effects of larval densities and the duration since larval infestation on the host-searching behavior...
TRANSCRIPT

ARTICLE
Effects of larval densities and the duration since larval infestationon the host-searching behavior of Diadegma semiclausum,a parasitoid of diamondback moth larvae on plants
Yoshitsugu Ohara • Junji Takabayashi
Received: 17 May 2011 / Accepted: 17 January 2012 / Published online: 31 January 2012
� Japan Ethological Society and Springer 2012
Abstract The host-searching behavior of Diadegma
semiclausum, a parasitoid of diamondback moth (DBM)
larvae, was studied in a wind tunnel. Wasps flew prefer-
entially to a cabbage plant, freshly infested by 1 DBM
larva, or one freshly infested by 10 DBM larvae, than to an
intact plant. There was no significant difference between
wasps’ responses to plants infested by different DBM lar-
vae densities. We also found that the duration since the last
infestation by 10 DBM larvae (1 or 3 days) negatively
affected the attractiveness of infested plants. We also
studied the time wasps spent searching for larvae on a
cabbage plant (residence time). The wasps spent ca. 400 s
on a plant freshly infested by 1 larva. Residence time
was significantly longer (ca. 1,200 s) on a plant freshly
infested by either 5 or 15 DBM larvae. Residence time of
D. semiclausum on a plant freshly infested by 5 DBM
larvae was significantly longer than on a previously
infested plant (1 or 3 days after the last infestation). These
results showed that host densities on a plant and the
duration since their last infestation affected the host-
searching behavior of D. semiclausum.
Keywords Diadegma semiclausum � Plutella xylostella �Cabbage plant � Olfactory responses � Residence time
Introduction
Carnivorous natural enemies of herbivorous arthropods are
known to use volatiles produced by herbivore-infested
leaves in their prey finding (e.g., Takabayashi and Dicke
1996; Horiuchi et al. 2003; Sabelis et al. 2007; Arimura
et al. 2009). Studies show that herbivore species and their
developmental stages affect the olfactory response of car-
nivores to prey-infested plant volatiles. Carnivores
responded preferentially to plant volatiles induced by their
prey over those induced by non-prey (e.g., Shiojiri et al.
2000a; De Moraes et al. 1998; Horiuchi et al. 2003). Fur-
thermore, they prefer volatiles from a plant that is infested
by prey of suitable developmental stages (Takabayashi
et al. 1994; Yoneya et al. 2009). Such studies focusing on
the specificity of tritrophic systems are essential for
understanding the ecological significance of tritrophic
interactions.
Once on a prey-infested plant, carnivores start searching
for their prey by using volatile/contact chemicals of either
plant or prey origin, or the combined residue of both left on
prey-infested plants (Sato 1979; Takabayashi et al. 1985;
Takabayashi and Takahashi 1986; Shiojiri et al. 2000b;
Steidle and Fischer 2000; Steidle and Ruther 2000; Maeda
and Takabayashi 2001a; Ohara et al. 2003b; Uefune et al.
2011). Herbivore species also affects the residence time of
prey searching behavior. For example, Sato (1979) reported
that Cotesia glomerata, a parasitoid of cabbage white
butterfly larvae (Pieris rapae), showed characteristic
antennal searching behavior to the open edge of a host-
infested leaf. The search duration was longer than to a non-
host-infested open edge or an artificially damaged edge.
The contact chemicals responsible for this host-specific
response by the wasps are comprised of leaf juice and host
regurgitant (Sato 1979; Horikoshi et al. 1997). Similarly,
Y. Ohara
Mishima High School, Sunto-Gun, Shizuoka 411-0944, Japan
J. Takabayashi (&)
Center for Ecological Research, Kyoto University,
2-509-3 Hirano, Otsu, Shiga 520-2113, Japan
e-mail: [email protected]
123
J Ethol (2012) 30:295–300
DOI 10.1007/s10164-012-0326-0

C. vestalis, a parasitoid of diamondback moth (DBM)
larvae, showed characteristic antennal searching behavior
only to host-infested open leaf edge (Shiojiri et al. 2000b).
