emergence of escherichia coli critically buckled motile helices … · fig. 1. (a) confocal image...
TRANSCRIPT

BIO
PHYS
ICS
AN
DCO
MPU
TATI
ON
AL
BIO
LOG
Y
Emergence of Escherichia coli critically buckled motilehelices under stressTrung V. Phana, Ryan J. Morrisb, Ho Tat Lama, Phuson Hulammc, Matthew E. Blackd, Julia Bose, and Robert H. Austina,1
aDepartment of Physics, Princeton University, Princeton, NJ 08544; bSchool of Physics and Astronomy, University of Edinburgh, Edinburgh EH8 9YL,Scotland; cNikon Instruments Inc., Melville, NY 11742; dLewis-Sigler Institute for Integrative Genomics, Princeton University, Princeton, NJ 08544; andeDepartment of Genomes and Genetics, Pasteur Institute, Paris, France
Contributed by Robert H. Austin, October 18, 2018 (sent for review June 4, 2018; reviewed by Raymond E. Goldstein and Yuhai Tu)
Bacteria under external stress can reveal unexpected emergentphenotypes. We show that the intensely studied bacteriumEscherichia coli can transform into long, highly motile helicalfilaments poized at a torsional buckling criticality when exposedto minimum inhibitory concentrations of several antibiotics. Whilethe highly motile helices are physically either right- or left-handed,the motile helices always rotate with a right-handed angularvelocity ~ω, which points in the same direction as the translationalvelocity ~vT of the helix. Furthermore, these helical cells do notswim by a “run and tumble” but rather synchronously flip theirspin ~ω and thus translational velocity—backing up rather thantumbling. By increasing the translational persistence length,these dynamics give rise to an effective diffusion coefficient upto 20 times that of a normal E. coli cell. Finally, we propose anevolutionary mechanism for this phenotype’s emergence wherebythe increased effective diffusivity provides a fitness advantage inallowing filamentous cells to more readily escape regions of highexternal stress.
bacteria | helical | motile | buckled | emergent
S tudies of bacterial morphology are typically conducted undernonphysiological conditions that partly or completely immo-
bilize the cell on an agar surface or in a confined channel (1),which puts constraints on cellular morphology (2, 3). Likewise,the swimming behavior of Escherichia coli is usually studiedwith prototypically healthy (short), unstressed cells. Such stud-ies of unstressed cells have given rise to the description of E.coli’s swimming behavior as “run and tumble” dynamics whereinunidirectional “runs” are intermittently disrupted by randomreorientations (“tumbles”) (4).
But even well-studied bacteria such as E. coli can exhibitremarkably emergent and unexpected phenotypes when exposedto external stressors such as antibiotics. The physics and bio-logical fitness of these emergent phenotypes can be surprising.Previous work has shown that both genotoxic (ciprofloxacin) (5,6) and cell wall-targeting (cephalexin) antibiotics (7) give riseto filamentous E. coli. Here we use the nongenotoxic antibioticcephalexin because cephalexin impairs division septum forma-tion but does not disrupt normal DNA replication as doesciprofloxacin. In our experiments, we used the common labo-ratory E. coli strains MG1655 and W3110. Except where notedwe used for detailed analysis a strain of W3110 E. coli that con-stitutively expresses GFP throughout the cytoplasm, allowing foraccurate tracking of the bacteria under the microscope. Filamen-tous E. coli were produced by incubation in lysogeny broth (LB)with 10 µg/mL of the antibiotic cephalexin at 30oC. We notethat while antibiotics can lead to near-complete emergence offilamented E. coli throughout the population, even in the com-plete absence of antibiotics, a small percentage of E. coli arefilamentous, indicating the emergence of the phenomena even atzero stress.
Data were recorded on swimming filaments using a high-speedresonant scanning confocal microscope. After 24 h of incubationat a dose of 10 µg/mL of the antibiotic cephalexin (a final OD of
1.0), we transferred 5 µL of the culture to a concave glass slideat either room temperature T ∼ 25oC (highly motile filaments)or near 0oC (greatly slowed motile filaments) and imaged themusing a Nikon A1R inverted confocal epifluorescence micro-scope (Nikon Instruments). The objective used was a Nikon CFIPlan Apo Lambda 60× Oil. Mechanical helices (springs) wereimaged in the Nikon A1R to correct for any mirror reflection orcamera readout inversion of actual helicity handedness.
