effects of light deprivation on recovery from neglect and extinction
TRANSCRIPT

Research report
Effects of light deprivation on recovery from neglect and extinctioninduced by unilateral lesions of the medial agranular cortex and
dorsocentral striatum
T.M. Van Vleet a, S.A. Heldt a, B. Pyter a, J.V. Corwin a,*, R.L. Reep b
a Department of Psychology, Northern Illinois University, De Kalb, IL 60115, USAb Department of Physiological Sciences, University of Florida, Gainesville, FL 32610, USA
Received 5 April 2002; received in revised form 6 August 2002; accepted 6 August 2002
Abstract
A number of previous studies have indicated that an environmental manipulation, 48 h of light deprivation (LD), produces
virtually complete and permanent behavioral recovery of function from neglect induced by medial agranular cortex (AGm) lesions.
LD-induced behavioral recovery from neglect is correlated with physiological changes in the dorsolateral striatum, an area that
contains the projection zone of AGm efferents in the dorsocentral striatum (DCS). In this study, the behavioral effects of 48 h of LD
on subjects with either unilateral DCS, AGm, or combined AGm/DCS lesions were investigated to examine whether the integrity of
the DCS is crucial for behavioral recovery from neglect and whether LD will have a therapeutic effect on extinction deficits. Subjects
were tested for extinction to bilateral simultaneous stimulation of the forepaws, and visual, auditory and tactile neglect. Forty-eight
hours of LD failed to produce behavioral recovery from neglect in rats with DCS lesions, or a therapeutic affect on extinction
deficits in any of the groups. The results of this study further support the crucial role of the DCS in recovery from neglect induced by
AGm lesions and suggests that the DCS may be the crucial site for the mechanisms leading to LD-induced recovery. Further, the
ineffectiveness of LD on extinction suggests that components of the neglect syndrome are dissociable and may require different
therapeutic interventions.
# 2002 Elsevier Science B.V. All rights reserved.
Keywords: Neglect; Light deprivation; Attention; Prefrontal; Extinction; Recovery
1. Introduction
Neglect is a complex and devastating human neurop-
sychological disorder characterized by a failure to attend
to novel or meaningful stimuli presented to the side
contralateral to a brain lesion, in the absence of a
primary sensory or motor dysfunction. Some manifesta-
tion of neglect is found in approximately 40% of all
cases of brain damage [22]. The vast majority (80�/90%)
of all cases of neglect are produced by destruction of one
of the three cortical regions: the dorsolateral prefrontal
cortex (area 8), the cingulate cortex, or the inferior
parietal lobule [22]. The disorder goes far beyond a lack
of responsiveness to contralesional stimuli to include
dramatic attentional and cognitive spatial deficits. The
failure to respond to contralesional stimuli has devastat-
ing effects for the patients’ abilities to demonstrate
independent daily living or to return to work, and the
presence of persistent neglect leads to a poor prognosis
[16,18,25]. Recovery, when it occurs, is spontaneous
over the course of weeks to months, but often it is
incomplete. Many patients continue to ignore or neglect
contralesional stimuli for months or years post-lesion
[20,23]. They are often unaware of their neurological
status (anosognosia), or are unconcerned and effectively
flat (anosodiaphoria) [22]. This lack of awareness and
changes in motivation interfere with successful physical
and occupational rehabilitation [40].
A rodent model of neglect has been developed to
examine the basic mechanisms of neglect and the
potential for recovery of function [2�/4,8,10]. The
* Corresponding author. Tel.: �/1-815-753-7088; fax: �/1-815-753-
8088
E-mail address: [email protected] (J.V. Corwin).
Behavioural Brain Research 138 (2003) 165�/178
www.elsevier.com/locate/bbr
0166-4328/02/$ - see front matter # 2002 Elsevier Science B.V. All rights reserved.
PII: S 0 1 6 6 - 4 3 2 8 ( 0 2 ) 0 0 2 4 6 - 2

symptoms of neglect in rodents following unilateral
lesions of the medial agranular cortex (AGm), the
rodent analog of area 8, are similar to those found in
human neglect patients and include: severe neglect ofvisual, tactile, and auditory stimulation, extinction,
allesthesia/allokinesia, and disorders of spatial proces-
sing [1,8,14,15,27,28,45].
In rats, recovery from AGm-induced neglect has been
found to occur in three contexts: (1) as in humans,
dopamine (DA) agonists produce acute recovery of
function [10,17,19,24,26]; (2) as found in humans,
rodents with neglect demonstrate spontaneous althoughincomplete recovery over the course of weeks to months
[10,26,28]; and (3) exposure to 48 h of light deprivation
(LD) at 4 h post-surgery produces dramatic immediate
recovery [2,15].
Several recent studies in rodents strongly indicate that
the striatum may play a critical role in neglect, and both
drug-induced and spontaneous recovery from neglect
produced by unilateral AGm lesions [50,53�/55,58,59].Vargo and Marshall [53,54] using immediate early gene
expression (IEG) methods found that spontaneous
recovery from neglect was correlated with symmetrical
IEG expression in the dorsolateral striatum, and in a
subsequent study [55] correlated with changes in gluta-
mate receptors in the dorsolateral striatum. The im-
portance of the striatum in AGm lesion-induced neglect
was also suspected because apomorphine (a DA recep-tor agonist) can produce acute recovery [10,26], and
spiroperidol (a DA receptor antagonist) can reinstate
neglect [57]. The role of DA in cortical [10,12,26] and
subcortical neglect [30�/34], led Fleet et al. [17] and
Hurford et al. [24] to examine the effects of bromocrip-
tine, a D2 receptor agonist, in human patients with
chronic neglect. Bromocriptine produced recovery
across a range of measures. When treatment wasterminated, neglect worsened. Recently, Geminiani et
al. [19] found that apomorphine can also produce acute
recovery from neglect in humans.
These findings led to a series of studies designed to
investigate the role of the striatum in recovery from
neglect induced by unilateral destruction of the AGm.
Van Vleet et al. [50] examined the effects of unilateral
destruction of the striatal projection zone of the AGm inthe dorsocentral striatum (DCS) [38]. The results
indicated that unilateral axon-sparing DCS lesions
produced severe neglect and that the DCS operates did
not demonstrate spontaneous recovery, even in subjects
tested at 96 days post-lesion. Further, apomorphine did
not produce a therapeutic effect in the DCS operates. In
another study, Van Vleet et al. [49] examined the effects
of direct infusion of apomorphine into the DCS or alaterally adjacent region of the striatum in subjects with
severe neglect induced by unilateral AGm lesions. Direct
infusion of apomorphine into the DCS produced a
dramatic acute recovery, while infusion into the adjacent
region of the striatum did not produce recovery [49].
The results of these studies suggest that the DCS may be
a crucial region for recovery from neglect induced by
unilateral AGm destruction.Recovery from neglect produced by unilateral AGm
lesions can also be produced by an environmental
manipulation, 48 h of LD. LD produces dramatic,
accelerated recovery from severe neglect that lasts for
the duration of behavioral testing [2,9,14,15]. Vargo et
al. [59] examined the behavioral and neural correlates of
LD-induced recovery and found that recovery from
AGm-induced neglect was inversely correlated withbehavioral activity and with the degree of asymmetry
in c-fos expression in the right and left dorsolateral
striata. LD decreased the lesion-induced asymmetry in
the dorsolateral striatum and produced recovery. The
region described as the dorsolateral striatum in the
Vargo et al. [59] study encompassed much of the DCS.
The Vargo et al. [59] study pointed to the potential
importance of the DCS in LD-induced recovery fromneglect produced by unilateral AGm lesions, but the
data were correlational and included the entire dorso-
lateral striatum rather than the site of the AGm
projections to the DCS. In this study, we directly
examined the therapeutic effectiveness of LD on severe
neglect produced by unilateral AGm and/or DCS
lesions. In addition, we extended what is known about
LD-induced recovery by examining the effects of LD onone of the major deficits associated with the neglect
syndrome in humans, extinction to bilateral simulta-
neous stimulation. Examination of the effects of LD on
extinction deficits is important because extinction defi-
cits often persist in patients who have recovered from
neglect [25].
2. Methods
2.1. Subjects
Subjects were 43 male Long-Evans hooded rats bred
from stock purchased from Harlan Sprague�/Dawley.
Prior to the surgical procedures, the subjects were
handled daily (1�/2 min per day) for 21�/28 days to
gentle them for subsequent behavioral testing. Animalswere tested between 12:00 and 17:00 h. All procedures
conformed to institutional guidelines for the care and
use of animals.
