early evolution of metazoan serine/threonine and tyrosine kinases
TRANSCRIPT

Early Evolution of Metazoan Serine/Threonine and Tyrosine Kinases: Identification of Selected Kinases in Marine Sponges
Michael C-use, Isabel M. Miiller, and Werner E. G. Miiller Institut fur Physiologische Chemie, Abteilung fur “Angewandte Molekularbiologie,” Universitat, Mainz, Germany
The phylum Porifera (sponges) was the first to diverge from the common ancestor of the Metazoa. In this study, six cDNAs coding for protein-serine/threonine kinases (PS/TKs) are presented; they have been isolated from libraries obtained from the demosponges Geodia cydonium and Suberites domuncula and from the calcareous sponge Sycon ruphanus. Sequence alignments of the catalytic domains revealed that two major families of PWTK, the “conven- tional” (Ca*+-dependent) protein kinase C (PKC), the cPKC subfamily, as well as the “novel” (Ca*+-independent) PKC (nPKC), form two separate clusters. In each cluster, the sequence from S. ruphunus diverges first. To approach the question about the origin of protein-tyrosine kinases (PTK), which are found only in Metazoa, we analyzed two additional PS/TKs which have been cloned from S. domunculu: the stress-responsive protein kinase (KRS_SD) and the protein-kinase-C-related kinase (PRICSD). The construction of the phylogenetic tree, comprising the eight PSFI’Ks and the PTK cloned previously from G. cydonium, revealed that the PTK derived from the branch including the KRS_SD kinase. These data facilitate the first molecular approach to elucidate the origin of metazoan PTK within the PWTK superfamily.
Introduction
It is characteristic of all organisms to recognize ex- tracellular signals, initiating intracellularly appropriate, adaptative biological responses. In unicellular and mul- ticellular animals, extracellular cues cause an activation of protein kinases which are involved in signal trans- duction pathways (Stabel and Parker 1991; Van der Geer, Hunter, and Lindberg 1994). Among protein ki- nases, two classes are predominant: the protein-serine/ threonine kinases (PS/TKs), which are phosphotransfer- ases with a protein alcohol group as acceptor, and the protein-tyrosine kinases (PTKs), which are phospho- transferases with a protein phenolic group as acceptor (Hunter 1991).
The PS/TK superfamily encompasses the AGC group kinases, which include the cyclic nucleotide-de- pendent family, the protein kinase C family, the p-ad- renergic receptor kinase family, and the ribosomal S6 kinase family (Hardie and Hanks 1995). The PTK group comprises the enzymes, which phosphorylate tryrosine residues only, and the dual-specific enzymes, which phosphorylate both tryrosine and serine/threonine. The first group of PTKs is further subdivided into two major groups, the receptor tyrosine kinases (RTKs), which are membrane-spanning molecules with similar overall structural topologies, and the nonreceptor TKs (Hardie and Hanks 1995).
The evolution of the protein kinases has not been studied in detail. The major reason for this is the fact that only a few amino acid (aa) sequences are known which allow conclusive phylogenetic analyses. Refering to animals, PS/TKs have been identified in vertebrates; in a limited number of invertebrate species, including the echinoderm Lytechinus pictus, Drosophila melano-
Key words: sponges, serine/threonine kinases, tyrosine kinases, phylogeny, molecular systematics, molecular evolution.
Address for correspondence and reprints: Werner E. G. Mtiller, Institut fur Physiologische Chemie, Abteilung fur “Angewandte Mo- lekularbiologie,” Universitat, Duesbergweg 6, D-55099 Mainz, Ger- many. E-mail: [email protected].
Mol. Biol. Evol. 14(12):1326-1334. 1997 0 1997 by the Society for Molecular Biology and Evolution. ISSN: 0737-4038
1326
gaster, Caenorhabditis elegans, and the sponge Geodia cydonium; and in the protozoan organism Trypanosoma brucei (reviewed in Kruse et al. 1996). These PS/TKs have been classified into the protein kinase C (PKC) family, which covers three subfamilies: the “conven- tional” (Ca 2+-dependent) PKC (cPKC) subfamily, the “novel” (Ca2+ -independent) protein kinase C (nPKC) subfamily, and the “atypical” PKC subfamily (Hardie and Hanks 1995).
Until now, PTKs, with the characteristic tyrosine kinase domain, were found only in metazoan (multicel- lular) organisms (Hardie and Hanks 1995); nom-elated tyrosine-phosphorylating enzymes have been identified in yeast (Hunter and Plowman 1997). The phylogenet- ically oldest member of PTK, an RTK, has been iden- tified in the sponge G. cydonium (Schacke et al. 1994a, 1994b, 1994~; Mtiller and Schacke 1996). The RTK from G. cydonium is a true receptor TK, as confirmed by immunochemical localization of the protein and by subcellular distribution in the sponge tissue (Skorokhod et al. 1997).
All of the known PS/TKs and the PTKs share a related catalytic domain of approximately 270 aa (Hanks, Quinn, and Hunter 1988), which is further sub- divided into 12 smaller subdomains. The consensus se- quences of the subdomains are highly similar in PS/T.Ks and PTKs (Hardie and Hanks 1995). Hence, based on the available sequence data, it can be concluded that the PTKs and the PS/TKs have a common ancestor.
In order to investigate the phylogenetic relationship of the PSflKs and PTKs, selected cDNAs encoding pro- tein kinases belonging to these superfamilies have been isolated from different sponge species. These animals have been used for our studies, because they belong to the oldest extant metazoan phylum. The phylum Porifera (sponges) is divided into (1) Demospongiae, (2) Calca- rea, and (3) Hexactinellida (Bergquist 1978). Member(s) of Demospongiae (G. cydonium and S. domuncula) and Calcarea (Sycon raphanus) have been used for our stud- ies. cDNAs of cPKC and nPKC have been identified from these three species. In addition, the cDNA of a
Downloaded from https://academic.oup.com/mbe/article-abstract/14/12/1326/1273002by gueston 21 March 2018

Evolution of Metazoan Protein Kinases 1327
PKC-related kinase (PRK) (Palmer, Ridden, and Parker 1995) was isolated from S. domunculu, and another one from the same species, belonging to the group of stress- responsive protein kinases (KRSs) (Creasy and Chernoff 1995), has been cloned. The new sequences are de- scribed. The major emphasis is put on a solution of the open questions about the phylogenetic relationships within these PS/TKs and about the origin of the meta- zoan PTK.
