effect of color hue and brightness of artificial oviposition substrates on the selection of...
TRANSCRIPT

Effect of color hue and brightness of artificial oviposit ion substrates on the selection of oviposit ion site by Dacus oleae
B. I. Katsoyannos, G. Patsouras & M. Vrekoussi University of Thessaloniki, Department of Agriculture, Laboratory of Applied Zoology and Parasitology, 54006 Thessaloniki, Greece
Keywords: Dacus oleae, host selection, oviposition behavior, color, vision
Abstract
In laboratory experiments of the choice type, olive fruit flies, Dacus oleae (Gmelin), wild and /o r olive- reared for a few generations, showed strong preference for oviposition in yellow and orange colored ceresin wax domes 18 mm in diameter. Green colored domes of two different hues were less preferred, while red, blue, black and white domes received a limited number of eggs when a preferred color was present. The females more frequently visited the domes of the preferred colors, which indicates that they selected them before arrival. The observed preference for certain colors like yellow and orange depended primarily on the color hue and not on the intensity of the total reflected light or on the degree to which the colored domes contrasted with the background. The females responded positively to hues reflecting maximally between 560 and 610 nm, opt imum at ca. 580 nm. Hues reflecting maximally between 520 and 550 nm or above 610 nm were not very attractive while hues reflecting below 520 nm and especially between 400 and 480 nm appeared to be rather unattractive. There were indications that not only the spectral purity of a hue but also the quantity of the energy reflected within the narrow spectral region which corresponds to that hue are important for the selection of the oviposition site by this fly.
Introduction
It is well documented that for a large number of phytophagous insects, including many frugivorous fruit flies (Diptera, Tephritidae), physical and chem- ical stimuli emitted by host and non-host plants and their fruits are of great importance for host selec- tion and host acceptance (Prokopy, 1977).
In order to better understand the host selection behavior of the olive fruit fly, Dacus oIeae which is the most important olive pest in the Mediterranean region, we have been investigating the influence of various physical and chemical stimuli on the selec- tion of the oviposition site, the olive fruit. Part of the investigation was dedicated to examining the effect of the size of artificial fruits and the effect of the presence of natural host fruits upon this behav- ior (Katsoyannos & Pittara, 1983). It was found
that D. oleae females readily accepted as oviposi- tion substrates orange colored ceresin wax domes with diameters ranging f rom 12 mm to 100 ram, preferring those of 35 mm.
In addition to shape and size, the color properties of the fruit are important in the selection of fruits suitable for oviposition by some fruit flies (Pro- kopy, 1977; Prokopy & Owens, 1983). The re- sponses of D. oleae flies to colored objects in gener- al, in most cases to sticky coated rectangles, have been investigated in the field (Orphanides & Soul- tanopoulos, 1962; Girolami & Cavalloro, 1973; P rokopy & Economopoulos, 1975; Prokopy et al., 1975; Prokopy & Haniotakis, 1975, 1976). It was found that the flies responded positively to particu- lar colors and were attracted most strongly to yel- low colored rectangles. P rokopy et al. (1975) de- monstrated that the field responses of the flies to
Entomol. exp. appl. 38,205-214 (1985). �9 Dr W. Junk Publishers, Dordrecht. Printed in the Netherlands.

206
rectangles of certain colors was a positive attraction due primarily to the hue and not to the intensity of reflected light. Only limited and contradictory in- formation exists concerning the role of fruit color in the location and acceptance of the oviposition site by D. oleae. Thus, Sacantanis (1953) reported that the females showed no differences in oviposition preference between olives of different coloration, while Orphanides et al. (1959) and Cirio (1971), found that the female prefers to oviposit in green rather than in dark colored olives. Finally, Prokopy & Haniotakis (1976), concluded on the base of cap- tures of flies on sticky-coated wooden olive models of different colors that colorper se of olive s, and /or the degree of contrast against the background, are cues utilized by the flies in locating olives.
A color is usually defined by three attributes: hue i.e. dominant wavelength of reflected light, satura- tion (chroma or tint) i.e. spectral purity of reflected
light, and brightness (value, intensity, shade) i.e. intensity of perceived reflected light.
In the present study we report on the effect of color hue and brightness of artificial oviposition substrates on the preference for oviposition by D. oleae, and discuss some findings on the effect of color-contrast against the background. We also give some information on whether this preference is expressed before arrival on the substrate, after arri- val, or both.
