embryonic development of connections in turtle pallium
TRANSCRIPT

Embryonic Development of Connectionsin Turtle Pallium
PATRICIA CORDERY AND ZOLTAN MOLNAR*University Laboratory of Physiology, University of Oxford,
Oxford OX1 3PT, United Kingdom
We are interested in similarities and conserved mechanisms in early development of thereptilian and mammalian thalamocortical connections. We set out to analyse connectivity inembryonic turtle brains (Pseudemys scripta elegans, between stages 17 and 25), by usingcarbocyanine dye tracing. From the earliest stages studied, labelling from dorsal and ventralthalamus revealed backlabelled cells among developing thalamic fibres within the lateralforebrain bundle and striatum, which had similar morphology to backlabelled internalcapsule cells in embryonic rat (Molnar and Cordery, 1999). However, thalamic crystalplacements did not label cells in the dorsal ventricular ridge (DVR) at any stage examined.Crystal placements into both dorsal and lateral cortex labelled cells in the DVR and,reciprocally, DVR crystal placements labelled cells in the dorsal and lateral cortices.Retrograde labelling revealed that thalamic fibres arrive in the DVR and dorsal cortex bystage 19. The DVR received projections from the nucleus rotundus and the dorsal cortexexclusively from the perirotundal complex (including lateral geniculate nucleus). Thalamicfibres show this remarkable degree of specificity from the earliest stage we could examine withselective retrograde labelling (stage 19). Our study demonstrates that axons of similar cellsare among the first to reach dorsal and ventral thalamus in mammals and reptiles. Ourconnectional analysis in turtle suggests that some cells of the mammalian primitive internalcapsule are homologous to a cell group within the reptilian lateral forebrain bundle andstriatum and that diverse vertebrate brains might use a highly conserved pattern of earlythalamocortical development. J. Comp. Neurol. 413:26–54, 1999. r 1999 Wiley-Liss, Inc.
Indexing terms: dorsal ventricular ridge; thalamus; internal capsule; lateral forebrain bundle;
lateral cortex; dorsal cortex
As a method of understanding the logic of early develop-mental steps, it could be important to compare pallialdevelopment and identify homologous cell groups betweenreptiles and mammals. Comparison of immature stagesmay reveal features of evolution that are otherwise ob-scured by the complexity of the mature brain, and may alsoreveal that many early cells are transient during develop-ment. To look at development from an evolutionary perspec-tive helps to focus on the most biologically relevant mecha-nisms. Examination of early connectivity could revealsome general building principles related to the evolution-ary origin of the cerebral cortex (Northcutt and Kaas,1995; Karten, 1997; Butler and Molnar, 1998).
On the basis of Golgi preparations of embryonic cerebralcortex, Ramon y Cajal (1909–1911) and Marin-Padilla(1971, 1972, 1988) pointed out that subplate and marginalzone cells in mammals have morphologic similarities withreptilian dorsal cortical cells. Marin-Padilla (1971) pro-posed that the mammalian neocortex develops by means ofa reptilian-style framework. It is possible that subplateand marginal zone cells were present in ancestral amni-
otes and serve as a blueprint for brain development in allextant amniotes; the embryonic mammalian brain mayuse these early networks for its development. The pres-ence of these cells and their transient connections isextremely widespread, as they were described in everymammalian species examined (see Allendoerfer and Shatz,1994; Molnar, 1998). Their homologous cell groups inreptiles, however, is not established.
Arguements based on immunohistochemical studies inadult turtles suggest that there are some similaritiesbetween mammalian primordial plexiform zone (preplate)and dorsal cortex cells in turtle, but several lines of
Grant sponsor: Medical Research Council, United Kingdom; Grantnumber: G9706008.
Dr. Molnar’s present address is: Institut de biologie cellulaire et demorphologie, Rue de Bugnon 9, 1005 Lausanne, Switzerland.
*Correspondence to: Zoltan Molnar, Department of Human Anatomy andGenetics, University of Oxford, South Parks Road, Oxford OX1 3QX,United Kingdom. E-mail: [email protected] orzoltan.molnar@human_anatomy.oxford.ac.uk
Received 15 July 1998; Revised 16 April 1999; Accepted 28 May 1999
THE JOURNAL OF COMPARATIVE NEUROLOGY 413:26–54 (1999)
r 1999 WILEY-LISS, INC.

evidence (Reiner, 1991) argue against a simple homologybetween these structures as proposed by Marin-Padilla(1971). On the basis of the comparison of neurotransmit-ter- and neuropeptide-specific neuronal cell types found inadult mammalian cortex to those found in adult turtle andlizard cortex, Reiner (1991) proposed that only neuronshomologous to those in isocortical layer 5 and 6 werepresent in the dorsal cortex of the reptiles, which areancestral to mammals. Reiner (1991, 1993) postulated theaddition of new cell types, mostly found in layers 2–4 inmammals, to the reptile-mammal lineage during the evolu-tion of the mammalian isocortex. Unfortunately, there arevery few similar comparative studies at early developmen-tal stages, and it is not yet established whether the largelytransient marginal zone or subplate cells of the developingmammalian cortex have homologous cell groups in rep-tiles.
In mammals, there are numerous studies on the earlyconnectivity in developing forebrain, which describe anetwork of transient connections involving cells of thethalamic reticular nucleus, the medial and lateral gangli-onic eminence, in addition to preplate cells in all mamma-lian species examined: cat (McConnell et al., 1989), ferret(Mitrofanis, 1994; Clasca et al., 1995), rat (Mitrofanis,1992; Adams and Baker, 1995; Molnar et al., 1998a; andMolnar and Cordery, 1999), mouse and hamster (Metinand Godement, 1996), Monodelphis domestica (Molnar etal., 1998b), and human (Kostovic and Rakic, 1990; Leviticand Kostovic, 1996). These cell groups are more prominentduring development than in adult (Rakic, 1976; Luskinand Shatz, 1985a,b; Liu and Graybiel, 1992; Mitrofanis,1992; Earle and Mitrofanis, 1996), and a large number ofthem are believed to disappear by preferential cell deathduring embryonic and early postnatal stages. Unfortu-nately, the extent and possible selectivity of their deathhas only been quantified for subplate cells in a few species(see e.g., Luskin and Shatz, 1985; Price et al., 1997). Theextent and timing of the cell death in internal capsule andin the thalamic reticular nucleus is not known. It ispossible that there are several subtypes within these cellgroups, perhaps with different evolutionary origin andonly a subpopulation of the cells are effected. The largelytransient internal capsule cells express chemoattractants(Richards et al., 1997; Metin et al., 1997) and are thoughtto act as guide post cells for the descending early connec-tions (Mitrofanis and Guillery, 1993; Ramcharan andGuillery, 1997; Molnar and Cordery, 1999). Why does early
pallial development use these largely transient cells in agreat variety of mammalian species? Could they be a partof a highly conserved developmental pattern inheritedfrom ancestors? Identifying the homologous cell groups innonmammalian developing brains and understanding thepossible origin of these cells, may help to answer thesequestions. Chelonians are generally believed to be derivedfrom ancestors closest to the stem reptiles, which also gaverise to synapsids and eosuchians. Therefore, they couldprovide an interesting system to study these possiblyconserved mechanisms, because they might show moresimilarities to the presumed ancestors from which themajor phylogenetic categories started to diverge (Goffinet,1983; Reiner, 1993). There are numerous connectionalstudies in the adult turtle (Hall and Ebner, 1970; Reiner etal., 1980; Balaban and Ulinski, 1981a,b; Belekhova et al.,1985, 1992; Belekhova, 1994; Hall et al., 1997), but thereare none during embryonic development. Early hodologicrelationships might be more obvious during development.It is conceivable that some of the cells, their early connec-tions, or both, may be transient and substantially different atembryonic ages from the ones described in adult reptiles.
We set out to extend our studies on thalamocorticaldevelopment of mammals (rodents and marsupials) toreptiles to study the hypothesis that the first diencephalicand telencephalic connections in mammals develop accord-ing to a pattern based on extant reptilian organization.Information about the embryonic development of the mam-malian pallium is increasing rapidly (see Molnar andCordery, 1999, for references), and some comparativestudies on nonmammals have also been performed (seeButler and Hodos, 1996). Unfortunately, most of thesestudies are in adults, and we need developmental studiesin nonmammalian amniotes to compare with the develop-mental steps observed in mammals. The advance of theembryonic tracing techniques (Godement et al., 1987)makes it relatively easy to examine the early connectivityin the developing pallium and facilitates the understand-ing of the general developmental mechanisms in variousspecies. We have studied the development of thalamocorti-cal connections and early pallial organization in turtle sothat we might compare the results with our similar studyin mammals (see Molnar and Cordery, 1999). We wereinterested in examining whether cells in homologoussubdivisions of the developing diencephalon and palliumtend to develop connections with respectively homologouscell groups in mammals and reptiles. One of our valuable
Abbreviations
ADVR anterior dorsal ventricular ridgeBDVR basal dorsal ventricular ridgeCO commissure opticDC dorsal cortexDiI 1,1’-dioctadecyl-3,3,3’,3’-tetramethylindocarbocyanine per-
chlorateDLA dorsolateral anterior nucleusDlh nucleus dorsolateralis hypothalamiDMA dorsomedial anterior nucleusDP dorsal peduncleDT dorsal thalamusDVR dorsal ventricular ridgeHB habenulaHT hypothalamusIC internal capsuleLC lateral cortexLFB lateral forebrain bundle
LGN dorsal lateral geniculate nucleusMC medial cortexMZ marginal zoneOT optic tractPH periventricular hypothalamic nucleusPRN perireticular nucleusPRC perireticular complexPT pallial thickeningRE nucleus reuniensRT rotundus nucleusS septumSP subplateST striatumTRN thalamic reticular nucleusV nucleus ventralisVP ventral peduncleVT ventral thalamus
EMBRYONIC CONNECTIONS IN TURTLE PALLIUM 27
reference points was the thalamus, because the reptilianthalamus has a similar organization to that of mammals(Butler, 1994a, 1995; Pritz, 1995). We were particularlyinterested in the possibility of finding a group of cellssimilar to the ones projecting to the dorsal thalamus inmammals during embryonic development. We focused onthe earliest connections established between thalamusand dorsal cortex and dorsal ventricular ridge (DVR)between stages 17 and 25. We also traced projections fromlateral cortex and lateral forebrain bundle and striatum.
MATERIALS AND METHODS
The study was performed by using 110 eggs from differ-ent embryonic ages (10 each from 22- and 30-day-old and15 each from 20-, 25-, 35-, 40-, 45-, and 50-day-old eggs) ofthe fresh water red-eared slider turtle (Pseudemys scriptaelegans). Development of the turtle embryo is temperaturedependent, and the duration of prehatching is normally 60days at room temperature (25°C). After obtaining permis-sion from the Louisiana Department of Agriculture andForestry, eggs were purchased from Harvey Kliebert’sReptile Farm in Louisiana and shipped to the UnitedKingdom. All experimental protocols were approved byand in accord with the regulations and guidelines of theHome Office (United Kingdom) and University of Oxford.
Fixation
To anaesthetise the embryos, the eggs were chilled inice-water for 15–30 minutes. The embryos were thenremoved from the shell and decapitated. The skulls werehemisected above the brain to provide better access for thefixative, and then the whole heads were fixed in 4%paraformaldehyde in 0.1 mM phosphate buffered saline(PBS), 4°C, pH 7.4) and kept in fixative until the histologicprocessing.
Staging
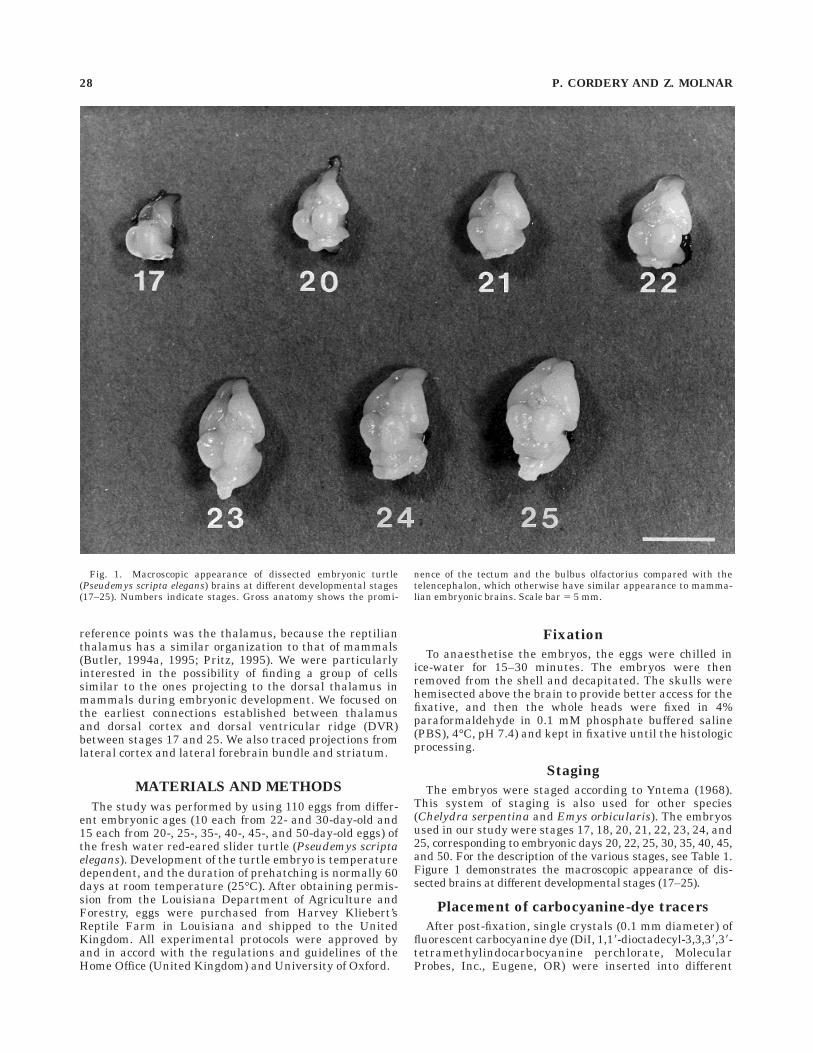
The embryos were staged according to Yntema (1968).This system of staging is also used for other species(Chelydra serpentina and Emys orbicularis). The embryosused in our study were stages 17, 18, 20, 21, 22, 23, 24, and25, corresponding to embryonic days 20, 22, 25, 30, 35, 40, 45,and 50. For the description of the various stages, see Table 1.Figure 1 demonstrates the macroscopic appearance of dis-sected brains at different developmental stages (17–25).
Placement of carbocyanine-dye tracers
After post-fixation, single crystals (0.1 mm diameter) offluorescent carbocyanine dye (DiI, 1,18-dioctadecyl-3,3,38,38-tetramethylindocarbocyanine perchlorate, MolecularProbes, Inc., Eugene, OR) were inserted into different
Fig. 1. Macroscopic appearance of dissected embryonic turtle(Pseudemys scripta elegans) brains at different developmental stages(17–25). Numbers indicate stages. Gross anatomy shows the promi-
nence of the tectum and the bulbus olfactorius compared with thetelencephalon, which otherwise have similar appearance to mamma-lian embryonic brains. Scale bar 5 5 mm.
28 P. CORDERY AND Z. MOLNAR

parts of the diencephalon or telencephalon (see Godementet al., 1987). Table 1 and the Nissl-stained sections illus-trated in Figure 2 describe the number and location ofcrystal placement sites for each of the various experi-ments.
For crystal placements in the diencephalon (dorsal andventral thalamus), the brainstem was first transectedcoronally, rostral to the tectum, to expose the posteriorthalamus. For the lateral forebrain bundle and the ante-rior DVR (ADVR), crystal placement was slightly morerostral to thalamic ones (Fig. 2C,D). For tracing from thedorsal and lateral cortices, crystals were placed directlyinto the surface of the brain after the removal of the skulland the dura mater. A single carbocyanine crystal wasinserted into a small slit made in the tissue with the tip ofa fine pair of forceps. After crystal insertion, the brainswere stored at room temperature (22°C) or at 37°C infixative or in phosphate-buffered saline (PBS) containing0.1% sodium-azide (to prevent contamination of the tis-sue), for 1–2 weeks. After incubation, the brains wereembedded in 5% agar, and 70- to 100-µm coronal sectionswere cut on a Vibratome (General Scientific, RedhillSurrey, UK). Sections were counterstained with bisbenz-imide (10 minutes in 2.5 µg/ml solution in PBS) to revealmajor boundaries and to confirm the presence of chromatinin backlabelled cells. The sections were cover-slipped inPBS and then sealed before being examined under epifluo-rescence illumination and photographed. Some of theserial sections were drawn with a camera lucida.
The tracing method we used was with a carbocyaninedye DiI. In fixed tissue, carbocyanine dyes diffuse antero-gradely and retrogradely. The dye travels ‘‘anterogradely’’along the fibres of labelled cells adjacent to the crystalplacement or along fibres passing through the crystalplacement site (Godemant et al., 1987). Cells extendingprojections into or through the crystal placement also arelabelled through these connections ‘‘retrogradely.’’ Thisproperty enables the carbocyanine dyes to label fibresoriginating from cells in the area of the crystal placementsand cells projecting into the crystal placements.