Materials and Methods Materials
Restriction endonucleases and other enzymes for recombinant DNA techniques and vectors were obtained from Stratagene (Heidelberg), Boehringer Mannheim (Mannheim), Epicentre Technologies (Madison, WISC), and USB (Cleveland, Ohio).
Sponges
Specimens of Geodia cydonium (Jameson) (Pori- fera, Demospongiae, Tetractinomorpha, Astrophorida, Geodiidae), Suberites domuncula (Olivi) (Porifera, De- mospongiae, Tetractinomorpha, Hadromerida, Suberiti- dae), and Sycon raphanus (Schmidt) (Porifera, Calcarea, Calcarona, Leucosoleniida, Sycettidae) were collected near Rovinj (Croatia). The material was immediately frozen in liquid nitrogen until use.
Preparation of cDNA Libraries
cDNA libraries of G. cydonium, S. domuncula, and S. ruphanus were prepared in lambda ZAP Express@@ (Stratagene). cDNA-inserts were integrated in ZAP Ex- press@’ between the EcoRI and Xho I restriction sites and packaged in vitro using MaxPlaxm Packaging Ex- tract (Epicentre Technologies). The resulting numbers of independent clones were 5.2 X lo6 for G. cydonium, 1.5 X lo7 for S. domuncula, and 8.7 X lo6 for S. raphanus.
Isolation of Protein-Ser/Thr Kinases
cDNAs coding for PS/TK were isolated from G. cydonium, S. domuncula, and S. raphanus cDNA li- braries by homology screening with digoxygenin- ll- dUTP-labeled DNA-probes (DIG random primed DNA labeling kit; Boehringer Mannheim). Screening of the G. cydonium library was performed by using a complete mouse PKCa (Rose-John, Dietrich, and Marks 1988) as a probe (Ausubel et al. 1995). Within the conserved regions of the kinase domains, the nt identities between the mouse cDNA and the sponge cDNAs are >75%. The kinase domains of the resulting two PKCs of G. cydonium (CPKC-GC, accession number X87683, and NPKC,GC, accession number X87684; Kruse et al. 1996) were subsequently used as probes to screen S. domuncula and S. ruphanus cDNA libraries. Screening of all libraries was performed under low-stringency hy- bridization conditions of plaque lifts from 3 X lo5 pfu on nitrocellulose. Filters were hybridized at 37°C over- night in 35% formamide, 5 X SSC, 0.02% SDS, 0.1% N-laurylsarcosine, and 1% blocking reagent (Boehringer Mannheim) containing 10 ng/ml of the DIG-labeled probes). Filters were washed twice in 2 X SSC, 0.1%
SDS (5 min, room temperature), followed by two ad- ditional washes in 0.1 X SSC, 0.1% SDS (15 min, 42°C). Positive clones were detected with an alkaline phosphatase conjugated anti-digoxygenin antibody us- ing BCIP/NBT as substrate (Blake et al. 1984). All cDNAs have been obtained from at least two different cDNA libraries, resulting in at least three independent clones each.
DNA sequencing was performed with an automatic DNA sequenator (Li-Cor 4000s).
Sequence Analysis
Prediction of sites and signatures was performed with programs available in PC/GENE ( 1995). Homolo- gy searches were done via the E-mail servers at the Eu- ropean Bioinformatics Institute, Hinxton Hall, U.K. ([email protected] and [email protected]) and the National Center for Biotechnology Information, National Insti- tutes of Health, Bethesda, Md. ([email protected]. gov). Phylogenetic trees were constructed on the basis of aa sequence alignment by neighbor-joining applying the “Neighbor” program from the PHYLIP package (Felsenstein 1993). The degree of support for internal branches was further assessed by bootstrapping (Felsen- stein 1993). The distance matrix was calculated as de- scribed (Dayhoff, Schwartz, and Orcutt 1978). The graphical output of the bootstrap figure was produced by the “Treeview” program (Roderic D. M. Page, Uni- versity of Glasgow, U.K.; HTTP://http://taxono- my.zoology.gla.ac.uWsoftware.html#Treeviewing). Mul- tiple alignment using the default options was performed with CLUSTAL W version 1.6 (Thompson, Higgins, and Gibson 1994), and the graphic presentation was composed with GeneDoc (Nicholas and Nicholas 1996).
Results Cloning of Sponge cDNAs Encoding cPKC Suberites domuncula
The cDNA clone CPKCSD, isolated from the cDNA library of S. domunculu, is 2,321 bp long. The open reading frame (ORF) consists of 2,019 bp (starting at nt 17) and encodes a protein of 673 aa; the deduced aa sequence is termed CPKC-SD. The characteristic sites of this kinase are indicated in figure 1. The PS/TK catalytic domain of CPKC-SD with its 12 subdomains shows all the hallmarks known from sequences of PS/TK isolated from higher metazoa (Hardie and Hanks 1995) (fig. 2). The GTPDYIAPE motif (marked in fig. 1) and the absence of a tyrosine between DFG and APE triplets are “indicators” that CPKCSD is a PS/TK (Hanks, Quinn, and Hunter 1988). These latter indicator sequences are present in all PS/TKs described in this paper.