The filaments have a high motility and rotate as they swim,although in the 2D projection of a conventional microscopethey appear to wiggle. Because of the highest rotational ratefR = 10 Hz of the filamentous bacteria at 25oC , direct 3D confo-cal visualization of the filament topology was difficult. Instead,we continuously imaged (30 Hz single z plane frame rate) inthe z plane nearest the coverslip and measured the 2D pro-jected length, the translational speed vF , and the rotationalfrequency fF in that z plane. Although we imaged in a singleconfocal z plane for maximum speed, distance from the planecould be determined by the relative brightness of the object. Wemeasured the intensity recorded by the confocal detector of 42nm-diameter Flash Red fluorescent beads (Bangs Laboratories)stuck to the coverslip as a function of distance from the cover-slip plane by moving the stage known distances in the verticaldirection; this brightness vs. distance calibration scale was thenused to determine the physical displacement of sections of thefilament from the coverslip plane as they rotated in the fluid.
Significance
We have discovered an emergent mechanism by whichEscherichia coli can escape high-stress regions, such as near-lethal concentrations of antibiotics, by forming long motilehelical filaments that are poized at the critical shear buck-ling point: 2π twist rotations independent of the length ofthe filament. All filaments, independent of length, have thesame twist, indicating that this is a highly evolved response.The helices do not tumble as nonstressed bacteria do butrather, over a period of tens of seconds, reverse direction. Theresult is that the persistence length of the filaments’ transla-tional motion is much larger than in unstressed normal-sizebacteria, giving filaments an extremely large effective diffu-sion coefficient, allowing the bacteria to escape high existen-tial stress.
Author contributions: T.V.P., R.J.M., M.E.B., J.B., and R.H.A. designed research; T.V.P.,R.J.M., H.T.L., P.H., M.E.B., J.B., and R.H.A. performed research; T.V.P., P.H., and R.J.M.contributed new reagents/analytic tools; T.V.P., R.J.M., H.T.L., P.H., M.E.B., J.B., and R.H.A.analyzed data; and T.V.P., R.J.M., M.E.B., J.B., and R.H.A. wrote the paper.y
Reviewers: R.E.G., University of Cambridge; and Y.T., IBM Thomas J. Watson ResearchCenter.y
The authors declare no conflict of interest.y
Published under the PNAS license.y1 To whom correspondence may be addressed. Email: [email protected]
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1809374115/-/DCSupplemental.y
Published online November 29, 2018.
www.pnas.org/cgi/doi/10.1073/pnas.1809374115 PNAS | December 18, 2018 | vol. 115 | no. 51 | 12979–12984
Dow
nloa
ded
by g
uest
on
Feb
ruar
y 18
, 202
0
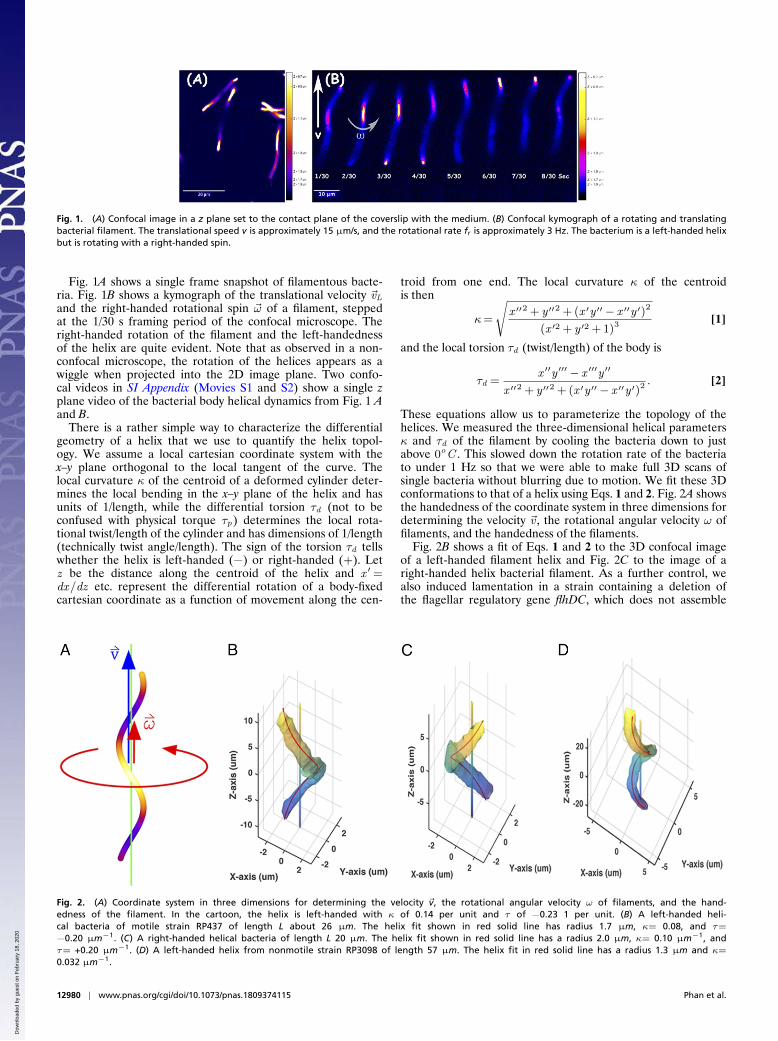
Fig. 1. (A) Confocal image in a z plane set to the contact plane of the coverslip with the medium. (B) Confocal kymograph of a rotating and translatingbacterial filament. The translational speed v is approximately 15 µm/s, and the rotational rate fr is approximately 3 Hz. The bacterium is a left-handed helixbut is rotating with a right-handed spin.