To clarify the nomenclature of the neuroanatomic areasof our description of our crystal placement and location oflabelled cells and fibres, we used terminology of Balabanand Ulinski (1981a); please refer to Figure 2A–D fordetails. Crystal placement sites into dorsal thalamus (DT)may have included both nucleus rotundus (RT) and theperirotundal complex; i.e., lateral geniculate nucleus(LGN), dorsomedial anterior nucleus (DMA), dorsolateralanterior nucleus (DLA), and nucleus reuniens (RE) (Fig.2A,B). Ventral thalamic crystal placements included thenucleus ventralis (V) and the dorsal and ventral peduncle(DP, VP; Fig. 2A). Hypothalamic tracing sites were near
the ventricle and may have included periventricular hypo-thalamic nucleus (PH) and nucleus dorsolateralis hypo-thalami (Dlh; Fig. 2A). Cortical placements were groupedinto dorsal and lateral cortex (DC and LC; Fig. 2C,D). Sitesin dorsal ventricular ridge were always anterior, thus, intothe ADVR (Fig. 2C). Lateral forebrain bundle (LFB) wasused, because it appeared to be in a similar location to thatof the internal capsule in mammals (Fig. 2C). Fibres andcells were localised in many cytoarchitectural areas, notmentioned above, including the basal dorsal ventricularridge (BDVR) in caudal sections of telencephalon, Figure2A,B; striatum (ST), Figure 2C; medial cortex (MC) andseptum (S), Figure 2D.
RESULTS
We present our results according to crystal placementsites: (1) dorsal and ventral thalamus; (2) hypothalamus;(3) lateral forebrain bundle (internal capsule); (4) anteriordorsal ventricular ridge; (5) dorsal cortex; (6) lateralcortex. Within each section the pattern of fibre labellingand the location of backlabelled cells are discussed inseparate subsections. This format draws attention toregions where no reciprocal projections were found (i.e., nobacklabelled cells were found among the labelled fibres).
Dorsal and ventral thalamiccrystal placements
In five different stages, all together 19 dorsal and 16ventral thalamic DiI crystal placements were performed(see Table 1). The dorsal thalamic crystal placementsincluded cases of crystal placements into the nucleusrotundus, perirotundal complex or cases where the place-ment involved both nuclei (Fig. 2B). Because it is ratherdifficult to achieve selective and exclusive labelling fromthese nuclei at these early stages, the specificity of theemerging connections will be examined from retrogradelabelling experiments (see tracing from DVR and dorsalcortex, below). The ventral thalamic crystal placementsranged from sites near the ventral edge of the dorsalthalamus to the border between thalamus and hypothala-mus.
Distribution of labelled fibres
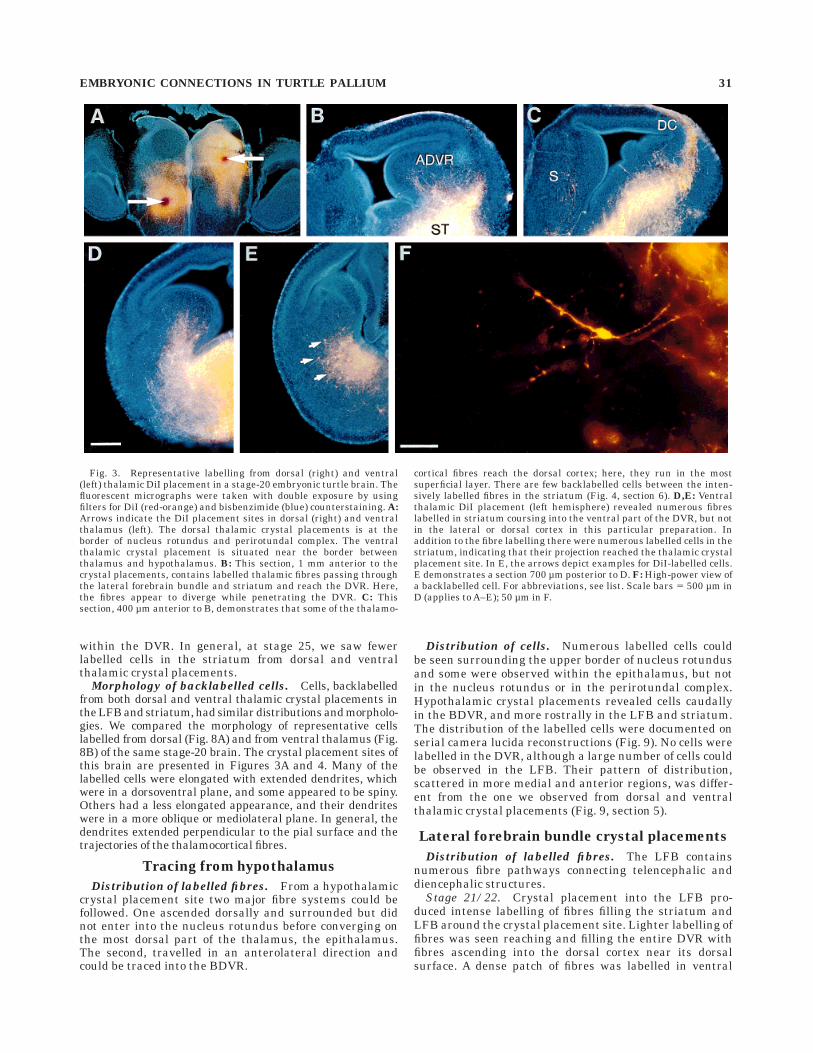
Stage 20/21. Dorsal thalamic crystal placement re-vealed thalamic fibres passing through the lateral fore-brain bundle (LFB) and striatum and reaching the lateralDVR. The labelled fibre bundle is split into two parts. Oneappears to have advanced and turned toward the ventralhalf of the DVR (Fig. 3B), and the other reached dorsalcortex where the fibres ran in the most superficial molecu-lar layer. These fibres could be best seen in the mostanterior sections (Fig. 3C).
Ventral thalamic crystal placement (Fig. 3A) revealedsimilar fibre labelling pattern to that observed from dorsalplacements. The ventral crystal placements probably la-belled the dorsal thalamic projections en passant in addi-tion to possible projections specific for ventral thalamus.Numerous fibres were labelled travelling through thestriatum, LFB, and coursing into the ventral part of theDVR where the fibre bundle became less fasciculated, andindividual fibres diverged from one another as they pen-etrated DVR. Some of these fibres ended in branches.Fibres appeared to be restricted by a boundary between
TABLE 1. Carbocyanine Crystal Placement Sites1
Stage
THAL CORTEX
ADVR LFB HYPDors Ventr Lat Dors Med
17 4 4 — 2 — — — 418 3 — — — — — — 320 4 4 2 3 — — — 221 3 3 3 3 — — — —22 — — — — — 3 3 —23 — — 2 1 1 6 6 —24 2 2 2 2 — 2 — —25 3 3 3 3 1 — — —
1THAL, thalamus; Dors, dorsal; Ventr, ventral; Lat, lateral; Med, medial; ADVR,anterior dorsal ventricular ridge; LFB, lateral forebrain bundle; HYP, hypothalamus.
EMBRYONIC CONNECTIONS IN TURTLE PALLIUM 29

dorsal DVR and dorsal cortex. The fibres labelled fromventral thalamus, in general, were more apparent in moreposterior sections than those fibres labelled from dorsalthalamic crystal placements (see Figs. 4, 5 for serialreconstruction).
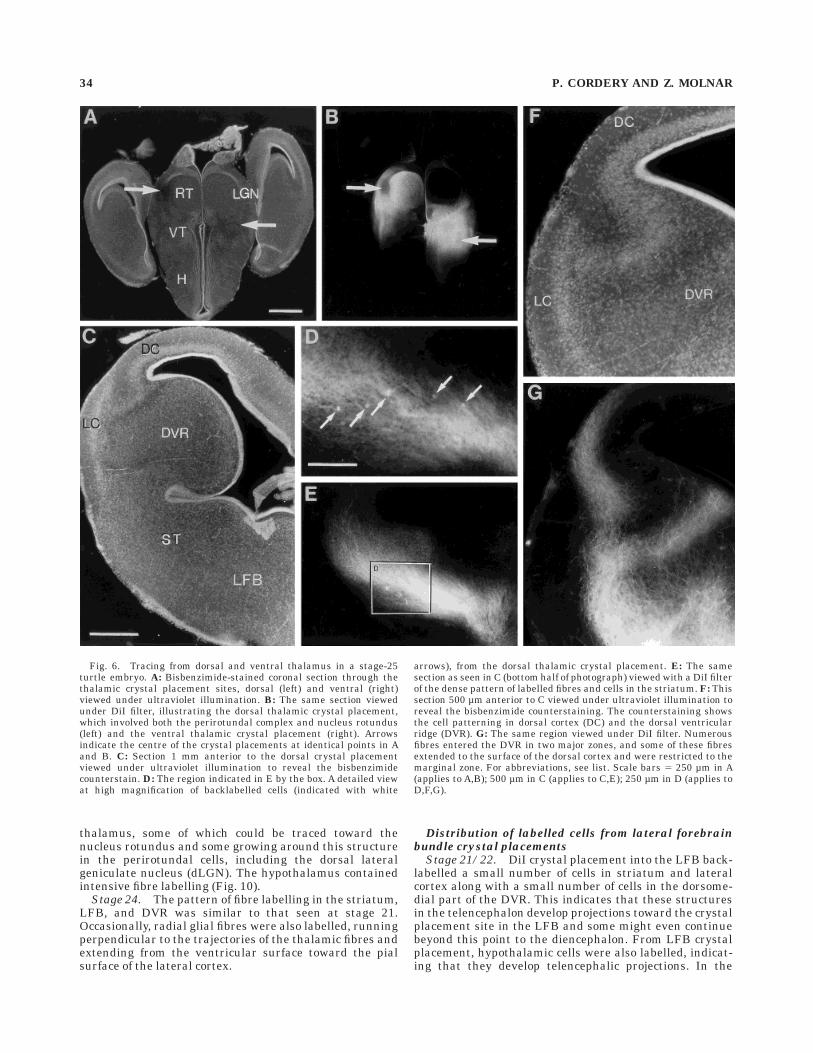
Stage 25. Dorsal thalamic crystal placement labelledeven larger numbers of fibres filling the striatum, LFB,and entering the DVR in two major zones (Fig. 6F,G).Numerous fibres extended into and were seen to almost fillthe ventral part of the DVR, with a narrower band of fibrescrossing the dorsal part and reaching the ependymalsurface. In addition to this, a wide tract of fibres extendedto the surface of the dorsal cortex. This pattern of labelling(involving both DVR and dorsal cortex) was very generalwhen the crystal placement involved both the nucleusrotundus and the perirotundal complex from even earlierstages (Figs. 3, 6). At stage 25, ventral thalamic crystalplacements revealed a similar fibre labelling pattern seenat stage 20/21 (Fig. 7).
Distribution of backlabelled cells from dorsal and
ventral thalamic crystal placements. In addition tolabelled fibres, backlabelled cells were seen in striatum,LFB, and, in some cases, near the ventral border of theDVR, indicating that their projection reached the thalamiccrystal placement sites. Some regions (DVR and dorsalcortex) only contained fibres, but not labelled cells. Al-
though individual cases showed considerable variation,the average number of cells labelled seemed to decreaseslightly over the period studied.
Stage 20/21. At stage 20, dorsal thalamic crystalplacement revealed a small number of scattered backla-belled cells in the striatum. At stage 21, however, crystalplacements (involving both nucleus rotundus and LGN)labelled a noticeably larger number of backlabelled cellswithin the LFB and striatum, but similar to earlier stages,no labelled cells were seen within the body of the DVR.
Ventral thalamic crystal placement showed noticeablymore numerous backlabelled cells in the striatum and LFBcompared with the dorsal at both stages (compare the twosides in the sections presented in Figs. 4, 5). In moreanterior sections, a small number of labelled cells wereseen consistently at the ventral border of the DVR, but notwithin the DVR itself.
Stage 25. From similar dorsal thalamic crystal place-ment (involving both nucleus rotundus and perirotundalcomplex as presented above in Figs. 4, 5), a relativelysparser number of cells were labelled, but the overalldistribution remained similar to that observed at stage20/21 (Fig. 7). The labelled cells were apparent in the LFBand striatum, but not in DVR (Fig. 7). From ventralthalamic crystal placement very few cells were seen in theLFB and striatum, but again no labelled cells were seen
Fig. 2. Major subdivisions of the embryonic turtle brain at stage 22 illustrated on 50-µm-thickVibrotome sections, cut in coronal plane, and Nissl stained. From the serial sections, every fifth section ispresented in anteroposterior sequence (A–D). For abbreviations, see list. Scale bar 5 500 µm.
30 P. CORDERY AND Z. MOLNAR
within the DVR. In general, at stage 25, we saw fewerlabelled cells in the striatum from dorsal and ventralthalamic crystal placements.
Morphology of backlabelled cells. Cells, backlabelledfrom both dorsal and ventral thalamic crystal placements inthe LFB and striatum, had similar distributions and morpholo-gies. We compared the morphology of representative cellslabelled from dorsal (Fig. 8A) and from ventral thalamus (Fig.8B) of the same stage-20 brain. The crystal placement sites ofthis brain are presented in Figures 3A and 4. Many of thelabelled cells were elongated with extended dendrites, whichwere in a dorsoventral plane, and some appeared to be spiny.Others had a less elongated appearance, and their dendriteswere in a more oblique or mediolateral plane. In general, thedendrites extended perpendicular to the pial surface and thetrajectories of the thalamocortical fibres.
Tracing from hypothalamus
Distribution of labelled fibres. From a hypothalamiccrystal placement site two major fibre systems could befollowed. One ascended dorsally and surrounded but didnot enter into the nucleus rotundus before converging onthe most dorsal part of the thalamus, the epithalamus.The second, travelled in an anterolateral direction andcould be traced into the BDVR.
Distribution of cells. Numerous labelled cells couldbe seen surrounding the upper border of nucleus rotundusand some were observed within the epithalamus, but notin the nucleus rotundus or in the perirotundal complex.Hypothalamic crystal placements revealed cells caudallyin the BDVR, and more rostrally in the LFB and striatum.The distribution of the labelled cells were documented onserial camera lucida reconstructions (Fig. 9). No cells werelabelled in the DVR, although a large number of cells couldbe observed in the LFB. Their pattern of distribution,scattered in more medial and anterior regions, was differ-ent from the one we observed from dorsal and ventralthalamic crystal placements (Fig. 9, section 5).
Lateral forebrain bundle crystal placements
Distribution of labelled fibres. The LFB containsnumerous fibre pathways connecting telencephalic anddiencephalic structures.
Stage 21/22. Crystal placement into the LFB pro-duced intense labelling of fibres filling the striatum andLFB around the crystal placement site. Lighter labelling offibres was seen reaching and filling the entire DVR withfibres ascending into the dorsal cortex near its dorsalsurface. A dense patch of fibres was labelled in ventral
Fig. 3. Representative labelling from dorsal (right) and ventral(left) thalamic DiI placement in a stage-20 embryonic turtle brain. Thefluorescent micrographs were taken with double exposure by usingfilters for DiI (red-orange) and bisbenzimide (blue) counterstaining. A:Arrows indicate the DiI placement sites in dorsal (right) and ventralthalamus (left). The dorsal thalamic crystal placements is at theborder of nucleus rotundus and perirotundal complex. The ventralthalamic crystal placement is situated near the border betweenthalamus and hypothalamus. B: This section, 1 mm anterior to thecrystal placements, contains labelled thalamic fibres passing throughthe lateral forebrain bundle and striatum and reach the DVR. Here,the fibres appear to diverge while penetrating the DVR. C: Thissection, 400 µm anterior to B, demonstrates that some of the thalamo-
cortical fibres reach the dorsal cortex; here, they run in the mostsuperficial layer. There are few backlabelled cells between the inten-sively labelled fibres in the striatum (Fig. 4, section 6). D,E: Ventralthalamic DiI placement (left hemisphere) revealed numerous fibreslabelled in striatum coursing into the ventral part of the DVR, but notin the lateral or dorsal cortex in this particular preparation. Inaddition to the fibre labelling there were numerous labelled cells in thestriatum, indicating that their projection reached the thalamic crystalplacement site. In E, the arrows depict examples for DiI-labelled cells.E demonstrates a section 700 µm posterior to D. F: High-power view ofa backlabelled cell. For abbreviations, see list. Scale bars 5 500 µm inD (applies to A–E); 50 µm in F.
EMBRYONIC CONNECTIONS IN TURTLE PALLIUM 31

Fig. 4. The distribution of backlabelled cells and the pattern offibre labelling from dorsal and ventral thalamic crystal placements isillustrated on serial camera lucida drawings from every third section(presented in posterior-anterior order) of the same brain shown atFigure 3 (stage 20). Black circles illustrate the crystal placement sites,darkly shaded regions contain heavy fibre labelling, and lighter
shading illustrates regions with less intensive labelling. Small dotsrepresent backlabelled cells. Dorsal crystal placements revealed onlyfew backlabelled cells in the striatum (sections 5 and 6), whereasnumerous cells were seen from ventral crystal placement (sections2–7). From sections 3 and 6, a small number of backlabelled cells wereseen in the most ventral part of the DVR. Scale bar 5 1 mm.
