dr. kalet león monzón
DESCRIPTION
Modeling the role of IL2 on CD4+T cell dynamics: On the effect of IL-2 modulating therapies (2). Dr. Kalet León Monzón. Summer school Leeds (2009). R. R. R. T. T. T. Crosstalk model of dominant tolerance to self The key role of IL-2. - PowerPoint PPT PresentationTRANSCRIPT

Dr. Kalet León Monzón
Modeling the role of IL2 on CD4+T cell dynamics: On the effect of IL-2 modulating therapies (2)
Summer school Leeds (2009)

Crosstalk model of dominant tolerance to self The key role of IL-2
T
R
T
T
R
RRegulatory T cells inhibit the activation and proliferation of helper T cells by an interaction which requires their colocalized activation at the APCs.
Helper T cells produce a growth factor that Regulatory T cells use to proliferate and survive
Leon et al J. Theor. Biol. 2000 Leon et al J. Immunol. 2001
Leon et al 2002 pHD thesis
IL-2 is a good candidate growth factor

In vitroRegulatory cells do not express IL-2
Regulatory cells over-express CD25
Regulatory cells proliferate when stimulated in the presence of IL-2
Regulatory cells inhibit IL-2 mRNA expression in their targets
In co-cultures, regulatory T cells proliferate/survive by using the IL-2 produced by their targets. Moreover they consume this IL-2 starving the normal T cell
In vivoAnimals deficient in IL-2, CD25, IL-2Rb chains exhibit lympho-proliferative disorders
and autoimmunity
These autoimmune disorders, in receptor deficient mice, can be prevented by transfer of wild-type Tregs. But in Il-2 deficient mice, they can be prevented by the addition of Il-2 or
transfer of IL-2 producing cells.
In-vivo depletion of IL-2 induces autoinmunity in 10-20 day thymectomized BalB/c mice.

Our Master Modelling Plan
To extend the Cross-Regulation model to include IL2 dynamics
-Check in which conditions the extended models conserve the key dynamical features of the original cross-regulation model.
-Compare models of different hypotheses about the specific role of IL2 in T cell mediated suppression
To study the effect of IL2 modulating therapies currently used in clinical or preclinical studies
-Study the impact of IL2 depletion treatments.
-Study the impact of IL2 administration treatments.

Adding IL-2 in the cross-regulation model
Basic interactions for T cells
-E cells produce IL-2 upon activation at the APCs.
-E cells need both APCs and c cytokine signal to proliferate and survive in vivo.
-E cells use in vivo either IL2 or another homeostatic cytokine (denoted IL-) to proliferate and/or survive.
-R cells do not produce IL-2
-R cells need both APC and IL-2 signals to proliferate and survive in vivo.
-R cells inhibit E cell activation and production of IL2 by a mechanism requiring their co-localized activation at the APCs
-R cells constitutively over express the Alpha chain of IL2R (CD25), while E cells up regulate this molecule transiently after activation.
-The main sources of IL2 in vivo are the helper CD4+ FoxP3- T cells (our E cells)
-IL-2 concentration is reduced in vivo by both by renal elimination and cell internalization upon binding to the IL2 receptor
ER
APC
IL-2(+)
(+)IL- (-)
(-)
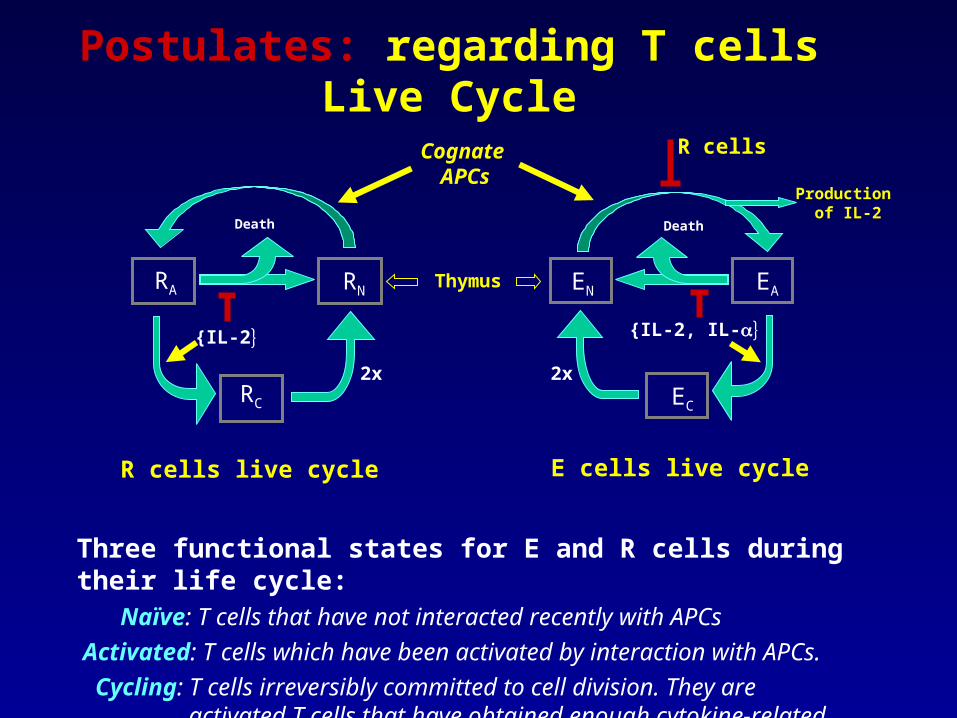
Postulates: regarding T cells Live Cycle
EN EA
R cells
Production of IL-2
EC
{IL-2, IL-
2x
Death
ThymusRNRA
Cognate APCs
RC
{IL-2
2x
Death
R cells live cycle E cells live cycle
Three functional states for E and R cells during their life cycle:
Naïve: T cells that have not interacted recently with APCs
Activated: T cells which have been activated by interaction with APCs.
Cycling: T cells irreversibly committed to cell division. They are activated T cells that have obtained enough cytokine-related signals

Postulates: regarding IL2 “life Cycle”
EN
R cells
Cognate APCs
EA
Production binding to IL2r Internalization
+degradation
Degradation/ elimination
IL2 interactions
unbinding of IL2r
F2IL TB2IL
Processes regarding IL2:
IL2 is produced by E cells upon activation on the APCs
IL2 is degraded by renal elimination while not bound to receptor.
IL2 is internalized and degraded by T cells, following its binding to receptors.

