effects of minerals on feed degradation and protein
TRANSCRIPT

HAL Id: hal-00900299https://hal.archives-ouvertes.fr/hal-00900299
Submitted on 1 Jan 1999
HAL is a multi-disciplinary open accessarchive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come fromteaching and research institutions in France orabroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, estdestinée au dépôt et à la diffusion de documentsscientifiques de niveau recherche, publiés ou non,émanant des établissements d’enseignement et derecherche français ou étrangers, des laboratoirespublics ou privés.
Effects of minerals on feed degradation and proteinsynthesis by rumen micro-organisms in a dual effluent
fermenterLaurent Broudiscou, Yves Papon, Anne F. Broudiscou
To cite this version:Laurent Broudiscou, Yves Papon, Anne F. Broudiscou. Effects of minerals on feed degradation andprotein synthesis by rumen micro-organisms in a dual effluent fermenter. Reproduction NutritionDevelopment, EDP Sciences, 1999, 39 (2), pp.255-268. �hal-00900299�

Original article
Effects of minerals on feed degradation and protein synthesisby rumen micro-organisms in a dual effluent fermenter
Laurent-Philippe Broudiscou Yves Papon Anne F. Broudiscoub
a Département de l’élevage et nutrition des animaux, Inra, Paris, Franceb LPRAI, 31190 Miremont, France
(Received 10 July 1998; accepted 27 January 1999)
Abstract - In dual outflow continuous fermenters on a 75:25 hay/barley diet, feed degradationand protein synthesis by mixed rumen microbes were tested in relation to the concentrations ofHPO4z-, HC03 and Cl- and Na+/K+ ratio in artificial saliva, by applying a 16-run Franquart design,and by fitting second-order polynomial models. The HP04 2-, HC03-1 Cl- concentrations and Na+/K+ratio ranged from 0.1 to 4 g-L-1, from 0.5 to 7 g-L-1, from 0.1 to 0.5 g.L! ’ and from 0.5 to 15 g-g-1, ’,respectively. Buffer salts, particularly HP04 2-, were the major factors while Cl- concentration had neg-ligible effects on microbial metabolism. Maximal neutral detergent fibre, acid detergent fibre andorganic matter degradabilities occurred at intermediate values of HP04 2- and HC03- concentrations.The outflow of microbial protein and the efficiency of microbial protein synthesis, which variedfrom 26.2 to 37.1 g.N.kg-1 of organic matter truly degraded, reached minima at the centre of theexperimental domain. @ Inra/Elsevier, Paris.
mineral / rumen / micro-organism / proteosynthesis
Résumé ― Effet des minéraux sur la dégradation des aliments et la synthèse de protéines parles microbes du rumen en fermenteur à double effluent. Les effets des concentrations de HP04 2-HC03 , Cl- et du rapport Na’/K+, dans la salive artificielle, sur la dégradation des aliments et la syn-thèse de protéines par les microbes du rumen ont été quantifiés dans des fermenteurs à effluentdouble recevant une ration 75 % foin/25 % orge, en appliquant une matrice de Franquart et à l’aidede modèles polynomiaux du second degré. Les concentrations de HP04z-, HC03-, Cl- et le rapportNa+/K+ ont respectivement varié de 0,1 à 4 g.L -1, de 0,5 à 7 g!L-!, de 0,1 à 0,5 g-L-1 et de 0,5 à 15 g-g-1.Les facteurs les plus influents ont été les minéraux tampons, particulièrement HPO i-. Les dégrada-bilités du neutral detergent fibre, de l’acid detergent fibre et de la matière organique ont été maximalespour des valeurs centrales des concentrations de HPO 4 2- et HC03 Le flux de protéines microbiennes
* Correspondence and reprintsLaboratoire de nutrition et alimentation, LN.A.P.G., 16, rue Claude-Bernard, 75231 Paris cedex O5,FranceE-mail: [email protected]

et l’efficacité de leur synthèse, de 26,2 à 37,1 g N-kg-’ de matière organique réellement dégradée, ontété minimaux au centre du domaine expérimental. © Inra/Elsevier, Paris.
minéral / rumen / micro-organisme / synthèse de protéines
1. INTRODUCTION
The role of minerals in regulating themetabolism of rumen micro-organisms isrecognised [20, 39] and their effects havebeen investigated in diverse experimentalsituations. In husbandry conditions, the sup-plementation of ruminants with buffer salts,to counteract the detrimental effects of high-concentrate diets [41], or the changesinduced by sodium and potassium on rumenfunctions [35] have been evaluated. In orderto formulate adequate culture media [11,13], microbiologists have studied the effectof specific ions on bacterial growth in pureculture. Continuous culture systems offeropportunities to gain information on thedirect response of mixed rumen microbesto a change in mineral supply. In animals, asimilar treatment would affect the host
digestive physiology (e.g. the rumen con-tent’s dilution rate and absorption rates ofmetabolites) and would also have indirectconsequences on rumen microbes. More-over, dual effluent continuous fermenters,well suited to the design of the experiments,allow data collection on a large set of factorswithin a single trial at reasonable expense.
