effects of free atp, citrate, and bicarbonate on rat liver mitochondrial atpase
TRANSCRIPT

ARCHIVES OF BIOCHEMISTRY AND BIOPHYSICS Vol. 196, No. 1, August, pp. l-6, 1979
Effects of Free ATP, Citrate, and Bicarbonate on Rat Liver Mitochondrial ATPase
ESTEBAN SANTIAGO, ANA J. IRIARTE, MARfA J. LOPEZ-ZABALZA, AND NATALIA LOPEZ-MORATALLA
Department of Biochemistry, University of Navarra, Pamplonu, Spain
Received February 7, 1979
The effects of citrate, free ATP, and bicarbonate on the activity of isolated rat liver mitochondrial ATPase have been studied. Citrate behaved as a weak noncompetitive inhibitor; free ATP at low concentrations was found to be an activator, whereas at high concentrations behaved as an inhibitor. Citrate competed with free ATP. Neither citrate nor free ATP competed with bicarbonate. The results obtained suggest the existence of at least two catalytic sites with ATP hydrolyzing activity and at least three regulatory sites: one for bicarbonate, and two for free ATP, one with low and another with high affinity. Citrate would compete with free ATP for the same sites. The interaction of free ATP with the high affinity site activated the enzyme, whereas its interaction with the low affinity site would lead to an inhibition.
Pedersen (1) and Ebel and Lardy (2) have carried out a series of kinetic studies on rat liver mitochondrial ATPase and have suggested the existence of two sites with catalytic activity on ATP hydrolysis with different KM values. However, the existence of only one catalytic site together with a separate modifying site has not been absolutely excluded (2). The presence of a number of nucleotide binding sites on ATPase has been reported by different authors (3-10). On the other hand, mitochondrial ATPase from different sources has been shown to exhibit a maximum activity which depends on an optimum ratio of ATP and Mg2+ concentrations of approxi- mately 1.5 (11-13). Since ATP-Mg2+ is the actual substrate of the enzyme (14), it is reasonable to think that free ATP might exert some regulatory function on mito- chondrial ATPase.
The studies here reported, in which varying concentrations of free ATP were used in a wide range of those of ATP-Mg2+, showed that free ATP, depending on its concentration, exhibited an activating or an inhibitory effect. The kinetic data obtained would be compatible with the existence of two catalytic sites for the hydrolysis
of ATP-Mg2+ and two modifying sites with different affinity for free ATP. Bicarbonate, which is known to be an activating anion for ATPase (1, 2, 15, 16), was found to act independently of free ATP, thus suggesting a different binding site. Another physiological anion tested was citrate, which was found to be a noncompetitive inhibitor “versus” substrate ATP-Mg2+; however, citrate exhibited a competitive pattern of inhibition “versus” free ATP. The inhibitory effect of citrate and the activating effect of bicarbonate were always additive.
MATERIALS AND METHODS
Isolation of mitochndria. Rat liver mitochondria were isolated as described by Hogeboom (1’7) in 250 mM sucrose.
Preparation of mitochondrial ATPase. F,-ATPase was prepared from rat liver mitochondria by the procedure of Lambeth and Lardy (16).
Assay of ATPase activity. ATPase was assayed by measuring the release of inorganic phosphate essentially as described by Pullman et al. (13) in the absence of an ATP-regenerating system. Aliquots of the F,-ATPase were preincubated for 5 min at 30°C in 0.8 ml of a medium containing 5 pmol of Tris-acetate, pH 7.4. The reaction was initiated by the addition of substrate (ATP-MgZ+) at the appropriate concentrations. The incubation was continued for 2 min and stopped by the
1 0003-9861/79/090001-06$02.00/O Copyright 0 1979 by Academic Press, Inc. All rights of reproduction in any form reserved.

2 SANTIAGO ET AL.
addition of 0.1 ml of 50% trichloroacetic acid. Inorganic phosphorus was determined according to Fiske and Subbarow (18). Reagent and enzyme blanks were determined in each experiment.