Postulates: regarding interactions with APCs
T cell get signals from APCs upon direct conjugation. Conjugation to APCs is assumed to be a fast process and taken to quasi-steady state equilibrium.
Cells in different states (naïve, activated, cycling) have different conjugation constants.
Regulatory cells suppress E cell activation or IL2 production by a mechanism dependent on co-localized activation on APCs.
EN
R
EA
EN
(-)
RA
EN
RN
RA
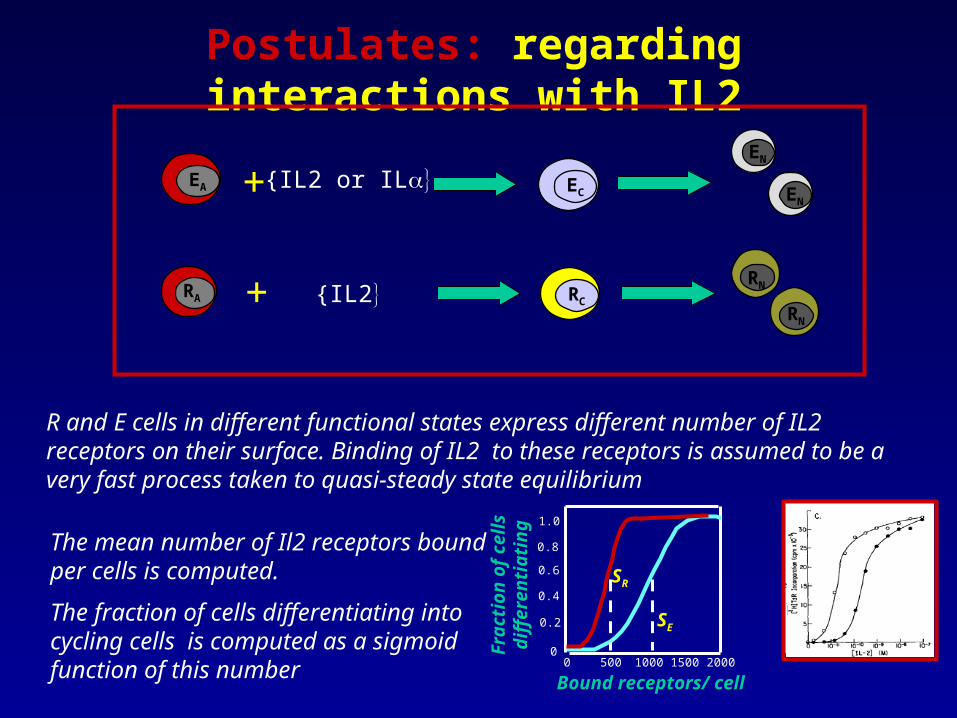
Postulates: regarding interactions with IL2
EA
RA
+ {IL2 or IL EC
EN
EN
{IL2+ RC
RN
RN
R and E cells in different functional states express different number of IL2 receptors on their surface. Binding of IL2 to these receptors is assumed to be a very fast process taken to quasi-steady state equilibrium
The mean number of Il2 receptors bound per cells is computed.
The fraction of cells differentiating into cycling cells is computed as a sigmoid function of this number 0 500 1000 1500 2000
0
0.2
0.4
0.6
0.8
1.0
SE
Bound receptors/ cell
Fra
ctio
n o
f ce
lls
diff
eren
tiat
ing
SR

Model Equations
)RIL+RIL+R(ILK
)EIL+E(ILKILKAs
REK=
dt
ILd
CCR
BAAR
BNNR
BRin
CCE
BAAE
BEin
id
)(sBTB
Nipi
222
22212
1
CRPAhR
Bh
R
hRBR
SC RKR
)(Il+)(S)(Il
K=dtRd
A
A
22
ARS
BN
RA
A RKRK=dt
Rd
FN
RdAhR
Bh
R
hRR
SCRp
BN
RAR
N RKR)(IL+)(S
)(SKα+RK+RK=
dt
RdA
22
CEPAhhE
Bh
E
hhEBE
SC EKE
(Ilα+)(Il+)(SIlα+(Il
K=dtEd
A
A
)2)()2
AES
)η(sBTB
NEA
A EKAs
REK=
dt
Ed
1
1
FN
EdAhhE
Bh
E
hEE
SC
)η(sBTB
NEAE
N EKE(Ilα+)(Il+)(S
)(SKαEE
PK+As
REK=
dt
EdA
)221
1E
ffec
tors
Cel
lsR
egul
ator
y C
ells
IL2
Plus algebraic relations to compute the number of cells bound to APCs and of Il2 bound to its receptor

Handling parameters
The models have 30 parameters, thus although we are interested in generic properties not dependent on specific parameter values, we try to find a good reference set of parameters based on existing experimental data.
The idea is to use this set of values as a reasonable reference to study model properties, but then study robustness of model behavior to parameter changes always around these reference values.
Thus we do a partial calibration parameterization of the model

Lymph node volume VNL=10 -6 L Personal communication
# of dendritic cells. APCS= 104 – 105 Personal communication
T cell division rates Cycling time (E=R) = ln2/tc1/2 ; tc1/2=5 h
are taken such that the mean doubling time is between 6-20 h
Stoll, Science, 2002. 296: 1873
(Confocal microsc)
T cell death rate Kd= ln2/t1/2 ; t1/2=7-30 días Surh et al, 2006
# of High affinity IL2 receptors
700 – 1000R up to 10 time more
Smith KA, Cell Res, 2006. 16: 11
Conjugation IL2 to IL2Rs alpha: Kon= 107 M-1s-1 ;
Koff= 10-1 s-1
Beta-Gamma: Kon= 105 M-1s-1 ;
Koff= 10-4 s-1
Wang H, J Exp Med, 1987, 166:1055
IL2/IL2R internalization rate
= ln2/t1/2 ; t1/2=15 min
Fallon, J Biol Chem, 2000. 275: 6790
Free IL2 degradation Rate = ln2/t1/2 ; t1/2=20 min Donohue et al, 1984
Most parameters values are bounded from existent dataMost parameters values are bounded from existent data
;K;KEAK R
A ;KES
RS
=KEPK R
P
K id
=KEinK R
in
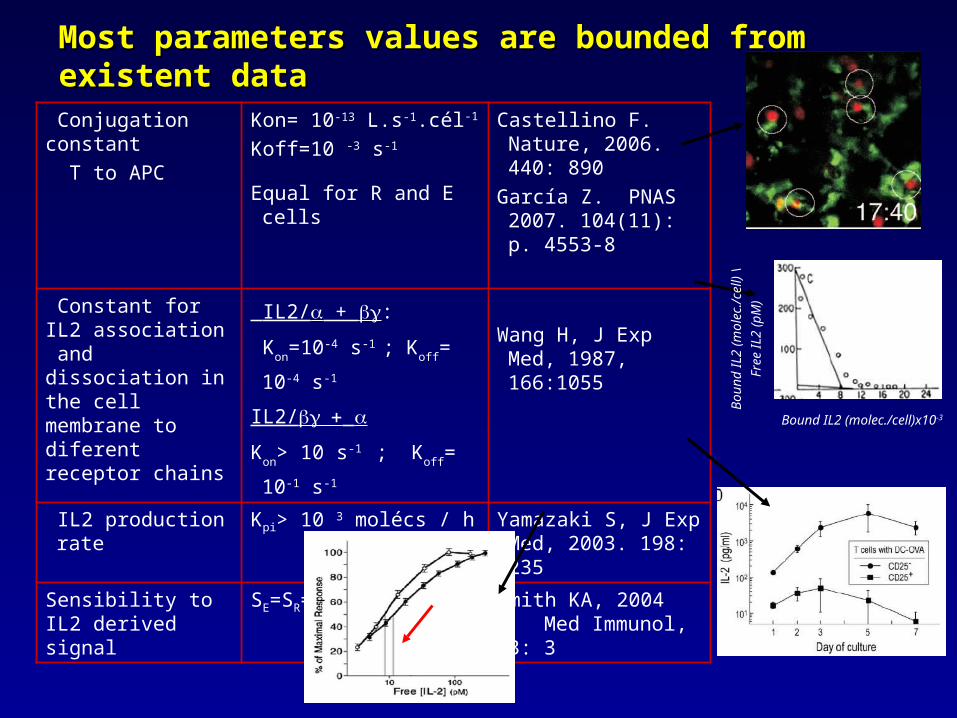
Most parameters values are bounded from existent dataMost parameters values are bounded from existent data
Conjugation constant T to APC
Kon= 10-13 L.s-1.cél-1
Koff=10 -3 s-1
Equal for R and E cells
Castellino F. Nature, 2006. 440: 890
García Z. PNAS 2007. 104(11): p. 4553-8
Constant for IL2 association and dissociation in the cell membrane to diferent receptor chains
IL2/ + :
Kon=10-4 s-1 ; Koff= 10-4
s-1
IL2/
Kon> 10 s-1 ; Koff= 10-1
s-1
Wang H, J Exp Med, 1987, 166:1055
IL2 production rate
Kpi> 10 3 molécs / h Yamazaki S, J Exp Med, 2003. 198: 235
Sensibility to IL2 derived signal
SE=SR=500; h=4 Smith KA, 2004 Med Immunol, 3: 3
Bound IL2 (molec./cell)x10-3
Bou
nd IL
2 (m
olec
./cel
l) \
Fre
e IL
2 (p
M)