2.2. Surgical groups
Prior to surgery, subjects were assigned to one of the
surgical groups. There were six groups. Five groupsreceived 48 h of LD: (1) a combined unilateral AGm and
DCS lesion group (AGm/DCS; n�/8); (2) a group with
combined AGm and control lesions of the area laterally
T.M. Van Vleet et al. / Behavioural Brain Research 138 (2003) 165�/178166

adjacent to the DCS (AGm/LSC; n�/6) which acted as a
control for extent of cortical and striatal damage; (3) a
unilateral DCS lesion group (n�/8); (4) a unilateral
AGm lesion group (AGm/LD; n�/5); and (5) a uni-lateral AGm lesion group that did not receive extinction
testing (AGm; n�/5). Prior studies have only examined
the effects of LD on neglect [2]; therefore, this AGm
group, which was taken from a prior study from our
laboratory [2], was used to control for the possible
effects of combined neglect and extinction testing on the
therapeutic effectiveness of LD. A sixth group which
received a unilateral AGm lesion was tested for bothneglect and extinction, but did not experience LD
(AGm/noLD; n�/8). This group was included in the
analysis of the extinction data only, and served as a
baseline comparison group to examine the affects of LD
on extinction. Finally, to examine the possibility that the
effective dose of LD may differ for the AGm and the
DCS, three additional unilateral DCS operates were
given 72 h of LD. A prior study has indicated that 72 hrather than 48 h of LD is necessary to consistently
produce recovery in 24-month-old AGm operates with
severe neglect [7].
2.3. Preoperative extinction testing
In order to assess for a preoperative paw preference,
subjects were given a test commonly used to examine
extinction to bilateral simultaneous stimulation[29,43,51]. The subjects, while still in their home cage,
were placed on a table in a room illuminated by dim red
light for 1 min of adaptation. After adaptation, round
adhesive-backed papers (1.3 cm diameter labels) (Avery
International) were attached bilaterally to the radial
aspect of the wrist. The distal part of the label covered
approximately 1 mm of the hairless part of the forepaw.
After both labels were attached, they were pressedsimultaneously, and the animal was replaced in its
home cage. Latency to contact and latency to remove
each label were recorded. If the animal failed to remove
one or both of the labels within 120 s of attachment, the
trial was terminated. Each animal was given five trials
per test session, with each trial separated by 1 min.
2.4. Surgical procedures
2.4.1. AGm lesions
A detailed description of the surgical procedures can
be found in Burcham et al. [2,4]. Briefly, animals were
anesthetized using sodium pentobarbital (50 mg/kg,
i.p.). Under aseptic conditions, a midline incision was
made and the skull revealed. A skull ‘‘window’’ extend-
ing from 5 mm rostral to �/2 mm caudal to bregma and2 mm lateral to the sagittal sinus was removed on the
intended side of the lesion. The AGm was then removed
via gentle subpial aspiration. Once hemostasis was
achieved, Gelfoam was placed in the wound and the
incision closed with sterile wound clips and treated with
Neomycin ointment. The subjects were kept warm and
monitored prior to behavioral testing.
2.4.2. DCS and LSC lesions
Animals were anesthetized using sodium pentobarbi-
tal (50 mg/kg, i.p.). When unresponsive (absence of
corneal reflex and responsiveness to a mild tail pinch),
the subjects were placed in a stereotaxic apparatus using
blunt-tipped ear bars. Under aseptic conditions, a small
hole was drilled in the skull at 0.5 mm rostral and 3.4mm lateral to bregma. Drugs were infused using a
programmable infusion pump (Stoelting) attached to a
0.5 ml, 26-gauge Hamilton syringe. For the DCS group,
the cannula was lowered 4.2 mm ventral to the dura. For
the LSC group, the hole was drilled at 0.5 mm rostral,
4.4 mm lateral relative to bregma, and the cannula
lowered to 5.2 mm ventral to the dura. The 0.1 ml dose
of NMDA was infused at a rate of 0.1 ml/2 min, andallowed to diffuse for an additional 4 min before the
syringe was removed. The incision was closed with
sterile wound clips, treated with neomycin ointment,
and the subjects kept warm and monitored prior to
being returned to the colony.
2.4.3. Combined lesions
In the AGm/DCS and AGm/LSC groups in which the
subjects received both surgeries in the same session, thesubjects received the appropriate striatal lesion first,
followed by subpial aspiration of the AGm as described
earlier for each of the individual procedures. The
subjects tolerated the combined procedures quite well,
and could be tested for neglect at 4 h post-injection.
2.5. Pre-LD behavioral testing
2.5.1. Circling behavior
The subject was placed in its home cage on the testing
platform, and the number of ipsi- and contralesional
turns was counted to the nearest 1/2 turn for a 2 min
period prior to behavioral testing for neglect [10,15]. A
1/2 turn was recorded when both of the subject’s hind
feet crossed over an imaginary line 908 from the starting
position.
2.5.2. Orientation testing
All testing were conducted during the light phase of
the light/dark cycle in a room with standard overhead
fluorescent lighting. Orientation testing was a modified
version of that developed by Crowne et al. [15] and was
designed to reflect simple bedside testing for neglect in
humans. These procedures have been used in a numberof studies from my laboratory [2,4,50] and others [59].
After the 2 min assessment of circling behavior, the
animal was taken out of its cage and placed directly on
T.M. Van Vleet et al. / Behavioural Brain Research 138 (2003) 165�/178 167

the test platform marked to delineate 08, 308, 458, and
608 angles in either direction from a central line running
the length of the testing board. The subject was gently
restrained by hand from behind without restricting headmovement, and aligned with the centerline. Stimuli were
presented only when there was no evidence of strug-
gling, no asymmetry of body posture, and when the
head was oriented in direct line with the body. Typically,
the animal’s body had to be realigned several times
during testing. The early extensive handling minimized
struggling and ‘‘freezing’’. Visual, tactile, and auditory
stimuli were presented in turn. The visual stimulusconsisted of the presentation of a silver metallic rod
10.0 cm in length (8 mm in width), which was waved in a
small circle (approximately 5.0 cm in diameter) five
times within the animal’s visual field at a distance of
7.5�/10.0 cm from the animal. Care was taken not to
contact the vibrissa while waving the metallic rod. The
auditory stimulus was a single 114 dB (SPL) click
generated by a clicking device held at mid-body,approximately 5 cm from the subject. The tactile
stimulus was a single caudal-to-rostral stroke through
the vibrissa with a 15 cm Puritan applicator (Harkwood
Products Co., No. 807). Although such stimulation has
a visual component, testing under red light conditions
yielded virtually identical results (Corwin, unpublished).
Three cycles of testing comprised one test session. One
cycle consisted of a single presentation of each of thethree stimuli to each body side in turn. Stimuli were
presented in the order: visual, tactile, and then auditory.
We have found that order of presentation does not
influence performance (Corwin, unpublished). The ex-
perimenter rated the degree of head turning toward or
away from (allesthetic responding) the stimuli as mea-
sured by the position of the tip of the snout over the test
platform markings. A head turn of less than 308 receiveda zero score, between 308 and 458 as 1.0, between 458and 608 as 1.5, and greater than 608 as 2.0. Orientations
later than 2 s after stimulus presentation received a zero
score. Orientations to the visual stimulus after the third
revolution (3 s) received a maximum score of 1.5. The
maximum score for each body side is 6.0 for each of the
three modalities, 18.0 in total. In prior studies, this
rating scale produced an interrator reliability of 1.0 forthe direction of orientation, and above 0.9 for the
magnitude of orientation [10]. The experimenter testing
the animals was blind with respect to the group
affiliation of the subjects.
A total neglect ratio was derived from the formula:
total contralesional responsiveness/total ipsilesional re-
sponsiveness, and collapsed across all modalities
[2,4,10]. The neglect ratio is a measure of relativeresponding to each body side and takes into account
total responsiveness; it indicates asymmetries in orienta-
tion behavior. Neglect ratios were also calculated to
compare the non-neglected to the neglected body side
for the visual, tactile, and auditory modalities. Because
neglect ratios give no indication of whether an asym-
metry in orientation results from lower contralesional
(non-neglect side), or higher ipsilesional (neglect side)scores separate analyses of raw scores for ipsi- and
contralesional responding was conducted to assess pre-
vs. post-manipulation responsiveness [57].
2.5.3. Allesthesia/allokinesia
Responses to the inappropriate side, away from the
side of stimulation (allesthetic responses), were rated
identically. All behavioral testing occurred between07:00 and 19:00 h of the light portion of the 12/12
day�/night cycle.
All LD subjects were tested for neglect 4 h after the
appropriate surgical procedures. As in prior studies, we
used animals with severe neglect: (total contralesional
responsiveness/total ipsilesional responsiveness)5/0.33,
in order to conservatively test the effects of LD [2,4,9].
The time post-injection when neglect is first assessed (4h) was chosen with two concerns in mind: (1) that as
short an interval post-surgery as possible be chosen that
would allow for behavioral assessment of neglect and (2)
severe neglect demonstrated at the first test would
predict severe neglect on subsequent tests. Both these
criteria have been met in previous studies of LD [2,4,9].
Severe neglect at 4 h post-injection has invariably
predicted severe neglect at 48 h post-surgery [2,9]. Inthe AGm and AGm/LSC groups, our criteria for severe
neglect was met by approximately 80% of all operates.
In contrast to prior studies [2,9], we have found that
DCS and AGm/DCS subjects consistently exhibit severe
chronic neglect at 48 h post-surgery regardless of the
severity of neglect during the Pre-LD test at 4 h post-
injection [51]. This may due to the possibility that the
neurotoxicity produced by NMDA requires a longertime course to produce severe neglect. In this study, only
50% of the subjects in the AGm/DCS and DCS groups
demonstrated severe neglect at 4 h post-injection.
