embryology: gastrulation and neurulation · pdf fileendoderm) into a trilaminar disc...
TRANSCRIPT

Dr. Raymond Colello
EMBRYOLOGY: Gastrulation and Neurulation
Recommended Reading: Larsen, pp. 53-102 Langman, pp. 53-66, 67-83 Moore, pp. 61-72 OBJECTIVES: Following the lecture the students should be able to:
1.) Define the following terms: gastrulation, primitive streak, notochord, somites and neural tube.
2.) Describe the tissues and organs formed from the three germ
layers- ectoderm, mesoderm and endoderm.
3.) Identify common malformations arising as a result of the disruption of gastrulation.
4.) Describe the time course of neurulation.
5.) Describe the difference between primary neurulation and
secondary neurulation.
6.) List the cell type/tissue derivatives of the neural crest cells.
7.) Describe common malformations of neural tube closure.
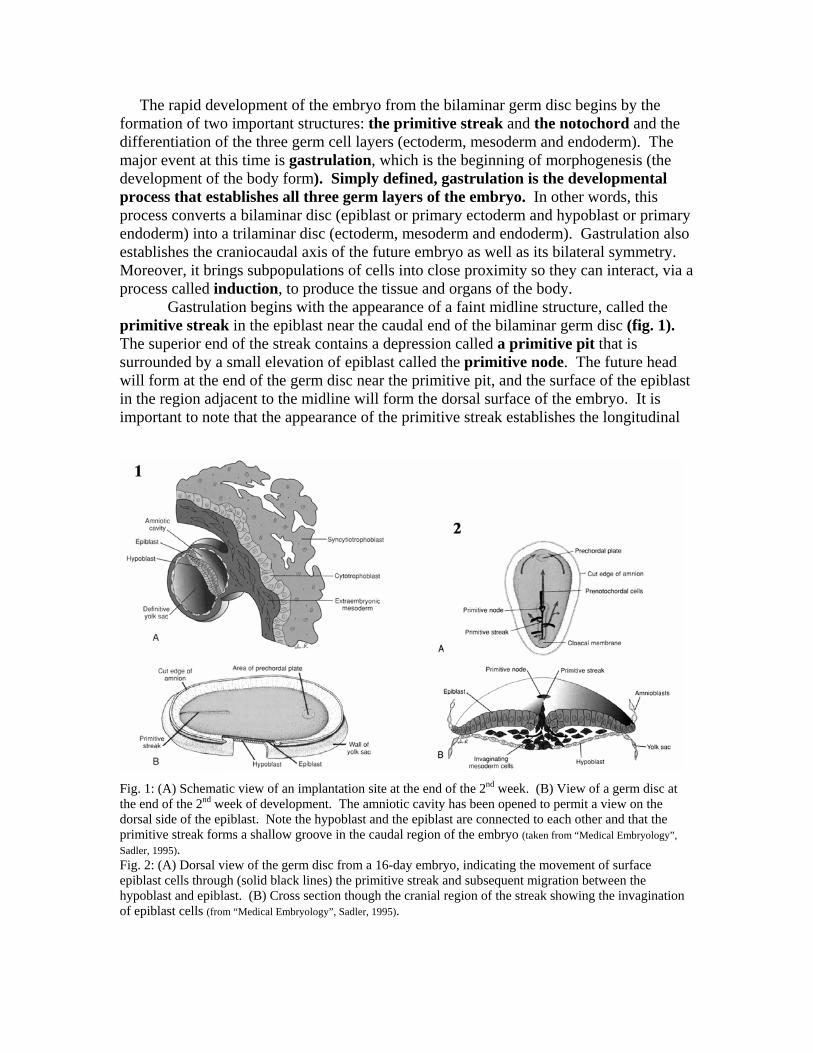
The rapid development of the embryo from the bilaminar germ disc begins by the formation of two important structures: the primitive streak and the notochord and the differentiation of the three germ cell layers (ectoderm, mesoderm and endoderm). The major event at this time is gastrulation, which is the beginning of morphogenesis (the development of the body form). Simply defined, gastrulation is the developmental process that establishes all three germ layers of the embryo. In other words, this process converts a bilaminar disc (epiblast or primary ectoderm and hypoblast or primary endoderm) into a trilaminar disc (ectoderm, mesoderm and endoderm). Gastrulation also establishes the craniocaudal axis of the future embryo as well as its bilateral symmetry. Moreover, it brings subpopulations of cells into close proximity so they can interact, via a process called induction, to produce the tissue and organs of the body.
Gastrulation begins with the appearance of a faint midline structure, called the primitive streak in the epiblast near the caudal end of the bilaminar germ disc (fig. 1). The superior end of the streak contains a depression called a primitive pit that is surrounded by a small elevation of epiblast called the primitive node. The future head will form at the end of the germ disc near the primitive pit, and the surface of the epiblast in the region adjacent to the midline will form the dorsal surface of the embryo. It is important to note that the appearance of the primitive streak establishes the longitudinal
Fig. 1: (A) Schematic view of an implantation site at the end of the 2nd week. (B) View of a germ disc at the end of the 2nd week of development. The amniotic cavity has been opened to permit a view on the dorsal side of the epiblast. Note the hypoblast and the epiblast are connected to each other and that the primitive streak forms a shallow groove in the caudal region of the embryo (taken from “Medical Embryology”, Sadler, 1995). Fig. 2: (A) Dorsal view of the germ disc from a 16-day embryo, indicating the movement of surface epiblast cells through (solid black lines) the primitive streak and subsequent migration between the hypoblast and epiblast. (B) Cross section though the cranial region of the streak showing the invagination of epiblast cells (from “Medical Embryology”, Sadler, 1995).