It is adaptive for carnivores to find a plant currently
infested by a certain number of prey. Thus, prey density on
a plant (e.g., Maeda and Takabayashi 2001b; Gols et al.
2003; Shiojiri et al. 2010) and duration since the last
infestation (Hanyu et al. 2009; Kugimiya et al. 2010;
Mandour et al. 2011) would be important factors for for-
aging carnivores. We recently reported host-density
dependent/independent response of parasitic wasps to host-
infested plant volatiles (Shiojiri et al. 2010). Cabbage plants
attract more parasitoids (Cotesia glomerata) when there are
more herbivores on the plant (Shiojiri et al. 2010). How-
ever, when cabbage plants are attacked by DBM (Plutella
xylostella) larvae, parasitoids of the larvae (Cotesia ves-
talis) fail to discriminate between herbivore-rich and her-
bivore-poor plants (Shiojiri et al. 2010). Effects of the
duration since the last infestation on the olfactory responses
to infested plants have been reported in tritrophic systems of
corn plants: common armyworms (Mythimna separata) and
parasitic flies Exorista japonica (Hanyu et al. 2009); cab-
bage plants: P. xylostella larvae and C. vestalis (Kugimiya
et al. 2010); and corn plants: common armyworms and
parasitic wasps Cotesia kariyai (Mandour et al. 2011).
Diadegma semiclausum, a solitary parasitoid of
P. xylostella larvae, is thought to be an effective biological
control agent of P. xylostella. In 1993, the wasp was
introduced into Japan from Taiwan to test its effectiveness
(Noda et al. 2000). We previously showed that the wasp
was attracted to herbivore-induced volatiles emitted by
larvae-infested cabbage plants in a wind tunnel (Ohara
et al. 2003a). On the infested plant, the wasp showed
characteristic antennal contact with the host-infested edge
but not with an artificially damaged edge (Ohara et al.
2003a). Both the regurgitant of the host larvae and the juice
of a host plant are essential to elicit the wasp’s antennal
searching on a plant (Ohara et al. 2003a). Effects of host
densities and duration since the last infestations that would
affect foraging behavior of D. semiclausum remain to be
studied. The objectives of this study were to test whether
olfactory response and antennal host-searching behavior of
D. semiclausum is affected by: (1) the number of DBM
larvae on a plant, and (2) the history of infestations.
Materials and methods
Plants and insects
Cabbage plants (Brassica oleracea cv. Shikidori) were
grown separately in 300-ml plastic pots with soil in an
incubator (25 ± 3�C, 60 ± 10% RH, L16:D8) for
approximately 1 month. Plants of ca. 20 cm in height were
used for the experiments.
Eggs of DBM P. xylostella were obtained from the stock
culture of Tohoku National Agricultural Experiment Sta-
tion, and mass-reared on potted plants in a climate-con-
trolled room (25 ± 3�C, 60 ± 10% RH, L16:D8). A
population of D. semiclausum was also obtained from the
stock culture of Tohoku National Agricultural Experiment
Station. Adults of the parasitoid species were fed honey
and contained in a plastic cup (12 cm diameter 9 5 cm
height) in a climate-controlled room (20 ± 2�C, 50–70%
RH, L16:D8) for 2 days to ensure mating. Females were
then kept in a climate-controlled room (15 ± 2�C, 50–70%
RH, L16:D8) until they were tested at 5–12 days old. At
least 1 h prior to each experiment, oviposition-inexper-
ienced females were transferred to another climate-con-
trolled room (25 ± 2�C, 50–70% RH, L16:D8).
Flight response of D. semiclausum to host-infested
plants under different conditions
Experiments were performed in a wind tunnel [50 cm
diameter 9 150 cm length; refer to Ohara et al. (2003a) for
details] in a climate-controlled room (25 ± 2�C, 50–70%
RH). Two potted plants with different treatments were set
20 cm apart at the upwind end of the wind tunnel as odor
sources. For each repetition, a wasp was released down-
wind, from a glass tube (3 cm diameter 9 9 cm length).
