior cardinal vein, the intersegmental vessels, andspecific head arteries25 (Figure 5B). These defects wereobserved in only 9% (n�32) of Vegf-c-depleted/hCD34�
cell-injected embryos (Figure 5C). Interestingly, pheno-type rescue was complete even in vascular regions whereCD34� cells failed to localize, suggesting complementa-tion of Vegf-c knockdown defects by soluble factorssecreted by human cells. Finally, hCD14� cell injection inVegf-c-depleted embryos did not rescue defective bloodvessel formation (100% of vascular abnormalities, n�5embryos injected with Vegf-c MO RNA and hCD14� cells;data not shown). To identify paracrine factors participatingin the rescue effect by injected cells, the secretion ofinterleukins, chemokines, and angiogenic growth factorswas assessed by Luminex assay (Table). The resultsshowed production of several factors, including chemo-kines, proinflammatory, and proangiogenic cytokines, sug-gesting a potent paracrine effect by hCD34� cells.
Figure 3. Flow cytometry analysis of blastula-injected hCD34� cells phenotype at 28 and 48 hours post fertilization (hpf). Tg(fli1:enhancedgreen fluorescent protein [EGFP])y1 embryos were injected with hCD34� cells and then dissociated and labeled with human anti-bodies recognizing hematopoietic and endothelial cell markers for analysis by flow cytometry. A to C, Human cells were recog-nized in green vs red fluorescence dot plots by comparison with uninjected embryos. These cells were logically gated for their redfluorescence by use of 488 nm (A) and 561 nm (C) lasers and analyzed for the expression of each of the human markers. B,Quantification of results shown in A and C for hematopoietic (top) and endothelial (bottom) markers in hCD34� cells before injec-tion (open bars) and at 28 (gray bars) 48 hpf (black bars). *P�0.05 by 1-way ANOVA with Newman-Keuls post hoc analysis; n�3.Orange Tracker indicates Orange Cell Tracker.
1592 Arterioscler Thromb Vasc Biol July 2011
by IMED Korea on October 10, 2011http://atvb.ahajournals.org/Downloaded from

84
monocytes or primary endothelial cells, irrespective of theirhematopoietic lineage programming, detected by CD45expression.
Interestingly, a number of blastula-transplanted humancells were enclosed in the endothelial layer of the dorsal aortaat 28 hpf (Figures 2D and 4). This is consistent with thetiming of EHT occurrence in the dorsal aorta of the develop-ing zebrafish embryos.13,16 To check whether the presence ofhuman cells alters the normal behavior of multipotent endo-thelial progenitors lining the dorsal aorta, we performed EHTimaging in KDR-EGFP embryos, which showed no abnor-malities in the budding process of definitive HSCs (Supple-mental Figure VII, Supplemental Movie I). The lack ofsubstantial perturbation of the EHT process is an indication ofa physiological integration of human cells into zebrafishdeveloping vascular network, even if future work will benecessary to establish whether human cells actively partici-pate in EHT in this system.
Paracrine Activity of hCD34� Cells in theZebrafish EmbryoThe angiogenic activity of human bone marrow–derived,2
PB-derived,44 or CB-derived40 progenitors has been demon-strated in several studies based on injection of these cells inpreclinical models of ischemic disease. Except for a fewreports describing results of intravital microscopy analyses ofstem cells homing to mouse vessels,45 so far only indirect
evidence of homing events in ischemic tissues, mostly basedon tissue sectioning, has been provided.26
The results of our hCD34� cell injection in postgastrulation-stage(48 hpf) embryos, showed that these cells circulated throughoutthe organism and produced early and late homing events,associated with abnormal or ectopic embryonic vasculatureformation (Supplemental Figure VIII). Interestingly, homing ofhCD34� cells was observed in several districts of the vascula-ture; this was different from the behavior of the human PB-derived CD14� cells, which were used as a negative control(Supplemental Figure VIII), or that of human melanoma and
Table. Quantification of Human Cytokines in Culture MediumConditioned by hCD34� Cells
Factor (pg/ml per 105 Cells) Mean SE (n�4)
Interleukins
Hu IL-1ra 25.5 14
Hu IL-5 13 1.8
Hu IL-9 2.9 0.8
Hu IL-8 7220 2468
Hu IL-10 3.0 1.8
Hu IL-12 1.1 0.76
Hu IL-13 11.2 3.0
Hu IL-15 0.13 0.09
Hu IL-17 1.3 0.54
Chemokines
Hu IP-10 196 103
Hu MCP-1 162 101
Hu MIP-1a 119 54
Hu MIP-1b 419 271
Hu RANTES 72.5 45
Proangiogenic factors
Hu PDGF-bb 2.0 1.3
Hu FGF basic 0.43 0.2
Hu G-CSF 14.8 5.7
Hu GM-CSF 12.3 5.9
Hu VEGF 19.1 18
Hu indicates human; IL, interleukin; PDGF, platelet-derived growth factor;FGF, fibroblast growth factor; G-CSF, granulocyte colony-stimulating factor;GM-CSF, granulocyte-macrophage colony-stimulating factor.
Figure 6. hCD34� cells accelerate vascular repair in immuno-suppressed adult zebrafish. An adult zebrafish fin amputationmodel was used to quantify vascular regeneration in regenerat-ing fin rays. A, Regenerating caudal fin of an adult fish at 7 daysafter vehicle (top) or unlabeled hCD34� cells (bottom) intraperi-toneal injection. The newly formed vascular network is moredeveloped in hCD34� cell–injected zebrafish than in controls.Dashed lines indicate the fin amputation level. B, Histogramshows the quantification of the enhanced green fluorescent pro-tein (EGFP)� area (pixel2), corresponding to the vascular net-work in confocal images of regenerating caudal fins in vehicleand hCD34� cell–injected adult fishes. fr indicates fin rays.*P�0.05 by unpaired Student t test; n�3.