32 P. CORDERY AND Z. MOLNAR
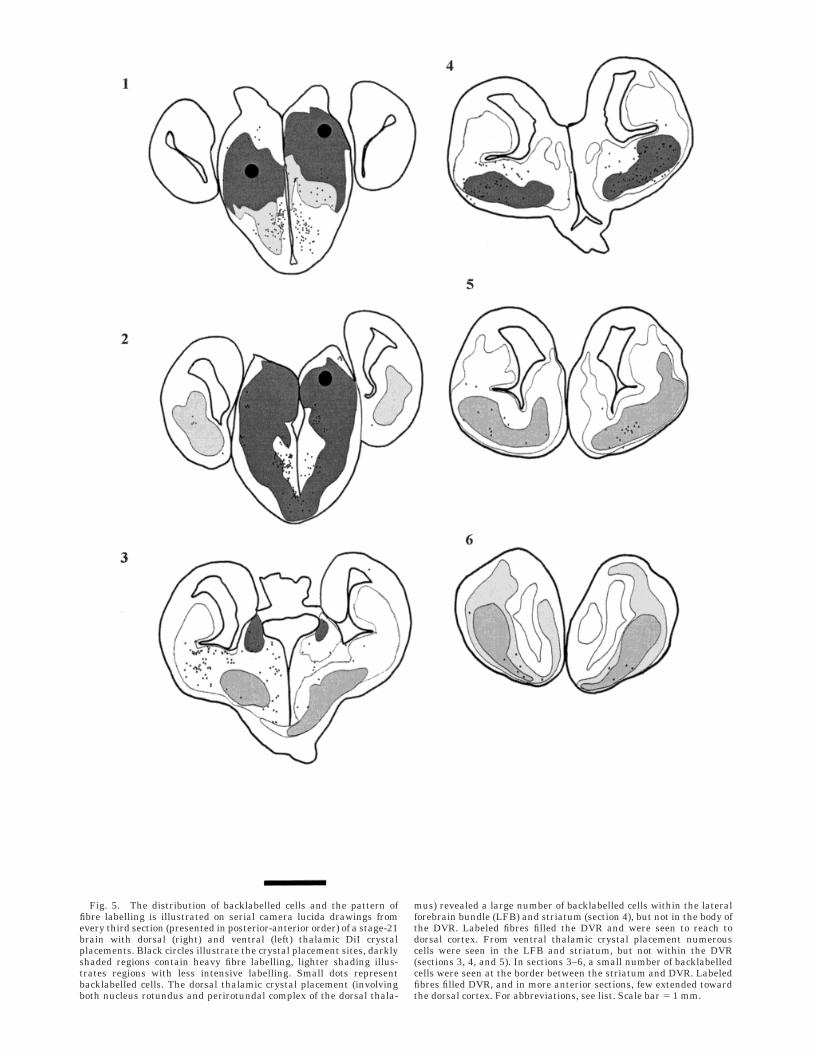
Fig. 5. The distribution of backlabelled cells and the pattern offibre labelling is illustrated on serial camera lucida drawings fromevery third section (presented in posterior-anterior order) of a stage-21brain with dorsal (right) and ventral (left) thalamic DiI crystalplacements. Black circles illustrate the crystal placement sites, darklyshaded regions contain heavy fibre labelling, lighter shading illus-trates regions with less intensive labelling. Small dots representbacklabelled cells. The dorsal thalamic crystal placement (involvingboth nucleus rotundus and perirotundal complex of the dorsal thala-
mus) revealed a large number of backlabelled cells within the lateralforebrain bundle (LFB) and striatum (section 4), but not in the body ofthe DVR. Labeled fibres filled the DVR and were seen to reach todorsal cortex. From ventral thalamic crystal placement numerouscells were seen in the LFB and striatum, but not within the DVR(sections 3, 4, and 5). In sections 3–6, a small number of backlabelledcells were seen at the border between the striatum and DVR. Labeledfibres filled DVR, and in more anterior sections, few extended towardthe dorsal cortex. For abbreviations, see list. Scale bar 5 1 mm.
thalamus, some of which could be traced toward thenucleus rotundus and some growing around this structurein the perirotundal cells, including the dorsal lateralgeniculate nucleus (dLGN). The hypothalamus containedintensive fibre labelling (Fig. 10).
Stage 24. The pattern of fibre labelling in the striatum,LFB, and DVR was similar to that seen at stage 21.Occasionally, radial glial fibres were also labelled, runningperpendicular to the trajectories of the thalamic fibres andextending from the ventricular surface toward the pialsurface of the lateral cortex.
Distribution of labelled cells from lateral forebrain
bundle crystal placements
Stage 21/22. DiI crystal placement into the LFB back-labelled a small number of cells in striatum and lateralcortex along with a small number of cells in the dorsome-dial part of the DVR. This indicates that these structuresin the telencephalon develop projections toward the crystalplacement site in the LFB and some might even continuebeyond this point to the diencephalon. From LFB crystalplacement, hypothalamic cells were also labelled, indicat-ing that they develop telencephalic projections. In the
Fig. 6. Tracing from dorsal and ventral thalamus in a stage-25turtle embryo. A: Bisbenzimide-stained coronal section through thethalamic crystal placement sites, dorsal (left) and ventral (right)viewed under ultraviolet illumination. B: The same section viewedunder DiI filter, illustrating the dorsal thalamic crystal placement,which involved both the perirotundal complex and nucleus rotundus(left) and the ventral thalamic crystal placement (right). Arrowsindicate the centre of the crystal placements at identical points in Aand B. C: Section 1 mm anterior to the dorsal crystal placementviewed under ultraviolet illumination to reveal the bisbenzimidecounterstain. D: The region indicated in E by the box. A detailed viewat high magnification of backlabelled cells (indicated with white
arrows), from the dorsal thalamic crystal placement. E: The samesection as seen in C (bottom half of photograph) viewed with a DiI filterof the dense pattern of labelled fibres and cells in the striatum. F: Thissection 500 µm anterior to C viewed under ultraviolet illumination toreveal the bisbenzimide counterstaining. The counterstaining showsthe cell patterning in dorsal cortex (DC) and the dorsal ventricularridge (DVR). G: The same region viewed under DiI filter. Numerousfibres entered the DVR in two major zones, and some of these fibresextended to the surface of the dorsal cortex and were restricted to themarginal zone. For abbreviations, see list. Scale bars 5 250 µm in A(applies to A,B); 500 µm in C (applies to C,E); 250 µm in D (applies toD,F,G).
34 P. CORDERY AND Z. MOLNAR
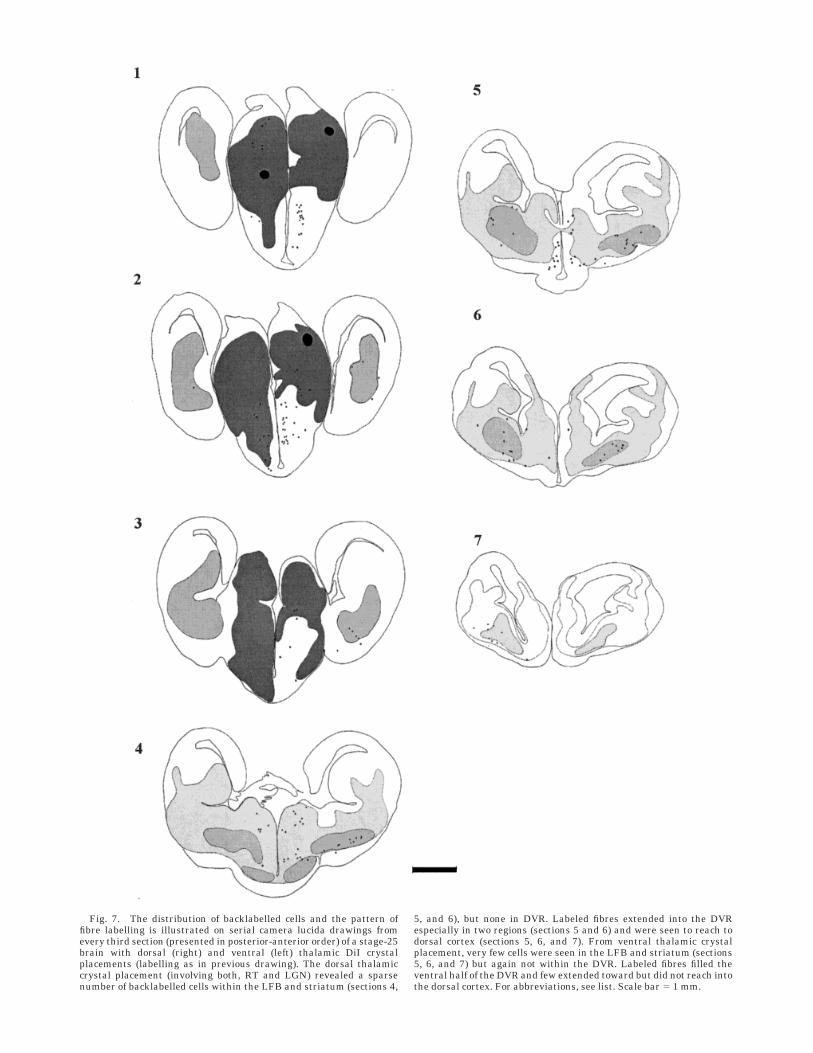
Fig. 7. The distribution of backlabelled cells and the pattern offibre labelling is illustrated on serial camera lucida drawings fromevery third section (presented in posterior-anterior order) of a stage-25brain with dorsal (right) and ventral (left) thalamic DiI crystalplacements (labelling as in previous drawing). The dorsal thalamiccrystal placement (involving both, RT and LGN) revealed a sparsenumber of backlabelled cells within the LFB and striatum (sections 4,
5, and 6), but none in DVR. Labeled fibres extended into the DVRespecially in two regions (sections 5 and 6) and were seen to reach todorsal cortex (sections 5, 6, and 7). From ventral thalamic crystalplacement, very few cells were seen in the LFB and striatum (sections5, 6, and 7) but again not within the DVR. Labeled fibres filled theventral half of the DVR and few extended toward but did not reach intothe dorsal cortex. For abbreviations, see list. Scale bar 5 1 mm.
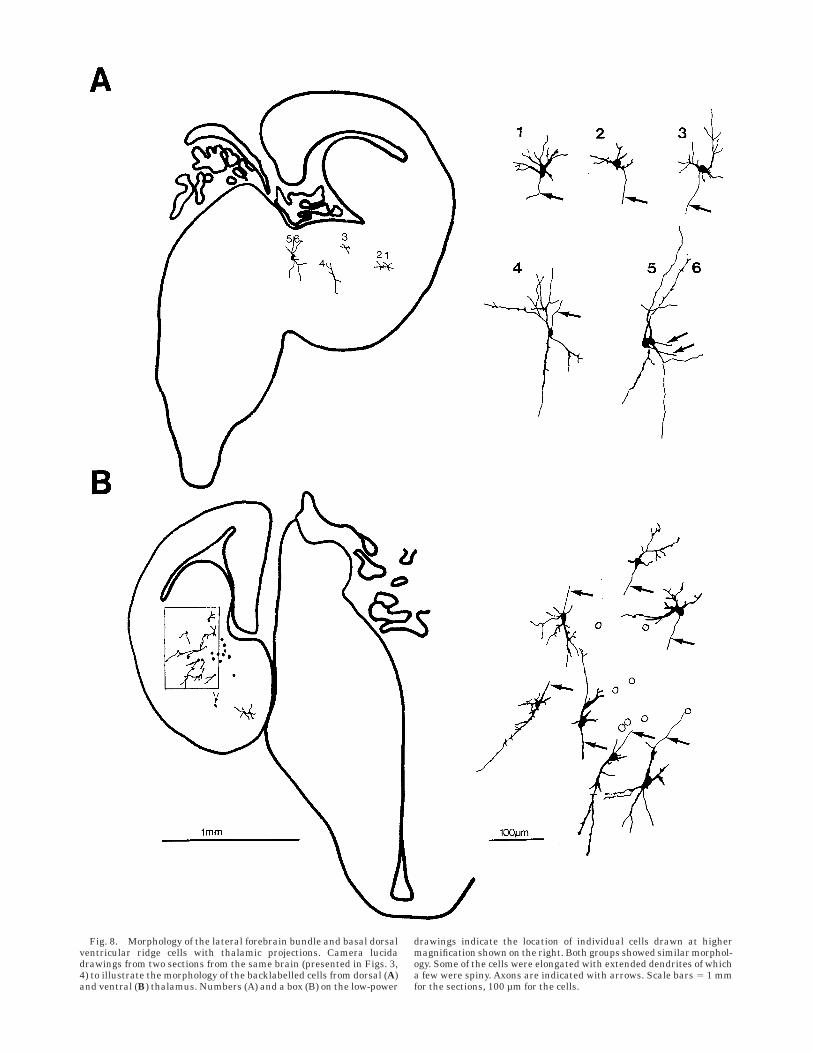
Fig. 8. Morphology of the lateral forebrain bundle and basal dorsalventricular ridge cells with thalamic projections. Camera lucidadrawings from two sections from the same brain (presented in Figs. 3,4) to illustrate the morphology of the backlabelled cells from dorsal (A)and ventral (B) thalamus. Numbers (A) and a box (B) on the low-power
drawings indicate the location of individual cells drawn at highermagnification shown on the right. Both groups showed similar morphol-ogy. Some of the cells were elongated with extended dendrites of whicha few were spiny. Axons are indicated with arrows. Scale bars 5 1 mmfor the sections, 100 µm for the cells.
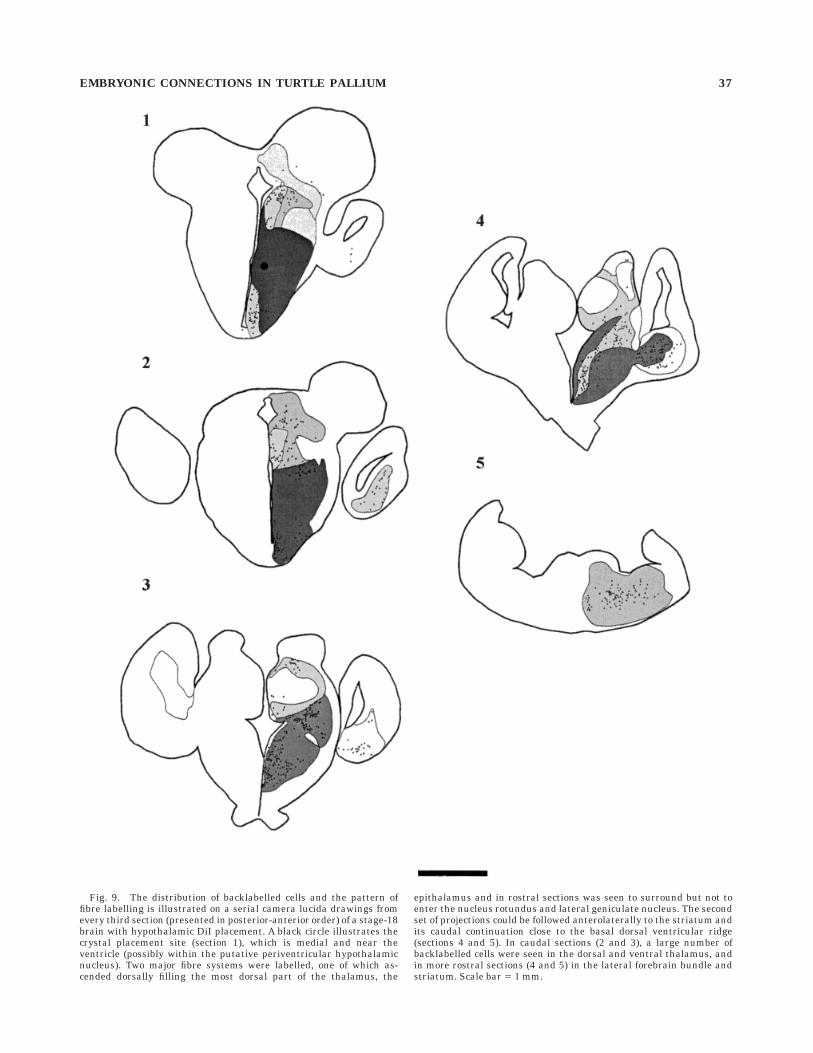
Fig. 9. The distribution of backlabelled cells and the pattern offibre labelling is illustrated on a serial camera lucida drawings fromevery third section (presented in posterior-anterior order) of a stage-18brain with hypothalamic DiI placement. A black circle illustrates thecrystal placement site (section 1), which is medial and near theventricle (possibly within the putative periventricular hypothalamicnucleus). Two major fibre systems were labelled, one of which as-cended dorsally filling the most dorsal part of the thalamus, the
epithalamus and in rostral sections was seen to surround but not toenter the nucleus rotundus and lateral geniculate nucleus. The secondset of projections could be followed anterolaterally to the striatum andits caudal continuation close to the basal dorsal ventricular ridge(sections 4 and 5). In caudal sections (2 and 3), a large number ofbacklabelled cells were seen in the dorsal and ventral thalamus, andin more rostral sections (4 and 5) in the lateral forebrain bundle andstriatum. Scale bar 5 1 mm.
