effects of roundup (glyphosate) on gut microorganisms of ...library.au.dk/fileadmin/ ·...
TRANSCRIPT

Charlotte Lynggaard Katholm Master Thesis, Agrobiology – Health and Welfare May 2015 – January 2016 60 ECTS
Matriculation number: 20095408 Department: Department of Animal Science, Research Centre Foulum, Aarhus University Principal supervisor: Senior scientist Ole Højberg Co – supervisor: Senior scientist Martin Tang Sørensen
Effects of Roundup (glyphosate) on gut microorganisms of farm animals
Effekter af Roundup (glyfosat) på mikroorganismer fra husdyr


Preface and Acknowledgement This master thesis is completed as a final part of the master degree, Agrobiology – Animal Health and Welfare,
at Aarhus University.
The thesis corresponds to 60 ECTS and consists of a literature review combined with an experimental part.
The experiments were conducted in the laboratory facilities at Research Centre Foulum, Aarhus University.
The focus is whether Roundup (glyphosate) affects gut microorganisms of farm animals. First and foremost, special thanks goes to Senior Scientist, Ole Højberg, Department of Animal Science –
Immunology and Microbiology and Senior Scientist Martin Tang Sørensen, Department of Animal Science –
Animal Health, Welfare and Nutrition, both Research Centre Foulum, Aarhus University. They have both been
of great help and guidance throughout the process; making themselves available, answering questions all the
way from the beginning, and right until the end.
Thanks also goes to the Immunology and Microbiology group, and especially to the laboratory staff; Trine
Poulsen, Thomas Rebsdorf and Karin Durup. I could not have done the laboratory work without their help.
Furthermore, thanks to Nuria Canibe, Department of Animal Science – Immunology and Microbiology and
Leslie Foldager, Department of Animal Science – Behaviour and Streesbiology, for statistical guidance.
Last, but definitely not least, thanks to Ann-Sofie Riis Poulsen for always making herself available and to
Rasmus Krarup, Katrine Bjørn and Christina Katholm, for help proofreading the thesis.
Aarhus University, January 31th 2016
Charlotte Lynggaard Katholm


Abstract
Glyphosate, is a broad-spectrum, nonselective, systemic herbicide, existing in the form of the acid itself or
formulated as a salt. Roundup consists of the glyphosate isopropylamine (IPA) salt, water and a surfactant,
often a polyethoxylated tallowamine, POEA.
Glyphosate inhibits 5-enolpyruvylshikimate-3-phosphate synthase (EPSPS) in the shikimate-pathway, which
leads to formation of aromatic amino acids. As the shikimate-pathway is present in plants, fungi and bacteria,
and not in animals, usually glyphosate has been considered as non-toxic to animals. However, recent studies
have raised concerns about the effects of glyphosate on gut microbiota, indirectly affecting farm animals,
when fed feed, containing residues of glyphosate. It has been proposed that glyphosate has a potential
inhibiting effect on growth of commensal bacteria, normally occupying the gut of farm animals, whereas
potential pathogens in general, should be more tolerant.
The present study was conducted to investigate effects of glyphosate on growth of selected commensals and
potential pathogens, in lab media. In addition, effects on the overall activity of microbiota in stomach, cecum
and colon contents from pigs, as well as in rumen contents from cows, were investigated. Glyphosate acid,
glyphosate IPA salt, Roundup and POEA, were included to differentiate eventual effects between different
compounds, included in commercial glyphosate formulations.
We were able to show that glyphosate suppressed growth of both commensals and potential pathogens, and
that inhibition differed between gram-positive and gram-negative bacteria; gram-negative being more
tolerant towards all treatments. In addition, glyphosate changed fermentation pattern in the gut of pigs and
cows and overall, the effect of glyphosate depended on the chemical formulation (acid, salt, mixture and
surfactant) used. In general, glyphosate acid were the least toxic of the treatments, whereas toxicity levels
of glyphosate IPA salt, Roundup and POEA were higher and more similar. The concentrations, at which we
observed an effect on gut content, were much higher than the concentrations affecting individual bacteria,
indicating that gut microbiota is complex.
As pigs and cows are not fed glyphosate directly, feeding trials should be performed, to give a better
indication of how, and if, the animals are affected by residues of glyphosate in feed.


Sammendrag
Glyfosat er et bredspektret, ikke-selektivt, systemisk herbicid, der findes som en syre eller formuleret som et
salt. Roundup består af glyfosat isopropylamin (IPA) saltet, vand og et overfladeaktivt middel, ofte et
polyethoxyleret amin, POEA.
Glyfosat inhiberer 5-enolpyruvylshikimat-3-phosphat-syntasen (EPSPS) i shikimate-pathway’en, som fører til
dannelsen af aromatiske aminosyrer. Shikimate pathway’en er tilstede i planter, svampe og bakterier, men
ikke i dyr, og derfor er glyfosat normalt blevet betragtet som ikke-toksisk overfor dyr. Imidlertid har nyere
undersøgelser udtrykt bekymring overfor virkningerne af glyfosat på tarmens mikroflora, som indirekte kan
have indflydelse på husdyrenes ve og vel, hvis de får foder, der indeholder rester af glyfosat. Det er blevet
foreslået, at glyfosat har en potentiel hæmmende effekt på væksten af kommensale bakterier mens
potentielle patogener overordnet er mere tolerante.
Den foreliggende undersøgelse blev udført for at undersøge virkningerne af glyfosat på væksten af udvalgte
kommensale og potentielle patogener i laboratorie medier. Derudover blev virkninger på den samlede
aktivitet af mikrofloraen i mave -, blindtarms - og tyktarmsindhold fra svin, såvel som i vomindhold fra køer,
undersøgt. Både glyfosat syren, glyfosat IPA saltet, Roundup og POEA, blev inkluderet for at skelne mellem
eventuelle virkninger mellem de forbindelser, der normalt indgår i kommercielle glyfosat-produkter.
Vi var i stand til at vise, at glyfosat (Roundup) hæmmede væksten af både kommensale og potentielle
patogene bakterier, og at hæmningen var forskellig for gram-positive og gram-negative bakterier; gram-
negative var mere tolerante overfor alle behandlingerne. Derudover ændrede glyfosat (Roundup)
forgæringsmønster i mave, blind – og tyktarm hos grise og i vommen hos køer. Overordnet afhang virkningen
af, hvilke formuleringer (syre, salt, Roundup og POEA) der blev anvendt. Generelt var glyfosat syren den
mindst toksiske af behandlingerne, mens toksisiteten af glyfosat IPA saltet, Roundup og POEA var større, og
generelt mere ens. De koncentrationer, hvor vi observerede en effekt på mave -, tarm – og vomindhold, var
meget højere end de koncentrationer, der påvirkede de enkelte bakterier, hvilket indikerer, at den
mikrobielle flora er en kompleks størrelse.
Da grise og køer ikke fodres direkte med glyfosat, bør der udføres fodringforsøg, for at give en bedre
indikation af, hvordan, og hvis, dyrene påvirkes af rester af glyfosat i foder.


Content Theory ................................................................................................................................................................ 1
Introduction ................................................................................................................................................... 1
Working hypotheses .................................................................................................................................. 2
Objectives .................................................................................................................................................. 2
Glyphosate formulations ............................................................................................................................... 3
The chemical properties of glyphosate ..................................................................................................... 3
Surfactants in Roundup ............................................................................................................................. 4
Herbicidal mechanism of glyphosate ............................................................................................................ 5
5-enolpyruvylshikimate-3-phosphate synthase ........................................................................................ 6
The main target of glyphosate – the plant .................................................................................................... 7
Degradation of glyphosate and subsequent effects on soil .......................................................................... 8
Production of glyphosate resistant (GR) crops .............................................................................................. 9
The success of glyphosate ........................................................................................................................... 11
Possible pesticide residues in crops treated with glyphosate ..................................................................... 13
Toxicity of glyphosate .................................................................................................................................. 15
Glyphosate as a carcinogen? ................................................................................................................... 15
Toxicity of POEA ....................................................................................................................................... 16
Toxicity of glyphosate towards microorganisms ......................................................................................... 16
Changes in gut microbiota with respect to glyphosate ............................................................................... 17
Potential levels of glyphosate in gut ....................................................................................................... 17
The effect of glyphosate on poultry microbiota ...................................................................................... 18
The effect of glyphosate on dairy cow microbiota .................................................................................. 19
Microbial fermentation in the gut of pigs and dairy cows .......................................................................... 20
Experimental setup ...................................................................................................................................... 21
Materials and Methods ................................................................................................................................... 22
Chemicals ..................................................................................................................................................... 22
Bacteria used in the experiment ................................................................................................................. 22
Media ........................................................................................................................................................... 22
Techniques used .......................................................................................................................................... 22
Determination of viable counts by use of Drop Plate Procedure............................................................ 22
Quantification of VFA analysis ................................................................................................................. 23
Gas measurements .................................................................................................................................. 23
Influence of glyphosate acid, glyphosate IPA salt, Roundup and POEA on growth of bacterial cultures ... 23

Influence of glyphosate IPA salt, Roundup and POEA on gut microbiota from pigs and cows ................... 25
Slaughter pigs .......................................................................................................................................... 25
Dairy cows ............................................................................................................................................... 26
Influence of glyphosate IPA salt on growth of L. sobrius in stomach content from pigs ............................ 27
Statistical analyses ....................................................................................................................................... 28
Results ............................................................................................................................................................. 29
Influence of glyphosate acid, glyphosate IPA salt, Roundup and POEA on growth of bacterial cultures ... 29
Commensal bacteria ................................................................................................................................ 29
Potential pathogens ................................................................................................................................ 30
Influence of glyphosate IPA salt, Roundup and POEA on gut microbiota from pigs and cows ................... 32
Pigs ........................................................................................................................................................... 32
Cows ........................................................................................................................................................ 35
Influence of glyphosate IPA salt on growth of L. sobrius in stomach content from pigs ............................ 37
Discussion ........................................................................................................................................................ 38
Influence of glyphosate acid, glyphosate IPA salt, Roundup and POEA on growth of bacterial cultures ... 38
Objective 1 ............................................................................................................................................... 38
Objective 2 ............................................................................................................................................... 39
Influence of glyphosate IPA salt, Roundup and POEA on gut microbiota from pigs and cows ................... 41
Objective 3 ............................................................................................................................................... 41
Influence of glyphosate IPA salt on growth of L. sobrius in stomach content from pigs ............................ 45
Objective 4 ............................................................................................................................................... 45
Comparisons of the three different experiments ....................................................................................... 45
Additional work, not included in the thesis ................................................................................................ 46
Conclusion ....................................................................................................................................................... 47
Perspectives and future considerations .......................................................................................................... 49
References ....................................................................................................................................................... 50

1
Theory
Introduction
N-(phosphonomethyl)glycine, commonly known as Glyphosate, is a broad-spectrum, nonselective, systemic
herbicide (Franz et al., 1997). As an analogue of the amino acid glycine, glyphosate can exist in different ionic
states, depending on pH (Chamberlain et al., 1996; Herold et al., 2013). In addition, it can exist in the form of
the acid itself (CAS number 1071-83-6, C3H8NO5P, M = 169.1 g/mol) or formulated as a salt (Budavari, 1996;
Giesy et al., 2000), as isopropylamine (IPA) (CAS number 38641-94-0, C6H17N2O5P, M = 228 g/mol), which is
the one, found in original Roundup products (Malik et al., 1989). In addition to glyphosate IPA salt, Roundup
is made up of water and a surfactant, referred to as inert (Giesy et al., 2000). The surfactant most often used,
is a polyethoxylated tallowamine, POEA (CAS number 61791-26-2)(Giesy et al., 2000).
The herbicidal properties of glyphosate was discovered in the beginning of the 1970’s (Franz, 1974) and since
the commercial introduction of glyphosate formulations in 1974 (Franz et al., 1997; Monsanto, 2015),
glyphosate has become the most dominant herbicide in the world (Grube et al., 2011). The main reasons for
this progress are its broad-spectrum characters and the introduction of glyphosate resistant (GR) crops,
which entered the market in 1996, under the brand ‘Roundup Ready®’ (Monsanto, 2015). The expiry of the
last patent protection for glyphosate took place in 2000, and since, multiple products with glyphosate, as the
active ingredient, have entered the market. This has, in addition to increased use of Roundup Ready crops,
led to an even more widespread use of glyphosate, throughout the years (Duke and Powles, 2008).
Crops treated with glyphosate, either by pre-harvest applications or treatment of GR-crops, during the
growing season, can contain residues that end up in feed (Arregui et al., 2004; Bøhn et al., 2014;
Miljøstyrelsen, 2014; Plantedirektoratet, 2010). As glyphosate exerts the same mechanisms on some
microorganisms, as it does on non-GR-crops (plants), glyphosate residues in feed may have the capacity to
affect gut microorganisms when ingested by livestock and, potentially, reaching critical levels in the gut.
Previous studies have found that glyphosate may affect the growth characteristics of dominating
microorganisms in gut of monogastrics and ruminants, in potential favor of pathogenic bacteria, affecting
health parameters negatively (Ackermann et al., 2015; Krüger et al., 2013b; Shehata et al., 2013a; Shehata et
al., 2013b). These studies have typically worked either with glyphosate acid or with Roundup.
In the beginning of the present study, we overlooked the fact that commercial Roundup formulations
typically contain glyphosate formulated as an IPA salt, instead of the acid itself; a mistake, we have observed
other researchers to have made as well. However, do to this misunderstanding, both glyphosate acid and
glyphosate IPA salt have been included and tested in the present study.
The aim of the project was to compare the effects of different chemical formulations of glyphosate
(glyphosate acid, glyphosate IPA salt and Roundup) as well as the POEA surfactant, and to investigate, at
which concentrations of these formulations, in vitro growth of gut bacteria was affected, if affected at all. In
addition, effects of glyphosate IPA salt, Roundup and POEA, added to stomach, cecum and colon contents
from slaughter pigs and rumen contents from dairy cows, were investigated. Here, the possible in situ effects
on the microbiota composition and activity were measured, by analyzing changes in bacterial cell counts,
production of volatile fatty acids (VFA), changes in pH and production of methane (rumen content only).
Finally, effects of glyphosate IPA salt on growth of a Lactobacillus sobrius strain, inoculated into stomach

2
content (pH=5), was studied. This last part was included to investigate whether glyphosate IPA salt had any
effects on bacterial growth, independent of pH in solution.
The first part of this thesis comprises a literature review. I have intended to give an overview of the working
mechanisms behind glyphosate; how and why it has succeeded to be the most used herbicide in the world
and which challenges, spraying possibly can create. Subsequently, the experimental part follows, and even
though the focus here is narrower, compared to the focus in the literature review, I chose to include a
relatively broad spectrum of literature, to put my research objectives into a broader context.
Working hypotheses
1. Glyphosate (Roundup) will suppress growth of gut bacteria
2. Different gut bacteria will have different susceptibilities towards glyphosate (Roundup)
3. Glyphosate (Roundup) will change fermentation pattern in the gut of pigs and cows
4. The effect of glyphosate will depend on the chemical formulation (acid, salt, mixture and surfactant)
Objectives
To investigate if glyphosate acid, glyphosate IPA salt, Roundup and POEA inhibits bacterial growth.
To investigate if inhibiting effects of glyphosate acid, glyphosate IPA salt, Roundup and POEA differs between
commensal - and potential pathogenic bacteria.
To investigate if glyphosate IPA salt, Roundup and POEA show any effects on the composition and activity of
gut microbiota, when added to stomach, cecum and colon content from slaughter pigs and to rumen content
from dairy cows.
To investigate if growth of a strain of Lactobacillus sobrius in stomach content is affected by glyphosate IPA
salt, when pH is held constant (pH=5).

3
Glyphosate formulations
Glyphosate exists as an acid or in formulation with different salts (Budavari, 1996; Giesy et al., 2000) and
glyphosate IPA salt is the one, primarily used in formulations (Malik et al., 1989). In glyphosate acid,
intermolecular hydrogen bonding between -OH and phosphono oxygens, and between -NH and phosphono
oxygens, creates a strong crystalline matrix, leaving the compound relatively insoluble in water (Knuuttila
and Knuuttila, 1985; Shoval and Yariv, 1981). At 25° the solubility is only 12 mg/ml (acid) (Budavari, 1996)
whereas glyphosate IPA salt has a solubility of 900 mg/ml. Here, hydrogen bonding only forms between three
IPA groups (NH4+) and the phosphono oxygens (PO3
2-), leaving the compound less resistible to breakage and
therefore more soluble in water (Shoval and Yariv, 1981). This is the main reason why, glyphosate is
formulated as the IPA salt in Roundup.
Glyphosate is the active ingredient (a.i) in Roundup (Franz et al., 1997). When comparing the effect of
different glyphosate-products, acid equivalent (a.e.) is more useful, than a.i. Active ingredient includes the
weight of the salt, formulated with glyphosate acid, and as the salt does not have herbicidal activity, active
equivalent only includes the amount of glyphosate, present as acid. EQ1, outlines the relationship between
glyphosate acid and glyphosate IPA salt:
EQ1: a. i. = Molecular mass of the acid−1
Molecular mass of the salt∗ 100 =
169−1
228∗ 100 = 74 %
As an example (EQ2), it is given on the product sheet of Roundup® 2000 that the product contains 400 g/L of
glyphosate acid and 541 g/L of glyphosate IPA salt. This is consistent with EQ1, as the amount of glyphosate
acid can be calculated based on the amount of glyphosate IPA salt:
EQ2: 541g/L ∗ 0.74 = 400 g/L
In general, the formulations contains different concentrations of the IPA salt, even though 360 g/L (a.e.) is
one of the most frequently used (Giesy et al., 2000).
When I refer to glyphosate concentration, throughout the rest of the literature review, as well as the
experimental part, concentrations are given as mg/ml glyphosate acid equivalent, no matter which
compound, glyphosate acid originates from. This eases comparisons.
The chemical properties of glyphosate
Glyphosate is an amphoteric molecule, with a complex ionization pattern. It has four ionizable functional
groups; pKa1 = 0.8 (1st phosphoric), pKa2 = 2.3 (carboxylate), pKa3 = 6 (2nd phosphoric) and pKa4 = 11 (amine),
see Figure 1 (Chamberlain et al., 1996; Herold et al., 2013).

4
Figure 1. Structures and pKa values of glyphosate, modified after Chamberlain et al. (1996); Sprankle et al. (1975)
In its solid, crystalline state, glyphosate exists as a zwitterion (Knuuttila and Knuuttila, 1985). Here, one
proton from the 1st phosphonic group is dissociated and will associate with the extra proton on the amine
group, creating a dipolar molecule. This is evident between pKa 0.8 and 2.3 and as the overall charge of the
molecule equals zero, this is the most stable form, in which glyphosate can exist, see Figure 2.
Figure 2. Structure formula of glyphosate acid (zwitterion). Modified after Chamberlain et al. (1996).
In contrast to the acid, the most stable form of glyphosate IPA salt is present between pKa 2.3 and 6. Here,
the overall charge is zero, and the amine group in IPA will associate with the phosphono group in glyphosate,
see Figure 3.
Figure 3. Structure formula of glyphosate IPA salt. Modified after Chamberlain et al. (1996); Shoval and Yariv (1981)
Surfactants in Roundup
As mentioned, POEA is the surfactant, most used in Roundup, Figure 4 (Giesy et al., 2000). POEA is composed
of tallowamine and two chains of ethoxylate (di-ethoxylates) and is characterized based on the average
oxide/tallowamine ratio (Brausch and Smith, 2007; Mesnage et al., 2013). Tallowamine is a derivative of
tallow, containing a complex mixtures of different fatty acids. These include oleic acid (37-43%), palmitic acid
(24-32%), stearic acid (20-25%), myristic acid (3-6%) and linoleic acid (2-3%) (Diamond and Durkin, 1997). The

5
solubility properties of POEA depends on the length of the polyethylene segment, and the compound is
soluble in water when 12-15 ethylene units are added (Budavari, 1996).
Figure 4. Structure of POEA. The sum of x+y gives 15, and R is a mix of C16 and C18 alkyl and alkenyl chains. After Ahle (1985); Graham
et al. (2006)
Usually, POEA constitutes 15 %, or less, of the formulations, corresponding to 150 g/L (Giesy et al., 2000;
Sawada et al., 1988).
Herbicidal mechanism of glyphosate
In 1980, (Steinrucken and Amrhein) discovered that glyphosate is a potent inhibitor of 5-
enolpyruvylshikimate-3-phosphate synthase (EPSPS) (EC 2.5.1.19), Figure 5. EPSPS, catalyses the reversible
formation of 5-enolpyruvylshikimate-3-phosphate (EPSP) and inorganic phosphate from
phosphoenolpyruvate (PEP) and shikimate-3-phosphate (S3P) (Levin and Sprinson, 1964). In plants, EPSPS is
a nuclear-encoded enzyme, located in the chloroplasts (Della-Cioppa et al., 1986) where it catalyses the sixth
(penultimate) step in the shikimate-pathway, leading to the formation of chorismate (Bentley, 1990).
Chorismate is the precursor of the aromatic amino acids; phenylalanine, tyrosine and tryptophane, Figure 5
(Doy and Gibson, 1961; Gibson and Jackman, 1963; Gibson and Gibson, 1962; Gibson et al., 1962), but also
of vitamins K1 and B9 and the plant hormone salicylic acid, as reviewed by Maeda and Dudareva (2012). In
higher plants, the aromatic amino acids are used as precursors of secondary metabolites, which makes up a
substantial part of the total dry weight of the plant (Herrmann, 1995). According to Maeda and Dudareva
(2012), tryptophan is a precursor of alkolids, phytoalexins, indole glucosinolates and auxin. Isoquinoline
alkaloids, pigment betalains and quinones can be synthesized from tyrosine, and phenylalanine is the
common precursor for phenolic compounds, as flavonoids, condensed tannins and lignin. By inhibiting EPSPS,
the plant will no longer be able to synthesize the aromatic amino acids and secondary metabolites, all used
for growth and, consequently, the plant will die.
The shikimate-pathway is only present in plants, fungi and bacteria and not in animals and insects (Bentley,
1990; Franz et al., 1997; Herrmann, 1995; Kishore and Shah, 1988; Padgette et al., 1995a). In addition there
is evidence that apicomplexan parasites, like Toxoplasma gondii, Plasmodium falciparum (malaria) and
Cryptosporidium parvum (Roberts et al., 1998) and Archaea also possesses the shikimate pathway (Bult et
al., 1996; Daugherty et al., 2001; Graham et al., 2001). As the pathway is absent in animals, they need to
obtain the essential aromatic amino acids through their diet (Franz et al., 1997; Herrmann, 1995; Kishore and
Shah, 1988; Padgette et al., 1995a), although tyrosine can be synthesized from phenylalanine (Herrmann,
1995).