In a previous paper [9], we described theinfluence of salivary salts on fermentationvariates, in a continuous culture of mixedrumen microbes maintained on a standarddiet. In the present report, we are consider-ing the relationships between the same min-eral compounds and the utilisation of feedsby microbes to produce biomass, in partic-ular proteins. Some preliminary results arein the process of publication in abstract form[ 10].
2. MATERIALS AND METHODS
2.1. Experimental strategy
The major inorganic components of naturaland artificial salivas are potassium or sodiumsalts of bicarbonate, of hydrogen and dihydro-gen phosphate, and of chloride. In vitro, the pHmay be adjusted by addition of KOH or NaOH.The main characteristics of any saliva can, thus,be defined by four variables: 1) the amount ofphosphates, expressed as hydrogen phosphateequivalent (HPO42-); 2) the amount of carbon-ates expressed as bicarbonate equivalent(HC03 ); 3) the amount of chloride (CI-); and4) the ratio of sodium to potassium amounts(Na+/K+). These variables were used as experi-mental factors in our study, and we aimed atdetermining their quantitative effect on rumenmicrobe metabolism. The quantities of HPO42-,HCO 3- and Cl- in our trial (table n reproducedphysiological variations around the average val-ues reported by Clarke [14] and Durand andKawashima [20]. A distinctive feature of theNa+/K+ factor was the location of the values of
greatest interest at the periphery of the experi-mental domain rather than at its centre, since theNa+/K+ ratios in most salivas were close to either14 or 2 [1, 4, 24, 27, 38, 45, 51]. We thus dimen-sioned the variation interval of this factor toinclude both sets of values.
Published data have clearly demonstrated lin-ear or quadratic responses of bacterial growth[2, 12, 16, 29, 30, 43, 47] or enzymatic activity [2,26] to physiological changes in mineral supply,in accordance with the law of ecological toleranceformulated by V.E. Shelford in 1913. Therefore,we modelled the relationships between experi-mental factors and microbial metabolism param-eters using second-order polynomial equations.Response surface methodology, extensivelyreviewed in the statistics literature [5, 15], pro-vided an efficient experimental strategy to deter-mine the relationships between the four inde-pendent variables and the responses relative tomicrobial metabolism. We explored the spherical

domain comprised within the values given intable I, using an experimental design publishedby Franquart [23]. This design was found usingsimulated annealing with D-optimality for cri-terion. Rather than selecting a central compositedesign for this trial, we employed a Franquartdesign, which also led to a valid estimate of theresponse surface model, though with a muchsmaller number of experiments. We have com-pared the main characteristics of both designs intable II, as summarised by Peissik [44], sincethe Franquart design has not yet been publishedin the agronomic or statistical literature. Maximalinflation factors are equally satisfactory (below4) and the Franquart design is close to orthogo-nality. Using this design leads to an adequateprediction of dependent variables (or responses),with a maximal variance function dMax of 1.
The Franquart design is almost rotatable, asshown by the Khuri index of above 98 % [36].Our experimental worksheet is shown in tableIII. All runs, except #16, are evenly distributed atthe edge of the four-dimensional experimentaldomain. Run #16, at the centre, was applied threetimes to give an estimation of experimental error.The 18 runs were randomly assigned to six inde-pendent fermenters, identically assembled, whichwere operated for three 7-day contiguous exper-imental periods. The randomisation was justi-fied by a preliminary trial, where the fermenterswere run for two 1 1-day experimental periodswith no statistically significant differencesbetween periods or between fermenters for mostmeasurements (Broudiscou, unpublished data).
2.2. Incubation procedure
The rumen contents used as inoculum werewithdrawn after a 24-h fasting period from twowethers, fed twice daily 1 000 g’d*’ chopped hayand 200 g-d-I ground and pelleted barley. Theinoculation protocol was described in Broudiscouet al. [8]. Each fermenter, maintained at 39 °C,was continuously infused with one of the 16 testedsalivas. All the salivas were supplemented with0.4 g-L-1 HCl-cysteine as reducing agent. Thefermentation broths were separately supple-mented with 31.7 mg-d-I CaCl2, 47.5 mg-d-IMgCl2 and 0.755 g!dw (NH4)ZS04. Ten gramsof a pelleted diet made of 750 g-kg-I orchard-
grass hay (dry matter content of 92.] %) and250 g’kg’’ ground barley (dry matter content of
91.1 %) was supplied to the fermenters at l Oh00and 20 g at 18h00. The composition of feeds isgiven in table IV. The dilution rates of particleand liquid phases were set at 0.03 h-I and0.06 h-’, respectively. The procedures followedto control the two turnover rates, and for the dailycollection of deplaced and filtered effluents, wereidentical to Broudiscou et al. [8].