Fig. 1A shows a single frame snapshot of filamentous bacte-ria. Fig. 1B shows a kymograph of the translational velocity ~vLand the right-handed rotational spin ~ω of a filament, steppedat the 1/30 s framing period of the confocal microscope. Theright-handed rotation of the filament and the left-handednessof the helix are quite evident. Note that as observed in a non-confocal microscope, the rotation of the helices appears as awiggle when projected into the 2D image plane. Two confo-cal videos in SI Appendix (Movies S1 and S2) show a single zplane video of the bacterial body helical dynamics from Fig. 1 Aand B.
There is a rather simple way to characterize the differentialgeometry of a helix that we use to quantify the helix topol-ogy. We assume a local cartesian coordinate system with thex–y plane orthogonal to the local tangent of the curve. Thelocal curvature κ of the centroid of a deformed cylinder deter-mines the local bending in the x–y plane of the helix and hasunits of 1/length, while the differential torsion τd (not to beconfused with physical torque τp) determines the local rota-tional twist/length of the cylinder and has dimensions of 1/length(technically twist angle/length). The sign of the torsion τd tellswhether the helix is left-handed (−) or right-handed (+). Letz be the distance along the centroid of the helix and x ′=dx/dz etc. represent the differential rotation of a body-fixedcartesian coordinate as a function of movement along the cen-
troid from one end. The local curvature κ of the centroidis then
κ=
√x ′′2 + y ′′2 + (x ′y ′′− x ′′y ′)2
(x ′2 + y ′2 + 1)3[1]
and the local torsion τd (twist/length) of the body is
τd =x ′′y ′′′− x ′′′y ′′
x ′′2 + y ′′2 + (x ′y ′′− x ′′y ′)2. [2]