Pozzoli et al Human CD34� Cells in Zebrafish 1595
by IMED Korea on October 10, 2011http://atvb.ahajournals.org/Downloaded from
and KDR markers without counterselection for CD45 shouldgive rise only to CFU-ECs38 and support neovascularizationby paracrine interactions with resident endothelial cells. Thislatter hypothesis, however, is contradicted by several exper-imental findings showing that blood-borne stem cells giverise to endothelial cells in vitro and in ischemic tissuesirrespective of their hematopoietic origin2,3,26,29,33,39–42 and bya very recent report showing that CD34�/CD38�/CD45�/CD133� CB progenitors differentiate in culture into clonallyexpanding endothelial cells, which, similarly to ECFCs,participate in neovascularization by incorporating into newblood vessels.18
Transplantation of hCD34� Cells at Early Stagesof Embryonic Zebrafish Development Unravels anUnexpected Hemangioblast-Like BehaviorTransplantation into midblastulas was performed to reveal thefate of hCD34� cells throughout the earliest stages ofvascular development. These experiments showed thatblastula-transplanted hCD34�cells were mostly cosegregatedwith the presumptive zebrafish Scl� and Gata2� progenitorsin the anterior and posterior lateral mesoderm. Thus, hCD34�
cells migrated in concert with zebrafish mesoderm progeni-tors and colonized the anterior region, ie, the site whereembryonic uncommitted progenitors are known to establishprimitive hematopoiesis.43 During somitogenesis, hCD34�
cells migrated medially and posteriorly along the developingintermediate cell mass, to finally colonize the entire cardio-vascular system at the 28 hpf stage. As shown by flowcytometry and immunofluorescence (Figures 3 and 4), thesemorphogenetic events were associated with transient upregu-lation of KDR and overexpression of CD146 at all stages;importantly, this also coincided with downregulation ofhematopoietic progenitor–specific markers CD38 and CD48.It was also remarkable that hCD34� cells showed increasingamounts of monocyte marker CD14 expression; that CD45was not downregulated in KDR� human cells (Figures 3 and4); and that injection of hCD34�/CD45� cells, the putativeECFC progenitors,10 produced the same segregation patterninto various vascular districts (Supplemental Figure III).Taken together, these results suggest that blastula-injectedhCD34� cells were committed to multiple differentiation into
Figure 5. Rescue of angiogenic defects in Vegf-c-depletedembryos by blastula transplantation of hCD34� cells. Shown isin vivo knockdown of Vegf-c by injection of antisense MO RNAat 1 to 8 cell stage into Tg(fli1:enhanced green fluorescent pro-tein [EGFP])y1 embryos. A, Low-power views of a control (Fli-GFP) and 2 Vegf-c MO RNA–injected embryos (Fli-GFP�VEGF-C). Note the absence of the bifurcation between the 2 lateraldorsal aorta branches in midpanel (*) and the irregular spac-ing of the intersomitic vessels (arrows), caused by absence of
Vegf-c expression. B, Defective vascular phenotype caused byVegf-c in vivo knockdown. Panels on the left show low-powerviews of the head/yolk regions (top) and the trunk/tail regions(bottom). Dashed box in the top left panel (magnified in the topright) shows the absence of the dorsal aorta (da). aa: aorticarches; cd: Cuvier duct. Dashed box in the bottom left panel(magnified in the bottom right) shows the irregular spacing ofthe intersomitic vessels (iv) of a Fli-EGFP �VEGF-C embryo.*Regions of the trunk where defects in the organization of thesevessels and the dorsal lateral vein (dlv) were observed. da indi-cates dorsal artery; cvp, caudal vein plexus. C, Injection of red-labeled hCD34� cells into Vegf-c knockdown embryos rescued thevascular phenotype. Low-power (left) and high-power (right) viewsof regions similar to those represented in B show the presence ofhCD34� cells associated with normal formation of the 2 lateralbranches of the dorsal aorta (*, lda and lda�) and to regular spacingand branching of the intersomitic vessels (iv) in the trunk/tailregions. Note the presence of several red-labeled cells in theCuvier duct (cd), and in the caudal vein plexus (cvp). dlv indicatesdorsal lateral vein; cv, cardinal vein; da, dorsal artery.
1594 Arterioscler Thromb Vasc Biol July 2011
by IMED Korea on October 10, 2011http://atvb.ahajournals.org/Downloaded from
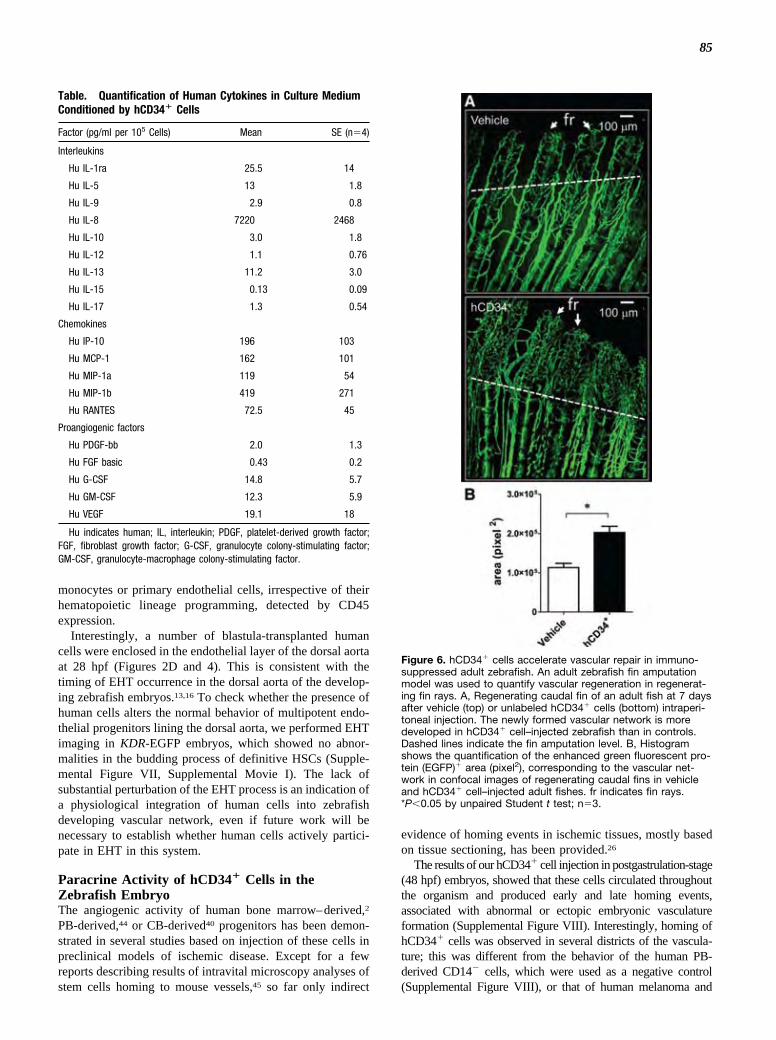
85
monocytes or primary endothelial cells, irrespective of theirhematopoietic lineage programming, detected by CD45expression.
Interestingly, a number of blastula-transplanted humancells were enclosed in the endothelial layer of the dorsal aortaat 28 hpf (Figures 2D and 4). This is consistent with thetiming of EHT occurrence in the dorsal aorta of the develop-ing zebrafish embryos.13,16 To check whether the presence ofhuman cells alters the normal behavior of multipotent endo-thelial progenitors lining the dorsal aorta, we performed EHTimaging in KDR-EGFP embryos, which showed no abnor-malities in the budding process of definitive HSCs (Supple-mental Figure VII, Supplemental Movie I). The lack ofsubstantial perturbation of the EHT process is an indication ofa physiological integration of human cells into zebrafishdeveloping vascular network, even if future work will benecessary to establish whether human cells actively partici-pate in EHT in this system.