Materials and Methods
All experiments were conducted in the laborato- ry at 25 ~ _+ 2~ 60 + 5% r,h. and L14 : D10 photoperiod, with the photophase between 06:00 and 20: 00. Light was provided by nine fluorescent tubes of the daylight type, which during most of the
LU 0 Z < p-
UJ ,-I U. UJ p,,
100
50 Ye I I o w ~ ~
/ / O r a n g e /
/ / < . , u o / / /
J,= / / ._-2-/<,,vo-o,~176 / 0 ~ - ' ~ ~ - - ~ '. "5. :' I " , ' - - t - 400 450 500 550 600 650
WAVELENGTH (nm)
Fig. 1. S p e c t r a l r e f l e c t a n c e c u r v e s o f c o l o r e d d o m e s .

207
photophase were supplemented by natural light en- tering from two glass-windows on the eastern and southern sides of the room. The light intensity ex- perienced by the flies, varied between 200 and 1000 lux, according to the outdoor conditions (cloudy, or sunny days). In the morning hours, during which most of the oviposition activity was observed, the experimental cages were protected from direct ex- posure to sunlight, although we have observed that females exposed directly to the sun showed the same preferences in selection to oviposition devices.
Flies, and homing conditions. A first series of experiments was:conducted between January and March 1982, using olive-reared flies (i.e. flies origi- nating f rom field-collected pupae in the area of Thessaloniki and subsequently reared in the labora- tory for 1-6 (mostly 3) generations on olives). The same series of experiments was repeated and ex-
panded between January and March 1984 using wild flies (i.e. flies that emerged f rom pupae from field-infested olives collected in the area of Thessa- loniki). The emerged flies were held until testing in groups of ca. 50-100 individuals of both sexes, in 30 • 30 • 30 cm cages, provided with water and food (Katsoyannos & Pittara, 1983). In some cases they also had access to oviposition devices.
Oviposition devices. All oviposition substrates were ceresin wax domes (Prokopy & B o l l e r , 1970), 18 mm in diameter. This size was chosen instead of the preferred 35 mm domes because it more closely corresponds to the size of natural host fruits, is more convenient for handling, is space saving, and when the females had no choice they accepted it well (Katsoyannos & Pittara, 1983).
To make domes of different hues, candle-wax dyes (from Wernle Drugs, Zurich, Switzerland, ex-
LU (D Z
f - t.) LU . 3 U. UJ e~
100
50
I I I I
White
f N8 J ~
j
N7
/ i ~ N 6 J J
N5
N4
0 4 0 0
I
- N 2
Y
450 500 550 600
W A V E L E N G T H ( n m )
Fig. 2. Spectral reflectance curve of black, white, and the eight neutrally colored domes (N 1 N8).
650

208
Fig. 3. Set-up of eight domes on lid of plastic petri dish which served as base of cage (= one replicate), with orange (O), yellow (Y), white (W) and five different neutral colored domes (N4-N8), as in E• B, Table 3.
cept brown and blue which were bought in the local market) were dissolved in 100 ml molten ceresin as follows: 0.4 g black dye to make black; 0.2 g yellow, orange, green, red and blue dyes to make the cor- responding hues; and 0.1 g yellow + 0.02 g brown + 0.015 g blue to make a hue called 'olive-green' mim- icking to some extent the color of green olives. Uncolored ceresin gave white domes. To investigate the effect of color brightness, a series of eight neu- tral colors (N1 to N8) was made, ranking in bright- ness f rom black to white, by adding the appropriate quantity of uncolored ceresin to a fixed quantity of black ceresin. The N1 neutral color was almost black, the N8 neutral almost white. Reflectance spectra (400 to 650 nm) of all colors (Fig. 1) and neutrals (Fig. 2), were measured as a percentage of the reflectance from a white magnesium carbonate standard, in a Bausch and Lomb 505 spectropho- tometer.
Exper imental procedure. All experiments were of the choice-type and conducted inside 1 1 cylindrical plastic cages, the same as those described by Kat- soyannos & Pit tara (1983). There were three domes of each color in the 2-choice tests, and eight domes, one of each color, in the multiple-choice tests. The domes were fixed in suitable holes cut in the lids of
9 cm diam. plastic petri dishes which served as a base for the cage; they were positioned in random order and equidistant from one another (Fig. 3). One female per cage (= one replicate), was tested, usually at the beginning of her oviposition period. The cages were placed on a large bench covered with white paper, which provided a whitish back- ground.
