effect of color properties on the selection of oviposition site by ceratitis capitata
TRANSCRIPT

Entomol. exp. appl. 42: 187-193, 1986 187 �9 Dr W. Junk Publishers, Dordrecht - Printed in the Netherlands
Effect of color properties on the selection of ov ipos i t ion site by Ceratitis capitata
B. I. Katsoyannos, K. Panagiotidou & I. Kechagia University of Thessaloniki, Department of Agriculture, Laboratory of Applied Zoology and Parasitology, GR-540 06 Thessaloniki, Greece
Keywords: Ceratitis capitata, host selection, oviposition behavior, color, vision
Abstract
In laboratory experiments of the choice type, wild Mediterranean fruit flies, Ceratitis capitata (Wiedemann), originating from infested figs collected in the island of Chios, Greece, showed strong prefer- ence for oviposition in black, blue and red colored ceresin wax domes 18 mm in diameter. Green and orange domes were less preferred, while yellow and white domes received the least number of eggs when a preferred color was present. The observed preference for certain colors was found to depend on both the color hue and the intensity of the total reflected light (brightness), while the degree the colored domes contrasted with the background had little if any effect. The females visited the domes of the preferred colors most frequently, which indicates that some selection occurred before arrival on the domes.
Introduction
Similarly to other fruit flies (Diptera, Tephriti- dae), the Mediterranean fruit fly Ceratitis capitata, utilizes chemical and physical stimuli provided by suitable host fruits to locate and select them for oviposition (Feron, 1962; Sanders, 1962). Among physical stimuli, those perceptible by vision such as fruit shape, size and color as well as the degree of the fruit's contrast to the background, have been reported to play an important role in this process (F6ron, 1962; Sanders, 1968; Nakagawa et al., 1978; Cytrynowicz et aL, 1982; Marchini & Wood, 1983; Prokopy et al., 1984).
Laboratory assessment of female ovipositional response in relation to the color properties of the oviposition substrate has been attempted in some cases using artificially-reared flies (F~ron, 1962; Sanders, 1968). Surprisingly, in view of the well known genetic variability and behavioral plasticity of this species, no thorough study of the oviposi- tional response of C. capitata to fruit color proper- ties has been conducted using wild flies of known origin and host plant.
Here, we report on the effect of color properties such as hue (=dominant wavelength), brightness (=intensity of the perceived total reflected light) and contrast with the background of artificial oviposition substrates, using wild Mediterranean fruit flies.
Material and methods
The materials and methods employed in this study were identical with those of a similar study with the olive fruit fly, Dacus oleae (Gmelin) (Kat- soyannos et al., 1985). All experiments were con- ducted in the laboratory at 25~176 60_+5 r.h. and L14:D10 photoperiod. Light was provided by fluorescent tubes of the daylight type supplement- ed by natural light entering from two glass- windows. The maximum daytime light intensity ex- perienced by the flies, varied between 200 and 1000 lux, according to the outdoor conditions (cloudy, or sunny days).
Flies and homing conditions. The experiments were

188
conducted between October and December 1984 and 1985, using wild flies originating from infested figs collected each year, in late September on the is- land of Chios, eastern Greece. The emerged flies were held until testing in groups of ca. 50-100 in- dividuals in 3 0 x 3 0 x 3 0 cm cages, provided with water, food (a mixture of yeast hydrolysate, sugar and water in the proportions 1:5:5) and oviposition devices of the seven colors same to those used for the experiments.
Oviposition devices. All oviposition substrates were ceresin wax domes (Prokopy & Boiler, 1970), 18 mm in diameter. The method of preparation of the 7 hues used and their spectral reflectance curves (from 400 to 650 nm) are given by Katsoyannos et al., (1985). To investigate the effect of brightness, 4 neutral colors (N1 to N4) were made, ranking on brightness from black to white, by adding the ap- propriate quantity of uncolored (white) ceresin to a fixed quantity of black-colored ceresin. The N1 neutral color was almost black, the N4 neutral al- most white.