We counted the number of wasps that landed on each of the
two treatments (plants). Observations ceased once the wasp
landed on one of the odor sources (treatments). If a wasp
did not land on one of the treatments within 5 min, we
stopped the bioassay. We regarded these wasps as having
made ‘no choice’. We replicated approximately 60 bioas-
says, using different wasp individuals. The experiments
were carried out over 6–7 experimental days. Fresh odor
sources were prepared on each experimental day. Cabbage
plants were inoculated with second and third stadia DBM
larvae, using a fine brush. Similarly, prior to each experi-
ment, DBM larvae were removed from plants with a fine
brush. Damaged leaves were washed with water prior to the
experiments. The following treatments were conducted on
plants for the wind tunnel bioassay:
Treatment (1). An intact cabbage plant.
Treatment (2). A cabbage plant damaged by one larva
for 1 day. We placed a larva on a leaf.
Treatment (3). A cabbage plant damaged by 10 larvae
for 1 day. We placed 2–3 larvae per leaf.
Treatment (4). A cabbage plant damaged according to
treatment (3) was kept in the climate room for 1 day.
Treatment (5). A cabbage plant damaged according to
treatment (3) was kept in the climate room for 3 days.
296 J Ethol (2012) 30:295–300
123

Treatments (1) to (3) were designed to test the flight
response of D. semiclausum to cabbage plants damaged by
different numbers of P. xylostella larvae (Experiment 1).
Plants of treatments (3) to (5) were designed to test the
flight response of D. semiclausum to cabbage plants that
were infested by P. xylostella larvae for different durations
(Experiment 2).
Residence times of D. semiclausum on leaves infested
by larvae at different densities and for different
durations
A potted cabbage plant (ca. 25 cm high) infested by
10 second stadium P. xylostella larvae for 1 day (upwind
odor source) was placed in the wind tunnel, and one cab-
bage leaf (patch: ca. 15 cm high) was placed 50 cm
downwind of the pot. A female wasp was placed in a
capped glass tube (3 cm diameter 9 9 cm length). To
introduce the wasp to a patch, we gently collected it in the
glass tube with an insect aspirator, and allowed it to walk
onto the patch. After release, we measured its residence
time. When the wasp flew away from the patch and landed
on the upwind odor source plant or on the wind tunnel wall,
it was recorded as having left the patch. This experiment
was replicated approximately 20 times, using different
wasps for each experiment. The experiments were con-
ducted over 3–4 days. A fresh odor source was prepared on
each experimental day. Second stadium larvae were care-
fully placed on, and removed from, leaves, using a fine
brush. Prior to the experiments, host larvae were removed.
Leaves were not washed with water for these experiments
because the wash has been shown to reduce the residence
time of wasps (Ohara et al. 2003b). The feces were not
removed either. Plants were treated in the following ways
for the wind tunnel bioassay:
Treatment (1). A cabbage leaf damaged by 1 second
stadium larva for 1 day.
Treatment (2). A cabbage leaf damaged by 5 second
stadium larvae for 1 day.
Treatment (3). A cabbage leaf damaged by 15 second
stadium larvae for 1 day.
Treatment (4). A cabbage leaf damaged according to
treatment (2) was kept in the climate room for 1 day.
Treatment (5). A cabbage leaf damaged according to
treatment (2) was kept in the climate room for 3 days.
Treatments (1) to (3) were to test the effects of the
number of DBM larvae in a patch on the residence time of
D. semiclausum (Experiment 3). Leaves of treatments (2),
(4) and (5) were to test the effect of the duration since the
last infestation of a patch, on the tendency of D. semi-
clausum to leave a patch (Experiment 4).
Statistics
Binomial testing was used for Experiment 1 and 2. Scheffe
post hoc test was used for Experiment 3 and 4.
Results
Experiment 1: flight responses of D. semiclausum
to cabbage plants damaged by different P. xylostella
larval densities
Diadegma semiclausum showed a significant preference for
plants damaged by either 1 or 10 larvae over the undam-
aged plant (Fig. 1; binomial test, P \ 0.0001). However,
wasps were equally distributed between the plants infested
by 1 larva versus those infested by 10 larvae (Fig. 1;
binomial test, P = 0.36).