2.3. Analytical methods
After a 5-day adaptation term, the deplacedand filtered effluents collected on days 6 and7 were pooled and kept at -20 °C until sub-sam-pled for dry matter (DM), volatile fatty acids(VFA) and ammonia nitrogen (NH3-N) deter-mination. The samples for VFA and NH3 N anal-ysis were mixed with 0.1 volume of H3P04 8.2 %
(w/w) and stored at -20 °C until analysis. Theremaining effluents were freeze-dried. At theend of the incubation period, the fermentationbroth was strained and the feed residue in the
liquid phase was separated by centrifugation at1 500 g for 10 min. The supernatant was cen-trifuged at 27 000 g for 30 min to isolate the liq-uid-associated bacterial (LAB) fraction. The bac-terial pellet was resuspended into 90 g-L-1 NaCI,
centrifuged at 27 000 g for 30 min and freeze-dried. Feeds and freeze-dried effluents were
ground prior to analysis using a Culatti grinderwith a 0.8-mm screen. Feed, effluent and bacte-rial samples were analysed for DM, organic mat-ter (OM) [7], neutral detergent fibre (NDF), andacid detergent fibre (ADF) [50]. Total nitrogenwas measured by the micro-Kjeldahl technique.NH3-N was determined as described by Daviesand Taylor [18]. VFA were determined asdescribed by Jouany [31 ]. Effluent and bacterialsamples were analysed for nucleobases accord-ing to Lassalas et al. [32] using a diode-arraydetector (Beckman Instruments, Fullerton). Phos-phorus in feeds was determined by AFNOR stan-dard method NF V 18-106.

2.4. Calculations
Spectral analysis of HPLC separation showedan impurity occurring within the peak of cyto-sine for most bacterial and effluent samples. Wethus discarded this pyrimidine base from the poolof nucleobases used as a microbial marker. Theoutflows of microbial OM and of microbial N(MN) were determined from nucleobases by twocalculation methods. In the first one, the micro-bial reference pool was the commonly used LAB.In the second one, the microbial reference wasalso calculated from the LAB chemical compo-sition but a fraction of nucleobase outflow wasattributed to solid-associated bacteria (SAB) byapplying the following hypotheses: the LAB rep-resents 20 % of the bacterial DM outflow, theash and the nucleobase contents of DM are both55 % lower in SAB than in LAB and the SABnitrogen contents is 1 g N/100 g DM lower thanin LAB. The outflows of microbial OM and Nresulting from the correction due to SAB biomasswere used to calculate the true OM and Ndegradabilities, and the efficiency of microbialprotein synthesis (EMPS). The true OM degrad-ability (tdoM) was determined by equation (1): ):
The daily amount of OM fermented (OMF)was estimated from outflows of individual VFA
by equation (2), derived from the relation givenby Demeyer and Van Nevel [19] to calculate theamounts of fermented hexose:
where C2, C3, IC4, C4, IC5 and C5 were thedaily outflows (mol-d-1) of acetate, propionate,isobutyrate, butyrate, isovalerate and valerate,respectively.
The amount of nucleic acids in LAB was esti-mated from nucleobase analysis by consideringthat the molar fractions of cytosine, adenine andguanine included into DNA equalled the ratio[thymine]/[thymine + uracil].
2.5. Statistical analyses
The results were submitted to stepwise regres-sion by an SAS procedure [46]. The followingsecond-order polynomial model was fitted todata:
where )!. were the four coded variables presentedin table 1, and bo, b; bij and b,, the 15 coefficientsto be estimated. The coded variables X!, used inmultiple linear regression, varied from -1 to +1.They were related to the experimental factors l!.,also called natural variables, by the equation:
in which N;° is the value at the centre of thedomain, and AN, is the interval of variation ofN;. For instance, ’No and AN equalled 2.05 and3.9 g-L-1, respectively, for variable HPO 42- . TheF statistic significance level for deleting or add-ing a variable was set to 0.15. We assessed thegoodness or the lack of fit of the models by visualanalysis of the response residuals. The resultswere also submitted to response surface polyno-mial regression, with no selection among modelterms [46).
3. RESULTS
All the regression analyses were per-formed on 17 runs, as one repetition ofexperiment #16 was stopped owing to tech-nical failure. The numerical results of step-wise regressions are given in the tables. Inthe model-fitting procedure, we did notapply the marginality principle - the termfor X;2 or X X could remain even if the termfor Xi or X had been excluded - becausenone of our variables was discrete [3]. Theaddition or the deletion of a term in themodel was only determined by the level ofsignificance of the F statistic. The coeffi-cients for excluded terms were set at zeroin the tables. The absence of a term in atable meant that it was deleted in all the
reported models. The degrees of freedomfor the error term, being easily determinedfrom the number of terms finally included inthe model for each response variable, werenot inserted in the tables. The outputs ofresponse surface analysis are presented inthe figures, owing to the large number ofcoefficients in the corresponding models.For all response variables, the model coef-ficients estimated by stepwise regression

and by full-model regression were very closeand led to similar conclusions.