Experimental procedure. All experiments were of the choice-type and conducted inside 1 liter cylin- drical transparent plastic cages, of the same type described by Katsoyannos & Pittara (1983), using the same procedures as those employed by Kat- soyannos et al. (1985). There were 3 domes of each color in the 2-choice tests, and 7 to 8 domes, one of each color or neutral, in the multiple-choice tests. The domes were fixed in suitable holes cut in the lids of 9 cm diam. plastic, transparent petri dishes which served as bases for the cages. The cages were placed in a random order, and equidis- tant from each other on large benches covered with white, gray or black paper to provide the appropri- ate background contrast. This was visible inside the cage because of the transparency of the petri dishes used as cage bases. Females were tested individual- ly, usually at the beginning of their oviposition period.
The main criteria used for determining oviposi- tion preference were the number of eggs and the number of egg batches (=number of eggs per oviposition) laid in the domes. Counts were made daily after which the position of the cages on the benches was rerandomized. Counts in each repli- cate were completed when ca. 50 to 100 egg batches
had been deposited. In addition to the egg counts, direct observations of ovipositing females were made, lasting 2 0 - 30 rain each. During these obser- vations, the number of visits to the domes and the number of ovipositions were recorded.
For the analysis of egg count data from multiple- choice experiments the test of Wilcoxon & Wilcox (1964) was used. For data from 2-choice tests, the Wilcoxon's signed rank test was used, applied to a discrimination coefficient calculated for each repli- cate as described by Katsoyannos & Pittara (1983). For statistical comparison of the oviposition data of the multiple-choice tests with the spectrophoto- metric data, we adopted the same procedures as Katsoyannos et al. (1985). This was accomplished through the calculation of correlation coefficients, (a) between the total numbers of egg batches deposited in the domes of each color and the total reflectance of each color (from 400 to 650 nm), and (b) between the total number of egg batches deposited in each color and the relative reflectances at all 10 nm wavelength intervals of those colors. As relative reflectance was considered to be the amount of light reflected by the while domes be- interval (e.g. within 500 and 510 nm) relative to the total amount of light reflected by the given color. The total reflectance values of colored domes and neutrals were calculated as percentages of the total amount of light-reflected by the white domes be- tween 400 and 650 nm. These values were: White = 100; yellow = 55.9; orange = 40.3; green = 8.4; red = 13.3; blue = 12.0 and black = 1.8. The total reflectance values of the 4 neutrals from N1 to N4 were: 3.7, 11.0, 40.1 and 81.2 respectively.
Results
Under our conditions, C. capitata females deposited an average of 1.3 eggs per batch. The results were almost identical when considering ei- ther the total number of eggs laid or the number of egg batches. For simplicity and because we consider that oviposition preference is more precisely ex- pressed by egg batches, only the results concerning batch counts are given.
The combined results of the multiple-choice tests using all seven colors (Table 1) and 2-choice tests (Table 2), clearly show that the most preferred colors were black, blue and red. These were equally
189
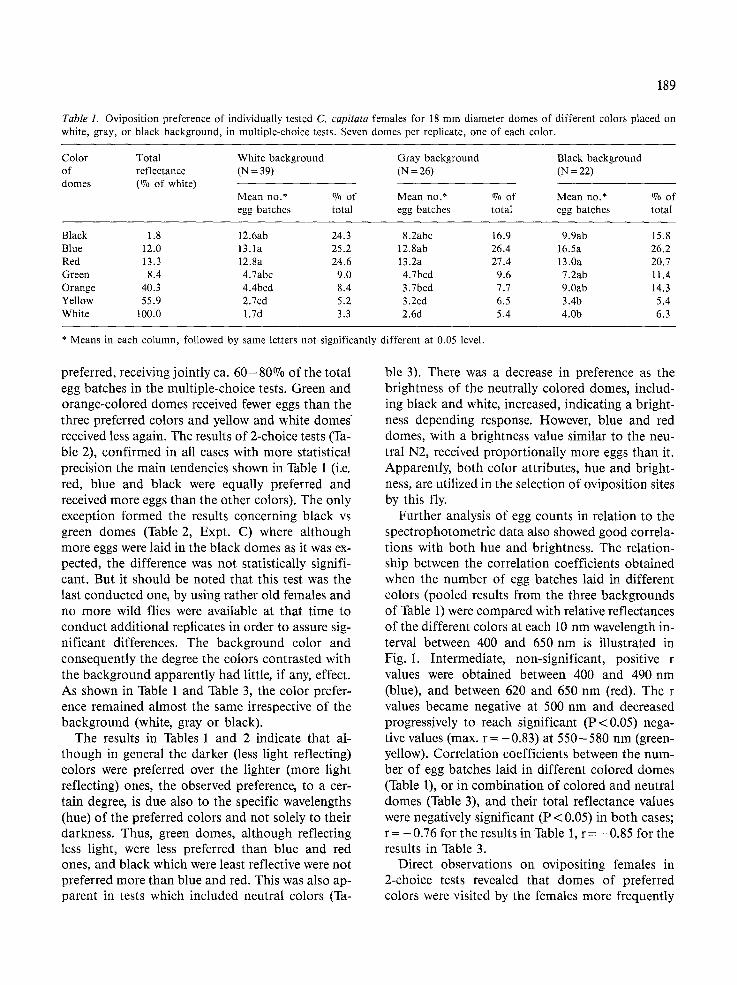
Table 1. Oviposition preference of individually tested C. capitata females for 18 mm diameter domes of different colors placed on white, gray, or black background, in multiple-choice tests. Seven domes per replicate, one of each color.