The main criterion used for determining oviposi- tion preference was the number of eggs laid in the domes. Counts of eggs were made daily and the position of the cages on the bench was re-random- ized daily. Egg counts in each replicate were com- pleted when ca. 50 to 100 eggs had been deposited.
In addition to the egg counts, direct observations of ovipositing females were made, lasting 20-30 min each. During these observations, the number of visits to the domes was recorded, as well as the number of ovipositions.
For the analysis of egg count data of multiple- choice experiments the test of Wilcoxon & Wilcox (1964) was used. For data of 2-choice experiments, the Wilcoxon's signed rank test was used, applied to a discrimination coefficient calculated for each rep- licate as described by Katsoyannos & Pittara (1983).
For statistical comparison of the oviposition da- ta with the spectrophotometric data in the cases where such comparisons were possible (i.e. multi- ple-choice tests), we adopted similar procedures to those used by Greany et al. (1977) and Cytrynowicz et al. (1982) for the analysis of capture data of fruit flies on colored objects. This was accomplished through the calculation of correlation coefficients, a) between the total numbers of eggs deposited in the domes of each color and the total reflectance of each color (from 400 to 650 nm), and b) between the total numbers of eggs deposited in each color and the relative reflectances at various wavelength in- tervals of these colors. As relative reflectance was considered the amount of light reflected within a given wavelength interval (e.g. within 570 and 580 nm) relative to the total amount of light reflected by the color. For further explanations on the differen- ces between total reflectance and relative reflec- tance, values, as well as on the rationale of this approach see Cytrynowicz et al. (1982).
For each color, relative reflectances were ob- tained by relating the amount of light reflected within a sequence of 10 nm wide wavelength inter-
209
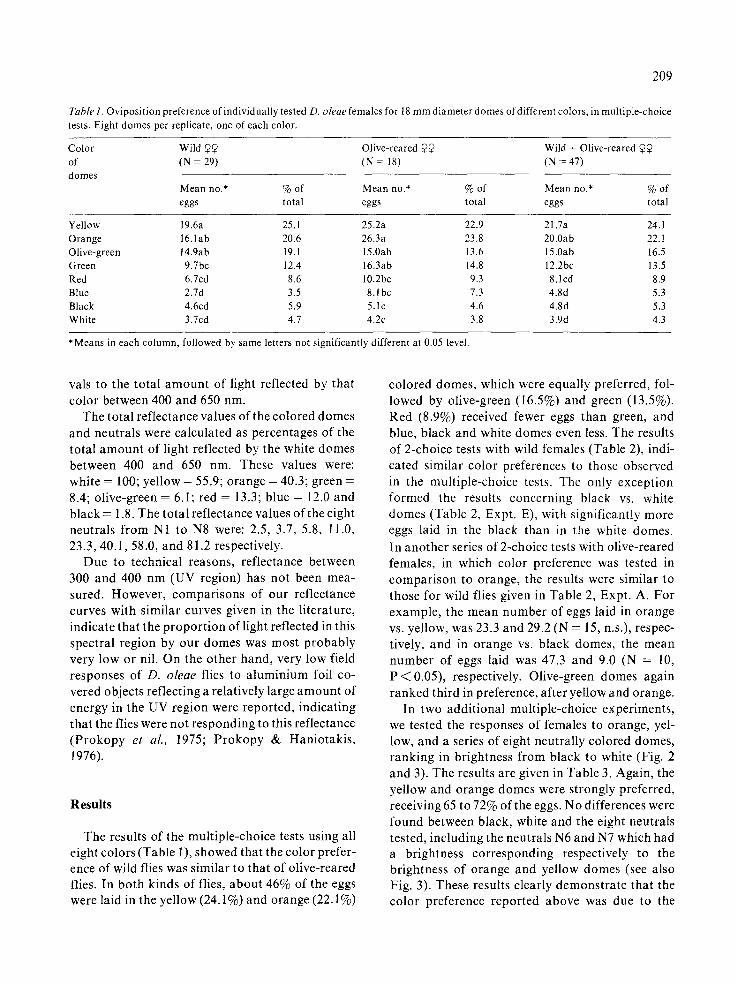
Table 1. Oviposition preference of individually tested D. oleae females for 18 mm diameter domes of different colors, in multiple-choice tests. Eight domes per replicate, one of each color.
