effects of maternal chlorpromazine on offspring nervous system development
TRANSCRIPT

Nrurophurnwcoloq~ Vol. 17. pp 5X3-587 0 Pergamon Press Ltd 197X. Printed in Great Britain
EFFECTS OF MATERNAL CHLORPROMAZINE ON OFFSPRING NERVOUS SYSTEM DEVELOPMENT*
J. W. DAILEY
Department of Pharmacology and Therapeutics. Louisiana State University, School of Medicine in Shreveport, P.O. Box 33932, Shreveport, Louisiana 71130. U.S.A.
(Accepted 10 January 1978)
Summary-Administration of chlorpromazine (7mg/kg/day) to rat dams from day 8 of pregnancy until the pups were weaned produced small but statistically significant decreases in litter size and birth weight. The difference in pup weight was no longer present at the time of weaning. The offspring of chlorpromazine-treated dams were less able to maintain body temperature in response to restrained cold stress as determined when they were 60-65 days of age. They also incorporated significantly less 14C from tyrosine into heart norepinephrine during acute cold exposure. No differences in incorpor- ation of 14C into norepinephrine from tyrosine were evident when the animals were not subjected to cold stress. Similarly treated offspring did not have an increase in superior cervical ganglion tyrosine hydroxylase activity after 48 hr of cold exposure while the offspring of control dams did have a signifi- cant increase in the activity of this enzyme. These data suggest that maternal administration of chlorpro- mazine produces a permanent alteration in the ability of the offspring to respond to cold stress and that this deficit is related to an alteration in nervous system development.
Drugs administered to the pregnant or lactating female are, in many cases, also presented to the foetus or nursing offspring. Long-lasting changes in brain monoamine levels and metabolism in the offspring of rats treated with methylamphetamine, chlorproma- zine and phencyclidine have been reported by Tonge (1972, 1973a,b,c). Other authors have found that ad- ministration of the neuroleptic penfluridol to rat dams on days 1, 3 and 5 of suckling can produce alterations in brain tyrosine hydroxylase activity and behaviour in the offspring at 4 weeks of age (Ahlenuis, Brown, Engel and Lundborg, 1973; Engel and Lund- berg, 1974). In addition, it has been reported that administration of relatively large doses of reserpine lo rats for specific periods during pregnancy can pro- duce permanent alterations in adrenal and brain tyro- sine hydroxylase activity and in adrenal dopamine-/I- hydroxylase activity in the -offspring (Bartolome, Seidler, Anderson and Slotkin, 1976; Bartolome and Slotkin, 1975).
In other studies it has been found that administra- tion of reserpine to rat dams from day 8 of pregnancy until the pups were weaned at approximately 20 days of age can produce a long-lasting and perhaps per- manent alteration in the ability of the offspring to maintain body temperature in response to restrained cold stress (Dailey, 1976, 1978). This altered capacity to maintain body temperature was accompanied by a decreased ability to increase norepinephrine syn- thesis in response to cold exposure.
*A preliminary report appeared in Pharmacologist 18: 231 (1976).
Key words: chlorpromazine, cold stress, norepinephrine synthesis, tyrosine hydroxylase activity.
The purpose of the present study was to determine if administration of chlorpromazine to pregnant and nursing rats would cause alterations in restrained cold stress response and in sympathetic nervous system ac- tivity.
METHODS
Drug treatment
Timed pregnant Sprague-Dawley rats (Flow Laboratories) were housed individually in plastic breeding cages. From day 8 of pregnancy until the pups were weaned at approximately 20 days of age, the treated dams received daily subcutaneous injec- tions of 7mg/kg of chlorpromazine (Thorazine; Smith, Kline & French). Control animals received daily injections of saline. Food consumption was not different between control and treated animals during pregnancy. The offspring were weighed within 24 hr of birth. At weaning, the pups were transferred to wire cages and when sex differences became readily distinguishable (30-40 days of age) the animals were segregated according to sex. All offspring were main- tained in their home cages and allowed free access to food and water until used in experiments.