Experiment 2: flight responses of D. semiclausum
to cabbage plants of different durations since the last
infestation by P. xylostella larvae
Wasps preferred the newly damaged plant to the damaged
plant 1 day after the last infestation (Fig. 2). The damaged
plant 1 day after the last infestation was favored over the
damaged plant 3 days after the last infestation (Fig. 2:
binomial test, P \ 0.01).
Experiment 3: effect of DBM larvae density
on patch-leaving decision by D. semiclausum
Wasps stopped at a host-damaged leaf edge and made
obvious antennal contact with the edge. The wasps spent a
significantly shorter residence time on a patch damaged by
NCPlants damaged by 10 larvae
Intact plants
Plants damaged by one larva
NC
19***
Intact plants
Plants damaged by 10 larvae Plants damaged by one larva
16
***
40 20 0 20
18NS
The number of D. semiclausum
Fig. 1 Response of D. semiclausum in a wind tunnel to cabbage
plants infested by host larvae versus intact cabbage plants.
***P [ 0.001; NS not significantly different (binominal test)
J Ethol (2012) 30:295–300 297
123
1 larva than on a patch damaged by 5 or 15 larvae (Fig. 3:
one-way ANOVA, F2,58 = 15.268, P \ 0.0001; Scheffe’s
test, P \ 0.0001). Consequently, it appears that these
wasps can evaluate the density of host larvae on a patch
and adapt their residence time accordingly.
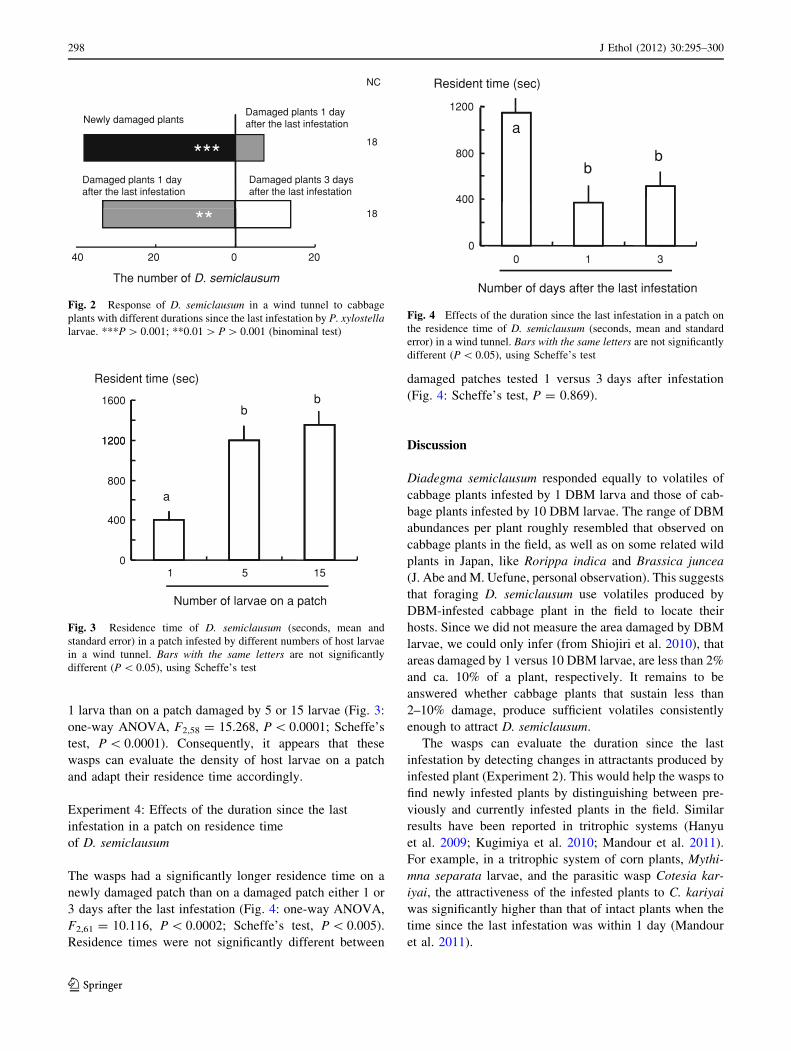
Experiment 4: Effects of the duration since the last
infestation in a patch on residence time
of D. semiclausum
The wasps had a significantly longer residence time on a
newly damaged patch than on a damaged patch either 1 or
3 days after the last infestation (Fig. 4: one-way ANOVA,
F2,61 = 10.116, P \ 0.0002; Scheffe’s test, P \ 0.005).