Fig. 3: Dorsal view of the embryonic disc early in the third week showing the invagination and migration of epiblast cells to form the trilaminar disc (from, “Before we were born”, Moore and Persaud, 1998). axis and, thus, the bilateral symmetry of the future adult. In other words, the tissues to the right of this structure give rise to the right side of the body, and the tissues to the left of it give, in general, to the left side of the body. While the form of the primary germ layers will change with folding of the embryo in the fourth week, the fundamental cranial/caudal, left/right, and ventral/dorsal axes of the body are established early in the third week.
Shortly after the primitive streak appears, at about day 16, the epiblast cells near the primitive streak begin proliferate, flatten and detach from each other. They then migrate through the streak into the space between the epiblast and hypoblast. (fig. 2B). This process of invagination is called gastrulation. The first epiblast cells to move inward displace the hypoblast and eventually replace the hypoblast layer with a new layer of cells, the definitive endoderm. The definitive endoderm will give rise to the lining of the gut and gut derivatives. Epiblast cells migrating into the space between the epiblast and the newly formed endoderm will form a third germ layer, the intraembryonic mesoderm. These cells can then migrate cranially to form a midline structure called the notochord process, which later hollows out to form the notochord (fig. 3). The notochord serves as a basis for the development of the axial skeleton (bones of the head and vertebral column). Other cells migrate laterally to form rod- and sheetlike structures on either side of the notochord. The mesoderm lying immediately to either side of the notochord forms the paraxial mesoderm, just lateral to this is the intermediate mesoderm and further lateral is the lateral plate mesoderm (fig. 4). These three mesodermal structures give rise to the following specific structures in the adult: Paraxial Mesoderm Intermediate mesoderm Lateral plate mesodermAxial skeleton Urinary System Epithelial covering of viscera Voluntary muscles Parts of the Genital System Parts of Limb Part of the dermis Most of Dermis Inner lining of body wall
Two midline regions devoid of mesoderm are the cranial, buccopharyngeal membrane, and the caudal, cloacal membrane (fig. 4). These membranes become the blind end of the gut tube, with the buccopharyngeal membrane breaking down in the 4th week to form the opening of the oral cavity and the cloacal membrane breaking down in the 7th week to form the openings of the anus and urogenital tracts.

Fig. 4: Sections through a 17-day embryo showing the differentiation of the mesoderm on either side of the midline (taken from “Human Embryology”, Larsen, 1993).
Fig. 5: Dorsal view of an embryo from 16 to 20 days of development. Fig. 6: The lateral edges of the neural folds first begin to fuse in the occipitocervical region on Day 22, leaving the cranial and caudal neuropores open at each end. The neural tube increases in length as it zippers up both cranially and caudally. The cranial neuropore closes on day 24 and the caudal neuropore closes on day 26 (taken from “Human Embryology”, Larsen, 1993).