Does the extended model does conserve the key dynamical features of the original
cross-regulation model ?
Bistability One stable state interpreted as tolerance and the other as immunity
Adaptive response to the quantity of APCsChange from tolerance to immunity with abrupt increase of APC numbersAdaptation the tolerant state upon slow increase of APC numbers.
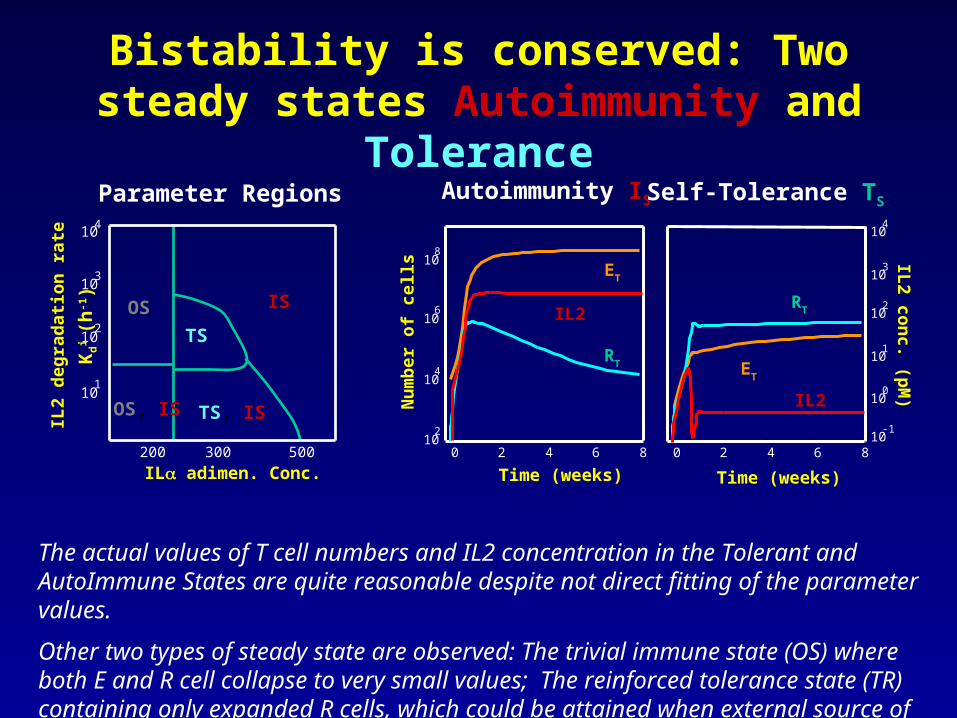
Bistability is conserved: Two steady states Autoimmunity and Tolerance
The actual values of T cell numbers and IL2 concentration in the Tolerant and AutoImmune States are quite reasonable despite not direct fitting of the parameter values.
Other two types of steady state are observed: The trivial immune state (OS) where both E and R cell collapse to very small values; The reinforced tolerance state (TR) containing only expanded R cells, which could be attained when external source of IL2 is added.
Parameter Regions Autoimmunity IS Self-Tolerance TS
Time (weeks)
IL2 co
nc. (p
M)
0 2 4 6 8
IL2
ET
RT
104
103
102
101
100
10-1
Time (weeks)
Nu
mb
er o
f ce
lls
IL2
ET
RT
106
108
102
104
0 2 4 6 8
IL2
deg
rad
atio
n r
ate
Kd
i (h
-1)
IL adimen. Conc.200 300 500
10
103
104
1
102
OS, IS
OS IS
TS
TS, IS
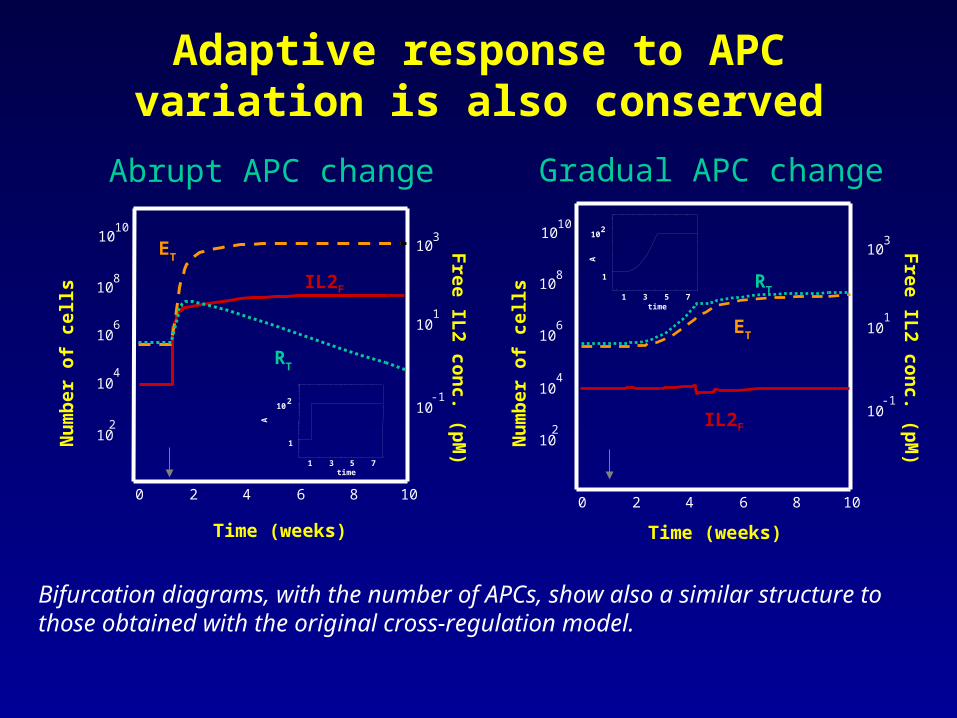
Adaptive response to APC variation is also conserved
0 2 4 6 8 10
10
106
108
2
104
Time (weeks)
1010
ET
RT
10
103
-1
101
A
1 3 5 7
1
102
time
Nu
mb
er o
f ce
lls
Free
IL2 c
on
c. (pM
)
0 2 4 6 8 10
10
106
108
2
104
1010
Time (weeks)
IL2F
ET
RT
10
103
-1
101
1 3 5 7
1
102
time
A
Nu
mb
er o
f ce
lls
Free
IL2 c
on
c. (pM
)
IL2F
Bifurcation diagrams, with the number of APCs, show also a similar structure to those obtained with the original cross-regulation model.
Abrupt APC change Gradual APC change

Three necessary parameter conditions for a reasonable model behavior
R cells use IL2 more effectively at low concentrations
E cells have net growth rate faster than R cells in IL2 excess
E cells have to use a cytokine alternative to IL2 to proliferate
AR
RAE
E RIS>RIS //
Rd
RA
REd
EA
E KKKKKK //
0Ilα