Color Wild 99 Olive-reared Q9 Wild + Olive-reared 99 of (N = 29) (N = 18) (N - 47) domes
Mean no.* % of Mean no.* % of Mean no.* % of eggs total eggs total eggs total
Yellow 19.6a 25.1 25.2a 22.9 21.7a 24. l Orange 16. Iab 20.6 26.3a 23.8 20.0ab 22.1 Olive-green 14.9ab 19.1 15.0ab 13.6 15.0ab 16.5 Green 9.7bc 12.4 16.3ab 14.8 12.2bc 13.5 Red 6.7cd 8.6 10.2bc 9.3 8.1 cd 8.9 Blue 2.7d 3.5 8.1bc 7.3 4.8d 5.3 Black 4.6cd 5.9 5.1c 4.6 4.8d 5.3 White 3.7cd 4.7 4.2c 3.8 3.9d 4.3
*Means in each column, followed by same letters not significantly different at 0.05 level.
vals to the total amoun t of light reflected by that color between 400 and 650 nm.
The total reflectance values of the colored domes and neutrals were calculated as percentages of the total amoun t of light reflected by the white domes between 400 and 650 nm. These values were: white = 100; yellow = 55.9; orange = 40.3; green = 8.4; olive-green = 6.1; red = 13.3; blue = 12.0 and black = 1.8. The total reflectance values of the eight neutrals f rom N1 to N8 were: 2.5, 3.7, 5.8, 11.0, 23.3, 40.1, 58.0, and 81.2 respectively.
Due to technical reasons, reflectance between 300 and 400 nm (UV region) has not been mea- sured. However , compar isons of our reflectance curves with similar curves given in the literature, indicate that the p ropor t ion of light reflected in this spectral region by our domes was most p robab ly very low or nil. On the other hand, very low field responses of D. oleae flies to a luminium foil co- vered objects reflecting a relatively large amoun t of energy in the UV region were reported, indicating that the flies were not responding to this reflectance ( P r o k o p y et al., 1975; P r o k o p y & Haniotakis , 1976).
Results
The results of the multiple-choice tests using all eight colors (Table 1), showed that the color prefer- ence of wild flies was similar to that of olive-reared flies. In both kinds of flies, about 46% of the eggs were laid in the yellow (24.1%) and orange (22.1%)
colored domes, which were equally preferred, fol- lowed by olive-green (16.5%) and green (13.5%). Red (8.9%) received fewer eggs than green, and blue, black and white domes even less. The results of 2-choice tests with wild females (Table 2), indi- cated similar color preferences to those observed in the multiple-choice tests. The only exception formed the results concerning black vs. white domes (Table 2, Expt. E), with significantly more eggs laid in the black than in the white domes. In another series of 2-choice tests with olive-reared females, in which color preference was tested in compar i son to orange, the results were similar to those for wild flies given in Table 2, Expt. A. For example, the mean number of eggs laid in orange vs. yellow, was 23.3 and 29.2 (N = 15, n.s.), respec- tively, and in orange vs. black domes, the mean number of eggs laid was 47.3 and 9.0 (N = 10, P < 0.05), respectively. Olive-green domes again ranked third in preference, after yellow and orange.
In two addit ional multiple-choice experiments, we tested the responses of females to orange, yel- low, and a series of eight neutrally colored domes, ranking in brightness f rom black to white (Fig. 2 and 3). The results are given in Table 3. Again, the yellow and orange domes were strongly preferred, receiving 65 to 72% of the eggs. No differences were found between black, white and the eight neutrals tested, including the neutrals N6 and N7 which had a brightness corresponding respectively to the brightness of orange and yellow domes (see also Fig. 3). These results clearly demonst ra te that the color preference reported above was due to the

210
Table 2. Oviposition preference of individually tested wild D. oleae c~ 9 for 18 mm diameter domes of different colors, in 2-choice tests.
Color of No. replic. Mean no.* % of domes eggs/replicate total
Expt. A Orange 10 29.3a 54.1 Yellow 24.9a 45.9 Orange 9 28.6a 54.8 Olive-green 23.6a 45.2 Orange 6 35.4a 60 .2 Green 19.5b 39.8 Orange 7 41.3a 80.7 Red 9.9b 19.3 Orange 6 48.2a 97.0 Blue 1.5b 3.0 Orange 6 41.8a 84.5 Black 7.7b 15.5 Orange 6 34.8a 87.8 White 4.8b 12.2
Expt. B Green 12 16.2a 35.3 Yellow 29.7b 64.7 Green 10 24.3a 47.2 Olive-green 27.2a 52.8 Green 6 41 .Sa 82.6 Red 8.8b 17.4 Green 6 35.5a 85.5 Blue 6.2b 14.5 Green 8 38.0a 70.0 Black 16.3b 30.0 Green 7 46.6a 86.2 White 7.4b 13.8
Expt. C Red 6 14.2a 27.9 Yellow 36.7b 72.1 Red 7 22.3a 49.2 Blue 23.0a 50.8 Red 8 23.1 a 51.4 Black 21.9a 48.6 Red 10 28.5a 56.5 White 21.9a 43.5
Expt. D Blue 10 21.4a 43.6 Black 27.7a 56.4 Blue 10 20.3a 43.8 White 26. I a 56.3
Expt. E White 12 16.3a 33.2 Black 32.9b 66.8
*Significance, see Table 1.