Sycon raphanus
A cDNA (3,244 bp long) encoding a PS/TK was isolated from the cDNA library of S. raphanus and termed CPKCSR; the ORF of 2,268 bp predicts a 756- aa-long protein (CPKC,SR). The characteristics of CPKC-SR are marked in figure 1. Downloaded from https://academic.oup.com/mbe/article-abstract/14/12/1326/1273002
by gueston 21 March 2018

1328 Kruse et al.
CPKC_CC : CPKC_SD : CPKC_SR :
CPKC_CC : CPKC_fD : CPKC_SR :
CPKC_GC : CPKC_fD : CPKC_SR :
CPKC_GC : CPKC_SD : CPKC_SR :
CPKC_GC : CPKC_SD : CPKC_SR :
CPKC_GC : CPKC_SD : CPKC_SR :
CPKC_GC I CPKC_SD : CPKC_fR :
CPKC_GC : CPKC_SD : CPKC_SR :
__1..._111..,~___~~~~.., ++++++++++++++++++++++++++++++++++++ pseudosubstrate Zn-1
----__a_---- ------------ c2
G--6ITUCN ---HEPHSS DPRSGFNDO
SLKO#@E@J&DTUK@RTDRGUOi$EDED---RGSRGTPE&@D@j-@- --~~TH*RILR~EERTKLIE$FRKR$ILDKRKWT~~@I*- E-~&!NUP~HEGP~OSRRSTRSI~GSS~PT~DED&J&@~R@$JS
- #--c _.
j l _ RF si ATP
kinase _________
Sermhr signature
: 82 : 78 : 99
: 177 : 174 : 198
: 271 : 267 : 281
: 357 : 353 : 376
: 453 : 449 : 472
: 549 : 545 : 568
: 645 : 641 : 663
677 673 756
FIG. I.-Alignment of the aa sequences deduced from the cDNAs coding for PKC group C from G. cydonium (CPKC_GC, accession number X87683; Kruse et al. 1996), S. domuncula (CPKCSD), and S. ruphanus (CPKC_SR). The following sites are marked: pseudosubstrate sequence ( ---), two zinc fingers (+ + +), C2 domain (= = =), ATP binding region (OOO), PS/TK active-site kinase signature (***), and the indicator signature for PS/TK (-). The boundaries of the catalytic domain are indicated (#+t#). Residues conserved in all sequences are shown in inverted type; those present in two sequences are shaded.
Since both sequences contain the C2 region, which cPKC from the sponge G. cydonium (k-use et al. 1996). is believed to be involved in Ca*+ binding (Stabel and The catalytic domains, displaying the highest sequence Parker 1991), they are grouped to cPKC. similarity, were compared (fig. 2). A quantitave analysis
The alignment in figure 1 shows, in addition to revealed an aa sequence identity (similarity) between the these new PS/TKs, the sequence of a recently described domains in S. d~muncula and G. cydokium of 77%
CPKC_GC : CPKC_SO : CPKC_SR : NPKC_CC : NPKC_SD : NPKC_SR :
CPKC_GC : CPKCfD : CPKC_SR : YPKC_CC : NPKC_SD : NPKC_SR :
CPKC_GC : CPKC_SD : CPKCfR : NPKC_GC : NPKC_SD c NPM_SR :
I l II l Ill a IV l V l Vla
W@ EKP);IH CPE#R @w.@
g:&
a Vlb 0 VII l VIII l IX . X
l
: 189 : 189 : 1es : 189 : 169 : 169
: 218 : 218 : 218 : 21a : 218 : 218
: 325 : 325 : 324 : 324 : 323 ; 323
FIG. 2.-Alignment of the catalytic domains of (i) the three sponge group C PS/TKs from G. cydonium (CPKC,GC), S. domunculu (CPKCLSD), and S. ruphunus (CPKC-SR) and (ii) the respective sponge group N PWTKs from G. cydonium (NPKC,GC), S. domunculu (NPKCSD), and S. ruphunus (NPKC_SR); the numbers refer to the aa residues of the respective catalytic domains. All subdomains are shown (I-XI); the nomenclature follows that of Hardie and Hanks (1995). Residues conserved in all sequences are shown in inverted type; those present in more than three sequences are shaded.
Downloaded from https://academic.oup.com/mbe/article-abstract/14/12/1326/1273002by gueston 21 March 2018

Evolution of Metazoan Protein Kinases 1329
HPKC_GC : NPKC_SD : NPKC_SR :
NPKCJX : NPKC_SD : NPKC_SR :
NPKC_GC : NPKC_SD : NPKC_SR :
NPKC_GC : NPKC_SD : NPKC_SR :
NPKC_GC : NPKC_SD : NPKC_SR :
NPKC_GC : NPKC_SD : NPKC_SR :
NPKC_GC T NPKC_SD : NPKC_SR :
NPKC_GC : NPKC_SD : NPKC_SR :
NPKC_GC : NPKC_SD : NPKC_SR :
______”
----NS
WRERGDD
ss---L UPCRDU DGURES
~E”IE----------
TRPGSDES------S SQKS6QPESRPPUP6
LImLI--v
pseudo-
RHUK _-_-_-ET UNRR ------SN
IR-- DKNUIKKS -CIeLIMmMLICICIwm ++++++++++++++++++++++++++++++++++
substrate Zn-1
++++++++++++++++++++++++++++++++++ Zn-2
SP6R KKST RQRS
6DT~N_______ ________^__
LUKLQDEQSBSH
: 78 : 82 : 88
: 156 : 164 : 175
: 237 : 245 : 261
: 325 : 332 : 348
: 398 : 418 : 381
: 478 : 566 : 467
: 568 : 594 : 555
kinase Ser/Thr signature
H : 73? - : 763 - : 724
FIG. 3.-Alignment of the newly described sponge nPKCs from S. domunculu (NPKC-SD) and S. ruphanus (NPKC-SR) with the published sequence from G. cydonium (NPKC-GC, accession number X87684; Kruse et al. 1996). The description of the sites is the same as in the legend to figure 1.
(88%) and one between the domains in S. domunculu and S. raphanus of 65% (80%).
Cloning of Sponge cDNAs Encoding nPKC
In addition to the previously reported nPKC from the sponge G. cydonium (Kruse et al. 1996), two further PKCs from this group have now been isolated from S. domuncula and S. raphanus. These PKCs are character- ized by the lack of a C2 domain signature and are clas- sified as nPKC (Hardie and Hanks 1995). Also, the new kinases have the typical indicator sites of a PS/TK, no tyrosine between the DFG and APE triplets and the GTPYIWAPE stretch (fig. 3).