Color Total White background Gray background Black background of reflectance (N = 39) (N = 26) (N = 22) domes (% of white)
Mean no.* % of Mean no.* ~ of Mean no.* ~ of egg batches total egg batches total egg batches total
Black 1.8 12.6ab 24.3 8.2abc 16.9 9.9ab 15.8 Blue 12.0 13.1a 25.2 12.8ab 26.4 16.5a 26.2 Red 13.3 12.8a 24.6 13.2a 27.4 13.0a 20.7 Green 8.4 4.7abc 9.0 4.7bcd 9.6 7.2ab 11.4 Orange 40.3 4.4bcd 8.4 3.7bcd 7.7 9.0ab 14.3 Yellow 55.9 2.7cd 5.2 3.2cd 6.5 3.4b 5.4 White 100.0 1.7d 3.3 2.6d 5.4 4.0b 6.3
* Means in each column, followed by same letters not significantly
preferred, receiving jointly ca. 60-80~ of the total egg batches in the multiple-choice tests. Green and orange-colored domes received fewer eggs than the three preferred colors and yellow and white domes received less again. The results of 2-choice tests (Ta- ble 2), confirmed in all cases with more statistical precision the main tendencies shown in Table 1 (i.e. red, blue and black were equally preferred and received more eggs than the other colors). The only exception formed the results concerning black vs green domes (Table 2, Expt. C) where although more eggs were laid in the black domes as it was ex- pected, the difference was not statistically signifi- cant. But it should be noted that this test was the last conducted one, by using rather old females and no more wild flies were available at that time to conduct additional replicates in order to assure sig- nificant differences. The background color and consequently the degree the colors contrasted with the background apparently had little, if any, effect. As shown in Table 1 and Table 3, the color prefer- ence remained almost the same irrespective of the background (white, gray or black).
The results in Tables 1 and 2 indicate that al- though in general the darker (less light reflecting) colors were preferred over the lighter (more light reflecting) ones, the observed preference, to a cer- tain degree, is due also to the specific wavelengths (hue) of the preferred colors and not solely to their darkness. Thus, green domes, although reflecting less light, were less preferred than blue and red ones, and black which were least reflective were not preferred more than blue and red. This was also ap- parent in tests which included neutral colors (Ta-
different at 0.05 level.
ble 3). There was a decrease in preference as the brightness of the neutrally colored domes, includ- ing black and white, increased, indicating a bright- ness depending response. However, blue and red domes, with a brightness value similar to the neu- traI N2, received proportionally more eggs than it. Apparently, both color attributes, hue and bright- ness, are utilized in the selection of oviposition sites by this fly.