EMBRYONIC CONNECTIONS IN TURTLE PALLIUM 37
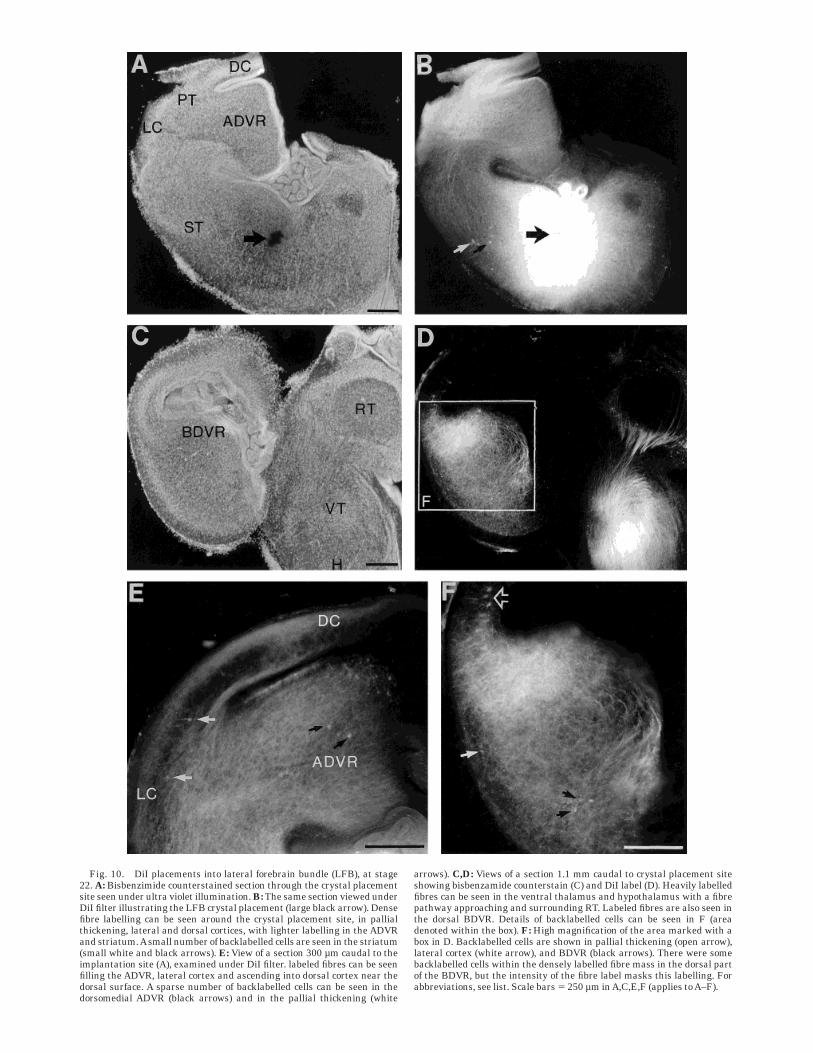
Fig. 10. DiI placements into lateral forebrain bundle (LFB), at stage22. A: Bisbenzimide counterstained section through the crystal placementsite seen under ultra violet illumination. B: The same section viewed underDiI filter illustrating the LFB crystal placement (large black arrow). Densefibre labelling can be seen around the crystal placement site, in pallialthickening, lateral and dorsal cortices, with lighter labelling in the ADVRand striatum.Asmall number of backlabelled cells are seen in the striatum(small white and black arrows). E: View of a section 300 µm caudal to theimplantation site (A), examined under DiI filter. labeled fibres can be seenfilling the ADVR, lateral cortex and ascending into dorsal cortex near thedorsal surface. A sparse number of backlabelled cells can be seen in thedorsomedial ADVR (black arrows) and in the pallial thickening (white
arrows). C,D: Views of a section 1.1 mm caudal to crystal placement siteshowing bisbenzamide counterstain (C) and DiI label (D). Heavily labelledfibres can be seen in the ventral thalamus and hypothalamus with a fibrepathway approaching and surrounding RT. Labeled fibres are also seen inthe dorsal BDVR. Details of backlabelled cells can be seen in F (areadenoted within the box). F: High magnification of the area marked with abox in D. Backlabelled cells are shown in pallial thickening (open arrow),lateral cortex (white arrow), and BDVR (black arrows). There were somebacklabelled cells within the densely labelled fibre mass in the dorsal partof the BDVR, but the intensity of the fibre label masks this labelling. Forabbreviations, see list. Scale bars 5 250 µm inA,C,E,F (applies toA–F).

thalamus, backlabelled cells were found below and aroundthe nucleus rotundus and in the perirotundal complex,including the region of the dLGN (Fig. 10).
Stage 24. Backlabelled cells continued to be seen in thestriatum and lateral forebrain bundle in sections morecaudal to the crystal placement, but interestingly, no cellswere seen backlabelled in the DVR. However, a smallnumber of cells were labelled in the lateral cortex. In theventral and dorsal thalamus, backlabelled cells were seenwithin an area of heavy fibre labelling and in addition,labelled hypothalamic cells were consistently observedventral to the thalamus. Backlabelled cells were also seenwithin and around the nucleus rotundus in a similarpattern to that at stage 21.
Labeling pattern from DiI crystal placementin ADVR
Distribution of labelled fibres from ADVR crystal
placements. The adult DVR has numerous subdivisions.We placed crystals in several regions within DVR, but atembryonic stages it was not possible to place DiI crystalsaccurately in any one specific subdivision. Some of ourlargest crystal placements involved one third of the ADVR(Fig. 11C).
Stage 21/22. A crystal placement into DVR stronglylabelled fibres within DVR and filled a large part of thenucleus. Some labelled fibres descended toward the stria-tum and some ascended toward the dorsal cortex (see Fig.11D). In the dorsal cortex, fibres could be followed to thesuperficial cortical layer beneath the pial surface. Thesesuperficial fibres extended toward the medial cortex. Inthe thalamus, very little fibre label was seen, except for abundle of fibres in the dorsal thalamus ascending towardthe RT (Fig. 11E,F).
Stage 23. The same pattern of fibre labelling could beobserved. Heavy fibre labelling was seen around theplacement site, which filled the DVR and ascended intodorsal cortex. Some labelled fibres descended into thestriatum and LFB. The labelled fibres of the anteriorcommissure could be seen additionally on more rostralsections. Sections more caudal showed fibre labelling inboth dorsal and ventral thalamus, around the RT and RE(Fig. 13, section 1 and 2).
Distribution of labelled cells from ADVR crystal
placements
Stage 21/22. DiI crystal placements in DVR backla-belled dorsal cortical cells, indicating that they developedprojections to the DVR (Figs. 11D, 12, 13). Some cells inlateral cortex and striatum were also labelled, indicatingconnections. We did not investigate the topography of DVRconnectivity with high resolution at these early stages, butwe noticed some subregional differences. The pattern andnumber of labelled cells in the above structures weredependent on the location of the DVR crystal placementsites. DiI placement into dorsomedial DVR produced ahigher number of labelled cells in lateral cortex, whereascentral and ventral DVR crystal placements revealed morebacklabelled cells in the striatum than crystal placementsinto the medial DVR. DVR crystal placements labelledcells in the nucleus rotundus and nucleus reuniens of thedorsal thalamus. There was a large number of backla-belled cells at the lower layer of the dorsal cortex, but theregion between the layers of backlabelled cells and thefibres was noticeably free of labelling (Fig. 11D).
Stage 23. DVR crystal placements continued to labelfew cells within the DVR itself, together with the processesand the endfeet of the radial glia near the ependymalsurface. Striatum and lateral forebrain bundle containedfew backlabelled cells together with numerous cells in thedorsal cortex and at the junction of the DVR and dorsalcortex. In sections more caudal, there were a large numberof backlabelled cells in the BDVR. In the thalamus,backlabelled cells were found exclusively in the nucleirotundus and reuniens. Most of our crystal placementsspanned the ventral and dorsal extent of the DVR.
Crystal placements into dorsal cortex
Distribution of labelled fibres from crystal place-
ments into dorsal cortex
Stages 19–21. Crystal placements labelled numerousfibres in the dorsal part of the DVR and two groups offibres, both travelling in thin segments near the pial andventricular surfaces of the dorsal and medial cortex anddescending toward the septum (Fig. 14B, 15). From theregion of intensive labelling, another larger group of fibresdescended toward the lateral forebrain bundle through theventral part of the DVR and the striatum.
Stage 24. Dorsal cortical crystal placement continuedto label two major fibre bundles, as in previous stages. Onebundle headed medial and travelled through the upperlayers of the medial cortex, whereas some of its compo-nents reached the midline, then descended within theseptum. The second bundle of fibres descended laterally,some passing through the lateral cortex and some enteringthe DVR. Fewer or no fibres at all were seen to be labelledbelow the border of the DVR and striatum. Fibres alsoreached the dorsal thalamus seen in sections posterior tothose with the crystal placement site. The labelled fibreshave an obvious twist as they enter the dLGN (Fig. 16D).
Distribution of labelled cells from dorsal cortical
crystal placements. Crystal placement in the dorsalcortex labelled cells within the neighbouring regions of thedorsal cortex itself, in the dorsolateral part of the DVR,and in the lateral cortex, but not within the striatum.From stages 19–20, cells of the dorsal thalamus werebacklabelled from dorsal cortical crystal placements, indi-cating that their projection had reached the crystal place-ment, confirming our description of the stage for thethalamocortical projections in turtles. The pattern andextent of thalamic labelling was dependent on the locationof the crystal placement site. In contrast with DVR, crystalplacements in dorsal cortex labelled cells exclusively in theperirotundal complex and not in nucleus rotundus (com-pare Fig. 11F with 16D).
Stage 21. Dorsal cortical crystal placements showedmost of the labelled cells in dorsal and lateral cortex alongwith cells in the dorsolateral part of the DVR. In some ofthe cases, numerous thalamic cells were backlabelled andseen exclusively in the dLGN, surrounding the nucleusrotundus, which did not contain labelling.
Stage 23. Most of the labelled cells were in dorsal andlateral cortex along with cells in the dorsolateral part ofthe DVR. DiI crystals placed into the dorsal cortex resultedin very different labelling patterns in the dorsal thalamusthan those from DVR placements. From the DVR, backla-belled thalamic cells appeared exclusively in the nucleusrotundus, whereas cells backlabelled from the dorsal cor-tex were exclusively in the perirotundal complex (Fig.16B,D). Lateral forebrain bundle crystal placements la-
EMBRYONIC CONNECTIONS IN TURTLE PALLIUM 39
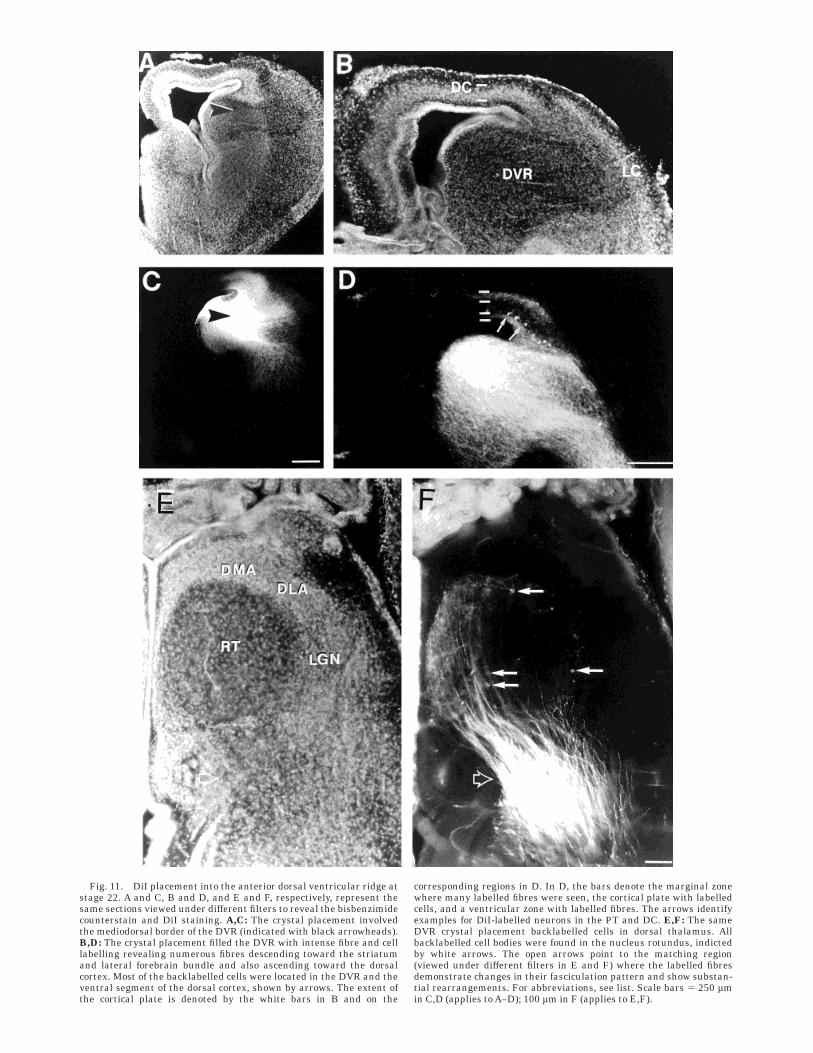
Fig. 11. DiI placement into the anterior dorsal ventricular ridge atstage 22. A and C, B and D, and E and F, respectively, represent thesame sections viewed under different filters to reveal the bisbenzimidecounterstain and DiI staining. A,C: The crystal placement involvedthe mediodorsal border of the DVR (indicated with black arrowheads).B,D: The crystal placement filled the DVR with intense fibre and celllabelling revealing numerous fibres descending toward the striatumand lateral forebrain bundle and also ascending toward the dorsalcortex. Most of the backlabelled cells were located in the DVR and theventral segment of the dorsal cortex, shown by arrows. The extent ofthe cortical plate is denoted by the white bars in B and on the
corresponding regions in D. In D, the bars denote the marginal zonewhere many labelled fibres were seen, the cortical plate with labelledcells, and a ventricular zone with labelled fibres. The arrows identifyexamples for DiI-labelled neurons in the PT and DC. E,F: The sameDVR crystal placement backlabelled cells in dorsal thalamus. Allbacklabelled cell bodies were found in the nucleus rotundus, indictedby white arrows. The open arrows point to the matching region(viewed under different filters in E and F) where the labelled fibresdemonstrate changes in their fasciculation pattern and show substan-tial rearrangements. For abbreviations, see list. Scale bars 5 250 µmin C,D (applies to A–D); 100 µm in F (applies to E,F).
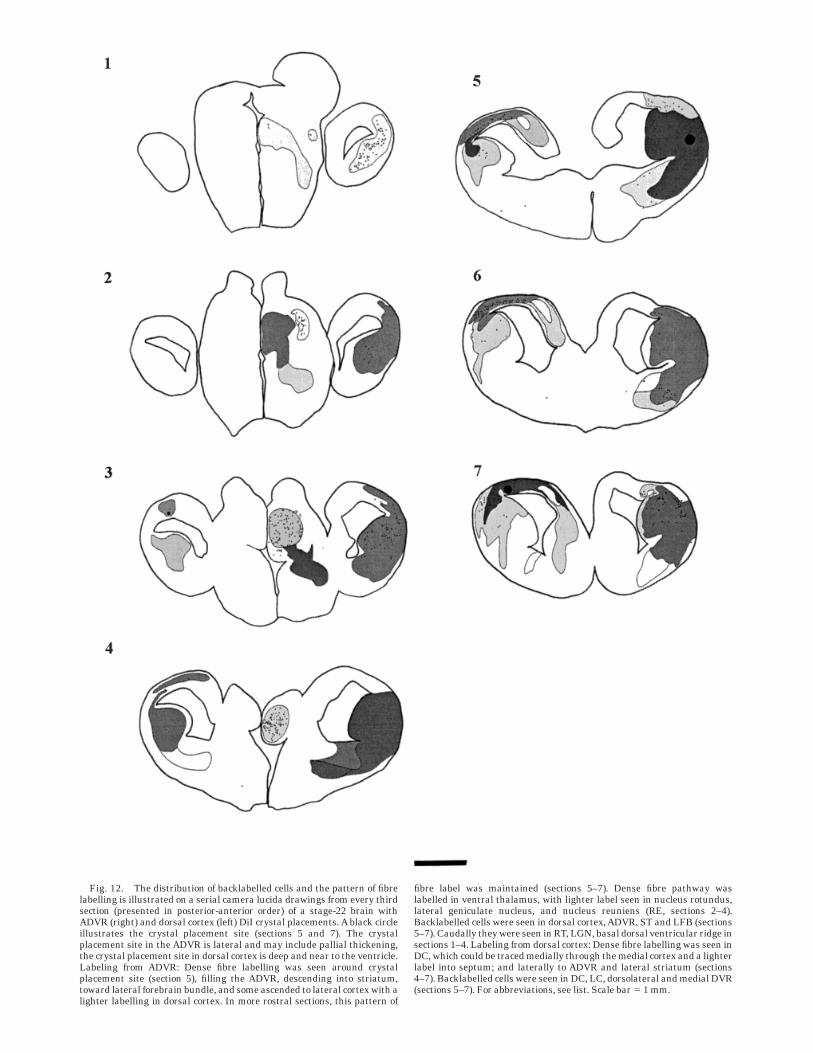
Fig. 12. The distribution of backlabelled cells and the pattern of fibrelabelling is illustrated on a serial camera lucida drawings from every thirdsection (presented in posterior-anterior order) of a stage-22 brain withADVR (right) and dorsal cortex (left) DiI crystal placements. A black circleillustrates the crystal placement site (sections 5 and 7). The crystalplacement site in the ADVR is lateral and may include pallial thickening,the crystal placement site in dorsal cortex is deep and near to the ventricle.Labeling from ADVR: Dense fibre labelling was seen around crystalplacement site (section 5), filling the ADVR, descending into striatum,toward lateral forebrain bundle, and some ascended to lateral cortex with alighter labelling in dorsal cortex. In more rostral sections, this pattern of
fibre label was maintained (sections 5–7). Dense fibre pathway waslabelled in ventral thalamus, with lighter label seen in nucleus rotundus,lateral geniculate nucleus, and nucleus reuniens (RE, sections 2–4).Backlabelled cells were seen in dorsal cortex,ADVR, ST and LFB (sections5–7). Caudally they were seen in RT, LGN, basal dorsal ventricular ridge insections 1–4. Labeling from dorsal cortex: Dense fibre labelling was seen inDC, which could be traced medially through the medial cortex and a lighterlabel into septum; and laterally to ADVR and lateral striatum (sections4–7). Backlabelled cells were seen in DC, LC, dorsolateral and medial DVR(sections 5–7). For abbreviations, see list. Scale bar 5 1 mm.