S. domuncula
The 2,618-bp-long cDNA clone is termed NPKCSD and the protein is termed NPKC-SD. This ki- nase displays the typical sites and signatures which are indicated in figure 3.
S. raphanus The nt sequence encoding an nPKC from this spe-
cies, termed NPKCSR, has a length of 3,455 bp; the predicted aa sequence is named NPKCSR. The char- acteristics of the NPKC-SR sequence are marked in fig- ure 3.
These two kinases show the PS/TK catalytic do- main, which can be subdivided into 12 subdomains (fig. 2). The two new nPKCs have been aligned with the corresponding sequence from G. cydonium (fig. 3); a high similarity, especially within the catalytic domain, is obvious. Again, the S. domunculu nPKC sequence displayed a higher aa sequence identity (similarity) to the respective G. cydonium sequence 76% (86%) than to the S. ruphanus sequence 56% (71%).
Sequence Comparison of Sponge PKC of Groups CandN
The respective three sponge sequences from the C group of PKC and the N group of PKC were subjected Downloaded from https://academic.oup.com/mbe/article-abstract/14/12/1326/1273002
by gueston 21 March 2018

1330 Kruse et al.
NST-l_HS : KRS_SD :
NST-l_HS : KM-SD :
MT-l_HS : KRS_SD :
NST-l_HS : KRS_SD :
NST-l_HS : KRS_SD :
NST-l_HS : KRS_SD :
: 83 : 81
: 188 : 184
: 219 : 247
: 332 : 318
: 418 : 388
: q81 : 478
FIG. 4.-Alignment of the stress-responsive protein kinases from human (MSTl-HS, Q15801; Creasy and Chemoff 1995) and S. domuncula (KRLSD). Residues conserved in the two sequences are shown in inverted type. The description of the sites is the same as in the legend to figure 1; in addition, the acidic region is indicated (R:::fi). The numbers of the subdomains composing the catalytic domain are listed above the alignment,
to phylogenetic analysis. The neighbor-joining analyses revealed that all nPKCs fall in one cluster, and all c- PKCs fall in a second one (fig. 6B). In addition, it be- comes obvious that the nPKC from S. raphanus branch- es off first from the unrooted tree. With respect to the three cPKC sequences, again, the sequence from S. raphanus branches off first from the two demospongian sequences. The robustness of the inferred phylogenies was tested with a bootstrap test, which revealed that the branching of the respective sequences has a significance of =lOO%.
Stress-Responsive Protein Kinase from S. domuncula
A stress-responsive protein kinase (KRS) was iso- lated from the S. domuncula cDNA library. The 1,567- bp-long cDNA KRSSD comprises a deduced polypeptide of 478 aa, termed KRS_SD. The similarity to the human homologous sequence Mst-1 (Creasy and Chernoff 1995) is high, especially within the catalytic domain (fig. 4). The overall aa sequence identity (similarity) be- tween the two sequences is 57% (74%).
The sponge KRS and the human MST-l have their PS/TK catalytic domain close to the N-terminus (Katz, Whalen, and Kehrl 1994; Creasy and Chernoff 1995), a feature which is indicative for the human KRS and the putative Caenorhabditis elegans Ste20 kinase and dif- ferent from other members of the Ste20 kinase family (Creasy and Chernoff 1995). The kinase domain of KRS-SD ranges from aa 25 to 297; the approximate lo- cations of the 12 subdomains are indicated in figure 4, following the suggestions of Katz, Whalen, and Kehrl (1994). The catalytic domain is followed by a short acid- ic region, as already reported for the human sequence (Taylor, Wang, and Erikson 1996).
PRK from S. domuncula From a human fetal brain library, cDNAs encoding
two PRKs, PRKl and PRK2, have been isolated and characterized (Palmer, Ridden and Parker 1995). A sim- ilar kinase was now isolated from S. domuncula. The deduced polypeptide from this sponge displays a se- quence identity (similarity) to the human sequence of 57% (74%).
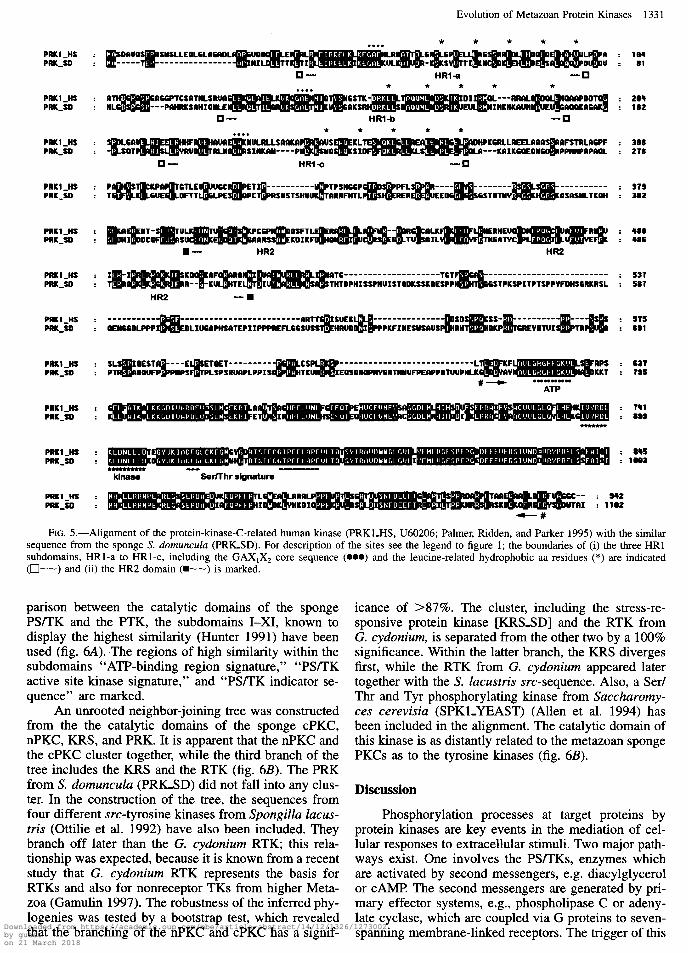
The sponge cDNA, PRKSD, has a size of 3,793 bp; it encodes the 1,102-aa-long putative PRK, PRKSD, which comprises the characteristic features (fig. 5). The catalytic domain of the PRK is preceded by the HR2 domain and the N-terminal HRl domain (Palmer, Rid- den, and Parker 1995). The HR2 domain contains at its C-terminus the pseudosubstrate site. The HRl-domain is subdivided into three shorter stretches, HRl-a-c, which are marked in figure 5. These subdomains, with the core sequence GAX1X2, where X1 is a charged res- idue and X2 is a polar residue, contain five repeated leucine or leucine-related hydrophobic residues.