Table 3. Oviposition preference of individually tested D. oleae females for 18 mm diameter domes of orange, yellow, black, white, and a series of neutral colors (NI to NS) in two multiple-choice experiments of 22 replicates each. Darkness of neutrals decreases from N 1 (almost black) to N8 (almost white). Eight domes per replicate, one of each color.
Color of domes Mean no. eggs*/ % of total replicate
Expt. A Orange 18.2a 27.8 Yellow 29.0a 44.3 Black 3.3b 5.0 White 2.6b 4.0 Nl 3.4b 5.2 N2 4.0b 6.1 N3 3.1b 4.7 N4 1 .Sb 2.8
Expt. B Orange 21.8a 35.4 Yellow 18.5a 30.0 White 4.2b 6.9 N4 3.6b 5.9 N5 4.0b 6.4 N6 3.2b 5.2 N7 4.0b 6.4 N8 2.3b 3.7
*Significance, see Table 1.
specific wavelengths (hue) of the preferred colors and not to the specific amount of light (brightness) reflected by them. The possibility that the observed preference was influenced by the degree to which the preferred colors contrasted (on the basis of intensity differences) with the whitish background can be excluded because a) two of the non-preferred neutrals (N6 and N7) had the same brightness as orange and yellow, and b) the most contrasting colors, such as black and blue, as well as the least contrasting such as white, were among the least preferred ones.
The results of the multiple-choice tests given in Table 1 were further analysed in relation to the spectrophotometric data (pooled numbers of eggs from wild and olive-reared females; total eggs = 4445). Calculations of correlations coefficients be- tween the number of eggs laid in each color and relative reflectances were made for all 10 nm wave- length intervals of each color between 400 and 650 nm (Fig. 4). Positive r values were obtained above 520 nm. The positive coefficients increased progressively and reached significant values in the 560-610 nm region. Maximal positive r values were

211
1 . 0
0 , 9
0 . 8
0 . 7
0 . 6
0 . 5
0 , 4 I.- z ua 0 . 3 ~
- 0 . 2 u. u.. ua 0 . 1 O
0,0
z - - 0 . 1 O
i.- -0.2 ,r w - - 0 . 3 ev ew O - - 0 . 4
- - 0 . 5
- 0 . 6
- 0 , 7
- - 0 . 8
- - 0 . 9
- - 1 . 0
, I , I , m I f m J J i r , i ~ , I 1 o..o.O...q. I 4
P ( 0 , 0 1 : ~ _J P( 0.05 ~ - /
6
D
_ _ P( 0 . 0 5
P ( 0 . 0 1
i i i ] i h , , I , , , , [ , i i i i i
4 0 0 4 5 0 5 0 0 5 5 0 6 0 0
W A V E L E N G T H ( n m )
Fig.4. Correlation coefficients between (a) number of eggs laid in domes of each color in multiple-choice test of Table 1 and (b) relative reflectance values of colored domes wherein the eggs were laid at every 10 nm interval, from 400 to 650 nm. �9 r values when data of all eight colors were included. �9 Same, but excluding data of black and white. Probability levels refer to all eight colors.