2.6. Light deprivation
The environmental manipulation took place in a 3
m�/3 m windowless room. At any one time, one to
three animals were in the experimental room. Care wastaken to ensure that sufficient food and water was given
for the duration of the manipulation. The door was
sealed and all extraneous light was blocked. It was
typical that different groups experienced the environ-
mental manipulation together to prevent order effects.
2.7. Post-LD behavioral testing
2.7.1. Orientation testing
Following the environmental manipulation, the door
to the room was opened, the lights turned on, and the
T.M. Van Vleet et al. / Behavioural Brain Research 138 (2003) 165�/178168

subjects allowed at least 15 min of adaptation to the
lighted room. After adaptation, the subjects were tested
for neglect in a fashion identical to that described for the
orientation test at 4 h post-injection. Thereafter, allsubjects were kept in the colony under a 12:12 light/dark
cycle. The experimenter was blind with respect to the
subjects’ group affiliation. Subjects were tested twice per
week and the effects of LD on neglect were evaluated for
2 weeks.
2.7.2. Extinction testing
Extinction testing procedures were identical to those
given prior to surgery. Extinction was considered to be a
tendency to touch and/or remove the tab attached to the
ipsilesional wrist before touching and/or removing the
contralesionally applied tab during an individual trial.Therefore, analyses of data on the extinction task took
place on the following measures: (1) frequency of
touching the ipsilesional tab first, (2) frequency of
removing the ipsilesional tab first, (3) frequency of
touching the contralesional tab first, and (4) frequency
of removing the contralesional tab first. Extinction was
assessed on alternate days when the subjects were not
being tested for orientation (neglect) [29]. The experi-menter was blind with respect to the group affiliation of
the subjects.
2.8. Histological procedures
After behavioral testing was completed, the subjects
were given an overdose of sodium pentobarbital (65 mg)
and when totally unresponsive (absence of a corneal
reflex, unresponsiveness to tail pinch, and cessation of
respiration), intracardially perfused with normal saline,
followed by 10% formalin. The brain was removed from
the skull, placed in 10% formalin for at least 3 days, andthen into 30% sucrose�/formalin until the brain sank.
The brain was frozen and sectioned in the coronal plane
at 50 mm. For the AGm operates every tenth section
through the extent of the lesion and every fifth section
through the thalamus was saved, mounted, and stained
with cresyl-violet. All lesions were examined to deter-
mine the extent of damage to adjacent areas and the
AGm. The thalamus was examined for any signs ofcalcification, gliosis, or shrinkage of nuclei. For the
DCS and LSC operates every section through the extent
of the lesion was saved, mounted, and stained with
cresyl-violet. Lesion sizes were measured by tracing the
extent of lesion and areas of gliosis through an image
analysis program (Optimus, BioScan). Lesion extents
were traced onto standard brain diagrams [37]. All
brains were identified by an arbitrary number, and notassociated with a particular group. All histological
processing and analyses were performed prior to the
behavioral analyses in order to assure blindness.
3. Results
3.1. Histology
The histological analysis revealed that the unilateralAGm lesions were virtually identical to those in other
studies of neglect [2,4,10,14,26,56,57]. As indicated in
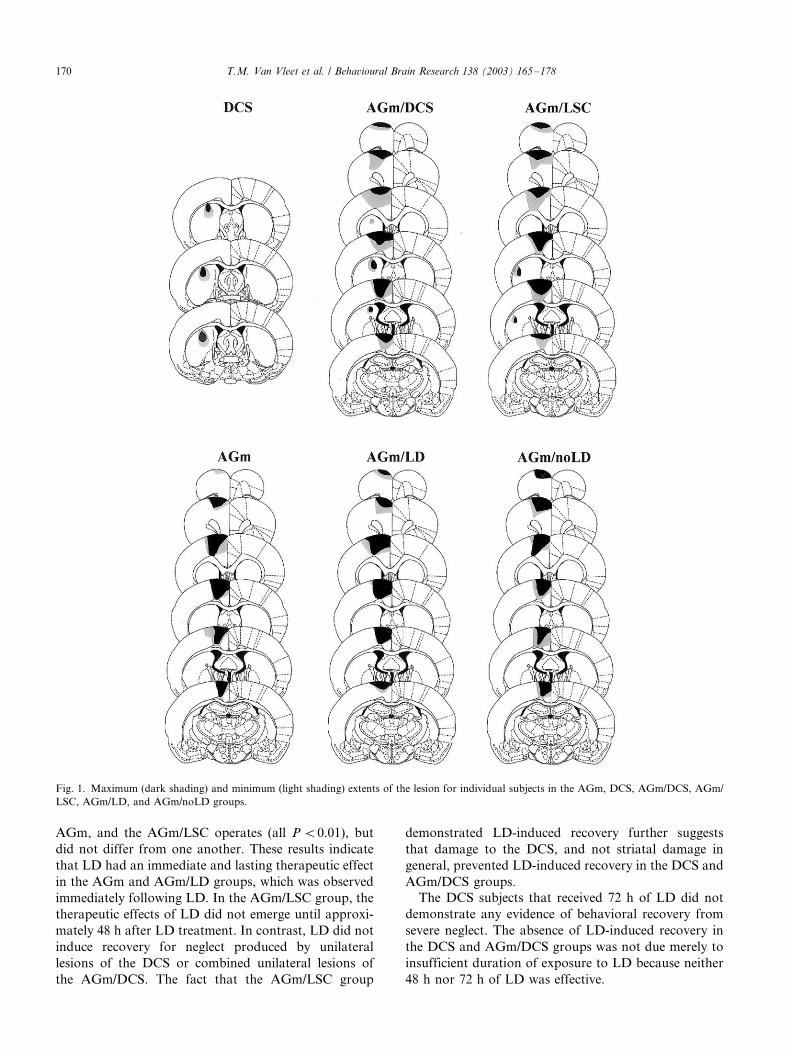
Fig. 1, in addition to the removal of AGm, typically
there was also some damage to the lateral agranular
cortex, the dorsal cingulate cortex, and the cingulum
bundle.
The size of the unilateral AGm lesions did not differ
among the groups and were virtually identical to thosein prior studies [2,4,9]. Likewise, DCS lesion size was
virtually identical across the groups and was virtually
identical to those in a prior study of the effects of DCS
lesions [50]. The DCS lesions in the AGm/DCS and the
DCS groups destroyed a significant portion of the
projection zone of the Agm; however, there was very
minor damage produced by the injection procedure
itself. The LSC lesions in the AGm/LSC group werevirtually identical in size to the DCS lesions. Image
analysis revealed no significant difference in lesion
extent between the AGm/DCS group and the AGm/
LSC group; therefore, any behavioral differences cannot
be explained by differences in lesion size.