ATPase was also assayed spectrophotometrically at 340 nm, and at 3O”C, by coupling the production of ADP to the oxidation of NADH via the pyruvate kinase and the lactic dehydrogenase, essentially as described by Pullman et al. (13). The reaction mixture contained in a final volume of 1 ml at pH 7.4, 50 mM Tris-acetate, 0.20 mM NADH, 2 mM
phosphoenolpyruvate, 13 pg of lactic dehydrogenase, and 32 pg of pyruvate kinase. ATP-Mg*+, pH 7.4 and F,-ATPase were added at concentrations indicated in the figures. The reactions were initiated by the addition of isolated ATPase appropriately diluted with 250 mM sucrose.
The K, was determined as follows: Using a constant concentration of the substrate ATP-Mg”+, plots of l/v-v, “versus” l/concentration of activator were constructed (where v = velocity in the presence and v0 = velocity in the absence of activator); the y-inter- cept represented l/V-v,; slope/intercept was defined as the K, for the activator (2). The different Ki values were calculated according to Cleland (19).
Determination of protein. Protein determination was carried out following the reaction of Lowry et al. (20). Crystalline bovine serum albumin was used as standard.
RESULTS AND DISCUSSION
ATPase Activity Catalyzed by Purified Mitochondrial ATPase
Since the true substrate of ATPase is the ATP-Mg2+ complex, the reaction rates were measured within the range of concen- trations from 0.06 to 6 mM and plotted against the corrected substrate concentra- tions taking into account the stability constant of the complex, in order to calculate the actual kinetic constants. The stability constant of ATP-Mg2+ complex is of the order of 20,000 M-l in Tris-HCl buffer (21,22). The kinetic constants were calculated from Eadie-Hofstee plots (23, 24) for high (from 0.5 to 6 IrIM ATP-MgZ+) and low (from 0.06 to 0.4 mM ATP-Mg2+) substrate concentrations and the following values were obtained: V, 8.6 -c 0.2 and 21.5 & 0.4 pmol of ATP hydrolyzed/min x mg; K,, 0.052 + 0.002 and 0.88 2 0.02 mM (see in Figs. 1A and B the curves corresponding to the controls). These values are well in agreement with those previously reported
in the literature (1). The correction after using the stability constant of the ATP-Mg2+ complex did not introduce any substantial change.
Effect of Free ATP
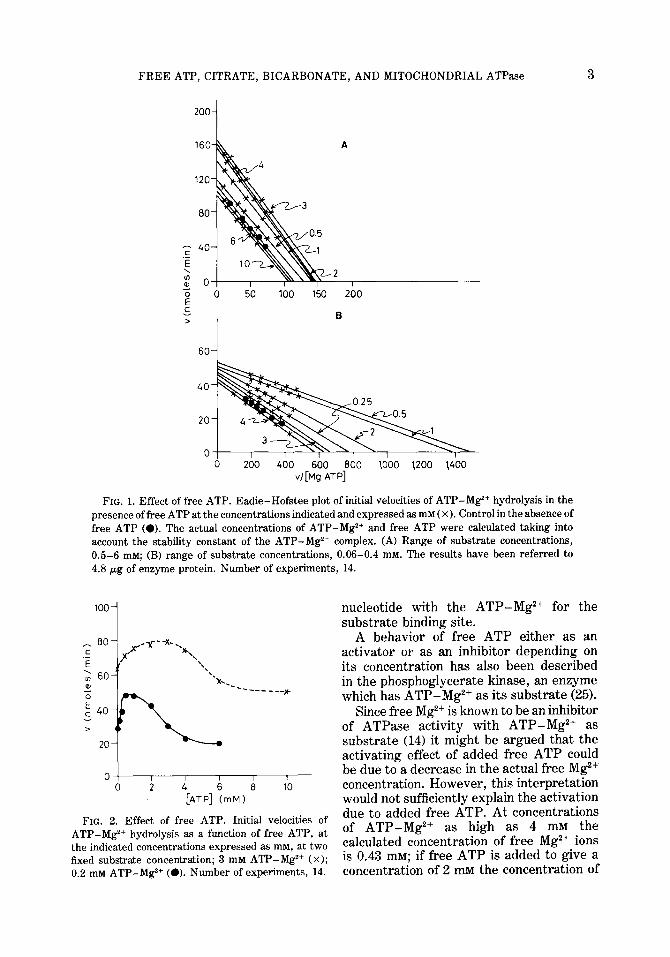
Figure 2 shows the effect of varying concentrations of free ATP on the reaction rate, while holding a constant concentration of the substrate ATP-Mg2+. It may be seen that a bell-shaped curve was obtained. In the presence of high concentrations of free ATP, the reaction rates became lower than those obtained in the absence of free ATP, thus indicating its inhibitory effect at high concentrations.