Paracrine Activity of hCD34� Cells in theZebrafish EmbryoThe angiogenic activity of human bone marrow–derived,2
PB-derived,44 or CB-derived40 progenitors has been demon-strated in several studies based on injection of these cells inpreclinical models of ischemic disease. Except for a fewreports describing results of intravital microscopy analyses ofstem cells homing to mouse vessels,45 so far only indirect
evidence of homing events in ischemic tissues, mostly basedon tissue sectioning, has been provided.26
The results of our hCD34� cell injection in postgastrulation-stage(48 hpf) embryos, showed that these cells circulated throughoutthe organism and produced early and late homing events,associated with abnormal or ectopic embryonic vasculatureformation (Supplemental Figure VIII). Interestingly, homing ofhCD34� cells was observed in several districts of the vascula-ture; this was different from the behavior of the human PB-derived CD14� cells, which were used as a negative control(Supplemental Figure VIII), or that of human melanoma and
Table. Quantification of Human Cytokines in Culture MediumConditioned by hCD34� Cells
Factor (pg/ml per 105 Cells) Mean SE (n�4)
Interleukins
Hu IL-1ra 25.5 14
Hu IL-5 13 1.8
Hu IL-9 2.9 0.8
Hu IL-8 7220 2468
Hu IL-10 3.0 1.8
Hu IL-12 1.1 0.76
Hu IL-13 11.2 3.0
Hu IL-15 0.13 0.09
Hu IL-17 1.3 0.54
Chemokines
Hu IP-10 196 103
Hu MCP-1 162 101
Hu MIP-1a 119 54
Hu MIP-1b 419 271
Hu RANTES 72.5 45
Proangiogenic factors
Hu PDGF-bb 2.0 1.3
Hu FGF basic 0.43 0.2
Hu G-CSF 14.8 5.7
Hu GM-CSF 12.3 5.9
Hu VEGF 19.1 18
Hu indicates human; IL, interleukin; PDGF, platelet-derived growth factor;FGF, fibroblast growth factor; G-CSF, granulocyte colony-stimulating factor;GM-CSF, granulocyte-macrophage colony-stimulating factor.
Figure 6. hCD34� cells accelerate vascular repair in immuno-suppressed adult zebrafish. An adult zebrafish fin amputationmodel was used to quantify vascular regeneration in regenerat-ing fin rays. A, Regenerating caudal fin of an adult fish at 7 daysafter vehicle (top) or unlabeled hCD34� cells (bottom) intraperi-toneal injection. The newly formed vascular network is moredeveloped in hCD34� cell–injected zebrafish than in controls.Dashed lines indicate the fin amputation level. B, Histogramshows the quantification of the enhanced green fluorescent pro-tein (EGFP)� area (pixel2), corresponding to the vascular net-work in confocal images of regenerating caudal fins in vehicleand hCD34� cell–injected adult fishes. fr indicates fin rays.*P�0.05 by unpaired Student t test; n�3.
Pozzoli et al Human CD34� Cells in Zebrafish 1595
by IMED Korea on October 10, 2011http://atvb.ahajournals.org/Downloaded from
and KDR markers without counterselection for CD45 shouldgive rise only to CFU-ECs38 and support neovascularizationby paracrine interactions with resident endothelial cells. Thislatter hypothesis, however, is contradicted by several exper-imental findings showing that blood-borne stem cells giverise to endothelial cells in vitro and in ischemic tissuesirrespective of their hematopoietic origin2,3,26,29,33,39–42 and bya very recent report showing that CD34�/CD38�/CD45�/CD133� CB progenitors differentiate in culture into clonallyexpanding endothelial cells, which, similarly to ECFCs,participate in neovascularization by incorporating into newblood vessels.18
Transplantation of hCD34� Cells at Early Stagesof Embryonic Zebrafish Development Unravels anUnexpected Hemangioblast-Like BehaviorTransplantation into midblastulas was performed to reveal thefate of hCD34� cells throughout the earliest stages ofvascular development. These experiments showed thatblastula-transplanted hCD34�cells were mostly cosegregatedwith the presumptive zebrafish Scl� and Gata2� progenitorsin the anterior and posterior lateral mesoderm. Thus, hCD34�
cells migrated in concert with zebrafish mesoderm progeni-tors and colonized the anterior region, ie, the site whereembryonic uncommitted progenitors are known to establishprimitive hematopoiesis.43 During somitogenesis, hCD34�
cells migrated medially and posteriorly along the developingintermediate cell mass, to finally colonize the entire cardio-vascular system at the 28 hpf stage. As shown by flowcytometry and immunofluorescence (Figures 3 and 4), thesemorphogenetic events were associated with transient upregu-lation of KDR and overexpression of CD146 at all stages;importantly, this also coincided with downregulation ofhematopoietic progenitor–specific markers CD38 and CD48.It was also remarkable that hCD34� cells showed increasingamounts of monocyte marker CD14 expression; that CD45was not downregulated in KDR� human cells (Figures 3 and4); and that injection of hCD34�/CD45� cells, the putativeECFC progenitors,10 produced the same segregation patterninto various vascular districts (Supplemental Figure III).Taken together, these results suggest that blastula-injectedhCD34� cells were committed to multiple differentiation into
Figure 5. Rescue of angiogenic defects in Vegf-c-depletedembryos by blastula transplantation of hCD34� cells. Shown isin vivo knockdown of Vegf-c by injection of antisense MO RNAat 1 to 8 cell stage into Tg(fli1:enhanced green fluorescent pro-tein [EGFP])y1 embryos. A, Low-power views of a control (Fli-GFP) and 2 Vegf-c MO RNA–injected embryos (Fli-GFP�VEGF-C). Note the absence of the bifurcation between the 2 lateraldorsal aorta branches in midpanel (*) and the irregular spac-ing of the intersomitic vessels (arrows), caused by absence of
Vegf-c expression. B, Defective vascular phenotype caused byVegf-c in vivo knockdown. Panels on the left show low-powerviews of the head/yolk regions (top) and the trunk/tail regions(bottom). Dashed box in the top left panel (magnified in the topright) shows the absence of the dorsal aorta (da). aa: aorticarches; cd: Cuvier duct. Dashed box in the bottom left panel(magnified in the bottom right) shows the irregular spacing ofthe intersomitic vessels (iv) of a Fli-EGFP �VEGF-C embryo.*Regions of the trunk where defects in the organization of thesevessels and the dorsal lateral vein (dlv) were observed. da indi-cates dorsal artery; cvp, caudal vein plexus. C, Injection of red-labeled hCD34� cells into Vegf-c knockdown embryos rescued thevascular phenotype. Low-power (left) and high-power (right) viewsof regions similar to those represented in B show the presence ofhCD34� cells associated with normal formation of the 2 lateralbranches of the dorsal aorta (*, lda and lda�) and to regular spacingand branching of the intersomitic vessels (iv) in the trunk/tailregions. Note the presence of several red-labeled cells in theCuvier duct (cd), and in the caudal vein plexus (cvp). dlv indicatesdorsal lateral vein; cv, cardinal vein; da, dorsal artery.