3.2. Neglect
A total neglect ratio was derived from the formula:
contralateral total neglect/ipsilateral total neglect
[2,4,10,50]. The total neglect ratio is a measure of
relative responding to each body side and takes into
account total responsiveness. The data from the five
tests, which included a Pre-LD test (Test 1) and 2 weeks
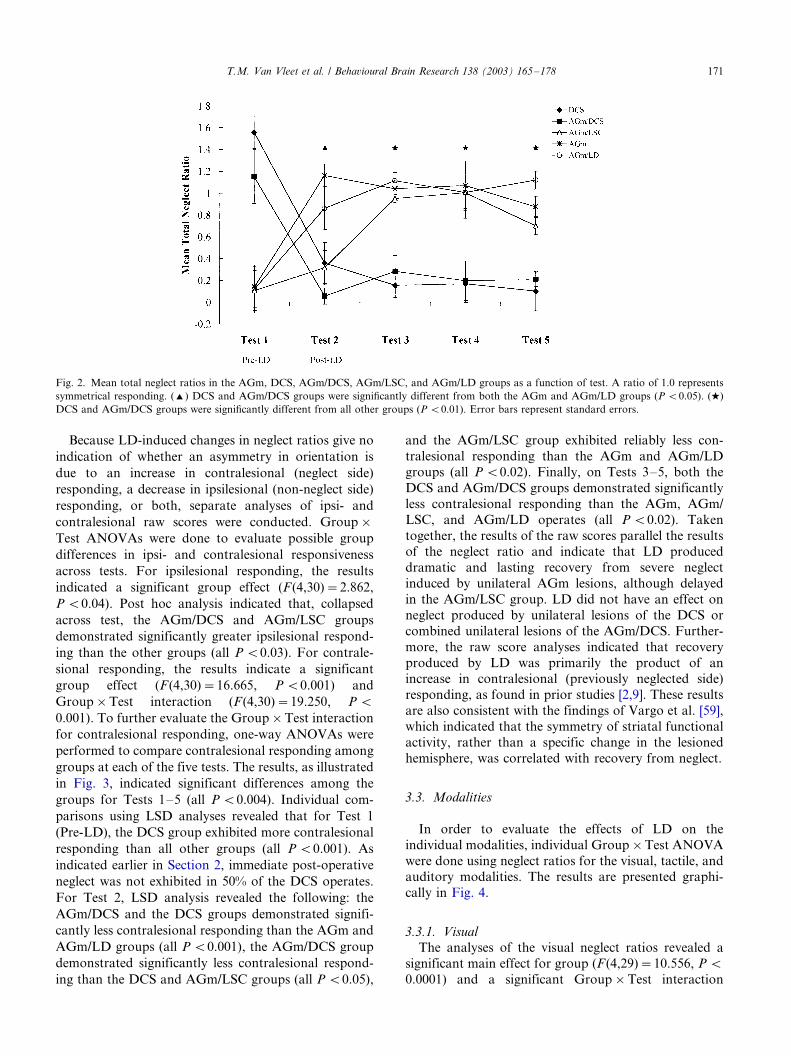
of post-operative testing (Tests 2�/5), were analyzed. A
Group�/Tests ANOVA comparing the total neglectratios for the AGm [2], DCS, AGm/LSC, AGm/DCS,
and the AGm/LD groups revealed no significant main
effects (all P �/0.05), but did indicate a significant
Test�/Group interaction (F (16,104)�/2.138, P B/
0.012). In order to explore the significant interaction,
individual one-way ANOVAs were done to compare the
groups at each of the five tests. The results, as illustrated
in Fig. 2, indicated significant differences among thegroups for Tests 2�/5 (all P B/0.001). The lack of a
significant difference on Test 1 (Pretest) reflects the
screening for severe neglect found in all but the DCS
groups. Individual comparisons using LSD analyses
revealed that on Test 2, both the DCS and AGm/DCS
groups, despite the absence of Pre-LD neglect, demon-
strated significantly more neglect than both the AGm
and AGm/LD (all P B/0.01), but did not differ fromeach other or the AGm/LSC group (all P �/0.05). On
Tests 3�/5, DCS and AGm/DCS groups demonstrated
significant neglect when compared with the AGm/LD,
T.M. Van Vleet et al. / Behavioural Brain Research 138 (2003) 165�/178 169
AGm, and the AGm/LSC operates (all P B/0.01), but
did not differ from one another. These results indicate
that LD had an immediate and lasting therapeutic effect
in the AGm and AGm/LD groups, which was observed
immediately following LD. In the AGm/LSC group, the
therapeutic effects of LD did not emerge until approxi-
mately 48 h after LD treatment. In contrast, LD did not
induce recovery for neglect produced by unilateral
lesions of the DCS or combined unilateral lesions of
the AGm/DCS. The fact that the AGm/LSC group
demonstrated LD-induced recovery further suggests
that damage to the DCS, and not striatal damage in
general, prevented LD-induced recovery in the DCS and
AGm/DCS groups.
The DCS subjects that received 72 h of LD did not
demonstrate any evidence of behavioral recovery from
severe neglect. The absence of LD-induced recovery in
the DCS and AGm/DCS groups was not due merely to
insufficient duration of exposure to LD because neither
48 h nor 72 h of LD was effective.
Fig. 1. Maximum (dark shading) and minimum (light shading) extents of the lesion for individual subjects in the AGm, DCS, AGm/DCS, AGm/
LSC, AGm/LD, and AGm/noLD groups.
T.M. Van Vleet et al. / Behavioural Brain Research 138 (2003) 165�/178170
Because LD-induced changes in neglect ratios give no
indication of whether an asymmetry in orientation is
due to an increase in contralesional (neglect side)
responding, a decrease in ipsilesional (non-neglect side)
responding, or both, separate analyses of ipsi- and
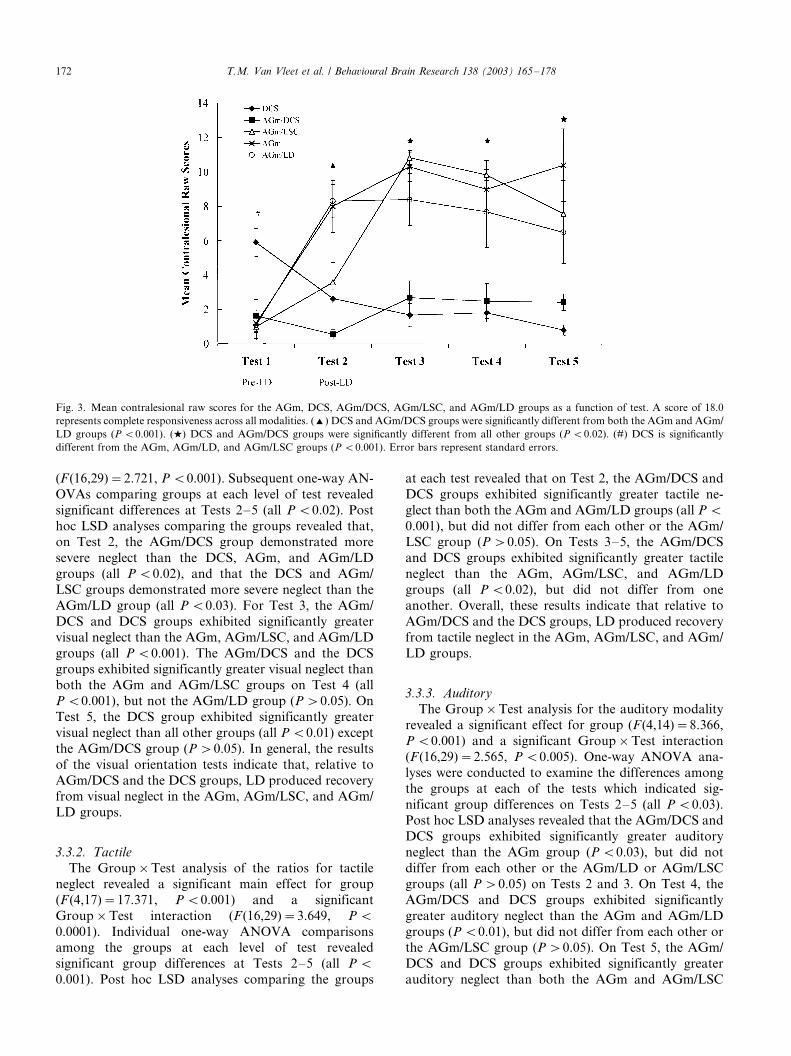
contralesional raw scores were conducted. Group�/
Test ANOVAs were done to evaluate possible group
differences in ipsi- and contralesional responsiveness
across tests. For ipsilesional responding, the results
indicated a significant group effect (F (4,30)�/2.862,
P B/0.04). Post hoc analysis indicated that, collapsed
across test, the AGm/DCS and AGm/LSC groups
demonstrated significantly greater ipsilesional respond-
ing than the other groups (all P B/0.03). For contrale-
sional responding, the results indicate a significant
group effect (F (4,30)�/16.665, P B/0.001) and
Group�/Test interaction (F (4,30)�/19.250, P B/
0.001). To further evaluate the Group�/Test interaction
for contralesional responding, one-way ANOVAs were
performed to compare contralesional responding among
groups at each of the five tests. The results, as illustrated
in Fig. 3, indicated significant differences among the
groups for Tests 1�/5 (all P B/0.004). Individual com-
parisons using LSD analyses revealed that for Test 1
(Pre-LD), the DCS group exhibited more contralesional
responding than all other groups (all P B/0.001). As
indicated earlier in Section 2, immediate post-operative
neglect was not exhibited in 50% of the DCS operates.
For Test 2, LSD analysis revealed the following: the
AGm/DCS and the DCS groups demonstrated signifi-
cantly less contralesional responding than the AGm and
AGm/LD groups (all P B/0.001), the AGm/DCS group
demonstrated significantly less contralesional respond-
ing than the DCS and AGm/LSC groups (all P B/0.05),
and the AGm/LSC group exhibited reliably less con-
tralesional responding than the AGm and AGm/LD
groups (all P B/0.02). Finally, on Tests 3�/5, both the
DCS and AGm/DCS groups demonstrated significantly
less contralesional responding than the AGm, AGm/
LSC, and AGm/LD operates (all P B/0.02). Taken
together, the results of the raw scores parallel the resultsof the neglect ratio and indicate that LD produced
dramatic and lasting recovery from severe neglect
induced by unilateral AGm lesions, although delayed
in the AGm/LSC group. LD did not have an effect on
neglect produced by unilateral lesions of the DCS or
combined unilateral lesions of the AGm/DCS. Further-
more, the raw score analyses indicated that recovery
produced by LD was primarily the product of anincrease in contralesional (previously neglected side)
responding, as found in prior studies [2,9]. These results
are also consistent with the findings of Vargo et al. [59],
which indicated that the symmetry of striatal functional
activity, rather than a specific change in the lesioned
hemisphere, was correlated with recovery from neglect.
3.3. Modalities
In order to evaluate the effects of LD on the
individual modalities, individual Group�/Test ANOVA
were done using neglect ratios for the visual, tactile, andauditory modalities. The results are presented graphi-
cally in Fig. 4.
3.3.1. Visual
The analyses of the visual neglect ratios revealed a
significant main effect for group (F (4,29)�/10.556, P B/
0.0001) and a significant Group�/Test interaction
Fig. 2. Mean total neglect ratios in the AGm, DCS, AGm/DCS, AGm/LSC, and AGm/LD groups as a function of test. A ratio of 1.0 represents
symmetrical responding. (') DCS and AGm/DCS groups were significantly different from both the AGm and AGm/LD groups (P B/0.05). (w)
DCS and AGm/DCS groups were significantly different from all other groups (P B/0.01). Error bars represent standard errors.