The effect of different concentrations of free ATP (from 0.25 to 10 InM), at several substrate concentrations, has been studied. The values obtained have been represented on Eadie-Hofstee plots, for high (Fig. 1A) and for low substrate concentrations (Fig. 1B). It may be observed that the activity of each of the two possible catalytic sites, as suggested by the plots, was modified by free ATP. At low concentrations free ATP had an activating effect, whereas increasing its concentration, the activating effect decreased and an inhibitory effect appeared. V and K, of each catalytic site were modified by free ATP. The K, value for free ATP at low substrate concentrations, K,,, was 2.6 2 0.09 x lop4 M; at high sub- strate concentrations, K,,, was 2.8 ? 0.09 x 10m4 M. The Ki values for free ATP were also different for high and low sub- strate concentrations: Ki, = 1.2 ? 0.06 x 10p2; Ki2 = 3.7 + 0.1 x 10m2 M. It is therefore obvious that the inhibitory effect of free ATP in both catalytic sites was much weaker than its activating effect.
These activating and inhibitory effects of free ATP could be interpreted as the result of an interaction of the nucleotide with two sites exhibiting different affinities. The interaction with the site of higher affinity would lead to an activation, whereas the interaction with the site of lower affinity would cause an inhibition of the enzyme. The inhibitory effect of free ATP at high concentrations could also be interpreted as a competition of the free
FREE ATP, CITRATE, BICARBONATE, AND MITOCHONDRIAL ATPase 3
60 -I
0 0 200 400 600 800 1000 1200 1400
v/ 14 AT PI
FIG. 1. Effect of free ATP. Eadie-Hofstee plot of initial velocities of ATP-Mg2+ hydrolysis in the presence of free ATP at the concentrations indicated and expressed as mM ( x). Control in the absence of free ATP (0). The actual concentrations of ATP-Mg2+ and free ATP were calculated taking into account the stability constant of the ATP-Mg2+ complex. (A) Range of substrate concentrations, 0.5-6 mM; (B) range of substrate concentrations, 0.06-0.4 mM. The results have been referred to 4.8 pg of enzyme protein. Number of experiments, 14.
0 1 I I I I I 0 2 4 6 0 10
[ATP] CmM)
FIG. 2. Effect of free ATP. Initial velocities of ATP-Mg2+ hydrolysis as a function of free ATP, at the indicated concentrations expressed as mM, at two fixed substrate concentration; 3 mM ATP-MgZ+ (x);
0.2 rIIM ATP-Mg2+ (0). Number of experiments, 14.
nucleotide with the ATP-MgZ+ for the substrate binding site.
A behavior of free ATP either as an activator or as an inhibitor depending on its concentration has also been described in the phosphoglycerate kinase, an enzyme which has ATP-Mg2+ as its substrate (25).
Since free MgZ+ is known to be an inhibitor of ATPase activity with ATP-Mg2+ as substrate (14) it might be argued that the activating effect of added free ATP could be due to a decrease in the actual free Mg2+ concentration. However, this interpretation would not sufficiently explain the activation due to added free ATP. At concentrations of ATP-Mg*+ as high as 4 mM the calculated concentration of free Mg2+ ions is 0.43 mM; if free ATP is added to give a concentration of 2 mM the concentration of