Determination of cold stress response
Male offspring of control and chlorpromazine- treated dams were exposed to restrained cold stress when they reached 665 days of age. For this pro- cedure, the animals were placed in tight-fitting wire restraining cages and had thermistor probes which were connected to a YSI telethermometer inserted ap- proximately 1Ocm into the rectum. They were trans- ferred to the cold room (6°C) where the drop in core
583

584 J. W. DAILEY
body temperature was monitored over 1 hr. In order to take into account small differences in body size which would affect the rate of cooling, the results are expressed as heat deficit per unit surface area or heat debt (Burton and Edholm, 1955).
Estimation of norepinephrine synthesis
In two experiments, the rate of synthesis of nor- epinephrine from labelled tyrosine was estimated in 7&80day-old male offspring of control and chlor- promazine-treated dams by measuring the specific activity of 14C tyrosine and i4C norepinephrine in the heart (Sedvall, Weise and Kopin, 1968). The 14C tyro- sine was purified by ion-exchange chromatography (Bio-Rad AG-50) before use. In the first experiment, offspring of control and chlorpromazine-treated dams were injected intravenously with 4OpCi of uniformly labelled i4C tyrosine (New England Nuclear). The animals were immediately transferred to the cold room (4°C) for 55 min. A total of 60min after the i4C tyrosine administration, the animals were decapi- tated. Their hearts were removed and homogenized in 10% trichloroacetic acid. Norepinephrine and its precursors and metabolites were separated by ion- exchange chromatography (Bio-Rad AG-50) as pre- viously described (Dailey, 1978). Labelled norepineph- rine and tyrosine were quantitated by liquid scintil- lation spectrometry. Endogenous tyrosine levels were determined by the method of Waalkes and Uden- friend (1957). Endogenous norepinephrine levels were measured by the automated trihydroxyindole pro- cedure of Viktora, Baukal and Wolff (1968) after this procedure had been optimized for quantitation of norepinephrine from rat tissues.
In a second experiment, 70-80-day-old offspring of control and chlorpromazine-treated dams were ad- ministered SO&i of 14C tyrosine via the tail vein and, after 60 min at room temperature (22”C), the ani- mals were sacrificed by decapitation. Immediately after sacrifice, their hearts were removed and specific activities of norepinephrine and tyrosine were deter- mined as described above.
Estimation of tyrosine hydroxylase activity
Offspring (70-80 days old) of control and chlorpro- mazine-treated dams were divided into two groups. One group which was composed of offspring of con-
trol and chlorpromazine-treated dams was placed in the cold room (4°C) for 48 hr in order to bring about induction of tyrosine hydroxylase (Hanbauer, Kopin and Costa, 1973). A second group of animals which was also composed of offspring of control and chlor- promazine-treated dams was housed at room tem- perature (22°C). At the end of 48 hr. the animals housed at room temperature and those housed in the cold were sacrificed and their superior cervical gang- lia were removed. Tyrosine hydroxylase activity was estimated in the superior cervical ganglia by a modifi- cation of the method of Nagatsu, Levitt and Uden- friend (1964). For the modified procedure, 6-methyl- 5,6,7,8-tetrahydropterine-HCl (Calbiochem) was used as the enzyme co-factor and the tritiated water was separated from other radioactive components by chromatography on charcoal.
