effect of aluminium on duodenal calcium transport in pregnant and lactating rats treated with...
TRANSCRIPT

JOURNAL OF
www.elsevier.com/locate/jinorgbio
Journal of Inorganic Biochemistry 101 (2007) 1270–1274
InorganicBiochemistry
Effect of aluminium on duodenal calcium transport in pregnantand lactating rats treated with bromocriptine
Daniel Orihuela *
Laboratorio de Investigaciones Fisiologicas Experimentales, Facultad de Bioquımica y Ciencias Biologicas,
Universidad Nacional del Litoral, Piso 4, Ciudad Universitaria, Paraje El Pozo, 3000 Santa Fe, Argentina
Received 7 April 2007; received in revised form 25 May 2007; accepted 5 June 2007Available online 12 June 2007
Abstract
The aim of present work was to study the effect of oral aluminium (Al) overload on intestinal calcium (Ca) absorption in the criticalstages of pregnancy and lactation of rats and to find out possible relationships with prolactin (PRL) and 17b-estradiol (E2) circulatinglevels. Adult female Wistar rats were orally treated from day 1 of pregnancy with 0 (control), or 50 mg elemental Al (as chloride)/kg bodyweight per day. Ca transport was determined by everted duodenal sacs technique using 2 lCi of 45CaCl2 as flux marker (JCams). Al treat-ment reduced JCams either in late pregnancy (day 19) or in middle lactation (day 9 postpartum). Oral administration of bromocriptine(BrC), an inhibitor of PRL secretion, at dose of 10 mg/kg body weight given 18 h before JCams measurements were done, produced asignificant decrease in the inhibitory effect of Al on JCams, expressed as percent of control, at day 9 of nursing (vehicle: 51 ± 7%,BrC: 28 ± 4%, P < 0.05). A positive correlation between Al effects on JCams and the physiological variations of E2 serum levels alongpregnancy and lactation in BrC-treated rats was also found (r2 = 0.277, P = 0.001). We conclude Al could reduce transcellular Caabsorption in the duodenum by interfering with physiological mechanisms of Ca transport partially mediated by serum level incrementsof E2 and PRL, observed in late pregnancy and mainly during middle lactation of rats.� 2007 Elsevier Inc. All rights reserved.
Keywords: Aluminium; Duodenal calcium transport; Bromocriptine; Prolactin; 17b-estradiol; Pregnancy; Lactation
1. Introduction
Even when aluminium (Al) toxicity was initially recog-nized as a neurological and skeletal disease which occurredin some dialysis populations, it has been demonstrated thatconcentrations of this element that are toxic to many bio-chemical processes are also found in a number of neurode-generative disorders affecting nonuremic and nondialyzedindividuals [1].
Al-containing antacids are widely used nonprescriptionmedications which have been administered for many yearsfor the treatment of various gastrointestinal disorders, par-ticularly peptic disorders, because of their acid-neutralizingand local protective properties [2]. It is now well established
0162-0134/$ - see front matter � 2007 Elsevier Inc. All rights reserved.
doi:10.1016/j.jinorgbio.2007.06.009
* Fax: +54 342 4595522.E-mail address: [email protected]
that small amounts of Al from these antacids are normallyabsorbed [3]. Especially relevant is the intake of largeamounts of Al by pregnant women. During pregnancy dys-pepsia is a common complaint and antacids are used toreduce the dyspectic symptoms, although it was suggestedthat high-dose antacids should not be consumed duringgestation [4,5].
Many studies have reported that Al reduces the vitaminD-dependent calbindin D-mediated transcellular Ca trans-port in the small intestine of humans, rodents and chickens[6–10].