T.M. Van Vleet et al. / Behavioural Brain Research 138 (2003) 165�/178 171
(F (16,29)�/2.721, P B/0.001). Subsequent one-way AN-
OVAs comparing groups at each level of test revealed
significant differences at Tests 2�/5 (all P B/0.02). Post
hoc LSD analyses comparing the groups revealed that,on Test 2, the AGm/DCS group demonstrated more
severe neglect than the DCS, AGm, and AGm/LD
groups (all P B/0.02), and that the DCS and AGm/
LSC groups demonstrated more severe neglect than the
AGm/LD group (all P B/0.03). For Test 3, the AGm/
DCS and DCS groups exhibited significantly greater
visual neglect than the AGm, AGm/LSC, and AGm/LD
groups (all P B/0.001). The AGm/DCS and the DCSgroups exhibited significantly greater visual neglect than
both the AGm and AGm/LSC groups on Test 4 (all
P B/0.001), but not the AGm/LD group (P �/0.05). On
Test 5, the DCS group exhibited significantly greater
visual neglect than all other groups (all P B/0.01) except
the AGm/DCS group (P �/0.05). In general, the results
of the visual orientation tests indicate that, relative to
AGm/DCS and the DCS groups, LD produced recoveryfrom visual neglect in the AGm, AGm/LSC, and AGm/
LD groups.
3.3.2. Tactile
The Group�/Test analysis of the ratios for tactile
neglect revealed a significant main effect for group
(F (4,17)�/17.371, P B/0.001) and a significant
Group�/Test interaction (F (16,29)�/3.649, P B/
0.0001). Individual one-way ANOVA comparisonsamong the groups at each level of test revealed
significant group differences at Tests 2�/5 (all P B/
0.001). Post hoc LSD analyses comparing the groups
at each test revealed that on Test 2, the AGm/DCS and
DCS groups exhibited significantly greater tactile ne-
glect than both the AGm and AGm/LD groups (all P B/
0.001), but did not differ from each other or the AGm/LSC group (P �/0.05). On Tests 3�/5, the AGm/DCS
and DCS groups exhibited significantly greater tactile
neglect than the AGm, AGm/LSC, and AGm/LD
groups (all P B/0.02), but did not differ from one
another. Overall, these results indicate that relative to
AGm/DCS and the DCS groups, LD produced recovery
from tactile neglect in the AGm, AGm/LSC, and AGm/
LD groups.
3.3.3. Auditory
The Group�/Test analysis for the auditory modality
revealed a significant effect for group (F (4,14)�/8.366,
P B/0.001) and a significant Group�/Test interaction
(F (16,29)�/2.565, P B/0.005). One-way ANOVA ana-
lyses were conducted to examine the differences among
the groups at each of the tests which indicated sig-
nificant group differences on Tests 2�/5 (all P B/0.03).Post hoc LSD analyses revealed that the AGm/DCS and
DCS groups exhibited significantly greater auditory
neglect than the AGm group (P B/0.03), but did not
differ from each other or the AGm/LD or AGm/LSC
groups (all P �/0.05) on Tests 2 and 3. On Test 4, the
AGm/DCS and DCS groups exhibited significantly
greater auditory neglect than the AGm and AGm/LD
groups (P B/0.01), but did not differ from each other orthe AGm/LSC group (P �/0.05). On Test 5, the AGm/
DCS and DCS groups exhibited significantly greater
auditory neglect than both the AGm and AGm/LSC
Fig. 3. Mean contralesional raw scores for the AGm, DCS, AGm/DCS, AGm/LSC, and AGm/LD groups as a function of test. A score of 18.0
represents complete responsiveness across all modalities. (') DCS and AGm/DCS groups were significantly different from both the AGm and AGm/
LD groups (P B/0.001). (w) DCS and AGm/DCS groups were significantly different from all other groups (P B/0.02). (#) DCS is significantly
different from the AGm, AGm/LD, and AGm/LSC groups (P B/0.001). Error bars represent standard errors.
T.M. Van Vleet et al. / Behavioural Brain Research 138 (2003) 165�/178172
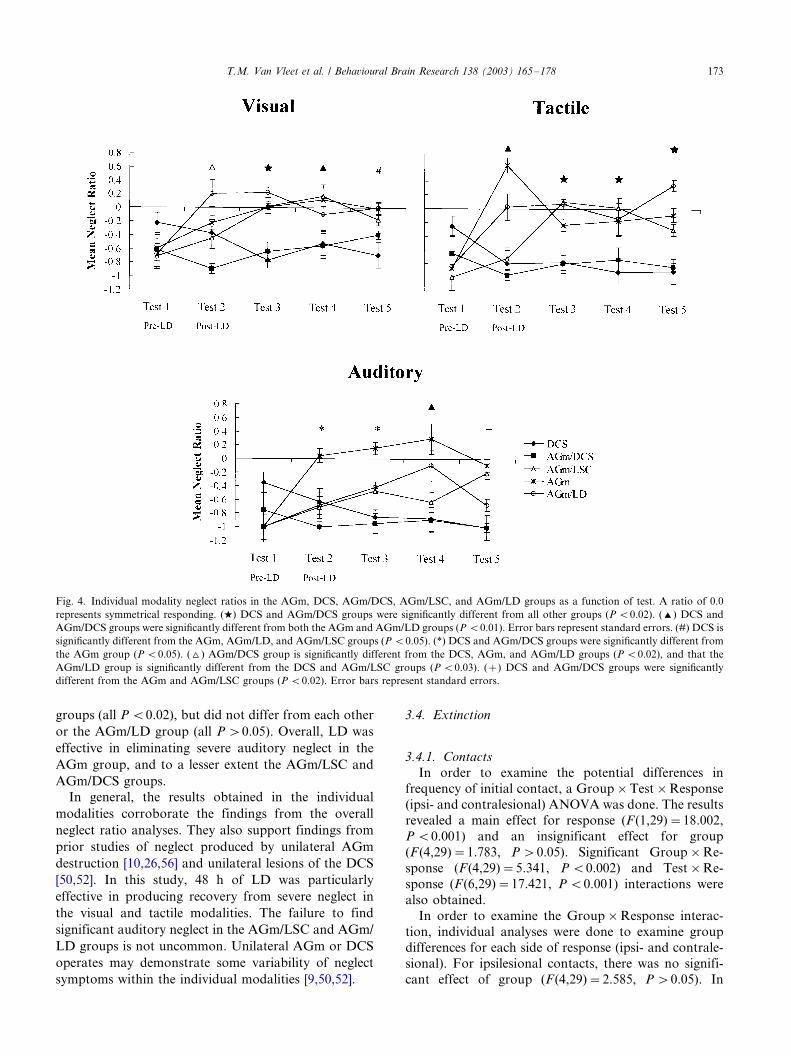
groups (all P B/0.02), but did not differ from each other
or the AGm/LD group (all P �/0.05). Overall, LD was
effective in eliminating severe auditory neglect in the
AGm group, and to a lesser extent the AGm/LSC and
AGm/DCS groups.
In general, the results obtained in the individual
modalities corroborate the findings from the overall
neglect ratio analyses. They also support findings from
prior studies of neglect produced by unilateral AGm
destruction [10,26,56] and unilateral lesions of the DCS
[50,52]. In this study, 48 h of LD was particularly
effective in producing recovery from severe neglect in
the visual and tactile modalities. The failure to find
significant auditory neglect in the AGm/LSC and AGm/
LD groups is not uncommon. Unilateral AGm or DCS
operates may demonstrate some variability of neglect
symptoms within the individual modalities [9,50,52].
3.4. Extinction
3.4.1. Contacts
In order to examine the potential differences in
frequency of initial contact, a Group�/Test�/Response
(ipsi- and contralesional) ANOVA was done. The resultsrevealed a main effect for response (F (1,29)�/18.002,
P B/0.001) and an insignificant effect for group
(F (4,29)�/1.783, P �/0.05). Significant Group�/Re-
sponse (F (4,29)�/5.341, P B/0.002) and Test�/Re-
sponse (F (6,29)�/17.421, P B/0.001) interactions were
also obtained.
In order to examine the Group�/Response interac-
tion, individual analyses were done to examine groupdifferences for each side of response (ipsi- and contrale-
sional). For ipsilesional contacts, there was no signifi-
cant effect of group (F (4,29)�/2.585, P �/0.05). In
Fig. 4. Individual modality neglect ratios in the AGm, DCS, AGm/DCS, AGm/LSC, and AGm/LD groups as a function of test. A ratio of 0.0
represents symmetrical responding. (w) DCS and AGm/DCS groups were significantly different from all other groups (P B/0.02). (') DCS and
AGm/DCS groups were significantly different from both the AGm and AGm/LD groups (P B/0.01). Error bars represent standard errors. (#) DCS is
significantly different from the AGm, AGm/LD, and AGm/LSC groups (P B/0.05). (*) DCS and AGm/DCS groups were significantly different from
the AGm group (P B/0.05). (^) AGm/DCS group is significantly different from the DCS, AGm, and AGm/LD groups (P B/0.02), and that the
AGm/LD group is significantly different from the DCS and AGm/LSC groups (P B/0.03). (�/) DCS and AGm/DCS groups were significantly
different from the AGm and AGm/LSC groups (P B/0.02). Error bars represent standard errors.
