effect of inhibitors trypsin-like proteolytic enzymes on...
TRANSCRIPT

JOURNAL OF BACTERuOLOGY, Feb. 1983, p. 700-7080021-9193/83/020700-0 2.00/0Copyright 0 1983, American Society for Microbiology
Vol. 153, No. 2
Effect of Inhibitors of Trypsin-Like Proteolytic Enzymes onBacillus cereus T Spore Germination
H. BOSCHWITZ,l Y. MILNER,' A. KEYNAN,' H. 0. HALVORSON,2* AND W. TROLL3Department ofBiological Chemistry, The Hebrew University ofJerusalem, Jerusalem, Israel'; Department of
Biology and Rosenstiel Basic Medical Sciences Research Center, Brandeis University, Waltham,Massachusetts 022542; and Department ofEnvironmental Medicine, New York University Medical Center,
New York, New York 100163
Received 1 March 1982/Accepted 26 November 1982
The germination of Bacillus cereus T spore suspensions is partially preventedby several inhibitors of trypsin-like enzymes. Leupeptin, antipain, and tosyl-lysine-chloromethyl ketone are effective inhibitors, whereas chymostatin, elasta-tinal, and pepstatin are inactive. A synthetic substrate of trypsin, tosyl-arginine-methyl ester, also inhibits germination. Its inhibitory effect decreases as afunction of incubation time in the presence of spores and is abolished by previoushydrolysis with trypsin. Germinating, but not dormant, spore suspensions hydro-lyze tosyl-arginine-methyl ester; its hydrolysis is insensitive to chloramphenicol,sulfhlydryl reagents, and EDTA. A crude extract of germinated B. cereus sporescontains a trypsin-like enzyme whose activity, as measured by hydrolysis ofbenzoyl-arginine p-nitroanilide, is sensitive to germination-inhibitory compoundssuch as leupeptin, tosyl-arginine-methyl ester, and tosyl-lysine-chloromethylketone. Spore suspensions exposed to the above inhibitors under germinationconditions lose only part of their heat resistance and some 10 to 30%o of theirdipicolinic acid content. Part of the germinating spore population becomes "phasegrey" under phase optics. Based on a study of the inhibition of germination byprotease inhibitors and the activity of a protease in germination spores and sporeextracts, it is suggested that the activity of a trypsin-like enzyme may be involvedin the mechanism of the breaking of dormancy in spores of B. cereus T.
Bacterial spores, when exposed to germinat-ing agents, irreversibly lose their spore proper-ties by an as yet obscure mechanism. However,it is generally accepted that germination occursthrough a series of sequential rather than simul-taneous steps (11, 13, 17).Once the initial "trigger reaction" has been
activated, germination continues in the absenceof the inducer (8). By pH and temperaturemanipulations, as well as the use of specificinhibitors, this trigger reaction can be separatedfrom the latter stages of germination (6, 21 26,28). According to Levinson and Hyatt (13), afterthe initiation of the process of germination bythe trigger reaction, the various spore propertiesare changed sequentially in the following order:loss of heat resistance, release of dipicolinic acid(DPA) and Ca2+ into the medium, increase inspore stainability, beginning of phase darkeningand decrease of the optical density of sporesuspension as cortex peptidoglycan is hydro-lyzed and the products released to the medium,and finally, the onset of metabolic activity asmeasured by oxygen uptake.A time lapse is detectable between the trigger
reaction, which starts with the addition of the
germinant, and the first visible changes ob-served microscopically in individual spores (lossof refractility). Based on these observations,Vary and Halvorson (27) divided germinationinto two processes: "microlag" and "microger-mination." Additional evidence for the exis-tence of sequential stages in germination stemsfrom the use of inhibitors that stop the laterstages of germination. Rossignol and Vary (20)reported that 1 mM HgCl2 permits the release ofDPA and partial loss of refractility but preventsthe restoration of oxygen uptake.Mutants have also been described which are
blocked at various stages of germination (4, 16).Some of these mutants undergo the early stagesof germination, such as the loss of heat resist-ance and partial loss of refractility, but areinhibited or delayed in the degradation of thespore cortex and release of hexosamine-contain-ing fragments (1, 30). Cheng et al. (1) describedone germination mutant which was altered in anintracellular protease. Our studies of this mutant(unpublished) suggested a possible involvementof proteases in the germination process. Evi-dence that a protease might be involved ingermination was also reported by Quesnel et al.
on June 22, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

INHIBITION OF B. CEREUS T SPORE GERMINATION 701
(18) based on the fact that in certain strains ofBacillus cereus and Bacillus subtilis, germina-tion can be induced under defined conditions bysubtilisin, a proteolytic enzyme isolated fromvarious strains of B. subtilis (12, 23). To test thehypothesis of the possible involvement of pro-
teolytic enzymes in the early stages of germina-tion, we investigated the effect of protease inhib-itors and substrates on the germination process.
In this paper, "germination" includes onlythose biochemical reactions which are responsi-ble for the irreversible loss of spore propertiesand which also occur in the presence of inhibi-tors of macromolecular synthesis and thereforeare not dependent on new synthesis. We presentevidence that (i) specific inhibitors of trypsin-like protease inhibit germination and probablyinterfere at some stage of the germination se-
quences between the trigger reaction and theloss of refractility and (ii) a trypsin-like proteo-lytic activity exists in germinating spores andspores extracts and is inhibited by the samecompounds that inhibit germination.