RESULTS
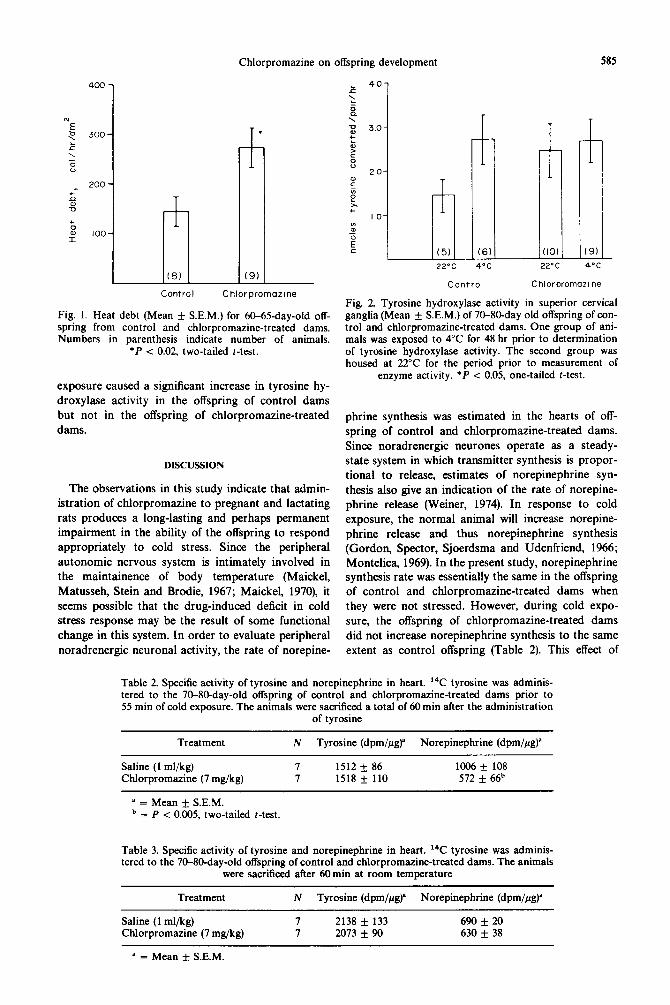
Chlorpromazine-treated dams had significantly fewer pups per litter and the average birth weights of their pups were lower than for saline-treated dams (Table 1). The body weight differences between the two groups of offspring were no longer present when the pups were weaned at approximately 20 days of age. Figure 1 illustrates the restrained cold stress re- sponse in these offspring. The 60-65-day-old offspring of chlorpromazine-treated dams had significantly higher heat debts than control offspring. The body temperature of the two groups of animals was not different prior to cold exposure.
The incorporation of 14C from tyrosine into heart norepinephrine was determined during cold exposure. The offspring of chlorpromazine-treated dams had a significantly lower heart norepinephrine specific ac- tivity than control offspring following 55 min of expo- sure to a temperature of 4°C even though the specific activity of heart tyrosine was not different between the two groups (Table 2). Norepinephrine synthesis was also estimated when similar groups of animals were housed at room temperature (22°C) prior to sac- rifice. Table 3 illustrates that there was no significant difference in the specific activity of either tyrosine or norepinephrine in the hearts of the two groups of rats housed at 22°C following intravenous injection of 14C tyrosine. Figure 2 shows that 48 hr of cold
Table 1. The litter size, birth weight and weaning weight of offspring from dams treated with chlorpromazine. Pregnant rats were given daily subcutaneous injections in the dosage indicated
from day 8 of pregnancy until the pups were weaned
Treatment Litters Litter size” Birth weight (g) Weaning weight (g)
Control (saline 1 ml/kg) Chlorpromazine (7 mg/kg)
12 10.83 + 0.46 6.78 f 0.19 45.92 & 1.57
11 9.18 k 0.69b 6.08 + 0.17’ 46.98 k 2.36
a = Mean f S.E.M. b = P < 0.05, two-tailed t-test. ' = P -e 0.02, two-tailed r-test.
Chlorpromazine on offspring development 585
NE ? 300 L
c 0 u
_ 200
2 %
+
z I
IOC -II (8)
1’ I-
(9)
Control Chlorpromazine
Fig. 1. Heat debt (Mean + S.E.M.) for 6&65-day-old off- spring from control and chlorpromazine-treated dams. Numbers in parenthesis indicate number of animals.