Further analysis of egg counts in relation to the spectrophotometric data also showed good correla- tions with both hue and brightness. The relation- ship between the correlation coefficients obtained when the number of egg batches laid in different colors (pooled results from the three backgrounds of Table 1) were compared with relative reflectances of the different colors at each 10 nm wavelength in- terval between 400 and 650 nm is illustrated in Fig. 1. Intermediate, non-significant, positive r values were obtained between 400 and 490 nm (blue), and between 620 and 650 nm (red). The r values became negative at 500 nm and decreased progressively to reach significant (P <0.05) nega- tive values (max. r= -0.83) at 550-580 nm (green- yellow). Correlation coefficients between the num- ber of egg batches laid in different colored domes (Table 1), or in combination of colored and neutral domes (Table 3), and their total reflectance values were negatively significant (P < 0.05) in both cases; r= -0 .76 for the results in Table 1, r= -0.85 for the results in Table 3.
Direct observations on ovipositing females in 2-choice tests revealed that domes of preferred colors were visited by the females more frequently

190
Table2. Oviposition preference of individually tested C. capitata females for 18 m m diameter domes of different colors placed on a white background, in 2-choice tests.
Color of No. repli- Mean no.* egg ~ of domes cates batches total
Expt. A Red 10 24.1a 42.7 Black 32.4a 57.3 Red 9 21.9a 42.0 Blue 30.2a 58.0 Red 10 40.0a 71.8 Green 15.7b 28.2 Red 7 45.9a 84.0 Orange 8.7b 16.0 Red 7 43.3a 84.6 Yellow 7.9b 16.3 Red 7 35.0a 72,7 White 13.1b 27,3 Expt. B Blue 10 28.6a 47.4 Black 31.7a 52.6 Blue 7 38.4a 74.3 Green 13.3b 25.7 Blue 7 37.6a 72.7 Orange 14. lb 27.3 Blue 7 46.7a 76.8 Yellow 14. lb 23.2 Blue 7 51.4a 95.2 White 2.6b 4.8 Expt. C Black 12 29.8a 57.2 Green 22.3a 42.8 Black 14 41.4a 74.6 Orange 14. lb 25.4 Black 7 46.6a 80.5 Yellow 11.3b 19.5 Black 7 42. la 84.0 White 8.0b 16.0
* Significance, see Table 1.
than the others (Table 4). These last results suggest that the females selected the preferred colors visual- ly before arriving on them. However, color effects after arrival on the domes cannot be excluded on the base of our experiments.
Discussion
In an earlier laboratory study on the oviposition behavior of C. capitata, F6ron (1962) concluded that differences in light intensity and quality may elicit variable response and the 'brilliancy' rather than the color of oviposition sites has the major in-
fluence on egg laying. On the other hand, Sanders (1968), reported that when females were given a choice between colored spheres against a monochromatic background, they preferred to oviposit in those contrasting most sharply against the background. However, when identical fruit models or real fruits were placed on both sides of a background divided in two halves (black and white or black and gray), the females preferred those placed in the darker part of the arena, ir- respective of the degree they contrasted to the back- ground. He concluded, that the color of the back- ground is more important than the degree of color-contrast. Both the aforementioned studies were conducted with flies of unknown origin, that had been laboratory reared for a large, unknown number of generations. In a more recent study (Marchini & Wood, 1983) colors were compared among two halves (upper and lower) of the same artificial fruit. It was found that yellow, green and orange were particularly favoured for oviposition with yellow being the most attractive. Black was much less favoured and red was the least attractive color of all. Unfortunately, details such as the flies' origin, holding conditions and experimental proce- dures are not given in their brief report, so it is not possible to make any detailed comparison with our results. Finally, Souza et al. (1984) reported that C. capitata females of a laboratory reared population preferred for oviposition red colored, 4 cm diam. agar spheres over green and yellow ones. Their results correspond to some of our findings. It is clear however, that there are some important differ- ences between our findings and most of those reported previously. These differences concern the order of color preference, as well as the relative im- portance of hue, brightness and color contrast.
A number of factors might be responsible for the differences between our results and those in the literature. One of them could be the different ex- perimental procedures, including the shape and size of the artificial oviposition substrates used. Howev- er, the main reason seems to be the origin and rear- ing conditions of the flies used (wild of known lo- cality and host plant vs artificially reared of unknown origin). In earlier studies, (B. I. Kat- soyannos and E. E Boller, unpubl.), considerable differences were observed in the color preferences of artificially reared and wild strains of C. eapitata using 18 mm ceresin wax domes similar those used

191
Table 3. Oviposition preference of individually tested C. capitata 9 9 for 18 m m diameter domes of black, blue, red, white and four neutral colors (N1 to N4) placed on white or black background, in multiple-choice tests, Darkness of neutrals decreases f rom N1 (al- most black) to N4 (almost white). Eight domes per replicate, one of each color.