On the other hand, during rat pregnancy occurs an earlyadaptive increase of intestinal calcium (Ca) absorptionto allow maternal skeleton accumulates Ca to meet thepeak of fetal demand by mid-pregnancy, and later in lacta-tion [11]. Several sexual hormones, including estradiol(E2) and prolactin (PRL) seem to be involved in this

D. Orihuela / Journal of Inorganic Biochemistry 101 (2007) 1270–1274 1271
physiological mechanism. A study performed in Wistar ratsdemonstrated that PRL may enhance intestinal calciumabsorption directly. This effect was absent in pregnant ratsbut was of physiological significance in lactating rats [12].During pregnancy or lactation estrogens have vitamin D-independent effects at the genomic level on active duodenalcalcium absorption mechanisms, mainly through a majorupregulation of the calcium influx channel CaT1. Theestrogen effects seem to be mediated solely by estrogenreceptor alpha [13].
We have previously showed that inhibitory effect of Alon intestinal calcium absorption is highly dependent onestrogen serum levels in virgin female rats. In ovariecto-mized female rats treated with estrogen the studies ofdose–response curves showed that the sensitivity to theeffect of Al on JCams was raised with increasing 17b-estra-diol serum levels, without changes in the maximumresponse [14].
Therefore, the goal of present work was to study theeffect of oral Al overload on intestinal Ca transport inthe critical stages of pregnancy and nursing of rats, andto examine possible relationships with PRL and E2 circu-lating levels.
2. Materials and methods
2.1. Chemicals
[45Ca]Cl2 (specific activity 20 mCi/mg) was obtainedfrom NEN Dupont (Boston, USA). Bromocriptine (2-bromo-a-ergocryptine) was obtained from Lab. Novartis(Argentina). AlCl3 Æ 6H2O was from Mallinckrodt (USA).Ultrapure type 1 deionised water with a conductivity<0.2 lS/cm2 (GT Lab., Rosario, Argentina) was employedto prepare solutions. All other chemicals were of analyticalgrade from Sigma Co. (St. Louis, USA). AlCl3 solution(0.6 M) was freshly prepared according to the standardprotocol proposed by Tapparo et al. [15].
2.2. Animals
Three-month old female nonmated previously Wistarrats, weighing 230 ± 10 g, were housed at controlled roomtemperature (22 ± 3 �C) on a 14 h light/10 h dark cycles.Rats were allowed to free access to balanced rodent chowcontaining �4 IU/g cholecalciferol, 1% Ca and 0.8%phosphorus.
Eight female rats per cage were mated with one provenreproducer male in a harem mode, thus the beginning ofgestation was determined by the presence of spermatozoain vaginal smears obtained daily from female rats [16].From that moment on, pregnant rats were separated in iso-lated cages. Delivery of rats occurred at day 22 of preg-nancy. Lactation period lasted 15 days.
The experimental procedures were conducted in accor-dance with international rules on the use of animals in tox-icological studies [17].
2.3. Experimental protocols
2.3.1. Al treatment
Rats were orally treated by gavage with 50 mg elementalAl (as chloride)/kg body weight per day, from day 1 ofpregnancy along pregnancy and postpartum period. Tak-ing into account mean body weight, 1 ml of 0.6 MAlCl3 Æ 6H2O freshly prepared solution in deionised water(autogenous pH � 4) was given to each rat per day. Ratsin the control groups received 1 ml of deionised water acid-ified with HCl (pH 4). This Al dose resembles the amountof Al people can consume in pharmaceutical products [9].The theoretical amount of Al absorbed from this dosethrough the gut of the rats could be estimated in �60 lgper day, taking as a reference that human intestinal Alabsorption is about 0.5% [3].
2.3.2. PRL-related experiments
Animals were orally administered (o.g.) with 10 mg/kgbody weight of bromocriptine (BrC) given as a single dose,18 h before duodenal Ca transport (JCams) measurementswere done. Respective controls received the drug vehicle(0.5 ml of normal saline via o.g.). BrC is a semisyntheticderivative of the ergot alkaloid family possessing dopamineagonist properties with a potent inhibitory effect on PRLsecretion [18,19].