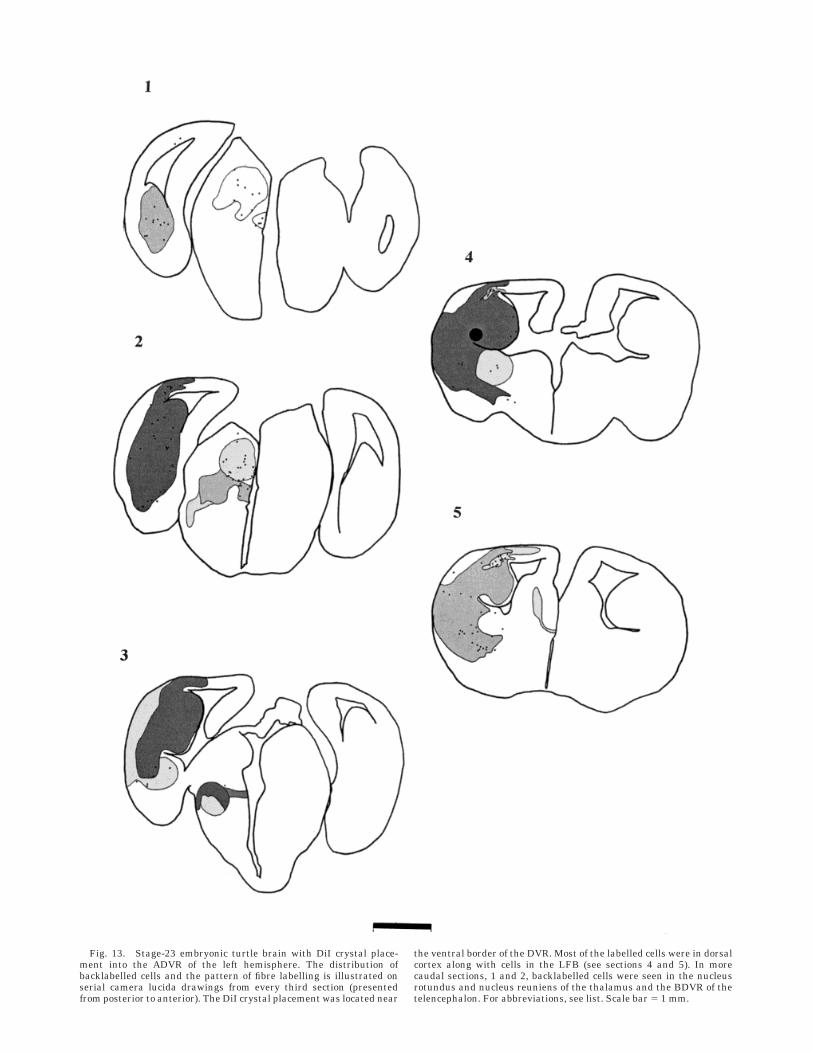
Fig. 13. Stage-23 embryonic turtle brain with DiI crystal place-ment into the ADVR of the left hemisphere. The distribution ofbacklabelled cells and the pattern of fibre labelling is illustrated onserial camera lucida drawings from every third section (presentedfrom posterior to anterior). The DiI crystal placement was located near
the ventral border of the DVR. Most of the labelled cells were in dorsalcortex along with cells in the LFB (see sections 4 and 5). In morecaudal sections, 1 and 2, backlabelled cells were seen in the nucleusrotundus and nucleus reuniens of the thalamus and the BDVR of thetelencephalon. For abbreviations, see list. Scale bar 5 1 mm.
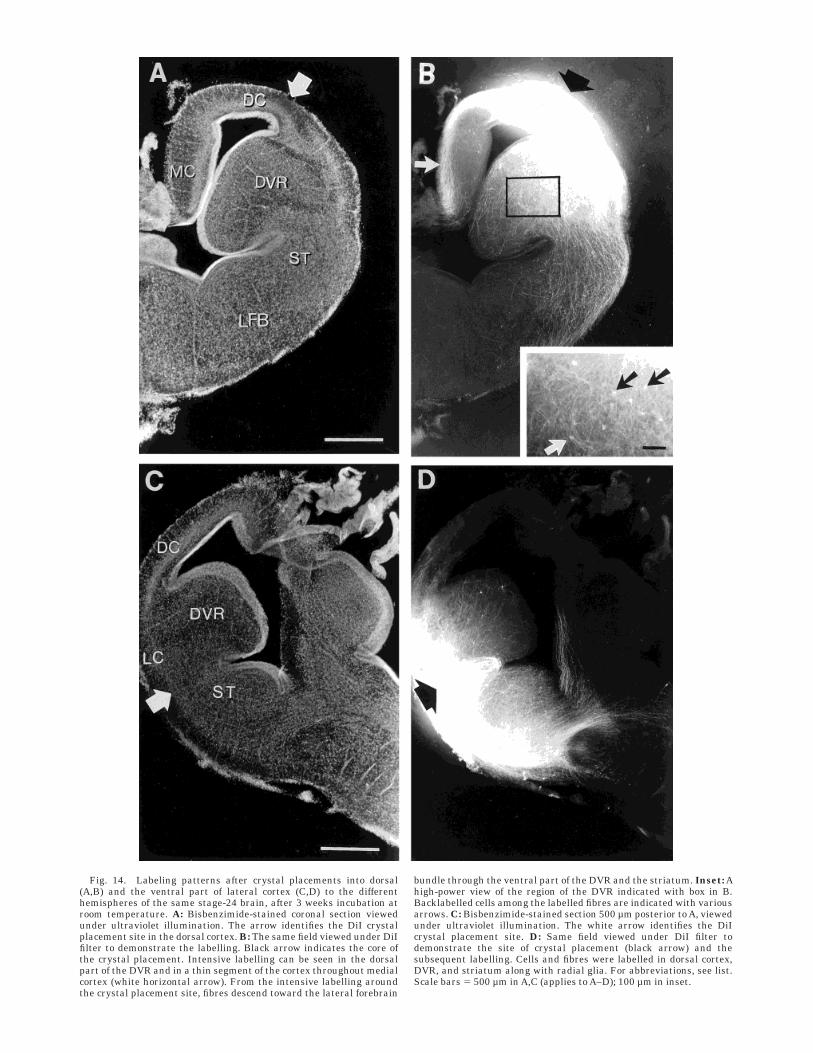
Fig. 14. Labeling patterns after crystal placements into dorsal(A,B) and the ventral part of lateral cortex (C,D) to the differenthemispheres of the same stage-24 brain, after 3 weeks incubation atroom temperature. A: Bisbenzimide-stained coronal section viewedunder ultraviolet illumination. The arrow identifies the DiI crystalplacement site in the dorsal cortex. B: The same field viewed under DiIfilter to demonstrate the labelling. Black arrow indicates the core ofthe crystal placement. Intensive labelling can be seen in the dorsalpart of the DVR and in a thin segment of the cortex throughout medialcortex (white horizontal arrow). From the intensive labelling aroundthe crystal placement site, fibres descend toward the lateral forebrain
bundle through the ventral part of the DVR and the striatum. Inset: Ahigh-power view of the region of the DVR indicated with box in B.Backlabelled cells among the labelled fibres are indicated with variousarrows. C: Bisbenzimide-stained section 500 µm posterior to A, viewedunder ultraviolet illumination. The white arrow identifies the DiIcrystal placement site. D: Same field viewed under DiI filter todemonstrate the site of crystal placement (black arrow) and thesubsequent labelling. Cells and fibres were labelled in dorsal cortex,DVR, and striatum along with radial glia. For abbreviations, see list.Scale bars 5 500 µm in A,C (applies to A–D); 100 µm in inset.

belled both groups of thalamic cells, indicating that bothsets of fibres join a similar section of the lateral forebrainbundle as they ascend toward their specific targets.
Stage 24. Crystal placement in the dorsal cortex re-vealed small numbers of backlabelled cells in the DVR (seeFig. 14B and inset) and in the dorsal geniculate (orperirotundal complex) of the dorsal thalamus; however,none were seen in any other region in the telencephalon(i.e., in lateral cortex or striatum).
Crystal placements into lateral cortex
Crystal placements in the lateral cortex labelled cells inthe dorsal cortex and lateral part of the striatum (Figs. 14,15).
Distribution of fibres labelled from DiI crystal place-
ments into lateral cortex
Stage 21. Lateral cortical crystal placement revealedintense labelling of fibres in the upper layer of dorsalcortex, together with descending fibres filling the striatumand the lateral part of the DVR (Figs. 14C,D, 15). Therewas intense labelling of radial glia fibres around theboundary of the DVR and striatum corresponding to thecentre of the crystal placement in the lateral cortex (Fig.14D).
Stage 24. Lateral cortical crystal placement revealed asimilar pattern of fibre labelling as seen at stage 21.Intense labelling of radial glia fibres occurred in a tractacross the ventral DVR to the ependymal surface. Labeledfibres were also seen in the dorsal cortex and in medialcortex from the lateral to medial border.
Distribution of labelled cells from lateral cortex.
Lateral cortical crystal placement revealed a small num-ber of backlabelled cells in the dorsomedial part of DVR,striatum, LFB, and radial glial cells at the border betweenthe striatum and DVR. Labeled cells were also seen in thedorsal cortex (Fig. 15, sections 4, 5, and 6). Many radialglia cells were also labelled from fibres extending acrossthe DVR with cell bodies on the ependymal surface. Fromlateral cortical crystal placements no thalamic cells werelabelled at stage 21.
DISCUSSION
In previous studies, we have investigated the possibledevelopmental role of largely transient cell populations inthe mammalian cortical subplate, marginal zone, internalcapsule, and thalamic reticular nucleus (Molnar and Blake-more, 1995; Molnar and Cordery, 1999). The aim of thisstudy is to investigate the similarities and differences inthalamocortical development in reptiles and mammalsand explore the possibility that some of the transient cellpopulations in the embryonic mammalian preplate andinternal capsule have reptilian homologues. Our resultsare summarised in Figure 17 and Table 2. We discuss herepossible homologous developmental algorithms betweenmammals and reptiles according to regions (thalamus,internal capsule, DVR, and lateral and dorsal cortex) andsuggest that some of the largely transient cells in thevertebrate pallium are part of a common evolutionaryblueprint.
Embryonic development of thalamocorticalprojections in turtle
Our study gives the first account of the timing andpattern of thalamocortical fibre innervation in embryonic
turtle. Fibres labelled from dorsal thalamus extend throughthe lateral forebrain bundle and striatum to reach theirtargets in dorsal cortex and DVR from stage 19–20. Thelabelled fibres extend along very specific paths to theirmajor targets; the ventral and dorsal part of the DVR andthe molecular layer of the dorsal cortex. It is interesting tonote that some regions like the central area of DVR, lateralcortex, or the lower cellular and subcellular layers ofdorsal cortex are avoided by thalamic fibres from earlystages of development, suggesting early specificity intarget selection. It was difficult to label selectively thedistinct projections from the different nuclei of the dorsalthalamus. Some of our crystal placements involved bothnucleus rotundus and the perirotundal complex and la-belled fibres both to DVR and dorsal cortex. Nevertheless,our backlabelling experiments from dorsal cortex and fromDVR showed that dorsal cortex selectively receives tha-lamic input from the perirotundal complex, whereas DVRselectively receives its input from nucleus rotundus. Thisis comparable with connections in adult turtles (Hall andEbner, 1970), with one known exception; the nuclei abovethe rotundus project to DVR as part of a nonspecificprojection system (Parent, 1976; Balaban and Ulinski,1981a,b). This distinct pattern of innervation was appar-ent from stage 20 and was unequivocal from the earlieststage at which thalamic fibres could be backlabelled. Theclear segregation of rotundal and perirotundal projectionsto DVR and to dorsal cortex must result from specific earlydevelopment. It is interesting to note that retinal labellingreaches the dorsal thalamus at stage 20 and that fibresalmost exclusively invade the LGN, but not the nucleusrotundus (Molnar and Sernagor, 1998). From stage 17, DiIplacements onto the colliculus labelled dense fibre net-works filling the nucleus rotundus (Molnar and Sernagor,1998), among other labelled fibres. These anatomicallydistinct embryonic fibre pathways might carry the prin-ciple adult organization. Because retinal projections reachthe dorsal thalamus and colliculus after stage 19 (Molnarand Sernagor, 1998) and it is not until stage 22 when theretinal ganglion cells develop their firing activity (Ser-nagor and Grzywacz, 1996), the early target finding ofthalamocortical connectivity in turtle (similarly to mam-mals) cannot possibly rely heavily on the early activitypatterns arriving from the peripheral sensory organs. Thisprocess most probably involves intrinsic guidance mecha-nisms, possibly including guide-post cells in the lateralforebrain bundle and striatum, similar to transient inter-nal capsule cell populations in mammals. The presence ofthese transient cells was described in several mammalianspecies including rodents, carnivores, marsupials, andhumans (Mitrofanis, 1992; Molnar et al., 1998a,b; Leviticand Kostovic, 1996). Our present study extends the inves-tigation to reptiles and suggests a possible homologous cellgroup in embryonic turtle.
Comparisons between reptilian andmammalian thalami during development
The homologies between reptilian and mammalian fore-brains are less controversial within the diencephalon, thanhomologies within the telencephalon. Hodologic and topo-logic evidence suggests that homologous dorsal thalamicnuclei are present in reptiles, birds, and mammals for therelay of visual, auditory, and somatosensory information tothe pallium and receive input from the thalamic reticularnucleus (see Karten, 1969; Shimizu and Karten, 1991;
44 P. CORDERY AND Z. MOLNAR
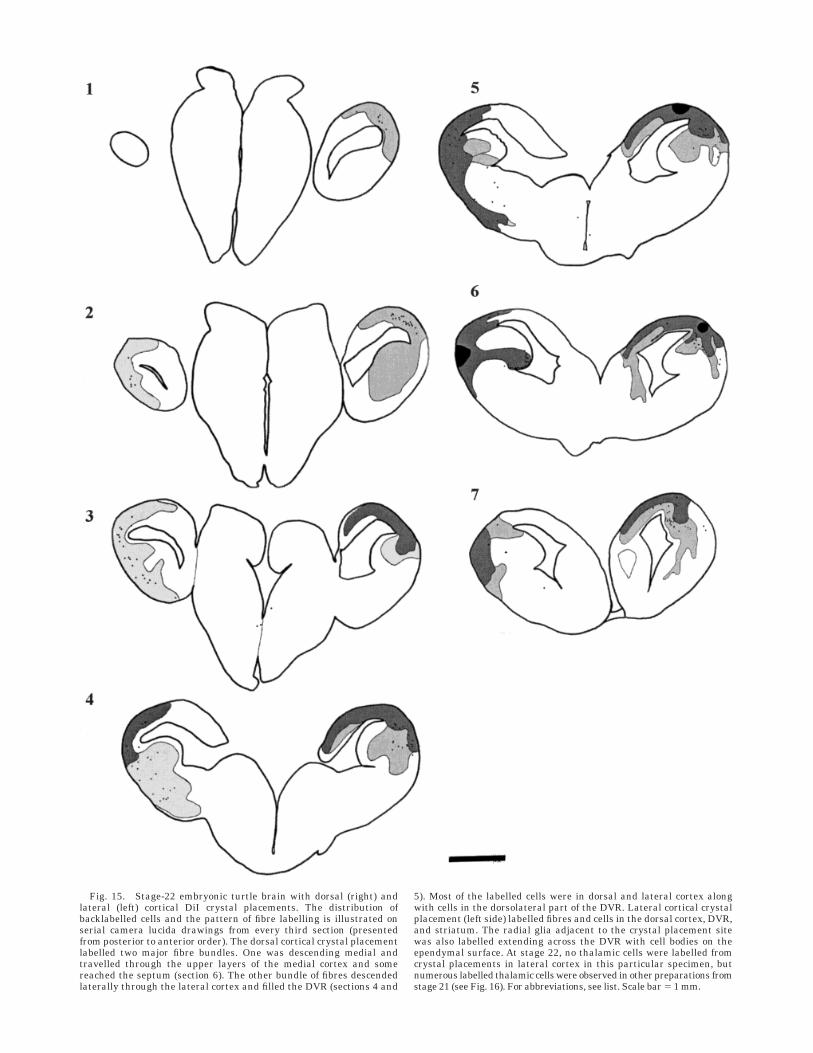
Fig. 15. Stage-22 embryonic turtle brain with dorsal (right) andlateral (left) cortical DiI crystal placements. The distribution ofbacklabelled cells and the pattern of fibre labelling is illustrated onserial camera lucida drawings from every third section (presentedfrom posterior to anterior order). The dorsal cortical crystal placementlabelled two major fibre bundles. One was descending medial andtravelled through the upper layers of the medial cortex and somereached the septum (section 6). The other bundle of fibres descendedlaterally through the lateral cortex and filled the DVR (sections 4 and
5). Most of the labelled cells were in dorsal and lateral cortex alongwith cells in the dorsolateral part of the DVR. Lateral cortical crystalplacement (left side) labelled fibres and cells in the dorsal cortex, DVR,and striatum. The radial glia adjacent to the crystal placement sitewas also labelled extending across the DVR with cell bodies on theependymal surface. At stage 22, no thalamic cells were labelled fromcrystal placements in lateral cortex in this particular specimen, butnumerous labelled thalamic cells were observed in other preparations fromstage 21 (see Fig. 16). For abbreviations, see list. Scale bar 5 1 mm.
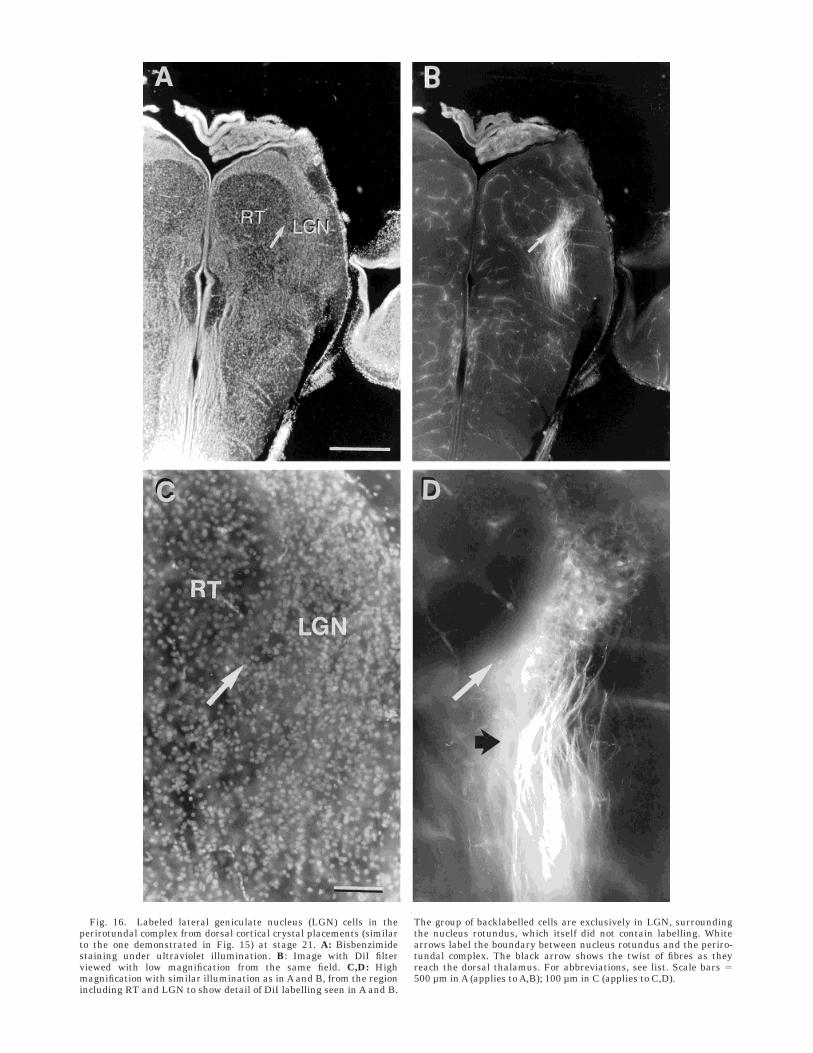
Fig. 16. Labeled lateral geniculate nucleus (LGN) cells in theperirotundal complex from dorsal cortical crystal placements (similarto the one demonstrated in Fig. 15) at stage 21. A: Bisbenzimidestaining under ultraviolet illumination. B: Image with DiI filterviewed with low magnification from the same field. C,D: Highmagnification with similar illumination as in A and B, from the regionincluding RT and LGN to show detail of DiI labelling seen in A and B.