MATERIALS AND METHODSMedia and strains. Most of the experiments were
conducted with B. cereus T, which was grown as
previously described (5). Spores were isolated bycentrifugation twice in 509%o urographin, resuspensionin water, and heat shocking by exposure to 60°C for 30min. In experiments in which protease activity wasmeasured on intact spores, the preparations were heatshocked at 70°C for 45 min.
Germination. A suspension of spores in water(108/ml) was heat shocked at 60°C for 30 min. Germi-nation experiments were performed at 36°C in 3 ml ofgermination medium which contained 1 mg of L-alanine, 0.5 mg of adenosine, and 1 ml of K+,Na+-phosphate buffer (pH 7.0). Germination was moni-tored by decrease in optical density (at 660 nm) in aBausch & Lomb Spectronic 20 spectrophotometer.Germinating suspensions were also examined with a
phase-contrast microscope (Zeiss) under oil immer-sion to check the changes occurring in the individualspores when germination was partly inhibited by pro-tease inhibitors. Germination rates were obtained fromthe slopes of semilogarithmic plots of the percentageof residual optical density as a function of time.To conserve the supply of some of the inhibitors,
several experiments were carried out in a total volumeof 0.3 ml. To 0.1 ml of a spore suspension in a 1-mlcuvette, 0.2 ml of germination medium, with or with-out inhibitor, was added. After 15 min of incubation at36°C, the germination mixture was diluted with waterto 1 ml, and the optical density (at 660 nm) in a
Beckman spectrophotometer was immediately com-
pared to a control of spores suspended in water.Making spores permeable. Spores were shaken in 5
M urea-7% (vol/vol) 3-mercaptoethanol (pH 6.2) at37°C for 5 h, washed four times with 0.05 M Tris (pH6.8) containing 0.1% Tween 80, and suspended inwater with 0.05% (vol/vol) Tween 80.DPA asys. DPA assays were performed according
to the method of Janssen et al. (10).
Inhibitors and chemicals. The following compoundswere of the highest purity commercially available:phenylmethylsulfonyl fluoride (PMSF), diisopropyl-fluorophosphate (DFP), soybean trypsin inhibitor, andtosyl-lysine-chloromethyl ketone (TLCK) (all pur-chased from Sigma Chemical Co., St. Louis, Mo.).Leupeptin and papain were supplied by one of us(W.T.). Chymostatin, pepstatin, and elastinal werepurchased from Peptide Institute Inc., Osaka, Japan.Tosyl-arginine-methyl ester (TAME) was purchasedfrom Schwarz/Mann, Orangeburg, N.Y. Dimethylsulfoxide (DMSO) was purchased from Frutatom, andbenzoyl-arginine p-nitroanilide (BANA) and lysozymewere purchased from Sigma Chemical Co. BANA andTAME show a single spot when analyzed in thin-layerchromatography on cellulose sheets (see below).
Protease assay in disrupted germinated spores. Dis-rupted spores were prepared as follows. Between 4and 30 ml of spore suspension in water was heatshocked for 45 min at 60 to 70°C and germinated for 30to 60 min at 37°C in a shaking bath in a mediumcontaining, per milliliter, 100 ,umol of K+,Na+-phos-phate buffer (pH 7.0), 2 x 109 spores, 2 mg of alanine,and 1 mg of adenosine. The germinated spores weresuspended in 3 mM EDTA-0.05% ,3-mercaptoethanol(vol/vol)-0.2% Triton X-100 (vol/vol). In some experi-ments, germination was conducted in the presence ofchloramphenicol (100 Rg/ml) to measure protease ac-tivity in normal or permeable spores in the absence ofprotein biosynthesis (19). The entire mixture was thenagitated in a B Braun rotator (Melsungen model 53030)in the cold with glass beads (0.10 to 0.11 mm; BraunMelsungen AG) for 20 to 30 s. This procedure wasrepeated six times. The degree of spore disintegrationor permeation was only partial and varied in differentexperiments. Permeation to BANA was achieved byheat treatment and germination as described above;however, the mechanical breakage was accomplishedin a French press with three cycles of rapid decom-pression from 7.35 kPa.
Trypsin-like activity was determined as follows. Afinal volume of 2.5 ml, containing 2 x 109 germinatedspores and 50 RI of DMSO was incubated with thetrypsin substrate BANA at 2 mM final concentration.The pH of the mixture was brought to 7.0 by theaddition of 0.1 M K+, Na+-phosphate buffer or 0.1 MTris-hydrochloride buffer (pH 7.1) and the suspensionwas incubated at 37°C for several hours. Color forma-tion indicating trypsin-like activity was measured at420 nm after centrifugation.
Qualitative protease assay with TAME as a substrate.Heat-treated spores were incubated in a 200-RI finalvolume with 50 mM Tris-hydrochloride (pH 7.1) and 5to 30 mM neutralized TAME. The mixture was incu-bated for 5 to 24 h at 37°C, and samples of 5 to 15 pulwere withdrawn and spotted on thin-layer cellulosesheets (Nagel-Machery, Heidelburg, West Germany;CEL-400) with continuous drying. TAME was sepa-rated from its hydrolysis product tosyl-arginine (TA)by chromatography in butanol-pyridine-acetic acid-H20 (50:45:15:60 [vol/vol]). Compounds containingarginine were detected on a thin-layer chromatographyplate by the modified Sakaguchi spray method (seeFig. 2C and SD for the separation achieved). Theplates were sprayed first in solution A containing 5 g ofurea, 10 g of KOH, and 100 mg of a-naphthol in 100 mlof ethanol. Then solution B (0.65 M Br2 and 5 g of
VOL. 153, 1983
on June 22, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

702 BOSCHWITZ ET AL.
KOH in 100 ml of water) was sprayed, the plates weredried, and the violet-red spots were marked.Lom of beat rsin of spores during n.