2.3.3. E2-related experiments
E2 influence was analyzed by comparing duodenal Catransport at two moments when it would be expected thatE2 serum levels are quite different, in BrC-treated rats tominimize likely PRL influence. During pregnancy werecompared day 5 JCams value (low E2 level expected)against day 19 JCams value (high E2 level expected). In lac-tation it was compared day 2 postpartum JCams value (lowE2 level expected) against day 9 postpartum JCams value(high E2 level expected) [20,21].
Summarizing, animals were assigned to six main exper-imental groups each containing two subgroups (Al-treatedand respective control) with at least five rats per group,taking into account both the time at which JCams weredetermined and the physiological condition:
Group 1: Day 5 of pregnancy treated with BrCGroup 2: Day 19 of pregnancyGroup 3: Day 19 of pregnancy treated with BrCGroup 4: Day 2 postpartum treated with BrCGroup 5: Day 9 postpartumGroup 6: Day 9 postpartum treated with BrC
2.4. Hormonal levels
Serum concentrations of 17b-estradiol were determinedby radioimmunoassay (Diagnostic Systems Laboratories,Texas, USA). Blood samples collection were performedby heart puncture between 10:00–12:00 AM to avoid

1272 D. Orihuela / Journal of Inorganic Biochemistry 101 (2007) 1270–1274
circadian fluctuations. Sera were frozen at �20 �C untilmeasurements were done.
2.5. Intestinal Ca absorption measurements
Since it has been demonstrated that Al effect on intesti-nal Ca transport occurs upon mucosa-to-serosa Ca flux(JCams) only at the duodenal level [14,22], this parameterwas determined at the following times: day 5 of pregnancy,day 19 of pregnancy, day 2 of nursing and at day 9 of nurs-ing in the respective experimental groups, by an evertedin vitro duodenal sacs technique as previously described[14]. Briefly, rats were anesthetized with sodium pentobar-bital (50 mg/kg, i.p.), the small intestine was removed fol-lowing a midline abdominal incision and transection ofthe intestine at the pylorus and cecum. The proximal10 cm segment (duodenum) was excised, rinsed in ice-cold0.9% sodium chloride solution, everted with a blunt glassrod and ligated at one end. A blunt-end needle was insertedthrough a second loosely tied ligature at the other end and0.3 ml of serosal buffer, pregassed with 95% O2/5% CO2
and warmed at 37 �C, were instilled. The needle wasremoved, the second ligature was tightened and each sacwas transferred to an individual flask containing pregassedwarmed mucosal buffer. The elapsed time followingremoval was 10 min. Sacs were incubated for 45 min inwater bath at 37 �C and the mucosal solutions were contin-uously gassed with 95% O2/5% CO2. At the end of the incu-bation period the serosal fluid was recovered from theinside of the intestinal sacs by draining its contents. Onlysacs with 80% recovery or more were considered. Sacs wereopened by longitudinal incision, blotted with filter-paperand the wet weight determined. All filled sacs were weighedbefore and after incubation.
The composition of mucosal and serosal buffer was125 mM NaCl, 4 mM KCl, 10 mM glucose, 1 mM CaCl2,30 mM Tris–HCl (pH 7.2). 2 lCi [45Ca]Cl2 was added intothe mucosal buffer. The 45Ca contents of the final serosalmedium, and that of the initial and final mucosal fluid,were determined by sampling 100 ll. JCams was calculatedfrom the following equation, adapted from Adler et al. [22]:
JCams ¼ ½45Ca�sf � V sf=SA � W � t
where [45Ca]sf is the final serosal 45Ca concentration (in dis-integrations/min/ml), Vsf is the final serosal volume(=0.3 Æ (Wf �W)/(Wi �W), in ml), SA is the mean specificactivity ((SAi + SAf)/2) of the mucosal medium (in disinte-grations/min/lmol), W is the wet weight of empty intesti-nal sac (in g) and t is the incubation time (in h). Wi andWf are the initial and final filled sac weight (in g),respectively.