Color Total A B Pooled A + B of reflectance White background Black background (N = 30) domes (070 of white) (N = 15) (N = 15
Mean no.* ~ of Mean no.* % of egg batches total egg batches total
Mean no.* ~ of egg batches total
Black 1.8 13.8a 20.1 10.5a 15.7 12.1a 17.9 Blue 12.0 11.5a 16.7 12.5a 18.7 12.0a 17,7 Red 13.3 13.9a 20.2 9. la 13.6 11.5ab 16.9 N1 3.7 12,8a 18.6 9.9a 14.9 11.4ab 16.8 N2 11.0 5.6ab 8.1 7.5a 11.2 6.5abc 9.7 N3 40.1 6,0ab 8.7 5.4a 8.1 5.7abc 8.4 N4 81.2 3.1b 4.6 7.0a 10.5 5.1bc 7.5 White 100.0 2. lb 3.0 4.9a 7.3 3.5c 5.1
* Significance, see Table 1.
O,
o
o.
o 5
0.4
~- 0.3 z
0 2
o.1
o o o
z - o . 1 o 7- - o . 2 <: _1 Lu - 0 3
O - 0 4 O
-0 5
- 0 6
-0 .7
- 0 . 8
- 0 9
-1 .0 4OO
8
7
6
. /
J l * l l l * l l I | l J J l t l i l l , l l t I
450 500 550 600 650
WAVELENGTH ( nrn }
Fig. 1. Correlation coefficients obtained when (a) number of egg batches laid in domes of each color in multiple choice tests (see Table 1) were compared with the (b) relative reflectance values of the colored domes wherein the eggs were laid, at each 10 n m interval from 400 to 650 nm.
in the present study. Whereas wild females from field-collected pupae in Costa Rica preferred or- ange colored domes over black ones for oviposi- tion, the reverse was true for F 1 flies of the same origin, reared on an artificial diet. A similar depen- dence of color preference on larval food of C. capitata has been indicated by F6ron (1962). B. I. Katsoyannos and B. Papadopoulou (unpubl.) also found that C. capitata flies of the same origin as those used in our experiments, reared for 1 to 3 generations on an artificial diet, showed a different color preference for oviposition sites than their wild ancestors. However, when subsequently reared on fruits (pears) for one generation, they again showed color preferences similar to their wild an- cestors.
As been pointed out in a recent review on the responses of fruit flies to shape, size and color (Katsoyannos, 1987), one important yet frequently neglected factor affecting female response to ovipo- sition sites, is the presence of oviposition-deterring pheromones. Such pheromones have been showed to be deposited on the substrate after oviposition by several fruit flies, including C. capitata (Prokopy et al., 1978). B. I. Katsoyannos & E. E Boller (un- publ.) have shown that C. capitata females deposit their oviposition-deterring pheromone also on the domes used as oviposition sites. Thus, in choice tests of the type we used, the real preference of fe- males could be partly masked by the deposition of

192
Table 4. Number of visits and ovipositions during 20- 30 min observations by individually cages C. capitata 9 Q in 2-choice tests. Three domes of each color per cage.
9 Color .of No. observed domes
Vis i t s Ovipositions
Black 13 1 White 6 0 Black 13 3 White 8 0 Black 17 3 White 10 0 Black, 13 2 Yellow 6 0 Black 21 2 Yellow 9 0 Black 24 10 Orange 7 1 Black 18 8 Orange 12 2 Orange 23 4 White 11 0
increasing amounts of pheromone on the initially preferred sites. As a result, less preferred sites may also receive a considerable number of eggs. Howev- er, in our experiments the initially preferred colors continued to be preferred for oviposition over the others, even when the number of eggs laid on them was very high (50-100 ovipositions) and, conse- quently, the concentration of the oviposition- deterring pheromone on them also high.