6
Figure 5. The shikimate pathway that leads to the formation of aromatic amino acids and the inhibition of EPSPS by glyphosate
(Pollegioni et al., 2011).
In addition to the inhibition of EPSPS, glyphosate has also been shown to target other plant enzymes,
indicating that it might have several sites of action (Bode et al., 1984; Ganson and Jensen, 1988; Lee, 1980;
Rubin et al., 1982). However, glyphosate is not shown to inhibit other PEP dependent reactions (Anton et al.,
1983; Steinrucken and Amrhein, 1984).
5-enolpyruvylshikimate-3-phosphate synthase
To understand how glyphosate exerts its effect on EPSPS, the three dimensional structure of the enzyme,
with and without ligand, have been studied. Stallings et al. (1991) were the first to elucidate the three
dimensional crystal structure of unliganded EPSPS (from E. coli). In short, they revealed a unique protein fold
with two globular hemispheric domains, connected to each other by two crossover chain segments. Some
years later, Schönbrunn et al. (2001) were able to identify the structure of E. coli EPSPS in its liganded form,
especially investigating the binding of S3P and glyphosate. They found, as already stated, that EPSPS is a two-
domain enzyme, but that binding of the ligand S3P, induces a conformational change in the enzyme, turning
it into a closed state. The active site is formed in the interdomain cleft, indicative of an induced-fit
mechanism. The closure leads to an accumulation of positive charges in the cleft, which will attract negatively

7
charged molecules to the active site, and glyphosate can bind. This makes glyphosate an uncompetitive
inhibitor with respect to S3P (Boocock and Coggins, 1983), as binding of S3P is a prerequisite for binding of
glyphosate (Schönbrunn et al., 2001). Schönbrunn et al. (2001) also concluded that glyphosate and PEP share
the same binding site, which is supported by Eschenburg et al. (2002). This makes glyphosate a competitive
inhibitor with respect to PEP (Anton et al., 1983; Boocock and Coggins, 1983).
The main target of glyphosate – the plant
As the main target of glyphosate is located intracellularly, glyphosate has to enter the cells of the plant, to
exert its mechanisms. In general, glyphosate is a polar, hydrophilic molecule, as reflected in its low log Kow
values, which Chamberlain et al. (1996) found to range between -3.39 and -4.85, depending on pH level.
Before spraying, a tank mix is prepared. As mentioned, glyphosate is mainly formed as the IPA salt in
formulations, due to its higher solubility in water (Shoval and Yariv, 1981). Glyphosate IPA salt is most stable
between pH 2.3 and pH 6 (see Figure 3), as the overall charge of the compound is zero. Here, glyphosate IPA
salt exist as two separate ions, weakly held together by hydrogen bonds (Shoval and Yariv, 1981). Therefore,
acidic conditions are most suitable in tank mixes.
Plant cells are covered by a cell wall, composed of cellulose, hemicellulose, pectin and, in many cases, also
lignin. Beneath is a single plasma membrane, surrounding the cytoplasm. The plasma membrane is a lipid
bilayer, embedded with proteins. It works as a semipermeable barrier between inner and outer surface of
the cell, and thereby regulate entry and exit of cells (Raven et al., 2005). The outermost cells of the plant
(upper epidermis) are covered with a waxy protection layer (cuticle). As mentioned, surfactants are a part of
Roundup and they aid in penetration of the waxy cuticle (Franz et al., 1997; Giesy et al., 2000). They reduce
surface tension of spray droplets on the plant (Relyea, 2005), enlarging the area of pesticide coverage and
thereby the penetration of glyphosate, through the plant surface. By improving retention of spray droplets,
they also minimize run-off (McCloskey). These properties enhances the effect of glyphosate, as its
penetration of the cuticle otherwise will be inadequate (Tsui and Chu, 2003).
After penetration of the cuticle, glyphosate is translocated via the vascular tissue (Raven et al., 2005)
throughout stems, leaves and roots of the entire plant; therefore the definition as a systemic herbicide (Franz
et al., 1997). Pline et al. (2002) suggested an increased sensitivity to glyphosate in reproductive tissue over
vegetative, as the shikimate pathway is most active in the growing parts of the plant. Therefore translocation
of glyphosate to the growing points is vital, as glyphosate blocks amino acids synthesis in the fast growing
parts of the plant, followed by inhibition in the other tissues (Pline et al., 2002).
Before glyphosate can inhibit the shikimate pathway, it has to enter the cells, by crossing cell wall and plasma
membrane. Until now, both passive diffusion and active transport have been suggested to be of importance
(Hetherington et al., 1998). A phosphate carrier is expected to be involved in the active transport, and
especially at low concentrations, the phosphate carrier might play a crucial role (Hetherington et al., 1998).
In addition, passive diffusion becomes more prominent at higher glyphosate concentrations (Denis and
Delrot, 1993).
Even though, glyphosate IPA salt exist as two separate ions in aqueous solution, they form an ion-pair, when
entering a lipohilic environment, as the plasma membrane. This facilitates an easier entrance, due to the
overall neutral charge (Krogh, 2016). When glyphosate IPA salt has entered the cell, it must be assumed that
the ion-pair splits again. In higher plants, the cytoplasm is slightly alkaline (7.4 – 7.5) (Gout et al., 1992),

8
leaving glyphosate IPA salt more unstable, due to the anionic character (see Figure 1). In addition to
glyphosate IPA salt, some glyphosate might also enter as the acid.
When glyphosate has exerted its effect, the plant dies slowly, and the process takes several days (Kishore
and Shah, 1988).
Degradation of glyphosate and subsequent effects on soil
One of the reasons why glyphosate is toxic to plants is that they have a limited ability to neutralize or degrade
glyphosate, compared to the soil environment (Franz et al., 1997).
When glyphosate enters soil, it can either be bound to soil particles, leached to subsoil or be degraded
through two principal pathways, mainly by microorganisms, Figure 6 (Borggaard and Gimsing, 2008). In the
first pathway, glyphosate is cleaved by glyphosate oxidoreductase, yielding glyoxylate and AMPA
(aminomethylphosphonic acid). AMPA can further be metabolized to inorganic phosphate and methylamine,
which ultimately is converted to CO2 and NH3, supporting growth of microorganisms. The second pathway
leads to the cleavage of inorganic phosphate from glyphosate, by C-P lyase, and sarcosine is formed.
Sarcosine can be further degraded, yielding formaldehyde and glycine, and both these products can also be
converted, and eventually utilized, by microorganisms (Barry et al., 1992; Borggaard and Gimsing, 2008; Duke
et al., 2012).
Figure 6. Microbial degradation of glyphosate. Modified after Borggaard and Gimsing (2008); Duke et al. (2012).
Even though glyphosate can be used as a nutrient source for some microorganisms (Partoazar et al., 2011;
Sviridov et al., 2015), additon of varying rates influences the soil microflora in different ways, as glyphosate
have shown toxicity towards some groups of microorganisms. Temporary effects were reported by Mijangos
et al. (2009), who observed increased activity and functional diversity within the culturable portion of
heterotrophic soil bacteria, after single glyphosate additions. Those short term effects might be due to a
rapid metabolization of glyphosate (Mijangos et al., 2009; Ratcliff et al., 2006). Lane et al. (2012) found
increased microbial activity in soils with a history of glyphosate applications, with additional glyphosate
applications, opposed to microbial activity in soils, with no prior glyphosate history. This was confirmed by
Dick and Quinn (1995), who compared a soil with no history of glyphosate applications, with a soil treated
with glyphosate, the three previous years. The latter contained approximately a 10 times higher microbial

9
population, even though it had a lower diversity of colonies. In contrast, Hart and Brookes (1996), observed
no long term differences in microbial biomass and activity in a soil, with 19 years of annual glyphosate
applications prior to the experiment. Mekwatanakarn and Sivasithamparam (1987) found glyphosate to
reduce bacterial numbers, at the glyphosate rate recommended for field use. This is supported by Kremer et
al. (2005), which observed a decrease in bacterial growth in root exudates from glyphosate-treated soybean
plants. However, Stratton and Stewart (1992) did not find any negative effects on numbers of bacteria, fungi
and actinomycetes, when applied at recommended rates.
Production of glyphosate resistant (GR) crops
To overcome the herbicidal effects of glyphosate on plants, different mechanisms for introduction of
glyphosate resistance (GR) has been explored; overexpression of EPSPS, detoxification of glyphosate and the
introduction of an insensitive form of EPSPS (Dill, 2005).
So far, overexpression of EPSPS has not generated any commercially available GR plants (Dill, 2005), but Shah
et al. (1986) succeeded in producing a Petunia plant, expressing higher levels of EPSPS, compared to normal,
which were able to tolerate increased levels of glyphosate. However, the growth rate was impaired and the
modified plant was not competitive compared to those, exhibiting the wild type.
Detoxification of glyphosate can be achieved by insertion of genes, which degrades glyphosate (Dill, 2005).
In combination with glyphosate insensitive EPSPS, the insertion of GOX (glyphosate oxidoreducatase) into
rape seed, has produced a commercial GR line. The detoxification of glyphosate itself, cannot provide
adequate resistance (Dill, 2005).

10
The method used to create the majority of GR crops on the market, is the insertion of a gene coding for a
naturally GR EPSPS (Dill, 2005). There have been several attempts to produce such, and some are summarized
in box below.
Agrobacterium sp. strain CP4 is naturally resistant towards glyphosate, and the majority of all commercially
GR crops, are produced from the insertion of EPSPS derived from Agrobacterium sp. strain CP4 (Barry et al.,
1997; Padgette et al., 1995a). EPSPS from Agrobacterium sp. strain CP4 was found from a screen of
microorganisms, and it was observed that the enzyme had favorable kinetics with respect to glyphosate and
PEP, indicating that EPSPS binds well to PEP (Padgette et al., 1995b). The reason for the insensitivity of CP4
EPSPS to glyphosate, is attributed to a single substitution from glycine to alanine in position 100 (Funke et
al., 2006). The methyl group of alanine interferes with one of the oxygen atoms in the phosphonate group of
glyphosate, which will result in a conformation change of glyphosate (Eschenburg et al., 2002; Funke et al.,
2006). Only the extended form of glyphosate has an inhibitory effect on EPSPS (Funke et al., 2006; Park et
al., 2004; Schönbrunn et al., 2001).
Sost and Amrhein (1990) found that a Gly96 to Ala substitution in the EPSPS from Klebsiella pneumonia,
reduced its affinity towards glyphosate. Later Eschenburg et al. (2002) revealed that it is the interference
between glyphosate and the additional methyl group of Ala96, protruding into the active site of the
enzyme, which confers the resistance, as glyphosate only interacts efficiently with EPSPS in its extended
conformation (Funke et al., 2006). As PEP is a smaller molecule than glyphosate, it can still fit in the
narrowed binding site. However, its translational freedom to S3P and the residues in the binding site
becomes limited, and therefore the affinity towards PEP also decreases (Eschenburg et al., 2002). The
affinity towards S3P remains unaltered (Eschenburg et al., 2002). In addition, one single site mutations
have conferred resistance towards glyphosate in Salmonella enterica Typhimurium. As in Klebsiella
pneumonia, it is one amino acid substitution, which is responsible for the resistance, although it is a
change from Pro to Ser at position 101, instead (Stalker et al., 1985). Pro101 is not located close to the
active site, but the alteration to Ser will change the orientation of Gly96 and Thr97 in the active site,
narrowing the binding site for glyphosate (Healy-Fried et al., 2007). Since PEP and glyphosate share the
same binding site, it is difficult to obtain an enzyme, which will bind PEP and exclude glyphosate too such
an extent that it will result in commercial levels of glyphosate resistance in normal developing plants.
According to Dill (2005) none of the singlesite mutations renders the enzyme with adequate kinetic
properties, even though they are advantageous. However, more favorable kinetic characteristics, were
observed for multisite mutations. One multisite mutation is used for the production of the commercially
available GR maize line (Lebrun et al., 2003), known as GA21 (Sidhu et al., 2000). The maize EPSPS is a
transgene, with the substitution of threonine with isoleucine at position 102 and proline with serine at
position 106 (Lebrun et al., 2003). Funke et al. (2009) studied the kinetic and structural effects of this
transgene EPSPS, using E. coli as a model. The two mutations induced the conformational changes
needed, to impair efficient glyphosate binding. As with the single site mutation P101S, the orientation of
Gly96 changes, interfering with binding of glyphosate. Ile97 (Thr97 before) will point away from the
binding site, facilitating the effective binding of PEP (Funke et al., 2009). The mutated EPSPS from E. coli
showed high resistance towards glyphosate while still maintaining high affinity for both PEP and S3P
(Funke et al., 2009), which is the reason for its commercial use.

11
The success of glyphosate
As mentioned above, glyphosate is a broad-spectrum systemic herbicide (Franz et al., 1997). It is used post-
harvest to target annual and perennial weeds in subsequent crops, but can also be used pre-harvest, as a
desiccating agent. Here, it acts to secure a uniform ripening of the crop, due to uneven soil conditions and
differences in microclimate (Miljøstyrelsen, 2014). According to Knowledge Center for Agriculture (SEGES)
and Institute for Agroecology, Aarhus University, the amount of glyphosate, used as a pre-harvest agent, has
increased since 2002, but to an extent which is not exactly known (Miljøstyrelsen, 2014). However, Djursing
(2013) estimates that about 10 % of the Danish areal with cereals are treated with glyphosate before harvest,
while it is 25 % of the areal with rapeseed. When used as a pre-harvest agent, glyphosate cannot be used
later than 10 days before harvest.
In addition to the use of glyphosate, in the traditional manner, introduction of glyphosate resistant (GR) crops
have changed the market considerably. One of the main reasons why adoption of GR crops have been faster,
than any other technology seen in agriculture (Sankula, 2006), is attributed to the substitution of several
different herbicides with glyphosate. Planting GR crops allow glyphosate to be spread during most of the
growing season, instead of limiting the application to pre-plant – and post-harvest periods. This makes it
possible to target weeds, as soon as they emerge from the soil and allows several applications throughout
the growing season (Benbrook, 2012; Waltz, 2010). In addition, less or no tillage is needed (Gianessi, 2005),
and combined, these factors create greater flexibility in timing of applications, improves soil
structure/moisture, reduces the risk of runoff and generally makes the control of a broad range of weeds,
including both annual grasses, broadleaves and perennials, more effective (Gianessi, 2005). Moreover,
adopting a less – or none tillage procedure is a less expensive alternative for weed control in comparison
with conventional tillage (e.g. reduces fuel and labor costs) (Sankula, 2006).
Even though growing GR crops have some advantages, compared to growing conventional crops, the strong
reliance on glyphosate have increased the resistant weed population (Benbrook, 2012). Until now,
glyphosate resistance have been reported in 32 weed species throughout the world, all emerging from 1996
and onwards, where GR crops were introduced (Heap, 2015). Unfortunately, this has led to a further increase
in the use of glyphosate worldwide, both in the context of more frequent spreading, but also with higher
doses, which further accelerates the emergence of resistant weeds, creating a vicious circle (Benbrook, 2012;
Binimelis et al., 2009).
Table 1 shows the consumption of glyphosate in the US, from 1987 and onwards. Since 2001, glyphosate has
been the most used active ingredient in agriculture (Grube et al., 2011). This is attributable to both the
invention and use of GR crops, but also to the increasing incidences of resistant weeds, and a steady decline
in cost of glyphosate, since the patent expired in 2000 (Duke and Powles, 2008).

12
Table 1. The consumption of glyphosate in the US, measured by million pounds of active ingredient, from 1987 to 2007. Modified
after Grube et al. (2011) and Kiely et al. (2004) (estimates).
The relationship between the use of glyphosate and GR crops becomes apparent, when looking at Figure 7,
which deals with one of the most widespread GR crop in the world, soybeans. Only few percentages of all
soybeans planted in 1996, were glyphosate resistant. The amount increased to almost 70% in 2001 and has
remained stable since 2007, representing approximately 93 % of all soybeans planted. The numbers
presented for the U.S. in Fejl! Henvisningskilde ikke fundet., corresponds to GM soybeans, and not directly
to GR. However, according to several references, including Duke and Powles (2009), this is practically the
same. As also presented in Fejl! Henvisningskilde ikke fundet., it is not only in the U.S., GR soybeans play a
crucial role. In Argentina, the first GM crop to be adopted was GR soybeans. The event took place in 1996
and since then, the area planted with GR soybeans has expanded dramatically - the increase has even
exceeded the increase observed in the U.S. Now, almost all the soybeans grown in Argentina are GR
soybeans. The same trend applies for GR soybeans in Brazil, as they occupied 92,4 % of the total soybean
area in 2013 (ISAAA, 2014). At the same time, these three countries are the leading producers of soybeans
in the world, as almost 80 % of the world’s production originates from here (FAO, 2015).
Figure 7. Adoption rate of GM soybeans in the US (according to Duke and Powles (2009), this corresponds to GR soybeans)
and adoption rate of GR soybeans in Argentina. Data from Duke and Powles (2009); USDA (2014) and ArgenBio (2015).
The EU has strict GM regulations (Davison, 2010) and only one GM maize line (MON810) and one GM potato
line (Amflora) are approved for commercial cultivation (Commission, 2011). Therefore, the use of glyphosate
Year Glyphosate (million pounds of a. i.)
1987 -8
1997 34-38
1999 67-73
2001 85-90
2003 128-133
2005 155-160
2007 180-185

13
in Denmark cannot be ascribed to the cultivation of GR crops, but agriculture is still the main responsible for
the consumption. Glyphosate is mainly used for weed killing, but a substantial amount is also used pre-
harvest, as a desiccating agent (Djursing, 2013). According to Figure 8, there has been a steady increase in
the sale of glyphosate in Denmark since 1993, with few exceptions, e.g. 2009. According to Miljøstyrelsen
(2013), the enhanced regulation of other herbicides has led to this relative abrupt increase in the sale of
glyphosate. In 2012, glyphosate constituted 23% of the total amount of active ingredients sold for agricultural
production, only surpassed by prosulfocarb with 34% (Miljøstyrelsen, 2013).
Figure 8. The amount of glyphosate, as an active ingredient, sold from 1991 to 2012. Modified after Middeldatabasen/SEGES (2015);
Miljøstyrelsen (2008, 2013)
Even though no GM crops are produced in Denmark, the adoption of, especially, GR soybeans in the rest of
the world, is of importance for Danish agriculture. Since 2000, the Danish soy import has exceeded 1,400,000
tons each year, with an import of 1,560,000 tons in 2014. The vast majority originates from countries, using
GR soybeans (Statistikbanken, 2015), which, together with glyphosate as a desiccating agent, leaves the
discussion of glyphosate residues in feed important in Danish agriculture.
Possible pesticide residues in crops treated with glyphosate
When applying glyphosate to crops, there is a risk that residues can end up in the feed. A pesticide residue is
defined as the combination of a pesticide and its metabolites, degradates and other transformation products
(FAO, 2009). To secure the lowest possible consumer and animal exposure to pesticides, Maximum Residue
Levels (MRL) are established. MRL is defined as the upper legal limits of a pesticide, allowed in food or feed,
expressed as mg/kg (EFSA, 2015b). MRL are based on toxicology data from animals, and toxicity on
microorganisms has not been included. MRL for glyphosate, determined by the EU and FAO/WHO, appears
from Table 2.

14
Table 2. MRL levels of glyphosate in different cereals and oilseeds (Codex Alimentarius, 2012; Commission, 2015)
As seen in Table 2Fejl! Henvisningskilde ikke fundet., the allowed MRLs differs between the different
feedstuffs and in addition the allowed limits differs between EU and FAO/WHO. Generally, EU are more
cautious in their determination than FAO/WHO.
Several investigations have sought to examine, whether MRL are met in different feedstuffs. Unpublished
data from Germany found glyphosate levels of 0.4 – 0.9 mg/kg in poultry and cattle feed samples (Shehata
et al., 2014), which is well below the MRL (Table 2). Arregui et al. (2004) detected AMPA residues in both
leaves and grains from GR soybeans and reported glyphosate residues ranging from 1,9 to 4,4 mg/kg in stems
and leaves, and from 0,1 to 1,8 mg/kg in grains, all below the MRL. The highest concentrations were obtained
after several glyphosate treatments, during the growing season, but a relationship between application rate
and residue level could not be established (Arregui et al., 2004). Bøhn et al. (2014) compared 31 soybean
batches, almost equally distributed between GR soybeans (10), conventional soybeans (10) and organic
soybeans (11), with respect to glyphosate and AMPA content. Bøhn et al. (2014) were able to show that only
GR soybeans contained residue levels of AMPA and glyphosate, with a mean value of 3.3 and 5.7 mg/kg,
respectively.
In a Danish survey from 2009, 20 batches of imported soy products were analyzed for glyphosate content.
16 were manufactured from GM soy, and all shipments originated from South America, with different export
countries involved (Plantedirektoratet, 2010). The batches were distributed between soy hulls, soybeans and
soybean meal, and glyphosate residues were only detected in GM - soy hull batches (4). In three of the four
batches, the residue levels exceeded the MRL of 20 mg/kg for soybeans (24,7 – 26,7 mg/kg) (Commission,
2015). However, MRL is set for soybeans and not for the processed products (Commission, 2015) and it
cannot directly be transferred to apply for soy-hulls (Plantedirektoratet, 2010).
Spraying pre-harvest can also influence the residues levels of glyphosate, found in seeds, after harvesting. As
application has to take place when the crop is mature, almost no translocation will occur in the plant.
Therefore residues, if any, will be present on the surface of the plant (Cessna et al., 1994). Miljøstyrelsen
(2014) have conducted a survey, investigating different feed samples where glyphosate has been used pre-
harvest. The results are presented in Table 3 and even though glyphosate was found in a large percentage of
the samples, none of these exceeded the MRL. However there is a tendency that pre-harvest treatment of,
especially barley, has increased during the last years, with the maximum residue level found to be 13 mg/kg.
MRL of glyphosate (mg/kg) (EU) MRL of glyphosate (mg/kg) (FAO/WHO)
Barley 20 30
Oat 20 30
Rye 10 30
Wheat 10 30
Maize/corn 1 5
Rapeseeds/canola seeds 10 30
Soybeans 20 20

15
Table 3. Overview of glyphosate residues in random samples from Danish produced barley and wheat (spelt and triticale included)
for animal feed. The quantification limit was set to be approx. 0,1 mg/kg. The survey is conducted by the Danish Veterinary and
Food Administration, Modified after Miljøstyrelsen (2014).
Summarizing the abovementioned results, glyphosate residues can be present in feed, regardless of whether
glyphosate is used in GR crops or as a desiccation agent in non-GR crops, even though the levels differs
between different investigations.
Toxicity of glyphosate The absence of the shikimate pathway in mammals, might explain the relatively low toxicology of glyphosate
towards mammals (Kier, 2015; Smith and Oehme, 1992; Williams et al., 2012; Williams et al., 2000). However,
glyphosate might target other pathways and toxicity of POEA has also gained more interest.
In a review by Samsel and Seneff (2013) it is stated that glyphosate has the ability to suppress cytochrome
P450 enzymes. It is a large group of enzymes playing crucial roles in primary and secondary metabolism
(Nelson et al., 1996). Lamb et al. (1998) discovered that glyphosate can inhibit plant cytochrome P450
(CYP71B1) and several studies, focusing on human cell lines, found glyphosate, and especially glyphosate
formulations, to be the disruptor of Aromatase cytochrome P450 activity (Gasnier et al., 2009; Richard et al.,
2005; Simpson et al., 2002; Simpson et al., 1994). Glyphosate decreased the hepatic levels of cytochrome
P450 in a study where rats were treated with glyphosate for two weeks (Hietanen et al., 1983) and Paganelli
et al. (2010) hypothesized that morphological changes in frog embryos after glyphosate addition, were
associated with inhibition of CYP26 enzymes (belongs to P450 superfamily).
Glyphosate is also patented as a broad-spectrum chelator, complexing divalent cationic nutrients (Fon and
Uhing, 1964), enabling it to bind a wide range of minerals. The very low levels of Mn and Co, found in the
urine samples from dairy cows, containing varying amounts of glyphosate, could possibly be ascribed to this
chelating effect. However, Cu and Se were within the reference level, not influenced by the presence of
glyphosate (Krüger et al., 2013a).
Glyphosate as a carcinogen?
In addition to the questioning about which components lead to the toxicity of glyphosate formulations, if
toxicity is observed at all, there has been some controversy about the carcinogenic potential of glyphosate.
In 1985, The United States Environmental Protection Agency (U.S. EPA) classified glyphosate as possibly
carcinogenic to humans (Group C), based on tumor studies in mice (EPA, 1985). However, when the study
was reevaluated by US EPA in 1991, the classification of glyphosate was changed to evidence of non-
carcinogenity in humans, instead (Group E) (IARC, 2015). In 2015, the International Agency for Research
Against Cancer (IARC), reclassified glyphosate as probably carcinogenic to humans (Group 2A) (Guyton et al.,
Crops Determination of glyphosate 2008 2009 2010 2011 2012 2013
Barley Number of samples 14 20 22 21 33 23
Samples with glyphosate present (%) 7 15 45 48 30 77
Highest level measured (mg/kg) 0.4 1.6 7.1 4.6 8.1 13
Average level (mg/kg) 0.03 0.17 0.79 0.5 0.85 1.86
Wheat, triticale, spelt Number of samples 13 15 22 12 22 15
Samples with glyphosate present (%) 23 27 36 58 32 47
Highest level measured (mg/kg) 0.9 0,8 3,6 1.2 4.1 2
Average level (mg/kg) 0.15 0.1 0.28 0.3 0.51 0.26

16
2015). This category is used for pesticides showing limited evidence of carcinogenity in humans (non-Hodgkin
lymphoma) but sufficient evidence in animals (Guyton et al., 2015; IARC, 2015). After a peer-review of the
carcinogenic potential of glyphosate, the European Food Safety Authority (EFSA) concluded that glyphosate
is unlikely to pose a carcinogenic threat to humans, in contrast to the evaluation by IARC (EFSA, 2015a).
However, IARC evaluated both glyphosate and glyphosate-formulations, whereas EFSA only included studies,
concerning the active substance, which might influence the outcome of the reviews (EFSA, 2015a).
Toxicity of POEA
The information regarding content of surfactants in Roundup, are usually kept confidential by the
manufacturer. This makes it difficult to determine exactly what is used in Roundup – and thereby to create
sufficient risk assessments. Mesnage et al. (2013) concluded that it was the ethoxylated surfactants of
glyphosate-based formulations that were the active principles of human cell (embryonic, placental and
hepatic) toxicity, with POE-15 displaying the greatest effect. In addition to this finding, all formulations were
more toxic than glyphosate itself. Results from Mesnage et al. (2014) supported these findings, as they found
Roundup to be 125 times more toxic to human cells (embryonic, placental and hepatic), than glyphosate was.
Benachour and Seralini (2009) concluded that surfactants, like POEA, changed the human cell permeability
of three different cell types (umbilical, embryonic and placental) and amplified the toxicity, already induced
by glyphosate, through apoptosis and necrosis. Other experiments sought to test the toxicities on aquatic
organisms. In a study by Folmar et al. (1979), POE-15 had approximately the same acute toxicity towards fish
and aquatic invertebrates, as Roundup had. However, the contribution of glyphosate to the toxicity of
Roundup, only ranged from 29% to 33% (Folmar et al., 1979). Other studies focusing on the aquatic habitat
(different fish species) also concluded that it were the surfactants, which where main responsible for the
toxicity observed (Mitchell et al., 1987; Servizi et al., 1987).
Toxicity of glyphosate towards microorganisms
Like plant cells, almost all prokaryotic cells, are surrounded by a cell wall, and most bacteria are divided based
on a gram – reaction. Gram-positive bacteria are covered by a thick layer of peptidoglycan, which constitutes
up to 90 % of the cell wall. In gram-negative bacteria, only approximately 10 % of the cell wall consists of
peptidoglycan, with the rest being composed of an outer membrane. For all bacteria, a plasma membrane is
located beneath the cell wall (Madigan et al., 2009). As well as for plants, glyphosate has to enter the cells to
exert its effects. To my knowledge, details about transport of glyphosate into bacterial cells are unclear, even
though some of the entrance mechanisms exerted in plants, might be evident. No matter of entrance
method, some studies have found inhibiting effect of glyphosate, towards different kinds of microorganisms.
Three food microorganisms, a fungus (Geotrichum candidum) and two bacteria (Lactococcus lactis subsp.
cremoris and Lactobacillus delbrueckii subsp. bulgaris) are used as starters in the dairy technology. Clair et
al. (2012) found all three microorganisms to be more sensitive to Roundup than to glyphosate alone. In an in
vitro study, the nitrogen fixating bacteria, Azotobacter vinelandii, showed a higher degree of sensitivity
towards glyphosate than Azotobacter chroococcum did (Santos and Flores, 1995). In soil samples, Busse et
al. (2000) observed an increase in total and viable bacteria after glyphosate addition, and especially
Arthrobacter, Pseudomonas, Xanthomonas and Bacillus spp. increased in population. Difference in
glyphosate tolerance between bacteria cultured in soil and in soil-free media might be explained by the
chemical properties of glyphosate. Glyphosate binds easily to soil particles, which leaves the compound
unable to exert its herbicidal effects (Borggaard and Gimsing, 2008; Duke et al., 2012).

