
LGN 5799 - SEMINÁRIOS EMGENÉTICA E MELHORAMENTO DE PLANTAS
Programa de Pós-Graduação em Genética e Melhoramento de Plantas
Departamento de GenéticaAvenida Pádua Dias, 11 - Caixa Postal 83, CEP: 13400-970 - Piracicaba - São Paulo - Brasil
Telefone: (0xx19) 3429-4250 / 4125 / 4126 - Fax: (0xx19) 3433-6706 - http://www.genetica.esalq.usp.br/semina.php
METABOLISMO DE AMINOÁCIDOS
Aluna: Bertha Dévora Agurto BerdejoOrientador: Ricardo Antunes Azevedo

SUMÁRIO
� Introdução� Aminoácidos
�Vias Metabólicas� Diferentes vias
�Via Metabólica do Aspartato� Quatro essenciais
�Estratégias de Pesquisa� Três formas
�Considerações Finais

�O que são aminoácidos� Molécula que contém os grupos funcionais amina e
ácido carboxílico. � Formam a estrutura das proteínas
�Aminoácidos Essencias� São vinte tipos de aminoácidos incorporados nas
proteínas, � nove (lisina, treonina, metionina, fenilalanina, triptofano,
isoleucina, leucina, valina e histidina) são denominados essenciais � não são sintetizados por humanos e animais monogástricos, sendo
adquiridos nas dietas alimentares.
INTRODUÇÃO

INTRODUÇÃO
� Características
� As enzimas que atuam na síntese destes aminoácidos estão normalmente localizadas dentro dos cloroplastos das folhas ou em plastídeos de órgãos não fotossintéticos, como raiz e sementes.
� Principais moléculas de acumulo de nitrogênio para as plantas.
� Mais de 300 aminoácidos não protéicos foram isolados de plantas, os quais tem papel importante na medicina, nutrição e agricultura

� Importância
� Desempenham importantes funções� transportadores de nitrogênio para diferentes partes nos vegetais,
� reguladores em diversos processos envolvidos em resposta a diferentes condições ambientais,
� relacionados também com a qualidade nutricional das proteínas presentes nas sementes.
INTRODUÇÃO

� Síntese
� São sintetizados nos vegetais em complexas vias metabólicas sujeitas a uma regulação muito complexa e restrita para evitar o desperdício de energia e de nutrientes importantes como, carbono, nitrogênio e enxofre.
� A regulação das vias metabólicas é feita por enzimas, substratos e pelos próprios aminoácidos obtidos como produtos finais
INTRODUÇÃO

� Assimilação de Nitrogênio
� Normalmente, o nitrato é a principal fonte de nitrogênio disponível para as plantas.
� O nitrato absorvido pelas raízes é reduzido e incorporado na célula através de uma série de reações catalisadas por enzimas assimilatórias.
� Inicialmente, o nitrato absorvido é reduzido a nitrito em uma reação catalisada pela enzima nitrato redutase (NR, EC 1.6.6.1).
� Posteriormente, o nitrito é reduzido a amônio pela ação da enzima nitrito redutase (NiR, EC 1.7.7.1),
� O amônio é convertido em aminoácidos pela ação das enzimas glutamina sintetase e glutamato sintase (GS - GOGAT; EC 6.3.1.2 e EC 1.4.7.1, respectivamente)
INTRODUÇÃO

INTRODUÇÃO

COO-
C H
R
+H3N
aminoácidosNH4
+
amônia
N2 Intermediáriosda via glicolítica
Intermediáriosdo ciclo doácido cítrico
Intermediáriosda via
das pentoses
COO-
C H
R
Esqueletocarbônico
BIOSÍNTESE DE AMINOÁCIDOS

Via metabólica dos aminoácidos

Ser Gly Cys

Ser Gly Cys
As sínteses de serina e glicina
estão intimamente relacionadas
com a via de fotorrespiração nos
vegetais. São transportados para
outros órgãos e usados como
precursores da cisteína e da
glutationa.
Na ausência de fotorrespiração a serina também é sintetizada a partir de fosfoglicerato, uma vez sintetizado, o aminoácido serina éconvertido em glicina.