Throughout the 3rd and 4th week of development, the paraxial mesoderm begins to form into a series of rounded structures called somitomeres. Formation of these structures occurs in a cranio-caudal progression starting with several pairs in the presumptive cranial region. The first seven pairs of somitomeres eventually give rise to the striated muscles of the face, jaw, and throat. The remaining somitomeres differentiate into the somites (42-44 pairs) flanking the notochord from the occipital region to the embryonic tail (figs. 5,6). The somites establish the segmental organization of the body by giving rise to most of the axial skeleton as well as the voluntary muscles of the neck, body wall, and limbs. Once the somites begin to form they each separate into three subdivisions, (myotomes, dermatomes, and sclerotomes) which give rise to specific mesodermal components. The myotomes differentiate into myogenic (muscle-producing) cells, the dermatomes differentiate into the dermis of the neck, the back, and the ventral and lateral trunk, and the sclerotomes develop into the vertebrae.
During the 4th week, the tissue layers differentiate to form the primitive structures of most of the body organs. Simultaneously, the embryonic disc undergoes a process of folding that creates the basic vertebrate body form (fig. 7). As a result of folding, the cephalic, lateral, and caudal edges of the germ disc are brought together along the ventral midline. Each of the germ layers then fuses to the corresponding layers on the opposite side to create a three-dimensional body form.
Fig. 7: Drawing of sagittal midline sections of embryos at various stages of development to demonstrate cephalocaudal folding and its effect on the position of the endoderm-lined cavity. (A) Presomite embryo, (B) Seven-somite embryo, (C) Fourteen somite embryo, (D) At the end of the 1st month (Taken from “Medical Embryology”, Langman, 1995).

Clinical Correlates The developmental period encompassing gastrulation (3rd week of development) is highly susceptible to agents (i.e. drugs or virus) that can lead to congenital abnormalities. One congenital abnormality associated with gastrulation is sirenomelia, which results in the fusion of the lower extremity limb buds during development leading to a “mermaidlike” appearance of the fetus. Neurulation During the 4th week, a process of folding called neurulation occurs. As you remember, the neural plate is a thickening in the epiblast along the midsagittal axis cranial to the primitive pit (fig. 5). The expanded cranial portion gives rise to the brain, whereas, the narrow caudal portion gives rise to the spinal cord. As a likely result of inductive signals by the closed apposed notochord, the neural plate begins to crease ventrally along its midline at the neural groove. (fig. 8). This action forms two neural folds, whose lateral lips meet dorsally to form a tube enclosing a space called the neural canal. This tube pinches itself free from the surface ectoderm, which fuses, and separates from it and sinks into the posterior body wall.
Fig. 8: Diagrammatic transverse sections though progressively older embryos, illustrating formation of the neural groove, the neural tube, and the neural crest up to the end of the fourth week (taken from “Before we were born”, Moore and Persaud. 1998).

Closure of the neural tube proceeds bidirectionally, ending with closure of the cranial and caudal neuropores. A failure of part of the neural tube to close disrupts both the differentiation of the CNS and the induction of the vertebral arches and can result in a number of anomalies. For example, if the vertebral arches remain underdeveloped and fail to fuse along the dorsal midline to enclose the vertebral canal, the result open vertebral canal is a condition called spinal bifida. Spinal bifida and related defects of cranial neuropore closure results in a range of malformations (fig. 9 and 10)
Fig. 9: Defective development of neural arches may result in formation of a cyst or cele.
Fig. 10: Types of Spina Bifida

During neurulation, neural crest cells detach from the lips of the neural fold and migrate to specific locations in the body, where they give rise to numerous structures (fig. 8). By utilizing cell tracing studies it has been found that cephalic neural crest give rise to diverse structures of the head and neck while occipital and spinal neural crest produce major components of the peripheral nervous system (see list below):
Cranial neural crest Spinal neural crest Some cranial nerve ganglia Preaortic ganglia Odontoblasts Adrenal medulla Connective tissue surrounding eye Dorsal root ganglia Pharyngeal arch cartilages Chain ganglia Dermis of face and neck Schwann cells of PNS Schwann cells of PNS Melanocytes Melanocytes Arachnoid and pia mater Arachnoid and pia mater Major changes in body form occurring from the 4th week through the 8th week result in an embryo now having a distinct human appearance (fig. 9). Although only 16.0 mm in length, all of the major organ systems have been formed and sexual differentiation of the embryo has taken place.
Fig. 9: Illustrations showing the development of the embryo from about 22 days of development (5-somite stage) to 56 days (taken from “Before we were born”, Moore and Persaud. 1998).