The loss of heat resistance was determined by plating adilution of spores after heating for 15 min at 80°C onnutrient agar plates.
RESULTSEffect of inhibitors of serine proteases. PMSF
and DFP inhibit serine hydrolytic enzymes, in-cluding proteases. Both compounds also inhibit-ed germination of B. cereus spores. Thesespores germinated completely within 70 min.However, in the presence of 4 mM PMSF (satu-rating solution), fewer than 50%o of the sporesgerminated within 60 min. A similar inhibitionoccurred in the presence of saturating concen-trations of DFP. The observed inhibitions byPMSF and DFP were reversible by centrifuga-tion and resuspension in fresh germination medi-um. Because of the low solubilities and instabil-ity of PMSF and DFP in water, it is difficult toquantitate their effect on germination. We there-fore investigated the effect of more soluble,stable, and specific inhibitors of proteolytic en-zymes on germination.Protease inhibitors of microbial origin.
Umezawa has isolated a variety of low-molecu-lar-weight protease inhibitors from streptomy-cetes (molecular weight, 400 to 700). Of thesecompounds, leupeptin and antipain are inhibi-tors of trypsin-like enzymes and cathepsins(trypsin, papain, plasmin, and cathepsin B),chymostatin inhibits chymotrypsin, elastinal in-hibits elastase, and pepstatin inhibits acid prote-
ases (25). Of these compounds, only leupeptinand antipain inhibited germination.At 30 mM, leupeptin inhibited both the rate
and the extent of germination of spores of B.cereus T by over 80% (Fig. 1). When the concen-tration was reduced to 7.5 mM, there was asmaller inhibitory effect. The Ki value of 11 mMwas obtained from a Dixon plot (2) constructedby plotting reciprocals of germination first-orderrate constant versus inhibitor concentration(Fig. 18).
Antipain has a similar effect, although it issomewhat less inhibitory than leupeptin; 17 mMantipain inhibits germination by 50%. At similarconcentrations, chymostatin, elastinal, and pep-statin had no effect on the kinetics or extent ofgermination.These substances are known to be reversible
protease inhibitors, and we found them to inhibitgermination reversibly. Germination was com-pletely inhibited by 40 mM leupeptin (Fig. 1).When the suspension was diluted threefold, ger-mination proceeded at 50% of the maximal rate(data not shown). The possibility that thesepeptides might compete for the site of the L-alanine-initiated trigger reaction was ruled outby the finding that increased amounts of L-alanine had no effect on the inhibitory propertiesof leupeptin or antipain (data not shown).To demonstrate that the inhibition by the
small-molecular-weight protease inhibitors isnot due to nonspecific binding of positivelycharged lysine- or arginine-containing peptidesor both to a nonenzymatic spore or membranesite, the effect of other similarly charged pep-
500
A ~~~~~~~~~B
0 20 40 60 7.5 15 22.5 30TIME , min [ LEUPEPTIN, mM
FIG. 1. Inhibition of germination of B. cereus T spores as a function of leupeptin concentration. (A) Sporeswere germinated at 36°C as described in the text in a volume of 3 ml containing 6 x 108 spores. Germination wasmeasured as the decrease of optical density (O.D.) at 660 nm as a function of time. 0, Control withoutgermination medium; *, control of spores in germimation medium without leupeptin. Germination medium withleupeptin: x, 40 mM; V, 30 mM; A, 25 mM; A, 12.5 mM; V, 7.5 mM. (B) Dixon plot of data similar to thoseshown in (A). Germination rate constants were obtained as described in the text. k, Definition.
J. BACTERIOL.
on June 22, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

INHIBITION OF B. CEREUS T SPORE GERMINATION 703
tides was tested. Putrescine (50 mM), histone(20 mM), and spermidine (20 mM) had no effecton germination (data not shown).