2.6. Radioactivity measurements
The activity of the radionuclide 45Ca, a pure b-emitter(t1/2 163 days, 0.257 MeV), was determined using a thinwindow GM detector coupled to a spectrometer (Alfa
Nuclear, Argentina). Liquid samples were arranged in5 cm diameter stainless steel compartments (1 ml final vol-ume). Measurements were performed in duplicate at 600 Voptimal voltage, with constant geometry at saturationthickness. The chemical quenching effect was corrected bysetting up in the instrument a quench curve constructedwith five standards prepared adding different amounts ofa quenching agent (carbon tetrachloride) to a 45Ca sampleof known absolute activity. Counting time for each samplewas set to maintain statistical error below 1% [9].
2.7. Statistical analysis
Results are presented as mean ± SEM. Statistical signif-icance of the differences between two groups was assessedby grouped Student’s t-test. One-way analysis of variancewith a post hoc Newman–Keuls test was used to determinesignificance of differences among more than two groups.
The inhibitory effect of Al on JCams was quantified aspercent of respective control applying the following for-mula: [(control value-Al-treated value)/control value] Æ 100.Linear regression analysis was performed by the leastsquares method. P values of 0.05 or less were consideredsignificant. Data were processed with a registered versionof the InStat/Prism 3.03 software (GraphPad SoftwareInc., San Diego, CA, USA).
3. Results
3.1. Al effect on JCams in rats treated with BrC
In Fig. 1, the influence of BrC administration in the Aleffect on duodenal JCams is depicted. In groups thatreceived vehicle, Al treatment significantly reduced JCams
both at day 19 of pregnancy and at day 9 of nursing ascompared to control. BrC treatment produced a significantincrement of JCams in Al-treated rats in both conditions(pregnancy: vehicle = 2.40 ± 0.15 vs. BrC = 2.90 ± 0.26lmol/h/g wet weight, P < 0.05; lactation: vehicle = 2.10 ±0.13 vs. BrC = 2.80 ± 0.21 lmol/h/g wet weight, P < 0.05).
Besides, BrC reduced the percentual inhibition of JCams
by Al on a greater extension in lactation (vehicle: 51 ± 7%vs. BrC: 28 ± 4%, P < 0.05) as compared to pregnancy(vehicle: 42 ± 8% vs. BrC: 31 ± 7%).