Residence times were not significantly different between
damaged patches tested 1 versus 3 days after infestation
(Fig. 4: Scheffe’s test, P = 0.869).
Discussion
Diadegma semiclausum responded equally to volatiles of
cabbage plants infested by 1 DBM larva and those of cab-
bage plants infested by 10 DBM larvae. The range of DBM
abundances per plant roughly resembled that observed on
cabbage plants in the field, as well as on some related wild
plants in Japan, like Rorippa indica and Brassica juncea
(J. Abe and M. Uefune, personal observation). This suggests
that foraging D. semiclausum use volatiles produced by
DBM-infested cabbage plant in the field to locate their
hosts. Since we did not measure the area damaged by DBM
larvae, we could only infer (from Shiojiri et al. 2010), that
areas damaged by 1 versus 10 DBM larvae, are less than 2%
and ca. 10% of a plant, respectively. It remains to be
answered whether cabbage plants that sustain less than
2–10% damage, produce sufficient volatiles consistently
enough to attract D. semiclausum.
The wasps can evaluate the duration since the last
infestation by detecting changes in attractants produced by
infested plant (Experiment 2). This would help the wasps to
find newly infested plants by distinguishing between pre-
viously and currently infested plants in the field. Similar
results have been reported in tritrophic systems (Hanyu
et al. 2009; Kugimiya et al. 2010; Mandour et al. 2011).
For example, in a tritrophic system of corn plants, Mythi-
mna separata larvae, and the parasitic wasp Cotesia kar-
iyai, the attractiveness of the infested plants to C. kariyai
was significantly higher than that of intact plants when the
time since the last infestation was within 1 day (Mandour
et al. 2011).
Newly damaged plantsDamaged plants 1 dayafter the last infestation
NC
Damaged plants 1 dayafter the last infestation
Damaged plants 3 daysafter the last infestation
18
***
18**40 20 0 20
The number of D. semiclausum
Fig. 2 Response of D. semiclausum in a wind tunnel to cabbage
plants with different durations since the last infestation by P. xylostellalarvae. ***P [ 0.001; **0.01 [ P [ 0.001 (binominal test)
1200
1600
Resident time (sec)
bb
400
800
1200
a
01 5 15
Number of larvae on a patch
Fig. 3 Residence time of D. semiclausum (seconds, mean and
standard error) in a patch infested by different numbers of host larvae
in a wind tunnel. Bars with the same letters are not significantly
different (P \ 0.05), using Scheffe’s test
Resident time (sec)
800
1200
b
a
b
0
400
0 1 3
Number of days after the last infestation
Fig. 4 Effects of the duration since the last infestation in a patch on
the residence time of D. semiclausum (seconds, mean and standard
error) in a wind tunnel. Bars with the same letters are not significantly
different (P \ 0.05), using Scheffe’s test
298 J Ethol (2012) 30:295–300
123

We have already reported that D. semiclausum uses
multiple cues (such as host feces, silks, and chemicals left
on the leaf surface by the host larvae) in the current patch
for their patch-leaving decision (Ohara et al. 2003b). The
response of the wasps to such complex cues reached a
plateau at 5 larvae per patch (Experiment 3). Thus, the
appeal of a 5 larvae-damaged patch and a 15 larvae-dam-
aged patch were roughly equivalent for the wasps. As
mentioned above, the abundances per plant roughly match
that observed in the field. Similar results were reported by
Maeda et al. (1998): the residence time of the predators in
patches with 40 eggs and those with 140 eggs did not differ
significantly.
It would clearly be adaptive for the wasps to stay a
shorter length of time in a patch where the host larvae had
departed more than 1 day previously than in a newly
infested patch (Experiment 4). The wasps’ residence times
did not differ significantly between patches of 1 versus
3 days after infestation. So it is suggested that the chemical
cues on leaf surfaces that wasps use to evaluate the dura-
tion since infestation in a patch probably decrease critically
within a day.