Effect of synthetic trypsin inhibitors and sub-strates. TAME is a synthetic substrate of trypsinwhich, by virtue of its relative high affinity (29),should effectively compete with normal "natu-ral" substrates of the putative spore protease(s).The effect of 40 mM TAME on germination ofspores ofB. cereus T is shown in Fig. 2A TAMEinhibited germination for at least 4 h (Fig. 2B).To test the specificity of TAME, it was firstincubated for 90 min with trypsin (33 ,ug/ml) at36°C (in 0.2 M phosphate buffer [pH 7.0]), andthe digest was then tested at the same concentra-tion (40 mM). As is also shown in Fig. SC,trypsin predigestion of TAME destroys its in-hibitory effect (Fig. 2A). The complete mole-cule, rather than its hydrolyzed products, is theeffective inhibitor. Under these conditions tryp-sin catalyzed the conversion of TAME to TA.Although trypsin iself has no effect on dor-
mant spores, it probably cannot enter the sporesbecause of its size (molecular weight, 25,000).Similarly, the failure of soybean trypsin inhibitorto effect the germination of spores of B. cereusmay be due to its high molecular weight (21,000).TAME was slowly hydrolyzed when in con-
tact with spores, leading to a loss of ability toinhibit germination. TAME (40 mM) inhibitedgermination for at least 4 h (Fig. 2A and B),
A B
whereas there was a slow recovery of germina-tion from 10 mM TAME (Fig. 2B). This suggeststhat TAME was hydrolyzed by germinatingspores. To test this possibility, various concen-trations of TAME were incubated for 5 h withspore suspensions, and the supernatant wasspotted on thin-layer chromatography plates(see Materials and Methods). TAME was hydro-lyzed by germinating spores at 37°C, as indicat-ed by the appearance of TA (Fig. 2C). Thehydrolysis rate of TAME is higher at 10 mMsubstrate than at 30 mM, probably due to theknown inhibition of protease action (22). Quanti-tative estimations show that after 5 h of incuba-tion of 10 mM TAME with spores, about 50 to60% of the TAME was hydrolyzed. This slowdecrease in TAME concentration could explainthe slow rate of germination seen in the presenceof 10 mM TAME (Fig. 2B). TAME hydrolysisoccurred only when it was added to germinatedspores. In the absence of germinating agents, noTAME hydrolysis was observed.The inhibitory effect ofTAME on the rate and
extent of germination of permeable and nonper-meable spores (see Materials and Methods) wasproportional to TAME concentration in therange of 1 to 20 mM, provided the spore concen-tration was kept constant (Fig. 3A and C).An experiment with low concentrations of
TAME is shown in Fig. 3A. When germinationrate constants of these permeabilized spores
C
6
0
0Il
TAME
TA
0 50 100 0 50 100 150 200 250TIME,min TIME,min mM TAME
FIG. 2. Effect on TAME on germination ofB. cereus T spores. O.D., Optical density. (A) Effect of incubationwith trypsin on inhibition of germination. Spores (2 x 108/ml) were germinated as described in the text. Symbols:0, control without germination; 0, control of spores in germination medium only; A, spores in the presence ofgermination medium containing 40 mM TAME; A, spores in the presence of germination medium containing 40mM TAME which had been incubated with trypsin (33 xLg/ml) at 36°C for 90 min. (B) Effect of low concentrationofTAME on inhibition of germination. Experimental conditions were the same as in (A). Symbols: 0, control,germination medium without inhibitor; A, germination medium with 40 mM TAME; A, germination mediumwith 10 mM TAME; 0, no germination medium, water only. (C) Hydrolysis of TAME by germinated spores.Spores (2 x 108/ml) were incubated in germination medium (see text) with various concentrations of neutralizedTAME as indicated. After 5 h of incubation at 37°C, ca. 0.1 ,umol was spotted on thin-layer chromatographyplates, chromatographed, developed, and sprayed for TAME and TA as described in the text. Black areasrepresent deep red-violet spots, shaded areas represent intermediate red intensity, and white areas representfaint red, the color of the lower arginine-containing material.
VOL. 153, 1983
on June 22, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
704 BOSCHWITZ ET AL.
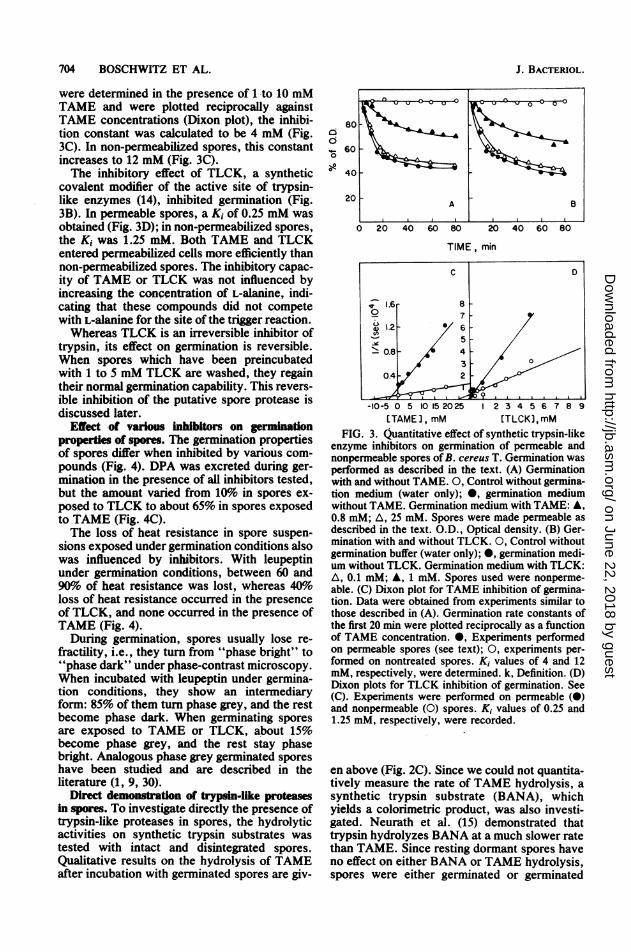
were determined in the presence of to 10 mMTAME and were plotted reciprocally againstTAME concentrations (Dixon plot), the inhibi-tion constant was calculated to be 4 mM (Fig.3C). In non-permeabilized spores, this constantincreases to 12 mM (Fig. 3C).The inhibitory effect of TLCK, a synthetic
covalent modifier of the active site of trypsin-like enzymes (14), inhibited germination (Fig.3B). In permeable spores, a Ki of 0.25 mM wasobtained (Fig. 3D); in non-permeabilized spores,the Ki was 1.25 mM. Both TAME and TLCKentered permeabilized cells more efficiently thannon-permeabilized spores. The inhibitory capac-ity of TAME or TLCK was not influenced byincreasing the concentration of L-alanine, indi-cating that these compounds did not competewith L-alanine for the site of the trigger reaction.Whereas TLCK is an irreversible inhibitor of
trypsin, its effect on germination is reversible.When spores which have been preincubatedwith 1 to 5 mM TLCK are washed, they regaintheir normal germination capability. This revers-ible inhibition of the putative spore protease isdiscussed later.Effectof various inhibitors on germination
properties of spores. The germination propertiesof spores differ when inhibited by various com-pounds (Fig. 4). DPA was excreted during ger-mination in the presence of all inhibitors tested,but the amount varied from 10%o in spores ex-posed to TLCK to about 65% in spores exposedto TAME (Fig. 4C).The loss of heat resistance in spore suspen-
sions exposed under germination conditions alsowas influenced by inhibitors. With leupeptinunder germination conditions, between 60 and90% of heat resistance was lost, whereas 40%oloss of heat resistance occurred in the presenceof TLCK, and none occurred in the presence ofTAME (Fig. 4).During germination, spores usually lose re-
fractility, i.e., they turn from "phase bright" to''phase dark" under phase-contrast microscopy.When incubated with leupeptin under germina-tion conditions, they show an intermediaryform: 85% of them turn phase grey, and the restbecome phase dark. When germinating sporesare exposed to TAME or TLCK, about 15%become phase grey, and the rest stay phasebright. Analogous phase grey germinated sporeshave been studied and are described in theliterature (1, 9, 30).