In control groups, BrC had no effect on JCams in latepregnancy, but significantly reduced the same parametermeasured during lactation (vehicle = 4.20 ± 0.25 vs. BrC =3.70 ± 0.20 lmol/h/g wet weight, P < 0.05).
3.2. Al effect on JCams in different stages of pregnancy and
nursing of the rats
Fig. 2 shows Al effects on JCams compared betweenearly (day 5) and late (day 19) pregnancy, in rats treatedwith BrC, to minimize the possible influence of PRL. Altreatment decreased JCams in both conditions. In Al-treated groups, JCams resulted to be lower at day 19 as
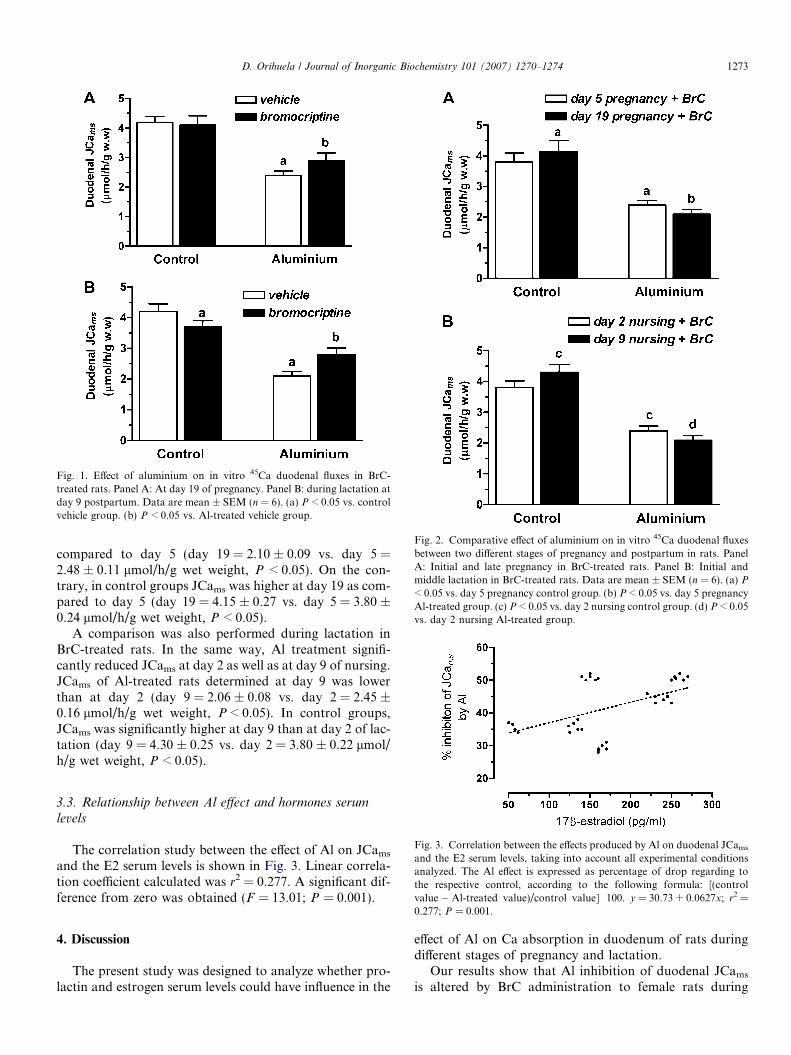
Fig. 1. Effect of aluminium on in vitro 45Ca duodenal fluxes in BrC-treated rats. Panel A: At day 19 of pregnancy. Panel B: during lactation atday 9 postpartum. Data are mean ± SEM (n = 6). (a) P < 0.05 vs. controlvehicle group. (b) P < 0.05 vs. Al-treated vehicle group.
Fig. 2. Comparative effect of aluminium on in vitro 45Ca duodenal fluxesbetween two different stages of pregnancy and postpartum in rats. PanelA: Initial and late pregnancy in BrC-treated rats. Panel B: Initial andmiddle lactation in BrC-treated rats. Data are mean ± SEM (n = 6). (a) P
< 0.05 vs. day 5 pregnancy control group. (b) P < 0.05 vs. day 5 pregnancyAl-treated group. (c) P < 0.05 vs. day 2 nursing control group. (d) P < 0.05vs. day 2 nursing Al-treated group.
D. Orihuela / Journal of Inorganic Biochemistry 101 (2007) 1270–1274 1273
compared to day 5 (day 19 = 2.10 ± 0.09 vs. day 5 =2.48 ± 0.11 lmol/h/g wet weight, P < 0.05). On the con-trary, in control groups JCams was higher at day 19 as com-pared to day 5 (day 19 = 4.15 ± 0.27 vs. day 5 = 3.80 ±0.24 lmol/h/g wet weight, P < 0.05).
A comparison was also performed during lactation inBrC-treated rats. In the same way, Al treatment signifi-cantly reduced JCams at day 2 as well as at day 9 of nursing.JCams of Al-treated rats determined at day 9 was lowerthan at day 2 (day 9 = 2.06 ± 0.08 vs. day 2 = 2.45 ±0.16 lmol/h/g wet weight, P < 0.05). In control groups,JCams was significantly higher at day 9 than at day 2 of lac-tation (day 9 = 4.30 ± 0.25 vs. day 2 = 3.80 ± 0.22 lmol/h/g wet weight, P < 0.05).