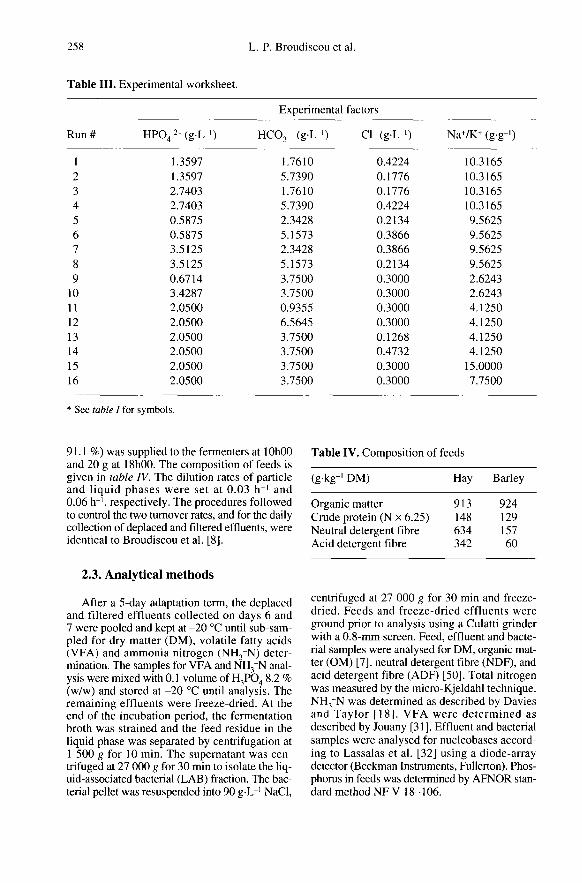
The model coefficients for apparent andtrue degradabilities of feed constituents arepresented in table V. NDF degradability(dNDF) ranged from 32.9 to 71.3 %, and aver-aged 49.3 %. The model provided a satis-factory fit to data, characterised mainly byquadratic effects of all four factors, the majorone being Hpo,2-. The maximal degrada-tion was thus attained in the central part ofthe experimental domain. Figure I showsthe contour plot of dNDF for HP04 2- andHC03 concentrations, at Cl- concentrationand Na+/K+ values of 0.3 g-L’’ and 7.75 g.,,-’,which are the co-ordinates of the central
point in our experimental domain. Whenusing the figures, one must remember thatthe precision of the prediction is substan-tially degraded outside of the spherical
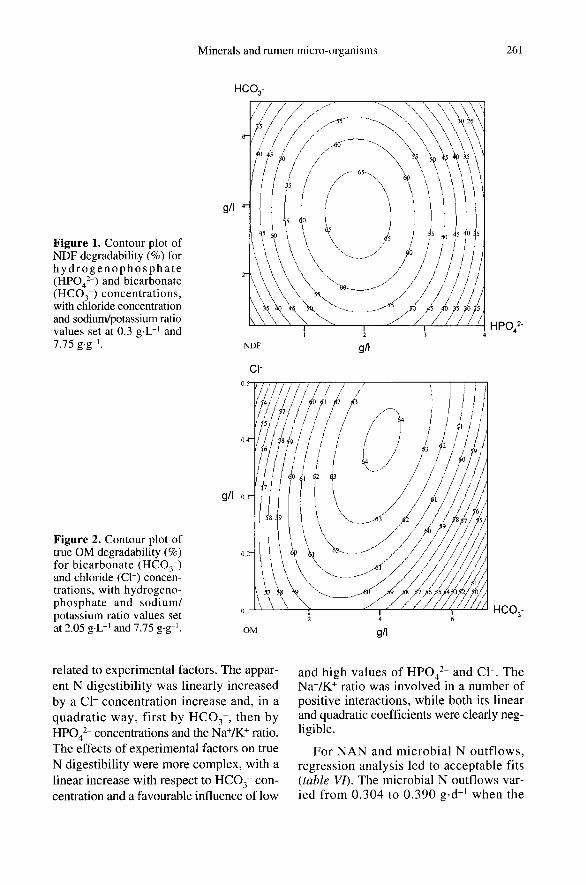
domain we have explored. The ADF degrad-ability (dall) was poorly explained by themodel, and was only influenced in aquadratic way by HP04z- and Na+/K+. Theapparent OM degradability (adoM) and thetrue OM degradability (tdoM) varied from19.1 to 37.7 % and from 52.6 to 64.0 %
(with an average value of 60.1 %), respec-tively. The degradation of OM demonstrateda conjugated action of HC03 -, in a quadraticway, and of Cl-. Figure 2 presents the con-tour plot of tdoM for HCO ! and Cl- con-centrations, at constant HPO4z- and Na+/K+values of 2.05 g-L-1 and 7.75 g-g-1. !.
The amount of fermented OM, with anadjusted R2 of 0.36, was minimal with highor low values of HCO!- and Hpo42- . Theapparent and true N degradabilities, rang-ing from 3.1 to 13.0 % and from 74.2 to85.9 %, respectively, were more closely
related to experimental factors. The appar-ent N digestibility was linearly increasedby a Cl- concentration increase and, in aquadratic way, first by HC03-, then byHP042- concentrations and the Na+/K+ ratio.
The effects of experimental factors on trueN digestibility were more complex, with alinear increase with respect to HC03 con-
centration and a favourable influence of low
and high values of HP042- and Cl-. TheNa+/K+ ratio was involved in a number of
positive interactions, while both its linearand quadratic coefficients were clearly neg-ligible.