1594 Arterioscler Thromb Vasc Biol July 2011
by IMED Korea on October 10, 2011http://atvb.ahajournals.org/Downloaded from

86
genitor for endothelial, myeloid, and lymphoid precursors in umbilicalcord blood. Circ Res. 2010;107:1460–1469.
19. Lawson ND, Weinstein BM. In vivo imaging of embryonic vasculardevelopment using transgenic zebrafish. Dev Biol. 2002;248:307–318.
20. Zhang XY, Rodaway AR. SCL-GFP transgenic zebrafish: in vivoimaging of blood and endothelial development and identification of theinitial site of definitive hematopoiesis. Dev Biol. 2007;307:179–194.
21. Chen MJ, Yokomizo T, Zeigler BM, Dzierzak E, Speck NA. Runx1 isrequired for the endothelial to haematopoietic cell transition but notthereafter. Nature. 2009;457:887–891.
22. North TE, Goessling W, Peeters M, Li P, Ceol C, Lord AM, Weber GJ,Harris J, Cutting CC, Huang P, Dzierzak E, Zon LI. Hematopoietic stemcell development is dependent on blood flow. Cell. 2009;137:736–748.
23. Lancrin C, Sroczynska P, Serrano AG, Gandillet A, Ferreras C, KouskoffV, Lacaud G. Blood cell generation from the hemangioblast. J Mol Med.2010;88:167–172.
24. Romagnani P, Annunziato F, Liotta F, Lazzeri E, Mazzinghi B, Frosali F,Cosmi L, Maggi L, Lasagni L, Scheffold A, Kruger M, Dimmeler S,Marra F, Gensini G, Maggi E, Romagnani S. CD14�CD34low cells withstem cell phenotypic and functional features are the major source ofcirculating endothelial progenitors. Circ Res. 2005;97:314–322.
25. Ober EA, Olofsson B, Makinen T, Jin SW, Shoji W, Koh GY, Alitalo K,Stainier DY. Vegfc is required for vascular development and endodermmorphogenesis in zebrafish. EMBO Rep. 2004;5:78–84.
26. Pesce M, Orlandi A, Iachininoto MG, Straino S, Torella AR, Rizzuti V,Pompilio G, Bonanno G, Scambia G, Capogrossi MC. Myoendothelialdifferentiation of human umbilical cord blood-derived stem cells in is-chemic limb tissues. Circ Res. 2003;93:e51–e62.
27. Gaipa G, Tilenni M, Straino S, Burba I, Zaccagnini G, Belotti D, Biagi E,Valentini M, Perseghin P, Parma M, Di Campli C, Biondi A, CapogrossiMC, Pompilio G, Pesce M. GMP-based CD133(�) cells isolationmaintains progenitor angiogenic properties and enhances standardizationin cardiovascular cell therapy. J Cell Mol Med. 2010;14:1619–1634.
28. Kocher AA, Schuster MD, Szabolcs MJ, Takuma S, Burkhoff D, WangJ, Homma S, Edwards NM, Itescu S. Neovascularization of ischemicmyocardium by human bone-marrow-derived angioblasts prevents car-diomyocyte apoptosis, reduces remodeling and improves cardiacfunction. Nat Med. 2001;7:430–436.
29. Botta R, Gao E, Stassi G, Bonci D, Pelosi E, Zwas D, Patti M, ColonnaL, Baiocchi M, Coppola S, Ma X, Condorelli G, Peschle C. Heart infarctin NOD-SCID mice: therapeutic vasculogenesis by transplantation ofhuman CD34� cells and low dose CD34�KDR� cells. FASEB J. 2004;18:1392–1394.
30. Kawamoto A, Iwasaki H, Kusano K, Murayama T, Oyamada A, Silver M,Hulbert C, Gavin M, Hanley A, Ma H, Kearney M, Zak V, Asahara T,Losordo DW. CD34-positive cells exhibit increased potency and safetyfor therapeutic neovascularization after myocardial infarction comparedwith total mononuclear cells. Circulation. 2006;114:2163–2169.
31. Ott I, Keller U, Knoedler M, Gotze KS, Doss K, Fischer P, Urlbauer K,Debus G, von Bubnoff N, Rudelius M, Schomig A, Peschel C, Oost-endorp RA. Endothelial-like cells expanded from CD34� blood cellsimprove left ventricular function after experimental myocardialinfarction. FASEB J. 2005;19:992–994.
32. Schatteman GC, Hanlon HD, Jiao C, Dodds SG, Christy BA. Blood-derived angioblasts accelerate blood-flow restoration in diabetic mice.J Clin Invest. 2000;106:571–578.
33. Madeddu P, Emanueli C, Pelosi E, Salis MB, Cerio AM, Bonanno G,Patti M, Stassi G, Condorelli G, Peschle C. Transplantation of low doseCD34�KDR� cells promotes vascular and muscular regeneration inischemic limbs. FASEB J. 2004;18:1737–1739.
34. Bayliss PE, Bellavance KL, Whitehead GG, Abrams JM, Aegerter S,Robbins HS, Cowan DB, Keating MT, O’Reilly T, Wood JM, RobertsTM, Chan J. Chemical modulation of receptor signaling inhibits regen-erative angiogenesis in adult zebrafish. Nat Chem Biol. 2006;2:265–273.
35. Huang CC, Lawson ND, Weinstein BM, Johnson SL. reg6 is required forbranching morphogenesis during blood vessel regeneration in zebrafishcaudal fins. Dev Biol. 2003;264:263–274.
36. Iovine MK. Conserved mechanisms regulate outgrowth in zebrafish fins.Nat Chem Biol. 2007;3:613–618.
37. Medvinsky A, Dzierzak E. Definitive hematopoiesis is autonomouslyinitiated by the AGM region. Cell. 1996;86:897–906.
38. Case J, Mead LE, Bessler WK, Prater D, White HA, Saadatzadeh MR,Bhavsar JR, Yoder MC, Haneline LS, Ingram DA. HumanCD34�AC133�VEGFR-2� cells are not endothelial progenitor cellsbut distinct, primitive hematopoietic progenitors. Exp Hematol. 2007;35:1109–1118.
39. Asahara T, Masuda H, Takahashi T, Kalka C, Pastore C, Silver M,Kearne M, Magner M, Isner JM. Bone marrow origin of endothelialprogenitor cells responsible for postnatal vasculogenesis in physiologicaland pathological neovascularization. Circ Res. 1999;85:221–228.