• Serina
O aminoácido glicina formado é, então, utilizado para a síntese de serina nas mitocôndrias.
• Glicina
O glicolato formado nos cloroplastos é oxidado a glioxalato e convertido a glicina nos peroxissomos.
• Cisteína
Regula a estrutura e a função das proteínas.
O mais importante constituinte da glutationa.
O esqueleto de carbono é cedido pela serina.
Ser Gly Cys

Tyr Phe Trp
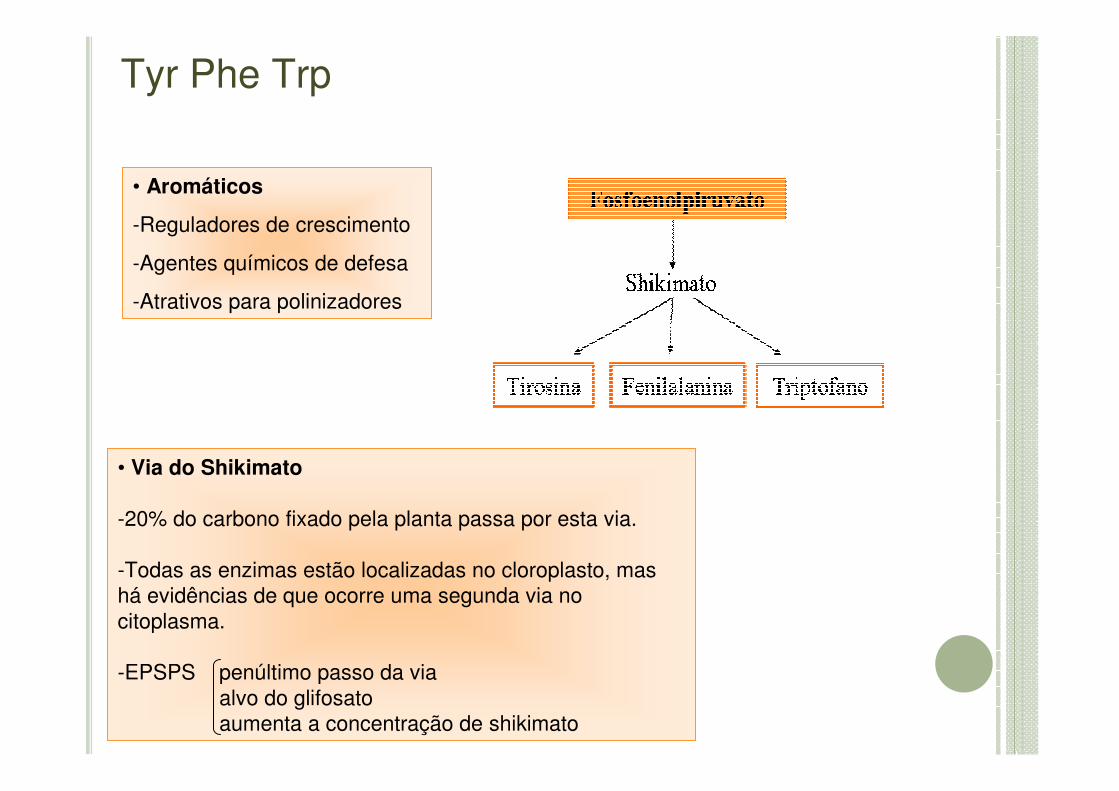
• Via do Shikimato
-20% do carbono fixado pela planta passa por esta via.
-Todas as enzimas estão localizadas no cloroplasto, mas há evidências de que ocorre uma segunda via no citoplasma.
-EPSPS penúltimo passo da viaalvo do glifosatoaumenta a concentração de shikimato
• Aromáticos
-Reguladores de crescimento
-Agentes químicos de defesa
-Atrativos para polinizadores
Tyr Phe Trp
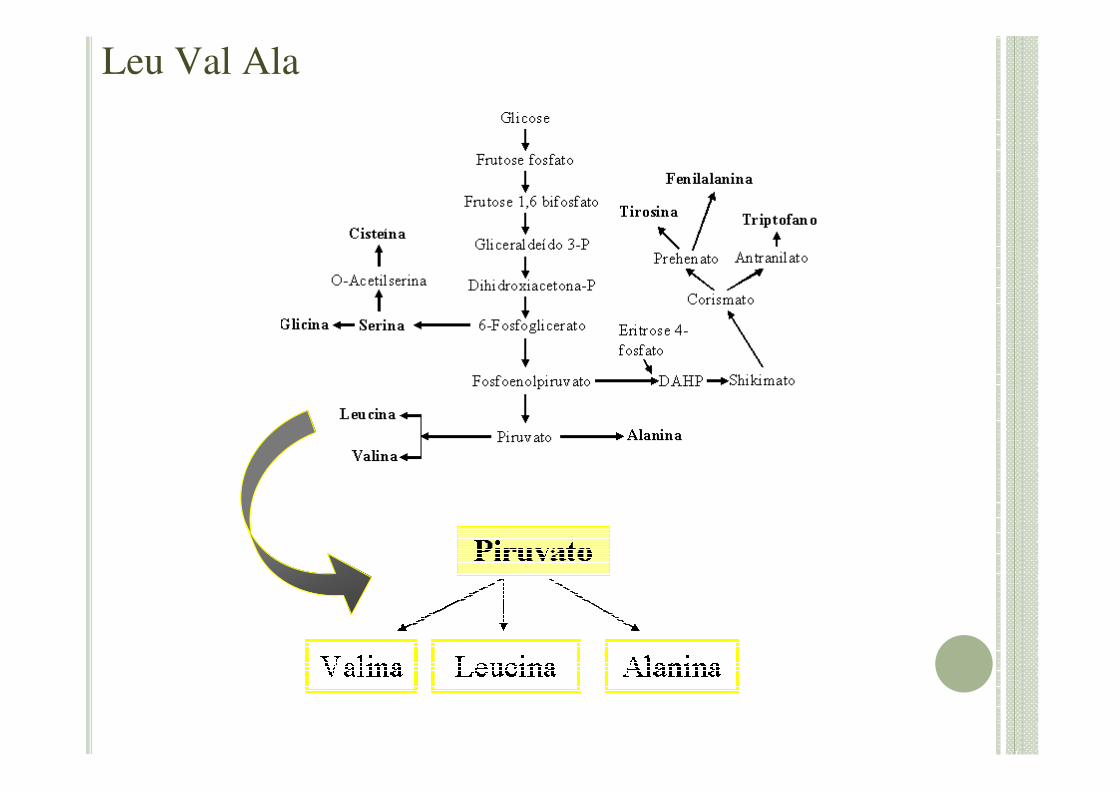
Leu Val Ala