m650
obtained between 560-590 nm, and the highest (r = 0.79), at ca. 580 nm. Significant negative r values were obtained below 480 nm. Because it is possible that the correlations observed were affected by the number of eggs laid in the black and the white domes, thus expressing differences between these versus the other colors (Cytrynowicz et al., 1982), we recalculated the correlation coefficients exclud- ing data of a) black, b) white and c) both black and white. When the data of only black or only white were excluded, the r values obtained were almost identical with those found when all colors were included in the calculations, throughout the wave- length interval sequence, as shown in Fig. 4. In the region of the spectrum where significant positive r values were obtained when all eight colors were
included in the calculations (560-610 nm), when the data of both black and white were excluded, the new r values obtained were even higher, reaching a maximum r = 0.96 at ca. 580 nm (Fig. 4, open circles). These results clearly show that the females responded positively to certain wavelengths, espe- cially to hues reflecting proportionally more energy between 560 and 610 nm, with peak reflection around 580 nm, and relatively little energy below 520 nm (yellow, orange). Hues reflecting most of their energy between 400 and 480 nm (blue) should, according to the r values obtained, evoke a negative female response. Finally, hues reflecting maximally above 610 nm (red) or between 520 and 560 nm (green), should evoke moderate positive responses. The behavioral data given in Tables 1 and 2 support these conclusions well, except that the negative re- sponse which should be evoked by blue is not clear. The high r values obtained within a relatively nar- row spectral region indicate that color hue purity, expressed as a high relative reflectance value in a given narrow wavelength interval is important for the selection of oviposition site by D. oIeae.
No correlation was found between the total re- flectance values of each color and the number of eggs laid in the corresponding domes during the multiple choice experiments (r -- 0.05). This is in accordance with the results of the choice tests using neutrals (Table 3) which showed that the brightness of the colors was not responsible for the preference observed. When, however, the data of black and white were excluded from the calculations, a posi- tive correlation coefficient value (r = 0.79) was obtained. This might indicate that not only hue purity but also the intensity of reflected light within the narrow spectral region corresponding to the preferred hue might be important for the selection of the oviposition site.
The equal preference for yellow and orange domes as shown in the tables, does not mean that all females tested laid equal numbers of eggs in both colors. In fact, in the 2-choice tests with orange vs. yellow domes, only 12 out of 25 individually tested females (pooled data from wild and olive-reared flies), (i.e. ca. 50%) laid almost equal numbers of eggs, (i.e. 50 _+ 5% of the eggs) in each color. From the remaining 13 females, 7 (i.e. ca. 25%) preferred the yellow and 6 (i.e. ca. 25%) the orange color. When a strongly preferred color was tested against a weekly preferred one, most of the females laid

212
Table 4. Number of visits and ovipositions during 20-30 min observations by individually caged D. oleae females in 2-choice tests. Three domes of each color per cage.
Female Color of No. observed domes
Visits Ovipositions
a Orange 28 1 Blue 14 0
b Orange 29 1 Blue 2 0
c Orange 11 4 Red 2 0
d Orange 13 1 Red 14 1
e Orange 32 7 Black 9 0
f Orange 28 2 White 2 0
more eggs in the preferred color, although in some cases a few females showed equal preference for either. Only in a few cases, females laid more eggs in the non-preferred color. Although the pooled data show a preference for a relatively wide spectral region, the responses of individual females indicate that in fact the individuals had a rather well devel- oped color discrimination ability.
Direct observations on ovipositing females in 2- choice tests showed a close correlation between the number of visits and the number of ovipositions in each color (Table 4). Colors receiving more eggs were considerably more frequently visited and vise versa. Usually, the females selected the domes by walking from one dome to another. Visits to non- preferred colors were in most cases very short, giv- ing the impression that the females were simply passing over them, either in their search for a pre- ferred substrate, or when abandoning a preferred one after an oviposition.
Discussion
According to the literature, the means by which frugivorous fruit flies detect individual fruits after arrival on a host plant is principally based on the physical characteristics of the fruit, such as shape and size, intensity of reflectance, and the degree to which the fruits contrast with the background (Prokopy, 1977; Prokopy & Owens, 1983 and refer- ences therein).
The present work demonstrates that D. oleae
females selected the artificial oviposition sites not only by shape and size as already shown (Prokopy & Haniotakis, 1976; Katsoyannos & Pittara, 1983), but also by their color properties. The observed preference for oviposition in domes of certain co- lors, especially yellow and orange, was apparently due to a positive response of the females to the specific wavelengths of these colors and not to the total amount of light reflected from them. This color preference was apparently not affected by the degree to which the colored domes contrasted with the background. The fact that in a 2-choice test the black domes were preferred over the white ones is not in disagreement with this conclusion because in an additional, similar test using a black back- ground, black domes were again preferred over white ones (B. I. Katsoyannos, unpubl.).
The results of direct observations on ovipositing females strongly suggest that the domes of the pre- ferred colors were visually selected before arrival. However, color effects after arrival on the domes cannot be excluded on the base of our experiments.