For NAN and microbial N outflows,regression analysis led to acceptable fits(table V17. The microbial N outflows var-ied from 0.304 to 0.390 g.d-I when the


microbial reference was LAB, and from0.429 to 0.555 g-d-I when the presence ofSAB was also considered. The estimationof EMPS ranged from 26.2 to 37.1 g g N.kg-IOMTD. Whatever the calculation method,the models for the amount of microbial pro-tein synthesised showed a curvilinear posi-tive effect of HPO42-, and opposed linearinfluences of HC03 and Cl- (figures 3 and4). The EMPS estimation (table VI) was sat-isfactorily modelled, with strong curvilin-ear positive influences of HPO4z- andHC03 concentrations (figure 5) and a neg-ative effect of Cl- interacting with HC03-.Maxima occurred for high values of HP042-and HC03 and low values of Cl-. The com-
position of LAB is presented in table VII.All the responses except the N contents weresatisfactorily modelled. Bacterial organicmatter ranged from 82.2 to 90.3 g/100 gDM. It showed a linear negative effect ofHP04z- and HC03-1 and was also lowered
by low or high values of Na+/K+. Thenucleic acid contents, the N contents andthe ratio [nucleic acids]/[nitrogen] werecomprised between 61.7 and 75.3 mg-g-1OM, 90.5 and 109.5 mg.g-I OM, 0.634 and0.734 g-g-1, respectively. With nucleic acids,
the most active factor was clearly the Na+/K+ratio, acting both in a curvilinear negativeway and through negative interactions withthe other factors. The nucleic acid contentwas the lowest for high Na+/K+ values. TheN content was merely influenced by Na+/K+and Cl-. The regression analysis for the ratio[nucleic acids]/[nitrogen] mainly demon-strated opposite curvilinear influences ofHC03 and Cl-.
4. DISCUSSION
The present paper, which deals with theeffects of mineral compounds on feed degra-dation and microbial growth in a continu-ous culture system, completes former reportsabout the action of these minerals on micro-bial fermentation in the same experiment([9]; Broudiscou et al., in press). Since alldata have been analysed, our experimentalscheme can now be thoroughly evaluatedas a strategy whose benefits make it superiorto a number of others. As Cl- effects
appeared to be for the most part negligible,would it have been wiser to screen all factors


and include only the most important onesin the polynomial model? With such anapproach, one first implements a factorial3 x 23 design, containing 12 experiments[25], to obtain all the main effects and two-factor interactions. One then adds at leastseven (preferably ten) experiments to theinitial matrix to generate a central compos-ite design and to model the actions ofHP04z-, HC03 and Na+/K+, which haveproved to be important factors. In thissequential strategy, the total number ofexperiments was at least 19, instead of 16 inthe Franquart matrix, to obtain similar infor-mation on the processes under study. Ourexperimental strategy was justified by therelatively small number of factors underconsideration.
The degradabilities and the EMPSreported in the present paper were all inaccordance with the values commonly mea-sured in animals. The difference between
tdoM and dNpF, in particular, accounted forthe extensive degradation of barley starch,almost absent from effluent samples. Themethod used to calculate tdoMin our exper-iment was justified by the imbalancebetween OM inflows and outflows observedin all our experimental runs. The most prob-able explanation was the presence of sig-nificant amounts of VFA in freeze-driedeffluent samples. In ruminants, most of theVFA produced is absorbed across theforestomach walls, without reaching theduodenal fistula [22]. In our fermenters, allthe fermentation end-products were col-lected. An analysis of water-extracts of driedeffluent samples clearly showed that VFAwere only partially eliminated throughfreeze-drying. These VFA are inadvertentlytaken for undegraded dietary OM (or DM) ifone applies a method adapted from in vivoexperiments to calculate tdoM - or true DMdigestibility, as described by Crawford etal. [17]. In our experiment, the equationused to estimate the OMF took into accountthe fermentation of pentoses releasedthrough pectin and hemicellulose degrada-tion [34], and the much less significant fer-
mentation of carbon chains resulting fromamino acid deamination. The mineral con-centrations in our experimental domain werecomprised within physiological limits [20].The ions linked to each experimental fac-tor have well-defined metabolic roles. Phos-
phates contribute to the buffering capacity ofthe medium, and phosphorus is a constituentof nucleic acids, phospholipids and coen-zymes. Bicarbonates are efficient buffers ata pH near 6.5, and methanogens use carbondioxide as an electron acceptor. Chloridesalts merely change osmotic pressure. Potas-sium is a major ion in microbial cells, espe-cially as a cofactor for such enzymes asphosphohexokinases. A number of halo-philic rumen bacterial species requiresodium [20].