• Leucina e Valina
As reações ocorrem no cloroplasto, o carbono é derivado apenas de duas moléculas de piruvato.
Provavelmente estão relacionados à degradação de aminoácidos.
As vias metabólicas de síntese de leucina e valina são consideradas bioquimicamente paralelas.
• Alanina
Um dos aminoácidos mais simples
Pouco reativo quimicamente
Reconhecimento de substratos em sítios ativos ou de regulação enzimática
Leu Val Ala

Glu Gln Pro Arg His
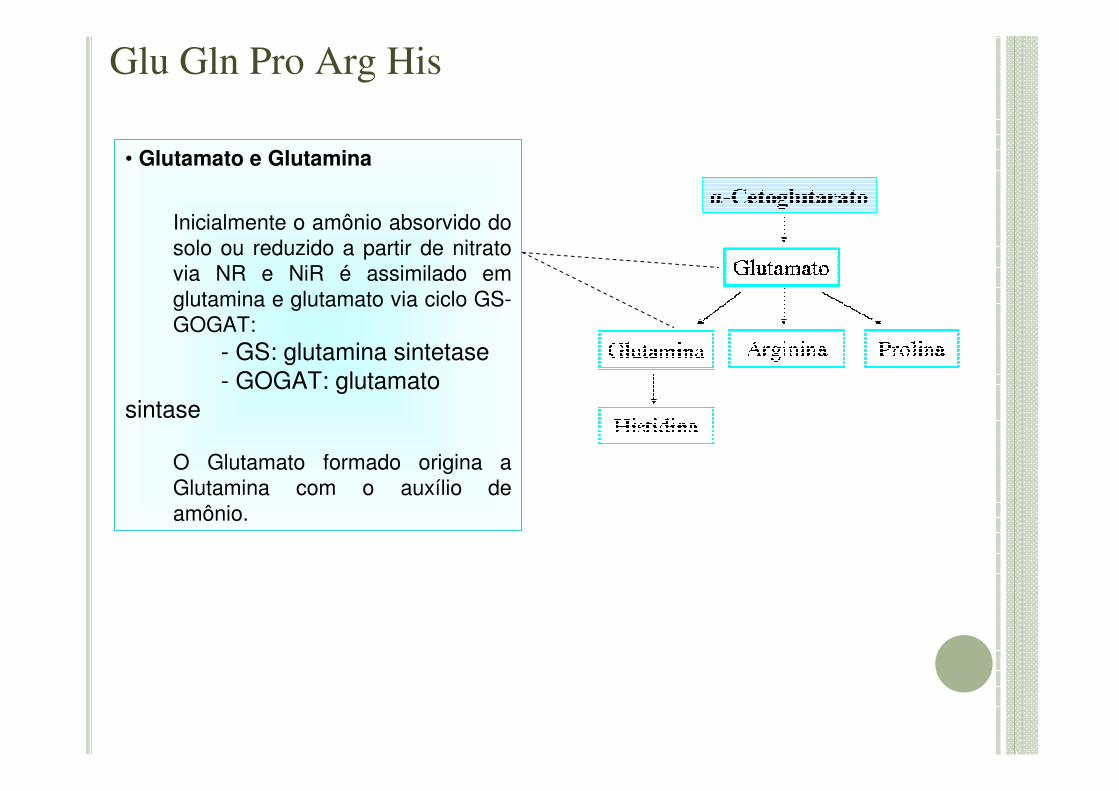
• Glutamato e Glutamina
Inicialmente o amônio absorvido do solo ou reduzido a partir de nitrato via NR e NiR é assimilado em glutamina e glutamato via ciclo GS-GOGAT:
- GS: glutamina sintetase- GOGAT: glutamato
sintase
O Glutamato formado origina a Glutamina com o auxílio de amônio.
Glu Gln Pro Arg His