Direct demonstration of trypsin-like proteasesin spores. To investigate directly the presence oftrypsin-like proteases in spores, the hydrolyticactivities on synthetic trypsin substrates wastested with intact and disintegrated spores.Qualitative results on the hydrolysis of TAMEafter incubation with germinated spores are giv-
J. BACTERIOL.
0-
0-
TIME, min
C D
1.6-80 i1.2 6
-~~~~~5
3 ~~~0
0.42
-10-5 05 10 15 2025 234 567 89[TAME I, mM tTLCK], mM
FIG. 3. Quantitative effect of synthetic trypsin-likeenzyme inhibitors on germination of permeable andnonpermeable spores ofB. cereus T. Germination wasperformed as described in the text. (A) Germinationwith and without TAME. 0, Control without germina-tion medium (water only); 0, germination mediumwithout TAME. Germination medium with TAME: A,0.8 mM; A, 25 mM. Spores were made permeable asdescribed in the text. O.D., Optical density. (B) Ger-mination with and without TLCK. 0, Control withoutgermination buffer (water only); 0, germination medi-um without TLCK. Germination medium with TLCK:A, 0.1 mM; A, 1 mM. Spores used were nonperme-able. (C) Dixon plot for TAME inhibition of germina-tion. Data were obtained from experiments similar tothose described in (A). Germination rate constants ofthe first 20 min were plotted reciprocally as a functionof TAME concentration. 0, Experiments performedon permeable spores (see text); 0, experiments per-formed on nontreated spores. Ki values of 4 and 12mM, respectively, were determined. k, Definition. (D)Dixon plots for TLCK inhibition of germination. See(C). Experiments were performed on permeable (0)and nonpermeable (0) spores. Ki values of 0.25 and1.25 mM, respectively, were recorded.
en above (Fig. 2C). Since we could not quantita-tively measure the rate of TAME hydrolysis, asynthetic trypsin substrate (BANA), whichyields a colorimetric product, was also investi-gated. Neurath et al. (15) demonstrated thattrypsin hydrolyzes BANA at a much slower ratethan TAME. Since resting dormant spores haveno effect on either BANA or TAME hydrolysis,spores were either germinated or germinated
on June 22, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

INHIBITION OF B. CEREUS T SPORE GERMINATION
0-L-o (.
0.4_(I) (I)
:O 0._ ,
0j
100
90
80
70
60
50
40
a b c a b cA B
a b cc
a b cD
a b c
FIG. 4. Effect on trypsin inhibitors on various spore germination characteristics. Various spores propertieswere measured during gennination of 2 x 108 spores per ml as described in the text. a, Optical density at 660 nm;b, heat resistance; c, DPA release. (A) Control without germination medium. (B) Control with germinationmedium. (C) Germination medium containing 30 mM TAME. (D) Germination medium containing 10 mMTLCK. (E) Germination medium containing 40 mM leupeptin.
and mechanically disruptured before assay. AsBANA itself inhibited germination either totallyor for prolonged periods, the following proce-dure was employed. To 2.5 ml of variouslytreated germinated spore suspension, 1 to 2 mML-BANA was added. As shown in Fig. 5, aprotease activity capable of hydrolyzing BANAcan be seen in these crude germinated sporesuspensions. Figure SA shows the time course ofBANA hydrolysis catalyzed by an enzyme pres-ent in germinated spores as well as in mechani-cally disrupted spores. The BANA hydrolysis isslow and continuous for at least 48 h of incuba-tion of 37°C. Most of the enzyme is retained inthe partially broken spores and is not leached tothe supematant (Fig. 5B). With either BANA(Fig. SB) or TAME (Fig. SC) as substrates, thepresence of chloramphenicol under a variety ofconditions had little effect on the hydrolyticactivity. Dormant spores did not hydrolyzeTAME (Fig. SCa) or BANA. Thus, the proteaseis cryptic in dormant spores. Germination ren-
ders this enzyme available to substrates, e.g.,TAME (Fig. 5Cc). When excess TAME was
used, germination was blocked, and little hydro-lysis of TAME was observed (Fig. 2C).