Relationship of Sponge PS/TK to Sponge PTK
Previously, we reported the first PTK from the low- est metazoan phylum, from the sponge G. cydonium. The cDNA encodes an RTK (Schacke et al. 1994a, 1994b, 1994~; Miiller and Schacke 1996). This RTK contains in its putative aa structure (1) the extracellular part with a Pro/Ser/Thr-rich region, and two complete immunoglobulin-like domains, (2) the transmembrane domain, (3) the juxtamembrane region, and (4) the cat- alytic tyrosine domain. The catalytic domain displays high sequence similarity to the corresponding domains of the PS/TKs in general (Hardie and Hanks 1995) and the sponge sequences, described here. For a direct com-
Downloaded from https://academic.oup.com/mbe/article-abstract/14/12/1326/1273002by gueston 21 March 2018
Evolution of Metazoan Protein Kinases 133 1
PRKl_li3 : PRK_so :
PRKl_R3 : PRK_30 :
PRKlJS : PRK_SD :
PRKl_tB : PRK_30 :
PRKl_HS : PRK_30 :
PRKlJB : PRK_SD :
PRKIJ3 : PRK,30 :
PRKl_HS : PRK_sD :
PRKlJf3 : PRK_SO :
PRKlJ3 : PRK_SD :
PRKl_HS : PRK_#) :
HR1-a
HR1-b
w- HR2 HR2
QEMRCDLPPPI
: 184 : 81
: tell : 182
: 388 : 278
: 373 : 382
: I80 z 488
: 537 : 587
: 837 : 715
: 845 : 1883
Ser/Thr signature
FIG. 5.-Alignment of the protein-kinase-C-related human kinase (PRKLHS, U60206; Palmer, Ridden, and Parker 1995) with the similar sequence from the sponge S. domunculu (PRICSD). For description of the sites see the legend to figure 1; the boundaries of (i) the three HRl subdomains, HRl-a to HRl-c, including the GAX,X* core sequence (0~) and the leucine-related hydrophobic aa residues (*) are indicated (O--) and (ii) the HR2 domain (m--) is marked.
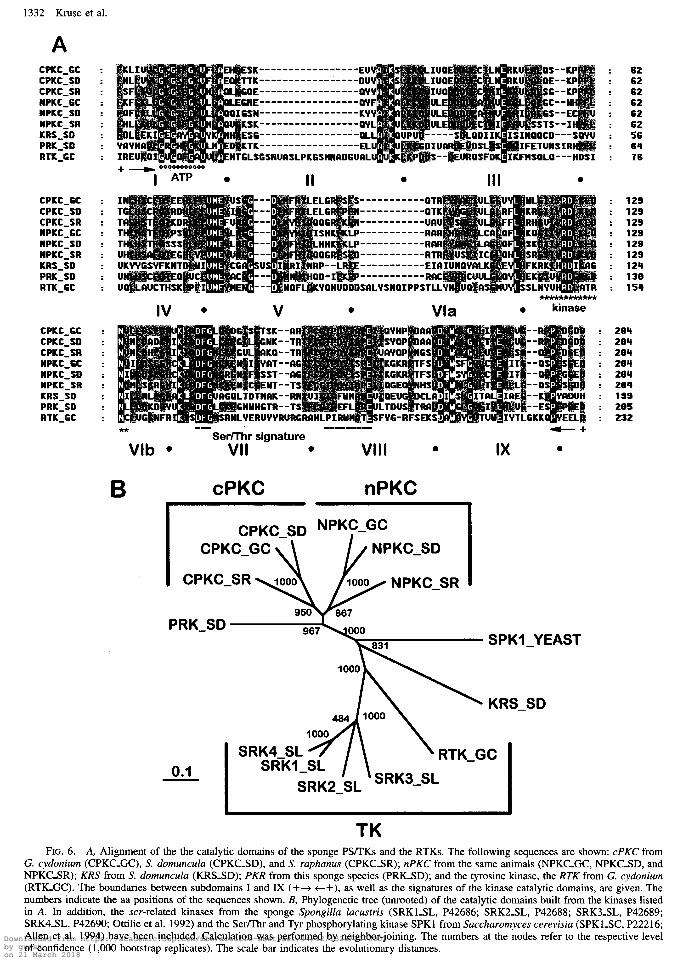
parison between the catalytic domains of the sponge PSLIK and the PTK, the subdomains I-XI, known to display the highest similarity (Hunter 1991) have been used (fig. 6A). The regions of high similarity within the subdomains ‘ ‘ATP-binding region signature,” “PS/TK active site kinase signature,” and “PS/TK indicator se- quence” are marked.