The effect of knocking down IL in the modelIL
2 d
egra
dat
ion
rat
e K
di (h
-1)
IL adimen. Conc.
200 300 500
10
103
104
1
102
OS, IS
OS IS
TS
TS, IS
If Il is forced to go to zero, the system reaches a parameter regime where only autoimmune or a trivial steady state could be attained
This result would predict a dramatic effect on T cells populations of knocking down this hypothetical cytokine
Interestingly the only gamma chain family cytokine whose knockout has a profound effect on the size of T cell population is Il7. Thus the model would predict it to be the best candidate for our IL cytokine.
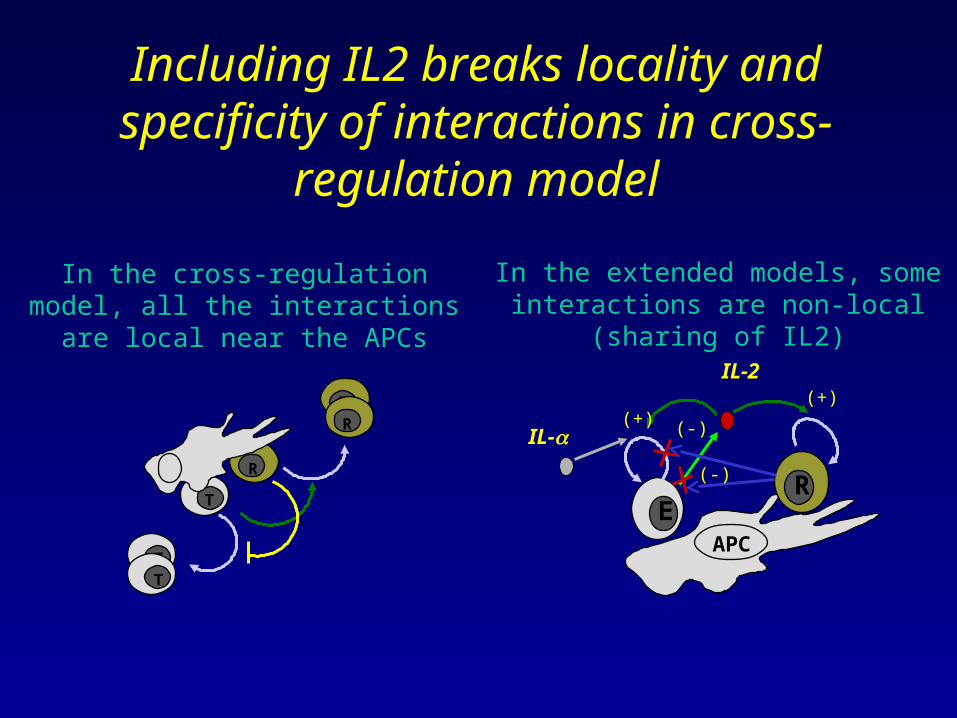
Including IL2 breaks locality and specificity of interactions in cross-
regulation model
ER
APC
IL-2(+)
(+)IL- (-)
(-)T
R
TT
RR
In the cross-regulation model, all the interactions are local near the APCs
In the extended models, some interactions are non-local (sharing of IL2)

Implications for independence of concomitant responses to different self-
antigen in a given lymph node
A1
A2
<- E1,R1
<- E2,R2
In the extended models there should be strong
interference between the immune reactions of
different clones
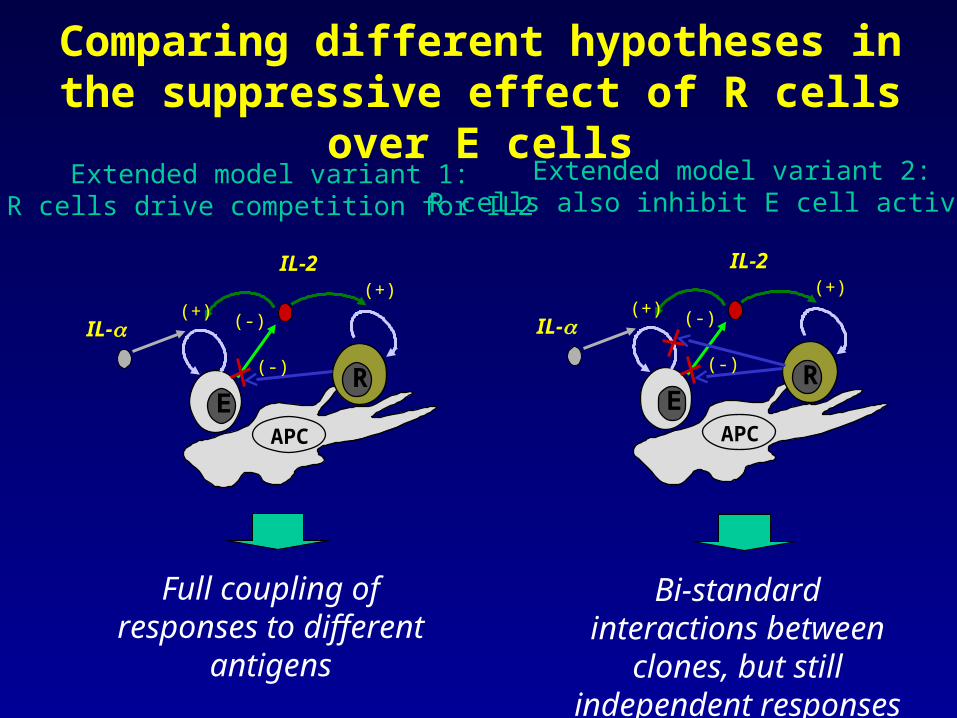
Comparing different hypotheses in the suppressive effect of R cells over E cells
IL-2
ER
APC
(+)(+)
IL- (-)
(-)
ER
APC
IL-2(+)
(+)IL- (-)
(-)
Extended model variant 1:R cells drive competition for IL2
Full coupling of responses to different
antigens
Extended model variant 2:R cells also inhibit E cell activation
Bi-standard interactions between clones, but still independent responses

But, another possible solution to be explored in the future is further locality on the use of
IL2 inside a given lymph node
A1
A2
<- E1,R1
<- E2,R2
But since the suppression mechanism relies on competition of E and R cells for IL2, the lymph node will have to be subdivided in sub-domains presenting different antigens