17
Tsui and Chu (2003) investigated whether POEA also accounted for the toxicity of Roundup to
microorganisms. Bacteria (Vibrio fischeri) and protozoa (Tetrahymena pyriformis and Euplotes vannus) had
more or less similar sensitivities towards Roundup. For the same species, POEA showed considerably lower
toxicity levels, and generally, glyphosate IPA salt was least toxic.
Based on these findings it become obvious, how important it is to investigate whole Roundup formulations.
In formulations, it is always important to know the individual toxicities of the different compounds – but also
their combined effects, as they might influence each other.
Changes in gut microbiota with respect to glyphosate
Potential levels of glyphosate in gut
As the overall objectives of this study are to investigating possible effects of glyphosate on microorganisms
in livestock, it is useful to calculate, which levels it is likely to obtain in the gut. Two scenarios are presented
below, one for pigs and one for cows.
Numbers presented in Table 4Fejl! Henvisningskilde ikke fundet., are calculated based on feeding pigs a diet
containing only barley or soybean meal. If the soybean meal and barley contain MRL of glyphosate allowed
in the EU (20 mg/kg), the residues in the gut would be between 0.005 and 0.019 mg/ml, based on the
assumptions made.
Table 4. Estimated glyphosate concentrations in the gut of pigs, fed a diet containing the MRL of glyphosate for barley and soybean
meal (20 mg/kg). Calculated by Martin Tang Sørensen.
In addition to Table 4, Table 5 shows which glyphosate concentrations it might be able to obtain in the gut of
cows, assumed that they are fed a diet, where 50 % is composed of soybean meal and barley, and MRL of
glyphosate are present in these feedstuffs. Compared to pigs, lower levels of glyphosate (from 0.0022-0.003
mg/ml) can be present in the different segments in cows, mainly because of the lower percentage of
concentrate in the feed ration.
Table 5. Estimated glyphosate concentrations in the gut of cows. Only half the diet originates from barley and soybean meal,
assumed to contain 20 mg glyphosate/kg. This means that the feed in average will contain 10 mg/kg when ingested. Calculated by
Martin Tang Sørensen
Feed DM1 Glyphosate content3
(pr. kg ingested feed) (pr. kg ingested feed)
Stomach 1.0 kg 4 liter (~25 % DM) 20 mg ~ 0.005 mg/ml
Small intestine (distal part) 0.3 kg 3 liter (~10 % DM) 14 mg ~ 0.005 mg/ml
Colon (feces) 0.15 kg 0.75 liter (~20 % DM) 14 mg ~ 0.019 mg/ml
Gut segmentGut content volume2
(pr. kg ingested feed)
Concentration of glyphosate
acid equivalents (mg/ml)
1Feed DM disappears through the GIT because of degradation and absorption (Anguita et al., 2006; Canibe and Knudsen, 2002)2The DM content in the gut segments is calculated based on results found in Canibe et al. (2005); Canibe and Jensen (2003)3In general it is assumed that glyphosate is not degraded in the GIT, but 30% is absorbed to the blood from the small intestine (Commission,
2002)
Feed DM1 Glyphosate content3
(pr. kg ingested feed) (pr. kg ingested feed)
Forestomach (at the end of the omasum) 0,65 kg 3.4 liter (~19.4 % DM) 10 mg ~ 0.003 mg/ml
Small intestine (distal part) 0.35 kg 3.2 liter (~10.8 % DM) 7 mg ~ 0.002 mg/ml
Colon (feces) 0.30 kg 2.3 liter (~13.3 % DM) 7 mg ~ 0.003 mg/ml
Gut segmentGut content volume2
(pr. kg ingested feed)
Concentration of glyphosate
acid equivalents (mg/ml)
1Feed DM disappears through the GIT because of degradation and absorption (Storm and Kristensen, 2010) 2The DM content in the gut segments is calculated based on results for cows in early lactation (63 days postpartum) found in Andrew et al. (1995)3In general it is assumed that glyphosate is not degraded in the GIT, but 30% is absorbed to the blood from the small intestine (Commission, 2002)

18
Overall, the assumptions made for the scenarios presented in Table 4 and Table 5, are based on feed rations
containing MRL of glyphosate. According to the findings of glyphosate residues in crops, presented earlier,
such levels are uncommon. However, some exceedances are observed (Plantedirektoratet, 2010), but the
risk that all feed ingredients in the ration has the same high level, might be (even) less plausible. In addition,
toxicity margins are included in MRL, meaning that it is not toxic in itself. Another important assumption used
for the calculation, which might be questioned, is that degradation of glyphosate does not take place and
that only 30 % is absorbed (Commission, 2002). Glyphosate might be degraded to AMPA in animals, as
observed in plants (Arregui et al., 2004; Bøhn et al., 2014). If that is the case, the residue levels will change.
So far, the toxicity of AMPA it is not known and therefore this also has to be taken into consideration, when
evaluating the toxic effects of glyphosate residues in livestock.
The effect of glyphosate on poultry microbiota
Shehata et al. (2013b) investigated the impact of glyphosate on pathogenic and commensal microorganisms,
in the gut of poultry, representative of a monogastric animal. The results are presented as the Minimum
Inhibitory Concentration (MIC), which is the lowest concentration of an inhibitory compound needed, to
completely block the growth of a microorganism. As seen in Table 6, most of the pathogenic bacteria were
highly tolerant to glyphosate (Roundup), with MIC values at 5.000 mg/ml (Clostridium perfringens,
Salmonella enterica Enteritidis, Salmonella enterica Gallinarium and Salmonella enterica Typhimurium),
while most of the commensal bacteria were moderate to highly sensitive (Bacillus badius, Bacillus cereus,
Bifidobacterium adolescentis, Enterococcus faecalis and Enterococcus faecium), with MIC values ranging from
0.075 to 0.300 mg/ml. Only few bacteria did not fit this characterization, as Campylobacter coli and
Campylobacter jejuni were highly sensitive (MIC = 0.150 mg/ml), while three Lactobacillus strains were more
resistant than the other beneficials (MIC = 0.600 mg/ml). In addition, Shehata et al. (2014) found MIC values
of glyphosate for B. adolescentis, B. badius and E. faecilis on 0.150, 0.300, and 0.300 mg/ml, respectively,
which all are slightly higher than the numbers presented in Table 6. Based on the two studies, referred to
above, glyphosate has the capability to suppress growth of Bifidobacteria substantially. Bifidobacteria can
create unfavorable conditions for pathogens, as Salmonella in the gut (Bielecka et al., 1998), and, therefore,
a decrease in the population of Bifidobacteria can indirectly increase growth of Salmonella and other
pathogens. The result is a disturbance in the gut microbiota, which possibly can lead to health issues.

19
Table 6. The MIC value of glyphosate on different pathogenic and beneficial bacteria. Based on Shehata et al. (2013b)
Minimum Effect Concentration (MEC) is the lowest concentration of an inhibitor needed to initiate the
inhibition of a microorganism (Arikan et al., 2001) and from data presented by Fredborg et al. (2013) it is
suggested that MEC can be calculated as 1/10 of the MIC value. This means, that the commensal bacteria,
found to be sensitive by Shehata et al. (2013b), already could be negatively affected by concentrations of
glyphosate, ranging from 0.0075 to 0.030 mg/ml, corresponding to a tenth of the MIC values.
The effect of glyphosate on dairy cow microbiota
To investigate the effect of glyphosate on the microbiota in dairy cattle, Ackermann et al. (2015) compared
two diets, one rich in crude fibers and one low in crude-fibers. Glyphosate, at very low concentrations (0.001
mg/ml), decreased the growth of Ruminococcus albus and Ruminococcus flavefaciens substantially (Leschine,
1995). In addition, cell counts of Euryarchaeota were significantly decreased at glyphosate concentrations of
0.01 and 0.1 mg/ml and for Streptococcus spp., a reduction was observed at 0.1 mg/ml. In both diets, cell
counts of the Clostridium histolyticum group, consisting of many species with pathogenic potential, increased
significantly at 0.1 mg/ml. The same effect was observed for Lactobacilli and Enterococci (Ackermann et al.,
2015). These results show that glyphosate causes a change in the microbiota, with a pronounced effect on
fiber degrading species. Fermentation of fibers enable other microbial species to exist and perform their role,
by providing important nutrients as VFA’s, lactate, gasses e.g. (Cunningham and Klein, 2007). Therefore, the
whole balance can change, even though not all species are affected directly. The authors suggest that the
significant reduction of Streptococcus spp. could be due to the inhibition of the cellulolytic species, and not
necessarily to glyphosate itself. It could also be the case for the reduction of Euryarchaeota, as especially
methanogens are associated with the abundance of some protozoal species (Lange et al., 2005), which also
were negatively affected by glyphosate (Ackermann et al., 2015).
Genus/species MIC value of glyphosate (mg/ml)
Pathogens Campylobacter coli 0.15
Campylobacter jejuni 0.15
Clostridium perfringens 5
Clostridium botulinum (type A and B) 1.2
Eschericia coli 1.2
Eschericia coli 1917 strain Nissle 1.2
Salmonella Enteritidis 5
Salmonella Gallinarium 5
Salmonella Typhimurium 5
Commensals Bacillus badius 0.15
Bacillus cereus 0.3
Bifidobacterium adolescentis 0.075
Enterococcus faecalis 0.15
Enterococcus faecium 0.15
Lactobacillus buchneri 0.6
Lactobacillus casei 0.6
Lactobacillus harbinensis 0.6

20
Compared to Table 5, most of the microorganisms described above, will not be negatively affected by the
glyphosate levels, expected in the gut of dairy cows. However, the growth of R. albus and R. flavefaciens
(found in rumen) decreased at 0.001 mg/ml. This does not directly refer to the MIC value, as few cells still
were able to grow, but if we assume it does, an inhibition would start at 0.0001 mg/ml, representing the MEC
value (Fredborg et al., 2013). This is lower than the concentration, which can be found in the rumen (0.003
mg/ml), and a possible inhibition of the two Ruminococcus strains could occur.
The effect of glyphosate on increased incidences of botulism in dairy cows
During the last years, there has been an increase in the incidences of C. botulinum associated diseases in
cattle. Strains of C. botulinum (spore forming, obligate anaerobic bacteria) generates neurotoxins (BoNT, A-
G) that blocks the release of acetylcholine in the neuromuscular junction, leading to botulism in cattle (Böhnel
et al., 2001). Lactic acid producing bacteria (LAB), as Lactobacilli, Lactococci and Enterococci are able to
produce bacteriocines, which are effective against Clostridium spp. and both E. faecalis and E. faecium
inhibited BoNT production by all C. botulinum strains, in a study by Krüger et al. (2013b). Also Shehata et al.
(2013a) discovered that E. faecalis, E. faecium and B. badius inhibited BoNT production while also reducing
the growth of all C. botulinum types tested (A, B, D and E).
Even though the reason for the increased incidences of botulism is unknown, a possible explanation is
proposed by Krüger et al. (2013b). They found that the inhibitory concentrations of glyphosate to E. faecalis
was 10-100 times lower than those, inhibiting the growth of C. Botulimum type B. The possible explanation
for the outbreaks of botulism might therefore refer to the loss of the antagonistic potential of Enterococci
towards C. botulinum, when glyphosate reaches the GIT of the cows. Unpublished data confirmed this
hypothesis; faeces samples from cows with C. botulinum associated diseases, were poorly colonized by
Enterococci, while the opposite was the case for cows, rarely showing any symptoms (of C. botulinum
associated diseases) (Krüger et al., 2013b).
The effect of glyphosate on fungi in dairy cows
Schrödl et al. (2014) studied the relationship between glyphosate excretion in dairy cows and ruminal fungi
of the order Mucorales (Lichtheimia corymbifera, Lichtheimia ramosa, Rhizopus and Mucor – all belonging to
the family Mucoraceae), as fungi are important members of the ruminal microbial community in cattle ().
Glyphosate concentrations in the urine ranged between 0.0 ng/ml and 164 ng/ml, and cows with the highest
glyphosate excretion (˃40 ng/ml) had significantly lower levels of total Mucorales, L. corymbifera and L.
ramosa, compared to cows with a lower concentration. In addition, unpublished data revealed that
Lichtheimia spp, Mucor spp. and Rhicopus spp. are highly resistant to glyphosate in in vitro trials (Schrödl et
al., 2014). The reason why Lichtheimia spp. are depressed in this trial could therefore reflect other imbalances
in the gut, not taken into account here.
Microbial fermentation in the gut of pigs and dairy cows
Microbial fermentation of organic compounds, occurs in cecum and colon of all farm animals, whether in
ruminants, most of the fermentation, takes place in the rumen (Sjaastad et al., 2010). Compared to rumen,
cecum and colon; stomach, and the beginning of the small intestine, only contains low numbers of bacteria,
ranging from 103 to 104 cells/ml, which compete with the animal for the most easily degradable nutrients
(Cunningham and Klein, 2007). The relatively low number is due to a less favorable environment in these

21
compartments, comprising a low pH in the stomach (Hao and Lee, 2004). In comparison, bacterial numbers
in rumen, cecum and colon, rages between 1010 and 1011 cells/ml (Cunningham and Klein, 2007).
Fermentation leads to production of VFA’s, the primary ones being acetate, propionate and butyrate
(Cunningham and Klein, 2007; McDonald et al., 2011). Acetate, propionate and butyrate mainly originates
from fermentation of cellulose, hemicellulose and pectin (Bergman, 1990) but can also be produced by
fermentation of proteins (Nery et al., 2012). The relative proportions of acetate, propionate and butyrate,
can change a lot, ranging between 75:15:10 and 40:40:20, depending on diet (Bergman, 1990). In addition to
acetate, propionate and butyrate, branched VFA’s can only be formed by fermentation of branched amino
acids; isobutyrate and isovalerate, are produced from fermentation of valine and leucine, respectively
(Blachier et al., 2007; McDonald et al., 2011). Valerate can be formed by fermentation of proline (McDonald
et al., 2011; Rasmussen et al., 1988) and by the condensation of acetate and propionate (Bergman, 1990).
Experimental setup
The experimental part of the thesis, comprises three different parts. First, the effect of glyphosate acid,
glyphosate IPA salt, Roundup and POEA on selected bacterial growth are investigated, and whether possible
effects differs between commensal - and potential pathogenic bacteria. In the second part, we investigate if
glyphosate IPA salt, Roundup and POEA show any effects on the composition and activity of gut microbiota,
when added to stomach, cecum and colon content from slaughter pigs and to rumen content from dairy
cows. In the last part, it is investigated, if growth of a strain of Lactobacillus sobrius in stomach content is
affected by glyphosate IPA salt, when pH is held constant (pH=5).

22
Materials and Methods
Chemicals
Jablo Glyfosat (glyphosate acid concentration 360 g/l, corresponding to a glyphosate IPA salt concentration
of 480 g/l), equivalent to Roundup, manufactured by Monsanto Crop Sciences, Denmark. Jablo is referred to
as Roundup, throughout the rest of the thesis. Glyphosate, both formulated as the acid, N-
(phosphonomethyl)glycine (CAS number 1071-83-6) and as the salt, N-(Phosphonomethyl)glycine,
monoisopropylamine salt solution (CAS number 38641-94-0) were purchased from SIGMA-ALDRICH® and
POEA (Tallowamine polyethoxylated, E17136000) from Dr. Ehrenstorfer GmbH.
Bacteria used in the experiment
Table 7 show a list of bacteria, used in these experiments. As the objective was to investigate the effect on
microbes, normally present in the GIT of pigs and cows, different culturable bacteria from the major genera
were chosen. Both the effects on selected commensals and potential pathogens were investigated.
Table 7. Information about bacterial species used in the experiment.
Media Table 7 lists the media used for preparation of overnight cultures of different bacteria. For detailed
information about these, and other media, used during the experiments, see Appendix 2.
Techniques used
Determination of viable counts by use of Drop Plate Procedure
In 96-well microtiterplates, 250 µl of sample (ex. glyphosate IPA salt, 10 mg/ml, T0) was loaded into the first
well in the first column (i), see Figure 9. 25 µl from column i was transferred to 225 µl medium (0.1 M sodium
phosphate buffer, pH 6.5) in column (i+1), mixed 10 times and the process repeated, until the wanted
dilutions were obtained. For each sample, 10 µl, from each of the dilutions of interest, were plated on MRS
agar and incubated anaerobically overnight, at 37°C. The plates were counted the following day and the
colony forming units (CFU) calculated.
Genus/Species Catalog number Origin Culture media Incubation conditions GramCommensals Bifidobacterium adolescentis DSM 20083 Adult intestine Anaerobic BHI 37°C/anaerobic +
Bifidobacterium longum DSM 20219 Adult intestine BHI 37°C/anaerobic +
Bifidobacterium longum DSM 20211 Pig faeces BHI 37°C/anaerobic +
Enterococcus faecalis O3 MRS 37°C/anaerobic +
Enterococcus faecium O4 MRS 37°C/anaerobic +
Escherichia coli K12 BHI 37°C/anaerobic -
Lactobacillus reuteri DSM 20016 Adult intestine MRS 37°C/anaerobic +
Lactobacillus sobrius DSM 16698 Feces, piglet MRS 37°C/anaerobic +Lactobacillus salivarius DSM 20555 Saliva MRS 37°C/anaerobic +
Ruminococcus albus DSM 20455 Ruminococcus media 37°C/anaerobic +
Ruminococcus flavefaciens DSM 25089 Ruminococcus media 37°C/anaerobic +
Streptococcus alactolyticus O10 MRS/colon 37°C/anaerobic +
Streptococcus hyointestinalis O18 MRS/colon 37°C/anaerobic +
Campylobacter coli Microaerophilic BHI 41°C/microaerophilic -
Campylobacter jejuni Microaerophilic BHI 41°C/microaerophilic -
Clostridium perfringens Anaerobic basal broth 37°C/anaerobic +
Escherichia coli K88 DSM 498 BHI 37°C/anaerobic -Salmonella enterica Typhimurium Anaerobic BHI 37°C/anaerobic -Salmonella enterica Entritidis Anaerobic BHI 37°C/anaerobic -
Staphylococcus aureus Clinical isolate 102480 – from OUH Anaerobic BHI 37°C/anaerobic +
Potential
pathogens

23
Figure 9. Microtiterplate used for dilution in preparation of drop plating. Modified after (Chen et al., 2003).
Quantification of VFA analysis
To quantify volatile fatty acids (VFA), 1 ml samples were extracted. To each sample (+ 1 ml standard mix and
1 ml blind sample), 100 μl intern standard was added. The acids were extracted by addition of 0.5 ml HCl and
addition of ether separated the acids from other components, leaving them in the ether phase. After
centrifugation at 2000 g for 10 min (5°C), 50 μl of the supernatant was transferred to microvials, containing
10 μl MTBSTFA, capped and mixed. After heating at 80°C for 20 min, microvials were stored in room
temperature for 48 hours, to ensure complete derivatization. The samples were run on the Gas
Chromatograph (HP 6890 Series GC System) (Richardson et al., 1989).
Gas measurements
For gas measurements, samples were analyzed by Gas Chromatography, using a ML GC82-22 (Mikrolab,
Aarhus, Denmark) with helium as carrier gas. Data were analyzed using the appurtenant PEAK359 software.
N2 gas in the bottles were used as internal standard for calculation of H2, CO2 and CH4 production (Poulsen
et al., 2013).
For further details, see Appendix 1.
Influence of glyphosate acid, glyphosate IPA salt, Roundup and POEA on growth of bacterial
cultures
To study the growth susceptibilities of selected gastrointestinal bacteria (Fejl! Henvisningskilde ikke fundet.)
to glyphosate acid, glyphosate IPA salt, Roundup and POEA, pure bacterial cultures were incubated in 96-
well mikrotiterplates, in the presence of different concentrations of the four treatments.
Samples from -80°C stocks of each bacterial culture were inoculated to broth and incubated overnight. The
overnight cultures were investigated in the microscope to check for purity, and diluted 100 times in broth, if
the growth was found sufficient (density of approx. 109 cells per ml). In a few cases, the overnight cultures
were less dense and only diluted 10 times. For preparation of the microtiterplates, stock solutions of

24
glyphosate acid, glyphosate IPA salt and Roundup were made. Each solution had a start concentration of 10
mg /ml glyphosate acid equivalents, independent of treatments, to ease comparisons (Appendix 2). For each
treatment, 325 µl of the solution was added to the first well in three columns, to make a replicate of three
(Figure 10). For POEA, another procedure was performed. 325 µl growth media was added to the first well in
three columns, as for the other treatments, but POEA was first added in the wells. Experimental
concentrations of POEA were calculated based on its expected concentrations in Roundup, compared to
glyphosate acid equivalents. It was expected that POEA constituted 15 % of Roundup (Giesy et al., 2000;
Sawada et al., 1988) leaving the ratio between glyphosate acid equivalents and POEA to be 2.4 to 1. This
corresponded to addition of 1,4 µl POEA in the first well, when the start concentration of glyphosate acid
equivalent was 10 mg/ml. However, due to a mistake, only 65 % of the concentrations, we expected to be
the pure product POEA, actually consisted of POEA. This left us with an expected proportion of POEA in
Roundup at 9.4 %, instead of 15%, thereby decreasing its proportions towards glyphosate acid equivalents.
In all other wells (row B-H), 225 µl pure media were added. To reach different concentrations of the
treatments, throughout the columns, serial 3.5 dilutions were made. The last well in all columns, served as
control. In each well, dilutions were followed by application of 25 µl diluted bacterial overnight culture,
ending with a total bacterial dilution of 1000 in the wells. After mixing bacteria and media, 50 µl sterilized
paraffin oil was added to each well, to prevent evaporation and limit exchange of oxygen.
Figure 10. Experimental setup of mikrotiterplates and the concentrations used, of the four different treatments
The microtiterplates were incubated in an ELISA spectrophotometer at 37°C and run on the ELISA
spectrophotometer until cultures reached stationary phase (18-24 h). Cell density (growth) was monitored
by measuring optical density at 650 nm (OD650). Bacterial growth curves were obtained by plotting OD650 data
against time. An output from the ELISA spectrophotometer is depicted on the left part of Figure 11. It shows
growth curves of a bacterial strain, in the presence of, glyphosate acid, glyphosate IPA salt, Roundup and
POEA. The concentrations of the individual treatments corresponds to the concentrations displayed in Figure
10. At the right part of Figure 11, growth curves, at different concentrations, are collected in one graph,
facilitating easier interpretation of the data.

25
To determine µmax (maximum growth rate), absorbance data were log-transformed, to investigate where
growth was logarithmic (Figure 12). For all bacteria, at each treatment, at each concentration, µmax was
calculated from the log-transformed data, and the values for the triplicates, averaged.
Figure 12. Draft of absorbance and log(absorbance) plotted against time.
For strict anaerobes, as R. albus and R. flavefaciens, mikrotiterplates were prepared in an anaerobic chamber.
For the potential pathogens, addition of bacteria to plates, were performed in a LAF bench.
For further details, see Appendix 2.
Influence of glyphosate IPA salt, Roundup and POEA on gut microbiota from pigs and cows
To study the effect of glyphosate IPA salt, Roundup and POEA, on microbiota in gut of slaughter pigs and
dairy cows, a total of four pig - and three cow studies were made.
Slaughter pigs
For pigs, the effects of Roundup and POEA were investigated in stomach (n=1) and colon content (n=1). In
addition, the effect of glyphosate IPA salt, Roundup and POEA have been investigated in stomach (n=2),
cecum (n=2) and colon content (n=2).
GIT content from 60 kg slaughter pigs, fed finely ground feed, were taken at slaughter (Research Center
Foulum), placed on ice, and immediately taken to the laboratory. With a cut-off syringe, 10 ml content were
added to 125 ml bottles, sealed with butyl rubber stoppers, containing 35 ml 0.1 M sterile anaerobic sodium-
Figure 11. Output from ELISA spectroscopy – example of growth curves of a bacterial strain in the presence of POEA, glyphosate acid and Roundup. You can choose to get the output as absorbance or as log absorbance on the y-axis. The figure at right, is a draft from the bachelor thesis of Camilla Koed.