An unrooted neighbor-joining tree was constructed from the the catalytic domains of the sponge cPKC, nPKC, KRS, and PRK. It is apparent that the nPKC and the cPKC cluster together, while the third branch of the tree includes the KRS and the RTK (fig. 6B). The PRK from S. domunculu (PRIGSD) did not fall into any clus- ter. In the construction of the tree, the sequences from four different src-tyrosine kinases from Spongillu Zucus- tris (Ottilie et al. 1992) have also been included. They branch off later than the G. cydonium RTK; this rela- tionship was expected, because it is known from a recent study that G. cydonium RTK represents the basis for RTKs and also for nonreceptor TKs from higher Meta- zoa (Gamulin 1997). The robustness of the inferred phy- logenies was tested by a bootstrap test, which revealed that the branching of the nPKC and cPKC has a signif-
icance of >87%. The cluster, including the stress-re- sponsive protein kinase [KRSSD] and the RTK from G. cydonium, is separated from the other two by a 100% significance. Within the latter branch, the KRS diverges first, while the RTK from G. cydonium appeared later together with the S. Zucustris src-sequence. Also, a Serl Thr and Tyr phosphorylating kinase from Succhuromy- ces cerevisiu (SPKLYEAST) (Allen et al. 1994) has been included in the alignment. The catalytic domain of this kinase is as distantly related to the metazoan sponge PKCs as to the tyrosine kinases (fig. 6B).
Discussion
Phosphorylation processes at target proteins by protein kinases are key events in the mediation of cel- lular responses to extracellular stimuli. Two major path- ways exist. One involves the PS/TKs, enzymes which are activated by second messengers, e.g. diacylglycerol or CAMP The second messengers are generated by pri- mary effector systems, e.g., phospholipase C or adeny- late cyclase, which are coupled via G proteins to seven- spanning membrane-linked receptors. The trigger of this Downloaded from https://academic.oup.com/mbe/article-abstract/14/12/1326/1273002
by gueston 21 March 2018
1332 Kruse et al.
A CPKC_GC CPKC_SD CPKC_SR NPKC_CC NPKC_SD NPKC_SR KRS_SD PRK_SD RTK_GC
CPKC_W CPKC_SD CPKC_SR NPKC_GC NPKC_SD NPKC_SR KRS_SD PRK_SD RTK_GC
CPKC_SC CPKC_SD CPKC_SR NPKC_SC NPKC_SD NPKC_SR KRS_SD PRK_SD RTK_GC
B e _
b--HDSI
ELCR ELGR QD6R ISHK LHHK DD6R RP-- DD-I KYQN
__ -_ __ __ mm
me
--DTR --OTK --URU --RRR
,--RRR ,--RTR ,--EIA &--RRC ‘PSTLL
Vla 0 kinase
** I__ -1BB11B-1
Ser/Thr signature
Vlb l VII 0 VIII .
B cPKC nPKC
PRK_SD _VEAST
.SD
TK
: 62 : 62 : 62 : 62 : 62 : 62 : 56 : 64 : 76
: 129 : 129 : 129 : 129 : 129 : 129 : 124 : 138 : 154
264 294 264 204 294 284 199 205 232
FIG. 6.-A, Alignment of the the catalytic domains of the sponge PS/TKs and the RTKs. The following sequences are shown: cPKC from G. cydonium (CPKCGC), S. domunculu (CPKCSD), and S. ruphanus (CPKC-SR); nPKC from the same animals (NPKC_GC, NPKC_SD, and NPKC-SR); KRS from S. domunculu (KRSSD); PKR from this sponge species (PRICSD); and the tyrosine kinase, the RTK from G. cydonium (RTECGC). The boundaries between subdomains I and IX (++ c+), as well as the signatures of the kinase catalytic domains, are given. The numbers indicate the aa positions of the sequences shown. B, Phylogenetic tree (unrooted) of the catalytic domains built from the kinases listed in A. In addition, the scr-related kinases from the sponge Spongillu lucustris (SRKLSL, P42686; SRKZSL, P42688; SRKISL, P42689; SRK4_SL, P42690; Ottilie et al. 1992) and the Ser/Thr and Tyr phosphorylating kinase SPKl from Succhuromyces cerevisiu (SPKl_SC, P22216; Allen et al. 1994) have been included. Calculation was performed by neighbor-joining. The numbers at the nodes refer to the respective level of confidence (1,000 bootstrap replicates). The scale bar indicates the evolutionary distances. Downloaded from https://academic.oup.com/mbe/article-abstract/14/12/1326/1273002
by gueston 21 March 2018

pathway is the interaction of a specific ligand with its receptor (reviewed in Nishizuka 1992). In the second pathway, the extracellular signals are transduced across the cellular membrane by transmembrane receptors, which have intrinsic enzymatic activity, as in the group of PTKs (reviewed in Van der Geer, Hunter, and Lind- berg 1994). Although the two kinases function at dif- ferent positions within the two signal transduction path- way chains, the catalytic domains of PS/TK and PTK are homologous (reviewed in Hunter 1991). The num- bers of kinases of both families discovered are growing exponentially; more than 200 kinases have been cloned (Hunter 1991).
PS/TKs are known from both eukaryotic and pro- karyotic organisms (reviewed in Hunter 1991). A com- mon ancestor for the kinase domain of these enzymes has recently been proposed (Kruse et al. 1996), while the overall composition of the domains of the PS/TKs differs between metazoan and nonmetazoan organisms (Kruse et al 1996). The class of serine/threonine kinases comprises the classical PS/TKs but also PS/TK-related kinases, e.g., the KRSs (Taylor, Wang, and Erikson 1996) or the PRKs (Palmer, Ridden, and Parker 1995).
In contrast to the PS/TKs, the appearance of the PTKs is a relatively recent process and occurred during the transition of the Protozoa to the Metazoa. The phy- logenetically oldest enzyme of the PTKs has been de- scribed from a member of the oldest metazoan phylum, the sponges (reviewed in Mtiller and Schacke 1996). The work summarized here establishes the emergence of the PTKs within the superfamily of PS/TKs and their related kinases. Members of PS/TKs of the family of cPKCS and nPKCs, as well as of the PUTK-related ki- nases, a KRS, and a PRK, have been cloned from the demosponges G. cydonium and S. domunculu and from the calcareous sponge S. ruphanus. Based on the anal- ysis of the catalytic domains of the PS/TKs, a phylo- gentic relationship could be established. Expectedly, the sequences belonging to the family of cPKC cluster to- gether and are separated from those classified as nPKC. The analysis showed in addition that in a third branch, the KRS from S. domunculu is together with the PRK from the same species. Interesting is the finding that the phylogenetically oldest PTK, the RTK from G. cydon- ium, falls into the same group. This finding is even more striking if the tree also includes the src-related kinases from S. Zucustris (Ottilie et al. 1992). The phylogenetic analysis revealed that the nonreceptor kinases branched off later than the G. cydonium RTK. Recently, it was summarized that budding yeasts have 113 conventional protein kinase genes, but none of them has true tyrosine kinase signatures, indicating that yeast tyrosine-phos- phorylating enzymes constitute a different group of ty- rosine kinases (Hunter and Plowman 1997).