In a study on the stimuli affecting oviposition in Rhagole t i s cerasi L., Prokopy &Boller (1971) re- ported that yellow was poor in eliciting oviposition after arrival on domes but did, in some cases, elicit more arrivals than some other colors. They sug- gested that color properties have an influence on whether oviposition after arrival will be attempted or not. However, owing to essential differences in the methodology employed by these authors and by us, such as using groups of flies of both sexes versus individual females, their results cannot be com- pared with ours.
It has been reported that D. oleae flies were at- tracted in the field to colored rectangles, particular- ly toward yellowish hues embodying the combined properties of highest amount of light reflected be- tween 520 and 580 nm and little reflectance below 520 nm (Prokopy et aL, 1975). Our findings suggest that the color response to fruit mimics of gravid females searching for oviposition sites is essentially similar to the reported general response of D. oleae flies to colored flat surfaces.
Olives in the field are suitable for oviposition for a long period, during which the chemical and physi- cal fruit properties change considerably. During ripening, the color of olive fruit gradually changes from olive-green, which matches the color of upper

surfaces of olive leaves, to yellowish, reddish or violet and almost black, depending upon the par- ticular olive variety. Olives of all these colors have been found suitable for oviposition in laboratory tests (Sacantanis, 1953), although, in general, green olives were preferred over black ones (Orphanides et al., 1959; Cirio, 1971). The preference for green over black olives reported by these authors, howev- er, represents a comparison of two extreme situa- tions. Anecdotal records including personal obser- vations of ours (B. I. Katsoyannos, unpubl.), indicate that ripening olives which are yellowish, are the most suitable for oviposition, in laboratory tests and in the field. Thus, our results correspond well with what we suspect to be the behavior of the flies in the field.
In a field study, Prokopy & Haniotakis (1976), assessed the importance of olive fruit color in the mechanism of olive detection by the flies. They found that D. oleae females were about equally attracted to black, red, and yellow sticky-coated wooden olive models, slightly less attracted to green and orange models and least attracted to blue, white, and aluminium foil covered models. The apparent difference in the order of color preference between our results and those reported by these authors is not surprising, because recent, more ex- tensive field experiments on the responses of D. oleae to colored spheres show that female response to sticky-coated fruit mimics does not necessarily indicate fruit-searching behavior for oviposition (B. I. Katsoyannos, unpubl.). During these experi- ments only a very low proport ion of the more than 3000 females captured were likely to have re- sponded to oviposition stimuli.
Yellow or orange colored olives of the same or closely similar hues as those used in our experi- ments do not occur in nature. Thus, while the mod- erate preference for olive-green and green domes can be explained by a certain similarity of these with green olives, the strong preference for the yel- low and orange domes might indicate that these hues constitute super-normal stimuli related to the fruit-seeking behavior of gravid females. These sti- muli in combination with other attractive cues, might prove useful in controlling the olive fruit fly by means of attractive devices.
213
Acknowledgements
We thank J. Tsitsipis and S. Daoussis, Nuclear Research Center 'Democri tos ' , for obtaining the spectral reflectance curves, E. Boiler and U. Re- mund, Swiss Federal Research Station for Arbori- culture, Viticulture and Horticulture, Wadenswil, for providing ceresin wax and dyes, and M. E. Tzanakakis, University of Thessaloniki, for criti- cism.
Zusammenfassung
Der Einfluss von Farbtdnen und Farbintensiti~t kiinstlicher Eiablagesubstrate auf ihre Auswahl fiir die Eiablage bei Dacus oleae
In Auswahlversuchen im Labor bevorzugten wilde und ftir einige Generafionen im Labor auf Oliven geztichtete Olivenfliegenweibchen zur Eiab- lage gelb und orange gef~irbte Ceresinwachsdome yon 18 mm Durchmesser. Grtingef~irbte Dome von zwei verschiedenen Farbt6nen waren weniger be- vorzugt. Rote, blaue, schwarze und weisse Dome wurden mit einer begrenzten Anzahl Eiern belegt, wenn eine der bevorzugten Farben dabei war. Die Weibchen besuchten 6tier die Dome mit den bevor- zugten Farben, was darauf hindeutet, dass sie diese vor der Ankunft ausw~ihlten.