26
phosphate buffer (pH=6.5), to reach a 20 % wt/vol suspension. Colon content were weighed out (10 g in each
bottle), as it was too solid to be handled with a syringe. In each bottle, 5 ml of glyphosate acid/glyphosate
IPA salt/Roundup/POEA solutions were added with a syringe, according to the desired concentrations (Table
8, Table 9Fejl! Henvisningskilde ikke fundet.). Due to the mistake, explained above, POEA concentrations
were underestimated, leaving the ratio of glyphosate acid to POEA, higher than expected. Final POEA
concentrations ended at 2.4, 0.24, 0.024 and 0.00024 mg/ml. For all treatments, 0 mg/ml served as control.
Table 8. Overview of experimental setup for glyphosate IPA salt and Roundup
Table 9. Overview of experimental setup for POEA
The gas phase of each serum bottle was changed to CO2 by three successive cycles, where the bottles were
evacuated and refilled, using a manifold, fitted to a vacuum pump and a gas tank. The bottles were removed
from the manifold under a slight overpressure and the pressure adjusted to atmospheric, using a needle. The
bottles were placed in a 37°C shaking water bath for 24 hours and samples were taken at 0, 3/4 and 24 hours.
With a syringe, 1 ml was withdrawn from each serum bottle for measurements of VFA (stored at -18°C until
analysis), 1 ml for DNA analysis (snap frozen in liquid nitrogen and stored at -80°C), 1 ml for immediately pH
measurements and 0.5 ml for Drop Plating Procedure.
Dairy cows
For cows, the effects of glyphosate IPA salt, Roundup and POEA were investigated in ruminal content (n=2).
Ruminal content were taken from normally fed, fistulated dairy cows, at Research Center Foulum. The
content was sieved through a cheesecloth to remove big particles, placed on ice, and immediately taken to
the laboratory. With a syringe, 10 ml content were added to each bottle, containing 35 ml 0.1 M sterile
anaerobic sodium-phosphate buffer (pH=6.5) to reach a 20 % wt/vol suspension. In each bottle, 5 ml of
glyphosate acid/glyphosate IPA salt/Roundup/POEA solutions were added with a syringe, according to the
desired concentrations (Table 8 and Table 9). Due to the mistake, explained above, POEA concentrations
were underestimated, leaving the ratio of glyphosate acid to POEA higher than expected. Final POEA
concentrations ended at 2.4, 0.24, 0.024 and 0.00024 mg/ml. For all treatments, 0 mg/ml served as control.
The gas phase of each serum bottle was changed to a gas mix consisting of 10% H2, 10% CO2 and 80% N2 by
three successive cycles, where the bottles were evacuated and refilled, using a manifold fitted to a vacuum
10 50 10 35 5 (100 mg/ml a.e.)
1 50 10 35 5 (10 mg/ml a.e.)
0.1 50 10 35 5 (1 mg/ml a.e.)
0.001 50 10 35 5 (0.01 mg/ml a.e.)
0 50 10 35 5 (0.00 mg/ml a.e.)
Concentration of glyphosate acid
equivalents (mg/ml)Total volume (ml) Content (ml) 0.1 M sodium-phosphate buffer (ml) Glyphosate IPA salt/Roundup (ml)
POEA concentration
(mg/ml)Total volume (ml) Content (ml) 0.1 M sodium-phosphate buffer (ml) POEA (ml)
2.6 50 10 35 5 (26 mg/ml)
0.26 50 10 35 5 (2.6 mg/ml)
0.026 50 10 35 5 (0.26 mg/ml)
0.00026 50 10 35 5 (0.0026 mg/ml)
0 50 10 35 5 (0.00 mg/ml)

27
pump and a gas tank. The bottles were removed from the manifold under a slight overpressure and the
pressure adjusted to atmospheric, using a needle. The bottles were placed in a 37°C shaking water bath for
24 hours and samples were taken at 0, 2, 4 and 24 hours. With a syringe, 1 ml was withdrawn from each
serum bottle for measurements of VFA (stored at -18°C until analysis), 1 ml for DNA analysis (snap frozen in
liquid nitrogen and stored at -80°C) and 1 ml for immediately pH measurements. 1 ml gas samples were taken
at 0, 2, 4, 6 and 24 hours.
For further details, see Appendix 3.
Influence of glyphosate IPA salt on growth of L. sobrius in stomach content from pigs
The effect of glyphosate IPA salt on the growth of L. sobrius in stomach content, at pH 5, was studied in vitro.
As the pH was held constant, the effect on growth of L. sobrius was attributed to glyphosate IPA salt.
A batch culture system was used, simulating the major environmental conditions in the porcine stomach. The
batch culture system involved 4 bioreactors, each with a working volume of 600-3000 ml. Stomach content
was collected and pooled from four 60 kg pigs, two fed a finely ground ration and two fed a coarse. The
content was stored at -20°C until use, and after thawing, the pooled content was diluted (1:1) in a sterile
anaerobic salt medium. For incubation, 1000 ml of diluted digesta was added to the vessel of each bioreactor.
The suspensions were stirred and kept under anoxic conditions by flushing with N2 gas in the headspace. pH
was maintained at the desired value (pH 5), using a pH-controller, regulated by 1M NaOH and 5M HCl.
Incubation temperature was kept at 37°C, by a circulating water bath. The suspension was inoculated with
10 ml of an overnight culture of L. sobrius, to reach a density of approximately 107 CFU/ml. Different amounts
of glyphosate IPA salt were added, to reach final glyphosate acid concentrations of 10, 1.0, 0.1 and 0.0 mg/ml,
Table 10
Table 10. Overview of experimental setup in bioreactors
During the incubation period, samples were removed from the bioreactors at 0, 2, 4 and 24 hours. 1 ml was
used for VFA determination, and stored at -18°C, until analyses began. Another 10 ml was used for
enumeration of bacteria and were transferred to sterile 125 ml serum bottles, with rubber stoppers,
containing 85 ml broth and 5 ml R. Each suspension was poured into a plastic bag and homogenized in a
stomacher for 2 minutes. Subsequently, enumeration of lactic acid bacteria were performed, using the Drop
Plate Procedure (Chen et al., 2003) (Figure 9).
For further details, see Appendix 4.
Bioreactor Glyphosate acid concentration (mg/ml) Glyphosate IPA salt (ml)
1 0 0
2 0.1 0.3375
3 1 3.375
4 10 33.75

28
Statistical analyses
Data were statistically analyzed using RStudio (Version 0.99.489 – © 2009-2015 RStudio, Inc.).
Growth of pure bacterial cultures were analyzed using the linear mixed effects model, with concentrations
and treatments considered as fixed effects and plate as random. Data from stomach, cecum and colon were
also analyzed using the linear mixed-effects model. Again, treatments and concentrations were considered
as fixed effects, as well as time, and individual was treated as random. In addition, data from rumen were
analyzed using the linear mixed-effects model and concentrations, treatments and time were treated as fixed
effects. Here, both day and cow were included as random effects. In all models, the interaction between
concentrations and treatments were included, as we otherwise assume that differences in responses
between concentrations, are the same between all treatments.
Following descriptions are used to demonstrate different significance levels: Non significant (NS) P>0.1,
†P≤0.1, *P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001
For further details about the statistical models, see Appendix 5.

29
Results To ease comparisons between treatments, all concentrations of glyphosate acid, glyphosate IPA salt and
Roundup, are presented in mg/ml of glyphosate acid equivalents (a.e.) throughout the text. However, even
though concentrations of POEA are lower than concentrations of glyphosate acid equivalents, see Table 9
and Figure 10, I also refer to glyphosate acid concentrations, when POEA is mentioned, as amount of POEA
originally was calculated to correspond to glyphosate acid equivalents.
Influence of glyphosate acid, glyphosate IPA salt, Roundup and POEA on growth of bacterial
cultures
Commensal bacteria
To investigate at which concentrations, the four treatments starts to affect growth of bacteria, if they affect
growth at all, all growth rates were compared to the control,
Growth of B. adolescentis decreased in the presence of glyphosate IPA salt (P ≤ 0.001) and POEA (P ≤ 0.01),
starting at 0.019 mg/ml. Roundup (P ≤ 0.001) decreased growth at 0.067 mg/ml, and in the presence of
glyphosate acid (P ≤ 0.001), growth decreased at 2.86 mg/ml, see Table 11. The same tendency was observed
for S. hyointestinalis, even though the decrease in growth, in the presence of glyphosate IPA salt (P ≤ 0.001),
Roundup (P ≤ 0.001) and POEA (P ≤ 0.001), started at 0.067 mg/ml. Glyphosate acid (P ≤ 0.001) decreased
growth at 10 mg/ml. Glyphosate IPA salt (P ≤ 0.001) and glyphosate acid (P ≤ 0.001) decreased growth of S.
alactolyticus at 0.067 mg/ml and 10 mg/ml, respectively. The growth of L. salivarius and L. reuteri decreased
at 0.82 mg/ml in the presence of glyphosate IPA salt (P ≤ 0.001), while decrease, in the presence of Roundup
(P ≤ 0.001) and POEA (P ≤ 0.001) started at 2.86 mg/ml. Glyphosate acid (P ≤ 0.001) decreased growth at 10
mg/ml. In the presence of glyphosate IPA salt (P ≤ 0.001), growth of L. sobrius decreased at 0.230 mg/ml, and
for Roundup (P ≤ 0.001) and POEA (P ≤ 0.001) decreases were observed at 0.820 mg/ml. Glyphosate acid (P
≤ 0.001) decreased growth at 10 mg/ml. Growth of E. faecium decreased at 0.820 mg/ml, in the presence of
glyphosate acid (P ≤ 0.05). Both glyphosate IPA salt (P ≤ 0.05) and POEA (P ≤ 0.05) decreased growth at 2.86
mg/ml, even though no effects were observed at 10 mg/ml. Roundup (P ≤ 0.05) decreased growth at 10
mg/ml. In general, only few significant effects were observed for R. albus and R. flavefaciens. Glyphosate acid
decreased growth of R albus (P ≤ 0.01) and R. flavefaciens (P>0.05) at 10 mg/ml. The same was evident for
Roundup, but only for R. albus (P ≤ 0.01).
Graphs are presented in Appendix 6.

30
Table 11. Growth rates of commensal bacteria in the presence of glyphosate acid, glyphosate IPA salt, Roundup and POEA.
Potential pathogens
Glyphosate IPA salt (P ≤ 0.001) decreased growth of C. perfringens at 0.067 mg/ml, while decreases in the
presence of Roundup (P ≤ 0.001) and POEA (P ≤ 0.001) were observed at 0.23 mg/ml, see Table 12.
Glyphosate acid (P ≤ 0.001) decreased growth at 2.86 mg/ml. Decrease in growth of S. aureus, in the presence
of glyphosate IPA salt (P ≤ 0.05), was observed at 0.019 mg/ml. Roundup (P ≤ 0.001) and POEA (P ≤ 0.001)
decreased growth from 0.067 mg/ml while glyphosate acid (P ≤ 0.001) decreased growth from 0.82 mg/ml.
For E. coli K88, S. enterica Enteritidis and S. enterica Typhimurium, glyphosate acid (P ≤ 0.001) decreased
growth at 2.86 mg/ml. In the presence of glyphosate IPA salt (P ≤ 0.001) growth of E. coli K88 and S. enterica
Typhimurium decreased at 10 mg/ml while decrease in growth of S. enterica Enteritidis was observed at 2.86
mg/ml, in the presence of glyphosate IPA salt (P ≤ 0.05). For all three strains, Roundup (P ≤ 0.001) decreased
growth at 10 mg/ml and no effects were found on growth, in the presence of POEA.
Graphs are presented in Appendix 7.
0 0.005 0.019 0.067 0.23 0.82 2.86 10 SEM n
B. adolescentis Glyphosate acid 1.74 1.71 1.83 1.82 1.79 1.6 0.55*** 0.28*** 0.12 4
Glyphosate IPA salt 1.77 1.35 0.14*** 0.11*** 0.11*** 0.13*** 0.26*** 0.05*** 0.16 2
Roundup 1.7 1.33 1.29 0.19*** 0.27*** 0.25*** 0.25*** 0.21*** 0.16 2
POEA 1.7 1.35 0.68** 0.33*** 0.26*** 0.26*** 0.24*** 0.25*** 0.16 2
E. faecium Glyphosate acid 0.47 0.46 0.44 0.42 0.36 0.28* 0.20*** 0.25** 0.04 5
Glyphosate IPA salt 0.42 0.38 0.33 0.33 0.31 0.21 0.20* 0.23 0.06 2
Roundup 0.5 0.47 0.49 0.43 0.43 0.43 0.27 0.24* 0.05 3
POEA 0.5 0.44 0.42 0.4 0.36 0.29 0.24* 0.27 0.05 3
L. reuteri Glyphosate acid 2.42 2.34 2.43 2.36 2.38 2.27 2.07 1.33*** 0.13 4
Glyphosate IPA salt 2.5 2.42 2.3 2.24 1.32*** 0.06*** -0.11*** 0.17 2
Roundup 2.34 2.35 2.34 2.27 2.27 2.09 0.40*** 0.23*** 0.17 2
POEA 2.34 2.31 2.29 2.23 2.24 1.6 0.28*** 0.20*** 0.17 2
L. sobrius Glyphosate acid 1.31 1.32 1.36 1.32 1.31 1.26 1.15 0.37*** 0.09 4
Glyphosate IPA salt 1.28 1.32 1.41 1.27 0.66*** 0.24*** 0.23*** 0.23*** 0.1 2
Roundup 1.34 1.36 1.43 1.42 1.4 0.19*** 0.19*** 0.21*** 0.1 2
POEA 1.34 1.35 1.42 1.4 1.4 0.18*** 0.19*** 0.23*** 0.1 2
L. salivarius Glyphosate acid 2.9 2.82 2.84 2.73 2.77 2.63 2.34 1.35*** 0.37 4
Glyphosate IPA salt 2.95 2.86 2.89 2.82 2.83 0.51*** 0.26*** 0.31*** 0.42 2
Roundup 2.84 2.82 2.86 2.84 2.81 2.58 0.26*** 0.35*** 0.42 2
POEA 2.84 2.96 3.01 2.99 2.96 2.64 0.19*** 0.45*** 0.42 2
S. alactolyticus Glyphosate acid 0.81 0.81 0.79 0.75 0.71 0.62 0.43 0.25*** 0.14 4
Glyphosate IPA salt 0.82 0.46 -0.08*** -0.08*** -0.07*** -0.10*** 0.03*** 0.16 2
Roundup 0.79 0.92 0.89 0.8 0.47 0.42 0.48 0.58 0.16 2
POEA 0.79 0.72 0.71 0.67 0.4 0.5 0.44 0.42 0.16 2
S. hyointestinalis Glyphosate acid 3.06 2.89 2.9 2.92 2.95 2.44 0.21*** 0.17 4
Glyphosate IPA salt 3 2.84 0.21*** 0.52*** 0.28*** 0.29*** 0.25*** 0.23 2
Roundup 3.12 2.62 0.12*** 0.16*** 0.19*** 0.09*** 0.05*** 0.23 2
POEA 3.12 2.83 0.10*** 0.14*** 0.13*** 0.21*** 0.18*** 0.23 2
R. albus Glyphosate acid 1.89 2.04 1.61 1.61 1.61 1.63 0.98 0.28** 0.28 4
Glyphosate IPA salt 1.33 1.99 1.39 2.11 1.69 1.4 1.56 0.52 0.36 2
Roundup 2.46 2.37 2.19 2.2 1.6 1.42 1.48 0.29** 0.36 2
POEA 2.46 2.36 3.08 2.73 2.56 1.76 2.1 2.82 0.36 2
R. flavefaciens Glyphosate acid 2.03 1.81 1.82 1.85 1.71 1.56 0.93 0.24* 0.43 4
Glyphosate IPA salt 1.53 1.54 2.11 2.04 1.69 1.48 1.66 0.64 0.53 2
Roundup 2.54 3.02 2.98 3.26 3.03 3.45 1.75 0.66 0.53 2
POEA 2.54 2.5 2.65 2.29 2.53 2.31 2.37 3.44 0.53 2
Concentration of glyphosate acid equivalents (mg/ml)

31
Table 12. Growth rates of potential pathogenic bacteria in the presence of glyphosate acid, glyphosate IPA salt, Roundup and POEA.
In general, commensals seem to be more tolerant to glyphosate acid than to any of the other treatments,
especially glyphosate IPA salt, see Table 13. In contrast, sensitivity of pathogens towards glyphosate acid
seems to be more pronounced. Values for glyphosate IPA salt, Roundup and POEA are widely dispersed. For
both commensals and potential pathogens, the levels of Roundup and POEA, where decreases are observed,
are very uniform.
Table 13. Overview of concentrations, where growth significantly decreased, in presence of glyphosate acid, glyphosate IPA salt,
Roundup and POEA for all tested bacteria.
The graphs, showing the growth rates for the individual bacteria, are placed in Appendix X. To ease the
interpretation, a non-linear x-axis are used. To explain the choice, two graphs with non-linear and linear x –
0 0.005 0.019 0.067 0.23 0.82 2.86 10 SEM n
C. perfringens Glyphosate acid 5.11 5 4.96 4.9 5.14 4.98 0.13*** 0.01*** 0.32 4
Glyphosate IPA salt 5.17 5.37 4.64 1.08*** 0.00*** 0.73*** -0.20*** -0.18*** 0.43 2
Roundup 5.04 4.79 4.85 4.8 0.29*** 0.43*** 0.33*** 0.35*** 0.43 2
POEA 5.04 5.15 5.19 3.55 0.32*** 0.69*** 0.58*** 0.53*** 0.43 2
E. coli K88 Glyphosate acid 4.63 4.8 4.77 4.72 4.58 4.29 2.89*** 0.20*** 0.31 4
Glyphosate IPA salt 4.72 4.95 4.54 4.86 4.78 4.49 3.78 1.30*** 0.38 2
Roundup 4.54 4.58 4.36 4.38 4.2 4.17 3.89 0.75*** 0.38 2
POEA 4.54 4.82 4.84 4.75 4.43 4.68 4.58 4.07 0.38 2
S. enterica Enteritidis Glyphosate acid 3.92 3.97 3.99 3.96 3.88 3.65 2.09*** 0.19*** 0.12 4
Glyphosate IPA salt 3.91 3.87 3.99 4.03 3.87 3.74 3.28* 1.76*** 0.14 2
Roundup 3.94 3.89 4.00 3.62 3.99 3.58 3.6 2.50*** 0.14 2
POEA 3.94 3.97 4.01 3.82 3.95 3.81 3.99 4.06 0.14 2
S. enterica Typhimurium Glyphosate acid 3.78 3.73 3.75 3.64 3.66 3.52 2.20*** 0.19*** 0.23 4
Glyphosate IPA salt 3.87 3.8 3.86 3.75 3.75 3.62 3.33 2.03*** 0.26 2
Roundup 3.7 3.68 3.63 3.36 3.47 3.6 3.39 2.47*** 0.26 2
POEA 3.7 3.64 3.62 3.42 3.67 3.43 3.71 3.76 0.26 2
S. aureus Glyphosate acid 2.69 2.67 2.72 2.69 2.6 2.09*** 0.61*** 0.20*** 0.09 4
Glyphosate IPA salt 2.73 2.79 2.11* 0.12*** 0.12*** 0.11*** 0.13*** 0.19*** 0.11 2
Roundup 2.64 2.74 2.49 1.79*** 0.96*** 0.37*** 0.27*** 0.52*** 0.11 2
POEA 2.64 2.71 2.28 1.27*** 0.15*** 0.17*** 0.19*** 0.20*** 0.11 2
Concentration of glyphosate acid equivalents (mg/ml)
Concentration of glyphosate acid equivalents (mg/ml)
Glyphosate acid Glyphosate IPA salt Roundup POEA
Commensals B. adolescentis 2.86 0.019 0.067 0.019
E. faecium 0.82 2.86 10 2.86
L. reuteri 10 0.82 2.86 2.86
L. salivarius 10 0.82 2.86 2.86
L. sobrius 10 0.23 0.82 0.82
S. alactolyticus 10 0.067 > 10 > 10
S. hyointestinalis 10 0.067 0.067 0.067
R. albus 10 > 10 10 > 10
R. flavefaciens 10 > 10 > 10 > 10
C. perfringens 2.86 0.067 0.23 0.23
E. coli 2.86 10 10 > 10
S. enterica Enteritidis 2.86 2.86 10 > 10
S. enterica Typhimurium 2.86 10 10 > 10
S. aureus 0.82 0.019 0.067 0.067
Pontential
pathogens

32
axes, respectively, are in Figure 13. When using the linear x-axis, all the results, at the low concentrations,
are aggregated to the left. This makes it difficult to see, at which concentrations the different treatments
have an effect, and therefore, a non-linear x-axis is used for all bacteria.
Figure 13. Growth rates of S. hyointestinalis. The graph to the left has a non-linear x-axis, and the graph to the right has a linear x –
axis.
Influence of glyphosate IPA salt, Roundup and POEA on gut microbiota from pigs and cows
Pigs
Stomach
For glyphosate acid, two replicates were made, while three were conducted for Roundup and POEA. This
leaves the values observed at 0 mg/ml the same for both Roundup and POEA.
There were no significant effects of glyphosate IPA salt and POEA on pH, even though the levels were lower
when glyphosate IPA salt was added, see Table 14. Roundup (P ≤ 0.05) decreased pH from 5.91 at 0 mg/ml
to 5.31 at 10 mg/ml.
No effects were observed on overall VFA concentrations, relative levels of acetate and propionate and the
lactate concentrations, in the presence of any of the three treatments. In addition, no effects were observed
on the numbers of lactic acid bacteria.
Graphs are presented in Appendix 8.
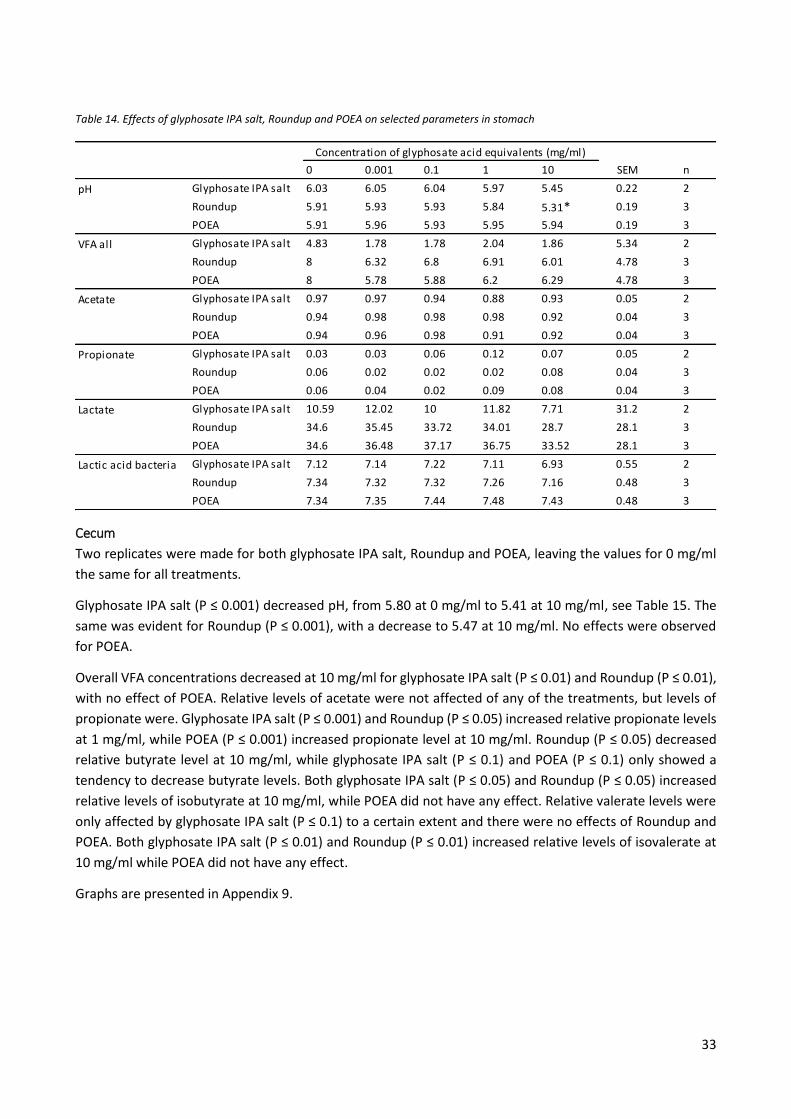
33
Table 14. Effects of glyphosate IPA salt, Roundup and POEA on selected parameters in stomach
Cecum
Two replicates were made for both glyphosate IPA salt, Roundup and POEA, leaving the values for 0 mg/ml
the same for all treatments.
Glyphosate IPA salt (P ≤ 0.001) decreased pH, from 5.80 at 0 mg/ml to 5.41 at 10 mg/ml, see Table 15. The
same was evident for Roundup (P ≤ 0.001), with a decrease to 5.47 at 10 mg/ml. No effects were observed
for POEA.
Overall VFA concentrations decreased at 10 mg/ml for glyphosate IPA salt (P ≤ 0.01) and Roundup (P ≤ 0.01),
with no effect of POEA. Relative levels of acetate were not affected of any of the treatments, but levels of
propionate were. Glyphosate IPA salt (P ≤ 0.001) and Roundup (P ≤ 0.05) increased relative propionate levels
at 1 mg/ml, while POEA (P ≤ 0.001) increased propionate level at 10 mg/ml. Roundup (P ≤ 0.05) decreased
relative butyrate level at 10 mg/ml, while glyphosate IPA salt (P ≤ 0.1) and POEA (P ≤ 0.1) only showed a
tendency to decrease butyrate levels. Both glyphosate IPA salt (P ≤ 0.05) and Roundup (P ≤ 0.05) increased
relative levels of isobutyrate at 10 mg/ml, while POEA did not have any effect. Relative valerate levels were
only affected by glyphosate IPA salt (P ≤ 0.1) to a certain extent and there were no effects of Roundup and
POEA. Both glyphosate IPA salt (P ≤ 0.01) and Roundup (P ≤ 0.01) increased relative levels of isovalerate at
10 mg/ml while POEA did not have any effect.
Graphs are presented in Appendix 9.
0 0.001 0.1 1 10 SEM n
pH Glyphosate IPA salt 6.03 6.05 6.04 5.97 5.45 0.22 2
Roundup 5.91 5.93 5.93 5.84 5.31* 0.19 3
POEA 5.91 5.96 5.93 5.95 5.94 0.19 3
VFA all Glyphosate IPA salt 4.83 1.78 1.78 2.04 1.86 5.34 2
Roundup 8 6.32 6.8 6.91 6.01 4.78 3
POEA 8 5.78 5.88 6.2 6.29 4.78 3
Acetate Glyphosate IPA salt 0.97 0.97 0.94 0.88 0.93 0.05 2
Roundup 0.94 0.98 0.98 0.98 0.92 0.04 3
POEA 0.94 0.96 0.98 0.91 0.92 0.04 3
Propionate Glyphosate IPA salt 0.03 0.03 0.06 0.12 0.07 0.05 2
Roundup 0.06 0.02 0.02 0.02 0.08 0.04 3
POEA 0.06 0.04 0.02 0.09 0.08 0.04 3
Lactate Glyphosate IPA salt 10.59 12.02 10 11.82 7.71 31.2 2
Roundup 34.6 35.45 33.72 34.01 28.7 28.1 3
POEA 34.6 36.48 37.17 36.75 33.52 28.1 3
Lactic acid bacteria Glyphosate IPA salt 7.12 7.14 7.22 7.11 6.93 0.55 2
Roundup 7.34 7.32 7.32 7.26 7.16 0.48 3
POEA 7.34 7.35 7.44 7.48 7.43 0.48 3
Concentration of glyphosate acid equivalents (mg/ml)