• Prolina
A prolina é sintetizada em outro ramo em quatro reações enzimáticas;
O principal aminoácido das proteínas de sementes;
Papel importante como osmoprotetor na resistência à seca;
Estoque de energia para o crescimento do tubo polínico;
Fixação de nitrogênio nos nódulos das raízes de leguminosas.
Glu Gln Pro Arg His
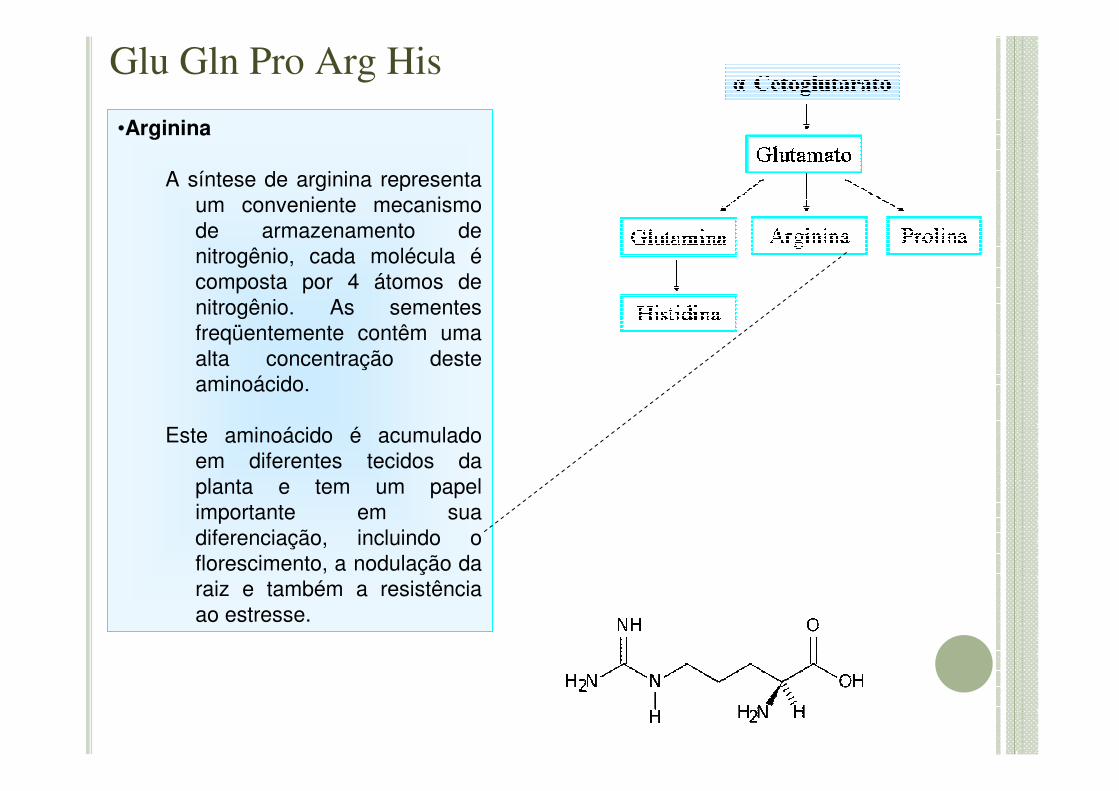
Glu Gln Pro Arg His
•Arginina
A síntese de arginina representa um conveniente mecanismo de armazenamento de nitrogênio, cada molécula écomposta por 4 átomos de nitrogênio. As sementes freqüentemente contêm uma alta concentração deste aminoácido.
Este aminoácido é acumulado em diferentes tecidos da planta e tem um papel importante em sua diferenciação, incluindo o florescimento, a nodulação da raiz e também a resistência ao estresse.