Since the sulfhydryl reagents N-ethylmalei-mide and dithionitrobenzoic acid and a metalchelator (EDTA) had no effect on the hydrolysisofTAME by germinating spores (Fig. SC), therewas no evidence for a metal or an -SH grouprequirement for activity.
If the BANA-hydrolyzing trypsin-like enzymewere identical to the TAME-hydrolyzing en-
zymes and were involved in germination, onewould expect its activity to be sensitive to thesame inhibitors of trypsin-like protease whichprevented germination. To test this relationship,spores were mechanically disintegrated (as de-scribed in Materials and Methods), and the rateof BANA hydrolysis was measured (Fig. SD).As can be seen, TLCK, TAME, and leupeptininhibited BANA hydrolysis. Because spore con-centrations were about 10 times higher than inthe germination experiments and the sporeswere only partially disintegrated, somewhathigher concentrations of inhibitors were used.For instance, with 25 mM TAME, 50% inhibi-tion of protease activity occurred when mea-sured by BANA hydrolysis, whereas the samedegree of inhibition of germination occurred atabout 13 mM.
DISCUSSIONIn this paper, experimental evidence is given
supporting the hypothesis that a proteolytic(most likely a trypsin-like) enzyme is involved inthe initial stages of germination. This hypothesisis based on three different experimental ap-proaches: (i) several different kinds of inhibitorsof trypsin-like proteases inhibit germination; (ii)germinating, but not dormant, spores hydrolyzea synthetic trypsin substrate (TAME); and (iii)disintegrated spores contain a trypsin-like pro-teolytic enzyme, as indicated by the hydrolysisof BANA. By these approaches, we have estab-lished that inhibitors of trypsin-like enzymesblock germination at a stage at which some heat
VOL. 153, 1983 705
on June 22, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

706 BOSCHWITZ ET AL.
E
0cJ
1-
zb
Incubation, hours D
B
t10
0)
z 20 a-C
a b c d e f ab c abc abc
FIG. 5. Tests for the presence of trypsin-like enzyme in B. cereus T spores. (A) Kinetics of BANAhydrolysis. Spores (80 mg per 4 ml of water; 5 x 1010 spores) were heat treated at 70°C for 45 min and thengerminated in 50 ml of 0.1 M Tris-hydrochloride buffer (pH 7.1) containing 50 mg of L-alanine, 25 mg ofadenosine, and 3 mM EDTA for 30 min at 37°C. The spores were then washed in Tris buffer, suspended to 5 x109 spores per ml, and incubated a further 60 min at 37°C in the presence of 1 mg of lysozyme per ml. Half of thespores were then incubated with 1 mM BANA and 1% DMSO (0). The second half was treated with a Braunrotator (see the text) and incubated with BANA as described above (0). Hydrolysis of BANA at variousincubation times was estimated from the optical density (O.D.) at 420 nm. All systems contained 500 ,ug ofchloramphenicol per ml, added with the BANA. (B) Effect of chloramphenicol on the spore protease. Sporeswere heat shocked and germinated as in (A), but half the quantity was used. The other half was germinated [as in(A)] in the presence of chloramphenicol (100 p.g/ml). After germination, the spores were washed in 0.1 M Trisbuffer (pH 7.1) containing 500 ,ug of chloramphenicol per ml, and part of the spores were disrupted by means of aFrench press at 500 atm. French press-treated or untreated spores were then incubated for 24 h at 37°C withBANA as described in (A). a, Spores, French Press treated; b, supernatant of a; c, spores, germinated andFrench press treated and in the presence of 100 ,ug of chloramphenicol per ml; d, supernatant of c; e, germinatedspores; f, spores germinated in the presence of 100 ,ug of chloramphenicol per ml. In all assays, 500 ,ug ofchloramphenicol per ml was added with the BANA to prevent bacterial growth. A unit of protease was defined asan optical density of 0.25 at 420 nm after 24 h of incubation. (C) Hydrolysis of TAME by germinated spores.Spore suspension (80 mg) in 2 ml of water (5 x 1010 spores) was heat treated as in (A). Germination and TAMEhydrolysis were performed at 109 spores per ml in 30 mM Tris-hydrochloride (pH 7.1), 1 mg of L-alanine per ml,0.5 mg of adenosine per ml, 10 mM TAME, and reagents as indicated. After S h of incubation at 37°C, sampleswere removed for chromatographic separation (see text). The figure shows tracings of the cellulose plateobtained after Sakaguchi spray (see text). a, Dormant spores in water; b, 2 SLg of trypsin per ml; c, germinatedspores; d to g, spores germinated in the presence of 100 ,ug of chloramphenicol per ml, 5 mM N-ethylmaleimide, 5
mM dithionitrobenzoic acid, or 5 mM EDTA. (D) Inhibition of BANA hydrolysis by trypsin-like proteaseinhibitors. Broken spores (final volume, 2.5 ml) (see text), equivalent to 5 x 109 spores, were incubated for 25 hat 37°C with 5 ,umol of BANA in 0.1 M phosphate buffer (pH 7.0) and 2% DMSO. The hydrolysis of BANA wasdetermined at 420 nm, and 100o activity was 0.25 optical density units at 25 h of incubation time. Theconcentrations of protease inhibitors tested were as follows: leupeptin, a, 40 mM; b, 12 mM; c, 5 mM; TAME, a,25 mM; b, 10 mM; c, 3 mM; TLCK, a, 10 mM; b, 3 mM; c, 1 mM.