40. Murohara T, Ikeda H, Duan J, Shintani S, Sasaki K, Eguchi H, OnitsukaI, Matsui K, Imaizumi T. Transplanted cord blood-derived endothelialprecursor cells augment postnatal neovascularization. J Clin Invest. 2000;105:1527–1536.
41. De Falco E, Porcelli D, Torella AR, Straino S, Iachininoto MG, OrlandiA, Truffa S, Biglioli P, Napolitano M, Capogrossi MC, Pesce M. SDF-1involvement in endothelial phenotype and ischemia-induced recruitmentof bone marrow progenitor cells. Blood. 2004;104:3472–3482.
42. Melero-Martin JM, Khan ZA, Picard A, Wu X, Paruchuri S, Bischoff J. Invivo vasculogenic potential of human blood-derived endothelial pro-genitor cells. Blood. 2007;109:4761–4768.
43. Zon LI. Developmental biology of hematopoiesis. Blood. 1995;86:2876–2891.
44. Kalka C, Masuda H, Takahashi T, Kalka-Moll WM, Silver M, KearneyM, Li T, Isner JM, Asahara T. Transplantation of ex vivo expandedendothelial progenitor cells for therapeutic neovascularization. Proc NatlAcad Sci U S A. 2000;97:3422–3427.
45. Mazo IB, Gutierrez-Ramos JC, Frenette PS, Hynes RO, Wagner DD, vonAndrian UH. Hematopoietic progenitor cell rolling in bone marrowmicrovessels: parallel contributions by endothelial selectins and vascularcell adhesion molecule 1. J Exp Med. 1998;188:465–474.
46. Nicoli S, Ribatti D, Cotelli F, Presta M. Mammalian tumor xenograftsinduce neovascularization in zebrafish embryos. Cancer Res. 2007;67:2927–2931.
47. Gnecchi M, Zhang Z, Ni A, Dzau VJ. Paracrine mechanisms in adult stemcell signaling and therapy. Circ Res. 2008;103:1204–1219.
48. Diez-Torre A, Andrade R, Eguizabal C, Lopez E, Arluzea J, Silio M,Arechaga J. Reprogramming of melanoma cells by embryonic microen-vironments. Int J Dev Biol. 2009;53:1563–1568.
49. Dovey MC, Zon LI. Defining cancer stem cells by xenotransplantation inzebrafish. Methods Mol Biol. 2009;568:1–5.
Pozzoli et al Human CD34� Cells in Zebrafish 1597
by IMED Korea on October 10, 2011http://atvb.ahajournals.org/Downloaded from
CACO-2 stem cells, whose presence was mostly associated withthe head; the caudal vein plexus, where they formed microme-tastases; or the yolk region, where they caused ectopic vesselsprouting (Supplemental Figure IV).46
Direct participation in primary angiogenesis was not theonly effect of hCD34� cell injection into developing ze-brafish embryos. In fact, hCD34� cells rescued vasculardefects caused by Vegf-c knockdown (Figure 6), and whentransplanted into immunosuppressed adults, they acceleratedregeneration of the vascular network in the amputated caudalfins. This activity was due to secretion of several cytokinesand chemokines implicated in (lympho)angiogenesis andinflammatory response (Supplemental Table I). Taken to-gether, and in line with the large existing literature on EPCs,these results suggest that transplanted hCD34� cells exert apotent paracrine effect47 that rescues vascular developmentaldefects due to knockdown of Vegf-C, deregulates formationof normal vascular network in postgastrulation zebrafishembryos, and accelerates wound repair in adults.
Concluding RemarksThe identity of human-derived endothelial progenitors is astill open issue. Although the distinction between the 2 EPCtypes10 has allowed reconciliation of the embryo with theadult stem cell worlds, it is still not clear whether, underpathological conditions or in developmental biology contexts,these cells may cross their lineage barriers and noncanoni-cally differentiate into alternative or mixed phenotypes. Ourfindings suggest that this might be the case and call for furtherinvestigation aimed at clarifying the molecular nature of thesepossible events.
To our knowledge, the present study represents the firstexample of successful human stem cell transplantation intodeveloping zebrafish embryos. In fact, apart from previousinvestigations showing the fate of human cancer cells in thedeveloping zebrafish48 and the recent demonstration of an-giogenic activity of mouse and human tumor cell lines,46 noother evidence exists that human stem cells colonize thedeveloping zebrafish embryos and respond to specific devel-opmental cues. In this context, our results may also haveimportant translational implication for in utero correction ofvascular defects in human embryos by injected stem cells.
It has been discussed that the developing zebrafish is aninteresting microenvironment in which modulatory effects onthe fate of injected cells are likely due to the presence ofdevelopmental mechanisms that are conserved among verte-brates.48 In addition, given the relative ease of zebrafishgenetic manipulation systems, xenotransplantation of humancells in zebrafish has been proposed as a reference techniqueto define the properties of human cancer (stem) cells.49 Ourstudy is in line with these emerging concepts and shows theversatility of this system to also address fundamental humandevelopmental and cell biology questions.
AcknowledgmentsWe thank Dr Mammoliti and the obstetrics group at Melzo Hospitalfor CB collection, and we thank Dr Nathan Lawson for theTg(fli1:EGFP)y1 line. We also thank Drs Serena Balasso and LauraMasala for fish breeding and transgenic line screenings and Drs
Gianluca Deflorian and Federica Pezzimenti of the Zebrafish Facilitycare at Istituto FIRC di Oncologia Molecolare Institute, Milan, Italy.
Sources of FundingThis work was supported by European Union–funded grants UlcerTherapy (contract no: LSHB-CT-2005-512102) and Thercord (contractno: LSHB-CT-2005-018817) and by Fondazione Cariplo NOBELProject “Tumour Stem Cells” 2006 (to F.C.).
DisclosuresNone.
References1. Asahara T, Murohara T, Sullivan A, Silver M, van der Zee R, Li T,
Witzenbichler B, Schatteman G, Isner JM. Isolation of putative progenitorendothelial cells for angiogenesis. Science. 1997;275:964–967.
2. Takahashi T, Kalka C, Masuda H, Chen D, Silver M, Kearney M, MagnerM, Isner JM, Asahara T. Ischemia- and cytokine-induced mobilization ofbone marrow-derived endothelial progenitor cells for neovascularization.Nat Med. 1999;5:434–438.
3. Gehling UM, Ergun S, Schumacher U, Wagener C, Pantel K, Otte M,Schuch G, Schafhausen P, Mende T, Kilic N, Kluge K, Schafer B,Hossfeld DK, Fiedler W. In vitro differentiation of endothelial cells fromAC133-positive progenitor cells. Blood. 2000;95:3106–3112.
4. Prater DN, Case J, Ingram DA, Yoder MC. Working hypothesis toredefine endothelial progenitor cells. Leukemia. 2007;21:1141–1149.