To use carnivores for pest management, it is important
to know whether, and if so how, carnivores use volatile/
contact infochemicals from plants, herbivores, and/or
plant–herbivore complex origins. In this study, we showed
that biotic factors such as host densities on a plant and the
duration since the last infestation affected the host-
searching behavior of D. semiclausum. Further, abiotic
factors would also affect carnivores’ responses (e.g.,
Takabayashi et al. 1994; Maeda et al. 2000). These factors
should also be studied for the effective use of D. semi-
clausum as a pest management agent.
Acknowledgments This research was partly supported by a Grant-
in-Aid for Scientific Research (S) (No. 19101009), JSPS Core-to-Core
project, and by the Global Center of Excellence Program ‘‘Formation
of a Strategic Base for Biodiversity and Evolutionary Research: from
Genome to Ecosystem’’ of Kyoto University.
References
Arimura G, Matsui K, Takabayashi J (2009) Chemical and molecular
ecology of herbivore-induced plant volatiles: proximate factors
and their ultimate functions. Plant Cell Physiol 50:911–923
De Moraes CM, Lewis WJ, Pare PW, Alborn HT, Tumlinson JH
(1998) Herbivore-infested plants selectively attract parasitoids.
Nature 393:570–573
Gols R, Roosjen M, Dijkman H, Dicke M (2003) Induction of direct
and indirect plant responses by jasmonic acid, low spider mite
densities, or a combination of jasmonic acid treatment and spider
mite infestation. J Chem Ecol 29:2651–2666
Hanyu K, Ichiki RT, Nakamura S, Kainoh Y (2009) Duration and
location of attraction to herbivore-damaged plants in the tachinid
parasitoid Exorista japonica. Appl Entomol Zool 44:371–378
Horikoshi M, Takabayashi J, Yano S, Yamaoka R, Ohsaki N, Sato Y
(1997) Cotesia glomerata female wasps use fatty acids from plant–
herbivore complex in host searching. J Chem Ecol 23:1505–1515
Horiuchi J, Arimura G, Ozawa R, Shimoda T, Takabayashi J,
Nishioka T (2003) A comparison of the responses of Tetrany-chus urticae (Acari: Tetranychidae) and Phytoseiulus persimilis(Acari: Phytoseiidae) to volatiles emitted from lima bean leaves
with different levels of damage made by T. urticae or
Spodoptera exigua (Lepidoptera: Noctuidae). Appl Entomol
Zool 38:109–116
Kugimiya S, Shimoda T, Tabata J, Takabayashi J (2010) Present or
past herbivory: a screening of volatiles released from Brassicarapa under caterpillar attacks as attractants for the solitary
parasitoid, Cotesia vestalis. J Chem Ecol 36:620–628
Maeda T, Takabayashi J (2001a) Patch-leaving decision of the
predatory mite Amblyseius womersleyi (Acari: Phytoseiidae)
based on multiple signals from both inside and outside a prey
patch. J Insect Behav 14:829–839
Maeda T, Takabayashi J (2001b) Production of herbivore-induced
plant volatiles and their attractiveness to Phytoseiulus persimilis(Acari: Phytoseiidae) with changes of Tetranychus urticae(Acari: Tetranychidae) density on a plant. Appl Entomol Zool
36:47–52
Maeda T, Takabayashi J, Yano S, Takafuji A (1998) Factors affecting
the residence time of the predatory mite Phytoseiulus persimilis(Acari: Phytoseiidae) in a prey patch. Appl Entomol Zool
33:573–576
Maeda T, Takabayashi J, Yano S, Takafuji A (2000) Effects of light
on the tritrophic interaction between kidney bean plants, two-
spotted spider mites and predatory mites, Amblyseius womersleyi(Acari : Phytoseiidae). Exp Appl Acarol 24:5–6
Mandour NS, Kainoh Y, Ozawa R, Takabayashi J (2011) Effects of
time after last herbivory on the attraction of corn plants infested
with common armyworms to a parasitic wasp Cotesia kariyai.J Chem Ecol 37:262–272
Noda T, Miyai S, Takashino K, Nakamura A (2000) Density
suppression of Plutella xylostella (Lepidoptera: Yponomeutidae)
by multiple releases of Diadegma semiclausum (Hymenoptera:
Ichneumonidae) in cabbage fields in Iwate, northern Japan.