resistance and DPA has been lost, but precedingtotal loss of refractility. Antipain and leupeptinare reversible inhibitors of germination. TAME,which has a high affinity for trypsin, competi-tively inhibits hydrolysis of substrates of loweraffinity and also inhibits germination. The inhibi-tion by TAME is temporary due to its hydrolysisby germinating spores. Similar observations hadbeen made with TAME as an inhibitor of clottingoffibrinogen by thrombin, leading to the conclu-sion that thrombin was a protease capable ofhydrolyzing TAME (22).TLCK, a known inhibitor of trypsin-like pro-
teolytic enzymes, is a reversible inhibitor ofgermination, in contrast to its irreversible inhibi-tion of trypsin. This remarkable difference re-quires further discussion. One possibility is thatTLCK recognizes a trypsin-like active site anddisplaces the natural spore substrate involved ingermination. Possibly the active site lacks thereactive nucleophile in proper distance and ori-entation to react with the chloromethyl group toform an irreversible alkyl enzyme derivative.Alternatively, germination could be dependenton a trypsin-like activated enzyme. In the pres-ence of TLCK, activation is inhibited; upon
0 0 $; -TAMEI-;~~~~~~T
a b c d e f g
J. BACTERIOL.
on June 22, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

INHIBITION OF B. CEREUS T SPORE GERMINATION 707
TLCK removal, activation of residual tryspin-like activity leads to activation of germinationand an apparent reversible inhibition.The experiments with TAME (a trypsin inhibi-
tor) suggest that germination leads to activationof a proteolytic enzyme. This enzyme seems tobe essential for normal germination because aslong as it is inhibited by higher concentrations ofTAME, germination is inhibited. As germinationproceeds, TAME is hydrolyzed and inhibition ofgermination is relieved. Since experiments withchloramphenicol demonstrate that the TAME-hydrolyzing enzyme is not synthesized duringgermination, we assume that it is activated dur-ing this process, or as spores germinate, TAMEbecomes increasingly accessible to an intracellu-lar enzyme. We have further shown that thisenzyme is neither a metal-requiring nor an SH-requiring enzyme. Although the nature of theputative trypsin-like enzyme will require its iso-lation, its properties resemble those of the en-zyme isolated by Srivastava and Aronson (24)from sporulating cells ofB. subtilis. The fact thatthis enzyme is not inhibited in its ability tohydrolyze TAME by EDTA or by inhibitors ofSH groups, as well as the finding that it isinhibited by antipain and leupeptide but notchymostatin or elastinal, is consistent with theassumption that it is a trypsin-like enzyme.Our data suggest that the inhibitors we used
are not acting at the level of the trigger reaction,since the release of some (but not all) DPAoccurs in spores exposed to all the differentinhibitors under conditions of germination.Also, high levels of germinants do not competefor the site of the trigger reaction with theinhibitors. Furthermore, some (although notcomplete) inhibition of changes in refractilityoccur with all inhibitors. There is no directcorrelation in the different experiments betweenloss of heat resistance and loss of DPA. This isin accordance with the findings of Hanson et al.(7), recently repeated by Dring and Gould (3),that DPA-negative mutants of B. cereus T maystill be as heat resistant as the wild type.
All our inhibitors of trypsin-like proteasesprevented the total loss of refractility, leading tovarious proportions of phase grey spores. Thesephase grey spores phenotypically resemble sev-eral germination mutants of B. cereus and B.subtilis (1, 28). Such germination mutants fre-quently lose their heat resistance and producespores interrupted during germination before thestage of complete excretion of hexosamine-con-taining cortex fragments (30). In at least onecase, such mutation was related to a defectiveprotease (1) which may be responsible for defec-tive coat processing, which in turn leads tointerrupted germination. Still, a direct involve-ment of these proteases in the germination proc-
ess cannot be eliminated. It is possible thatactivation of proteases during germination ofdormant bacterial spores could remove an inhib-itor of germination or activate a key enzymerequired for completion of germination. Othermechanisms might also be responsible for inter-rupted germination at the stage of phase greyspores (30). The mechanism of the trypsin-likeprotease action in germination of spores remainsto be elucidated.
ACKNOWLEDGMENTSThis research was supported by a grant from the Fritz
Thyssen Foundation to A.K., Public Health Service grant A110610 from the National Institutes of Health to H.O.H.,Public Health Service grant CA 16060 from the NationalInstitutes of Health to W.T., and grants from the National Airand Space Administration and the Foundation for Microbiolo-gy to the Microbial Ecology Program at the Marine BiologicalLaboratory, Woods Hole, Mass.The authors thank M. Yudkin, N. Sandler, and H. Levinson
for stimulating discussion, criticism, and advice.LITERATURE CITED
1. Cheng, Y. E., P. C. Ftz-James, and A. I. Aronson. 1978.Characterization of Bacillus cereus protease mutant de-fective in an early stage of spore germination. J. Bacteriol.133:336-344.
2. Cornish-Bowden, A. 1976. Principles of enzyme kinetics.Butterworth, London.
3. Dring, G. J., and G. W. Gould. 1981. DPA-negative heatresistant spore mutant of Hanson et al. Spore News.7:130-132.
4. Gallizzi, A., A. M. Albertini, M. L. Baldi, E. Ferrari, E.Isnenghl, and M. T. Zambelli. 1978. Genetic studies ofspore germination and outgrowth in Bacillus subtilis, p.150-157. In G. Chambliss and J. C. Vary (ed.), SporesVII. American Society for Microbiology, Washington,D.C.