Fig. 3. Correlation between the effects produced by Al on duodenal JCams
and the E2 serum levels, taking into account all experimental conditionsanalyzed. The Al effect is expressed as percentage of drop regarding tothe respective control, according to the following formula: [(controlvalue � Al-treated value)/control value] Æ 100. y = 30.73 + 0.0627x; r2 =
3.3. Relationship between Al effect and hormones serum
levels
The correlation study between the effect of Al on JCams
and the E2 serum levels is shown in Fig. 3. Linear correla-tion coefficient calculated was r2 = 0.277. A significant dif-ference from zero was obtained (F = 13.01; P = 0.001).
0.277; P = 0.001.
4. Discussion
The present study was designed to analyze whether pro-lactin and estrogen serum levels could have influence in the
effect of Al on Ca absorption in duodenum of rats duringdifferent stages of pregnancy and lactation.
Our results show that Al inhibition of duodenal JCams
is altered by BrC administration to female rats during

1274 D. Orihuela / Journal of Inorganic Biochemistry 101 (2007) 1270–1274
pregnancy and lactation (Fig. 1). In rats with PRL levelslowered by BrC administration, the effect of Al on JCams
was attenuated. This finding could indicate that Al inter-feres, in some way, with PRL-mediated Ca transport induodenum. It has been postulated that duodenum is oneof the target organs to prolactin action. A strong receptorgene expression, a high level of PRL receptors and numberof binding sites have been reported in duodenum. It isknown that PRL enhances, even in the range of physiolog-ical concentration, the transcellular active Ca transport inthe mucosa-to-serosa direction. This effect is exerted stim-ulating Ca entry possibly via channel-mediated pathwayand Ca extrusion via Ca-ATPase-mediated pathway [23].Since these effects of PRL on Ca transport are dose-depen-dent, the fact that inhibitory effect of Al on JCams accom-panied the fluctuations of PRL levels in a wide range ofvalues obtained in different physiological situations[24,25], would demonstrate an interaction of Al with sucha mechanisms. A feasible way by which Al could interactwith Ca-ATPase-related PRL mechanism is blocking thestimulative effect of calmodulin (CaM) on plasma mem-brane Ca pump [26], since it is known that Al is able toinduce conformational changes in CaM, a key protein inCa signaling, reducing its affinity to targets [27].
Analyzing the comparative data of two stages of preg-nancy and lactation (Fig. 2), the positive correlation foundbetween Al effect on JCams and E2 levels (Fig. 3), and dis-carding the PRL influence due to BrC treatment, we caninfer that one of the major candidates to be responsiblefor the variations observed in Al effect is E2. In this regard,the results of present work would extend previous findingsabout the relationship between the effect of Al on duodenalCa transport and circulating E2 levels in virgin rats [14].
Al would be able to interact with E2 mechanisms ofaction on duodenal Ca transport throughout pregnancyand lactation.
Other factors should be considered in the effect of Al onJCams. It is known that intestinal Ca transport is primarilyregulated by calcitriol, the active metabolite of vitamin D[28]. However, because of all rats ingested the same dietwith a normal Ca and vitamin D content, we can assumethat vitamin D-related factors influenced in the same wayall experimental groups, so the differences encounteredcan be assigned only to hormones under study.
In summary, we present evidences that Al could reducetranscellular Ca absorption in the duodenum by interferingwith physiological mechanisms of Ca transport partiallymediated by serum level increments of E2 and PRL, observedin late pregnancy and mainly during middle lactation of rats.
5. Abbreviations
BrC bromocriptinePRL prolactin
E2 17b-estradiolJCams mucosa-to-serosa calcium fluxCaM calmodulin
Acknowledgement
This work was supported by a Grant C.A.I+D/2005-12/B419-PROG 020, Universidad Nacional del Litoral,Argentina.
References
[1] D.R. McLachlan, T.P. Kruck, M.F. VanBerkum, Am. J. Kidney Dis.6 (1985) 322–329.