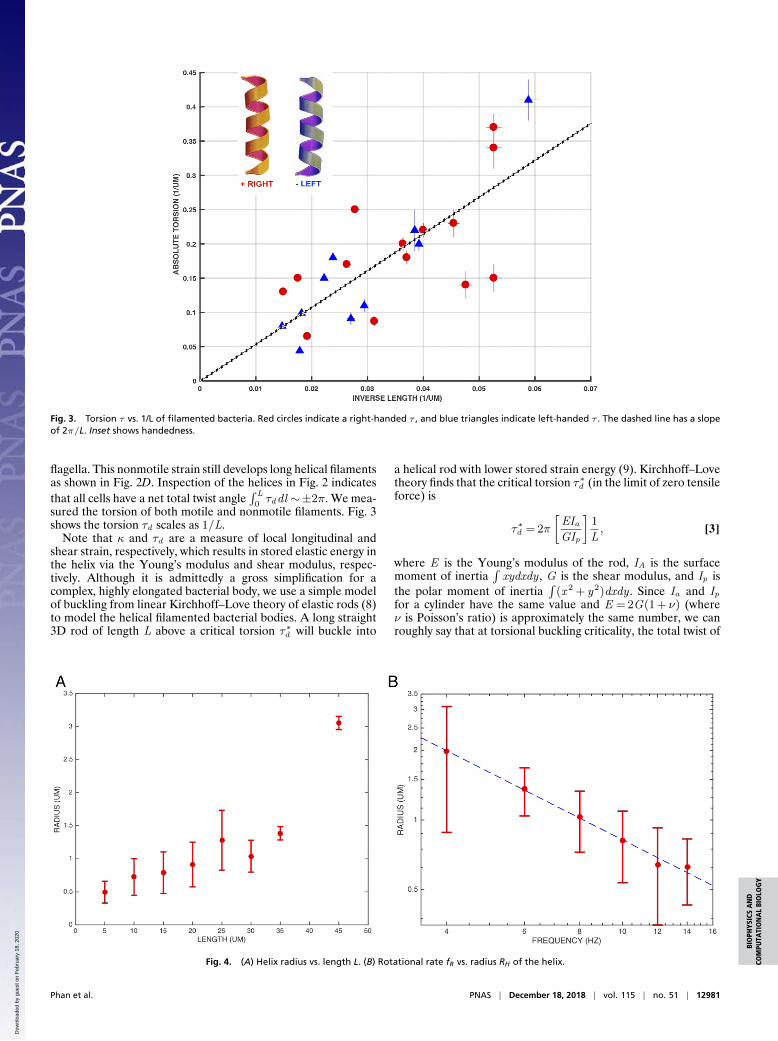
These equations allow us to parameterize the topology of thehelices. We measured the three-dimensional helical parametersκ and τd of the filament by cooling the bacteria down to justabove 0oC . This slowed down the rotation rate of the bacteriato under 1 Hz so that we were able to make full 3D scans ofsingle bacteria without blurring due to motion. We fit these 3Dconformations to that of a helix using Eqs. 1 and 2. Fig. 2A showsthe handedness of the coordinate system in three dimensions fordetermining the velocity ~v , the rotational angular velocity ω offilaments, and the handedness of the filaments.
Fig. 2B shows a fit of Eqs. 1 and 2 to the 3D confocal imageof a left-handed filament helix and Fig. 2C to the image of aright-handed helix bacterial filament. As a further control, wealso induced lamentation in a strain containing a deletion ofthe flagellar regulatory gene flhDC, which does not assemble
Fig. 2. (A) Coordinate system in three dimensions for determining the velocity ~v, the rotational angular velocity ω of filaments, and the hand-edness of the filament. In the cartoon, the helix is left-handed with κ of 0.14 per unit and τ of −0.23 1 per unit. (B) A left-handed heli-cal bacteria of motile strain RP437 of length L about 26 µm. The helix fit shown in red solid line has radius 1.7 µm, κ= 0.08, and τ=
−0.20 µm−1. (C) A right-handed helical bacteria of length L 20 µm. The helix fit shown in red solid line has a radius 2.0 µm, κ= 0.10 µm−1, andτ= +0.20 µm−1. (D) A left-handed helix from nonmotile strain RP3098 of length 57 µm. The helix fit in red solid line has a radius 1.3 µm and κ=
0.032 µm−1.
12980 | www.pnas.org/cgi/doi/10.1073/pnas.1809374115 Phan et al.