*P < 0.02, two-tailed t-test.
exposure caused a significant increase in tyrosine hy- droxylase activity in the offspring of control dams but not in the offspring of chlorpromazine-treated dams.
DISCUSSION
The observations in this study indicate that admin- istration of chlorpromazine to pregnant and lactating rats produces a long-lasting and perhaps permanent impairment in the ability of the offspring to respond appropriately to cold stress. Since the peripheral autonomic nervous system is intimately involved in the maintainence of body temperature (Maickel, Matusseh, Stein and Brodie, 1967; Maickel, 1970), it seems possible that the drug-induced deficit in cold s8tress response may be the result of some functional change in this system. In order to evaluate peripheral noradrenergic neuronal activity, the rate of norepine-
r (51
t
I (6: -
t
IO -
t
9) -
Control C hlorpromozl ne
Fig. 2. Tyrosine hydroxylase activity in superior cervical ganglia (Mean f S.E.M.) of 7&80-day old offspring of con- trol and chlorpromazine-treated dams. One group of ani- mals was exposed to 4°C for 48 hr prior to determination of tyrosine hydroxylase activity. The second group was housed at 22°C for the period prior to measurement of
enzyme activity. *P < 0.05, one-tailed t-test.
phrine synthesis was estimated in the hearts of off- spring of control and chlorpromazine-treated dams. Since noradrenergic neurones operate as a steady- state system in which transmitter synthesis is propor- tional to release, estimates of norepinephrine syn- thesis also give an indication of the rate of norepine- phrine release (Weiner, 1974). In response to cold exposure, the normal animal will increase norepine- phrine release and thus norepinephrine synthesis (Gordon, Spector, Sjoerdsma and Udenfriend, 1966; Montelica, 1969). In the present study, norepinephrine synthesis rate was essentially the same in the offspring of control and chlorpromazine-treated dams when they were not stressed. However, during cold expo- sure, the offspring of chlorpromazine-treated dams did not increase norepinephrine synthesis to the same extent as control offspring (Table 2). This effect of
Table 2. Specific activity of tyrosine and norepinephrine in heart. r4C tyrosine was adminis- tered to the 70-80-day-old offspring of control and chlorpromazine-treated dams prior to 55 min of cold exposure. The animals were sacrificed a total of 60 min after the administration
of tyrosine
Treatment N Tyrosine (dpm/pg)” Norepinephrine (dpm/pg)
Saline (1 ml/kg) 7 1512 f 86 1006 + 108 Chlorpromazine (7 mg/kg) 7 1518 + 110 572 f 66b
a = Mean f S.E.M. b = P < 0.005, two-tailed r-test.
Table 3. Specific activity of tyrosine and norepinephrine in heart. r4C tyrosine was adminis- tered to the 7&80day-old offspring of control and chlorpromazine-treated dams. The animals
were sacrificed after 60min at room temperature
Treatment N Tyrosine (dpm/pg) Norepinephrine (dpm/pg)
Saline (1 ml/kg) 7 2138 f 133 690*20 Chlorpromazine (7 mgjkg) 7 2073 + SO 630 f 38
a = Mean + S.E.M.

586 J. W. DAILEY
chlorpromazine on norepinephrine synthesis supports the concept that the deficit in cold stress response in offspring rats is due to an alteration in peripheral autonomic nervous system activity. Administration of reserpine (0.1 mg/kg per day) to pregnant rats has also been reported to produce an impairment in the ability of the offspring to maintain body temperature (Skol- nick, 1972: Dailey, 1976) and a decrease in the capa- city to increase norepinephrine synthesis in response to acute cold exposure (Dailey, 1978). Reserpine ad- ministration did not cause alterations in litter size and birth weight as was the case with chlorpromazine treatment.