The extended model can be used to study the effect of IL2 modulation therapies. i.e Therapies that deplete IL2 and Therapies that inject IL2
Let´s study whether these treatments can trigger a switch of steady states in the models: Tolerance to immunity and viceversa.
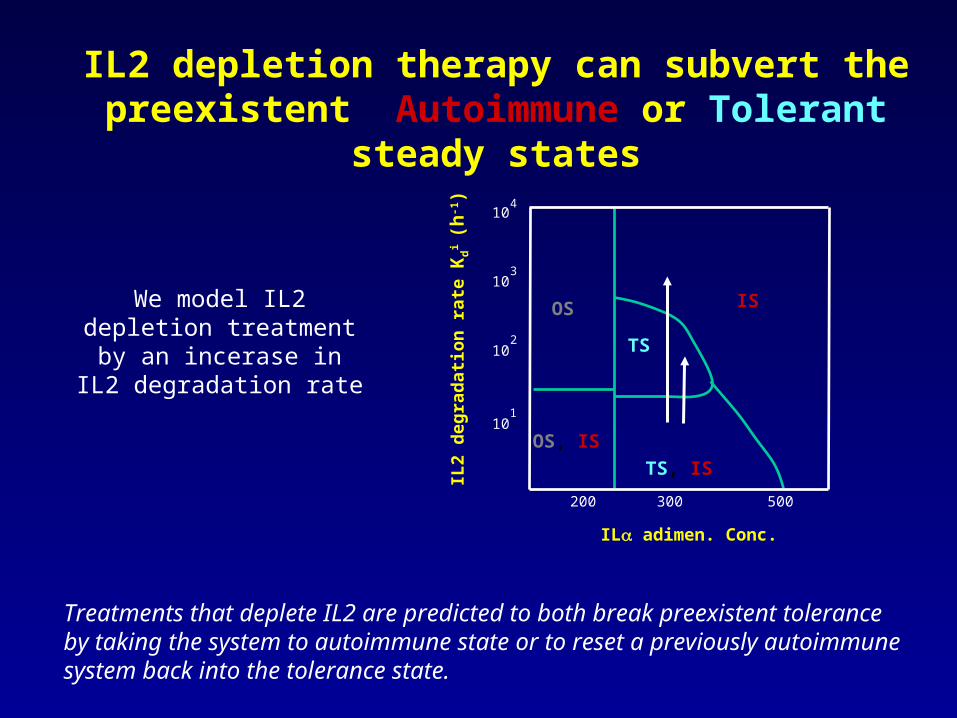
IL2
deg
rad
atio
n r
ate
Kd
i (h
-1)
IL adimen. Conc.
200 300 500
10
103
104
1
102
OS, IS
OS IS
TS
TS, IS
IL2 depletion therapy can subvert the preexistent Autoimmune or Tolerant steady states
We model IL2 depletion treatment by an incerase in
IL2 degradation rate
Treatments that deplete IL2 are predicted to both break preexistent tolerance by taking the system to autoimmune state or to reset a previously autoimmune system back into the tolerance state.

IL2 depletion can subvert the preexistent Autoimmune or Tolerant steady states
Breaking Tolerance
Reestablishing Tolerance
This supports applications of IL2 depletion both in the treatment of autoimmune diseases and cancer, but these applications strongly depend on treatment schedules
0 5 10 15 20
106
108
102
104
104
103
102
101
100
10-1
IL2 co
nc. (p
M)N
um
ber
of
cells
IL2
ET
RT
+
0 10 20 30 40
106
108
102
104
104
103
102
101
100
10-1
IL2 co
nc. (p
M)N
um
ber
of
cells
IL2
ET
RT
Time (weeks)
+ -
Dep
leti
on
Do
seD
eple
tio
n D
ose
104
102
1 2 3 4 5 6 7
103
IS
TS
6
1 2 3 40
104
10
102
105
103
IS
TS
Duration of treatment (w)
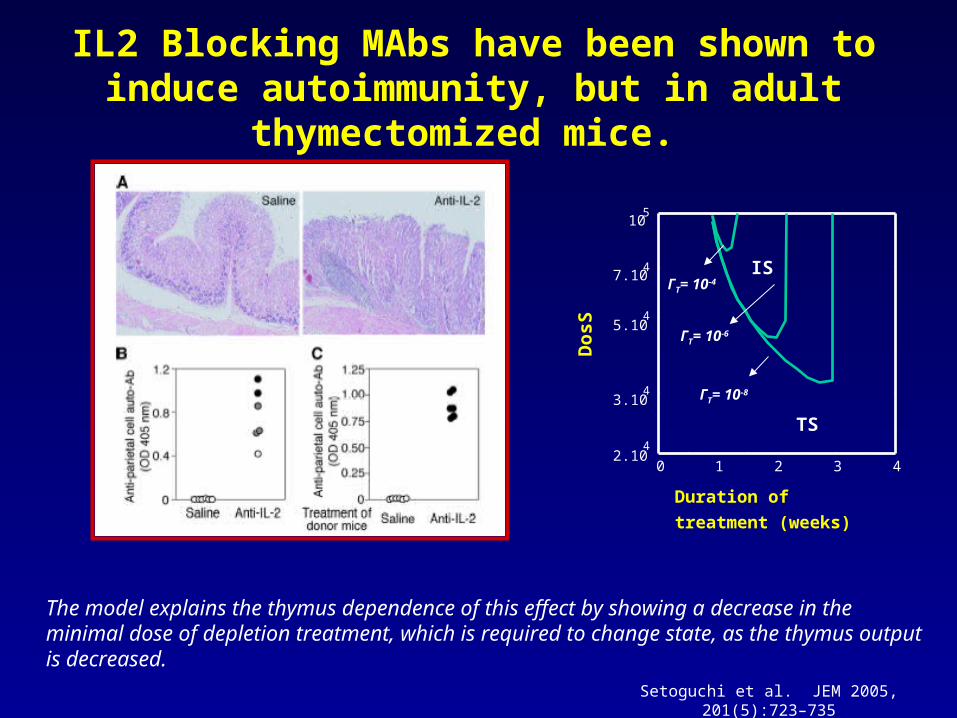
IL2 Blocking MAbs have been shown to induce autoimmunity, but in adult thymectomized mice.
The model explains the thymus dependence of this effect by showing a decrease in the minimal dose of depletion treatment, which is required to change state, as the thymus output is decreased.
5.104
105
2.104
7.104
3.104
1 2 3 40
Do
sS
IS
TS
ΓT= 10-4
ΓT= 10-6
ΓT= 10-8
Duration of treatment
(weeks)
Setoguchi et al. JEM 2005, 201(5):723–735

Anti-IL2 MAbs can induce tumor regression
1 mg AcM IL-2 iv
5x105 IU/Kg sc IL-2
5 d2 d
F10 F10 irradiated
Montero et al. 2005 unpublished results