•Histidina:
- Aminoácido não essencial que apresenta funções essenciais como a manutenção do status de oxi-redução(Boldyrev, 1999).
- A quelação dos metais através de biomolécula serve como um mecanismo de defesa utilizado por algumas espécies para resistir a exposição a íons de metais tóxicos. Nas plantas denominadas hiperacumuladoras, por exemplo, a histidina é conhecido por iniciar a principal função da quelação e transporte de metais como zinco, níquel, cobalto e cobre lidando com a intolerância do metal (Lasat, 2000).
Glu Gln Pro Arg His

VIA METABÓLICA DO ÁCIDO ASPÁRTICO
� Devido a baixa concentração de lisina e treonina nas sementes de cereais, e sua importância como aminoácidos essenciais, estudos tem sido realizados para se obter um melhor entendimento da regulação da via metabólica.

• O AspartatoAspartato atua como
precursor comum em duas
vias metabólicas
A primeira conduz
à síntese do aminoácido
asparagina,
A segunda conduz à
síntese de quatro dos nove
aminoácidos essenciais:
lisina, treonina, metionina e
isoleucina.
Aspartato
β-Aspartil fosfato
AsparaginaAK
HSDHDHDPS
Lisina
Homoserina
Treonina
Metionina
Isoleucina
TS
β-aspartil fosfato
Asn Asp Lys Met Thr Ile
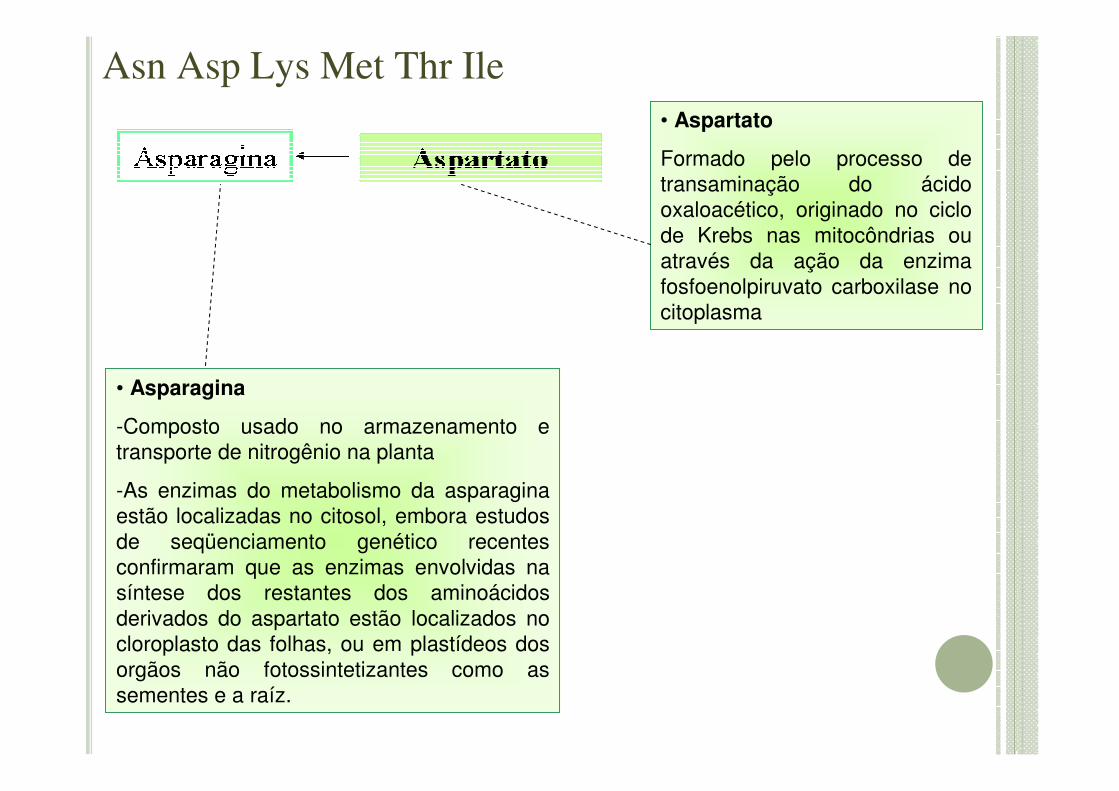
• Asparagina
-Composto usado no armazenamento e transporte de nitrogênio na planta
-As enzimas do metabolismo da asparaginaestão localizadas no citosol, embora estudos de seqüenciamento genético recentes confirmaram que as enzimas envolvidas na síntese dos restantes dos aminoácidos derivados do aspartato estão localizados no cloroplasto das folhas, ou em plastídeos dos orgãos não fotossintetizantes como as sementes e a raíz.
• Aspartato
Formado pelo processo de transaminação do ácido oxaloacético, originado no ciclo de Krebs nas mitocôndrias ou através da ação da enzima fosfoenolpiruvato carboxilase no citoplasma
Asn Asp Lys Met Thr Ile