34
Table 15. Effects of glyphosate IPA salt, Roundup and POEA on selected parameters in cecum
Colon
For both Roundup and POEA, two replicates were made, leaving all the values for 0 mg/ml the same.
Roundup (P ≤ 0.001) decreased pH from 5.97 at 0 mg/ml to 5.65 at 10 mg/ml and no effects were observed
for POEA.
Overall VFA concentrations were not affected by either Roundup or POEA, but relative acetate levels
decreased at 10 mg/ml in the presence of Roundup (P ≤ 0.05), see Table 16. POEA did not have any effect.
Relative propionate levels increased at 10 mg/ml for Roundup (P ≤ 0.01) and POEA (P ≤ 0.05) and no effects
were observed on relative butyrate levels. Roundup (P ≤ 0.001) increased relative isobutyrate, valerate and
isovalerate levels at 10 mg/ml, while no effects were observed for POEA.
Graphs are presented in Appendix 10.
0 0.001 0.1 1 10 SEM n
pH Glyphosate IPA salt 5.8 5.79 5.78 5.74 5.41*** 0.06 2
Roundup 5.8 5.84 5.85 5.78 5.47*** 0.06 2
POEA 5.8 5.82 5.8 5.82 5.8 0.06 2
VFA all Glyphosate IPA salt 241 225 232 232 206** 9.93 2
Roundup 241 232 229 240 209** 9.93 2
POEA 241 223 229 222 240 9.93 2
Acetate Glyphosate IPA salt 0.615 0.625 0.617 0.601 0.583 0.035 2
Roundup 0.615 0.626 0.635 0.617 0.603 0.035 2
POEA 0.615 0.628 0.629 0.633 0.608 0.035 2
Propionate Glyphosate IPA salt 0.227 0.226 0.228 0.256*** 0.266*** 0.018 2
Roundup 0.227 0.225 0.222 0.249* 0.262*** 0.018 2
POEA 0.227 0.225 0.224 0.224 0.263*** 0.018 2
Butyrate Glyphosate IPA salt 0.128 0.12 0.126 0.116 0.101† 0.019 2
Roundup 0.128 0.121 0.114 0.107 0.090* 0.019 2
POEA 0.128 0.118 0.118 0.115 0.100† 0.019 2
Isobutyrate Glyphosate IPA salt 0.0047 0.0046 0.0048 0.0049 0.0075* 0.001 2
Roundup 0.0047 0.0045 0.0048 0.0048 0.0075* 0.001 2
POEA 0.0047 0.0045 0.0046 0.0048 0.005 0.001 2
Valerate Glyphosate IPA salt 0.0217 0.0208 0.0213 0.0195 0.0353† 0.0035 2
Roundup 0.0217 0.0211 0.0206 0.0187 0.0299 0.0035 2
POEA 0.0217 0.0209 0.021 0.0197 0.0193 0.0035 2
Isovalerate Glyphosate IPA salt 0.0035 0.0034 0.0035 0.0035 0.0070** 0.0009 2
Roundup 0.0035 0.0034 0.0036 0.0034 0.0068** 0.0009 2
POEA 0.0035 0.0034 0.0034 0.0036 0.0038 0.0009 2
Concentration of glyphosate acid equivalents (mg/ml)

35
Table 16. Effects of Roundup and POEA on selected parameters in colon
Cows
Rumen
Two replicates were made for both glyphosate IPA salt, Roundup and POEA, leaving the values for 0 mg/ml
the same for all treatments
Glyphosate IPA salt (P ≤ 0.001) decreased pH from 6.47 at 0 mg/ml to 6.37 at 1 mg/ml and further to 5.68 at
10 mg/ml, see Table 17. The same was observed for Roundup (P ≤ 0.001) where pH decreased to 6.40 and
5.79, at 1 mg/ml and 10 mg/ml, respectively. POEA (P ≤ 0.05) increased pH to 6.50 at 10 mg/ml.
Glyphosate IPA salt (P ≤ 0.05) increased overall VFA concentration at 10 mg/ml while Roundup (P ≤ 0.05)
increased overall VFA concentration at 1 mg/ml. POEA did not have any effect. Glyphosate IPA salt (P ≤ 0.01)
and Roundup (P ≤ 0.001) decreased relative levels of acetate at 1 mg/ml and POEA (P ≤ 0.001) decreased
relative acetate level at 10 mg/ml. Glyphosate IPA salt (P ≤ 0.01) and Roundup (P ≤ 0.01) increased relative
levels of propionate at 1 mg/ml and POEA (P ≤ 0.01) increased relative propionate level at 10 mg/ml. None
of the treatments had any effects on the relative levels of butyrate, isobutyrate, valerate and isovalerate.
In addition, methane measurements were performed in all rumen samples. However, due to technical
problems, the results showed unexplainable variations, and no statistical analysis were conducted. Even
though the results of methane production varied considerable, one thing became clear: at 10 mg/ml, no
methane were produced, independently of the treatments. In addition, no production was observed at 1
mg/ml for Roundup.
Graphs are presented in Appendix 11.
0 0.001 0.1 1 10 SEM n
pH Roundup 5.97 5.98 5.98 5.94 5.65*** 0.06 2
POEA 5.97 5.97 5.97 5.98 6 0.06 2
VFA all Roundup 242 235 241 235 238 8.94 2
POEA 242 234 240 243 250 8.94 2
Acetate Roundup 0.59 0.6 0.59 0.59 0.55* 0.03 2
POEA 0.59 0.59 0.58 0.58 0.58 0.03 2
Propionate Roundup 0.228 0.226 0.228 0.237 0.258** 0.005 2
POEA 0.228 0.228 0.232 0.233 0.249* 0.005 2
Butyrate Roundup 0.13 0.129 0.133 0.128 0.123 0.022 2
POEA 0.13 0.133 0.138 0.136 0.119 0.022 2
Isobutyrate Roundup 0.0088 0.0089 0.009 0.0095 0.0127*** 0.0032 2
POEA 0.0088 0.0086 0.0089 0.009 0.0097 0.0032 2
Valerate Roundup 0.0347 0.0338 0.0344 0.0343 0.0485*** 0.0022 2
POEA 0.0347 0.0344 0.0349 0.0349 0.0331 0.0022 2
Isovalerate Roundup 0.0064 0.0065 0.0066 0.0058 0.0113*** 0.0021 2
POEA 0.0064 0.0063 0.0065 0.0067 0.0073 0.0021 2
Concentration of glyphosate acid equivalents (mg/ml)

36
Table 17. Effects of glyphosate IPA salt, Roundup and POEA on selected parameters in rumen
To give an overview of the effects of glyphosate IPA salt, Roundup and POEA in the different compartments,
Table 18 has been prepared. In general, VFA concentrations show varying results, pH, relative acetate - and
butyrate levels decrease while relative propionate-, isobutyrate-, valerate- and isovalerate levels increase.
Overall, effects in rumen start earlier than in stomach, cecum and colon and, in general, POEA is less toxic
than glyphosate IPA salt and Roundup.
0 0.001 0.1 1 10 SEM n
pH Glyphosate IPA salt 6.47 6.47 6.45 6.37*** 5.68*** 0.01 2
Roundup 6.47 6.48 6.46 6.40*** 5.79*** 0.01 2
POEA 6.47 6.47 6.47 6.47 6.50* 0.01 2
VFA all Glyphosate IPA salt 110 113 112 112 116* 1.8 2
Roundup 110 107 111 115* 115* 1.8 2
POEA 110 109 110 112 110 1.8 2
Acetate Glyphosate IPA salt 0.667 0.668 0.665 0.651** 0.643*** 0.003 2
Roundup 0.667 0.669 0.669 0.650*** 0.643*** 0.003 2
POEA 0.667 0.665 0.667 0.664 0.650*** 0.003 2
Propionate Glyphosate IPA salt 0.184 0.181 0.184 0.201** 0.202** 0.005 2
Roundup 0.184 0.182 0.183 0.203** 0.202** 0.005 2
POEA 0.184 0.183 0.184 0.187 0.203** 0.005 2
Butyrate Glyphosate IPA salt 0.119 0.119 0.12 0.119 0.126 0.004 2
Roundup 0.119 0.119 0.117 0.119 0.126 0.004 2
POEA 0.119 0.12 0.118 0.118 0.119 0.004 2
Isobutyrate Glyphosate IPA salt 0.0091 0.0095 0.0093 0.0084 0.0087 0.001 2
Roundup 0.0091 0.0091 0.0092 0.0083 0.0086 0.001 2
POEA 0.0091 0.0093 0.0092 0.0092 0.0084 0.001 2
Valerate Glyphosate IPA salt 0.0176 0.0176 0.0173 0.016 0.0165 0.0009 2
Roundup 0.0176 0.0172 0.0171 0.0157 0.0165 0.0009 2
POEA 0.0176 0.0178 0.0176 0.0171 0.0162 0.0009 2
Isovalerate Glyphosate IPA salt 0.0042 0.0049 0.0047 0.0035 0.0037 0.0021 2
Roundup 0.0042 0.0042 0.0044 0.0035 0.0036 0.0021 2
POEA 0.0042 0.0046 0.0045 0.0045 0.0035 0.0021 2
Concentration of glyphosate acid equivalents (mg/ml)

37
Table 18. Overview of concentrations, where selected parameters are significantly affected by glyphosate IPA salt, Roundup and POEA
in stomach, cecum, colon and rumen
Influence of glyphosate IPA salt on growth of L. sobrius in stomach content from pigs The experiment was performed as a pilot study, and no statistical analyses were conducted due to lack of
replicates. However, the results for lactic acid bacteria indicate that the cell count was lower at 10 mg/ml,
both after 4 and 24 hours, even though no statistical tests were performed. Due to a mistake, values for 0
and 2 hours are missing, but only for enumeration of lactic acid bacteria. The VFA results showed diverging
results, and no tendencies were visualized.
Graphs are presented in Appendix 12.
Glyphosate IPA salt Roundup POEA
Stomach pH - 10 -
VFA all - - -
Acetate - - -
Propionate - - -
Lactate - - -
Lactic acid
bacteria- - -
Cecum pH 10 10 -
VFA all 10 10 -
Acetate - - -
Propionate 1 1 10
Butyrate 10† 10 10†
Isobutyrate 10 10 -
Valerate 10† - -
Isovalerate 10 10 -
Colon pH 10 -
VFA all - -
Acetate 10 -
Propionate 10 10
Butyrate - -
Isobutyrate 10 -
Valerate 10 -
Isovalerate 10 -
Rumen pH 1 1 10
VFA all 10 1 -
Acetate 1 1 10
Propionate 1 1 10
Butyrate - - -
Isobutyrate - - -
Valerate - - -
Isovalerate - - -
Concentration of glyphosate acid equivalents (mg/ml)

38
Discussion N-(phosphonomethyl)glycine, commonly known as Glyphosate, is a broad-spectrum, nonselective, systemic
herbicide (Franz et al., 1997), existing as an acid or formulated as a salt. Normally, glyphosate isopropylamine
salt is the form of glyphosate, primarily used in Roundup formulations(Malik et al., 1989). In addition,
Roundup also contains water and a surfactant, usually a polyethoxylated tallowamine (POEA) (Giesy et al.,
2000).
Glyphosate, is the active ingredient in Roundup, inhibiting 5-enolpyruvylshikimate-3-phosphate synthase
(EPSPS) in the shikimate-pathway, which leads to formation of aromatic amino acids (Levin and Sprinson,
1964). The shikimate-pathway is present in plants, fungi and bacteria, but not in animals (Bentley, 1990; Franz
et al., 1997; Herrmann, 1995; Kishore and Shah, 1988; Padgette et al., 1995a).
Recently, studies have raised concerns about the effects of glyphosate on gut microbiota (Ackermann et al.,
2015; Krüger et al., 2013b; Shehata et al., 2013a; Shehata et al., 2013b). Results have shown that glyphosate
has a potential inhibiting effect on growth of commensal bacteria, normally occupying the gut of farm
animals. Opposed to the effect on commensals, potential pathogens were, in general, more tolerant towards
glyphosate. If these effects can be recovered in vivo, changes in microbiota, in favor of potential pathogens,
can possibly lead to negative effects of health and productivity of farm animals.
The present study was conducted to investigate effects of glyphosate on the microbiota, found in gut of
slaughter pigs and dairy cows. In the present experiment, glyphosate acid, glyphosate IPA salt, Roundup and
POEA, were included to differentiate eventual effects on microbiota between the different compounds,
included in commercial glyphosate formulations.
Influence of glyphosate acid, glyphosate IPA salt, Roundup and POEA on growth of bacterial
cultures
Objective 1
The first objective was to investigate if glyphosate acid, glyphosate IPA salt, Roundup and POEA inhibited
bacterial growth. We hypothesized that glyphosate (Roundup) would suppress growth of gut bacteria and
that gut bacteria would have different susceptibilities towards glyphosate (Roundup). In addition, we
hypothesized that the effect of glyphosate would depend on the chemical formulation (acid, salt, mixture
and surfactant). Throughout the rest of the thesis, glyphosate acid, glyphosate IPA salt, Roundup and POEA
are referred to as ‘treatments’.
Overall, growth of most bacteria were inhibited by one or more of the four treatments, even though the
effect of treatments were not similar, between different strains, confirmed by Shehata et al. (2013b). In
addition, different treatments also showed differences in the degree of inhibition. However, we had
difficulties in culturing Enterococcus faecium, Ruminococcus albus and Ruminococcus flavefaciens, and
therefore, the inhibition of treatments, or lack hereof, should be questioned. In addition Enterococcus
faecalis, Bifidobacterium longum, Campylobacter jejuni and Campylobacter coli were cultured, but because
of inadequate growth, which we expected were due to poor overall growth conditions, culturing did not
succeed, and results are not included.
The lowest concentration, completely inhibiting bacterial growth, is defined as the Minimum Inhibitory
Concentration (MIC). We found MIC values in the range from 0.019 mg/ml to 10 mg/ml, depending on

39
bacteria and treatments used, indicative of considerable changes in sensitivities. However, even more
interestingly than MIC, Minimum Effect Concentration (MEC), calculated as 1/10 of MIC (Fredborg et al.,
2013), is the lowest concentration of an inhibitor needed, to initiate inhibition of a microorganism (Arikan et
al., 2001). If MIC values range between 0.019 mg/ml and 10 mg/ml, MEC values will range from 0.0019 mg/ml
to 1 mg/ml. To get an estimate of whether it is possible to observe MEC values of glyphosate acid equivalents
in vivo, we sat up two different scenarios, calculating possible concentrations of glyphosate acid equivalents
in the gut. Bifidobacterium adolescentis had a MIC value of 0.019 mg/ml, corresponding to the level possible
to find in the colon of pigs (Table 4). However, the inhibiting effect of B. adolescentis would already start at
0.0019 mg/ml, meaning, that it could also be affected in both stomach and small intestine, if present, as
concentrations of 0.005 mg/ml glyphosate acid equivalents could be present here. In addition to B.
adolescentis, Staphylococcus aureus also had the potential to be affected in colon of pigs, as to a MEC value
of 0.0067 mg/ml. However, calculations of the different possible concentrations of glyphosate acid
equivalents are based on different assumptions, which can be discussed. They are based on feed rations
containing maximum residue levels (MRL) of glyphosate, and according to the findings of glyphosate residues
in plants/crops, presented earlier, such levels are uncommon (Arregui et al., 2004; Bøhn et al., 2014;
Miljøstyrelsen, 2014). However, some exceedances have been observed (Plantedirektoratet, 2010), but the
risk that all feed ingredients in the ration has the same high level, is unlikely. Another important assumption,
which might be questioned, is that degradation of glyphosate does not take place in the gut and that only 30
% is absorbed (Commission, 2002). Glyphosate might be degraded to AMPA in the animals, like observed in
plants (Arregui et al., 2004; Bøhn et al., 2014) and if that is the case, the residue levels will change. So far, the
toxicity of AMPA it is not known and therefore it is difficult to include it in the evaluation of the overall toxicity
of glyphosate residues in farm animals. Due to these assumptions, lower concentrations than the ones
presented in Table 4 and Table 5 might be more realistic to find. However, as to MEC values, bacteria still
have the potential to be affected, even though lower levels of glyphosate acid equivalents are found in vivo.
To further examine exact MIC values, and thereby possible MEC values, different concentrations for each
bacteria, according to the results found in our study, should be further investigated. For instance, growth of
Streptococcus hyointestinalis, in the presence of Roundup, are non-existing at a concentration from 0.067
mg/ml to 10 mg/ml. Therefore, it is more interesting to investigate several concentrations from 0.019 mg/ml
to 0.067 mg/ml, as it is here, inhibition occurs. For other bacteria, as Salmonella enterica Typhimurium, MIC
value for glyphosate acid was not yet determined at 10 mg/ml, and therefore it could be an advantage to
further increase concentrations.
Objective 2
In addition to the investigation, of whether glyphosate acid, glyphosate IPA salt, Roundup and POEA inhibited
bacterial growth, we were also interested in investigating if inhibition differed between commensal - and
potential pathogenic bacteria. We hypothesized that gut bacteria would have different susceptibilities
towards glyphosate (Roundup) and that the effect of glyphosate would depend on the chemical formulation
(acid, salt, mixture and surfactant).
Results for nine commensal - and five potential pathogenic bacteria are included. Important to note is that
independent of bacteria, there were big differences in the effects of glyphosate acid, glyphosate IPA salt,
Roundup and Jablo.

40
Overall differences between commensals and potential pathogens
In general, commensal bacteria were tolerant to glyphosate acid, sensitive towards glyphosate IPA salt, and
sensitive - to moderately tolerant towards Roundup and POEA. As already mentioned, we had difficulties in
culturing E. faecium, R. albus and R. flavefaciens. In general, growth rates of E. faecium were low, even in
control wells, compared to growth rates for other bacteria. We might assume that before an inhibitor can
affect growth, bacteria need to be physiologically active, as growth only can be affected, if growth is present.
This could explain missing effects on E. faecium, and as Shehata et al. (2013b) found E. faecium to be highly
sensitive to Roundup, this further underpin the growth-theory. Not mentioned above, Streptococcus
alactolyticus, also had a lower growth rate. Even though effects of glyphosate acid and glyphosate IPA salt
were evident, it is noteworthy that S. hyointestinalis were sensitive towards both Roundup and POEA, not
affecting S. alactolyticus at all. Overall growth rates of R. albus and R. flavefaciens did not differ considerably
from the others, but as they are strictly anaerobic, even a very small amount of air, entering the well, can
possibly impair growth. Opposed to our findings, Ackermann et al. (2015) reported a high sensitivity (0.001
mg/ml) of R. albus and R. flavefaciens, towards glyphosate acid whereas both strains were highly tolerant
towards glyphosate acid in our study.
Compared to commensals, potential pathogens were less tolerant to glyphosate acid. Clostridium perfringens
and Staphylococcus aureus were both sensitive towards glyphosate IPA salt, Roundup and POEA, whereas
Escherichia coli K88, Salmonella enterica Enteritidis and S. enterica Typhimurium were tolerant towards all
treatments. In contrast, Shehata et al. (2013b), found E. coli K88 to be sensitive towards Roundup, but, as we
reported, they observed a high tolerance of S. enterica Enteritidis and S. enterica Typhimurium towards
Roundup, with MIC values of 5.00 mg/ml (Shehata et al., 2013b). Escherichia and Salmonella are closely
related, showing a genomic hybridization of about 50 % (Madigan et al., 2009). This could explain the
relatively similar results found in our study. Further, species belonging to Escherichia and Salmonella are
gram-negative, and in addition, E. coli K88, S. enterica Enteritidis and S. enterica Typhimurium are the only
gram-negative bacteria, tested in our study. Even though we do not know the exact details of how glyphosate
enter bacterial cells, these results indicate that gram-negative cell wall is more protective, than gram-
positive, and therefore considerably more difficult to penetrate. In general, most pathogens are gram-
negative bacteria (Madigan et al., 2009).
Interactions between commensals and potential pathogens
Adhesion of bacteria to intestinal mucosa is usually a prerequisite for colonization in the gut, as to the
continuously moving environment. Lactic acid bacteria are able to compete for adhesion with many
pathogenic species and their production of different antimicrobial substances is observed as mechanisms to
suppress growth of pathogens (Hao and Lee, 2004) including Salmonella, E. coli and Clostridia (Blomberg et
al., 1993; Bomba et al., 1997; Coconnier-Polter et al., 2005; Naber et al., 2004; Yun et al., 2009). Therefore,
an increased colonization of pathogens can indirectly be due to a decrease in the proliferation of Lactic acid
bacteria. Enterococcus faecalis, E. faecium and Bifidobacterium badius has the ability to inhibit BoNT
production, while reducing growth of Clostridium botulinum (A, B, D and E) and Krüger et al. (2013b) observed
that both glyphosate acid and Roundup were inhibitory to E. faecalis, at considerably lower concentrations,
by which they inhibited growth of C. botulinum Type B. Even though we did not succeed in culturing E.
faecalis, and therefore not were able to confirm or invalidate the results by Krüger et al. (2013b), a higher
sensitivity of glyphosate towards E. faecalis, could lead to increased proliferation of C. botulinum. Both due

41
to the loss of antagonistic effect of E. faecalis, against C. botulinum and to the tolerance of glyphosate
observed by C. botulinum itself.
Also, bifidobacteria can create unfavorable conditions for the colonization of Salmonella in the gut (Bielecka
et al., 1998), and therefore, sensitivity of B. adolescentis, towards glyphosate IPA salt, Roundup and POEA, in
our study, has the ability to increase proliferation of Salmonella, indirectly. As Salmonella strains themselves,
were tolerant towards all treatments in our study, addition of glyphosate has the possibility to create
disturbances in gut microbiota, in potential favor of some tolerant pathogens.
Overall effects of glyphosate depending on chemical formulation
In general, all bacteria displayed highest tolerance towards glyphosate acid. However, highest sensitivities
were observed for all gram-positive bacteria, when glyphosate was formulated as the IPA salt. To my
knowledge, no other studies on bacterial cultures have been conducted, investigated effects of glyphosate
IPA salt. However, in a study on cell cultures from the frog, Xenopus laevis, glyphosate IPA salt showed the
most severe effects, and at much lower levels, than the addition of glyphosate acid or Roundup did (Hedberg
and Wallin, 2010). Different toxicities could be explained by the fact, that more glyphosate enters plant cells,
when formulated as the IPA salt (Nalewaja et al., 1996). High tolerance observed for glyphosate acid,
compared to glyphosate IPA salt, might be attributed to its difficulties in crossing cell membranes, due to its
anionic character above pH 2.3 (Figure 1). Even though, glyphosate IPA salt exist as two separate ions in
aqueous solution, they form an ion-pair, when crossing a lipohilic environment, as a plasma membrane in
plant cells (Krogh, 2016). This facilitates an easier entrance, due to the resulting overall, neutral charge
(Krogh, 2016). As far as I am concerned, the exact entrance mechanisms into bacterial cells for glyphosate
acid and glyphosate IPA, have not yet been revealed. However, some of the mechanisms observed in plant
cells, might be applicable, suggesting that, it is the chemistry of the two different glyphosate forms, which
explains difference in toxicity, as to difference in how easy they enter cells.
If we choose to disregard the effect of Roundup on E. faecium and R. albus, Roundup and POEA were almost
equally toxic, towards the rest of the gram-positive bacteria. As Roundup is made up of glyphosate IPA salt
and POEA, the sensitivity towards Roundup should be equal or, eventually, lower than sensitivity observed
for glyphosate IPA salt and POEA, independently. All concentration levels are calculated to correspond to
each other and, subsequently, to the content of Roundup, in order to facilitate comparisons. Even though we
ended up investigating a lower amount of POEA, than the one originally intended, it might not have led to
any misleading results, as POEA also can constitute less than 15 % of Roundup (Giesy et al., 2000; Sawada et
al., 1988). The reason, why glyphosate IPA salt were more toxic than Roundup, in most cases, could be
explained by the fact that some of the glyphosate, in Roundup, could exist as the acid, even though the
majority exist as the salt. Toxicity of glyphosate acid were low compared to glyphosate IPA salt, leaving the
sensitivity of Roundup a bit lower that the sensitivity of glyphosate IPA salt.
Influence of glyphosate IPA salt, Roundup and POEA on gut microbiota from pigs and cows
Objective 3
In addition to in vitro effects on individual bacteria, we investigated whether glyphosate IPA salt, Roundup
and POEA showed any effects on the composition and activity of gut microbiota, when added to stomach,
cecum and colon content from slaughter pigs and to rumen content from dairy cows. Glyphosate acid was
not included, as only a certain amount of samples could be handled. As glyphosate IPA salt is the form found
in Roundup, we chose to use that instead.