Our finding indicates that the PTKs originated within the superfamily of PS/TKs, a step which occurred during the evolution of sponges. One question to be in- vestigated in the future is that of the relationship of the PS/TKs and PTKs to the tyrosine-threonine kinases, e.g., the MAP kinases, which are present in fungal, plant, and metazoan snecies and comprise signatures related to the
Evolution of Metazoau Protein Kinases 1333
PS/TK and PTK (Anderson et al. 1990; Blumer, John- son, and Lange-Carter 1994; Davis 1994).
A further important outcome of this study is the finding that, based on sequence data from cDNAs and encoding proteins- here the kinases-a phylogenetic positioning of the two sponge classes Calcarea and De- mospongiae appears to be possible. In earlier studies, sequence data from 28s rRNAs of Porifera revealed un- expected relationships (Lafay et al. 1992). Based on these results the phylogenetic relationships showed a separation of the Porifera into groups with different cor- respondence to the phyla Cnidaria and Ctenophora, hence allowing no conclusive branching order. Also, 18s rRNA sequence analyses did not prove to be suit- able to resolve deep branchings in the phylogenetic tree, including that of Porifera (Rodrigo et al. 1994). In a more recent study (Cavalier-Smith et al. 1996) the two classes of Porifera, Calcarea and Demospongiae, are grouped together; within this group the Calcarea are the more recent. These data are in conflict with embryolog- ical findings, which suggest that calcareous sponges are more simple than most siliceous sponges, especially with respect to organization of the germ layers (Boro- jevic 1970). Our data show that the catalytic domains of both the nPKCs and the cPKCs, cloned from the cal- careous sponge S. ruphunus, are phylogenetically older than the derived sequences from the two demosponges. The relationships are statistically robust, as confirmed by bootstrapping.
In conclusion, the present study indicates that, based on the deduced kinase data, the class Calcarea appeared earlier during evolution than the class Demos- pongiae. Lastly, the phylogenetic analyses strongly sug- gest that the PTKs have a common ancestor with the PS/TK superfamily, from which the G. cydonium RTKs diverged first.
Acknowledgments
This work was supported by grants from the Deutsche Forschungsgemeinschaft (Mii 348/14-l). and the International Human Frontier Science Program (RG- 333/96-M). The sequences reported here are deposited in the EMBL database: Suberites domunculu cPKC (CPKCSD), Y 13099; nPKC (NPKCSD), Y 13102; stress- responsive protein kinase (KRSSD), Y 13 10 1; protein-ki- nase-C-related kinase (PRKSD), Y 13 104; Sycon rupha- nus cPKC (CPKCSR), Y 13100; and nPKC (NPKCSR), Y13103.
LITERATURE CITED
ALLEN, J. B., Z. ZHOU, W. SIEDE, E. C. FRIEDBERG, and S. J. ELLEDGE. 1994. The SADUR4D53 protein kinase controls multiple checkpoints and DNA damage-induced transcrip- tion in yeast. Genes Dev. 8:2401-2415.
ANDERSON, N., J. MALLER, N. TONKS, and T. STURGILL. 1990. Requirement for integration of signals from two distinct phosphorylation pathways for activation of MAP kinase. Nature 343:65 l-653.
AUSUBEL, E M., R. BRENT, R. E. KINGSTON, D. D. MOORE, J. A. SMITH, J. G. SEIDMANN, and K. STRUHL. 1995. Current Downloaded from https://academic.oup.com/mbe/article-abstract/14/12/1326/1273002
by gueston 21 March 2018

1334 Kruse et al.
protocols in molecular biology. John Wiley & Sons, New York.
BERGQUIST, I? R. 1978. Sponges. Hutchinson, London. BLAKE, M. S., K. H. JOHNSTON, G. J. RUSSEL-JONES, and E.
C. GOTSCHLICH. 1984. A rapid, sensitive method for de- tection of alkaline phosphatase-conjugated anti-antibody on western blots. Anal. Biochem. 136: 175-179.
BLUMER, K. J., G. L. JOHNSON, and C. A. LANGE-CARTER. 1994. Mammalian mitogen-activated protein kinase kinase kinase (MEKK) can function in a yeast mitogen-activated protein kinase pathway downstream of protein kinase C. Proc. Natl. Acad. Sci. USA 91:4925-4929.
BOROJEVIC, R. 1970. Differentiation cellulaire dans l’embryogenese et la morphogenese chez les spongiaires. Symp. Zool. Sot. Lond. 25:467-490.
CAVALIER-SMITH, T., M. T. E. P ALLSOPP, E. E. CHAO, N. BOURY-ESNAULT, and J. VACELET. 1996. Sponge phylog- eny, animal monophyly, and the origin of the nervous sys- tem: 18s rRNA evidence. Can. J. Zool. 74:2031-2045.
CREASY, C. L., and J. CHERNOFF. 1995. Cloning and charac- terization of a human protein kinase with homology to Ste20. J. Biol. Chem. 270:21695-21700.
DAVIS, R. J. 1994. MAPKs: new JNK expands the group. Trends Biochem. Sci. 19:470-473.
DAYHOFF, M. O., R. M.SCHWARTZ, and B. C. ORCUTT. 1978. A model of evolutionary change in protein. Pp. 345-352 in M. 0. DAYHOFF, ed. Atlas of protein sequence and struc- ture. Vol. 5, suppl. 3. National Biomedical Research Foun- dation, Washington, D.C.