The group of backlabelled cells are exclusively in LGN, surroundingthe nucleus rotundus, which itself did not contain labelling. Whitearrows label the boundary between nucleus rotundus and the periro-tundal complex. The black arrow shows the twist of fibres as theyreach the dorsal thalamus. For abbreviations, see list. Scale bars 5500 µm in A (applies to A,B); 100 µm in C (applies to C,D).

Butler, 1994a, 1995; Pritz, 1995; Adams et al., 1997).Differences between the reptilian and mammalian thalamiare the lack of reciprocal telencephalic efferents and less orabsent interneurons (Pritz, 1995). Some theories of DVRevolution require sensory-relay dorsal thalamic nuclei tobe homologous to mammalian intralaminar nuclei (Dıaz etal., 1990; Bruce and Neary, 1995; Striedter, 1997). In bothmammals and reptiles, neurons surround the ventrolat-eral surface of the dorsal thalamus, some express immuno-reactivity for glutamic acid decarboxylase, parvalbumin,calretinin and possess projections to the dorsal thalamic
nuclei (Pritz, 1995). We observed that the projection todorsal thalamus develop during early embryonic develop-ment, most probably from the putative thalamic reticularnucleus in both mammals (Mitrofanis and Baker, 1993;Molnar et al., 1998a,b; Molnar and Cordery, 1999) andreptiles (present study). Because the thalamus in mam-mals and reptiles has numerous similarities from embry-onic stages and the comparative organisation is relativelywell understood (Butler, 1994a; Pritz, 1995), the thalamicconnections are very useful as reference points in searchfor possible homologous developmental processes.
Cells in the reptilian lateral forebrain bundleand striatum with early thalamic connections
Our study in the embryonic turtle describes backla-belled cells in the lateral forebrain bundle and the stria-tum after both dorsal and ventral thalamic carbocyanidedye placements from the earliest stage studied (stage 17).This result is similar to the one we and others observed invarious mammalian species during embryonic develop-ment (see Molnar and Cordery, 1999). The data areconsistent with the possibility that these cells are homolo-gous to the ones observed with thalamic projections in theembryonic internal capsule in hamster (Metin and Gode-ment, 1996), rat (Molnar et al., 1998a; Molnar and Cord-ery, 1999), and the marsupial Monodelphis domestica(Molnar et al., 1998b). Our suggestion, that the cells of themammalian embryonic internal capsule are homologous tothe cells of the lateral forebrain bundle and striatum inturtle is based on connectional analysis (Molnar andCordery, 1999; and present study, Fig. 17). This suggestionis supported by the similarity of the morphology of thebacklabelled cells and matching immunoreactivity forcalretinin and neuropeptide Y (NPY) in the correspondingregions of embryonic rat and embryonic turtle brains(Cordery et al., 1997). Both, in embryonic rat and turtle,neurons with early thalamic projections have fusiformmorphology with dendrites extending perpendicular to thetrajectories of the thalamocortical fibres (Ramcharan andGuillery, 1997; Molnar and Cordery, 1999; and presentstudy). Immunostaining of a stage-24 embryonic turtlebrain demonstrated calretinin- and NPY-positive cells inthe lateral forebrain bundle and striatum, whereas inembryonic rat internal capsule similar immunostainingreveals matching cells (Cordery et al., 1997). In mammals,these cells are thought to be transient and only presentduring development; shortly after birth, they are believedto undergo preferential cell death and only a fraction ofthem survive to adulthood (Mitrofanis and Guillery, 1993;Earle and Mitrofanis, 1996). The fate of the internal
Fig. 17. Schematic summary of our tracing study in embryonicturtle brains. A: Tracing from dorsal and ventral thalami. Dorsal andventral thalamic DiI crystal placements (large dots) revealed similarlabelling patterns (small dots represent backlabelled cells) in thehypothalamus, lateral forebrain bundle, and striatum. At early stages(stage 20), ventral crystal placements labelled a small number of cellsclose to the most ventral part of the DVR, but we observed no labelledcells in the DVR. Dorsal thalamic crystal placements revealed overallfewer labelled cells, but similarly, no cells in DVR at any stage. Ingeneral, from both dorsal and ventral thalamic crystal placements, wesaw more labelled cells in early stages particularly in the striatum.B: Tracing from lateral forebrain bundle and DVR. Lateral forebrainbundle placements (left) labelled a small number of cells in the dorsalDVR, cells in lateral cortex, hypothalamus, and dorsal thalamus (thislabelling included nucleus rotundus and perirotundal complex). DVRcrystal placement (right) labelled cells in dorsal and lateral cortex,BDVR, together with backlabelled cells in the nucleus rotundus of thethalamus. C: Tracing from dorsal and lateral cortex. Crystal place-ment in the dorsal cortex (left), labelled cells within the neighbouringregions of the dorsal cortex itself, in the dorsolateral part of DVR andin the lateral cortex but not in striatum. In caudal sections, somebacklabelled cells were seen in the ventral pallium, in BDVR (which issuggested to correspond to amygdala). From stage 21, thalamic cellswere backlabelled; however, this labelling was dependent on thecrystal placement site. In contrast with DVR crystal placements whichlabelled cells in the nucleus rotundus, dorsal cortical crystal place-ments labelled cells in a perirotundal complex and not in nucleusrotundus. Crystal placements in the lateral cortex (right) labelled cellsin the dorsal cortex, BDVR (or amygdala), and occasionally in thelateral part of the striatum. D: Tracing from hypothalamus andtermination of fibre pathways from the retina and the olfactory bulb.Numerous cells were seen in the hypothalamus, ventral thalamus,upper border of the nucleus rotundus, and epithalamus from hypotha-lamic DiI crystal placements, but no cells were backlabelled in thenucleus rotundus or the perirotundal complex. No cells were found inthe anterior dorsal ventricular ridge but there were many in the areaof the lateral forebrain bundle, striatum, and in BDVR. Olfactoryprojections target a distinct region of lateral cortex (lightly shadedareas) and retinal projections terminate in the perirotundal complex(black areas). For abbreviations, see list.
TABLE 2. Timing of Main Events in Forebrain Developmentin Embryonic Turtle1
Stage 17–19 Projections of lateral forebrain bundle and striatal cells reachdorsal thalamus
Stage 17–20 Rotundal projections reach DVRStage 17–20 Perirotundal projections reach dorsal cortexStage 19 Projections of DVR cells reach to dorsal cortexStage 19 Dorsal cortex projections reach to DVRStage 17 Tectal projections reach to nucleus rotundus2
Stage 19 Olfactory projections reach lateral cortex3
Stage 19–20 Retinal projections reach LGN of the perirotundal complex2
Stage 19–20 Retinal projections reach tectum2
Stage 22 Retinal ganglion cells begin spontaneous activity4
1For abbreviations, see list.2Molnar and Sernagor, 1998.3Cordery et al., 1998.4Sernagor and Grzywacz, 1996.
EMBRYONIC CONNECTIONS IN TURTLE PALLIUM 47

capsule cells with thalamic projections in reptiles is notdetermined. There are numerous immunohistochemicalstudies on adult turtle pallidum and striatum (Medina andReiner, 1995), but none during embryonic development. Inhamster Metin and Godement, (1996) described that fromE13.5, early projections from the ganglionic eminence tothe thalamus and cortex decrease. In rodents, we alsonoticed (Molnar and Cordery, 1999) that there are moreinternal capsule cells labelled at E14 than at E16 fromsimilar dorsal thalamic crystal placements. We noticed asimilar decline in turtle, with less lateral forebrain bundlecells labelled at later stages. This might reflect the simi-larly transient nature of these connections in both turtlesand mammals; nevertheless, we feel that additional stud-ies are required to establish the transient nature of thesecells.
There are several unknown factors that make it difficultto draw firm conclusions from our solely connectional dataand, thus, propose homologous relationships between theinternal capsule cells in mammals and lateral forebrainbundle and striatum in reptiles with dorsal thalamicprojections: (1) lack of quantification of cell death in thedifferent cell groups within and around internal capsule indeveloping mammals and reptiles, (2) no clear cytoarchitec-tonic borders present in the close neighbourhood of thedifferent cell groups of the embryonic internal capsule, (3)lack of detailed studies that used selective neurochemicalmarkers for the detection of similar cell groups on embry-onic turtle, comparable for embryonic and early postnatalmammals (Mitrofanis, 1992).
(1) Although, there is a general consensus that cells inthe developing mammalian internal capsule are moreprominent during development than in the adult (Liu andGraybiel, 1992; Mitrofanis, 1992; Metin and Godement,1996; Earle and Mitrofanis, 1996), their transient naturehas never been rigorously quantified even in mammals.Without further studies that use birth dating and celldeath markers in mammals and reptiles, it is not possibleto draw conclusions on their transient nature. It is possiblethat some types of internal capsule cells in mammals or inreptiles do not die.
(2) The thalamic reticular nucleus is continuous with acell group in the internal capsule, which Mitrofanis (1992)named the perireticular nucleus. Perireticular cells wereidentified by their small soma size and their markers,which are similar to the ones expressed by the thalamicreticular nucleus (g-aminobutyric acid, somatostatin, parv-albumin, calbindin, calretinin, CAT-301, pro-a-thyrotropin–releasing hormone; Mitrofanis, 1992; Crabtree and Kind,1993; Amadeo et al., 1998). Mitrofanis (1992) describedthat these cells have thalamic origin and are different fromthe adjacent globus pallidus or striatum, although duringearly development perireticular cells (identified by immu-nohistochemistry for pro-a-thyrotropin–releasing hor-mone) are scattered throughout the internal capsule andover other structures, including the globus pallidus, whichis known to derive from the medial ganglionic eminence.Cells of nucleus basalis (a nucleus that has telencephalicorigin) are known to project to the cortex in adult (Saper,1984). The lack of clear cytoachitectonic boundaries inembryonic brains makes it very difficult to use morespecific terms than ‘‘internal capsule’’ (e.g., Foster, 1998)while relying only on connectional data. Adams and Baker(1995) described that perireticular neurons in the ratbegin to develop projections to the cortex by E17, but based
on the combination of a similar DiI tracing study withacetylcholinesterase immunohistochemistry De Carlos etal. (1995) proposed that this early input originates fromcells of the substantia innominata and nucleus basalis ofMeynert (Dori and Parnavelas, 1989). Our study in the rat(Molnar and Cordery, 1999) also confirmed the observationthat it is not until E17 that the first internal capsule cellsreach dorsal cortex. In the same study, we demonstratedthat at E14, a DiI crystal placement into the perirhinalcortex consistently labelled a small number of cells amongthe fibres entering the internal capsule, whereas moredorsal crystal placement sites, not including the perirhinalcortex, did not. Additional and independent criteria isrequired for identifying these cell groups during earlystages in development in reptiles and mammals to resolvethese issues.
(3) There are numerous immunohistochemical studieson adult turtle pallidum (Reiner et al., 1980; Brauth et al.,1983; Medina and Smeets, 1991; Reiner, 1992; Marın et al.,1998), but very few during embryonic development (Cord-ery et al., 1997). In adult turtles and mammals, it is knownthat the globus pallidus is similarly organised and projectsto ventral anterior and ventral lateral thalamic nuclei(Saper, 1984; Medina and Reiner, 1995). Nevertheless, ourlocalised dorsal thalamic carbocyanine dye placements(limited to perirotundal complex and nucleus rotundus)revealed numerous cells scattered in the lateral forebrainbundle in embryonic turtle, similar to that seen from moreventral thalamic crystal placements. Although we report aslight decline in the number of labelled cells from stage 17to 25, it is not known whether the cells with dorsalthalamic projections at embryonic stages are transient. Itis conceivable that these are true homologues of ratperireticular cells in embryonic turtles and have thalamicorigin but are not transient. Thus, some turtle internalcapsule cells, similar to that of rat, may be pallidal;alternatively, those cells in the rat, could be truly transientand nonpallidal, whereas those in turtles may be pallidal.The basal ganglia, however, has been shown to have verysimilar organization in vertebrates (Medina and Reiner,1995; Marın et al., 1998). We used the term ‘‘neurons of the(primitive or embryonic) internal capsule’’ in mammalsand ‘‘lateral forebrain bundle and striatum’’ in turtle, but itis possible that future research combining connectionalanalysis and neurochemical markers will define subgroupsof these cells and a more specific nomenclature will bepossible.
Comparison of pallial organisationin adult reptiles and mammals, possible
corresponding boundaries in the developingreptilian and mammalian pallia
On the basis of our hodologic analysis in rat embryos, wesuggested that there is a boundary between the lateraledge of the striatum and cortical intermediate zone (Mol-nar and Cordery, 1999; Fig. 18). Dorsal and ventralthalamic crystal placements, from embryonic day (E) 14days of gestation, revealed labelled neurons in the internalcapsule below both medial and lateral ganglionic emi-nences, but not beyond the junction between the striatumand cortical intermediate zone. This boundary is interest-ing in mammalian thalamocortical development in severalrespects (see discussion of Molnar and Cordery, 1999). Inturtle embryos, there appeared to be a similar boundary(defined by our present connectional analysis) extending
48 P. CORDERY AND Z. MOLNAR
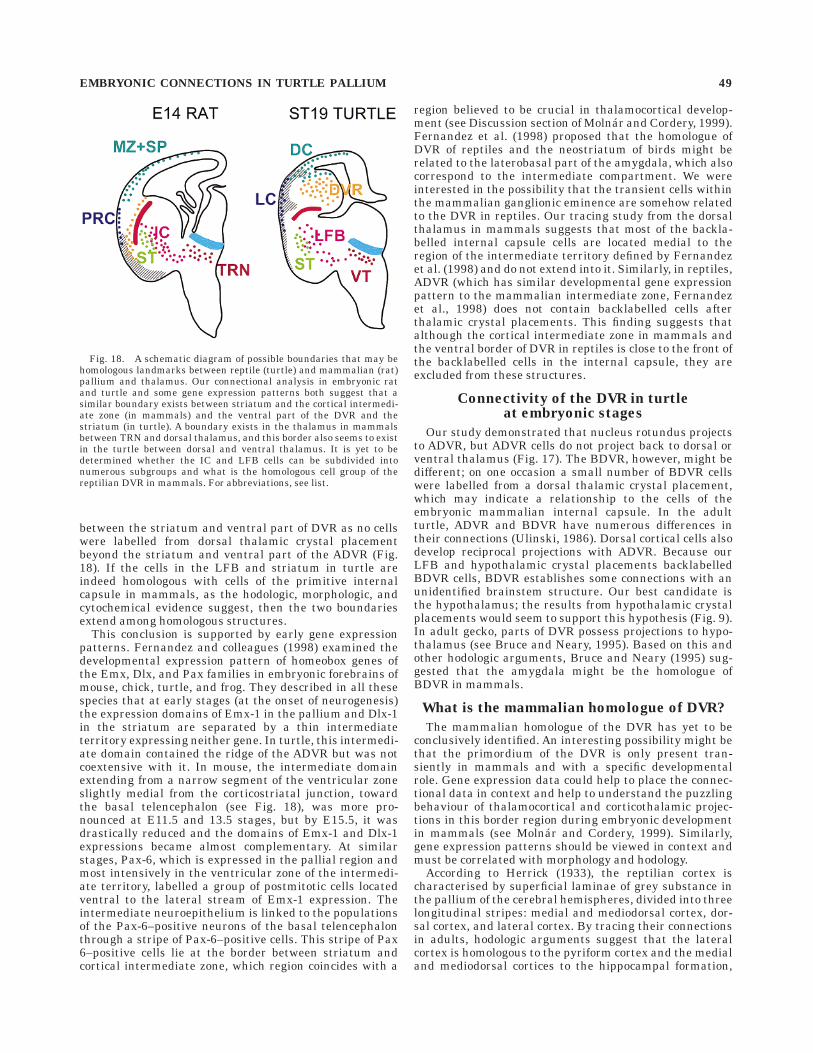
between the striatum and ventral part of DVR as no cellswere labelled from dorsal thalamic crystal placementbeyond the striatum and ventral part of the ADVR (Fig.18). If the cells in the LFB and striatum in turtle areindeed homologous with cells of the primitive internalcapsule in mammals, as the hodologic, morphologic, andcytochemical evidence suggest, then the two boundariesextend among homologous structures.