• HSDH
A enzima HSDH catalisa a síntese de homoserina
Foi primeiramente estudada em procariotos e posteriormente em vegetais.
Duas isoenzimas da HSDH têm sido observadas em vegetais
uma sensível a treonina, no citoplasmaoutra resistente à inibição por treonina.
• AK
Primeira enzima da via metabólica do ácido aspártico.
Inicialmente a AK foi estudada em microrganismos e posteriormente em diversas espécies vegetais como milho, arroz, sorgo
Duas isoenzimas distintas da AK têm sido descritas:
Isoenzima monofuncional e sensível à inibição por lisina, predominante
Presente em um polipeptídeo bifuncional apresentando um domínio de AK e outro de HSDH, sendo sensível à inibição por treonina
β-aspartil fosfato
Asn Asp Lys Met Thr Ile
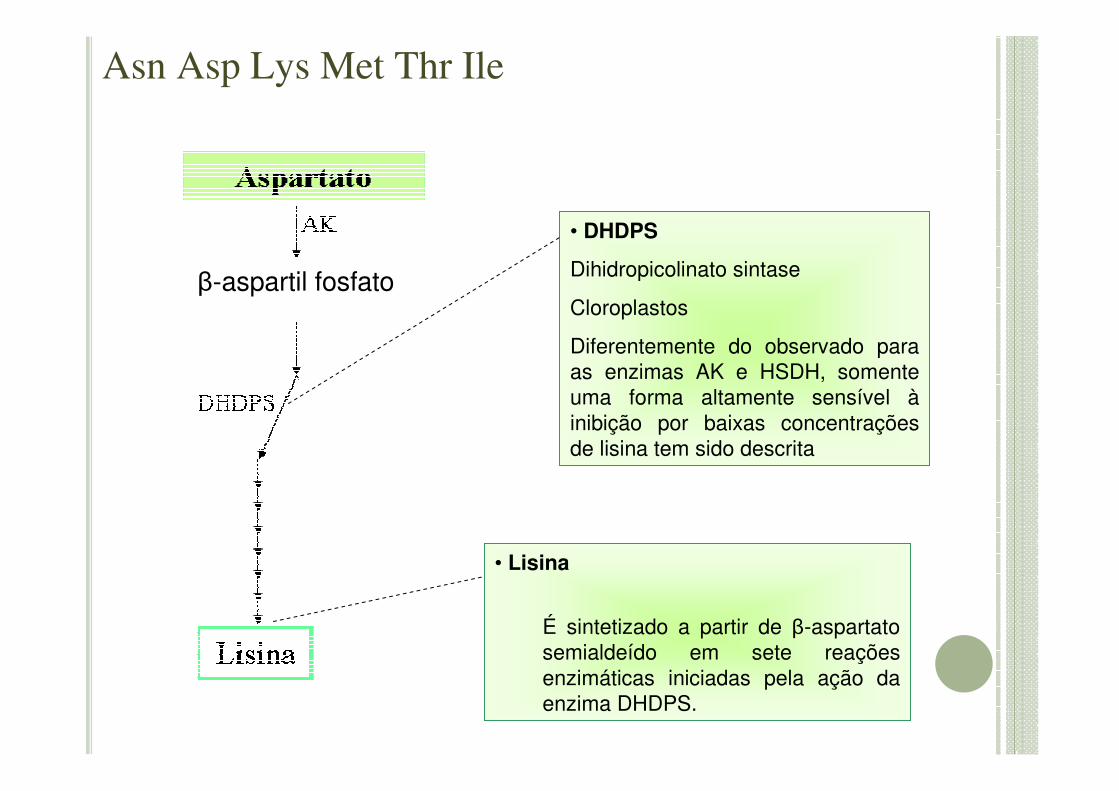
• DHDPS
Dihidropicolinato sintase
Cloroplastos
Diferentemente do observado para as enzimas AK e HSDH, somente uma forma altamente sensível àinibição por baixas concentrações de lisina tem sido descrita
β-aspartil fosfato
• Lisina
É sintetizado a partir de β-aspartatosemialdeído em sete reações enzimáticas iniciadas pela ação da enzima DHDPS.
Asn Asp Lys Met Thr Ile