5. Gollakota, K. G., and H. 0. Halvorson. 1960. Biochemicalchanges occurring during sporulation of Bacillus cereus:inhibition of sporulation by a-picolinic acid. J. Bacteriol.79:1-8.
6. Halmann, M., and A. Keynan. 1962. Stages in germinationof spores of Bacillus licheniformis. J. Bacteriol. 84:1187-1193.
7. Hanson, R. S., M. V. Curry, J. V. Garner, and H. 0.Halvorson. 1971. Mutants of Bacillus cereus strain thatproduce thermoresistant spores lacking dipicolinate andhave low level of calcium. Can. J. Microbiol. 18:1139-1143.
8. Harrell, W. K., and H. 0. Halvorson. 1955. Studies on therole of L-alanine in the germination of spores of Bacillusterminalis. J. Bacteriol. 69:275-279.
9. Hashlmoto, T., W. R. Frieben, and S. F. Contd. 1969.Germination of single bacterial spores. J. Bacteriol.98:1011-1020.
10. Janssen, F. W., A. J. Lund, and L. E. Anderson. 1958.Colorimetric assay for dipicolinic acid in bacterial spores.Science 127:26-27.
11. Keynan, A., and H. 0. Halvorson. 1962. Calcium dipico-linic acid-induced germination of Bacillus cereus spores.J. Bacteriol. 83:100-105.
12. Levinson, H. S. 1957. Non-oxidative enzymes of sporeextracts, p. 120-135. In H. 0. Halvorson (ed.), Spores.American Institute of Biological Sciences, Washington,D.C.
13. Levinson, H. S., and M. T. Hyatt. 1966. Sequence ofevents during Bacillus megaterium spore germination. J.Bacteriol. 91:1811-1818.
14. Means, G. J., and R. Feeney. 1971. Chemical modificationof proteins. Holden-Day Inc., San Francisco.
VOL. 153, 1983
on June 22, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

708 BOSCHWITZ ET AL.
15. Neurath, H., and G. W. Schwert. 1950. The mode ofaction of the crystalline pancreatic proteolytic enzymes.Chem. Rev. 46:69-153.
16. PFggot, P. J., A. Moir, and D. A. Smith. 1981. Advances inthe genetics of Bacilus subtilis differentiation, p. 29-39.In H. S. Levinson, A. L. Sonenshein, and D. J. Tipper(ed.), Sporulation and germination. American Society forMicrobiology, Washington, D.C.
17. Powell, E. 0. 1957. The appearance of bacterial sporesunder phase-contrast illumination. J. Appi. Bacteriol.70:342-348.
18. Quesnel, L. B., J. A. Owers, V. E. Farmer, and D. Coupes.1977. Subtilisin induced germination of Bacillus cereusPX spores and effects of dimethylsulphoxide, p. 753-770.In A. N. Barker, J. Wolf, D. J. Ellar, G. J. Dring, and G.W. Gould (ed.), Spore research. Academic Press, Inc.,London.
19. Rodenberg, S., W. Steaberg, J. PIer, K. Nickerson, J.Vay, R. Epsten, and H. 0. Halvormon. 1968. Relationshipbetween protein and ribonucleic acid synthesis duringoutgrowth of spores of B. cereus. J. Bacteriol. 96:492-500.
20. Ros1gnol, D. P., and J. C. Vary. 1977. A unique methodfor studying the initiation of Bacillus megaterium sporegermination. Biochem. Biophys. Res. Commun. 79:1098-1103.
21. Scott, I. R., G. S. A. B. Stewart, M. A. Koncewcz, D. J.Ellar, and A. Crafta-Lghty. 1978. Sequence of biochemi-cal events during germination of Bacillus megaterium
J. BACTERIOL.
spores, p. 95-103. In G. Chambliss and J. C. Vary (ed.),Spores VII. American Society for Microbiology, Wash-ington, D.C.
22. Sberry, S., and Troll, W. 1954. The action of thrombin onsynthetic substrates. J. Biol. Chem. 2018:95-105.
23. Serra, G. 1967. Germination of bacterial endospores withsubtilopeptidases. Can. J. Microbiol. 13:489-501.
24. Srlvasava, D. P., and A. I. Aronson. 1981. Isolation andcharacterization of a unique protease from sporulatingcells of Bacillus subtilis. Arch. Microbiol. 129:227-232.
25. Umeawa, H. 1976. Structure and activities of proteaseinhibitors of microbial origin. Methods Enzymol. 45:678-695.
26. Vary, J. C. 1978. Glucose-initiated germination in Bacillusmegaterium spores, p. 104-108. In G. Chambliss and J. C.Vary (ed.), Spores VII. American Society for Microbiolo-gy, Washington, D.C.
27. Vary, J. C., and H. 0. Halvorson. 1965. Kinetics ofgermination of Bacillus spores. J. Bacteriol. 89:1340-1347.
28. Vary, J. C., and A. Kornberg. 1970. Biochemical studiesof bacterial sporulation and germination. XXI. Tempera-ture-sensitive mutants for initiation of germination. J.Bacteriol. 101:327-329.
29. Wals, K. A. 1970. Trypsinogen and trypsins of variousspecies. Methods Enzymol. 19:41-44.
30. Warburg, R. J., and A. Moir. 1981. Properties of mutantof Bacillus subtilis 168 in which spore germination isblocked at a late stage. J. Gen. Microbiol. 124:243-253.
on June 22, 2018 by guesthttp://jb.asm
.org/D
ownloaded from