FELSENSTEIN, J. 1993. PHYLIP Version 3.5. University of Washington, Seattle.
GAMULIN, V., A. SKOROKHOD, V. KAVSAN, I. M. MILLER, and W. E. G. MULLER. 1997. Experimental indication against blockwise evolution of metazoan protein molecules: ex- ample, receptor tyrosine kinase gene from the sponge Geo- dia cydonium. J. Mol. Evol. 44:242-252.
HANKS, S. K., A. M. QUINN, and T. HUNTER. 1988. The pro- tein kinase family: conserved features and deduced phylog- eny of the catalytic domains. Science 241:42-52.
HARDIE, G., and S. HANKS. 1995. The protein kinase facts- book: protein-tyrosine kinases. Academic Press, London.
HUNTER, T. 199 1. Protein kinase classification. Methods En- zymol. 200: 3-8 1.
HUNTER, T., and G. D. PLOWMAN. 1997. The protein kinases of budding yeast: six score and more. Trends Biochem. Sci. 22: 18-22.
KATZ, I?, G. WHALEN, a.1 J. H. KEHRL. 1994. Differential expression of a novel protein kinase in human B lympho- cytes. J. Biol. Chem. 269: 16802-16809.
-USE, M., V. GAMULIN, H. CETKOVIC, Z. PANCER, I. M. M~_?LLER, and W. E. G. MILLER. 1996. Molecular evolution of the metazoan protein kinase C multigene family. J. Mol. Evol. 43:374-383.
LAFAY, B., N. BOURY-ESNAULT, J. VACELET, and R. CHRISTEN. 1992. An analysis of partial 28s ribosomal RNA sequences suggests early radiation of sponges. BioSystems 28: 139- 151.
MUELLER, W. E. G., and H. SCHACKE. 1996. Characterization of the receptor protein-tyrosine kinase gene from the marine sponge Geodia cydonium. Prog. Mol. Subcell. Biol. 17: 183-208.
NICHOLAS, K. B., and H. B. NICHOLAS JR. 1996. GeneDoc. Version 1.1.004. Distributed by the author, cris.com/ -ketchup/genedoc.shtml; INTERNET (1997).
NISHIZUKA, Y. 1992. Intracellular signaling by hydrolysis of phospholipids and avtivation of protein kinase C. Science 258:607-614.
OTTILIE, S., E RAULF, A. BARNEKOW, G. HANNIG, and M. SCHARTL. 1992. Multiple src-related kinase genes, s&l-4, in the fresh water sponge Spongilla Zacustris. Oncogene 7: 1625-1630.
PALMER, R. H., J. RIDDEN, and l? J. PARKER. 1995. Cloning and expression patterns of two members of a novel protein- kinase-C-related kinase family. Eur. J. B&hem. 227:344- 351.
PC/GENE. 1995. Data Banks CD-ROM. Release 14.0. IntelliGenetics, Mountain View, Calif.
RODRIGO, A. G., I? R. BERGQUIST, I? L. BERGQUIST, and R. A. REEVES. 1994. Are sponges animals? An investigation into the vagaries of phylogenetic interference. Pp. 47-54 in R. W. M. v. SOEST, T. M. G. v. KEMPENA, and J. C. BRAEK- MAN, eds. Sponges in time and space. Balkema Press, Rot- terdam.
ROSE-JOHN, S., A. DIETRICH, and I? MARKS. 1988. Molecular cloning of mouse protein kinase C (PKC) cDNA from Swiss 3T3 fibroblasts. Gene 74:465-47 1.
SCH&XE, H., W. E. G. MILLER, V. GAMULIN, and B. RINK- EVICH. 1994a. The Ig superfamily includes members from the lowest invertebrates to the highest vertebrates. Immunol. Today 15497-498.
SCH~CKE, H., B. RINKEVICH, V. GAMULIN, I. M. MUELLER, and W. E. G. MULLER. 1994b. Immunoglobulin-like domain is present in the extracellular part of the receptor tyrosine ki- nase from the marine sponge Geodia cydonium. J. Mol. Re- cognit. 7:272-276.
SCHWCKE, H., H. C. SCHR~DER, V. GAMULIN, B. RINKEVICH, I. M. MUELLER, and W. E. G. MUELLER. 1994~. Molecular cloning of a receptor tyrosine kinase from the marine sponge Geodia cydonium: a new member of the receptor tyrosine kinase class II family in invertebrates. Mol. Membr. Biol. ll:lOl-107.
SKOROKHOD, A., H. SCH&XE, B. DIEHL-SEIFERT, R. STEFFEN, A. HOFMEISTER, and W. E. G. MILLER. 1997. Immuno- chemical localization of the phylogenetically oldest receptor tyrosine kinase: existence in the marine sponge Geodia cy- donium. Cell. Mol. Biol. 43:509-519.
STABEL, S., and l? J. PARKER. 1991. Protein kinase C. Phar- macol. Ther. 51:7 l-95.
TAYLOR, L. K., H. C. R. WANG, and R. L. ERIKSON. 1996. Newly identifed stress-responsive protein kinases, Krs- 1 and Krs-2. Proc. Natl. Acad. Sci. USA 93:10099-10104.
THOMPSON, J. D., D. G. HIGGINS, and T. J. GIBSON. 1994. CLUSTAL W improving the sensitivity of progressive mul- tiple sequence alignment through sequence weighting, po- sitions-specific gap penalties and weight matrix choice. Nu- cleic Acids Res. 22:4673-4680.
VAN DER GEER, I?, T. HUNTER, and R. A. LINDBERG. 1994. Receptor protein-tyrosine kinases and their signal transduc- tion pathways. Annu. Rev. Cell Biol. 10:251-337.
CLAUDIA KAPPEN, reviewing editor
Accepted September 12, 1997
Downloaded from https://academic.oup.com/mbe/article-abstract/14/12/1326/1273002by gueston 21 March 2018