J Appl Entomol Zool 35:557–563
Ohara Y, Takafuji A, Takabayashi J (2003a) Response to host-infested
plants in females of Diadegma semiclausum Hellen (Hymenop-
tera: Ichneumonidae). Appl Entomol Zool 38:157–162
Ohara Y, Takafuji A, Takabayashi J (2003b) Factors affecting the
patch-leaving decision of the parasitic wasp Diadegma semi-clausum (Hymenoptera: Ichneumonidae). App Entomol Zool
38:211–214
Sabelis M, Takabayashi J, Janssen A, Kant M, van Wijk M, Sznajder
B, Aratchige N, Lesna I, Belliure B, Schuurink R (2007) Ecology
meets plant physiology: herbivore-induced plant responses and
their indirect effects on arthropod communities. In: Ohgushi T,
Craig T, Price P (eds) Ecological communities: plant mediation
in indirect interaction webs. Cambridge University Press,
Cambridge, pp 188–217
Sato Y (1979) Experimental studies on parasitization by Apantelesglomeratus. V. Factors leading a female to the host. Physiol
Entomol 4:63–70
Shiojiri K, Takabayashi J, Yano S, Takafuji A (2000a) Flight
response of parasitoid toward plant–herbivore complexes: a
comparative study of two parasitoid-herbivore systems on
cabbage plants. Appl Entomol Zool 35:87–92
Shiojiri K, Takabayashi J, Yano S, Takafuji A (2000b) Herbivore-
species-specific interactions between crucifer plants and parasitic
wasps (Hymenoptera: Braconidae) that are mediated by info-
chemicals present in areas damaged by herbivores. Appl
Entomol Zool 35:519–524
J Ethol (2012) 30:295–300 299
123

Shiojiri K, Ozawa R, Kugimiya S, Uefune M, van Wijk M, Sabelis
MW, Takabayashi J (2010) Herbivore-specific, density-depen-
dent induction of plant volatiles: honest or ‘‘Cry Wolf’’ signals?
PLoS One e12161
Steidle JLM, Fischer A (2000) Quantity does matter: how feces are
used for host stage selection by granary weevil parasitoid
Lariophagus distinguendus. J Chem Ecol 26:2657–2664
Steidle JLM, Ruther J (2000) Chemicals used for host recognition by
the granary weevil parasitoid Lariophagus distinguendus.
J Chem Ecol 26:2665–2675
Takabayashi J, Dicke M (1996) Plant–carnivore mutualism through
herbivore-induced carnivore attractants. Trends Plant Sci 1:
109–113
Takabayashi J, Takahashi S (1986) Effect of kairomones in the host
searching behavior of Apanteles kariyai Watanabe (Hymenoptera,
Braconidae), a parasitoid of the common arymworm, Pseudaletiaseparata Walker (Lepidoptera, Noctuidae). 3. Synthesis and
bioassay of arrestants and related compounds. Appl Entomol Zool
21:519–524
Takabayashi J, Noda T, Takahashi S (1985) Effect of kairomones in
the host searching behavior of Apanteles kariyai Watanabe
(Hymenoptera, Braconidae), a parasitoid of the common arym-
worm, Pseudaletia separata Walker (Lepidoptera, Noctuidae). 1.
Presence of arresting stimulants produced by the host larvae.
Appl Entomol Zool 20:484–489
Takabayashi J, Dicke M, Posthumus MA (1994) Volatile herbivore-
induced terpenoids in plant–mite interactions: variation caused
by biotic and abiotic factors. J Chem Ecol 20:1324–1354
Uefune M, Kugimiya S, Sano K, Takabayashi J (2011) Herbivore-
induced plant volatiles enhance the ability of parasitic wasps to
find hosts on a plant. J Appl Entomol 136:133–138
Yoneya K, Kugimiya S, Takabayashi J (2009) Can herbivore-induced
plant volatiles inform predatory insects about the most suitable
stage of its prey? Physiol Entomol 34:379–386
300 J Ethol (2012) 30:295–300
123