5. Ingram DA, Caplice NM, Yoder MC. Unresolved questions, changingdefinitions, and novel paradigms for defining endothelial progenitor cells.Blood. 2005;106:1525–1531.
6. Ingram DA, Mead LE, Tanaka H, Meade V, Fenoglio A, Mortell K,Pollok K, Ferkowicz MJ, Gilley D, Yoder MC. Identification of a novelhierarchy of endothelial progenitor cells using human peripheral andumbilical cord blood. Blood. 2004;104:2752–2760.
7. Yoder MC, Mead LE, Prater D, Krier TR, Mroueh KN, Li F, Krasich R,Temm CJ, Prchal JT, Ingram DA. Redefining endothelial progenitor cellsvia clonal analysis and hematopoietic stem/progenitor cell principals.Blood. 2007;109:1801–1809.
8. Piaggio G, Rosti V, Corselli M, Bertolotti F, Bergamaschi G, Pozzi S,Imperiale D, Chiavarina B, Bonetti E, Novara F, Sessarego M, Villani L,Garuti A, Massa M, Ghio R, Campanelli R, Bacigalupo A, Pecci A,Viarengo G, Zuffardi O, Frassoni F, Barosi G. Endothelial colony-forming cells from patients with chronic myeloproliferative disorders lackthe disease-specific molecular clonality marker. Blood. 2009;114:3127–3130.
9. Hur J, Yoon CH, Kim HS, Choi JH, Kang HJ, Hwang KK, Oh BH, LeeMM, Park YB. Characterization of two types of endothelial progenitorcells and their different contributions to neovasculogenesis. ArteriosclerThromb Vasc Biol. 2004;24:288–293.
10. Timmermans F, Plum J, Yoder MC, Ingram DA, Vandekerckhove B,Case J. Endothelial progenitor cells: identity defined? J Cell Mol Med.2009;13:87–102.
11. Huber TL, Kouskoff V, Fehling HJ, Palis J, Keller G. Haemangioblastcommitment is initiated in the primitive streak of the mouse embryo.Nature. 2004;432:625–630.
12. Vogeli KM, Jin SW, Martin GR, Stainier DY. A common progenitor forhaematopoietic and endothelial lineages in the zebrafish gastrula. Nature.2006;443:337–339.
13. Kissa K, Herbomel P. Blood stem cells emerge from aortic endotheliumby a novel type of cell transition. Nature. 2010;464:112–115.
14. Boisset JC, van Cappellen W, Andrieu-Soler C, Galjart N, Dzierzak E,Robin C. In vivo imaging of haematopoietic cells emerging from themouse aortic endothelium. Nature. 2010;464:116–120.
15. Swiers G, Speck NA, de Bruijn MF. Visualizing blood cell emergencefrom aortic endothelium. Cell Stem Cell. 2010;6:289–290.
16. Bertrand JY, Chi NC, Santoso B, Teng S, Stainier DY, Traver D. Haema-topoietic stem cells derive directly from aortic endothelium during devel-opment. Nature. 2010;464:108–111.
17. Pelosi E, Valtieri M, Coppola S, Botta R, Gabbianelli M, Lulli V,Marziali G, Masella B, Muller R, Sgadari C, Testa U, Bonanno G,Peschle C. Identification of the hemangioblast in postnatal life. Blood.2002;100:3203–3208.
18. Ramos AL, Darabi R, Akbarloo N, Borges L, Catanese J, Dineen SP,Brekken RA, Perlingeiro RC. Clonal analysis reveals a common pro-
1596 Arterioscler Thromb Vasc Biol July 2011
by IMED Korea on October 10, 2011http://atvb.ahajournals.org/Downloaded from

87
genitor for endothelial, myeloid, and lymphoid precursors in umbilicalcord blood. Circ Res. 2010;107:1460–1469.
19. Lawson ND, Weinstein BM. In vivo imaging of embryonic vasculardevelopment using transgenic zebrafish. Dev Biol. 2002;248:307–318.
20. Zhang XY, Rodaway AR. SCL-GFP transgenic zebrafish: in vivoimaging of blood and endothelial development and identification of theinitial site of definitive hematopoiesis. Dev Biol. 2007;307:179–194.
21. Chen MJ, Yokomizo T, Zeigler BM, Dzierzak E, Speck NA. Runx1 isrequired for the endothelial to haematopoietic cell transition but notthereafter. Nature. 2009;457:887–891.
22. North TE, Goessling W, Peeters M, Li P, Ceol C, Lord AM, Weber GJ,Harris J, Cutting CC, Huang P, Dzierzak E, Zon LI. Hematopoietic stemcell development is dependent on blood flow. Cell. 2009;137:736–748.
23. Lancrin C, Sroczynska P, Serrano AG, Gandillet A, Ferreras C, KouskoffV, Lacaud G. Blood cell generation from the hemangioblast. J Mol Med.2010;88:167–172.
24. Romagnani P, Annunziato F, Liotta F, Lazzeri E, Mazzinghi B, Frosali F,Cosmi L, Maggi L, Lasagni L, Scheffold A, Kruger M, Dimmeler S,Marra F, Gensini G, Maggi E, Romagnani S. CD14�CD34low cells withstem cell phenotypic and functional features are the major source ofcirculating endothelial progenitors. Circ Res. 2005;97:314–322.
25. Ober EA, Olofsson B, Makinen T, Jin SW, Shoji W, Koh GY, Alitalo K,Stainier DY. Vegfc is required for vascular development and endodermmorphogenesis in zebrafish. EMBO Rep. 2004;5:78–84.
26. Pesce M, Orlandi A, Iachininoto MG, Straino S, Torella AR, Rizzuti V,Pompilio G, Bonanno G, Scambia G, Capogrossi MC. Myoendothelialdifferentiation of human umbilical cord blood-derived stem cells in is-chemic limb tissues. Circ Res. 2003;93:e51–e62.
27. Gaipa G, Tilenni M, Straino S, Burba I, Zaccagnini G, Belotti D, Biagi E,Valentini M, Perseghin P, Parma M, Di Campli C, Biondi A, CapogrossiMC, Pompilio G, Pesce M. GMP-based CD133(�) cells isolationmaintains progenitor angiogenic properties and enhances standardizationin cardiovascular cell therapy. J Cell Mol Med. 2010;14:1619–1634.
28. Kocher AA, Schuster MD, Szabolcs MJ, Takuma S, Burkhoff D, WangJ, Homma S, Edwards NM, Itescu S. Neovascularization of ischemicmyocardium by human bone-marrow-derived angioblasts prevents car-diomyocyte apoptosis, reduces remodeling and improves cardiacfunction. Nat Med. 2001;7:430–436.