4 SANTIAGO ET AL.
free Mg2+ ions would become 0.1 mM. Experiments have been carried out to determine the inhibitory effect of added Mg2+ ions in the presence of 4 mM ATP-Mg2+. In the range of 0.3-0.5 mM free Mg2+ the inhibition was of the order of 5-Y%, whereas the activating effect of the 2 mM concentration of free ATP added was over 30%. At concentrations of 0.1 mM ATP-Mg2+ the calculated free Mgl+ con- centration is approximately 0.05 mM. After adding free ATP to give a 0.05 mM concen- tration the calculated free Mg2+ concentra- tion would be equal to 0.01 mM. The activity has also been determined for this substrate concentration with added free Mg2+. It was observed that at concentration of free Mg2+ of the order of 0.05 InM the inhibitory effect was negligible, whereas the activating effect of free 0.5 mM ATP was approxi- mately 60%.
Effect of Bicarbonate in the Presence of Free ATP
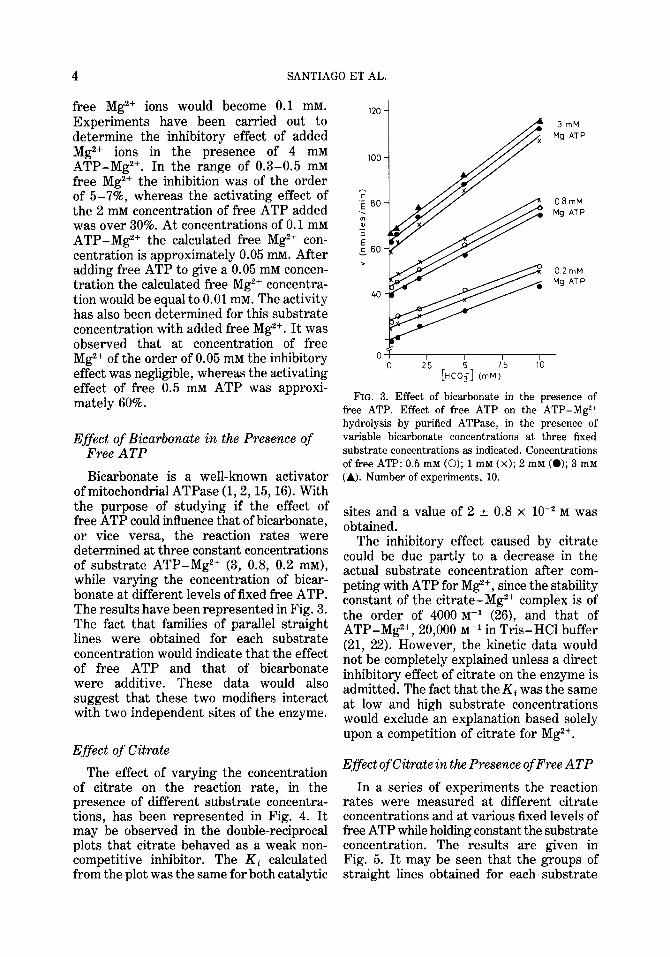
Bicarbonate is a well-known activator of mitochondrial ATPase (1,2, 15,161. With the purpose of studying if the effect of free ATP could influence that of bicarbonate, or vice versa, the reaction rates were determined at three constant concentrations of substrate ATP-Mg2+ (3, 0.8, 0.2 mM), while varying the concentration of bicar- bonate at different levels of fixed free ATP. The results have been represented in Fig. 3. The fact that families of parallel straight lines were obtained for each substrate concentration would indicate that the effect of free ATP and that of bicarbonate were additive. These data would also suggest that these two modifiers interact with two independent sites of the enzyme.
Effect of Citrate
The effect of varying the concentration of citrate on the reaction rate, in the presence of different substrate concentra- tions, has been represented in Fig. 4. It may be observed in the double-reciprocal plots that citrate behaved as a weak non- competitive inhibitor. The Ki calculated from the plot was the same for both catalytic
3 rnM Mg ATP
08mM Mq ATP
02mM Mq ATP
04 1 I I I 0
25 [HCO:,] (mMT5 lo
FIG. 3. Effect of bicarbonate in the presence of free ATP. Effect of free ATP on the ATP-M$+ hydrolysis by purified ATPase, in the presence of variable bicarbonate concentrations at three fixed substrate concentrations as indicated. Concentrations of free ATP: 0.5 mM (0); 1 ml (X); 2 mM (0); 3 mM
(A). Number of experiments, 10.
sites and a value of 2 + 0.8 x 10T2 M was obtained.