42
Stomach
Compared to cecum, colon and rumen, a lower fermentation rate was found in stomach, in our study.
However, VFA concentrations differed considerably between the stomach replicates, leading to exclusion of
results for 24 hours, due to unexplainable variation, between this time and the two remaining time points.
Theoretically, of the gut segments investigated, least fermentation takes place in stomach, due to acidic
conditions (Cunningham and Klein, 2007). pH is low usually lower than 3.6 (Yen, 2001), keeping bacterial
numbers low (Cunningham and Klein, 2007; McDonald et al., 2011). Most of the bacteria, inhabiting stomach,
are lactic acid bacteria (Cunningham and Klein, 2007; McDonald et al., 2011), as they, in general, are more
acid tolerant than other bacteria (Madigan et al., 2009). We were not able to detect any effects of glyphosate
IPA salt, Roundup or POEA, independent of concentrations, on levels of acetate and propionate, as well as
on production of lactate and on the cell count of lactic acid bacteria. The only significant effect was observed
for Roundup, which decreased pH, at 10 mg/ml. As no other effects was evident for Roundup, at the same
concentration, there was no evidence that lowering pH influenced any of the other parameters, measured in
stomach. As stomach content already is acidic and lactic acid bacteria are the main inhabitants, it makes
sense that lactate production, and lactic acid cell count, were not affected by a decrease in pH, due to their
high tolerance towards acid.
Cecum and colon
The main fermentation sites in pigs are cecum and colon, both compartments maintaining a big, complex
microbial population, composed of both aerobic and anaerobic bacteria (Bergman, 1990; Cunningham and
Klein, 2007).
As a starting point, both cecum and colon had the same production of total VFA’s, approximately 240
mmol/kg sample each. VFAs in cecum and colon have been measured in the range between 30-240 mmol/L,
but according to Bergman (1990), an average of 70 – 120 mmol/L is more common. However, we measured
VFA production over 24 hours, possibly explaining the higher level.
In cecum, the overall fermentation pattern changed after addition of glyphosate IPA salt and Roundup. Total
VFA concentrations decreased, while propionate levels increased, for both treatments. Whether changes
observed, were due to direct effects of glyphosate IPA salt and Roundup, or whether the effects were
mediated through a decrease in pH, can be discussed. Normally, pH is close to neutral in cecum and colon
(Cunningham and Klein, 2007) and tolerance towards fluctuations in pH, differs between bacterial groups. As
cellulolytic bacteria, preferentially producing acetate (McDonald et al., 2011; Sjaastad et al., 2010), are more
sensitive than amylolytic, mostly producing lactate and thereby propionate (McDonald et al., 2011; Sjaastad
et al., 2010), fermentation leading to a higher production of propionate would not be unlikely, if pH drops
(Sjaastad et al., 2010). However, decreased pH was probably not the reason for the changes we observed in
propionate levels, as the effects on propionate were evident before pH dropped, and as pH were below six,
independent of all concentrations. Results for POEA supports this theory, as it did not affect pH, while still
increasing relative levels of propionate.
Even though, only low levels of isobutyrate, valerate and isovalerate were present in cecum, levels increased
by glyphosate IPA salt and Roundup. As branched VFA’s only are produced from amino acids, they are
representatives of protein fermentation (Nery et al., 2012). Therefore, we might propose that glyphosate IPA
salt and Roundup, had the ability to stimulate activity of proteolytic bacteria, or that pH level indirectly
affected it. Another parameter, possibly changing fermentations patterns, could be degradation of

43
glyphosate by gut bacteria. Some soil microorganisms are able to degrade glyphosate, releasing nutrients
available for growth (Barry et al., 1992; Borggaard and Gimsing, 2008; Duke et al., 2012). Even though,
degradation has only been investigated in soils, it seems plausible that some gut bacteria, also have the
abilities to degrade glyphosate, releasing nutrients, possibly changing composition of microbiota.
In colon, only results for Roundup and POEA were included, as glyphosate acid and glyphosate IPA salt, only
were tested once, and therefore had to be excluded from the final dataset. Roundup did not have any effect
on total VFA concentration, but decreased acetate levels and increased propionate levels, meaning that
acetate levels were counterbalanced by the increase in propionate levels. As for cecum, Roundup decreased
pH and even though increase in propionate probably were independent of pH level, it cannot be rejected
that decrease in acetate correlates with a decrease in pH, as cellulolytic bacteria are pH sensitive (Sjaastad
et al., 2010). Even though relative proportions of isobutyrate, valerate and isovalerate were low, significant
increases were evident in presence of Roundup. Usually, the sum of isobutyrate, valerate and isovalerate
constitute less than 10 % of the VFA’s produced in colon, as most protein degradation takes place in stomach
and small intestine, and only a small amount escapes to colon, compared to undigestible carbohydrates
(Rasmussen et al., 1988). As for cecum, there might be an increase in proliferation of proteolytic bacteria as
to an increased fermentation of amino acids. In addition, some bacteria might be able to degrade glyphosate,
thereby changing fermentation patterns, as mentioned for cecum.
In addition to glyphosate IPA salt and Roundup, POEA only increased relative propionate levels. In several
experiments it has been concluded that POEA was main responsible for the toxicity observed for Roundup
(Benachour and Seralini, 2009; Folmar et al., 1979; Mesnage et al., 2013; Mesnage et al., 2014; Mitchell et
al., 1987; Servizi et al., 1987) but we were not able to reproduce these results. However, many of these
studies tested the effect on single cells, and here we have to do with a more complex system. In addition,
even though we are aware that POEA facilitates penetration of glyphosate to plants, we do not know how
POEA penetrate cells and how it exerts its mechanism, which would be on interest when explaining the
effects, present or missing, we observed here. The effects of glyphosate IPA salt and Roundup were observed
at same concentrations, in most cases, which implies that POEA did not contribute considerably to the effect
exerted by Roundup on cecum and colon content.
Other than propionate levels in cecum, which were affected at 1 mg/ml, the rest of the significant differences,
were not observed before addition of 10 mg/ml. Both concentrations are way above the calculated
theoretical, concentrations we presented in Table 4, meaning that fermentation patterns will not be affected
in vivo, based on our results.
Rumen
The rumen is the main fermentation site in cows, maintaining a big, complex microbial population, mostly
composed of anaerobic bacteria (Bergman, 1990; Cunningham and Klein, 2007).
In a feeding study, Huther et al. (2005) investigated the effect of Roundup on ruminal fermentation patterns
and they found total VFA concentrations ranging between 64 and 92 mmol/L, depending on time after
feeding. Our total VFA concentrations were slightly higher, increasing in the presence of glyphosate IPA salt
and Roundup. Acetate levels decreased for all treatments, even though effects of glyphosate IPA salt and
Roundup, were evident before POEA. No matter which mechanisms are involved in the changes, more
propionate were produced, at the expense of acetate, indicative of change in the overall fermentation
pattern, from cellulolytic - to amylolytic bacteria (McDonald et al., 2011; Sjaastad et al., 2010). Also in rumen,

44
we observed pH effects, but as mentioned in both cecum and colon, pH effects did not have any direct effects
on neither acetate – nor propionate levels. Normally, pH in rumen is close to neutral (Cunningham and Klein,
2007), but glyphosate IPA salt and Roundup both decreased pH, starting at 1 mg/ml. A drop in pH (below 6)
can inhibit the activity of cellulolytic bacteria, degrading structural carbohydrates as cellulose, hemicellulose
and pectin (McDonald et al., 2011; Sjaastad et al., 2010). However, even though pH decreased at 1 mg/ml, it
only decreased slightly, still ranging between 6.37 and 6.40. This pH level were too high to inhibit cellulolytic
bacteria, and as pH increased (even though only 0.03) in the presence of POEA, while acetate levels
decreased, the effect of pH on acetate levels, probably can be rejected. Levels of propionate also increased,
independent of pH, as observed in cecum and colon. As for cecum and colon, rumen might inhabit bacteria,
which are able to degrade glyphosate, possibly explaining some of the changes in fermentation patterns.
However, further studies needs to highlight this.
As no changes were observed for isobutyrate, valerate and isovalerate, fermentation of proteins were
unchanged, irrespective of treatment. In rumen, these three acids usually constitute less than 5 % of the total
VFA pool (Bergman, 1990), which also corresponds to our findings.
To my knowledge, few studies have been conducted, investigating the effect of glyphosate acid, on rumen
fermentation patterns. Reuter et al. (2007) were able to show that addition of more than 8.45 mg/ml
inhibited fermentation, and that 10.14 mg/ml decreased acetate, butyrate and total VFA concentrations,
compared to control. Concentrations of propionate fluctuated more, but after incubation for 48 hours,
propionate concentrations were higher than control, both at 1.69 mg/ml and at 10.14 mg/ml (Reuter et al.,
2007). Even though we used another form of glyphosate, we also observed decreases in acetate levels, as
well as increase in propionate levels. In addition, Huther et al. (2005) observed no changes in ruminal
fermentation pattern after addition of glyphosate acid at 1.67 mg/ml (Huther et al., 2005). Even though
rumen were the component where significant effects were evident at lowest concentrations, these
concentrations were considerably higher than the theoretical concentrations, we calculated in Table 5.
Overall, the effect of POEA was observed later than the effect of glyphosate IPA salt and Roundup, but the
effects were also greater here, than in cecum and colon. Again, several experiments concluded that POEA
was main responsible for the toxicity observed for Roundup, but we were not able to reproduce these results.
In addition to the missing effects of POEA, the effects of glyphosate IPA salt and Roundup were similar, in
most cases, which implies that POEA did not contribute considerably to the effect of Roundup.
Methane
Even though the results of methane production varied considerable, and no statistical analyses were
performed, one thing became clear: at 10 mg/ml, no methane were produced, independent of the
treatments. In addition, no production was observed at 1 mg/ml for Roundup. It has been suggested that a
feed ration, mainly composed of starch, lowers pH, which might inhibit the growth and activity of the
methanogens (Boadi et al., 2004; Martin et al., 2010). This could explain the effect we observed for Roundup
and glyphosate IPA salt, due to the lowering of pH at 10 mg/ml. Nevertheless, when comparing these to
POEA, other factors might be involved, as pH increases in the presence of POEA. On the other hand,
production of acetate and butyrate leads to methane production, while propionate decreases it (Cunningham
and Klein, 2007). The decreased acetate to propionate ratio, we observed, could also have led to the
inhibition of the methane production. However, as archaea also possesses the shikimate pathway (Bult et al.,

45
1996; Daugherty et al., 2001; Graham et al., 2001), glyphosate can have a direct, inhibbiting effect on
methanogens.
Influence of glyphosate IPA salt on growth of L. sobrius in stomach content from pigs
Objective 4
In addition to the direct effect of glyphosate on bacterial cultures, and the microflora, found in gut, we
wanted to investigate whether growth of a strain of Lactobacillus sobrius, in stomach content, was affected
by glyphosate IPA salt, when pH was held constant (pH=5). As this part of the experiment was conducted, as
a pilot study, no statistical tests were done.
In general, it seems like lactobacilli cell count were lowered at 10 mg/ml. Inhibition was due to the direct
effect of glyphosate IPA salt, and not pH, as it was constant, no matter concentration level of glyphosate IPA
salt. Changes in VFA’ concentrations were difficult to interpret, and no other clear effects, than an increase
in lactate – and succinate production over time, were evident. When L. sobrius were tested in lab media, the
MIC value for glyphosate acid were 0.23 mg/ml, considerably lower than the effect, observed here. The
differences between lab media and stomach content could, most likely, have had an influence on the effects
observed, as we do not know the exact composition of stomach content.
Comparisons of the three different experiments
Even though, especially glyphosate IPA salt, Roundup and POEA were able to inhibit growth of some bacterial
cultures, at considerably low concentrations, we were not able to demonstrate the same effects on gut
contents from pigs and cows, as well as on a single culture in stomach content. In cecum, colon and rumen
content, activity of microorganisms were measured through VFA production; the first effects of glyphosate
IPA salt and Roundup were observed at 1 mg/ml, and effects of POEA at 10 mg/ml. In addition, possible
effects of glyphosate IPA salt on L. sobrius, in stomach content, were observed at a considerably higher
concentration, than effects, observed in lab media. Some of the bacterial MIC - and MEC values found, would
be realistic to observe in vivo, based on the theoretical calculations made. However, none of the
concentrations, by which we observed an effect on gut content, can be found in vivo, at least not due to our
assumptions and calculations. This indicates that gut microbiota is complex, and that effects of treatments,
on individual bacteria in lab media, cannot be transferred to affecting bacteria in the same way, when present
in gut, as to a complex interaction pattern between the microorganisms (Madigan et al., 2009; McDonald et
al., 2011; Sjaastad et al., 2010). However, even though in vivo concentrations, would not affect activities of
microbiota, some changes in composition could be evident. Sequencing samples would have been helpful to
highlight, possible differences in composition of microbiota, but due to time limitations, activity was only
included in this experiment.
When tested on individual strains, bacteria were more or less equally sensitive towards glyphosate IPA salt,
Roundup and POEA. For glyphosate IPA salt and Roundup, these effects were also observable in gut contents,
even though concentrations, where effects were evident, increased considerable. However, when tested on
gut content, POEA was least toxic of all treatments. Compared to glyphosate, we do not know much about
the exact composition of POEA and therefore it is not clear, how it enters cells and exerts its mechanisms.
Other studies should be performed to investigate this further.
Even though pH decreased in both stomach, cecum, colon and rumen for glyphosate IPA salt and/or
Roundup, results for L. sobrius in stomach content, indicate that glyphosate IPA salt exert its mechanisms,

46
independent of pH, as it was held constant. However, as this experiment was only conducted, as a pilot study,
replicating the experiment could be advantageously.
Additional work, not included in the thesis
In total, I have run 73 plates and presented results from 44, in the thesis. The reason why I chose to exclude
the others is that they were used to investigate which glyphosate acid concentrations that were appropriate
to test, how the plates could be prepared as uniform as possible and which bacteria it was possible to grow
in the conditions, we were able to supply. As mentioned, some bacteria would not grow, even though they
were tested in different media.
To test the treatments on animal digesta, a pilot study was performed. This was both done to test how the
protocol worked, and at which concentrations, and in which compartments, it was relevant to further
investigate the effect of the treatments. To begin with, glyphosate acid was used instead of glyphosate IPA
salt, and this is the reason why there are missing values for glyphosate IPA salt in some of the datasets. In
total, five trials were performed with digesta from slaughtered pigs.

47
Conclusion Overall, we showed that glyphosate acid, glyphosate IPA salt, Roundup and POEA all had the ability to inhibit
bacterial growth, even though MIC values differed between treatments and bacteria (objective 1). In
addition, inhibiting effects of glyphosate acid, glyphosate IPA salt, Roundup and POEA differed between
gram-positive and gram-negative bacteria; gram-negative being more tolerant towards all treatments. We
cannot conclude that commensals and potential pathogens are affected differently, just due to this division,
but as most gram-negative bacteria are pathogens, this is a more reasonable explanation for observed
differences between commensals and potential pathogens. Instead of differentiating between commensals
and potential pathogens, differentiation between gram-positive and gram-negative would be a more correct
determination, according to our findings (objective 2).
Other than affecting growth of bacteria directly, glyphosate also has the potential to affect growth indirectly.
Some commensals are able to suppress growth of some potential pathogens, and if the commensals are
sensitive towards glyphosate, and the pathogens tolerant, addition of glyphosate has the potential to
eliminate the inhibiting effect, the commensal has towards the potential pathogen. This, in addition to the
direct effect of glyphosate, can create disturbances in the gut, possibly leading to health problems for the
animal.
Glyphosate IPA salt, Roundup and POEA, were able to change fermentation patterns in cecum, colon and
rumen, indicative of changes in microbial activity, even though effects failed to appear in stomach (objective
3). The earliest effects were evident at 1 mg/ml being considerably higher than the possible concentrations
we calculated theoretically, in the different compartments. Overall, addition of glyphosate IPA salt and
Roundup led to more changes, than POEA did.
Even though the results of methane production in rumen varied considerable, and no statistical analyses
were performed, no methane were produced at 10 mg/ml independent of the treatments. In addition, no
production was observed at 1 mg/ml for Roundup. The decreased acetate to propionate ratio, observed for
all treatments in rumen, could have led to the inhibition, but a decrease of pH might also play a role for
glyphosate IPA salt and Roundup.
The effect of glyphosate IPA salt on the growth of L. sobrius in stomach content, at pH 5, was investigated in
a pilot study. Due to time limitations, the experiment was only performed ones, and therefore, data were
not analyzed statistically. Overall, it seemed like, lactic acid cell count were lowered at 10 mg/ml, and no
other parameters were affected. However, the experiment have to be repeated, to detect eventual
differences (objective 4).
Overall, fermentation is the action of the entire microbial biomass and need to be considered like an overall
process, not focusing on individuals (Cunningham and Klein, 2007; McDonald et al., 2011). This means that
even though we found considerably effects of glyphosate IPA salt, Roundup and POEA, when treating
individual bacterial strains, effects on the overall microbiota, observed in the second part of the experiment,
is more important, as it better reflects how different treatments affect microbiota in vivo.

48
In conclusion, we have been able to accept, all the four of our working hypothesis. First of all, glyphosate
(Roundup) suppressed growth of gut bacteria, different gut bacteria had different susceptibilities towards
glyphosate (Roundup), glyphosate (Roundup) changed fermentation pattern in the gut of pigs and dairy cows
and the effect of glyphosate depended on the chemical formulation (acid, salt, mixture and surfactant) used.

49
Perspectives and future considerations Even though glyphosate does not have the ability to affect animals directly, it is now evident, through
different studies, that it has the potential to affect microbiota, and thereby the animals, they inhabits.
As our results were not conclusive, it would be interesting to further investigate, whether the effects of
glyphosate IPA salt and Roundup in gut contents, are attributed to the treatments directly, or indirectly, due
to a lowering of pH by the compounds. This can be done in bioreactors, by either keeping pH, or
concentrations, constants. In addition to VFA measurements, sequencing would help determine eventual
changes in microbial composition.
In this study, glyphosate etc. was added directly to gut contents. However, pigs and cows are not fed
glyphosate directly; plants, which have been sprayed with glyphosate, are fed to them. This could influence
the effects in vivo, and therefore, feeding trials should be performed, to give a better indication of how, and
if, the animals are affected. In addition, feeding trials has the ability to highlight long-term effect, as it is
possible that microorganisms can develop resistance towards glyphosate or that glyphosate, on the other
hand has an additive effect, increasing toxicity, the longer the period the animal are fed feed, containing
residues. Effects, of different residue levels, should also be determined. Other than measuring microbial
activity and composition, animal performance parameters are important to include. Through the effect on
microbiota, glyphosate can possibly effect livestock productions, which are the main interest for farmers.

50
References Ackermann, W., Coenen, M., Schrodl, W., Shehata, A. A., and Kruger, M. (2015). The Influence of
Glyphosate on the Microbiota and Production of Botulinum Neurotoxin During Ruminal Fermentation. Current Microbiology 70, 374-382.
Ahle, J. L. (1985). Enhancement of herbicidal activity of tetraaluminum salts of N-phosphonomethylglycine. Google Patents.
Anton, D. L., Hedstrom, L., Fish, S. M., and Abeles, R. H. (1983). Mechanism of Enolpyruvyl Shikimate-3-phosphate Synthase Exchange of Phosphoenolpyruvate with Solvent Protons Biochemistry 22, 5903-5908.
ArgenBio (2015). Gráfico de evolución de las superficies sembradas con OGM en la Argentina, en porcentajes (Graph showing the areas planted with GM crops in Argentina). http://www.argenbio.org/adc/uploads/imagenes_doc/planta_stransgenicas/1_Grafico_de_evolucion_de_superficie_cultivada_OGM_en_Arg_en_porcentaje.pdf.
Arikan, S., Lozano-Chiu, M., Paetznick, V., and Rex, J. H. (2001). In Vitro Susceptibility Testing Methods for Caspofungin against Aspergillus and Fusarium Isolates. Antimicrobial Agents and Chemotherapy 45, 327-330.
Arregui, M. C., Lenardon, A., Sanchez, D., Maitre, M. I., Scotta, R., and Enrique, S. (2004). Monitoring glyphosate residues in transgenic glyphosate-resistant soybeans. Pest Management Science 60, 163-166.
Barry, G., Kishore, G., Padgette, S., Taylor, M., Kolacz, K., Weldon, M., Re, D., Eichholtz, D., Fincher, K., and Hallas, L. (1992). Inhibitors of amino acid biosynthesis: Strategies for imparting glyphosate tolerance to crop plants. In "Current Topics in Plant Physiology an American Society of Plant Physiologists Series; Biosynthesis and molecular regulation of amino acids in plants" (B. K. Singh, H. E. Flores and J. C. Shannon, eds.), Vol. 7, pp. 139-145.
Barry, G. F., Kishore, G. M., Padgette, S. R., and Stallings, W. C. (1997). Glyphosate-tolerant 5-enolpyruvylshikimate-3-phosphate synthases. pp. 2616.
Benachour, N., and Seralini, G. E. (2009). Glyphosate Formulations Induce Apoptosis and Necrosis in Human Umbilical, Embryonic, and Placental Cells. Chemical Research in Toxicology 22, 97-105.
Benbrook, C. M. (2012). Impacts of genetically engineered crops on pesticide use in the US - the first sixteen years. Environmental Sciences Europe 24, 24.
Bentley, R. (1990). The Shikimate Pathway - A Metabolic Tree with Many Branches. Critical Reviews in Biochemistry and Molecular Biology 25, 307-384.
Bergman, E. N. (1990). Energy contributions of volatile fatty acids from the gastrointestinal tract in various species. Physiological Reviews 70, 567-590.
Bielecka, M., Biedrzycka, E., Biedrzycka, E., Smoragiewicz, W., and Smieszek, M. (1998). Interaction of Bifidobacterium and Salmonella during associated growth. International Journal of Food Microbiology 45, 151-155.
Binimelis, R., Pengue, W., and Monterroso, I. (2009). "Transgenic treadmill": Responses to the emergence and spread of glyphosate-resistant johnsongrass in Argentina. Geoforum 40, 623-633.
Blachier, F., Mariotti, F., Huneau, J. F., and Tome, D. (2007). Effects of amino acid-derived luminal metabolites on the colonic epithelium and physiopathological consequences. Amino Acids 33, 547-562.
Blomberg, L., Henriksson, A., and Conway, P. L. (1993). Inhibition of Adhesion of Escherichia-Coli K88 to Piglet Ileal Mucus by Lactobacillus spp. Applied and Environmental Microbiology 59, 34-39.
Boadi, D., Benchaar, C., Chiquette, J., and Massé, D. (2004). Mitigation strategies to reduce enteric methane emissions from dairy cows: Update review. Canadian Journal of Animal Science 84, 319-335.
Bode, R., Ramos, C. M., and Birnbaum, D. (1984). Inhibition of tyrosine-sensitive 3-deoxy-D-arabino-heptulosonate 7-phosphate synthase by glyphosate in Candida maltosa Fems Microbiology Letters 23, 7-10.

51
Bøhn, T., Cuhra, M., Traavik, T., Sanden, M., Fagan, J., and Primicerio, R. (2014). Compositional differences in soybeans on the market: Glyphosate accumulates in Roundup Ready GM soybeans. Food Chemistry 153, 207-215.
Böhnel, H., Schwagerick, B., and Gessler, F. (2001). Visceral botulism: A new form of bovine Clostridium botulinum toxication. Journal of Veterinary Medicine Series A 48, 373-383.
Bomba, A., Kravjansky, I., Kastel, R., Herich, R., Juhasova, Z., Cizek, M., and Kapitancik, B. (1997). Inhibitory effects of Lactobacillus casei upon the adhesion of enterotoxigenic Escherichia coli K99 to the intestinal mucosa in gnotobiotic lambs. Small Ruminant Research 23, 199-206.
Boocock, M. R., and Coggins, J. R. (1983). Kinetics of 5-enolpyruvylshikimate-3-phosphate synthase inhibition by glyphosate. Febs Letters 154, 127-133.
Borggaard, O. K., and Gimsing, A. L. (2008). Fate of glyphosate in soil and the possibility of leaching to ground and surface waters: a review. Pest Management Science 64, 441-456.
Brausch, J. M., and Smith, P. N. (2007). Toxicity of three polyethoxylated tallowamine surfactant formulations to laboratory and field collected fairy shrimp, Thamnocephalus platyurus. Archives of Environmental Contamination and Toxicology 52, 217-221.
Budavari, S. (1996). "The Merck Index. An Encyclopedia of Chemicals, Drugs and Biologicals. Twelth Edition. ," 12th/Ed. Merck & Co. Inc, Whitehouse Station, N.J. : Merck & Co. Inc., 1996.
Bult, C. J., White, O., Olsen, G. J., Zhou, L. X., Fleischmann, R. D., Sutton, G. G., Blake, J. A., FitzGerald, L. M., Clayton, R. A., Gocayne, J. D., Kerlavage, A. R., Dougherty, B. A., Tomb, J. F., Adams, M. D., Reich, C. I., Overbeek, R., Kirkness, E. F., Weinstock, K. G., Merrick, J. M., Glodek, A., Scott, J. L., Geoghagen, N. S. M., Weidman, J. F., Fuhrmann, J. L., Nguyen, D., Utterback, T. R., Kelley, J. M., Peterson, J. D., Sadow, P. W., Hanna, M. C., Cotton, M. D., Roberts, K. M., Hurst, M. A., Kaine, B. P., Borodovsky, M., Klenk, H. P., Fraser, C. M., Smith, H. O., Woese, C. R., and Venter, J. C. (1996). Complete genome sequence of the methanogenic archaeon, Methanococcus jannaschii. Science 273, 1058-1073.
Busse, M. D., Ratcliff, A. W., Shestak, C. J., and Powers, R. F. (2000). Non-Target Effects of Glyphosate on Soil Microbes. Proceedings of the California Weed Science Society 52, 146-150.
Cessna, A. J., Darwent, A. L., Kirkland, K. J., Townleysmith, L., Harker, K. N., and Lefkovitch, L. P. (1994). Residues of glyphosate and its metabolite AMPA in wheat seed and foilage following preharvest applications. Canadian Journal of Plant Science 74, 653-661.
Chamberlain, K., Evans, A. A., and Bromilow, R. H. (1996). 1-Octanol/Water Partition Coefficient (Kow) and pKa for Ionisable Pesticides Measured by apH-Metric Method. Pesticide Science 47, 265-271.
Chen, C. Y., Nace, G. W., and Irwin, P. L. (2003). A 6 x 6 drop plate method for simultaneous colony counting and MPN enumeration of Campylobacter jejuni, Listeria monocytogenes, and Escherichia coli. Journal of Microbiological Methods 55, 475-479.
Clair, E., Linn, L., Travert, C., Amiel, C., Seralini, G., and Panoff, J. (2012). Effects of Roundup (R) and Glyphosate on Three Food Microorganisms: Geotrichum candidum, Lactococcus lactis subsp. cremoris and Lactobacillus delbrueckii subsp. bulgaricus. Current Microbiology 64, 486-491.
Coconnier-Polter, M. H., Moal, V. L. L., and Servin, A. L. (2005). A Lactobacillus acidophilus strain of human gastrointestinal microbiota origin elicits killing of enterovirulent Salmonella enterica serovar typhimurium by triggering lethal bacterial membrane damage. Applied and Environmental Microbiology 71, 6115-6120.
Codex Alimentarius, C. (2012). Report of the 44th session of the Codex Committee on Pesticide Residues. FAO/WHO Food Standards Programme REP12/PR.
Commission, E. (2002). "Review report for the active substance glyphosate. Glyphosate 6511/VI/99-final," http://ec.europa.eu/food/plant/protection/evaluation/existactive/list1_glyphosate_en.pdf.
Commission, E. (2011). "Questions and answers on the evaluation of the European Union's GMO legislation," European Commission, Press Release Database.
Commission, E. (2015). EU Pesticide Database - Current MRL values. Vol. 2015. European Commission, http://ec.europa.eu/food/plant/pesticides/eu-pesticides-database/public/?event=pesticide.residue.CurrentMRL&language=EN.