Dow
nloa
ded
by g
uest
on
Feb
ruar
y 18
, 202
0
BIO
PHYS
ICS
AN
DCO
MPU
TATI
ON
AL
BIO
LOG
Y
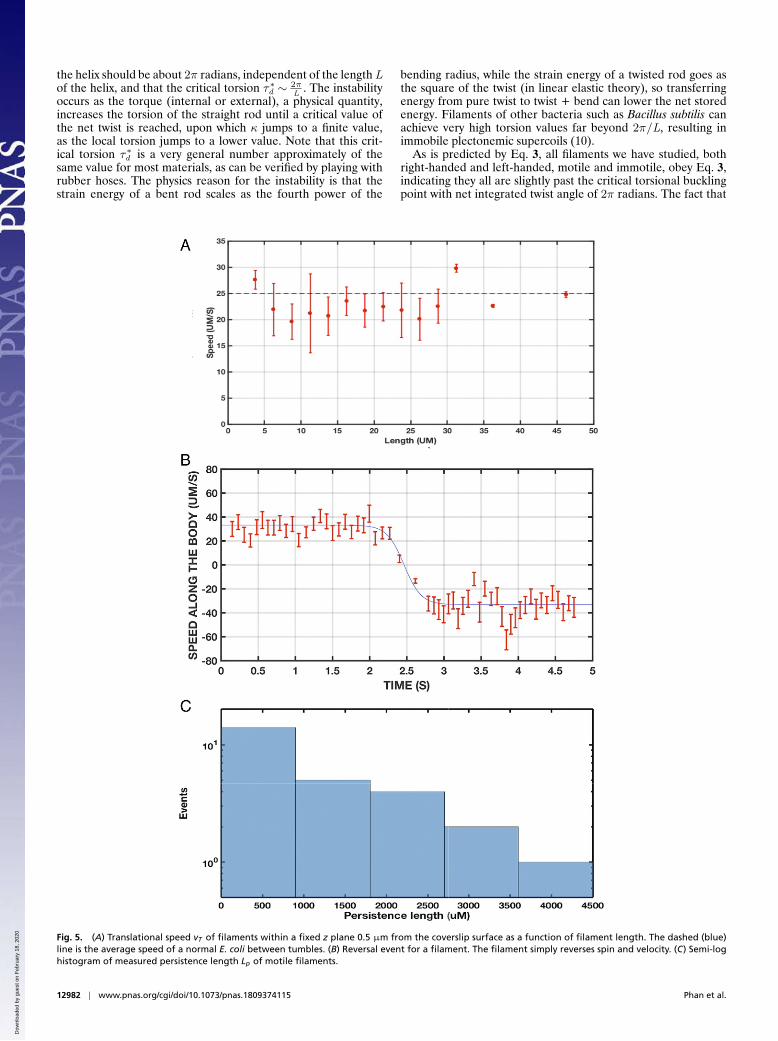
Fig. 3. Torsion τ vs. 1/L of filamented bacteria. Red circles indicate a right-handed τ , and blue triangles indicate left-handed τ . The dashed line has a slopeof 2π/L. Inset shows handedness.
flagella. This nonmotile strain still develops long helical filamentsas shown in Fig. 2D. Inspection of the helices in Fig. 2 indicatesthat all cells have a net total twist angle
∫ L
0τddl ∼±2π. We mea-
sured the torsion of both motile and nonmotile filaments. Fig. 3shows the torsion τd scales as 1/L.
Note that κ and τd are a measure of local longitudinal andshear strain, respectively, which results in stored elastic energy inthe helix via the Young’s modulus and shear modulus, respec-tively. Although it is admittedly a gross simplification for acomplex, highly elongated bacterial body, we use a simple modelof buckling from linear Kirchhoff–Love theory of elastic rods (8)to model the helical filamented bacterial bodies. A long straight3D rod of length L above a critical torsion τ∗d will buckle into
a helical rod with lower stored strain energy (9). Kirchhoff–Lovetheory finds that the critical torsion τ∗d (in the limit of zero tensileforce) is
τ∗d = 2π
[EIaGIp
]1
L, [3]
where E is the Young’s modulus of the rod, IA is the surfacemoment of inertia
∫xydxdy , G is the shear modulus, and Ip is
the polar moment of inertia∫
(x2 + y2)dxdy . Since Ia and Ipfor a cylinder have the same value and E = 2G(1 + ν) (whereν is Poisson’s ratio) is approximately the same number, we canroughly say that at torsional buckling criticality, the total twist of
Fig. 4. (A) Helix radius vs. length L. (B) Rotational rate fR vs. radius RH of the helix.
Phan et al. PNAS | December 18, 2018 | vol. 115 | no. 51 | 12981
Dow
nloa
ded
by g
uest
on
Feb
ruar
y 18
, 202
0
the helix should be about 2π radians, independent of the length Lof the helix, and that the critical torsion τ∗d ∼ 2π
L. The instability
occurs as the torque (internal or external), a physical quantity,increases the torsion of the straight rod until a critical value ofthe net twist is reached, upon which κ jumps to a finite value,as the local torsion jumps to a lower value. Note that this crit-ical torsion τ∗d is a very general number approximately of thesame value for most materials, as can be verified by playing withrubber hoses. The physics reason for the instability is that thestrain energy of a bent rod scales as the fourth power of the
bending radius, while the strain energy of a twisted rod goes asthe square of the twist (in linear elastic theory), so transferringenergy from pure twist to twist + bend can lower the net storedenergy. Filaments of other bacteria such as Bacillus subtilis canachieve very high torsion values far beyond 2π/L, resulting inimmobile plectonemic supercoils (10).