T.M. Van Vleet et al. / Behavioural Brain Research 138 (2003) 165�/178 173

contrast, a significant group effect was obtained for
contralesional contacts (F(4,29)�/7.162, P B/0.001).
Pairwise comparisons revealed that the frequency of
contacting the contralesional tab first was significantly
greater in the DCS operates when compared with all
other groups (all P B/0.007), reflecting the absence of an
extinction deficit. In addition, it was also found that the
AGm/LSC group exhibited significantly fewer contrale-
sional contacts than the AGm/noLD group (P B/0.04).
No other differences were detected among the AGm/
noLD, AGm/DCS, AGm/LSC, and AGm/LD groups
(all P �/0.05).
To evaluate the Test�/Response interaction, separate
comparisons were done for ipsi- and contralesional
contacts collapsed across group. The results indicated
that both ipsilesional (F (6,24)�/6.378, P B/0.017) and
contralesional (F (6,24)�/16.701, P B/0.0001) initial
contacts changed significantly over tests. Pairwise com-
parisons revealed the frequency of contacting the
contralesional tab was greater for Test 1 (Pretest) than
Tests 2�/7 (all P B/0.0001). For ipsilesional contacts, the
frequency on Test 1 (Pretest) was significantly smaller
than Tests 2�/7 (all P B/0.0001).
Analyses of the Test�/Response interaction indicated
that when groups are analyzed together, all operates
displayed a significant increase in ipsilesional contacts
and significant decrease in contralesional contacts after
surgery. However, the Group�/Response interaction
revealed that the DCS operates, unlike the other groups,
did not change their preoperative contralesional paw
preference and thus displayed significantly more con-
tacts to the contralesional side than the other groups.
Taken together, the results suggest that all groups,
except the DCS, exhibited a post-surgical change in
contact paw preference, indicative of an extinction
deficit.
Fig. 5. (A) Mean frequency of contralesional contacts in the AGm, DCS, AGm/DCS, AGm/LSC, AGm/LD, and AGm/noLD groups as a function
of test. The maximum number of potential contacts is 5.0. With the exception of the DCS group, all groups demonstrated a significant and lasting
reduction in the frequency of contralesional contacts. Error bars represent standard errors. (B) Mean frequency of contralesional removals in the
AGm, DCS, AGm/DCS, AGm/LSC, AGm/LD, and AGm/noLD groups as a function of test. The maximum number of potential removals is 5.0.
With the exception of the DCS and AGm/DCS groups, all groups demonstrated a significant and lasting reduction in the frequency of contralesional
removals. Error bars represent standard errors.
T.M. Van Vleet et al. / Behavioural Brain Research 138 (2003) 165�/178174

In order to determine whether individual groups
displayed a change in paw preference for contact, a
series of within-subject analyses were done which
separately examined contra- and ipsilesional frequencyof contact for each group across tests. With regard to
ipsilesional tab contacts, the AGm/noLD, AGm/LD,
AGm/DCS, and AGm/LSC groups demonstrated a
significant test effect (all P B/0.02). In these groups,
paired sample t-tests were done to compare Test 1
(Pretest) to subsequent Post-LD tests. For the AGm/
noLD group, the results indicated that Test 1 differed
significantly from Tests 2, 3, 4, and 6 (all P B/0.03); anindication that ipsilesional tab contact increased follow-
ing the AGm lesion. In a similar fashion, the AGm/DCS
group significantly increased ipsilesional contacts on
Tests 2 and 3, as did the AGm/LSC group on Test 3 (all
P B/0.01). For the DCS group, the analysis revealed no
significant change in ipsilesional contacts.
The analysis for the contralesional contacts revealed a
significant test effect for the AGm/noLD, AGm/LD,AGm/DCS, and AGm/LSC groups (all P B/0.008). As
seen in Fig. 5A, the analyses comparing Test 1 with the
subsequent tests showed that the AGm/noLD, AGm/
LD, AGm/DCS, and AGm/LSC lesions produced a
significant decrease in contacts to the contralesional
preoperatively preferred paw on Tests 2�/7 (all P B/
0.04). Taken together, the within-subject analyses in-
dicated that all groups except the DCS group demon-strated extinction post-surgically, as evidenced by a
significant increase in ipsilesional contacts and a sig-
nificant decrease in contralesional contacts. Further,
results from this study support recent findings from our
lab which demonstrate that DCS lesions alone do not
produce extinction [52].
3.4.2. Removals
A Group�/Test�/Response (ipsi- and contralesional)ANOVA was done to examine the pattern of tab
removals. The results indicated that there were no
significant main effects (all P �/0.23), but there was a
significant Test�/Response (F (6,150)�/8.372, P B/
0.0001) interaction. The Response�/Group interaction,
although not significant, was noteworthy (F (4,25)�/
2.686, P B/0.054).
In order to evaluate the Test�/Response (ipsi- andcontralesional) interaction, individual comparisons were
done for ipsi- and contralesional removals collapsed
across group. The results indicated that both ipsilesional
(F (6,24)�/10.344, P B/0.0001) and contralesional
(F (6,24)�/8.996, P B/0.0001) removals changed signifi-
cantly over the duration of testing. Follow-up compar-
isons revealed that when, collapsed across all groups,
Test 1 differed significantly from Tests 2�/7 for bothipsilesional (all P B/0.005) and contralesional (all P B/
0.008) removals. These results suggest that, when
collapsed across all groups, there was a significant
increase in ipsilesional removal and a significant de-
crease in contralesional removal after surgery.
In order to compare more directly ipsi- and contrale-
sional removals, additional within-subject analyses weredone to separately examine contra- and ipsilesional
responses for each group across the tests. With regard
to ipsilesional tab removals, only the AGm/DCS and
AGm/LD groups demonstrated a significant test effect
(all P B/0.049). Paired sample t-tests comparing Test 1
(Pretest) to subsequent post-surgical tests indicated that
the number of Pretest ipsilesional tab removals in both
the AGm/LD and AGm/DCS groups significantlyincreased on post-surgical Tests 2, 3, 4, 6, and 7 (all
P B/0.04).
With regard to the contralesional tab removals, the
AGm/noLD, AGm/LD, and AGm/DCS groups demon-
strated a significant test effect (P B/0.002). Paired
sample t -tests comparing Test 1 (Pretest) to subsequent
post-surgical tests indicated that the number of con-
tralesional tab removals in the AGm/noLD, AGm/LD,and AGm/DCS lesion groups decreased post-surgically
(all P B/0.04). Overall, the removal within-subject
analyses indicated that the AGm/DCS, AGm/LD, and
AGm/noLD groups exhibited significant extinction, as
evidenced by a significant change in paw preference
post-surgery. The AGm/DCS and AGm/LD groups
demonstrated a significant increase in ipsilesional re-
movals and a significant decrease in contralesionalremovals post-surgically. The AGm/noLD groups also
demonstrated a significant decrease in contralesional
responding post-surgery (see Fig. 5B). The results also
corroborate the findings from the contact analyses. The
failure of the AGm/LSC group to demonstrate any
significant change in ipsi- or contralesional removals is
likely due to a somewhat lower pretest removal
frequency in this group, as illustrated in Fig. 5B.
3.5. Allesthesia/allokinesia
Responses to the inappropriate side, away from the
side of stimulation (allesthetic/allokinetic responses),
were rated identically to appropriate orientations.
Because of the large number of zero scores, the total
number of allesthetic responses for the ipsi- and
contralesional sides were compared across the initialfive tests for each of the five groups using a Wilcoxon
Signed-Ranks test. There were no significant differences
between ipsi- and contralesional allesthesia/allokinesia
responding in any of the groups (all P �/0.05).
3.6. Circling
Wilcoxon Signed-Ranks tests were conducted tocompare the amount of ipsilesional vs. contralesional
circling in the five groups. The analyses of circling
behavior revealed no significant differences between
T.M. Van Vleet et al. / Behavioural Brain Research 138 (2003) 165�/178 175

ipsi- and contralesional circling within any of the groups
(all P �/0.05). Thus, any neglect or extinction deficits
cannot be explained by a circling bias or postural
asymmetries [44].
4. Discussion
Crowne et al. [14,15] found that animals that experi-
enced eye closure or 48 h of LD did not demonstrate
neglect following unilateral AGm lesions. In both
studies, LD was effective in producing virtually com-
plete sparing from AGm-induced neglect. Further, theywere able to demonstrate that the sparing of function
was due to the therapeutic effects of LD and not the
result of auditory or activity changes in the colony [15].
Similarly, LD has also been found to produce sparing
and recovery of function from sensory motor deficits
produced by lesions of the lateral hypothalamus [21,42].
However, LD was not effective in producing sparing of
neglect induced by unilateral lesions of the superiorcolliculus [15]. Corwin and Vargo [9] extended the
findings of Crowne et al. on sparing of function
[14,15] by demonstrating that LD can produce dramatic
recovery from severe neglect if administered within 4 h.