Nagakawa et al. (1978), reported that C. capitata flies were attracted to 7.5 cm diam spheres in the field, with black and yellow being the most attrac- tive of eight colors tested. They suggested that the flies were highly attracted to black spheres because these .stood out in stronger contrast against the background. Similar results were reported b y Cytrynowicz et al. (1982) and interpreted as a re- sponse of ready, to-oviposit females to ripening fruits. Finally, Katsoyannos (1986). working, in the same locality (Chios) and using the same trees (fig) as those from which the experimental flies used in the laboratory studies were collected as larvae, found that wild gravid females were strongly at- tracted to yellow, 7.0 cm diam spheres suspended on these trees. Orange, green, red and black spheres were less attractive than yelloW but equal to each
other, while blue and white were the least attractive. The response to the colored spheres was strongest during late afternoon and was highly correlated with observed feeding activity on ripe fruits. He concluded that the observed response to colored spheres, especially to yellow ones, was a feeding re- sponse to super normal ripe-fruit type stimuli and not a response to oviposition site type stimuli. This suggests, that any attempt to compare our labora- tory findings on oviposition behavior with the re- sponse to colored spheres observed in the field is probably risky.
The fact that C capitata attacks a large variety of ripening fruits, the colors of which are extremely variable, depending on fruit species and variety also makes it difficult to relate the observed color preference for the domes with the coloration of ripening host fruits. In nature, color stimuli of host fruits suitable for oviposition might be less impor- tant than other visual and/or chemical stimuli emitted by these fruits and thus the color prefer- ence found in the present study might be masked by them.
In a similar investigation on Dacus oleae, the sole fruit fly which was tested by using almost iden- tical to the present study experimental procedures and colors, a quite different color preference for oviposition sites was found (Katsoyannos et al., 1985). It is known however, that the compound eyes of both species, D. oleae and C. capitata, show similar spectral sensitivities, when measured elec- trophysiologically using electroretinograms, with a broad major peak of sensitivity at 485-500 nm and a secondary peak at 365 nm (Agee et al., 1982). The differences in the response found between the two species, apparently support the suggestion of Agee et al. (1982) that specific colors to which a species is most responding need not to correspond with the wavelengths to which the photoreceptors of that species are most sensitive, i.e., response may be controlled by higher order of neural input than the first order receptor level.
Acknowledgments
We thank Prof. M. E. Tzanakakis for comments. Supported in part by a grant from the Greek Ministery of Research and Technology, Program of Free Research 1984-85.

Zusammenfassung
Der Ein f luss yon Farbeigenschaf ten a u f die Aus - wahl von Eiablagesubstraten bei Cera t i t i s c a p i t a t a
In A u s w a h l v e r s u c h e n i m L a b o r ze ig ten wi lde
M i t t e l m e e r f r u c h t f l i e g e n , Ceratitis capitata (Wiede -
m a n n ) , die aus in fes t i e r t en Fe igen der Inse l Ch ios ,
G r i e c h e n l a n d , s t a m m t e n , e ine s tarke B e v o r z u g u n g
fiir schwarze, b l a u e u n d ro te Ce res in W a x d o m e m i t
18 m m Durchmesse r . G r t i n e u n d o r a n g e f a r b e n e
D o m e w a r e n wen ige r b e v o r z u g t , w/ ih rend ge lbe u n d
weisse D o m e m i t n u r w e n i g e n E i e r n be leg t w u r d e n ,
w e n n eine der b e v o r z u g t e n F a r b e n a n w e s e n d war.
D i e b e o b a c h t e t e B e v o r z u g u n g fi ir gewisse F a r b e n
h / ing t v o m F a r b t o n u n d v o m to t a l r e f l ek t i e r t en
L i c h t (He l l igke i t ) ab, j e d o c h ha t t e die St~trke des
Kon t ra s t e s der f a r b i g e n D o m e v o m U n t e r g r u n d we-
n ig o d e r ke inen E in f luss . D ie W e i b c h e n b e s u c h t e n
6 f t e r die D o m e m i t b e v o r z u g t e n Fa rben , was dar -
a u f h i n d e u t e t , dass sic sic vo r ihrer A n k u n f t aus-
w/ihl ten.