It has also been shown that exposure to cold will cause induction of tyrosine hydroxylase in the super- ior cervical ganglia of rats (Thoenen, 1970; Thoenen, Kettler, Burkard and Saner, 1971; Hanbauer et al., 1973). This induction is thought to be brought about by a greater rate of nerve impulse traffic from higher centres as a result of the increased central nervous system activity that occurs in response to cold stress. The statistically significant increase in tyrosine hy- droxylase activity in the cold exposed control animals in the present study indicates that the stress employed was severe enough to cause induction of the enzyme in untreated animals (Fig. 2). This cold stress did not, however, cause the offspring of chlorpromazine- treated dams to increase their superior cervical gang- lion tyrosine hydroxylase activity above that for chlorpromazine otfpsring housed at room tempera- ture. The superior cervical ganglion tyrosine hydroxy- lase activity for offspring of chlorpromazine-treated dams housed at room temperature may already have been partially induced when compared with that of control offspring housed at room temperature but this difference was not statistically significant. The failure of cold exposure to increase superior cervical gang- lion tyrosine hydroxylase activity in the offspring of chlorpromazine-treated dams is consistent with the hypothesis that the drug-induced deficit is present in peripheral adrenergic neurones. Perhaps chlorproma- zine produces an alteration such that the peripheral adrenergic nerves are unable to synthesize a sufficient number of active molecules of tyrosine hydroxylase and thereby to keep pace with the increased demand for norepinephrine release which occurs when the rate of impulse traffic from higher centres increases in re- sponse to cold exposure.
However, the data presented in this report are not sufficient to confirm the aforementioned hypothesis and are equally as consistent with the hypothesis that central nervous system function may be altered in the offspring of chlorpromazine-treated dams. That is, if chlorpromazine causes the hypothalamic and/or spinal cord tracts that participate in the response to cold stress (Bligh, 1973) to function abnormally, the appropriate stimuli from these higher centres would not reach the peripheral noradrenergic neurones. Thus, these animals would not be able to bring about an appropriate increase in norepinephrine synthesis
or an induction of superior cervical ganglion tyrosine hydroxylase activity in response to cold stress. As a result of these deficits, the animals would be less able to maintain body temperature in response to re- strained cold stress.
Acknowledgements-The skilled technical assistance of Messrs Stephen Ikeda, Larry Thompson and Fleet Cook is greatly appreciated. The work was supported in part by a grant from the National Foundation, March of Dimes.
REFERENCES
Ahlenius, S., Brown, R., Engel, J. and Lundborg, P. (1973). Learning deficits in 4-week-old offspring of the nursing mothers treated with the neuroleptic drug penfluridol. Naunyn-Schmiedeberg’s Arch. exp. Path. Pharmak. 279: 31-37.
Bligh, J. (1973). Temperature Regulation in Mammals and Other Vertebrates. North-Holland, Amsterdam.
Bartolome, J. and Slotkin, T. A. (1975). Prenatal reserpine administration: Permanent changes in adrenal tyrosine hydroxylase and dopamine-/I-hydroxylase. Psychopharm. Comm. 1: 46472.
Bartolome, J., Seidler, F. J., Anderson, T. R. and Slotkin, T. A. (1976). Effects of prenatal reserpine administration on development of the rat adrenal medulla and central nervous system. J. Pharmac. exp. Ther. 197: 293-302.
durton, A. C. and Edholm, A. G. (1955). Man In A Cold Enoironment. Arnold, London.
Dailev. J. W. (1976). Effect of reseruine on the cold stress . ~ I
response in rats and its possible-relation to adrenergic nervous system function. Fedn Proc. Fedn Am. Sots exp. Biol. 35: 585.
Dailey. J. W. (1978). Effects of maternally administered res- erpine on the development of the cold stress response and its possible relation to adrenergic nervous system function. Res. Communs. them. Pathol. Pharmac. 19: 389402.
Engel, J. and Lundborg, P. (1974). Regional changes in monoamine levels and in the rate of tyrosine and trypto- phan hydroxylation in 4-week-old offspring of nursing mothers treated with the neuroleptic penfluridol. Naunyn-Schmiedeberg’s Arch. exp. Path. Pharmak. 282 327-334.