This conclusion is supported by early gene expressionpatterns. Fernandez and colleagues (1998) examined thedevelopmental expression pattern of homeobox genes ofthe Emx, Dlx, and Pax families in embryonic forebrains ofmouse, chick, turtle, and frog. They described in all thesespecies that at early stages (at the onset of neurogenesis)the expression domains of Emx-1 in the pallium and Dlx-1in the striatum are separated by a thin intermediateterritory expressing neither gene. In turtle, this intermedi-ate domain contained the ridge of the ADVR but was notcoextensive with it. In mouse, the intermediate domainextending from a narrow segment of the ventricular zoneslightly medial from the corticostriatal junction, towardthe basal telencephalon (see Fig. 18), was more pro-nounced at E11.5 and 13.5 stages, but by E15.5, it wasdrastically reduced and the domains of Emx-1 and Dlx-1expressions became almost complementary. At similarstages, Pax-6, which is expressed in the pallial region andmost intensively in the ventricular zone of the intermedi-ate territory, labelled a group of postmitotic cells locatedventral to the lateral stream of Emx-1 expression. Theintermediate neuroepithelium is linked to the populationsof the Pax-6–positive neurons of the basal telencephalonthrough a stripe of Pax-6–positive cells. This stripe of Pax6–positive cells lie at the border between striatum andcortical intermediate zone, which region coincides with a
region believed to be crucial in thalamocortical develop-ment (see Discussion section of Molnar and Cordery, 1999).Fernandez et al. (1998) proposed that the homologue ofDVR of reptiles and the neostriatum of birds might berelated to the laterobasal part of the amygdala, which alsocorrespond to the intermediate compartment. We wereinterested in the possibility that the transient cells withinthe mammalian ganglionic eminence are somehow relatedto the DVR in reptiles. Our tracing study from the dorsalthalamus in mammals suggests that most of the backla-belled internal capsule cells are located medial to theregion of the intermediate territory defined by Fernandezet al. (1998) and do not extend into it. Similarly, in reptiles,ADVR (which has similar developmental gene expressionpattern to the mammalian intermediate zone, Fernandezet al., 1998) does not contain backlabelled cells afterthalamic crystal placements. This finding suggests thatalthough the cortical intermediate zone in mammals andthe ventral border of DVR in reptiles is close to the front ofthe backlabelled cells in the internal capsule, they areexcluded from these structures.
Connectivity of the DVR in turtleat embryonic stages
Our study demonstrated that nucleus rotundus projectsto ADVR, but ADVR cells do not project back to dorsal orventral thalamus (Fig. 17). The BDVR, however, might bedifferent; on one occasion a small number of BDVR cellswere labelled from a dorsal thalamic crystal placement,which may indicate a relationship to the cells of theembryonic mammalian internal capsule. In the adultturtle, ADVR and BDVR have numerous differences intheir connections (Ulinski, 1986). Dorsal cortical cells alsodevelop reciprocal projections with ADVR. Because ourLFB and hypothalamic crystal placements backlabelledBDVR cells, BDVR establishes some connections with anunidentified brainstem structure. Our best candidate isthe hypothalamus; the results from hypothalamic crystalplacements would seem to support this hypothesis (Fig. 9).In adult gecko, parts of DVR possess projections to hypo-thalamus (see Bruce and Neary, 1995). Based on this andother hodologic arguments, Bruce and Neary (1995) sug-gested that the amygdala might be the homologue ofBDVR in mammals.
What is the mammalian homologue of DVR?
The mammalian homologue of the DVR has yet to beconclusively identified. An interesting possibility might bethat the primordium of the DVR is only present tran-siently in mammals and with a specific developmentalrole. Gene expression data could help to place the connec-tional data in context and help to understand the puzzlingbehaviour of thalamocortical and corticothalamic projec-tions in this border region during embryonic developmentin mammals (see Molnar and Cordery, 1999). Similarly,gene expression patterns should be viewed in context andmust be correlated with morphology and hodology.
According to Herrick (1933), the reptilian cortex ischaracterised by superficial laminae of grey substance inthe pallium of the cerebral hemispheres, divided into threelongitudinal stripes: medial and mediodorsal cortex, dor-sal cortex, and lateral cortex. By tracing their connectionsin adults, hodologic arguments suggest that the lateralcortex is homologous to the pyriform cortex and the medialand mediodorsal cortices to the hippocampal formation,
Fig. 18. A schematic diagram of possible boundaries that may behomologous landmarks between reptile (turtle) and mammalian (rat)pallium and thalamus. Our connectional analysis in embryonic ratand turtle and some gene expression patterns both suggest that asimilar boundary exists between striatum and the cortical intermedi-ate zone (in mammals) and the ventral part of the DVR and thestriatum (in turtle). A boundary exists in the thalamus in mammalsbetween TRN and dorsal thalamus, and this border also seems to existin the turtle between dorsal and ventral thalamus. It is yet to bedetermined whether the IC and LFB cells can be subdivided intonumerous subgroups and what is the homologous cell group of thereptilian DVR in mammals. For abbreviations, see list.
EMBRYONIC CONNECTIONS IN TURTLE PALLIUM 49

i.e., dentate gyrus and Ammon’s horn, respectively (re-viewed by Ulinski, 1983). The mammalian neocortex (iso-cortex) is most probably related, at least in part, to thedorsal pallium (Goffinet, 1983, 1984). The DVR is includedhere with cortical areas because it contains several sen-sory fields and is presumably phylogenetically related topallial structures, but its evolutionary origin is not at allclear (Ulinski, 1983; Striedter, 1997). Other authors havepointed out that the DVR of reptiles lies deep to pyriformcortex in mammals. On this basis, authors such as Striedter(1997) have claimed that DVR might be homologous toclaustrum/endopyriform cortex in mammals.
The out-group and the recapitulation hypotheses try toexplain how the neocortex originated from the roof of thecerebral hemisphere and provide different views on theorigin of the DVR (see Northcutt and Kaas, 1995). Accord-ing to the ‘‘out-group hypothesis,’’ the neocortex arose byan independent enlargement of the dorsal pallium, and thecerebral hemispheres in living amphibians are comparableto those of putative ancestral tetrapods. This hypothesisassumes that the DVR arose independently in reptiles(and birds) from part of the lateral pallium. According tothe ‘‘recapitulation hypothesis’’ (Karten, 1969; Shimizuand Karten, 1991), the DVR represents a primitive ances-tral condition and has been retained in living reptiles andbirds, and mammalian isocortex arose by the dual differen-tiation of the dorsal cortex and migration of the cells of thedorsal ventricular ridge. In this case, the dorsal ventricu-lar ridge and dorsal cortex of reptiles are consideredhomologous to mammalian isocortex. This hypothesis alsoassumes that during mammalian development some of thecortical cells are generated in the lateral ganglionic emi-nence and migrate into the cortex. In the literature, thereis support for both theories, and other possibilities (Dıaz etal., 1990; Reiner, 1993; Butler, 1994b; Bruce and Neary,1995; De Carlos et al., 1996; Striedter, 1997; Tamamaki etal., 1997; Anderson et al., 1997; Fernandez et al., 1998;Lavdas et al., 1998). The comparative study of cell lineageand migration together with the analysis of early connec-tivity will eventually answer these questions.
Similarities between reptilian lateral cortexand mammalian perirhinal cortex
The lateral cortex is the recipient of the olfactoryprojections in reptiles, as is perirhinal cortex in mammals(Reiner and Karten, 1985). In embryonic turtle, theseconnections are present from stage 19 (Cordery et al.,1998). In mammals, perirhinal cortex develops early con-nectivity with the internal capsule cells. It seems thatconnectivity with neocortex in mammals is delayed severaldays in this respect. In rat, cells in the internal capsuleproject to perirhinal and more ventral cortices by E14, butdo not seem to extend projections into dorsal cortex untilE17–18. At E14, a small crystal in the primitive internalcapsule labels not only perireticular nucleus (PRN) cells(within the lateral part of the internal capsule) but alsoneurons of the perirhinal cortex and the preplate of theadjacent ventrolateral cortical segment. In embryonicturtle, cells of the lateral forebran bundle have projectionsto both dorsal and lateral cortices (Fig. 10). Lateral cortexprojects to the lateral forebrain bundle but not to dorsalthalamus (Fig. 15). Tracing from dorsal thalamus reveals
thalamic fibres avoiding the lateral cortex (Fig. 6G). Thesehodologic similarities support the hypothesis of homologyof reptilian lateral cortex and olfactory-recipient cortex inmammals.
Evidence for and against homologybetween reptilian dorsal cortex
and mammalian preplate
In embryonic turtles, from stage 19–20 dorsal cortexreceives selective thalamic input from the perirotundalcomplex and develops specific reciprocal connections withDVR, lateral forebrain bundle, and lateral cortex. Inembryonic mammals, the preplate of the neocortex, re-ceives the first thalamocortical projections and projects tonumerous cortical and subcortical targets (Allendoerferand Shatz, 1994). There are two main lines of argumentsfor homology between the reptilian dorsal cortex and themammalian preplate: (1) ontogenetic and (2) morphologic/histochemical, but it is clear that the simple view ofmammalian preplate cells being equivalent to cells of thereptilian dorsal cortex is not sustainable.
(1) Regarding the ontogenetic similarity between reptil-ian and mammalian cortices, there are similar tangentialand radial gradients of development in mammalian pre-plate and reptilian dorsal cortex. Reptilian cortices de-velop in an outside-first, inside-last manner (Goffinet etal., 1986). In mammals, the cortical plate assembles as a‘‘sandwich,’’ with the cortical plate itself splitting theprimordial plexiform zone into two layers and developingin an inside-first, outside-last pattern (Marin-Padilla,1971; Luskin and Shatz, 1985a,b). However, the primor-dial plexiform zone itself (before it is split into subplateand marginal zone by the cortical plate) is generated in the‘‘reptilian’’ outside-first, inside-last manner (Bayer andAltman, 1991). There is a period in ferret cortical develop-ment when birth dating labels only subplate and nomarginal zone cells (Noctor et al., 1997). In reptiles, anoutside-first, inside-last morphogenetic gradient has beendemonstrated at the ultrastructural level (Goffinet, 1990):synapses appear first in the molecular layer, then in thesubcortex, and finally within the cortical ribbon. Thetangential gradients of reptilian and mammalian corticaldevelopment also follow a similar pattern of development.In the medial, dorsal, and lateral supraventricular corticalzones of reptiles, neurogenesis follows a lateral to medialand an anterior to posterior gradient (Goffinet, 1990),similar to gradients in mammalian embryos (reviewed byBayer and Altman, 1991). All these similarities suggesthomology with the mammalian preplate.
(2) Regarding the similarities in morphology and histo-chemical properties between reptilian and primordial plexi-form zone cells, Ramon y Cajal noticed the similaritiesbetween the mammalian polymorph cells of layer 1 andcells of the lizard cortex (Ramon y Cajal, 1909–1911). Thework of Marin-Padilla (1971) and Luskin and Shatz(1985a,b) demonstrated that these marginal zone cells areindeed descendants of the primordial plexiform zone.Marin-Padilla described the splitting of the first gener-ated, polymorphous cell layer (the primordial plexiformzone or preplate) into the marginal zone and the subplate(Marin-Padilla, 1971, 1972, 1988). He called these prema-ture zones the ‘‘the reptilian framework’’ of the developing
50 P. CORDERY AND Z. MOLNAR

mammalian brain. There is increasing evidence that MZcells are composed of various different populations withdifferent developmental origins, some only present tran-siently during early embryonic stages and some migratetangentially to the region from the olfactory cortex (Meyeret al., 1998; Soria et al., 1998). It is conceivable that somesubplate and some cortical plate cells also originate fromprogenitor cells of the ventricular and subventricularzones in the ganglionic eminence and subsequently mi-grate tangentially into the dorsal cortex (De Carlos et al.,1996; Anderson et al., 1997; Tamamaki et al., 1997; Lavdaset al., 1998). The lineage analysis of these cells is a highlyinteresting field. It would argue that cells of the mamma-lian SP and MZ have dual origin. Some residing inprimordial plexiform zone and some subsequently migrat-ing into it.
Origin of the mammalian isocortex
There are two major different views on the evolution ofcell types in mammals: (1) ancestral reptiles contained allmajor cells present in mammalian isocortex, and thesebecame rearranged into a six-layered structure; or (2) thecortex of ancestral reptiles possessed only some of themajor cell types of the mammalian isocortex and new celltypes evolved (reviewed in Reiner, 1993). Although severalsimilar cell types are present in adult turtle cortex and inmammalian isocortex, turtle cortex appears to lack thecholecystokinin, vasoactive intestinal polypeptide, andcholin acetyltransferase containing neurons. These typesof neurons are mostly found in granular and supragranu-lar layers of mammalian isocortex (Reiner, 1991). On thebasis of this observation, Reiner (1991, 1993) postulatedthe addition of new cell types, mostly found in layers 2–4 inmammals, to the reptile-mammal lineage during the evolu-tion of mammalian isocortex. Unfortunately, there arevery few similar comparative studies at early developmen-tal stages. Examining the development of calbindin, calreti-nin, g-aminobutyric acid, and reelin, Meyer and colleagues(1998) proposed that some of the marginal zone cellsoriginate from the retrobulbar ventricle (between thecortex and olfactory bulb) and reach the marginal zonethrough tangential subpial migration later in fetal life. Itis conceivable that different subpopulations of marginalzone cells are only preset in mammals, but not in reptiles.Interestingly, in the turtle dorsal cortex at stage 21, we didnot observe calretinin-positive cells, but there were numer-ous subplate and marginal zone cells labelled in an E16-day-old rat cortex (Cordery et al., 1997). At a similarlyearly stage in turtle (stage 21), NPY-immunoreactive cellswere scattered in all regions of ventral and dorsal pallium,including dorsal cortex, and the neuropil staining did notspare the dorsal and lateral cortex; where it appearedmore homogeneous. These findings do not support theargument for simple homology between all mammaliancells with preplate origin and all dorsal cortex cells ofturtle. We have not yet understood why the NPY-positivecells are present, but the calretinin-positive cells areabsent from the developing turtle cortex. Perhaps suchcells mature later in turtle. This process would seemunlikely, however, because the development of pathwaysin embryonic turtle is quite advanced when compared withrodent at an equivalent stage. Alternatively, calretinin-
positive cells may have been present in ancestral amniotesand subsequently lost in extant turtles. More specificmarkers are needed to resolve this issue. Goffinet andcolleagues described that the reeler gene is highly con-served in numerous amniotes (de Bergeyck et al., 1998).The gene was cloned (D’Arcangelo et al., 1995; Bar et al.,1995; Hirotsune et al., 1995) and shown to encode asecreted glycoprotein named reelin, expressed in the pre-plate of embryonic cortex by Cajal-Retzius cells. A reeler-associated antigen, CR-50, was also found to be expressedby embryonic Cajal-Retzius neurons and could be anepitope belonging to reelin (Ogawa et al., 1995).D’Arcangelo et al. (1997) recently reported that the CR-50antibody recognised reelin in vitro. This protein is ex-pressed in large levels in some cells of the marginal zone inmammals and could be an extremely useful marker toidentify these cells in other amniotes. In the reptiliantelencephalon. Bernier et al. (1998) recently demonstrated(with in situ hybridization of species specific reelin probes)that reelin is expressed in the lateral cortex, and a fewstrongly labelled cells were also present in the molecularlayer and subplate of the dorsal cortex, but less so inmedial cortex. DVR had relatively low level of reelinexpression. Dab1 is a recently identified gene essential forsignalling through reelin (see Goffinet, 1995). Dab1 isstrongly expressed by cortical plate neurons in mammals,and interestingly, Dab1 is also present in the dorsal cortexof reptiles and lizards (Goffinet, 1999). Thus, the detailedanalysis of immunoreactivity (calretinin, NPY, reelin,Dab1) does not support the notion of a simple homology ofall cells of reptilian cortex and preplate, and it is yet to beestablished whether the largely transient marginal zoneor subplate cells of the developing mammalian cortex havehomologous cell groups in reptiles. Reelin and Dab 1expression in turtle cortex suggests that the turtle cortexmight contain similar neurons to the developing mamma-lian cortex at a stage shortly after the appearance of thecortical plate. This suggestion is different from both Marin-Padilla (1971) and Reiner (1993). The combined study ofhighly specific gene expression patterns together withearly connectional and cell lineage studies could be used tocorrelate cell groups in developing mammalian and reptil-ian pallia and, thus, clarify possible homologies (seeFernandez et al., 1998; Rubenstein et al., 1998).
In conclusion, our studies in developing reptilian andmammalian forebrains reveal numerous similarities andsuggest a highly conserved developmental blueprint inthese diverse amniotes. In mammals, the transient cells ofthe marginal zone, subplate, and the internal capsule arepossibly homologous to cells in the reptilian dorsal cortexand lateral forebrain bundle. These cells are believed to belargely transient in mammals, but during developmentthey seem to have similar characteristics and follow asimilar developmental program to reptiles. Cells withsimilar morphology extend the first projections to thedorsal and ventral thalamus from the internal capsule inmammals and the lateral forebrain bundle and striatum inreptiles. Furthermore, thalamocortical projections extendtoward their targets along these cells, possibly using themfor their path finding. The development of diverse amniotebrains, like the development of many other organs, revealstraces of their common ancestry and reflects the course ofevolution.
EMBRYONIC CONNECTIONS IN TURTLE PALLIUM 51

ACKNOWLEDGMENTS
We thank Ann Butler, Andre Goffinet, Anthony Hannan,Alla Katsnelson, Paul Manger, and Michael Pritz fordiscussions and for their thoughtful comments on anearlier version of the manuscript and Colin Blakemore andRichard Adams for support. This study would not havebeen possible without the help of Evelyne Sernagor whointroduced the authors to chelonians. We are indebted toLaurence Waters, Lorraine Chappell, and William Hinkesfor their help with the photography. Z.M. held a JuniorResearch Fellowship at Merton College, Oxford, and anMRC Training Fellowship.
LITERATURE CITED
Adams NC, Baker GE. 1995. Cells of the perireticular nucleus project to thedeveloping neocortex of the rat. J Comp Neurol 359:613–626.
Adams NC, Lozsadi DA, Guillery RW. 1997. Complexities in the thalamocor-tical and corticothalamic pathways. Eur J Neurosci 9:204–209.