As is predicted by Eq. 3, all filaments we have studied, bothright-handed and left-handed, motile and immotile, obey Eq. 3,indicating they all are slightly past the critical torsional bucklingpoint with net integrated twist angle of 2π radians. The fact that
Fig. 5. (A) Translational speed vT of filaments within a fixed z plane 0.5 µm from the coverslip surface as a function of filament length. The dashed (blue)line is the average speed of a normal E. coli between tumbles. (B) Reversal event for a filament. The filament simply reverses spin and velocity. (C) Semi-loghistogram of measured persistence length Lp of motile filaments.
12982 | www.pnas.org/cgi/doi/10.1073/pnas.1809374115 Phan et al.
Dow
nloa
ded
by g
uest
on
Feb
ruar
y 18
, 202
0

BIO
PHYS
ICS
AN
DCO
MPU
TATI
ON
AL
BIO
LOG
Y
E. coli RP3098 (∆flhA–flhD) also is poized at criticality suggeststhat the buckling instability we see here is not due to a hydro-dynamic shear loading of the swimming filaments and thus isprobably not an externally imposed dynamical bucking transition(11) but rather due to internally generated strain registration.However, unlike the strain registration topology observed byMendelson (12) in stationary B. subtilis, in our case the buck-ling always stops exactly at the critical value independent of thelength of the bacterium.
The rotational handedness of the helices is also unusual: Thefilaments always rotate with angular velocity ~ω parallel to thetranslational velocity ~v . However, as seen in Fig. 2, not all fil-aments are physically right-handed as would be expected if thefilaments are “screwing” through the liquid at low Re (13). Thatis, a left-handed helix should rotate with ~ωL in the oppositedirection from ~vT if the handedness of the helix determinesthe direction of ~vL. But this is not observed: All helices havea right-handed ~ω when moving independent of their physicalhandedness.
Further, we found that the radius rh of the helices increases ina nonlinear way with the length of the helix L as shown in Fig. 4A.However, another constant of the motion, not as surprising as thecriticality of the buckling of the filament, can be found from therelation between the rotation rate of the helical bodies ωR andthe radius of the helix rh . Measurement of the rotational ratefR =ω(R)/2π shows that fR varies inversely with the radius rHof the helix as seen in Fig. 4B, which reveals that rH ∼ f −1.0
R . Thisimplies that the tangential velocity ~vT = ~ωR = 20 µm/ sec is bothconstant and independent of not only the length L of the filamentbut also the radius rH of the helix and equal to the translationspeed vT of the motile helices.
The ultimate propulsion mechanism of the helical E. coli fil-aments is believed to be left-handed rotating helical flagellabundles (14, 15). The motion of a normal 2 µm × 0.5 µmcylindrical E. coli cell in the absence of a chemical gradientis described by a simple, elegant random-walk model basedon alternating sequences of linear runs and random-directiontumbles (4). The helices of the flagella are left-handed and dur-ing linear runs form a single left-handed bundle that pushesthe bacterium by rotating with angular velocity ~ωF , which isdirected opposite to the translational velocity ~vL of the normalbacteria, because of the left-handed torsion τ of the flagellahelix bundle. Normal bacteria always swim with angular veloc-ity ~ωN of the flagella that is oppositely directed from ~vL becausethe bundle is left-handed. The reaction torque on the bac-terium body gives it a right-handed angular velocity ω, whichis always directed in the direction of ~vL for normal bacteria.Since the helical bacterial body filaments also always rotatewith angular velocity ~ω in the same direction as vL, presum-ably the same reaction torque drives their rotation, independentof the body torsion. This finding is in agreement with otherwork on flagella-driven helical swimming bacteria that indicatesthat the helical shape has very little influence on speed of thebacteria (16).