The inhibitory effect caused by citrate could be due partly to a decrease in the actual substrate concentration after com- peting with ATP for Mg2+, since the stability constant of the citrate-Mg2+ complex is of the order of 4000 M-’ (26), and that of ATP-Mg2+, 20,000 M-’ in Tris-HCl buffer (21, 22). However, the kinetic data would not be completely explained unless a direct inhibitory effect of citrate on the enzyme is admitted. The fact that the Ki was the same at low and high substrate concentrations would exclude an explanation based solely upon a competition of citrate for Mg2+.
Effect of Citrate in the Presence of Free ATP
In a series of experiments the reaction rates were measured at different citrate concentrations and at various fixed levels of free ATP while holding constant the substrate concentration. The results are given in Fig. 5. It may be seen that the groups of straight lines obtained for each substrate

FREE ATP, CITRATE, BICARBONATE, AND MITOCHONDRIAL ATPase 5
I I / I 1 I
-3 -2 -1 0 1 2 3 l/ [Mg ASP] (mM-‘)
-1‘6 -12 -8 -4 6 I,;, ATP] h+l’l:
FIG. 4. Effect of citrate. Lineweaver-Burk diagram of initial velocities of ATP-Mg2+ hydrolysis by purified ATPase as a function of substrate concentration, at different citrate concentrations. Citrate absent (0); 2.5 mM citrate (X); 5 mM citrate (0); 10 mM citrate (A). Number of experiments, 15.
concentrations have the same y-intercept. This competitive pattern would imply that the binding site for free ATP coincides with that of citrate.
Effect of Citrate in the Presence of Bicarbonate
The reaction rates were also determined at two constant substrate concentrations (3 and 0.2 InM) and by varying the bicarbonate concentration at different levels of citrate. The results have been represented in Fig. 6. It may be seen that groups of parallel straight lines were obtained for each substrate concentration. The simplest explanation for this behavior would be that the inhibitory effect of citrate and the activating effect of bicarbonate are in- dependent of each other, reflecting the interaction of these two modifiers on different sites of the enzyme.
The results now reported seem to support the existence of a binding site for free ATP per catalytic site of the enzyme. The
binding of the modulator free nucleotide would induce an increase in the hydrolysis rate. The inhibitory effect of high concen- trations of free ATP could be explained through a competition with the substrate ATP-Mg2+ for the substrate binding site. The existence of two possible types of nucleotide binding sites would be in agree- ment with the results of Adolfsen and Moudrianakis (27). These authors have suggested a “flip-flop” mechanism in which the enzyme would oscillate between two alternate but catalytically equivalent con- formational states. These data would also be in agreement with an alternating site model as proposed by Boyer (28) and by Hackney and Boyer (29). The kinetic data
0 0.1 0.2 0.3 0.4 l/[CITRATE] (mM-l)
FIG. 5. Effect of citrate in the presence of free ATP. Effect of free ATP on ATP-Mg2+ hydrolysis by purified ATPase in the presence of varying citrate concentrations and at two fixed substrate concentra- tions: (A) 3 mM ATP-M$+; (B) 0.2 mM ATP-Me+; free ATP absent (0); 0.5 mM free ATP (0); 1 mM
free ATP (0); 2 mM free ATP (x); 3 mM free ATP (A). Number of experiments, 12.

3mM Mg ATP
02mM 10. GARRET, N. E., AND PENEFSKY, H. S. (1974) Fed. Mcj ATP Proc. 33, 1399.