[2] J.L. Domingo, Neurotoxicol. Teratol. 17 (1995) 515–521.[3] R. Weberg, A. Berstad, Eur. J. Clin. Invest. 16 (1986) 428–432.[4] R. Weberg, A. Berstad, B. Ladehaug, Y. Thomassen, Acta Pharma-
col. Toxicol. 59 (1986) 63–65.[5] S.K. Agarwal, L. Ayyash, C.S. Gourley, J. Levy, K. Faber, C.L.
Hughes, Fd. Chem. Toxic. 34 (1996) 49–53.[6] A.J. Adler, G.M. Berlyne, Am. J. Physiol. 249 (1985) G209–G213.[7] M.A. Dunn, N.E. Johnson, M.Y.B. Liew, E. Ross, J. Nutr. 123
(1993) 1786–1793.[8] K.A. Cox, M.A. Dunn, J. Nutr. 131 (2001) 2007–2013.[9] D. Orihuela, V. Meichtry, M. Pizarro, J. Inorg. Biochem. 99 (2005)
1879–1886.[10] D. Orihuela, V. Meichtry, N. Pregi, M. Pizarro, J. Inorg. Biochem. 99
(2005) 1871–1878.[11] C.S. Kovacs, H.M. Kronenberg, Endocr. Rev. 18 (1997) 832–872.[12] N. Krishnamra, R. Thumchai, L. Limlomwongse, Bone Miner. 11
(1990) 31–41.[13] S.J. Van Cromphaut, K. Rummens, I. Stockmans, E. Van Herck,
F.A. Dijcks, A.G. Ederveen, P. Carmeliet, J. Verhaeghe, R. Bouillon,G. Carmeliet, J. Bone Miner. Res. 18 (2003) 1725–1736.
[14] D. Orihuela, C.E. Carnovale, J.A. Monti, M.C. Carrillo, Toxicol.Lett. 85 (1996) 165–171.
[15] A. Tapparo, L. Solda, G.G. Bombi, P. Zambenedetti, P.F. Zatta, R.Bertani, B. Corain, Analyst 120 (1995) 2425–2429.
[16] W.C. Adams, E.H. Frieden, Biol. Reprod. 33 (1985) 1168–1175.[17] Guiding Principles in the Use of Animals in Toxicology, Society of
Toxicology, Virginia, USA, revised March, 1999.[18] J.W. Kebabian, D.B. Calne, Nature 277 (1979) 93–96.[19] D. Valente, M. Delaforge, S. Urien, D. Guivarc’h, R. Vienet, J.M.
Grognet, E. Ezan, J. Pharmacol. Exp. Therap. 282 (1997) 1418–1424.[20] A.A. Shaikh, Biol. Reprod. 5 (1971) 297–307.[21] B.N. Roy, K.E. Wynne-Edwards, Biol. Reprod. 52 (1995) 855–863.[22] A.J. Adler, C. Zara, G.M. Berlyne, Am. J. Physiol. 257 (1989) G433–
G437.[23] N. Charoenphandhu, L. Limlomwongse, N. Krishnamra, Can. J.
Physiol. Pharmacol. 79 (2001) 430–438.[24] T. Yohkaichiya, A. O’Connor, D.M. de Kretser, Biol. Reprod. 44
(1991) 6–12.[25] A. Boass, J.A. Lovdal, S.U. Toverud, Am. J. Physiol. 263 (1992)
G127–G134.[26] B.D. Slaughter, R.J. Urbauer, J.L. Urbauer, C.K. Johnson, Bio-
chemistry 46 (2007) 4045–4054.[27] H. Kurita, A. Nakatomi, H. Shimahara, M. Yazawa, S.Y. Ohki,
Biochem. Biophys. Res. Commun. 333 (2005) 1060–1065.[28] J.G.J. Hoenderop, B. Nilius, R.J.M. Bindels, Physiol. Rev. 85 (2005)
373–422.