Gordon, R., Spector, S., Sjoerdsma, A. and Udenfriend, S. (1966). Increased synthesis of norepinephrine and epinephrine in the intact rat during exercise and expo- sure to cold. J. Pharmac. exp. Ther. 153: 440447.
Hanbauer, I., Kopin, I. J. and Costa, E. (1973). Mechan- isms involved in the transynaptic increase of tyrosine hydroxylase and dopamine+hydroxylase activity in sympathetic ganglia. Naunyn-Schmiedeberg’s Arch. exp. Path. Pharmak. 280. 39-48.
Maickel. R. P. (1970). Interaction of drugs with autonomic nervous function and thermoregulation. Fedn Proc. F edn Am. Sots exp. Biol. 2% 1973-1979.
Maickel, R. P., Matusseh, N., Stein, D. N. and Brodie, B. B. (1967). The sympathetic nervous system as a homeostatic mechanism. I. Absolute need for sympathe- tic nervous function in body temperature maintenance of cold-exposed rats. J. Pharmac. exp. Ther. 157: 103-l 10.
Montelica, I. (1969). Urinary excretion of catecholamines and vanilmandelic acid in rats exposed to cold. Acta physiol. Scand. 76: 393-395.
Nagatsu, T., Levitt, M. and Udenfriend, S. (1964). Tyrosine hydroxylase: The initial step in norepinephrine biosyn- thesis. J. biol. Chem. 239: 2910-2917.
Sedvall, G. C., Weise, V. K. and Kopin, I. J. (1968). The rate of norepinephrine synthesis measured in viuo during

Chlorpromazine on offspring development 587
short intervals: Influence of adrenergic nerve impulse ac- tivity. J. Pharmac. exp. Ther. 159: 274282.
Skolnick, P. (1972). The effects of maternally administered reserpine on the response to stress in offspring. Ph.D Thesis. The George Washington University.
Thoenen, H. (1970). Induction of TH in peripheral and central adrenergic neurons by cold exposure of rats. Nature, Lond. 22% 861-862.
Tonge, S. R. (1973b). Permanent alterations m catecholamine concentrations in discrete areas of brain in the offspring of rats treated with methylam- phetamine and chlorpromazine. Br. J. Phannac. 47: 4255427.
Thoenen, H.. Kettler, R., Burkard, W. and Saner, A. (1971). Neurally mediated control of enzymes involved in the synthesis of norepinephrine: Are they regulated as an operational unit? Naunyn-Schmiedeberg’s Arch. exp. Path. Pharmak. 270: 146-160.
Tonge, S. R. (1973~). Some persistent effects of the pre- and neonatal administration of psychotropic drugs on noradrenaline metabolism m discrete areas of rat brain. Br. J. Pharmac. 48: 364-365.
Viktora, J. K., Baukal, A. and Wollf, F. W. (1968). New automated fluorometric methods for estimation of small amounts of adrenaline and noradrenaline. Analyt. Bio- them. 23: 51S528.
Tonge, S. R. (1972). Permanent changes in brain mono- Waalkes, T. P. and Udenfriend, S. (1957). A flurormetric amine metabolism Induced by the admimstration of psy- method for the estimation of tyrosine in plasma and chotropic drugs during the pre- and neonatal periods ttssues. J. Lab. clin. Med. 50: 733-736. in rats. J. Pharm. Pharmac. 24: 149P. Weiner, N. (1974). A critical assessment of methods for
Tonge, S. R. (1973a). Catecholamme concentrattons in dis- the determination of monoamme synthesis and turnover crete areas of the rat brain after the pre- and neonatal rates in uivo. In: Neuropsychopharmacology of Mono- administration of phencychdine and imipramme. J. amines and Their Regulator Enzymes (Usdin, E.. Ed.), Pharm. Pharmac. 25: 164166. pp. 14%159. Raven Press, New York.