Allendoerfer KL, Shatz CJ. 1994. The subplate, a transient neocorticalstructure: its role in the development of connections between thalamusand cortex. Annu Rev Neurosci 17:185–218.
Amadeo A, De Biasi S, Frassoni C, Ortino B, Spreafico R. 1998. Immunohis-tochemical and ultrastructural study of the rat perireticular nucleusduring postnatal development. J Comp Neurol 392:390–401.
Anderson SA, Einsenstat DD, Shi L, Rubenstein JL. 1997. Interneuronmigration from basal forebrain to neocortex: dependence on Dlx genes.Science 278:474–476.
Balaban CD, Ulinski PS. 1981a. Organization of thalamic afferents toanterior dorsal ventricular ridge in turtles. I. Projections of thalamicnuclei. J Comp Neurol 200:95–124.
Balaban CD, Ulinski PS. 1981b. Organisation of thalamic afferents toanterior dorsal ventricular ridge in turtles. II. Properties of therotundo-dorsal map. J Comp Neurol 200:131–150.
Bar I, Lambert de Rouvroit C, Royaux I, Kritzman DB, Dernoncourt C,Ruelle D, Beckers MC, Goffinet AM. 1995. A YAC contig containing thereeler locus with preliminary characterization of candidate gene frag-ments. Genomics 26:543–549.
Bayer SA, Altman J. 1991. Neocortical development. New York: RavenPress.
Belekhova MG. 1994. Thalamo-amygdalar auditory-somatic connectionsare ancient conservative brain characteristics of amniotes (analysis ofpublished and personal data; hypothesis) J Evol Biochem Physiol30:454–473.
Belekhova MG, Zharskaja VD, Kachunts AS, Gardaenko GV, TumanovaNL. 1985. Connections of the mesencephalic, thalamic, and telence-phalic auditory centres in turtles. Some structural bases for audioso-matic interactions. J Hirnforsch 26:127–152.
Belekhova MG, Kosareva AA, Vesselkin NP, Ermakova TV. 1992. Telence-phalic afferent connections in the turtle Emys orbicularis: a peroxidasestudy. J Evol Biochem Physiol 15:97–103.
Bernier B, Goffinet AM, Lambert de Rouvroit C. 1998. Comparative studiesof reelin during brain development. Eur J Neurosci 10(Suppl 10):381.
Brauth SE, Reiner CA, Kitt CA, Karten HJ. 1983. A substance P-containingpathway from the basal ganglia to the midbrain tegmentum in reptiles.J Comp Neurol 219:305–327.
Bruce LL, Neary TJ. 1995. The limbic system of tetrapods: a comparativeanalysis of cortical and amygdalar populations. Brain Behav Evol46:224–234.
Butler AB. 1994a. The evolution of the dorsal thalamus of jawed verte-brates, including mammals: cladistic analysis and a new hypothesis.Brain Res Rev 19:29–65.
Butler AB. 1994b. The evolution of the dorsal pallium in the telencephalonof amniotes: cladistic analysis and a new hypothesis. Brain Res Rev19:66–101.
Butler AB. 1995. The dorsal thalamus of jawed vertebrates: a comparativeviewpoint. Brain Behav Evol 46:209–223.
Butler AB, Hodos W. 1996. Comparative vertebrate neuroanatomy. Evolu-tion and adaptation. New York, Chichester: Wiley-Liss.
Butler A, Molnar, Z. 1998. Development and evolution in nervous systems:development and evolution of ideas. Trends Neurosci 21:177–178.
Clasca F, Angelucci A, Sur M. 1995. Layer-specific programs of developmentin neocortical projection neurons. Proc Natl Acad Sci USA 92:11145–11149.
Cordery P, Blakemore C, Molnar Z. 1997. Comparison of mammalian andreptilian pallium during development. Soc Neurosci Abstr 23:75.
Cordery P, Butler A, Molnar Z. 1998. Development of the olfactory bulbprojections in embryonic turtle. Eur J Neurosci 10(Suppl 10):196.
Crabtree JW, Kind PC. 1993. Monoclonal antibody Cat-301 selectivelyidentifies a subset of nuclei in the cat’s somatosensory thalamus. JNeurocytology 22:903–912.
D’Arcangelo G, Miao GG, Chen SC, Soares HD, Morgan JI, Curran T. 1995.A protein related to extracellular matrix proteins deleted in the mousemutant reeler. Nature 374:719–723.
D’Arcangelo G, Nakajima K, Miyata T, Ogawa M, Mikoshiba K, Curran T.1997. Reelin is a secreted glycoprotein recognized by the CR-50monoclonal antibody. J Neurosci 17:23–31.
Davila JC, de la Calle A, Gutierrez A, Megias M, Andreu MJ, Guirado S.1991. Distribution of neuropeptide Y (NPY) in the cerebral cortex of thelizards Psammodromus algirus and Podarcis hispanica: co-localisationof NPY, somatostatin, and GABA. J Comp Neurol 308:397–408.
De Carlos JA, Schlaggar BL, O’Leary DDM. 1995. Development of acetylcho-linesterase-positive thalamic and basal forebrain afferents to embry-onic rat neocortex. Exp Brain Res 104:385–401.
De Carlos JA, Lopez-Mascaraque L, Valverde F. 1996. Dynamics of cellmigration from the lateral ganglionic eminence in the rat. J Neurosci16:6146–6156.
de Bergeyck V, Naerhuyzen B, Goffinet AM, Lambert de Rouvroit C. 1998. Apanel of monoclonal antibodies against Reelin, the extracellular matrixprotein defective in reeler mutant mice. J Neurosci Methods 82:17–24.
Dıaz C, Yanes C, Medina L, Monz—n M, Trujillo CM, Puelles L. 1990. Golgistudy of the anterior dorsal ventricular ridge in a lizard. I. Neuronaltypology in the adult. J Morphol 203:293–300.
Dori I, Parnavelas JG. 1989. The cholinergic innervation of the rat cerebralcortex shows two distinct phases in development. Exp Brain Res76:417–423.
Earle KL, Mitrofanis J. 1996. Genesis and fate of the perireticular thalamicnucleus during early development. J Comp Neurol 366:1–18.
Fernandez AS, Pieau C, Reperant J, Boncinelli E, Wassef M. 1998.Expression of the Emx-1 and Dlx-1 homeobox genes define threemolecularly distinct domains in the telencephalon of mouse, chick,turtle and frog embryos: implications for the evolution of telencephalicsubdivisions in amniotes. Development 101:2099–2111.
Foster GA. 1998. Chemical anatomy of the prenatal rat brain. Oxford, NewYork, Tokyo: Oxford Univeristy Press.
Frotscher M. 1998. Cajal-Retzius cells, Reelin, and the formation of layers.Curr Opin Neurobiol 8:570–575.
Godement P, Vanselow J, Thanos S, Bonhoeffer F. 1987. A study indeveloping visual systems with a new method of staining neurones andtheir processes in fixed tissue. Development 101:697–713.
Goffinet AM. 1983. The embryonic development of the cortical plate inreptiles: a comparative analysis. J Comp Neurol 219:10–24.
Goffinet AM. 1984. Events governing organization of postmigratory neu-rons: studies on brain development in normal and reeler mice. BrainRes Rev 7:261–296.
Goffinet AM. 1990. Cortical architectonic development: a comparativestudy in reptiles. Exp Brain Res 19:135–144.
Goffinet, AM. 1995. A real gene for reeler Nature, 374: 675–676.Goffinet AM. 1999. Evolution of mammalian cortical lamination: the
reelin/Dab1 pathway and cortical evolution. In: Bock G, Cardew G,editors. Evolutionary developmental biology of the cerebral cortex.Novartis Symposium 228. (in press).
Goffinet AM, Daumerie C, Langewerf B, Pieau C. 1986. Neurogenesis in
52 P. CORDERY AND Z. MOLNAR

reptilian cortical structures: 3H-thymidine autoradiographic analysis. JComp Neurol 243:106–116.
Hall JA, Foster RE, Ebner FF, Hall WC. 1997. Visual cortex in a reptile, theturtle (Pseudemys scripta and Chrysemys picta). Brain Res 120:197–216.
Hall WC, Ebner FF. 1970. Thalamotelencephalic projections in the turtle(Pseudemys scripta). J Comp Neurol 140:101–122.
Herrick CJ. 1933. The amphibian forebrain. VI. Necturus. J Comp Neurol58:1–288.
Hirotsune S, Takahara T, Sasaji N, Hirose K, Yoshiki A, Ohashi T,Kusakabe M, Murakami Y, Muramatsu M, Watanabe S, et al. 1995. Thereeler gene encodes a protein with an EGF-like motif expressed bypioneer neurons. Nat Genet 10:77–83.
Karten HJ. 1969. Comparative and evolutionary aspects of the vertebratecentral nervous system. Ann NY Acad Sci 169:164–179.
Karten HJ. 1997. Evolutionary developmental biology meets the brain: theorigins of mammalian cortex. Proc Natl Acad Sci USA 94:2800–2804.
Kostovic I, Rakic P. 1990. Developmental history of the transient subplatezone in the visual and somatosensory cortex of the macaque monkeyand human brain. J Comp Neurol 297:441–470.
Lavdas AA, Grigoriou M, Pachnis V, Parnavelas GJ. 1998. The medialganglionic eminence is a source of the early neurons of the developingcerebral cortex. Soc Neurosc Abstr 24:282.
Levitic K, Kostovic I. 1996. Transient neuronal population of the internalcapsule in the developing human cerebrum. Neuroreport 7:2159–2162.
Liu F-C, Graybiel AM. 1992. Transient calbindin-D28K-positive systems inthe telencephalon: ganglionic eminence, developing striatum and cere-bral cortex. J Neurosci 12:674–690.
Luskin MB, Shatz CJ. 1985a. Neurogenesis of the cat’s primary visualcortex. J Comp Neurol 242:611–631.
Luskin MB, Shatz CJ. 1985b. Studies of the earliest-generated cells of thecat’s visual cortex: cogeneration of subplate and marginal zones. JNeurosci 5:1062–1075.
Marın O, Smeets WJAJ, Gonzalez A. 1998. Evolution of the basal ganglia intetrapods: a new perspective based on recent studies in amphibians.Trends Neurosci 11:487–494.
Marin-Padilla M. 1971. Early prenatal ontogenesis of the cerebral cortex(neocortex) of the cat (Felis domestica): a Golgi study. I. The primordialneocortical organization. Z Anat Entwicklungsgesch 134:117–145.
Marin-Padilla M. 1972. Prenatal ontogenetic history of the principalneurons of the neocortex of the cat (Felis domestica): a Golgi study. II.Developmental differences and their significances. Z Anat Entwick-lungsgesch 136:125–142.
Marin-Padilla M. 1988. Early ontogenesis of the human cerebral cortex. In:Peters A, Jones EG, editors. Cerebral Cortex, Vol 7. Development andmaturation of cerebral cortex. New York, London: Plenum Press. p1–34.
McConnell SK, Gosh A, Shatz CJ. 1989. Subplate neurons pioneer the firstaxon pathway from the cerebral cortex. Science 245:978–982.
Medina L, Reiner A. 1995. Neurotransmitter organization and connectivityof the basal ganglia in vertebrates: implications for the evolution ofbasal ganglia. Brain Behav Evol 46:235–258.
Medina L, Smeets WJ. 1991. Comparative aspects of the basal ganglia-tectal pathways in reptiles. J Comp Neurol 308:614–629.
Metin C, Godement P. 1996. The ganglionic eminence may be an intermedi-ate target for corticofugal and thalamocortical axons. J Neurosci16:3219–3235.
Metin C, Deleglise D, Serafini T, Kennedy TE, Tessier-Lavigne M. 1997. Arole for netrin-1 in the guidance of cortical efferents. Development124:5063–5074.
Meyer G, Soria JM, Martınez-Galan JR, Martın-Clemente B, Fairen A.1998. Different origins and developmental histories of transient neu-rons in the marginal zone of the fetal and neonatal rat cortex. J CompNeurol 397:493–518.
Mitrofanis J. 1992. Patterns of antigenic expression in the thalamicreticular nucleus of developing rats. J Comp Neurol 320:161–181.
Mitrofanis J. 1994. development of the pathway from the reticular andperireticular nuclei to the thalamus in ferrets: a DiI study. Eur JNeurosci 6:1865–1882.
Mitrofanis J, Baker GE. 1993. Development of the thalamic reticular andperireticular nuclei in rats and their relationship to the course ofgrowing corticofugal and corticopetal axons. J Comp Neurol 338:575–587.
Mitrofanis J, Guillery RW. 1993. New views of the thalamic reticularnucleus in the adult and developing brain. Trends Neurosci 16:240–245.
Molnar Z. 1998. Development of thalamocortical connections. Berlin,Heidelberg: Springer-Verlag and Georgetown, Texas: RG Landes Com-pany.
Molnar Z, Blakemore C. 1995. How do thalamic axons find their way to thecortex? Trends Neurosci 18:389–397.
Molnar Z, Cordery PM. 1999. Connections between cells of the internalcapsule, thalamus and cerebral cortex in embryonic rat. J Comp Neurol413:1–25.
Molnar Z, Sernagor E. 1998. Embryonic development of the retinalprojections in the turtle (Pseudemys scripta elegans). J Anat 192:460–461.
Molnar Z, Adams R, Blakemore C. 1998a. Mechanisms underlying theestablishment of topographically ordered early thalamocortical connec-tions in the rat. J Neurosci 18:5723–5745.
Molnar Z, Knott GW, Blakemore C, Saunders NR. 1998b. Development ofthalamocortical projections in the south American grey short-tailedOpossum (Monodelphis Domestica).J Comp Neurol 398:491–514.
Noctor SC, Scholnicoff NJ, Juliano SL. 1997. Histogenesis of ferret somato-sensory cortex. J Comp Neurol 387:179–193.
Northcutt RG, Kaas JH. 1995. The emergence and evolution of mammalianneocortex. Trends Neurosci 18:373–379.
Ogawa M, Miyata T, Nakajima K, Yagyu K, Seike M, Ikenaka K, YamamotoH, Mikoshiba K. 1995. The reeler gene-associated antigen on Cajal-Retzius Neurons is a crucial molecule for laminar organization ofcortical neurons. Neuron 14:899–912.
Parent A. 1976. Striatal afferent connections in the turtle (Choysenys picta)as revealed by retrograde axonal tranport of horseradish peroxidase.Brain Res 108:25–36.
Price DJ, Aslam S, Tasker L, Gilles K. 1997. Fates of the earliest generatedcells in the developing murine neocortex. J Comp Neurol 377:414–422.
Pritz MB. 1995. The thalamus of reptiles and mammals: similarities anddifferences. Brain Behav Evol 46:197–208.
Rakic P. 1976. Prenatal genesis of connections subserving ocular domi-nance in the rhesus monkey. Nature 261:467–471.
Ramcharan EJ, Guillery RW. 1997. Membrane specializations in thedevelopmentally transient perireticular nucleus of the rat. J CompNeurol 365:435–448.
Ramon y Cajal S. 1909–1911. Histologie du Systeme Nerveux de l’Hommeet des Vertebres, 2 vols. trans L Azoulay), repr Instituto Ramony Cajaldel CSIC, Madrid, 1952–1955.
ReinerA. 1991.Acomparison of neurotransmitter-specific and neuropeptide-specific neuronal cell types present in the dorsal cortex in turtles withthose present in the isocortex in mammals: implications for theevolution of isocortex. Brain Behav Evol 38:53–91.
Reiner A. 1992. Neuropeptides in the nervous system. In: Gans C, UlinskiPS, editors. Sensorimotor integration biology of the Reptilia, vol 17.Chicago: University of Chicago Press. p 587–739.
Reiner A. 1993. Neurotransmitter organization and connections of turtlecortex: implications for the evolution of mammalian isocortex. CompBiochem Physiol 104:735–748.
Reiner A, Karten HJ. 1985. Comparison of olfactory bulb projections inpigeons and turtles. Brain Behav Evol 27:11–27.
Reiner A, Brauth SE, Kitt CA, Karten JK. 1980. Basal ganglionic pathwaysto the tectum: studies in reptiles. J Comp Neurol 193:565–589.
Richards LJ, Koester SE, Tuttle R, O’Leary DDM. 1997. Directed growth ofearly cortical axons in influenced by a chemoattractant released froman intermediate target. J Neurosci 17:2445–2458.
Rubenstein JL, Shimamura K, Martinez S, Puelles L. 1998. Regionaliza-tion of the prosencephalic neural plate. Annu Rev Neurosci 21: 445–477.
Saper CB. 1984. Organisation of cerebral cortical afferent systems in therat. II. Magnocellular basal nucleus. J Comp Neurol 222:313–342.
Sernagor E, Grzywacz NM. 1996. Influence of spontaneous activity and
EMBRYONIC CONNECTIONS IN TURTLE PALLIUM 53

visual experience on developing retinal receptive fields. Curr Biol6:1503–1508.
Shimizu T, Karten HJ. 1991. Multiple origins of neocortex: contributions ofthe dorsal ventricular ridge. In: Finlay BL, et al., editors. The neocor-tex. New York: Plenum Press.
Soria JM, Meyer G, Fairen A. 1998. Cortical pioneer cells acquire specifictangential distributions during rat corticogenesis. Eur J Neurosci10(Suppl. 10):8.
Striedter GF. 1997. The telencephalon of tetrapods in evolution. BrainBehav Evol 49:179–213.
Tamamaki N, Fujimori KE, Takauji R. 1997. Origin and route of tangen-tially migrating neurons in the developing neocortical intermediatezone. J Neurosci 17:8313–8323.
Ulinski PS. 1983. Dorsal ventricular ridge: a treatise on forebrain organiza-tion in reptiles and birds. New York: John Wiley & Sons.
Ulinski PS. 1986. The ecology and biology of mammalian-like reptiles.Holten N, et al., editors. Washington DC: Smithsonian Inst Press. p147–172.
Yntema CL. 1968. A series of stages in the embryonic development ofChelydra serpentina. J Morphol 125:219–252.
54 P. CORDERY AND Z. MOLNAR