52
Cunningham, J. G., and Klein, B. G. (2007). "Textbook of Veterinary Physiology," Saunders Elsevier. Daugherty, M., Vonstein, V., Overbeek, R., and Osterman, A. (2001). Archaeal shikimate kinase, a new
member of the GHMP-kinase family. Journal of Bacteriology 183, 292-300. Davison, J. (2010). GM plants: Science, politics and EC regulations. Plant Science 178, 94-98. Della-Cioppa, G., Bauer, S. C., Klein, B. K., Shah, D. M., Fraley, R. T., and Kishore, G. M. (1986). Translocation
of the precursor of 5-enolpyruvylshikimate-3-phosphate synthase into chloroplasts of higher plants in vitro. Proceedings of the National Academy of Sciences of the United States of America 83, 6873-6877.
Denis, M. H., and Delrot, S. (1993). Carrier-mediated uptake of glyphosate in broad bean (Vicia faba) via a phosphate transporter. Physiologia Plantarum 87, 569-575.
Diamond, G. L., and Durkin, P. R. (1997). "Effects of Surfactants on the Toxicity of Glyphosate, with Specific Reference to RODEO."
Dick, R. E., and Quinn, J. P. (1995). Glyphosate-degrading isolated from environmental samples: occurence and pathways of degradation. Applied Microbiology and Biotechnology 43, 545-550.
Dill, G. M. (2005). Glyphosate-resistant crops: history, status and future. Pest Management Science 61, 219-224.
Djursing, T. (2013). Landmænd sprøjter med Roundup kort før høst for at modne korn. Ingeniøren, http://ing.dk/artikel/landmaend-sproejter-med-roundup-kort-foer-hoest-modne-korn-161055.
Doy, C. H., and Gibson, F. (1961). Formation of 4-Hydroxyphenylpyruvic Acid and Phenylpyruvic Acid by Tryptophan Auxotrophs and Wild-type Aerobacter Aerogenes Considered in Relation to General Aromatic Pathway. Biochimica Et Biophysica Acta 50, 495-&.
Duke, S. O., Lydon, J., Koskinen, W. C., Moorman, T. B., Chaney, R. L., and Hammerschmidt, R. (2012). Glyphosate Effects on Plant Mineral Nutrition, Crop Rhizosphere Microbiota, and Plant Disease in Glyphosate-Resistant Crops. Journal of Agricultural and Food Chemistry 60, 10375-10397.
Duke, S. O., and Powles, S. B. (2008). Glyphosate: a once-in-a-century herbicide. Pest Management Science 64, 319-325.
Duke, S. O., and Powles, S. B. (2009). Glyphosate-resistant crops and weeds: now and in the future. AgBioForum 12, 346-357.
EFSA (2015a). Conclusion on the peer review of the pesticide risk assessment of the active substance glyphosate. EFSA Journal 13.
EFSA (2015b). Maximum residue levels. Vol. 2015. EFSA, http://www.efsa.europa.eu/en/pesticides/mrls. EPA, U. S. (1985). Consensus Review of Glyphosate. Eschenburg, S., Healy, M. L., Priestman, M. A., Lushington, G. H., and Schönbrunn, E. (2002). How the
mutation glycine96 to alanine confers glyphosate insensitivity to 5-enolpyruvyl shikimate-3-phosphate synthase from Escherichia coli. Planta 216, 129-135.
FAO (2009). Submission and evaluation of pesticide residues data for the estimation of maximum residue levels in food and feed. FAO Plant Production and Protection Paper, xi + 264 pp.
FAO (2015). FAOSTAT data. Vol. 2015, http://faostat3.fao.org/browse/Q/QC/E. Folmar, L. C., Sanders, H. O., and Julin, A. M. (1979). Toxicity of the herbicide glyphosate and several of its
formulations to fish and aquatic invertebrates. Archives of Environmental Contamination and Toxicology 8, 269-278.
Fon, T. A. D., and Uhing, E. H. (1964). Aminomethylenephosphinic acids, salts thereof, and process for their production. Google Patents, Stauffer Chemical Co.
Franz, J. E. (1974). N-phosphonomethyl-glycine phytotoxicant compositions. Google Patents. Franz, J. E., Mao, M. K., and Sikorski, J. A. (1997). "Glyphosate: a unique global herbicide." Fredborg, M., Andersen, K. R., Jorgensen, E., Droce, A., Olesen, T., Jensen, B. B., Rosenvinge, F. S., and
Sondergaard, T. E. (2013). Real-Time Optical Antimicrobial Susceptibility Testing. Journal of Clinical Microbiology 51, 2047-2053.

53
Funke, T., Han, H., Healy-Fried, M. L., Fischer, M., and Schönbrunn, E. (2006). Molecular Basis for the herbicide resistance of Roundup Ready Crops. Proceedings of the National Academy of Sciences of the United States of America 103, 13010-13015.
Funke, T., Yang, Y., Han, H., Healy-Fried, M., Olesen, S., Becker, A., and Schönbrunn, E. (2009). Structural Basis of Glyphosate Resistance Resulting from the Double Mutation Thr(97) -> Ile and Pro(101) -> Ser in 5-Enolpyruvylshikimate-3-phosphate Synthase from Escherichia coli. Journal of Biological Chemistry 284, 9854-9860.
Ganson, R. J., and Jensen, R. A. (1988). The Essential Role of Cobalt in the Inhibition of the Cytosolic lsozyme of 3-Deoxy-D-arabino-heptulosonate-7-phosphate Synthase from Nicotiana silvestris by Glyphosate. Archives of Biochemistry and Biophysics 260, 85-93.
Gasnier, C., Dumont, C., Benachour, N., Clair, E., Chagnon, M. C., and Seralini, G. E. (2009). Glyphosate-based herbicides are toxic and endocrine disruptors in human cell lines. Toxicology 262, 184-191.
Gianessi, L. P. (2005). Economic and herbicide use impacts of glyphosate-resistant crops. Pest Management Science 61, 241-245.
Gibson, F., and Jackman, L. M. (1963). Structure of Chorismic Acid, a New Intermediate in Aromatic Biosynthesis. Nature 198, 388-&.
Gibson, M. I., and Gibson, F. (1962). New Intermediate in Aromatic Biosynthesis Biochimica Et Biophysica Acta 65, 160-&.
Gibson, M. I., Gibson, F., Doy, C. H., and Morgan, P. N. (1962). The Branch Point in the Biosynthesis of the Aromatic Amino-Acids. Nature 195, 1173-1175.
Giesy, J. P., Dobson, S., and Solomon, K. R. (2000). Ecotoxicological risk assessment for Roundup (R) Herbicide. In "Reviews of Environmental Contamination and Toxicology, Vol 167" (G. W. Ware, ed.), Vol. 167, pp. 35-120.
Gout, E., Bligny, R., and Douce, R. (1992). Regulation of intracellular pH values in higher plant cells. Carbon-13 and phosphorus-31 nuclear magnetic resonance studies. J Biol Chem 267, 13903-9.
Graham, D. E., Kyrpides, N., Anderson, I. J., Overbeek, R., and Whitman, W. B. (2001). Genome of Methanocaldococcus (Methanococcus) jannaschii. In "Methods in Enzymology. Hyperthermophilic enzymes: Part A" (M. W. W. Adams and R. M. Kelly, eds.), Vol. 330, pp. 40-123.
Graham, J. A., Lennon, P. J., Wright, D. R., Brinker, R. J., Mahoney, M. D., Pallas, N. R., Becher, D. Z., Wideman, A. S., Agbaje, H. E., and Xu, X. C. (2006). Aqueous pesticide formulations and novel surfactants. Google Patents.
Grube, A., Donaldson, D., Kiely, T., and Wu, L. (2011). Pesticides Industry Sales and Usage 2006 and 2007 Market Estimates. Pesticides Industry Sales and Usage Report, 41 pp.
Guyton, K. Z., Loomis, D., Grosse, Y., El Ghissassi, F., Benbrahim-Tallaa, L., Guha, N., Scoccianti, C., Mattock, H., Straif, K., and Int Agcy Res Canc Monog, W. (2015). Carcinogenicity of tetrachlorvinphos, parathion, malathion, diazinon, and glyphosate. Lancet Oncology 16, 490-491.
Hao, W. L., and Lee, Y. K. (2004). Microflora of the gastrointestinal tract: a review. Methods Mol Biol 268, 491-502.
Hart, M. R., and Brookes, P. C. (1996). Soil microbial biomass and mineralisation of soil organic matter after 19 years of cumulative field applications of pesticides. Soil Biology and Biochemistry 28, 1641-1649.
Healy-Fried, M. L., Funke, T., Priestman, M. A., Han, H., and Schonbrunn, E. (2007). Structural basis of glyphosate tolerance resulting from mutations of Pro(101) in Escherichia coli 5-enolpyruvylshikimate-3-phosphate synthase. Journal of Biological Chemistry 282, 32949-32955.
Heap, I. (2015). Glyphosate Resistant Weeds. In "The International Survey of Herbicide Resistant Weeds", http://weedscience.org/summary/moa.aspx?MOAID=12.
Hedberg, D., and Wallin, M. (2010). Effects of Roundup and glyphosate formulations on intracellular transport, microtubules and actin filaments in Xenopus laevis melanophores. Toxicology in Vitro 24, 795-802.
Herold, A. E., Beardmore, R. A., and Parrish, S. K. (2013). Herbicide compositions comprising suspension concentrate with glyphosate acid. Google Patents.

54
Herrmann, K. M. (1995). The Shikimate Pathway as an Entry to Aromatic Secondary Metabolism. Plant Physiology 107, 7-12.
Hetherington, P. R., Marshall, G., Kirkwood, R. C., and Warner, J. M. (1998). Absorption and efflux of glyphosate by cell suspensions. Journal of Experimental Botany 49, 527-533.
Hietanen, E., Linnainmaa, K., and Vainio, H. (1983). Effects of Phenolxyherbicides and Glyphosate on the hepatic and Intestinal Biotransformation Activities in the Rat. Acta Pharmacologica Et Toxicologica 53, 103-112.
Huther, L., Drebes, S., and Lebzien, P. (2005). Effect of glyphosate contaminated feed on rumen fermentation parameters and in sacco degradation of grass hay and corn grain. Archives of Animal Nutrition 59, 73-79.
IARC (2015). IARC Monographs Volume 112: evaluation of five organophosphate insecticides and herbicides. Vol. 2015. International Agency for Research on Cancer.
ISAAA (2014). "Biotech Facts and Trends - Brazil." ISAAA, https://www.isaaa.org/resources/publications/biotech_country_facts_and_trends/download/Facts%20and%20Trends%20-%20Brazil.pdf.
Kier, L. D. (2015). Review of genotoxicity biomonitoring studies of glyphosate-based formulations. Critical Reviews in Toxicology 45, 209-218.
Kishore, G. M., and Shah, D. M. (1988). Amino Acid Biosynthesis Inhibitors as Herbicides Annual Review of Biochemistry 57, 627-663.
Knuuttila, P., and Knuuttila, H. (1985). Molecular and crystalline structure of glyphosate. In "The Herbicide Glyphosate" (E. Grossbard and D. Atkinson, eds.), pp. 18-22. Buttersworth & Co.
Kremer, R., Means, N., and Kim, S. (2005). Glyphosate affects soybean root exudation and rhizosphere micro-organisms. International Journal of Environmental Analytical Chemistry 85, 1165-1174.
Krogh, S. (2016). Personal Communication. Krüger, M., Schrödl, W., Neuhaus, J., and Shehata, A. A. (2013a). Field Investigations of Glyphosate in Urine
of Danish Dairy Cows. J. Environ. Anal. Toxicol. 3, 1000186. Krüger, M., Shehata, A. A., Schroedl, W., and Rodloff, A. (2013b). Glyphosate suppresses the antagonistic
effect of Enterococcus spp. on Clostridium botulinum. Anaerobe 20, 74-78. Lamb, D. C., Kelly, D. E., Hanley, S. Z., Mehmood, Z., and Kelly, S. L. (1998). Glyphosate is an inhibitor of
plant cytochrome P450: Functional expression of Thlaspi arvensae cytochrome P45071B1/Reductase fusion protein in Escherichia coli. Biochemical and Biophysical Research Communications 244, 110-114.
Lane, M., Lorenz, N., Saxena, J., Ramsier, C., and Dick, R. P. (2012). The effect of glyphosate on soil microbial activity, microbial community structure, and soil potassium. Pedobiologia 55, 335-342.
Lange, M., Westermann, P., and Ahring, B. K. (2005). Archaea in protozoa and metazoa. Applied Microbiology and Biotechnology 66, 465-474.
Lebrun, M., Sailland, A., Freyssinet, G., and DeGryse, E. (2003). Mutated 5-enolpyruvylshikimate-3-phosphate synthase, gene coding for said protein and transformed plants containing said gene.
Lee, T. T. (1980). Characteristics of glyphosate inhibition of growth in soybean and tobacco callus cultures. Weed Research 20, 365-369.
Leschine, S. B. (1995). Cellulose Degradation in Anaerobic Environments. Annual Review of Microbiology 49, 399-426.
Levin, J. G., and Sprinson, D. B. (1964). The Enzymatic Formation and Isolation of 3-enolpyruvylshikimate 5-phosphate Journal of Biological Chemistry 239, 1142-&.
Madigan, M. T., Martinko, J. M., Dunlap, P. V., and Clark, D. P. (2009). "Biology of Microorganisms," 12th/Ed. Pearson Education, Inc.
Maeda, H., and Dudareva, N. (2012). The Shikimate Pathway and Aromatic Amino Acid Biosynthesis in Plants. Annual Review of Plant Biology, Vol 63 63, 73-105.
Malik, J., Barry, G., and Kishore, G. (1989). The Herbicide Glyphosate. Biofactors 2, 17-26.

55
Martin, C., Morgavi, D. P., and Doreau, M. (2010). Methane mitigation in ruminants: from microbe to the farm scale. Animal 4, 351-365.
McCloskey, W. B. Effect of Surfactants and Adjuvants on Postemergens Herbicide Efficacy. Department of Plant Sciences, University of Arizona.
McDonald, P., Edwards, R. A., Greenhalgh, J. F. D., Morgan, C. A., Sinclair, L. A., and Wilkinson, R. G. (2011). "Animal Nutrition," 7th/Ed. Pearson Education.
Mekwatanakarn, P., and Sivasithamparam, K. (1987). Effect of certain herbicides on soil microbial populations and their influence on saprophytic growth in soil and pathogenicity of take-all fungus. Biology and Fertility of Soils 5, 175-180.
Mesnage, R., Bernay, B., and Seralini, G. E. (2013). Ethoxylated adjuvants of glyphosate-based herbicides are active principles of human cell toxicity. Toxicology 313, 122-128.
Mesnage, R., Defarge, N., Vendomois, J. S. d., and Seralini, G. E. (2014). Major pesticides are more toxic to human cells than their declared active principles. BioMed Research International 2014, Article ID 179691.
Mijangos, I., Becerril, J. M., Albizu, I., Epelde, L., and Garbisu, C. (2009). Effects of glyphosate on rhizosphere soil microbial communities under two different plant compositions by cultivation-dependent and -independent methodologies. Soil Biology and Biochemistry 41, 505-513.
Miljøstyrelsen (2013). "Bekæmpelsesstatistik 2012. Behandlingshyppighed og pesticidbelastning, baseret på salgsstatistik og sprøjtejournaldata." Miljøministeriet.
Miljøstyrelsen (2014). "Anvendelse af glyphosat før høst i korn." Miljøministeriet. Mitchell, D. G., Chapman, P. M., and Long, T. J. (1987). Acute Toxicity of Roundup and Rodeo Herbicides to
Rainbow Trout, Chinook, and Coho Salmon. Bulletin of Environmental Contamination and Toxicology 39, 1028-1035.
Monsanto (2015). Company History. Vol. 2015, http://www.monsanto.com/whoweare/pages/monsanto-history.aspx.
Naber, P., Smidt, I., Stsepetova, J., Brilene, T., Annuk, H., and Mikkelsaar, M. (2004). Inhibition of Clostridium difficile strains by intestinal Lactobacillus species. Journal of Medical Microbiology 53, 551-554.
Nalewaja, J. D., Devilliers, B., and Matysiak, R. (1996). Surfactant and salt affect glyphosate retention and absorption. Weed Research 36, 241-247.
Nelson, D. R., Koymans, L., Kamataki, T., Stegeman, J. J., Feyereisen, R., Waxman, D. J., Waterman, M. R., Gotoh, O., Coon, M. J., Estabrook, R. W., Gunsalus, I. C., and Nebert, D. W. (1996). P450 superfamily: Update on new sequences, gene mapping, accession numbers and nomenclature. Pharmacogenetics 6, 1-42.
Nery, J., Goudez, R., Biourge, V., Tournier, C., Leray, V., Martin, L., Thorin, C., Nguyen, P., and Dumon, H. (2012). Influence of dietary protein content and source on colonic fermentative activity in dogs differing in body size and digestive tolerance. Journal of Animal Science 90, 2570-2580.
Padgette, S. R., Kolacz, K. H., Delannay, X., Re, D. B., Lavallee, B. J., Tinius, C. N., Rhodes, W. K., Otero, Y. I., Barry, G. F., Eichholtz, D. A., Peschke, V. M., Nida, D. L., Taylor, N. B., and Kishore, G. M. (1995a). Development, Identification, and Characterization of a Glyphosate-Tolerant Soybean Line. Crop Science 35, 1451-1461.
Padgette, S. R., Re, D. B., Barry, G. F., Eichholtz, D. E., Delannay, X., Fuchs, R. L., Kishore, G. M., and Fraley, R. T. (1995b). New weed control opportunities: development of glyphosate-tolerant soybeans. In "Herbicide Resistant Crops" (S. O. Duke, ed.).
Paganelli, A., Gnazzo, V., Acosta, H., Lopez, S. L., and Carrasco, A. E. (2010). Glyphosate-Based Herbicides Produce Teratogenic Effects on Vertebrates by Impairing Retinoic Acid Signaling. Chemical Research in Toxicology 23, 1586-1595.
Park, H., Hilsenbeck, J. L., Kim, H. J., Shuttleworth, W. A., Park, Y. H., Evans, J. N., and Kang, C. H. (2004). Structural studies of Streptococcus pneumoniae EPSP synthase in unliganded state, tetrahedral intermediate-bound state and S3P-GLP-bound state. Molecular Microbiology 51, 963-971.

56
Partoazar, M., Hoodaji, M., and Tahmourespour, A. (2011). The effect of glyphosate application on soil microbial activities in agricultural land. African Journal of Biotechnology 10, 19419-19424.
Plantedirektoratet (2010). Undersøgelse af pesticider, mykotoksiner, bly og cadmium i sydamerikanske sojaprodukter til foder. 1-22.
Pline, W. A., Wilcut, J. W., Duke, S. O., Edmisten, K. L., and Wells, R. (2002). Tolerance and accumulation of shikimic acid in response to glyphosate applications in glyphosate-resistant and nonglyphosate-resistant cotton (Gossypium hirsutum L.). Journal of Agricultural and Food Chemistry 50, 506-512.
Poulsen, M., Schwab, C., Jensen, B. B., Engberg, R. M., Spang, A., Canibe, N., Højberg, O., Milinovich, G., Fragner, L., Schleper, C., Weckwerth, W., Lund, P., Schramm, A., and Urich, T. (2013). Methylotrophic methanogenic Thermoplasmata implicated in reduced methane emissions from bovine rumen. Nat Commun 4, 1428.
Rasmussen, H. S., Holtug, K., and Mortensen, P. B. (1988). Degradation of amino acids to short-chain fatty acids in humans. Scandinavian Journal of Gastroenterology 23, 178-182.
Ratcliff, A. W., Busse, M. D., and Shestak, C. J. (2006). Changes in microbial community structure following herbicide (glyphosate) additions to forest soils. Applied Soil Ecology 34, 114-124.
Raven, P. H., Evert, R. F., and Eichhorn, S. E. (2005). "Biology of Plants," 7th/Ed. Sara Tenney, W. H. Freeman and Company.
Relyea, R. A. (2005). The lethal impact of roundup on aquatic and terrestrial amphibians. Ecological Applications 15, 1118-1124.
Reuter, T., Alexander, T. W., Martinez, T. F., and McAllister, T. A. (2007). The effect of glyphosate on digestion horizontal gene transfer during in vitro ruminal fermentation of genetically modified canola. Journal of the Science of Food and Agriculture 87, 2837-2843.
Richard, S., Moslemi, S., Sipahutar, H., Benachour, N., and Seralini, G. E. (2005). Differential effects of glyphosate and roundup on human placental cells and aromatase. Environmental Health Perspectives 113, 716-720.
Richardson, A. J., Calder, A. G., Stewart, C. S., and Smith, A. (1989). Simultaneous determination of volatile and non-volatile acidic fermentation products of anaerobes by capillary gas chromatography. Letters in Applied Microbiology 9, 5-8.
Roberts, F., Roberts, C. W., Johnson, J. J., Kyle, D. E., Krell, T., Coggins, J. R., Coombs, G. H., Milhous, W. K., Tzipori, S., Ferguson, D. J. P., Chakrabarti, D., and McLeod, R. (1998). Evidence for the shikimate pathway in apicomplexan parasites. Nature 393, 801-805.
Rubin, J. L., Gaines, C. G., and Jensen, R. A. (1982). Enzymological Basis for Herbicidal Action of Glyphosate Plant Physiology 70, 833-839.
Samsel, A., and Seneff, S. (2013). Glyphosate's Suppression of Cytochrome P450 Enzymes and Amino Acid Biosynthesis by the Gut Microbiome: Pathways to Modern Diseases. Entropy 15, 1416-1463.
Sankula, S. (2006). "Quantification of the Impacts on US Agriculture of Biotechnology-Derived Crops Planted in 2005." National Center for Food and Agricultural Policy, www.ncfap.org.
Santos, A., and Flores, M. (1995). Effects of glyphosate on nitrogen fixation of free-living heterotrophic bacteria Letters in Applied Microbiology 20, 349-352.
Sawada, Y., Nagai, Y., Ueyama, M., and Yamamoto, I. (1988). Probable Toxicity of Surface-Active Agent in Commercial Herbicide Containing Glyphosate. Lancet 1, 299-299.
Schönbrunn, E., Eschenburg, S., Shuttleworth, W. A., Schloss, J. V., Amrhein, N., Evans, J. N. S., and Kabsch, W. (2001). Interaction of the herbicide glyphosate with its target enzyme 5-enolpyvuvylshikimate 3-phosphate synthase in atomic detail. Proceedings of the National Academy of Sciences of the United States of America 98, 1376-1380.
Schrödl, W., Krüger, S., Konstantinova-Muller, T., Shehata, A. A., Rulff, R., and Krüger, M. (2014). Possible Effects of Glyphosate on Mucorales Abundance in the Rumen of Dairy Cows in Germany. Current Microbiology 69, 817-823.
Servizi, J. A., Gordon, R. W., and Martens, D. W. (1987). Acute Toxicity of Garlon 4 and Roundup Herbicides to Salmon, Daphnia, and Trout. Bulletin of Environmental Contamination and Toxicology 39, 15-22.