Finally, these helical filaments do not tumble in random direc-tions every second or so but rather simply reverse direction,as shown in Fig. 5; have the same swimming speed vL as nor-mal cells independent of the filament length L, as shown in Fig.5A; and reverse direction much less frequently than normal E.coli. Tracking individual filaments allowed us to determine theswimming persistence length Pf of the filaments using
< cos θ >=e−s/PF , [4]
where s is the arc length of a path of a filament and < cos θ >is the average value of the angle θ between a vector that istangent to the path at a distance s away from the initial posi-tion. Fig. 5C shows the distribution histogram of persistence
lengths recorded using this tracking method. There is a largerange of persistence lengths, with basically an exponential dis-tribution. Using this distribution of the persistence lengths andknowing the constant swimming speed of the filaments, we cancompute the effective diffusion constant DF of the filament.For example, in our dataset, for PF ∼ 500 µm , KF = 2PF (theKuhn length of the filamentous E. coli motion) and d = 3dimensional space:
tF =KF
vF, DF =
K 2F
2dtF=
vFPF
d∼ 3000 µm2/s. [5]
We do not yet know the underlying biological and physical mech-anisms of how the helicity is generated, but the evidence is thatinternal strain buckles the filaments. The programmatic natureof filamentous response to heat stress was originally studied inthe classic work of Mendelson and Cole (17), and we assumea related response occurs in E. coli under antibiotic stress. Cellshape in both gram-negative and -positive bacteria is determinedby the rigid peptidoglycan (PG) cell wall. The PG cell wall isconstructed of glycan strands covalently cross-linked by peptideunits that generate a material that can withstand the internalturgor pressure of the cell and maintain its shape (18). It wasdemonstrated that cells grown in confined geometries retainedtheir shape after release but relaxed after periods of growth (19).How the PG layer is modified during growth and division hasbeen a long-studied problem. MreB is a cytoskeletal protein thatis a homolog of actin and has been shown to form helical con-structs within the cell that direct and recruit the synthesis of PG(20–22). Several papers have shown that during PG synthesis,MreB induces a chiral ordering and physical twisting of the PGwithin the cell wall. The handedness of the chiral ordering/twisthas been observed to be both left- and right-handed, with oftensome bias toward one or the other (23–25). Computational workhas shown that defects or spatial variation in the cross-linkingof the PG can result in different cellular morphologies (26, 27).Indeed, the shape of the helical bacteria Helicobacter pyloriis a result of directed relaxation of cross-links in the PG cellwall (28).
It is very possible that the helices we observe in this workare a result of both the intrinsic chiral twist that is generatedduring PG synthesis and possible defects that result from thestress imposed by the environment. Interestingly, highly spi-raled/helical filaments have been observed in certain mutant E.coli that lack machinery for PG synthesis and septation (29).These higher spiraled structures are reminiscent of different,noncritical modes of a torsional instability of a thin rod andmay provide another route in understanding how the molecu-lar machinery of PG deposition couples to the resultant physicalmorphology of the cell.
Ultimately, emergent phenotypes in biology must have a fit-ness advantage. The extremely large filament diffusion coeffi-cients, up to 20 times that of a normal bacterium, mean thatthe filaments cover much larger regions of space for a fixedtime than a normal bacterium despite their similar translationalspeeds. We suggest that motile filamentous E. coli can exploreenvironments much faster and more widely than normal-lengthbacteria. Helical motile filamentation is an emergent property(30, 31): Genetically, the helical bacterial filaments are identicalto their parents in the absence of mutations, yet their phenotypeis greatly different from the parent’s with a possibly substantiallychanged fitness mission.
ACKNOWLEDGMENTS. The authors thank Mike Nelson and Neal Barlow ofNikon Optics for assistance with the Nikon A1R; Thomas Gregor for suggest-ing data presentations and time on a Nikon A1R; Sujit S. Datta for use ofhis Nikon A1R; and Josh Shaevitz, Angela Dawson, Vincent A. Martinez, andCait E. MacPhee for lively discussions. This work was supported by NSF GrantPHY-1659940.
Phan et al. PNAS | December 18, 2018 | vol. 115 | no. 51 | 12983
Dow
nloa
ded
by g
uest
on
Feb
ruar
y 18
, 202
0

1. Caspi Y (2014) Deformation of filamentous Escherichia coli cells in a microfluidicdevice: A new technique to study cell mechanics. PLoS One 9:e83775.
2. Young KD (2007) Bacterial morphology: Why have different shapes? Curr OpinMicrobiol 10:596–600.
3. Jiang HY, Si FW, Margolin W, Sun SX (2011) Mechanical control of bacterial cell shape.Biophys J 101:327–335.
4. Berg HC, Turner L (1995) Cells of Escherichia coli swim either end forward. Proc NatlAcad Sci USA 92:477–479.
5. Zhang Q, et al. (2011) Acceleration of emergence of bacterial antibiotic resistance inconnected microenvironments. Science 333:1764–1767.
6. Bos J, et al. (2015) Emergence of antibiotic resistance from multinucleated bacterialfilaments. Proc Natl Acad Sci USA 112:178–183.