As for fermentation variates (Broudis-cou et al., in press), buffers (HC03-, HP042-)were the most active factors on substrate
hydrolysis and microbial synthesis. How-ever, while fermentation and OM degrada-tion were mainly influenced by HC03-1HP04z- appeared to be a major factor in thedegradation of cell wall constituents andproteins. It has been clearly established [37]that a deficiency in phosphorus inhibits cel-lulolysis. In our experiment, phosphoruswas supplied in a readily available form andapparently met microbial requirementsexpressed in terms of fermentable energyin all runs, with at least 20 mg-g-I OMF
[21 ]. Nevertheless, the explored domain wasprobably wide enough to induce, in a num-ber of experimental runs, phosphorus con-centrations low enough to be slightly harm-ful. The effects of high values have beenpoorly documented and their cause wasunclear. As expected, NDF degradabilityfollowed the same pattern as acetate pro-duction (Broudiscou et al., in press). Bicar-bonate probably influenced cell wall degra-dation by changes in pH. Stewart [48]reported a similar relationship between pHand cotton degradation by mixed rumenmicrobes. The nature of cations influencedfibre degradation as well, in accordance withthe findings of Hubbert et al. [28] on cellu-

lose digestion in batch incubations of mixedsuspensions of rumen micro-organisms. StOmer and Roberts [49], however, found noeffect of potassium on crude fibre apparentdigestibility in experiments with heifers.The model fit to true nitrogen degradabil-ity presented a specific feature - significantpositive interactions between the Na+/K+ratio and other factors - which might bemerely interpreted as a positive effect ofsmall amounts of potassium on proteinhydrolysis. Mackie and Therion [39] havestressed the greater sensitivity of microbesto higher concentrations of potassium thansodium. In our experiments, the concentra-tions of Na+ and K+ in the saliva varied from2.1 to 4.4 g-L-1 and from 0.22 to 1.3 g!L-!,respectively. One should note that the natureof cations also affected the branched chainVFA production rate (Broudiscou et al., inpress). Both observations suggest specificmetabolic requirements for a number of pro-teolytic bacterial strains. In general, our dataclearly demonstrated that most of the vari-ables related to feed degradation were keptconstant in the central part of our experi-mental domain.
The microbial matter flowing out of thefermenters was mostly of bacterial origin.Taking into account the protozoa bio-vol-umes, chemical composition and outflowrate reviewed by Williams and Coleman[52], one can estimate from protozoa counts[9] that these micro-organisms accountedfor 2-5 % of the daily microbial nitrogenoutflow. The correction for the portion ofbacterial matter outflow of SAB origin wasbased on the bibliography relating to SABchemical composition [6, 33, 40, 42, 54]and to SAB/LAB ratio in the ruminal orduodenal bacterial matter [33, 53, 54] on amixed diet. This calculation method led tomore sensible values for MN outflow, tdoMand EMPS, but did not modify the conclu-sions on the influence of experimental fac-tors, which were only based on observationsof LAB. In our experiment, the daily out-flow of microbial N was kept relatively con-stant in a large central area of the experi-
mental domain but tended to vary inverselyto OM degradation as a function of experi-mental factors. This trend led to a minimi-sation of the EMPS at intermediate values of
mineral inputs. Nevertheless, one must notethat EMPS, although well modelled, variedwithin a rather narrow range. Within our
experimental conditions, the practical con-sequences of mineral supply on microbialprotein synthesis were hardly noticeable.The comparison of the effect of HP04z- onmicrobial protein synthesis with its effecton fibre degradation, in particular, was con-sistent with the findings of Komisarczuk etal. [37] on the different phosphorus require-ments for cellulose digestion and for pro-tein synthesis. The chemical compositionof bacterial matter was markedly altered byminerals. The ash content of LAB, althoughpartly related to the mineral content of washsolution used in the isolation procedure [40],varied directly as buffer salt concentrationsin the culture medium. Moreover, the natureand amount of cations strongly influencedthe nucleic acid content in bacterial organicmatter and to a lesser degree, the N content,which might be interpreted both as a changein the specific composition of bacterial pop-ulations and as an effect on bacterial growthrates.
In our experimental conditions, the influ-ences of minerals on feed degradation andprotein synthesis were reduced in scale.Fibre degradability was the only variablewhich was simultaneously well modelledand exhibited extended variations through-out the experimental domain. Moreover,most of the parameters investigated werekept relatively constant within a large cen-tral area of the domain. In conclusion, theartificial saliva formulated in a former paper[9] and containing intermediate amounts ofbuffer salts should warrant an acceptablestability for in vitro studies of microbial pro-cesses in the rumen.

ACKNOWLEDGEMENTS
The authors would like to thank Dr S. Komis-
arzuk-Bony for analysis of phosphorus in feeds,Mr R. Bergeault for fibre determination, MrsB. Lassalas for other laboratory analyses, andL. L’Hotelier for taking care of experimental ani-mals.
REFERENCES
[1] Aafjes J.H., Nijhof J.K., A simple artificialrumen giving good production of volatile fattyacids, Br. Vet. J. 123 (1967) 436-445.
[2] ] Atlas R.M., Bartha R., Microbial Ecology, Fun-damentals and Applications, Benjamin/Cum-mings Publishing Company, Redwood City,1993.
[3] ] Benoist D., Tourbier Y., Germain-Tourbier S., .,
Plans d’experiences : Construction et Analyse,Lavoisier, Paris, 1994.
[4] Bowie W.C., In vitro studies of rumen micro-organisms, using a continuous-flow system, Am.J. Vet. Res. 23 (1962) 858-868.
[5] Box G.E.P., Hunter W.G., Hunter J.S., Statis-tics for Experimenters: An Introduction toDesign, Data Analysis and Model Building, JohnWiley, New York 1978.
[6] Broudiscou L.-P., Introduction d’un hydrolysatd’huile de soja ou d’une huile de lin dans laration de moutons : influence sur la digestiondes aliments, these, Institut national agrono-mique Paris-Grignon, 1988.