57
Shah, D. M., Horsch, R. B., Klee, H. J., Kishore, G. M., Winter, J. A., Tumer, N. E., Hironaka, C. M., Sanders, P. R., Gasser, C. S., Aykent, S., Siegel, N. R., Rogers, S. G., and Fraley, R. T. (1986). Engineering Herbicide Tolerance in Transgenic Plants. Science 233, 478-481.
Shehata, A., Schrodl, W., Neuhaus, J., and Kruger, M. (2013a). Antagonistic effect of different bacteria on Clostridium botulinum types A, B, D and E in vitro. Veterinary Record 172, 47.
Shehata, A. A., Kuehnert, M., Haufe, S., and Krüger, M. (2014). Neutralization of the antimicrobial effect of glyphosate by humic acid in vitro. Chemosphere 104, 258-261.
Shehata, A. A., Schrodl, W., Aldin, A. A., Hafez, H. M., and Krüger, M. (2013b). The Effect of Glyphosate on Potential Pathogens and Beneficial Members of Poultry Microbiota In Vitro. Current Microbiology 66, 350-358.
Shoval, S., and Yariv, S. (1981). Infrared study of the fine structures of glyphosate and Roundup. Agrochimica 25, 377-386.
Sidhu, R. S., Hammond, B. G., Fuchs, R. L., Mutz, J. N., Holden, L. R., George, B., and Olson, T. (2000). Glyphosate-tolerant corn: The composition and feeding value of grain from glyphosate-tolerant corn is equivalent to that of conventional corn (Zea mays L.). Journal of Agricultural and Food Chemistry 48, 2305-2312.
Simpson, E. R., Clyne, C., Rubin, G., Boon, W. C., Robertson, K., Britt, K., Speed, C., and Jones, M. (2002). Aromatase - A brief overview. Annual Review of Physiology 64, 93-127.
Simpson, E. R., Mahendroo, M. S., Means, G. D., Kilgore, M. W., Hinshelwood, M. M., Grahamlorence, S., Amarneh, B., Ito, Y. J., Fisher, C. R., Michael, M. D., Mendelson, C. R., and Bulun, S. E. (1994). Aromatase Cytochrome-P450, the Enzyme Responsible for Estrogen Biosynthesis. Endocrine Reviews 15, 342-355.
Sjaastad, V. Ø., Sand, O., and Hove, K. (2010). "Physology of Domestic Animals," 2nd/Ed. Scandinavian Veterinary Press, Oslo.
Smith, E. A., and Oehme, F. W. (1992). The biological activity of glyphosate to plants and animals: a literature review. Veterinary and Human Toxicology 34, 531-543.
Sost, D., and Amrhein, N. (1990). Substitution of Gly-96 to Ala in the 5-Enolpyruvylshikimate 3-Phosphate Synthase of Klebsiellapneumoniae Results in a Greatly Reduced Affinity for the Herbicide Glyphosate. Archives of Biochemistry and Biophysics 282, 433-436.
Sprankle, P., Meggitt, W. F., and Penner, D. (1975). Adsorption, Mobility, and Microbial Degradation of Glyphosate in the Soil. Weed Science 23, 229-234.
Stalker, D. M., Hiatt, W. R., and Comai, L. (1985). A Single Amino Acid Substitution in the Enzyme 5-enolpyruvylshikimate-3-phosphate Synthase Confers Resistance to the Herbicide Glyphosate. Journal of Biological Chemistry 260, 4724-4728.
Stallings, W. C., Abdel-Meguid, S. S., Lim, L. W., Shieh, H. S., Dayringer, H. E., Leimgruber, N. K., Stegeman, R. A., Anderson, K. S., Sikorski, J. A., Padgette, S. R., and Kishore, G. M. (1991). Structure and topological symmetry of the glyphosate target 5-enol-pyruvylshikimate-3-phosphate synthase: A distinctive protein fold. Proceedings of the National Academy of Sciences of the United States of America 88, 5046-5050.
Statistikbanken (2015). Import af oliekager og andre faste restprodukter fra udvinding af sojaolie, også formalede eller i form af piller pellets. Vol. 2015, http://www.statistikbanken.dk/statbank5a/default.asp?w=1680.
Steinrucken, H. C., and Amrhein, N. (1980). The Herbicide Glyphosate is a Potent Inhibitor of 5-enolpyruvylshikimic acid-3-phosphate Synthase. Biochemical and Biophysical Research Communications 94, 1207-1212.
Steinrucken, H. C., and Amrhein, N. (1984). 5-Enolpyrulvylshikimate-3-phosphatesynthase of Klebsiella pneumonia. 2. Inhibition by glyphosate (N-(phosphonomethyl)glycine) European Journal of Biochemistry 143, 351-357.
Stratton, G. W., and Stewart, K. E. (1992). Glyphosate effects on microbial biomass in a coniferous forest soil. Environmental Toxicology and Water Quality 7, 223-236.

58
Sviridov, A. V., Shushkova, T. V., Ermakova, I. T., Ivanova, E. V., Epiktetov, D. O., and Leontievsky, A. A. (2015). Microbial Degradation of Glyphosate Herbicides (Review). Applied Biochemistry and Microbiology 51, 188-195.
Tsui, M. T. K., and Chu, L. M. (2003). Aquatic toxicity of glyphosate-based formulations: comparison between different organisms and the effects of environmental factors. Chemosphere 52, 1189-1197.
USDA (2014). Genetically engineered varieties of corn, upland cotton, and soybeans, by State and for the United States, 2000-14. In "Adoption of Genetically Engineered Crops in the U.S.", http://www.ers.usda.gov/data-products/adoption-of-genetically-engineered-crops-in-the-us.aspx.
Waltz, E. (2010). Glyphosate resistance threatens Roundup hegemony. Nature Biotechnology 28, 537-538. Williams, A. L., Watson, R. E., and DeSesso, J. M. (2012). Developmentan and Reproductive Outcomes in
Humans and Animals After Glyphosate Exposure: A Critical Analysis. Journal of Toxicology and Environmental Health-Part B-Critical Reviews 15, 39-96.
Williams, G. M., Kroes, R., and Munro, I. C. (2000). Safety evaluation and risk assessment of the herbicide Roundup and its active ingredient, glyphosate, for humans. Regulatory Toxicology and Pharmacology 31, 117-165.
Yen, J. T. (2001). Anatomy of the Digestive System and Nutritional Physiplogy. In "Swine Nutrition" (A. J. Lewis and L. L. Southern, eds.).
Yun, J. H., Lee, K. B., Sung, Y. K., Kim, E. B., Lee, H. G., and Choi, Y. J. (2009). Isolation and characterization of potential probiotic lactobacilli from pig feces. Journal of Basic Microbiology 49, 220-226.

Appendix 1
Determination of viable counts by use of Drop Plate Procedure Add 250 µl sample to the first well in the first column (i), in a In 96-well microtiterplates
Add 225 µl medium in the wells, where dilutions are wanted
Transfer 25 µl sample from column i to 225 µl medium (0.1 M sodium phosphate buffer, pH 6.5) in
column (i+1)
Mixed sample and buffer 10 times
Repeat process until wanted dilutions are obtained
Plate 10 µl, from each of the dilutions of interest, for each sample, on MRS agar
Incubate MRS plates anaerobically overnight, at 37°C.
Count plates the next day
Calculate colony forming units (CFU)
Quantification of VFA concentrations Add 1 ml sample (1 ml standard mix and 1 ml blind (H2O) to SCFA tubes
Add 100 µl intern standard to each sample
Add 0,5 ml HCl to each sample
Add 2 ml ether to each sample
Mix samples for 30 sec on Vibrax
Centrifuge samples at 2000 g for 10 min (5°C)
Add 10 µl MTBSTFA to microvials
Add 50 µl of the supernatant (from the samples) to microvials
Close microvials
Mix microvials on vortex’er
Place microvials on heating block (80°C) for 20 min
Store microvials in room temperature for 48 hours
Run samples on the Gas Chromatograph (Thomas Rebsdorf)
Gas measurements Extract 1 ml gas from each sample of interest
Eject 0.5 ml of the gas into a Gas Chromatography
Analyze data using the appurtenant PEAK359 software.

Appendix 2
Influence of glyphosate acid, glyphosate IPA salt, Roundup and POEA on growth of bacterial
pure cultures
Preparation of MRS
Mix contents
Autoclave at 110°C for 20 min (sometimes 100° in 15 min, just to melt it, as it easily burns off)
Stir and cool media, while infusing CO2
Distribute media into VFA tubes while infusing CO2
Autoclave tubes at 121 °C for 15 min (118° for 15 min)
Preparation of BHI
Mix contents
Autoclave at 121°C for 15 min
Stir and cool media
If anaerobic, infuse CO2 while stirring
If microaerophilic, infuse a gas, with a low oxygen level
Distribute into VFA tubes
Autoclave tubes at 121 °C for 15 min
Preparation of RM02/Ruminococcus media
Kim (2012), 1Widdel et al. (1983), 2Tschech and Pfennig (1984)1
1Kim, C. C. (2012). "Identification of rumen methanogens, characterization of substrate requirements and measreument of hydrogen thresholds: a
thesis presented in partil fulfilment of the requirements for the degree of Master's in Microbiology." Massey University, New Zealand. Tschech, A., and Pfennig, N. (1984). Growth yield increase linked to caffeate reduction in Acetobacterium woodii. Archives of Microbiology 137, 163-
167. Widdel, F., Kohring, G. W., and Mayer, F. (1983). Studies on dissimilatory sulfate-reducing bacteria that decompose fatty acids. Archives of
Microbiology 134, 286-294.
Broth media 52.4 g
Agar 0.5 g (0.05 % of 1 L)
Demineralized water 1 L
Broth media 37.1 g
Agar 0.5 g (0.05 % of 1 L)
Demineralized water 1 L
KH2PO4 1.4 g
(NH4)2SO4 0.6 g
KCl 1.5 g
Sodium acetate 1.6 g
Trace element solution (SL10)1 1 ml
Selenite/tungstate solution2 2 ml
Resazurin 5-7 drops
Distil led water 950 ml

Mix contents
Boil in the microwave - go on high for 4 minutes. Look for bubbles - once bubbles appear, keep
going for around a minute. Increase time if needed.
Place media on ice, while infusing CO2 until it reaches room temp or just a little warmer
After cooling, add 4 g NaHCO3 and 0.50 g L-cysteine·HCl·H2O
Adjust pH to 6.8 with 5.6 M NaOH
Disperse 7.6 ml media into each VFA tube while infusing CO2
Autoclave tubes at 121°C for 15 min
Store in the dark for 24 hours
Before use, add 0.4 ml CRF, yeast extract and vitamin mix, 0.4 ml glucose, cellobiose, xylose and
arabinose mix (40 mM each) and 0.2 ml reducing agent
Preparation of colon
Mix contents
Autoclave at 121°C for 10 min
Stir and infuse CO2
Add 0.200 ml Vitamin K1 and 1.0 g L-cysteinehydrochloride while stirring
Distribute media in VFA tubes while infusing CO2
Autoclave tubes at 121 °C for 15 min
Preparation of anaerobic basal broth
Mix contents
Autoclave at 121°C for 15 min
Stir and cool media while infusing CO2
Distribute media into VFA tubes under CO2 infusion
Autoclave tubes at 121 °C for 15 min
Solution A 167 ml
Solution B 167 ml
Colon-juice? 200 ml
Sugar solution 40 ml
Yeast extract (Merck 1.03753) 5.0 g
Trypticase peptone (Merck 1.07213) 5.0 g
NaHCO3 5.0 g
Resazurin 1 ml
Hemin 10 ml
Elga water Add up to 1 L
Broth media 35.4 g
Agar 0.5 g (0.05 % of 1 L)
Demineralized water 1 L

Preparation of microtiterplates
Bacteria cultures are inoculated to broth media and incubated overnight, at either 37°C or 41°C
Overnight culture is investigated in microscope, to check for purity
Overnight culture is diluted 100 times in broth media, if growth is found sufficient (density of approx.
109 cells per ml)
Overnight culture is diluted 10 times in broth media, if growth is less dense than approx. 109 cells per
ml
325 µl media + 1,4 µl POEA is added to each well; 1, 2 and 3, in row A, to make a replicate of three
Stock solution of glyphosate acid is made (10 mg/ml a.e.). 325 µl is added to each well; 4, 5 and 6 in
row A, to make a replicate of three
Stock solution of glyphosate IPA salt is made (10 mg/ml a.e.). 325 µl is added to each well; 7, 8 and 9
in row A, to make a replicate of three
Stock solution of Roundup is made (10 mg/ml a.e.). 325 µl is added to each well; 10,11 and 12 in row
A, to make a replicate of three
225 µl media is added to each well in row B-H
For each well in row A, solution is mixed 10 times with a pipette and 100 µl is transferred to the next
well (row B) and mixed 10 times. The procedure is repeated until row G, where the extra 100 µl is
thrown away. A 3.5 dilution is now made from each well to the next.
Row H corresponds to the control
In each well, 25µl bacterial dilution is added. Mix culture on vortex’er before use.
Bacteria and media is mixed, operating from high to low dilution
50 µl sterilized paraffin oil is added to each well
The microtiterplate is incubated in an ELISA spectrophotometer at 37°C for 18-24 h

Glyphosate acid: Add 80 mg glyphosate acid to 8 ml media to end with a final concentration of 10 mg/ml a.e.
Glyphosate IPA salt: Add 270 µl glyphosate IPA salt to 8 ml media to end with a final concentration of 10
mg/ml a.e.
Calculations:
Glyphosate IPA solutions: 40 wt % in H20, corresponds to 400 mg/ml.
0.74 ∗ IPA salt = glyphosate acid
glyphosate IPA salt = glyphosate acid
0.74=
10 mg/ml
0.74= 13,51 mg/ml
13.51 mg/ml
400 mg/ml= 0,0338 ml IPA pr ml media
0.0338 ml ∗ 8 ml = 0.270 ml = 270 µl
Roundup: Add 222 µl Roundup to 8 ml media to end with a final concentration of 10 mg/ml a.e.
Calculations:
Roundup contains 360 g/L glyphosate acid (a.e.):
10 mg/ml
360 mg/ml= 0.0278 ml Roundup pr ml media
0.0278 ml ∗ 8 ml = 0.222 ml = 222 µl
POEA: Add 1.4 µl POEA to 325 µl media to end with a final concentration of 4.17 mg/ml POEA.
Calculations:
We expect Roundup to contain 360 g/L glyphosate acid and 150 g/L POEA
150 mg/mL
360 mg/ml= 0.417
Based on the assumption that 1 mg POEA is the same as 1 ml, I have to add 1 µl POEA for every 250 µl.
Every time we have 10 mg/ml glyphosate acid, we have 4.17 mg/ml POEA.

Appendix 3
Influence of glyphosate IPA salt, Roundup and POEA on gut microbiota from pigs and cows
Preparation of 0.1 M sodium-phosphate buffer (pH 6.5)
Add 13,8 g NaH2PO4 * H2O to 1 L ELGA water
Stir the suspension on the magnetic stirrer, while measuring pH. Add 5.6 M NaOH to reach a final pH
of 6.5
Transfer 35 ml solution to 125 ml bottles and seal them with butyl rubber stoppers
Flush bottles with N2 by three successive cycles, where bottles are evacuated and refilled, using a
manifold, fitted to a vacuum pump and a gas tank.
Adjust pressure to atmospheric with a needle
Autoclave bottles at 121 °C for 15 min
Digesta samples from slaughter pigs and dairy cows:
Prepare glyphosate IPA salt, Roundup and POEA solutions according to Table 1. Mix on vortex’er
Pigs: Collect stomach, cecum and colon content at slaughter
Cows: Collect rumen content from fistulated dairy cows. Sieve content through a cheesecloth to
remove big particles
Place samples on ice, and take them to the laboratory immediately.
Add 10 ml content to each bottle, containing 35 ml 0.1 M sterile anaerobic sodium-phosphate
buffer (pH=6.5), with a cut-off syringe. A final suspension of 20 % wt/vol is reached.
Add 5 ml of glyphosate IPA salt/Roundup/POEA solution to each bottle, with a syringe, to reach
desired concentrations (Table 2, Table 3).
Flush bottles with CO2 by three successive cycles, where bottles are evacuated and refilled, using a
manifold, fitted to a vacuum pump and a gas tank.
Adjust pressure to atmospheric with a needle
Pigs: from each bottle, extract 1 ml for VFA measurements (store at -18°C until analysis), 1 ml for
DNA analysis (snap freeze in liquid nitrogen and store at -80°C), 1 ml for immediately pH
measurement and 0.5 ml for Drop Plating Procedure, at 0, 3/4 and 24 hours, respectively. Extraction
is done with a syringe to avoid contamination with air.
Cows: from each bottle, extract 1 ml for VFA measurements (store at -18°C until analysis), 1 ml for
DNA analysis (snap freeze in liquid nitrogen and store at -80°C) and 1 ml for immediately pH
measurement, at 0, 2, 4 and 24 hours, respectively. 1 ml gas samples, from each bottle are extracted
at 0, 2, 4, 6 and 24 hours, respectively. Extraction is done with a syringe to avoid contamination with
air.
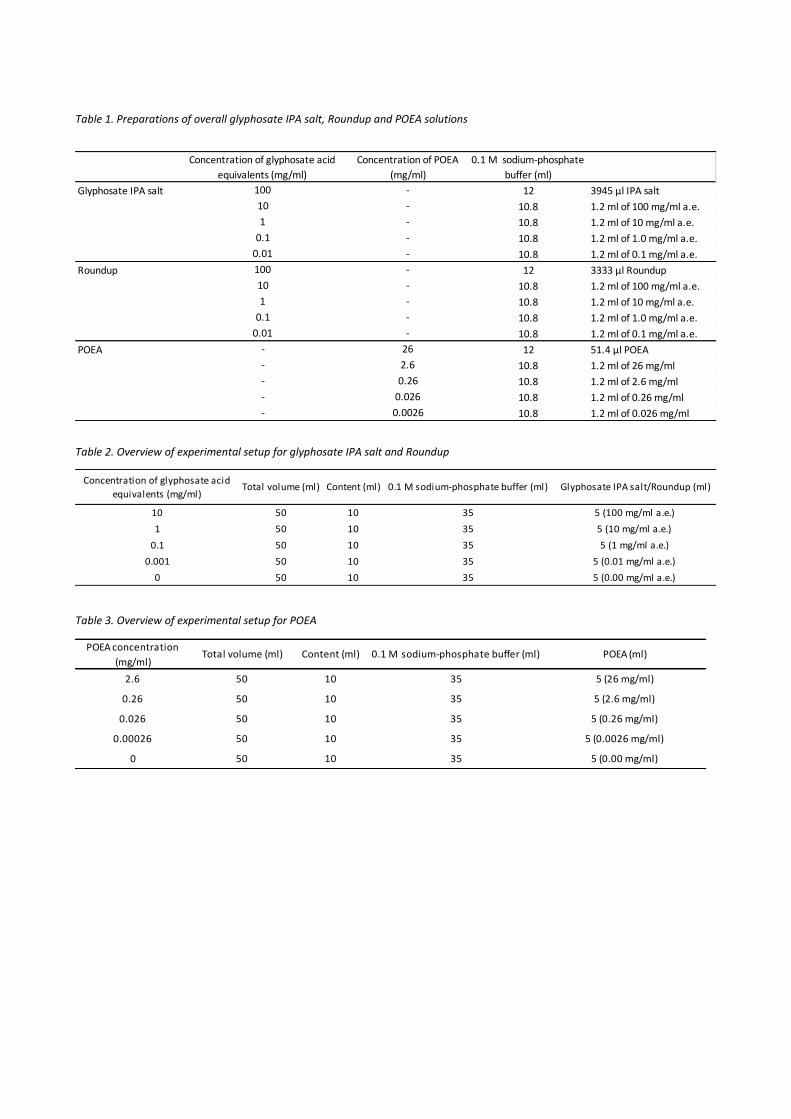
POEA concentration
(mg/ml)Total volume (ml) Content (ml) 0.1 M sodium-phosphate buffer (ml) POEA (ml)
2.6 50 10 35 5 (26 mg/ml)
0.26 50 10 35 5 (2.6 mg/ml)
0.026 50 10 35 5 (0.26 mg/ml)
0.00026 50 10 35 5 (0.0026 mg/ml)
0 50 10 35 5 (0.00 mg/ml)
Concentration of glyphosate acid
equivalents (mg/ml)
Concentration of POEA
(mg/ml)
0.1 M sodium-phosphate
buffer (ml)
Glyphosate IPA salt 100 - 12 3945 µl IPA salt
10 - 10.8 1.2 ml of 100 mg/ml a.e.
1 - 10.8 1.2 ml of 10 mg/ml a.e.
0.1 - 10.8 1.2 ml of 1.0 mg/ml a.e.
0.01 - 10.8 1.2 ml of 0.1 mg/ml a.e.
Roundup 100 - 12 3333 µl Roundup
10 - 10.8 1.2 ml of 100 mg/ml a.e.
1 - 10.8 1.2 ml of 10 mg/ml a.e.
0.1 - 10.8 1.2 ml of 1.0 mg/ml a.e.
0.01 - 10.8 1.2 ml of 0.1 mg/ml a.e.
POEA - 26 12 51.4 µl POEA
- 2.6 10.8 1.2 ml of 26 mg/ml
- 0.26 10.8 1.2 ml of 2.6 mg/ml
- 0.026 10.8 1.2 ml of 0.26 mg/ml
- 0.0026 10.8 1.2 ml of 0.026 mg/ml
10 50 10 35 5 (100 mg/ml a.e.)
1 50 10 35 5 (10 mg/ml a.e.)
0.1 50 10 35 5 (1 mg/ml a.e.)
0.001 50 10 35 5 (0.01 mg/ml a.e.)
0 50 10 35 5 (0.00 mg/ml a.e.)
Concentration of glyphosate acid
equivalents (mg/ml)Total volume (ml) Content (ml) 0.1 M sodium-phosphate buffer (ml) Glyphosate IPA salt/Roundup (ml)
Table 1. Preparations of overall glyphosate IPA salt, Roundup and POEA solutions
Table 2. Overview of experimental setup for glyphosate IPA salt and Roundup
Table 3. Overview of experimental setup for POEA

Appendix 4
Influence of glyphosate IPA salt on growth of L. sobrius in stomach content from pigs
Preparation of anaerobic salt media
Mix contents
Autoclave at 121°C for 15 minutes.
Preparation of broth
Add yeast extract and peptone to 200 ml ELGA water
Add solution A, B, resazurin, hemin and tween 80 and the rest of the water
Autoclave at 110°C for 10 min.
Cool to room temperature while flushing with CO2.
Add 85 ml to 125 ml infusion serum bottles, while flushing with CO2
Flush sealed bottles with CO2
Autoclave the bottles at 121°C for 15 min.
The day of slaughter
Collect stomach content from pigs at slaughter
Pool stomach content
Store stomach content at -18°C until use
The day before start of experiment
Prepare an overnight culture of L. sobrius
Remove stomach content from freezer
On the day of the experiment
Add 5 ml R to the bottles with 85 ml broth
Solution A 167 ml
Solution B 167 ml
Hemin 5 ml
Elga water Add up to 1 L in total
Yeast extract 2.5 g
Peptone from casein (Merck 1.07213) 2.5 g
Solution A 167 ml
Solution B 167 ml
Resazurin 1 ml
Hemin 5 ml
Tween 80 1 ml
Elga water Add up to 1 L in total

Dilute 500 ml content in 500 ml sterile anaerobic salt media (1:1) and stomach the suspension. To
ease handling, each suspension is prepared in three successive steps, due to limitations of the
stomacher
Add 1000 ml of suspension to each bioreactor
Wait until the temperature has reached 37°C.
Add glyphosate IPA salt, to reach the desired concentrations (Tabel 4), in each bioreactor
Add 10 ml of L. sobrius overnight culture to each bioreactor
Suspension is stirred and kept under anoxic conditions by flushing with N2 gas in the headspace.
pH is maintained, using a pH-controller, regulated by 1M NaOH and 5M HCl.
Incubation temperature is kept at 37°C, by a circulating water bath.
Remove 12 ml samples from each bioreactors at 0, 2, 4 and 24 hours
Of the 12 ml, transfer 1 ml to SCFA tubes, for VFA determination and store at -18°C
Of the 12 ml, transfer 10 ml to sterile 125 ml serum bottles, with rubber stoppers, containing 85 ml
broth and 5 ml R. Pour each suspension from the bottles into a plastic bag and homogenize in a
stomacher for 2 minutes.
Perform the Drop Plate Procedure (Appendix 1)
Tabel 4. Preparation of glyphosate IPA salt concentrations
Bioreactor Glyphosate acid concentration (mg/ml) Glyphosate IPA salt (ml)
1 0 0
2 0.1 0.3375
3 1 3.375
4 10 33.75

Appendix 5
Statistical models
Required packages library(lme4)
library(lsmeans)
library(pbkrtest)
Example – Plates (B.adolescentis) adol=read.csv("C:/Users/Charlotte/Desktop/Speciale/Results/B. adolescentis .csv", header=T)
usedat1 <- reshape(adol,varying=names(adol)[3:ncol(adol)],
v.names = "resp",timevar="treat",
times=names(adol)[3:ncol(adol)],
direction = "long")
usedat1 <- usedat1[!is.na(usedat1$resp),]
adol.lmer <- lmer(resp ~ conc * treat + (1|plate),data=usedat1)
tmp <- lsmeans(adol.lmer,list(pairwise~conc:treat))
tmp[1]
Example - Cecum (representative for pigs)
pH ph = read.csv("C:/Users/Charlotte/Desktop/Speciale/Results/Statistics/Cecum/pH_all.csv",header=T)
ph$individ <- as.factor(ph$individ)
ph$timegrp <- as.factor(ph$time)
treatments <- unique(ph$treatment)
concentrations <- unique(ph$conc)
ph.lmer <- lmer(pH~1+treatment*conc+timegrp+(1|individ),data=ph)
ph.lsm <- lsmeans(ph.lmer,~conc|treatment)
ph.lsm

VFA all vfa = read.csv("C:/Users/Charlotte/Desktop/Speciale/Results/Statistics/Cecum/VFA_all.csv",header=T)
vfa$individ <- as.factor(vfa$individ)
vfa$timegrp <- as.factor(vfa$time)
treatments <- unique(vfa$treatment)
concentrations <- unique(vfa$conc)
all.lmer <- lmer(all~1+treatment*conc+timegrp+(1|individ),data=vfa)
all.lsm <- lsmeans(all.lmer,~conc|treatment)
all.lsm
Example - Rumen
pH ph = read.csv("C:/Users/Charlotte/Desktop/Speciale/Results/Rumen/pH_all.csv",header=T)
ph$day <- as.factor(ph$day)
ph$cow <- as.factor(ph$cow)
ph$timegrp <- as.factor(ph$time)
treatments <- unique(ph$treatment)
concentrations <- unique(ph$conc)
ph.lmer <- lmer(pH~1+treatment*conc+timegrp+(1|day)+(1|cow),data=ph)
ph.lsm <- lsmeans(ph.lmer,~conc|treatment)
ph.lsm
VFA vfa = read.csv("C:/Users/Charlotte/Desktop/Speciale/Results/Cow
trials/Statistics/Rumen/VFA_all.csv",header=T)
vfa$day <- as.factor(vfa$day)
vfa$cow <- as.factor(vfa$cow)
vfa$timegrp <- as.factor(vfa$time)
all.lmer <- lmer(all~1+treatment*conc+timegrp+(1|day)+(1|cow),data=vfa)
all.lsm <- lsmeans(all.lmer,~conc|treatment)
all.lsm

Appendix 6 Growth of commensal bacteria

Appendix 7 Growth of potential pathogenic bacteria

Appendix 8 Stomach
Roundup
RoundupRoundup
Roundup

Appendix 9 Cecum
RoundupRoundup
RoundupRoundup
RoundupRoundup

Appendix 10 Colon
Roundup Roundup
RoundupRoundup
RoundupRoundup

Appendix 11 Rumen
RoundupRoundup
RoundupRoundup
RoundupRoundup
Appendix 12 Bioreactors