7. White MA, Eykelenboom JK, Lopez-Vernaza MA, Wilson E, Leach DR (2008)Non-random segregation of sister chromosomes in Escherichia coli. Nature 455:1248–1250.
8. Love A (1944) A Treatise on the Mathematical Theory of Elasticity (Dover Publications,Mineola, NY).
9. Snir Y, Kamien RD (2005) Entropically driven helix formation. Science 307:1067.10. Powers TR, Goldstein RE, Wiggins CH (1999) Supercoiling bacterial filaments. Biol Phys
487:271–278.11. Goldstein RE, Goriely A (2006) Dynamic buckling of morphoelastic filaments. Phys Rev
E 74:010901.12. Mendelson NH (1976) Helical growth of bacillus subtilis: A new model of cell growth.
Proc Natl Acad Sci USA 73:1740–1744.13. Purcell EM (1977) Life at low Reynolds number. Am J Phys 45:3–11.14. Huitorel P (1988) From cilia and flagella to intracellular motility and back again:
A review of a few aspects of microtubule-based motility. Biol Cell 63:249–258.
15. Darnton NC, Turner L, Rojevsky S, Berg HC (2007) On torque and tumbling inswimming Escherichia coli. J Bacteriol 189:1756–1764.
16. Constantino MA, Jabbarzadeh M, Fu HC, Bansil R (2016) Helical and rod-shaped bac-teria swim in helical trajectories with little additional propulsion from helical shape.Sci Adv 2:e1601661.
17. Mendelson NH, Cole RM (1972) Genetic regulation of cell division initiation in Bacillussubtilis. J Bacteriol 112:994–1003.
18. Typas A, Banzhaf M, Gross CA, Vollmer W (2011) From the regulation of pep-tidoglycan synthesis to bacterial growth and morphology. Nat Rev Microbiol10:123–136.
19. Takeuchi S, DiLuzio WR, Weibel DB, Whitesides GM (2005) Controlling the shape offilamentous cells of Escherichia coli. Nano Lett 5:1819–1823.
20. Jones LJ, Carballido-Lopez R, Errington J (2001) Control of cell shape in bacteria:Helical, actin-like filaments in bacillus subtilis. Cell 104:913–922.
21. Vats P, Shih YL, Rothfield L (2009) Assembly of the mreB-associated cytoskeletal ringof Escherichia coli. Mol Microbiol 72:170–182.
22. Kruse T, Bork-Jensen J, Gerdes K (2005) The morphogenetic mreBCD proteins ofEscherichia coli form an essential membrane-bound complex. Mol Microbiol 55:78–89.
23. Wang S, Furchtgott L, Huang KC, Shaevitz JW (2012) Helical insertion of peptido-glycan produces chiral ordering of the bacterial cell wall. Proc Natl Acad Sci USA109:E595–E604.
24. Ouzounov N, et al. (2016) MreB orientation correlates with cell diameter inEscherichia coli. Biophys J 111:1035–1043.
25. Tropini C, et al. (2014) Principles of bacterial cell-size determination revealed by cell-wall synthesis perturbations. Cell Rep 9:1520–1527.
26. Huang KC, Mukhopadhyay R, Wen B, Gitai Z, Wingreen NS (2008) Cell shape andcell-wall organization in gram-negative bacteria. Proc Natl Acad Sci USA 105:19282–19287.
27. Huang KC, Ehrhardt DW, Shaevitz JW (2012) The molecular origins of chiral growthin walled cells. Curr Opin Microbiol 15:707–714.
28. Sycuro LK, et al. (2010) Peptidoglycan crosslinking relaxation promotes Helicobacterpylori’s helical shape and stomach colonization. Cell 141:822–833.
29. Varma A, Young KD (2004) Ftsz collaborates with penicillin binding proteins togenerate bacterial cell shape in Escherichia coli. J Bacteriol 186:6768–6774.
30. Anderson PW (1972) More is different—Broken symmetry and nature of hierarchicalstructure of science. Science 177:393–396.
31. Laughlin RB, Pines D, Schmalian J, Stojkovic BP, Wolynes P (2000) The middle way.Proc Natl Acad Sci USA 97:32–37.
12984 | www.pnas.org/cgi/doi/10.1073/pnas.1809374115 Phan et al.
Dow
nloa
ded
by g
uest
on
Feb
ruar
y 18
, 202
0