LOR
SDH
Lisina
Sacaropina
2-aminoadipato semialdeído
+
glutamato
• Catabolismo de Lisina
Duas enzimas estão envolvidas no catabolismo de lisina
LOR, lisina 2-oxoglutarato redutaseSDH, sacaropina desidrogenase
Embora as enzimas LOR e SDH possam estar presentes em vegetais como enzimas monofuncionais, a maior atividade da LKR e SDH está presente em um polipeptídeo bifuncional
O aminoácido lisina pode regular seu próprio catabolismo com as enzimas sendo moduladas diferencialmente em uma cascata de sinais intracelular envolvendo principalmente cálcio e um processo de fosforilação-desfosforilação da proteína.
Asn Asp Lys Met Thr Ile
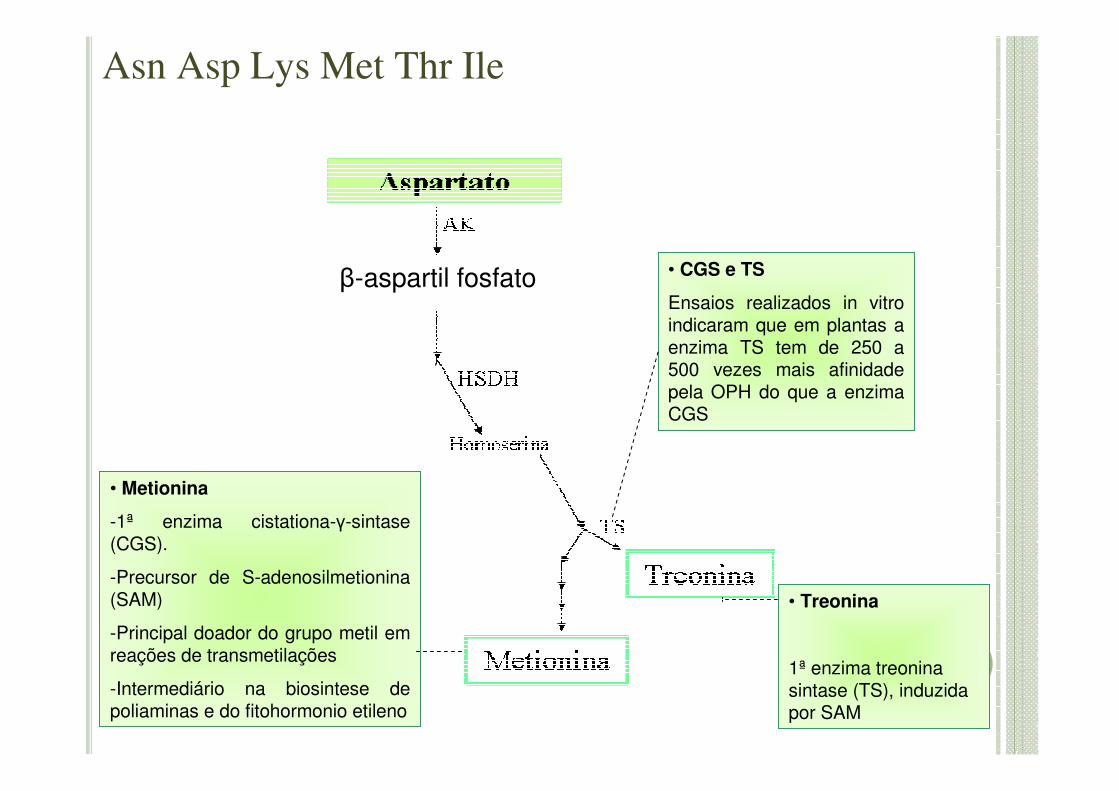
β-aspartil fosfato • CGS e TS
Ensaios realizados in vitroindicaram que em plantas a enzima TS tem de 250 a 500 vezes mais afinidade pela OPH do que a enzima CGS
• Metionina
-1ª enzima cistationa-γ-sintase(CGS).
-Precursor de S-adenosilmetionina(SAM)
-Principal doador do grupo metil em reações de transmetilações
-Intermediário na biosintese de poliaminas e do fitohormonio etileno
• Treonina
1ª enzima treonina sintase (TS), induzida por SAM
Asn Asp Lys Met Thr Ile