29. Botta R, Gao E, Stassi G, Bonci D, Pelosi E, Zwas D, Patti M, ColonnaL, Baiocchi M, Coppola S, Ma X, Condorelli G, Peschle C. Heart infarctin NOD-SCID mice: therapeutic vasculogenesis by transplantation ofhuman CD34� cells and low dose CD34�KDR� cells. FASEB J. 2004;18:1392–1394.
30. Kawamoto A, Iwasaki H, Kusano K, Murayama T, Oyamada A, Silver M,Hulbert C, Gavin M, Hanley A, Ma H, Kearney M, Zak V, Asahara T,Losordo DW. CD34-positive cells exhibit increased potency and safetyfor therapeutic neovascularization after myocardial infarction comparedwith total mononuclear cells. Circulation. 2006;114:2163–2169.
31. Ott I, Keller U, Knoedler M, Gotze KS, Doss K, Fischer P, Urlbauer K,Debus G, von Bubnoff N, Rudelius M, Schomig A, Peschel C, Oost-endorp RA. Endothelial-like cells expanded from CD34� blood cellsimprove left ventricular function after experimental myocardialinfarction. FASEB J. 2005;19:992–994.
32. Schatteman GC, Hanlon HD, Jiao C, Dodds SG, Christy BA. Blood-derived angioblasts accelerate blood-flow restoration in diabetic mice.J Clin Invest. 2000;106:571–578.
33. Madeddu P, Emanueli C, Pelosi E, Salis MB, Cerio AM, Bonanno G,Patti M, Stassi G, Condorelli G, Peschle C. Transplantation of low doseCD34�KDR� cells promotes vascular and muscular regeneration inischemic limbs. FASEB J. 2004;18:1737–1739.
34. Bayliss PE, Bellavance KL, Whitehead GG, Abrams JM, Aegerter S,Robbins HS, Cowan DB, Keating MT, O’Reilly T, Wood JM, RobertsTM, Chan J. Chemical modulation of receptor signaling inhibits regen-erative angiogenesis in adult zebrafish. Nat Chem Biol. 2006;2:265–273.
35. Huang CC, Lawson ND, Weinstein BM, Johnson SL. reg6 is required forbranching morphogenesis during blood vessel regeneration in zebrafishcaudal fins. Dev Biol. 2003;264:263–274.
36. Iovine MK. Conserved mechanisms regulate outgrowth in zebrafish fins.Nat Chem Biol. 2007;3:613–618.
37. Medvinsky A, Dzierzak E. Definitive hematopoiesis is autonomouslyinitiated by the AGM region. Cell. 1996;86:897–906.
38. Case J, Mead LE, Bessler WK, Prater D, White HA, Saadatzadeh MR,Bhavsar JR, Yoder MC, Haneline LS, Ingram DA. HumanCD34�AC133�VEGFR-2� cells are not endothelial progenitor cellsbut distinct, primitive hematopoietic progenitors. Exp Hematol. 2007;35:1109–1118.
39. Asahara T, Masuda H, Takahashi T, Kalka C, Pastore C, Silver M,Kearne M, Magner M, Isner JM. Bone marrow origin of endothelialprogenitor cells responsible for postnatal vasculogenesis in physiologicaland pathological neovascularization. Circ Res. 1999;85:221–228.
40. Murohara T, Ikeda H, Duan J, Shintani S, Sasaki K, Eguchi H, OnitsukaI, Matsui K, Imaizumi T. Transplanted cord blood-derived endothelialprecursor cells augment postnatal neovascularization. J Clin Invest. 2000;105:1527–1536.
41. De Falco E, Porcelli D, Torella AR, Straino S, Iachininoto MG, OrlandiA, Truffa S, Biglioli P, Napolitano M, Capogrossi MC, Pesce M. SDF-1involvement in endothelial phenotype and ischemia-induced recruitmentof bone marrow progenitor cells. Blood. 2004;104:3472–3482.
42. Melero-Martin JM, Khan ZA, Picard A, Wu X, Paruchuri S, Bischoff J. Invivo vasculogenic potential of human blood-derived endothelial pro-genitor cells. Blood. 2007;109:4761–4768.
43. Zon LI. Developmental biology of hematopoiesis. Blood. 1995;86:2876–2891.
44. Kalka C, Masuda H, Takahashi T, Kalka-Moll WM, Silver M, KearneyM, Li T, Isner JM, Asahara T. Transplantation of ex vivo expandedendothelial progenitor cells for therapeutic neovascularization. Proc NatlAcad Sci U S A. 2000;97:3422–3427.
45. Mazo IB, Gutierrez-Ramos JC, Frenette PS, Hynes RO, Wagner DD, vonAndrian UH. Hematopoietic progenitor cell rolling in bone marrowmicrovessels: parallel contributions by endothelial selectins and vascularcell adhesion molecule 1. J Exp Med. 1998;188:465–474.
46. Nicoli S, Ribatti D, Cotelli F, Presta M. Mammalian tumor xenograftsinduce neovascularization in zebrafish embryos. Cancer Res. 2007;67:2927–2931.
47. Gnecchi M, Zhang Z, Ni A, Dzau VJ. Paracrine mechanisms in adult stemcell signaling and therapy. Circ Res. 2008;103:1204–1219.
48. Diez-Torre A, Andrade R, Eguizabal C, Lopez E, Arluzea J, Silio M,Arechaga J. Reprogramming of melanoma cells by embryonic microen-vironments. Int J Dev Biol. 2009;53:1563–1568.
49. Dovey MC, Zon LI. Defining cancer stem cells by xenotransplantation inzebrafish. Methods Mol Biol. 2009;568:1–5.
Pozzoli et al Human CD34� Cells in Zebrafish 1597
by IMED Korea on October 10, 2011http://atvb.ahajournals.org/Downloaded from
CACO-2 stem cells, whose presence was mostly associated withthe head; the caudal vein plexus, where they formed microme-tastases; or the yolk region, where they caused ectopic vesselsprouting (Supplemental Figure IV).46
Direct participation in primary angiogenesis was not theonly effect of hCD34� cell injection into developing ze-brafish embryos. In fact, hCD34� cells rescued vasculardefects caused by Vegf-c knockdown (Figure 6), and whentransplanted into immunosuppressed adults, they acceleratedregeneration of the vascular network in the amputated caudalfins. This activity was due to secretion of several cytokinesand chemokines implicated in (lympho)angiogenesis andinflammatory response (Supplemental Table I). Taken to-gether, and in line with the large existing literature on EPCs,these results suggest that transplanted hCD34� cells exert apotent paracrine effect47 that rescues vascular developmentaldefects due to knockdown of Vegf-C, deregulates formationof normal vascular network in postgastrulation zebrafishembryos, and accelerates wound repair in adults.