[7] Broudiscou L.-P., Pochet S., Poncet C., Effect oflinseed oil supplementation on feed degrada-tion and microbial synthesis in the rumen of cil-iate-free and refaunated sheep, Anim. Feed Sci.Technol, 49 (1994) 189-202.
[8] Broudiscou L.-P., Papon Y., Fabre M., Broud-iscou A.F., Maintenance of rumen protozoa pop-ulations in a dual outflow continuous fermenter,J. Sci. Food Agric. 75 ( 1997) 273-280.
[9] Broudiscou L.-P., Papon Y., Broudiscou A.F.,Effect of minerals on methane production andprotozoa numbers in continuous culture ofrumen micro-organisms. Reprod. Nutr. Dev.suppi. 1 (1997) 70-71. 1.
[ 10] Broudiscou, L.-P., Papon, Y., Broudiscou, A.F.,Effet de la supplementation minerale sur le m6ta-bolisme azot6 des micro-organismes du rumen,étude en fermenteur à effluent double, in: Pro-ceedings of the 5th Rencontres RecherchesRuminants, 2-3 December 1998, Inra Editions,Paris, 1998.
[11] Bryant M.P., Robinson I.M., Chu H., Observa-tions on the nutrition of Bacteroides succino-
genes - a ruminal cellulolytic bacterium, J. DairySci. 42 (1959) 1831-1847.
[ 12] Caldwell D.R., Keeney M., Barton J.S., Kelley J.F.,Sodium and other inorganic growth require-ments of Bacteroides amylophilus, J. Bacteriol.114 (1973) 782-789.
[13] Caldwell D.R., Arcand C., Inorganic and metal-organic growth requirements of the genus Bac-teroides, J. Bacteriol. 120 ( 1974) 322-333.
[14] Clarke R.T.J., Methods for studying gutmicrobes, in: Clarke R.T.J., Bauchop T. (Eds.), ),Microbial Ecology of the Gut, Academic Press,London, 1977, pp. (-33.
[15] Cochran W.G., Cox G.M., ExperimentalDesigns, John Wiley, New York, 1957.
[ 16] Cote R.J., Gherna R.L., Nutrition and media,in: Gerhardt P., Murray R.G.E., Wood W.A.,Krieg N.R. (Eds.), Methods for General andMolecular Bacteriology, ASM, Washington,1994, pp. 155-177
[17] Crawford R.J., Hoover W.H., Knowlton P.H.,Effects of solids and liquid flows on fermenta-tion in continuous cultures. 1. Dry matter andfiber digestion, VFA production and protozoanumbers, J. Anim. Sci. 51 (1980) 975-985.
[18] Davies A.W., Taylor, K., Application of theautoanalyser in a river authority laboratory, inSymposium Technicon, Technicon, Tarrytown,1965, pp. 294-300.
[ l9] Demeyer D.I., Van Nevel C.J., Methanogenesis,an integrated part of carbohydrate fermentationand its control, in: McDonald I.W., WarnerA.C.I. (Eds.), Digestion and Metabolism in theRuminant, University of New England Pub-lishing, Armidale, 1975, pp. 366-382.
[20] Durand M., Kawashima R., Influence of min-erals in rumen microbial digestion, in: Rucke-bush Y., Thivend P. (Eds.), Digestive Physiologyand Metabolism in Ruminants, MTP Press, Lan-caster, 1980, pp. 375-408.
[21 ] Durand M., Komisarczuk S., Influence of majorminerals on rumen microbiota, J. Nutr. 1 18 8
( 1988) 249-260.
[22] France J., Siddons R.C., Volatile fatty acid pro-duction, in: Forbes J.M., France J. (Eds.), Quan-titative Aspects of Ruminant Digestion andMetabolism, C.A.B. International, Wallinford,1993, pp. 107-121.
[23] Franquart P., Optimisations multicritères etm6thodologie de la recherche exp6rimentale,these, universit6 d’Aix-Marseille, 1992.
[24] Gray F.V., Weller R.A., Pilgrim A.F., Jones,G.B., A stringent test for the artificial rumen,Austr. J. Agric. Res. 13 (1962) 343-349.
[25] Haaland P.D., Experimental Design in Biotech-nology, Marcel Dekker, New York, 1989.
[26] Halpern Y.S., Barash H., Dover S., Druck K.,Sodium and potassium requirements for activetransport of glutamate by Escherichia coli K-12, J. Bacteriol. 114 (1973) 53-58.

[27] Hino T., Sugiyama M., Okumura K., Mainte-nance of protozoa and methanogens, and fiberdigestion in rumen-simulating continuous cul-ture, J. Gen. Appl. Microbiol. 39 ( 1993) 35-45.
[28] Hubbert F., Cheng E., Burroughs W., The influ-ence of potassium, sodium, rubidium, lithiumand cesium on in vitro cellulose digestion byrumen micro-organisms with observations uponsodium and potassium influences in lamb fat-tening rations, J. Anim. Sci. 17 (1958) 576-585.
[29] Iijima T., Diesterhaft M.D., Freese E., Sodiumeffect of growth on aspartate and genetic analysisof a Bacillus subtilis mutant with high aspar-tase activity, J. Bacteriol 129 (1977) 1440-1447.