IL2 injection therapy can not subvert the preexistent Autoimmune or Tolerant steady states
We model IL2 injection treatment by an increase in the
external source of IL2 (i) Γi
0.1 1 10 100 1000 10000
10
106
108
2
104
Ilα
TS, IS
RT, IS
OS, IS
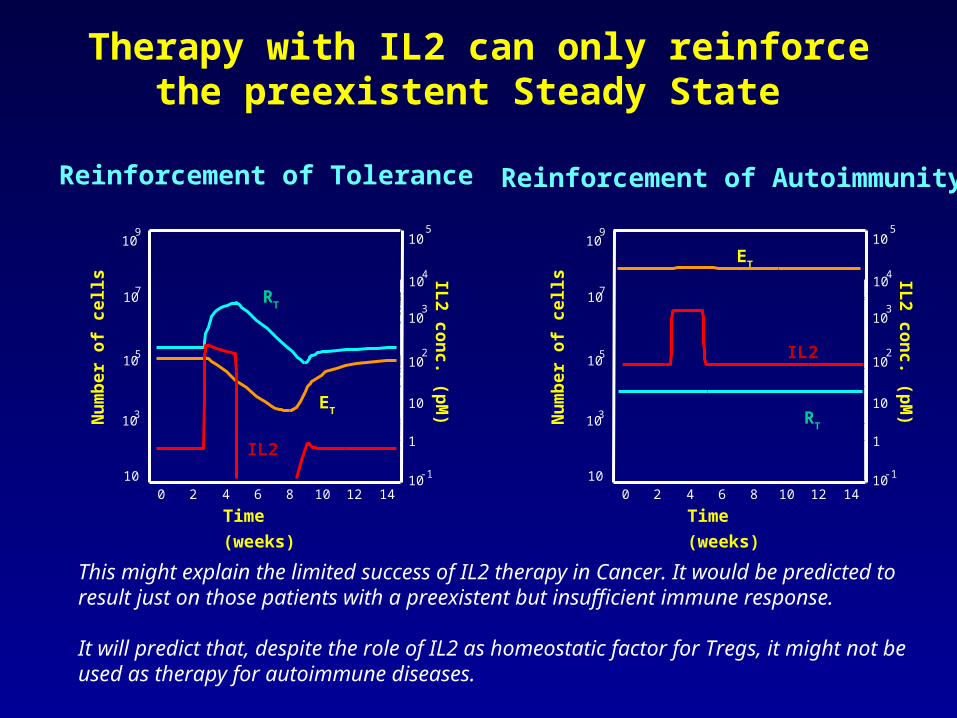
Therapy with IL2 can only reinforce the preexistent Steady State
Reinforcement of Tolerance Reinforcement of Autoimmunity
This might explain the limited success of IL2 therapy in Cancer. It would be predicted to result just on those patients with a preexistent but insufficient immune response.
It will predict that, despite the role of IL2 as homeostatic factor for Tregs, it might not be used as therapy for autoimmune diseases.
ET
Time (weeks)IL
2 con
c. (pM
)Nu
mb
er o
f ce
lls
0 2 4 6 8 10 12 1410
107
109
105
103
RT
102
104
1
10-1
105
10
103
IL2
ET
Time (weeks)
IL2 co
nc. (p
M)N
um
ber
of
cells
0 2 4 6 8 10 12 1410
107
109
105
103 R
T
102
104
1
10-1
105
10
103
IL2
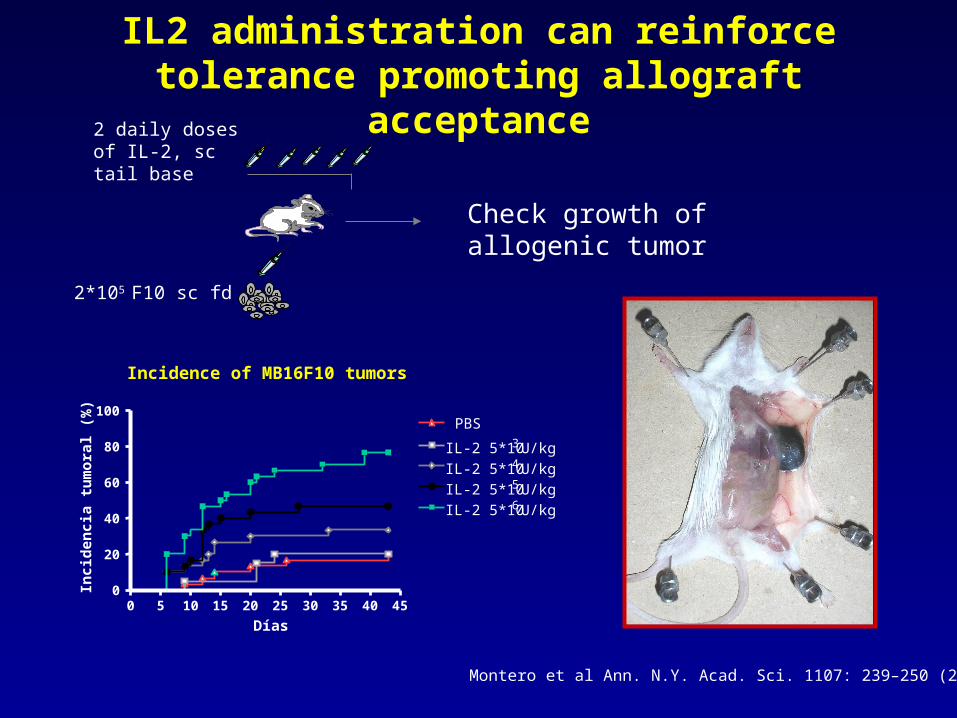
IL2 administration can reinforce tolerance promoting allograft acceptance
2*105 F10 sc fd
Check growth of allogenic tumor
2 daily doses of IL-2, sc tail base
Incidence of MB16F10 tumors
0 5 10 15 20 25 30 35 40 450
20
40
60
80
100PBS
IL-2 5*103 U/kg
IL-2 5*104 U/kg
IL-2 5*105 U/kg
IL-2 5*106 U/kg
Días
Inci
den
cia
tum
ora
l (%
)
Montero et al Ann. N.Y. Acad. Sci. 1107: 239–250 (2007).