Most recently, Vargo et al. [59] have suggested that the
crucial site for LD-induced recovery may be the
dorsolateral striatum, by demonstrating that recovery
was correlated with alterations in functional activity inthis region.
The results of this study extend these findings and
indicate that the striatal projection zone of the AGm,
the DCS, may be a crucial site within the dorsolateral
striatum for the mechanisms of LD-induced recovery
from neglect induced by unilateral AGm destruction.
Combined unilateral destruction of the AGm/DCS
prohibited recovery, whereas combined unilateralAGm/LSC lesions equivalent in size, but outside of the
projection zone of AGm did not prohibit the therapeutic
effects of LD on severe neglect. These findings support
prior studies which indicate that the DCS may play a
critical role in recovery from neglect induced by
unilateral AGm lesions. In studies examining sponta-
neous recovery, AGm-induced neglect and recovery
were correlated with changes in immediate gene expres-sion in the dorsolateral striatum [54]. In a subsequent
study, Vargo and Marshall [55] found that changes in
NMDA and kainate receptors in this same region were
correlated with spontaneous recovery from AGm-in-
duced neglect. Recent observations also support the role
of the DCS in acute drug-induced recovery from neglect.
Systemic administration of apomorphine has been
found to produce acute recovery from AGm lesion-induced neglect [10,26]. The likely site of action for the
therapeutic effects of apomorphine is the DCS [49].
Direct infusion of apomorphine into the DCS in rats
with severe neglect produced by AGm lesions produced
virtually the same therapeutic effects as systemic injec-
tions [49]. Infusion into the more lateral striatum did not
produce a therapeutic effect [49]. Further, it has beendemonstrated that apomorphine is ineffective in produ-
cing recovery from severe neglect in unilateral DCS
operates [50]. These findings on the role of the DCS in
spontaneous and drug-induced recovery, when taken
together with the present results, indicate that the
integrity of the DCS may be necessary for recovery
from AGm-induced neglect.
Recent anatomical findings in rats indicate that theDCS is a convergence zone for projections from cortical
association areas including the AGm, PPC, and the
ventrolateral orbital cortex [5,39]. Damage to any one of
these interconnected cortical regions produces neglect
and deficits in spatial processing [2,4,8,11,14,15,26�/29].
In addition, the DCS has been implicated as a multi-
modal convergence region of striatum [35]. Previous
work in primates has also suggested that interconnectedcortical areas, that have been implicated in spatial and
attention behavior, are likely to have converging corti-
costriatal projections [36,46]. The importance of corti-
costriatal relationships in rats has also been implicated
in attentional deficits following bilateral lesions of
striatum [41], or via disconnection studies [6].
A second major finding of this study is that the
therapeutic effects of LD do not extend to extinction. Inthe same subjects that demonstrated recovery from
neglect as a result of exposure to LD, there was no
evidence that the effects generalized to extinction. For
example, the AGm and AGm/LSC groups demonstrated
dramatic LD-induced multimodal recovery from severe
neglect by the second test after LD, but a severe
extinction deficit was unaffected. These findings
strongly suggest that deficits associated with the neglectsyndrome likely have distinct or dissociable anatomical
and pharmacological substrates [47,48,52], and that the
eventual treatment of these components of the neglect
syndrome will be quite complex [25]. Recent pilot data
from our laboratory supports this contention by de-
monstrating that apomorphine produces a therapeutic
effect on neglect but not extinction in rats with
unilateral AGm lesions (Pyter et al., unpublished).These findings provide support for the dissociation of
neglect symptoms as found in an earlier study, which
compared the effects of rostral and caudal AGm lesions.
Unilateral destruction of the rostral AGm produced
significant allesthesia/allokinesia without severe neglect,
whereas caudal AGm lesions produced severe neglect
without significant allesthesia [26]. Taken together, these
studies suggest that the rat model may be useful fordetermining the neural substrates, which underlie the
specific deficits that comprise the neglect syndrome.
The present findings have potential relevance for the
treatment of neglect. Behavioral treatments rarely gen-
T.M. Van Vleet et al. / Behavioural Brain Research 138 (2003) 165�/178176

eralize outside of the therapeutic context or across tasks
within the same therapeutic context [13,40]. Based on
findings from rodent models of neglect, drugs have been
administered to patients, but only those with chronicneglect and stable behavioral baselines because of the
concern that drug effects may interfere with ongoing
recovery [17,19,24]. Fleet et al. [17] and Hurford et al.
[24] examined the effects of bromocriptine, a D2
receptor agonist, in patients with chronic neglect.
Bromocriptine produced recovery across a range of
measures, and when treatment was terminated, neglect
worsened. Recently, Geminiani et al. [19] found thatapomorphine can also produce acute recovery from
neglect in humans. While some hope is raised by these
studies, the specific mechanism or site of action to
account for these effects was unknown.
The evidence discussed above strongly suggests that
the DCS plays a crucial role in neglect induced by
cortical lesions, and that the DCS may be the critical site
for the mechanisms leading to spontaneous recovery[50,54,55], the therapeutic effects of DA receptor
agonists [49], and LD-induced recovery [59] in rodents.
The results of these studies further our understanding of
the role of DCS as a crucial site in a cortical�/subcortical
network which mediates directed attention, and within
which the dynamic changes that lead to recovery from
neglect may occur. Further, these results may help to
provide a rational basis for the therapeutic administra-tion of drugs or environmental manipulations in neglect
patients. The finding that LD does not have a ther-
apeutic effect on extinction points to the need for a more
detailed understanding of the neural substrates, which
underlie extinction. This is particularly compelling
because many patients that recover from neglect still
exhibit extinction deficits [25].
Acknowledgements
The authors wish to express their appreciation to
Kevin Harris for help with the histological processing
and behavioral testing. This work was supported by the
NIMH grant MH60399.
References
[1] Barth TM, Jones TA, Schallert T. Functional subdivisions of the
rat somatic sensorimotor cortex. Behav Brain Res 1990;39(1):73�/
95.
[2] Burcham KJ, Corwin JV. Effects of delay and duration of light
deprivation on recovery of function from neglect induced by
unilateral medial agranular prefrontal cortex lesions in rats.
Psychobiology 1998;263:216�/60.
[3] Burcham KJ, Corwin JV, Stoll ML, Reep RL. Disconnection of
medial agranular and posterior parietal cortex produces multi-
modal neglect in rats. Behav Brain Res 1997;86:41�/7.
[4] Burcham KJ, Corwin JV, Van Vleet TM. Light deprivation
produces behavioral recovery of function from multimodal
neglect following unilateral posterial parietal cortex lesions in
rats. Behav Brain Res 1998;90:187�/97.
[5] Cheatwood JL, Reep RL, Corwin JV. The associative striatum:
cortical and thalamic projections to the dorsocentral striatum in
rats, Brain Research, submitted for publication.
[6] Christakou A, Robbins TW, Everitt BJ. Functional disconnection
of a prefrontal cortical�/dorsal striatal system disrupts choice
reaction time performance: implications for attentional function.
Behav Neurosci 2001;115:812�/25.
[7] Corwin JV, Burcham KJ. Light deprivation and apomorphine
produce recovery from neglect induced by medial granular cortex
lesions in aged rats. Neurosci Abstr 1997;23:230.
[8] Corwin JV, Reep RL. Rodent posterior parietal cortex as a
component of a cortical network mediating directed spatial
attention. Psychobiology 1998;26:87�/102.
[9] Corwin JV, Vargo JM. Light deprivation produces accelerated
behavioral recovery of function from neglect produced by
unilateral medial agranular prefrontal cortex lesions in rats.
Behav Brain Res 1993;56:187�/96.
[10] Corwin JV, Kanter S, Watson RT, Heilman KM, Valenstein E,
Hashimoto A. Apomorphine has a therapeutic effect on neglect
produced by unilateral dorsomedial prefrontal cortex lesions in
rats. Exp Neurol 1986;94:683�/9.
[11] Corwin JV, Fussinger M, Meyer RC, King VR, Reep RL.
Bilateral destruction of the ventrolateral orbital cortex produces
allocentric but not egocentric spatial deficits in rats. Behav Brain
Res 1994;61:79�/86.
[12] Corwin JV, Burcham KJ, Hix GI. Apomorphine produces acute
dose-dependent therapeutic affects on neglect produced by
unilateral destruction of the posterior parietal cortex in rats.
Behav Brain Res 1996;79:41�/9.
[13] Couvier W, Bua B, Blanton P, Urey J. Behavioral changes
following visual scanning training: observation of five cases. Int
J Clin Neuropsychol 1987;9:74�/80.
[14] Crowne DP, Pathria MN. Some attentional effects of unilateral
frontal lesions in the rat. Behav Brain Res 1982;6:25�/39.
[15] Crowne DP, Richardson CM, Ward G. Brief deprivation of vision
after unilateral lesions of the frontal eye field prevents contral-
ateral inattention. Science 1983;220:527�/30.
[16] Denes G, Semenza C, Stoppa E, Lis A. Unilateral spatial neglect
and recovery from hemiplegia: a follow-up study. Brain
1982;105:543�/52.