FIG. 6. Effect of citrate in the presence of bicarbonate. Citrate inhibition of the ATP-Mg*+ hydrolysis catalyzed by purified ATPase in the presence of variable bicarbonate concentrations and at two fixed substrate concentrations as indicated. Citrate concen- trations: 2.5 mM (0); 5 mM (X); 10 mM (0). Number of experiments, 12.
now reported would also be compatible with the recent molecular model for oxida- tive phosphorylation recently proposed by Santiago and Lopez-Moratalla (30). According to their proposal each of the catalytic sites present in mitochondrial ATPase would require as an essential component a pair of iron atoms which may bind the substrate or substrates and modulators in order to carry out either the oxidative phosphorylation or the hydrolysis of ATP.
REFERENCES
1. PEDERSEN, P. L. (1976) J. Biol. Chem. 251, 934-940.
2. EBEL, R. E., AND LARDY, H. A. (1975) J. Biol. Chm. 250, 191-196.
3. HARRIS, D. A., ROSING, J., VAN DE STADT, R. J., AND SLATER, E. C. (1973) Biochem. Biophys. Acta 314, 149-153.
4. PENEFSKY, H. S. (1974) J. Biol. Chem. 249, 3579-3585.
5. CATTERALL, W. A., AND PEDERSEN, P. L. (1972)
SANTIAGO ET AL.
6. SCHUSTER, S. M., EBEL, R. E., AND LARDY, H. A. (1975) J. Bid. Chem. 250, 7448-7853.
7. PEDERSEN, P. L. (1975) Biochem. Biophys. Res. Commun. 64, 610-616.
8. WAGENVOORD, R. J., VAN DER KRAAN, I., AND KEMP, A. (1977) Biochim. Biophys. Acta 460, 17-24.
9. SENIOR, A. E. (1973) B&him. Biophys. Acta 301, 249-277.
11. HOUSTEK, J., AND DRAHOTA, Z. (1977) Biochim. Biophys. Acta 484, 127-139.
12. DI PIETRO, A., G~DINOT, C., BOUILLANT, M. L., AND GAUTHERON, D. C. (1975) Biochimie 57, 959-967.
13. PULLMAN, M. E., PENEFSKY, H. S., DATTA, A., AND RACKER, E. (1960) J. Biol. Chem. 235, 3322-3329.
14. SELWYN, M. J. (1967) Biochem. J. 105,279-288. 15. FANESTIL, D. P., HASTING, A. B., AND
MAHOWALD, T. A. (1963) J. Biol. Chem. 238, 836-842.
16. LAMBETH, D. O., AND LARDY, H. A. (1971) Eur. J. Biochem. 22, 355-363.
17. HOGEBOOM, G. H. (1955)in Methods in Enzymology (Collowick, S. P., and Kaplan, N. O., eds.), Vol. 1, p. 16, Academic Press, New York.
18. FISKE, C. N., AND SUBBAROUW, Y. (1925) J. Biol. Chem. 66, 375-400.
19. CLELAND, W. W. (1963) Biochim. Biophys. Acta 67, 173-187.
20. LOWRY, 0. H., ROSENBROUGH, N. J., FARR, A. L., AND RANDALL, R. J. (1951) J. Biol. Chem. 193, 265-275.
21. O’SULLIVAN, W. J., AND PERRIN, D. D. (1964) Biochemistry 3, 18-26.
22. BURTON, K. (1959) Btichem. J. 71, 388-395. 23. EADIE, G. S. (1952) Science 116, 688. 24. HOFSTEE, B. H. J. (1952) Science 116, 329-331. 25. LARSSON-RAZNIKIEWICZ, M., AND SCHIERDECK,
B. (1977) Biochim. Biophys. Acta 481,283-287. 26. BLAIR, J. McD. (1969) Eur. J. Biochem. 8,
287-291. 27. ADOLFSEN, R., AND MOUDRIANAKIS, E. N.
(1976) Arch. Biochem. Biophys. 172, 425-433. 28. BOYER, P. D. (1977) Annu. Rev. Biochem. 46,
957-966. 29. HACKNEY, D. D., AND BOYER, P. D. (1978)
J. Biol. Chem. 253, 3169-3170. 30. SANTIAGO, E., AND LOPEZ-MORATALLA, N. (1978)
J. Biol. Chem. 247, 7969-7976. Rev. esp. Fisiol. 34, 477-480.