References
Agee, H. R., E. Boller, U. Remund, J. C. Davis & D. L. Cham- bers, 1982. Spectral sensitivities and visual attraetant studies on the Mediterranean fruit fly, Ceratitis capttata (Wiedemann), olive fly, Dacus oleae (Gmelin), and the Euro- pean cherry fruit fly, Rhagoletis eerasi (L.) (Diptera, Tephriti- dae). Z. ang. Entomol. 93: 403-412,
Cytrynowicz, M., J. S. Morgante & H. M. L. de Souza, 1982. Visual responses of South American fruit flies, Anastrepha fratereulus, and Mediterranean fruit flies, Ceratitis eapitata, to colored rectangles and spheres. Environ. Entomol. 11: 1202-1210.
F6ron, M., 1962. Uinstinct de reproduction chez la mouche m6diterran6enne des fruits Ceratitis capitata Wied. (Dipt., Trypetidae). Comportement sexuel. Comportement de ponte. Rev. Pathol. V6g. Entomol. Agric. France 41: 1-129.
Katsoyannos. B. I., 1986. Field responses of Mediterranean fruit flies to colored spheres suspended on fig, citrus and olive trees. Proc. 6th Int. Syrup. Insect-Plant Relationships, Pau,
193
France, 1-5 July 1986. Dr. W. Junk Publishers, Dordrecht, (In press).
Katsoyannos, B. I., 1987. Response to shape, size and color. In: A. S. Robinson & H. S. Hooper (eds), Fruit Flies: Their Biol- ogy, Natural Enemies and Control Elsevier Sc. Publ., Am- sterdam. (In press).
Katsoyannos, B. I. & I. S. Pittara, 1983. Effect of size of artifi- cial oviposition substrates and presence of natural host fruits on the selection of oviposition site by Dacus oleae. Entomol. exp. appl. 34: 326-332.
Katsoyannos, B. I., G. Patsouras & M. Vrekoussi, 1985. Effect of color hue and brightness of artificial oviposition substrates on the selection of oviposition site by Dacus oleae. Entomol. exp. appl. 38: 205-214.
Marchini, L. & R. J. Wood, 1983. Laboratory studies on ovipo- sition and on the structure of the ovipositor in the Mediterra- nean fruit fly Ceratitis capitata (Wied.). In: R. Cavalloro (ed.), Fruit Flies of Economic Importance. Proc. of the CEC/IOBC Int. Syrup., Athens, Nov. 1982. A. A. Balkema, Rotterdam: 113.
Nakagawa, S., R. J. Prokopy, T. T. Y. Wong, J. R. Ziegler, S. M. Mitchell, T. Urago & E. J. Harris, 1978. Visual orientation of Ceratitis capitata flies to fruit models. Entomol. exp. appl. 24: 193-198.
Prokopy, R. J. & E. E Boller, 1970. Artificial egging system for the European cherry fruit fly. J. Econ. Entomol. 63: 1413-1417.
Prokopy, R. J., J. R. Ziegler & T. T. Y. Wong, 1978. Deterrence of repeated oviposition by fruit-marking pheromone in Cera- titis capitata (Diptera: Tephritidae). J. Chem. Ecol. 4: 55- 63.
Prokopy, R. J., P. T. McDonald & T. T. Y. Wong, 1984. Inter- population variation among Ceratitis capitata flies in host ac- ceptance pattern. Entomol. exp. appl. 35: 65-69.
Sanders, W., 1962. Das Verhalten der Mittelmeerfruchtfliege Ceratitis capitata Wied. bei der Eiablage. Z. Tierpsychol. 19: 1-28.
Sanders, W., 1968. Die Eiablagehandlung der Mittelmeerfruch- fliege Ceratitis capitata Wied. Ihre Abh/ingigkeit von Farbe und Gliederung des Umfeldes. Z. Tierpsychol. 25: 588-607.
Souza, H. M. L. de, O. H. O. Pavan & I. D. Silva, 1984. Ovipo- sition and alightment behavior of Ceratitis capitata (Diptera, Tephritidae) on colored spheres. Revta bras. Entomol. 28: 11-14.
Wilcoxon, E & R. A. Wilcox, 1964. Some Rapid Approximate Statistical Procedures. Lederle Laboratories, Pearl River, New York.
Accepted May24, 1986,