IL2 administration can reinforce tolerance preventing EAE induction
Interestingly this tolerizing effect of IL2 or Il2+MAbs is only observed when it is administered before the onset the disease.
Montero et al Ann. N.Y. Acad. Sci. 1107: 239–250 (2007).
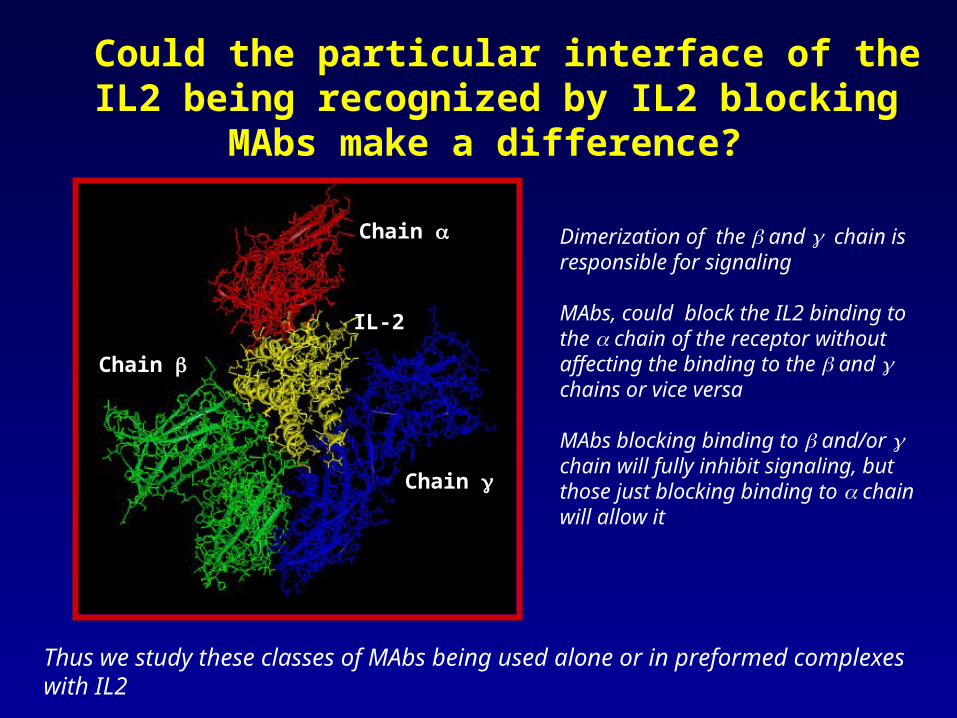
Could the particular interface of the IL2 being recognized by IL2 blocking MAbs make a difference?
Chain
Chain
Chain
IL-2
Dimerization of the and chain is responsible for signaling
MAbs, could block the IL2 binding to the chain of the receptor without affecting the binding to the and chains or vice versa
MAbs blocking binding to and/or chain will fully inhibit signaling, but those just blocking binding to chain will allow it
Thus we study these classes of MAbs being used alone or in preformed complexes with IL2
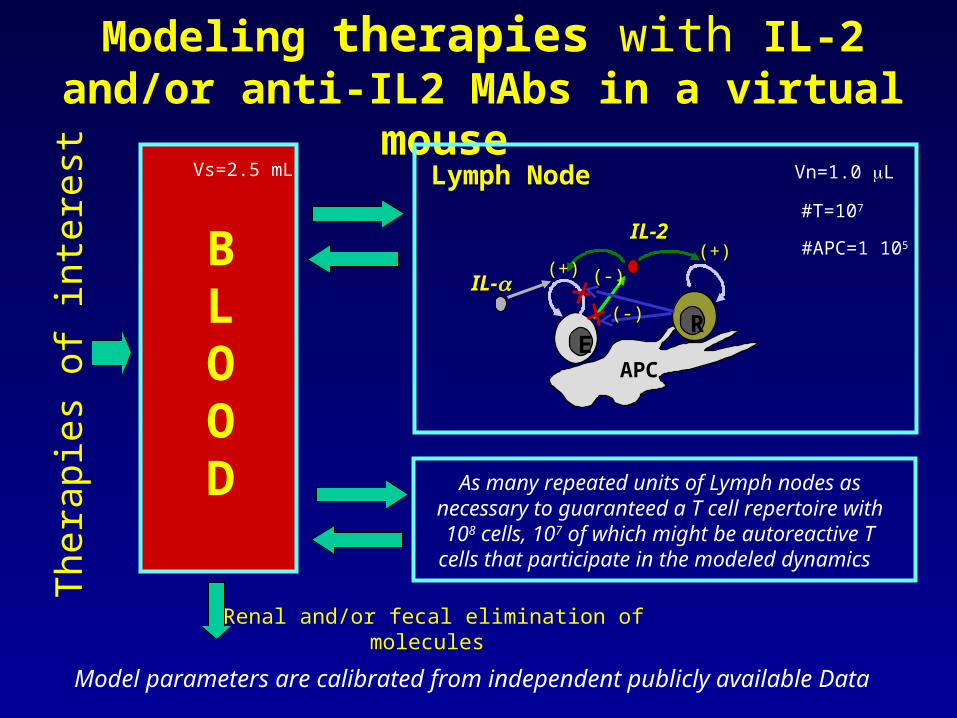
Modeling therapies with IL-2 and/or anti-IL2 MAbs in a virtual mouse
BLOOD
Vs=2.5 mL
ER
APC
IL-2(+)
(+)IL- (-)
(-)
As many repeated units of Lymph nodes as necessary to guaranteed a T cell repertoire with 108 cells, 107 of which might be autoreactive T cells that participate in
the modeled dynamics
Vn=1.0 LLymph Node
Renal and/or fecal elimination of molecules
#T=107
#APC=1 105
The
rapi
es o
f in
tere
st
Model parameters are calibrated from independent publicly available Data
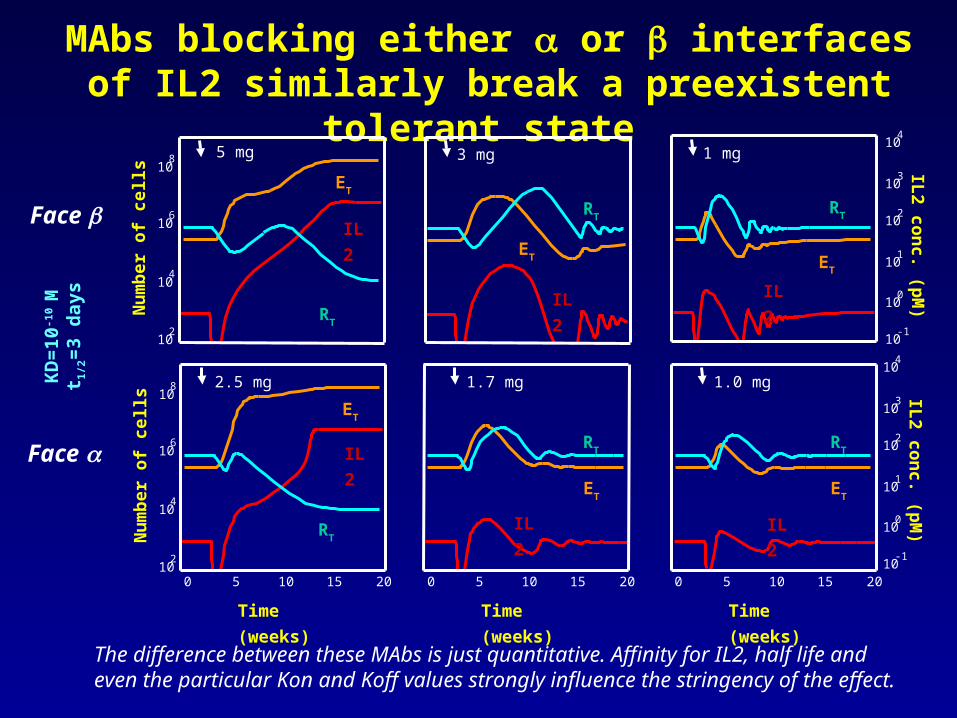
MAbs blocking either or interfaces of IL2 similarly break a preexistent tolerant state
106
108
102
104
Nu
mb
er o
f ce
lls
IL2
ET
RT
5 mg 104
103
102
101
100
10-1
IL2 co
nc. (p
M)
Face
Face
IL2
ET
RT
3 mg
IL2
ET
RT
1 mg
0 5 10 15 20
106
108
102
104
Nu
mb
er o
f ce
lls
IL2
ET
RT
Time (weeks)
2.5 mg
0 5 10 15 20
IL2
ET
RT
Time (weeks)
1.7 mg
0 5 10 15 20
104
103
102
101
100
10-1
IL2 co
nc. (p
M)IL2
ET
RT
Time (weeks)
1.0 mg
The difference between these MAbs is just quantitative. Affinity for IL2, half life and even the particular Kon and Koff values strongly influence the stringency of the effect.
KD=10-10 M
t1/2=3 days
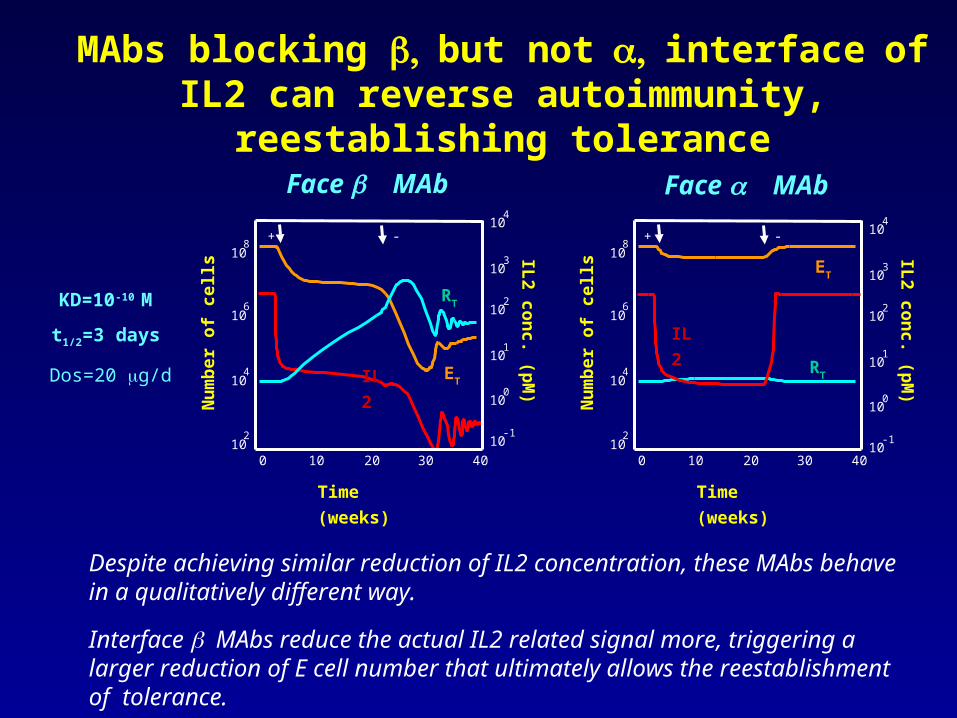
MAbs blocking but notinterfaceof IL2 can reverse autoimmunity, reestablishing tolerance
0 10 20 30 40
106
108
102
104
104
103
102
101
100
10-1
IL2 co
nc. (p
M)
Nu
mb
er o
f ce
lls
IL2
ET
RT
Time (weeks)
+ -
Face MAb
0 10 20 30 40
106
108
102
104
104
103
102
101
100
10-1
IL2 co
nc. (p
M)
Nu
mb
er o
f ce
lls
IL2 ET
RT
Time (weeks)
Face MAb
Despite achieving similar reduction of IL2 concentration, these MAbs behave in a qualitatively different way.
Interface MAbs reduce the actual IL2 related signal more, triggering a larger reduction of E cell number that ultimately allows the reestablishment of tolerance.
+ -
KD=10-10 M
t1/2=3 days
Dos=20 g/d