Die beobachtete Bevorzugung f~ir gewisse Far- ben wie gelb und orange hing hauptsgchlich von den Farbt6nen ab und nicht v o n d e r Intensitgt des total reflektierten Lichtes oder der St~trke des Kon- trastes der farbigen Dome vom Hintergrund. Die Weibchen reagierten positiv auf Farbt6ne mit einer maximalen Reflektion zwischen 560 und 610 nm mit einem Opt imum bei ca. 580 nm. Farbt~3ne, die maximal zwischen 520 und 550 nm oder bei mehr als 610 nm reflektierten, waren nicht sehr attraktiv und Farbt6ne mit einer Reflektion bei weniger als 520 nm, speziell zwischen400 und 480 nm, scheinen nicht at traktiv zu sein. Es bestehen Hinweise daftir, dass nicht nur die Farbreinheit sondern auch die Menge der reflektierten Energie des entsprechen- den Spektralabschnittes wichtig ft~r die Auswahl der Eiablagesubstrate ist.

214
References
Cirio, U., 1971. Reperti sul meccanismo stimolo-risposta nell' ovideposizione del Dacus oleae Gmelin (Diptera, Trypeti- dae). Redia 52: 577-600.
Cytrynowicz, M., J. S. Morgante & H. M. L. De Souza, 1982. Visual responses of South American fruit flies, Anastrepha fraterculus, and Mediterranean fruit flies, Ceratitis capitata, to colored rectangles and spheres. Environ. Entomol. l l: 1202 1210.
Girolami, V. & R. Cavalloro, 1973. Metodi cromotropici per indagini di popolazione degli adulti di Dacus oleae Gmelin. Note ed Appunti Sperimentali di Entomol. Agraria 14: 13-29.
Greany, P. D., H. R. Agee, A. K. Burditt & D. L. Chambers, 1977. Field studies on color preferences of the Caribbean fruit fly, Anastrepha suspensa (Diptera: Tephritidae). En- tomol, exp. appl. 21:63 70.
Katsoyannos, B. I. & I. S. Pittara, 1983. Effect of size of artificial oviposition substrates and presence of natural host fruits on the selection of oviposition site by Dacus oleae. Entomol. exp. appl. 34: 326-332.
Orphanides, P. S., E. A. Phytizas & A. A. Tsakmakis, 1959. Quelques observations sur l'intensit6 de l'attaque du Dacus, en fonction du degr6 de maturation de l'olive. Ann. Inst. Phytopath. Benaki, 2:144 148.
Orphanides, P. S. & C. D. Soultanopoulos, 1962. Some observa- tions concerning the influence of the colour and the number of traps per tree on the captures of adult flies of Dacus oleae Gruel. Ann. Inst. Phytopath. Benaki, 4:112-117.
Prokopy, R. J., 1972. Stimuli influencing trophic relations in Tephritidae. Colloq. Int. CNRS 265:305 336.
Prokopy, R. J. & E. F. Boiler, 1970. Artificial egging system for the European cherry fruit fly. J. Econ. Entomol. 63: 1413-7.
Prokopy, R. J. & E. F. Boiler, 1971. Stimulieliciting oviposition of European cherry fruit flies, Rhagoletis cerasi (Diptera: Tephritidae), into inanimate objects. Entomol. exp. appl. 14: 1-14.
Prokopy, R. J. & A. P. Economopoulos, 1975. Attraction of laboratory-cultured and wild Dacus oleae flies to sticky- coated McPhail traps of different colors and odors. Environ. Entomol. 4: 187-192.
Prokopy, R. J., A. P. Economopoulos & M. W. McFadden, 1975. Attraction of wild and laboratory-cultured Dacus oleae flies to small rectangles of different hues, shades, and tints. Entomol. exp. appl. 18:141 152.
Prokopy, R. J. & G. E. Haniotakis, 1975. Responses of wild and laboratory-cultured Dacus oleae to host plant color. Ann. Entomol. Soc. Am. 68: 73-77.
Prokopy, R. J. & G. E. Haniotakis, 1976. Host detection by wild and lab-cultured olive flies. Symp. Biol. Hung. 16:209 214.
Prokopy, R. J. & E. D. Owens, 1983. Visual detection of plants by herbivorous insects. Ann. Rev. Entomol. 28: 337-364.
Sacantanis, K., 1953. Facteurs dbterminant le comportement de Daeus oleae GmeI. vis-a-vis des vari6t6s d'oliviers. Rev. Path. V6g6t. Entomol. Agric. Fr. 32:50 57.
Wilcoxon, F. & R. A. Wilcox, 1964. Some rapid approximate statistical procedures. Lederle Laboratories, Pearl River, New York.
Accepted. February 7, 1985.