Concluding RemarksThe identity of human-derived endothelial progenitors is astill open issue. Although the distinction between the 2 EPCtypes10 has allowed reconciliation of the embryo with theadult stem cell worlds, it is still not clear whether, underpathological conditions or in developmental biology contexts,these cells may cross their lineage barriers and noncanoni-cally differentiate into alternative or mixed phenotypes. Ourfindings suggest that this might be the case and call for furtherinvestigation aimed at clarifying the molecular nature of thesepossible events.
To our knowledge, the present study represents the firstexample of successful human stem cell transplantation intodeveloping zebrafish embryos. In fact, apart from previousinvestigations showing the fate of human cancer cells in thedeveloping zebrafish48 and the recent demonstration of an-giogenic activity of mouse and human tumor cell lines,46 noother evidence exists that human stem cells colonize thedeveloping zebrafish embryos and respond to specific devel-opmental cues. In this context, our results may also haveimportant translational implication for in utero correction ofvascular defects in human embryos by injected stem cells.
It has been discussed that the developing zebrafish is aninteresting microenvironment in which modulatory effects onthe fate of injected cells are likely due to the presence ofdevelopmental mechanisms that are conserved among verte-brates.48 In addition, given the relative ease of zebrafishgenetic manipulation systems, xenotransplantation of humancells in zebrafish has been proposed as a reference techniqueto define the properties of human cancer (stem) cells.49 Ourstudy is in line with these emerging concepts and shows theversatility of this system to also address fundamental humandevelopmental and cell biology questions.
AcknowledgmentsWe thank Dr Mammoliti and the obstetrics group at Melzo Hospitalfor CB collection, and we thank Dr Nathan Lawson for theTg(fli1:EGFP)y1 line. We also thank Drs Serena Balasso and LauraMasala for fish breeding and transgenic line screenings and Drs
Gianluca Deflorian and Federica Pezzimenti of the Zebrafish Facilitycare at Istituto FIRC di Oncologia Molecolare Institute, Milan, Italy.
Sources of FundingThis work was supported by European Union–funded grants UlcerTherapy (contract no: LSHB-CT-2005-512102) and Thercord (contractno: LSHB-CT-2005-018817) and by Fondazione Cariplo NOBELProject “Tumour Stem Cells” 2006 (to F.C.).
DisclosuresNone.
References1. Asahara T, Murohara T, Sullivan A, Silver M, van der Zee R, Li T,
Witzenbichler B, Schatteman G, Isner JM. Isolation of putative progenitorendothelial cells for angiogenesis. Science. 1997;275:964–967.
2. Takahashi T, Kalka C, Masuda H, Chen D, Silver M, Kearney M, MagnerM, Isner JM, Asahara T. Ischemia- and cytokine-induced mobilization ofbone marrow-derived endothelial progenitor cells for neovascularization.Nat Med. 1999;5:434–438.
3. Gehling UM, Ergun S, Schumacher U, Wagener C, Pantel K, Otte M,Schuch G, Schafhausen P, Mende T, Kilic N, Kluge K, Schafer B,Hossfeld DK, Fiedler W. In vitro differentiation of endothelial cells fromAC133-positive progenitor cells. Blood. 2000;95:3106–3112.
4. Prater DN, Case J, Ingram DA, Yoder MC. Working hypothesis toredefine endothelial progenitor cells. Leukemia. 2007;21:1141–1149.
5. Ingram DA, Caplice NM, Yoder MC. Unresolved questions, changingdefinitions, and novel paradigms for defining endothelial progenitor cells.Blood. 2005;106:1525–1531.
6. Ingram DA, Mead LE, Tanaka H, Meade V, Fenoglio A, Mortell K,Pollok K, Ferkowicz MJ, Gilley D, Yoder MC. Identification of a novelhierarchy of endothelial progenitor cells using human peripheral andumbilical cord blood. Blood. 2004;104:2752–2760.
7. Yoder MC, Mead LE, Prater D, Krier TR, Mroueh KN, Li F, Krasich R,Temm CJ, Prchal JT, Ingram DA. Redefining endothelial progenitor cellsvia clonal analysis and hematopoietic stem/progenitor cell principals.Blood. 2007;109:1801–1809.
8. Piaggio G, Rosti V, Corselli M, Bertolotti F, Bergamaschi G, Pozzi S,Imperiale D, Chiavarina B, Bonetti E, Novara F, Sessarego M, Villani L,Garuti A, Massa M, Ghio R, Campanelli R, Bacigalupo A, Pecci A,Viarengo G, Zuffardi O, Frassoni F, Barosi G. Endothelial colony-forming cells from patients with chronic myeloproliferative disorders lackthe disease-specific molecular clonality marker. Blood. 2009;114:3127–3130.
9. Hur J, Yoon CH, Kim HS, Choi JH, Kang HJ, Hwang KK, Oh BH, LeeMM, Park YB. Characterization of two types of endothelial progenitorcells and their different contributions to neovasculogenesis. ArteriosclerThromb Vasc Biol. 2004;24:288–293.
10. Timmermans F, Plum J, Yoder MC, Ingram DA, Vandekerckhove B,Case J. Endothelial progenitor cells: identity defined? J Cell Mol Med.2009;13:87–102.
11. Huber TL, Kouskoff V, Fehling HJ, Palis J, Keller G. Haemangioblastcommitment is initiated in the primitive streak of the mouse embryo.Nature. 2004;432:625–630.
12. Vogeli KM, Jin SW, Martin GR, Stainier DY. A common progenitor forhaematopoietic and endothelial lineages in the zebrafish gastrula. Nature.2006;443:337–339.
13. Kissa K, Herbomel P. Blood stem cells emerge from aortic endotheliumby a novel type of cell transition. Nature. 2010;464:112–115.
14. Boisset JC, van Cappellen W, Andrieu-Soler C, Galjart N, Dzierzak E,Robin C. In vivo imaging of haematopoietic cells emerging from themouse aortic endothelium. Nature. 2010;464:116–120.
15. Swiers G, Speck NA, de Bruijn MF. Visualizing blood cell emergencefrom aortic endothelium. Cell Stem Cell. 2010;6:289–290.
16. Bertrand JY, Chi NC, Santoso B, Teng S, Stainier DY, Traver D. Haema-topoietic stem cells derive directly from aortic endothelium during devel-opment. Nature. 2010;464:108–111.
17. Pelosi E, Valtieri M, Coppola S, Botta R, Gabbianelli M, Lulli V,Marziali G, Masella B, Muller R, Sgadari C, Testa U, Bonanno G,Peschle C. Identification of the hemangioblast in postnatal life. Blood.2002;100:3203–3208.
18. Ramos AL, Darabi R, Akbarloo N, Borges L, Catanese J, Dineen SP,Brekken RA, Perlingeiro RC. Clonal analysis reveals a common pro-
1596 Arterioscler Thromb Vasc Biol July 2011
by IMED Korea on October 10, 2011http://atvb.ahajournals.org/Downloaded from