What about injecting Immune-complexes?
MAbs + IL2
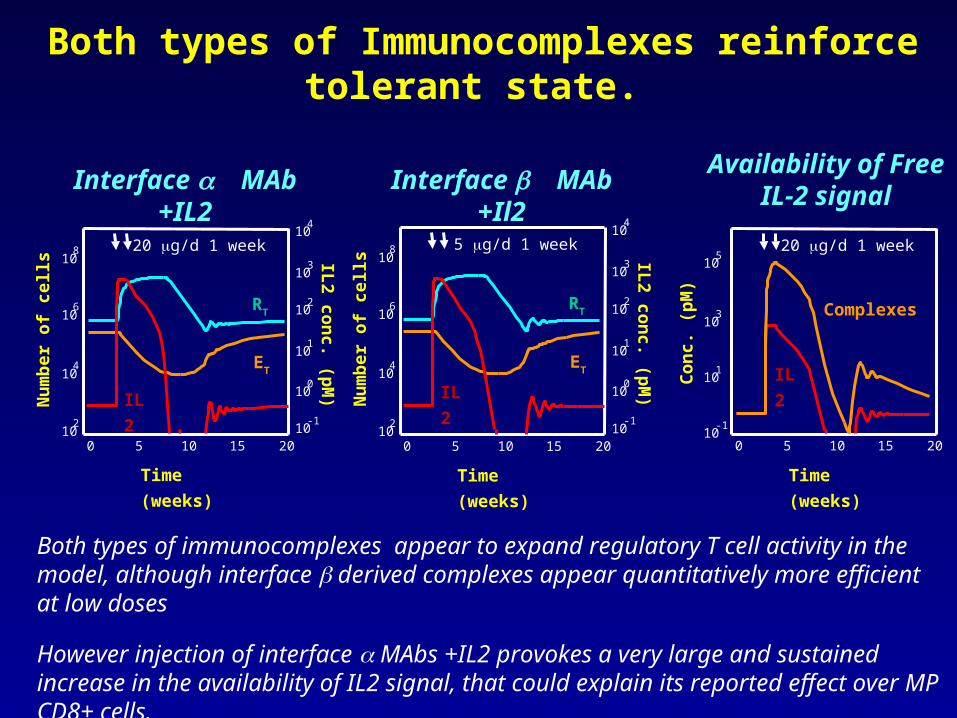
Both types of Immunocomplexes reinforce tolerant state.
Both types of immunocomplexes appear to expand regulatory T cell activity in the model, although interface derived complexes appear quantitatively more efficient at low doses
However injection of interface MAbs +IL2 provokes a very large and sustained increase in the availability of IL2 signal, that could explain its reported effect over MP CD8+ cells.
0 5 10 15 20
106
108
102
104
104
103
102
101
100
10-1
IL2 co
nc. (p
M)N
um
ber
of
cells
IL2
ET
RT
Time (weeks)
5 g/d 1 week
Interface MAb +IL2
Interface MAb +Il2
0 5 10 15 20
106
108
102
104
104
103
102
101
100
10-1
IL2 co
nc. (p
M)N
um
ber
of
cells
IL2
ET
RT
Time (weeks)
20 g/d 1 week
Availability of Free IL-2 signal
0 5 10 15 20
105
103
101
10-1
Co
nc.
(p
M)
IL2
Complexes
Time (weeks)
20 g/d 1 week
Immunocomplex with interface but notinterface anti- IL2 MAbs can reestablish Tolerant State.
106
108
102
104
104
103
102
101
100
10-1
IL2 co
nc. (p
M)
Nu
mb
er o
f ce
lls
IL2
ET
RT
Time (weeks)
0 10 20 30 40 50 60 70
0.3 mg/d 50week
Face MAb +IL2 Face MAb +IL2
106
108
102
104
104
103
102
101
100
10-1
IL2 co
nc. (p
M)
Nu
mb
er o
f ce
lls
IL2
ET
RT
Time (weeks)
0 10 20 30 40 50 60 70
0.3 mg/d 50week
Despite both types of immunocomplexes appearing to expand regulatory T cell activity in the model, only interface derived complexes appear capable of reversing the autoimmune state.
Conclusions through the Modeling relationship
ExperimentalObservations
Model
Postulates, Axioms
Predictions
),,,( 432111 NNNNF
dt
Nd
),,,( 432122 NNNNF
dt
Nd
),,,( 432133 NNNNF
dt
Nd

Model Predictions and explanationsIt explains:
It predicts:
• The Cross-regulation model is compatible with IL2 being a growth factor of both E and R cells.
• That in the absence of IL7 the immune system could be either in a steady state with a few E and R cells sustained by thymic output or in an autoimmune steady state.
• That IL2 depletion can induce autoimmunity and that the absence of thymus could favor this effect.
• That IL2 depletion treatments could favor tumor rejection or potentiate cancer vaccines.
• That IL2 injection treatment could be effective in a subset of cancer patients or can be good adjuvants for some vaccines.
• That Il2 injection treatment could reinforce tolerance and promote allograft acceptance if applied before graft implant.
1. That IL7 is the likely gamma chain family cytokine that helper cells use as an alternative to Il2 to growth in vivo
2. That Il2 depletion treatments can be used to reestablish tolerance in an autoimmune individual. (interface Mabs)
3. That IL2 treatment could only reinforce a preexistent immune state, thus is could be used to adjuvate vaccines, but applying it preferentially after the vaccine and not before.
4. That Il2 can not be used as a therapy to autoimmune diseases, despite being the main growth factors for Tregs. (The same goes for immunocomplexes)
5. That the lack of success of Il2 based therapies on cancer is a consequence of the heterogeneity of the preexistent immune response of patients to tumors.

Acknowledgements
CIMLic. Tania CarmenateM.Sc. Karina García Lic. Ileana GonzalezLic. Jose Antonio Dr. Enrique MonteroDr. Agustín Lage
IGC (Portugal)Dr. Jorge Carneiro
Financial support
IGC and CIM