[30] Ingram L.O., Thurston E.L., Potassium require-ments for cell division in Anacystis nidulans,J. Bacteriol. 125 ( 1976) 369-371. 1 .
[31J 1 Jouany J.-P., Volatile fatty acid and alcoholdetermination in digestive contents, silage juices,bacterial cultures and anaerobic fermentor con-
tents, Sci. Aliments 2 ( 1982) 131-144.
[32] Lassalas B., Jouany J.P., Broudiscou, L., Dosagedes bases puriques et pyrimidiques par chro-matographie liquide a haute performance, Ann.Zootech.42 (1993) 170-171. 1.
[33] Legay-Carmier F., Bauchart D., Distribution ofbacteria in the rumen contents of dairy cowsgiven a diet supplemented with soya-bean oil,Br. J. Nutr. 61 (1989) 725-740.
[34] Lin K.W., Patterson J.A., Ladisch M.R., Anaer-obic fermentations: microbes from ruminants,Enz. Microb. Technol. 7 ( 1985) 98-107.
[35] Khorasani G.R., Armstrong D.G., Effect ofsodium and potassium on overall digestibilityof a semi-purified diet and microbial proteinproduction in the rumen of sheep, LivestockProd. Sci. 24 (1990) 347-357.
[36] Khuri A.I., A measure of rotatability forresponse-surface designs, Technometrics 30(1988)95-104.
[37] Komisarczuk S., Merry R.J., McAllan A.B.,Effects of different levels of phosphorus onrumen microbial fermentation and synthesisdetermined using a continuous culture system,Br. J. Nutr. 57 (1987) 279-290.
[38] McDougall E.I., Studies on ruminant nutrition.I. The composition and output of sheep’s saliva,Biochem. J. 43 (1948) 99-109.
[39] Mackie R.L, Therion J.J., Influence of mineralinteractions on growth efficiency of rumen bac-teria, in: Gilchrist F.M.C., Mackie R.I. (Eds.),Herbivore Nutrition, Craighall: Science Press,London, 1984, pp. 455-477.
[40] Martin C., Williams A.G., Michalet-Doreau B., .,
Isolation and characteristics of the protozoaland bacterial fractions from bovine ruminal con-tents, J. Anim. Sci. 72 ( 1994) 2962-2968.
[41 ] Mees D.C, Merchen N.R., Mitchel C.J., Effectsof sodium bicarbonate on nitrogen balance, bac-terial protein synthesis and sites of nutrientdigestion in sheep, J. Anim. Sci. 61 (1985)985-994.
[42] Merry R.J., McAllan A.B., A comparison of thechemical composition of mixed bacteria har-vested from the liquid and solid fractions ofrumen digesta, Br. J. Nutr. 50 (1983) 701-709.
[43] O’Brien R.W., Stern J.R., Requirements forsodium in the anaerobic growth of Aerobacteraerogenes on citrate, J. Bacteriol. 98 ( 1969)388-393.
[44] Peissik A., M6thodologic de la recherche exp6-rimentale : propri6t6s et caract6ristiques desmatrices d’exp6riences pour les modèles poly-nomiaux du second degr6, these, universited’Aix-Marseille, 1995.
[45] Rufener W.H., Nelson W.O., Wolin M.J., Main-tenance of the rumen microbial population incontinuous culture, Appl. Microbiol. 1 (1963)196-201.
[46] SAS/STAT@, User’s Guide, Release 6.04, SASInstitute Inc, Cary, 1990.
[47] Sistrom W.R., A requirement for sodium in thegrowth of Rhodopseadomonas spheroides,J. Gen. Microbiol. 22 (1960) 778-785.
[48] Stewart C.S., Factors affecting the cellulolyticactivity of rumen contents, Appl. Environ.Microbiol. 33 (1977) 497-502.
149J St Omer V.V.E., Roberts W.K., Some effectsof dietary potassium upon digestibility, serumelectrolytes and utilization of potassium, sodium,nitrogen and water in heifers, Can. J. Anim. Sci.47 ( 1967) 39!6.
[50] Van Soest P.J., Wine R.H., Use of detergentsin the analysis of fibrous feeds. Determination ofplant cell-wall constituents, J. A. O. A. C. 50(1967) 50-55.
[5 1] ] Weller R.A., Pilgrim A.F., Passage of protozoaand volatile fatty acids from the rumen of thesheep and from a continuous in vitro fermenta-tion system, Br. J. Nutr. 32 (1974) 34I-351. 1.
1521 Williams A.G., Coleman G.S., The Rumen Pro-tozoa, Springer Verlag, New York, 1992.
[53] Wolstrup J., Jensen K., Adenosine triphosphateand deoxyribonucleic acid in the alimentary tractof cattle fed different nitrogen sources, J. Appl.Bacteriol. 45 (1978) 49-56.
[54] Yang W.Z., Etude cin6tique de la colonisationmicrobienne des aliments dans le rumen du mou-ton. Consequences sur la compartimentation dela biomasse et sur sa dynamique de sortie durumen dans le cas de diff6rents types de rations,these, universit6 de Clermont-Ferrand-11, 1992.