[17] Fleet WS, Valenstein E, Watson RT, Heilman KM. Dopamine
agonist therapy for neglect in humans. Neurology 1987;37:1765�/
70.
[18] Fullerton KJ, Mackenzie G, Stout RW. Prognostic indices of
stroke. Quart J Med 1988;66250:147�/62.
[19] Geminiani G, Bottini G, Sterzi R. Dopamine stimulation in
unilateral neglect. J Neurosurg Psychiatr 1998;65:345�/7.
[20] Halligan PW, Marshall JC. Current issues in spatial neglect: an
editorial introduction. Neuropsychol Rehabil 1994;4:103�/10.
[21] Harrell LE, Balagura S. The effects of dark and light on the
functional recovery following lateral hypothalamic lesions. Life
Sci 1974;15:2079�/88.
[22] Heilman KM, Watson RT, Valenstein E. Neglect and related
disorders. In: Heilman KM, Valenstein E, editors. Clinical
Neuropsychology, 3rd ed.. New York: Oxford University Press,
1993:279�/336.
[23] Henley S, Pettit P, Todd-Pokropek L, Tupper L. Who goes home?
Predictive factors in stroke recovery. J Neurol Neurosurg
Psychiatr 1985;48:1�/6.
[24] Hurford P, Stringer AY, Jann B. Neuropharmacologic treatment
of hemineglect: a case report comparing bromocriptine and
methylphenidate. Arch Phys Med Rehabil 1998;79:346�/9.
T.M. Van Vleet et al. / Behavioural Brain Research 138 (2003) 165�/178 177

[25] Kerkoff G. Spatial hemineglect in humans. Prog Neurobiol
2001;631:1�/27.
[26] King V, Corwin JV. Neglect following unilateral ablation of the
caudal but not the rostral portion of the medial agranular cortex
of the rat. Behav Brain Res 1990;37:169�/84.
[27] King V, Corwin JV. Spatial deficits and hemispheric asymmetries
in the rat following unilateral and bilateral lesions of the posterior
parietal or medial agranular cortices. Behav Brain Res
1992;50:53�/68.
[28] King V, Corwin JV. Comparisons of hemiinattention produced by
unilateral lesions of the posterior parietal or the medial agranular
prefrontal cortex in the rat. Behav Brain Res 1993;54:117�/31.
[29] King V, Corwin JV, Reep RL. Production and characterization of
neglect in rats with unilateral lesions of ventrolateral orbital
cortex. Exp Neurol 1989;105:289�/99.
[30] Kozlowski MR, Marshall JF. Plasticity of [14C]2-deoxyglucose
incorporation into neostriatum and related structures in response
to dopamine neuron damage and apomorphine replacement.
Brain Res 1980;197:167�/83.
[31] Kozlowski MR, Marshall JF. Plasticity of neostriatal metabolic
activity and behavioral recovery from nigrostriatal injury. Exp
Neurol 1981;74:318�/23.
[32] Kozlowski MR, Marshall JF. Recovery of function and basal
ganglia [14C]2-deoxyglucose uptake after nigrostriatal injury.
Brain Res 1983;259:237�/48.
[33] Marshall JF. Somatosensory inattention after dopamine-deplet-
ing intracerebral 6-OHDA injections: spontaneous recovery and
pharmacological control. Brain Res 1979;177:311�/24.
[34] Marshall JF, Gotthelf T. Sensory inattention in rats with 6-
hydroxdopamine-induced degeneration of ascending dopaminer-
gic neurons: apomorphine-induced reversal of deficits. Exp
Neurol 1979;65:398�/411.
[35] McGeorge AJ, Faull RLM. The organization of the projection
from the cerebral cortex to the striatum in the rat. Neuroscience
1989;29:503�/37.
[36] Parent A. New frontiers in basal ganglia research. Trends
Neurosci 1990;13:241�/4.
[37] Paxinos G, Watson C. The Rat Brain in Stereotaxic Coordinates,
4th ed.. New York: Academic Press, 1998.
[38] Reep RL, Corwin JV. Topographic organization of the striatal
and thalamic connections of rat medial agranular cortex. Brain
Res 1999;841:43�/52.
[39] Reep RL, Cheatwood JL, Corwin JV. Neural substrates of
directed attention in rats: dorsocentral striatum and its cortical
and thalamic connections. Neurosci Abstr 2001;27:31311.
[40] Robertson IH, Halligan PW, Marshall JC. Prospects for the
rehabilitation of unilateral neglect. In: Robertson IH, Marshall
JC, editors. Unilateral Neglect: Clinical and Experimental Stu-
dies. Hillsdale, NJ: Lawrence Erlbaum Associates, 1993:279�/92.
[41] Rogers RD, Baunez C, Everitt BJ, Robbins TW. Lesions of the
medial and lateral striatum in the rat produce differential deficits
in attentional performance. Behav Neurosci 2001;115:799�/811.
[42] Schallert T. Preoperative intermittent feeding or drinking regi-
mens enhance postlesion sensorimotor function. In: Schulkin J,
editor. Preoperative Events: Their Effects on Behavior Following
Brain Damage. New York: Lawrence Erlbaum Associates,
1989:1�/20.
[43] Schallert T, Whishaw IQ. Bilateral cutaneous stimulation of the
somatosensory system in hemidecorticate rats. Behav Neurosci
1984;98:518�/40.
[44] Schallert T, Upchurch M, Wilcox R, Vaughn DM. Posture-
independent sensorimotor analysis of inter-hemispheric receptor
asymmetries in neostriatum. Pharm Biochem Behav 1983;18:753�/
9.
[45] Schallert T, Hernandez T, Barth TM. Recovery of function after
brain damage: severe and chronic disruption by diazepam. Brain
Res 1986;379:104�/11.
[46] Selemon LD, Goldman-Rakic PS. Common cortical and sub-
cortical targets of the dorsolateral prefrontal and posterior
parietal cortices in the rhesus monkey: evidence for a distributed
neuronal network subserving spatially guided behavior. J Neu-
rosci 1988;8:4049�/68.
[47] Stone SP, Halligan PW, Marshall JC, Greenwood RJ. Unilateral
neglect: a common but heterogeneous syndrome. Neurology
1998;50:1902�/5.
[48] Vallar G, Rusconi ML, Bignamini L, Geminiani G, Perani D.
Anatomical correlates of visual and tactile extinction in humans: a
clinical CT scan study. J Neurol Neurosurg Psychiatr
1994;574:464�/70.
[49] Van Vleet T, Corwin JV, Burcham KJ. October infusion of
apomorphine into the dorsal central striatum produces acute
recovery from AGm-induced neglect in rats. Soc Neurosci Abstr
1999;25:1896.
[50] Van Vleet TM, Burcham KJ, Corwin JV, Reep RL. Unilateral
destruction of the medial agranular cortical projection zone in the
dorsocentral striatum produces severe neglect in rats. Psychobiol-
ogy 2000;28(1):57�/66.
[51] Van Vleet TM, Guerrettaz KR, Burcham KJ, Corwin JV, Reep
RL. Effects of light deprivation on neglect induced by unilateral
destruction of the medial agranular cortical projection zone in the
dorsocentral striatum rats. San Diego Neurosci Abstr 2001;27:85.
[52] Van Vleet TM, Heldt SA, Guerrettaz KR, Corwin JV, Reep RL.
Unilateral destruction of the dorsocentral striatum in rats
produces neglect but not extinction to bilateral simultaneous
stimulation, Behavioural Brain Research, in print.
[53] Vargo JM, Marshall JF. Time-dependent changes in dopamine
agonist-induced striatal fos immunoreactivity are related to
sensory neglect and its recovery after unilateral prefrontal cortex
injury. Synapse 1995;20:305�/15.
[54] Vargo JM, Marshall JF. Frontal cortex ablation reversibly
decreases striatal zif/268 and junB expression: temporal corre-
spondence with sensory neglect and its spontaneous recovery.
Synapse 1996;22:291�/303.
[55] Vargo JM, Marshall JF. Unilateral frontal cortex ablation
producing neglect causes time-dependent changes in striatal
glutamate receptors. Behav Brain Res 1996;77:189�/99.
[56] Vargo JM, Corwin JV, King V, Reep RL. Differential behavioral
effects of left and right hemisphere lesions of medial prefrontal
cortex in rats. Exp Neurol 1988;102:199�/209.
[57] Vargo JM, Richard-Smith M, Corwin JV. Spiroperidol reinstates
asymmetries in neglect in rats recovered from left vs right
dorsomedial prefrontal cortex lesions. Behav Neurosci
1989;103:1017�/27.
[58] Vargo JM, Bromberg BB, Best PJ, Corwin JV, Marshall JF. D1-
class dopamine receptor involvement in the behavioral recovery
from prefrontal cortical injury. Behav Brain Res 1996;72:39�/48.
[59] Vargo JM, Lai HV, Marshall JF. Light deprivation accelerates
recovery from frontal cortical neglect: relation to locomotion and
striatal fos expression. Behav Neurosci 1998;1122:387�/98.
T.M. Van Vleet et al. / Behavioural Brain Research 138 (2003) 165�/178178