β-aspartil fosfato
• Isoleucina
Treonina deaminase (TD) é a primeira enzima na síntese de Isoleucina.
Há duas isoformas de TD
-Biossintética: presente em tecido jovem em desenvolvimento, que sofre inibição por isoleucina
-Biodegradativa: presente em tecido velho, senescente, não sensível à inibição.
Asn Asp Lys Met Thr Ile
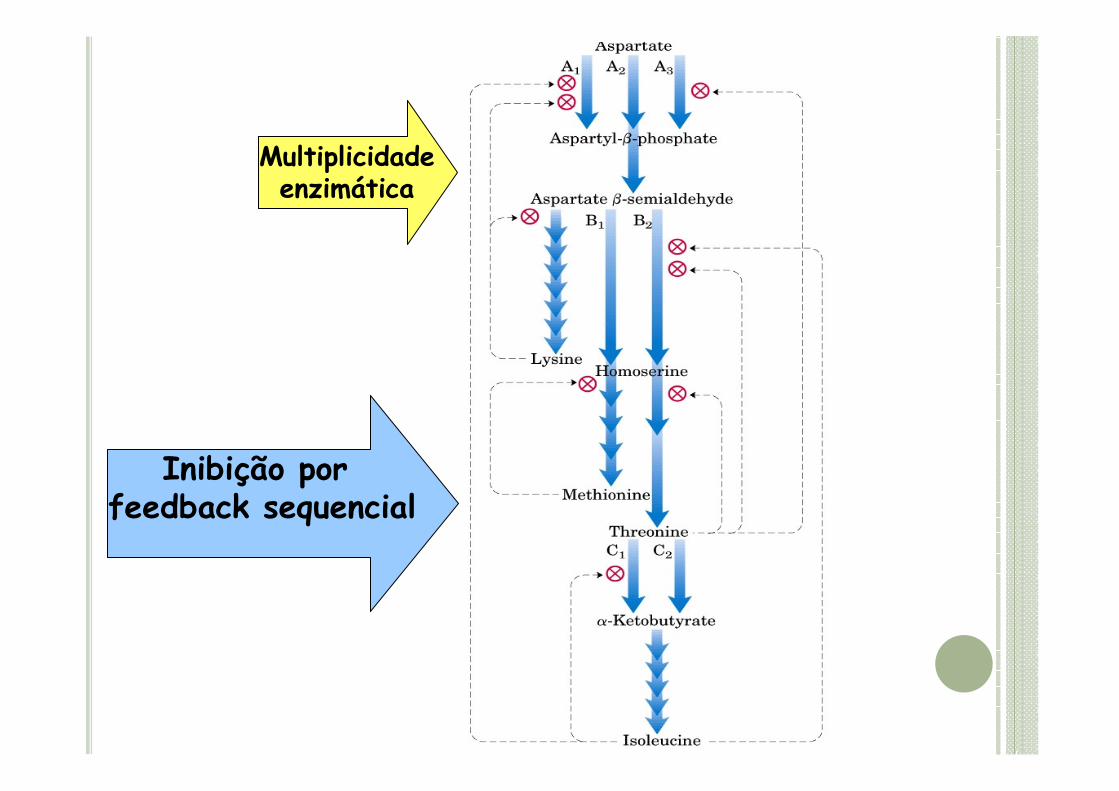
Inibição porfeedback sequencial
Multiplicidadeenzimática

� Os estudos bioquímicos, genéticos e moleculares da via do ácido aspártico levaram à compreensão de mecanismos importantes para a manipulação metabólica da via e para a elaboração de estratégias para produção de cereais com alto teor de lisina nas sementes.
Estratégias de Pesquisa

� Quatro principais estratégias têm sido utilizadas para produção de cereais que acumulam alta concentração de lisina� Melhoramento genético
� QPM
� Identificação de mutantes naturais
� Produção de plantas transgênicas
Estratégias de Pesquisa

• Objetivos
Foi realizado um estudo, em sementes imaturas, com as enzimas envolvidas na biossíntese de lisina, para obter o seu padrão de atividade.
• Quinoa
Importante fonte de proteínas devido a sua digestibilidade e composição balanceada de aminoácidos.
Pseudocereal
Identificação de Mutantes Naturais

As enzimas apresentaram um resultado similar nas três etapas de desenvolvimento indicando que qualquer uma das três etapas pode ser usado como fonte de atividade para o estudo das propriedades reguladoras das enzimas.
Assim para o estudo de atividade foram utilizadas as sementes com 20 DAP
Identificação de Mutantes Naturais
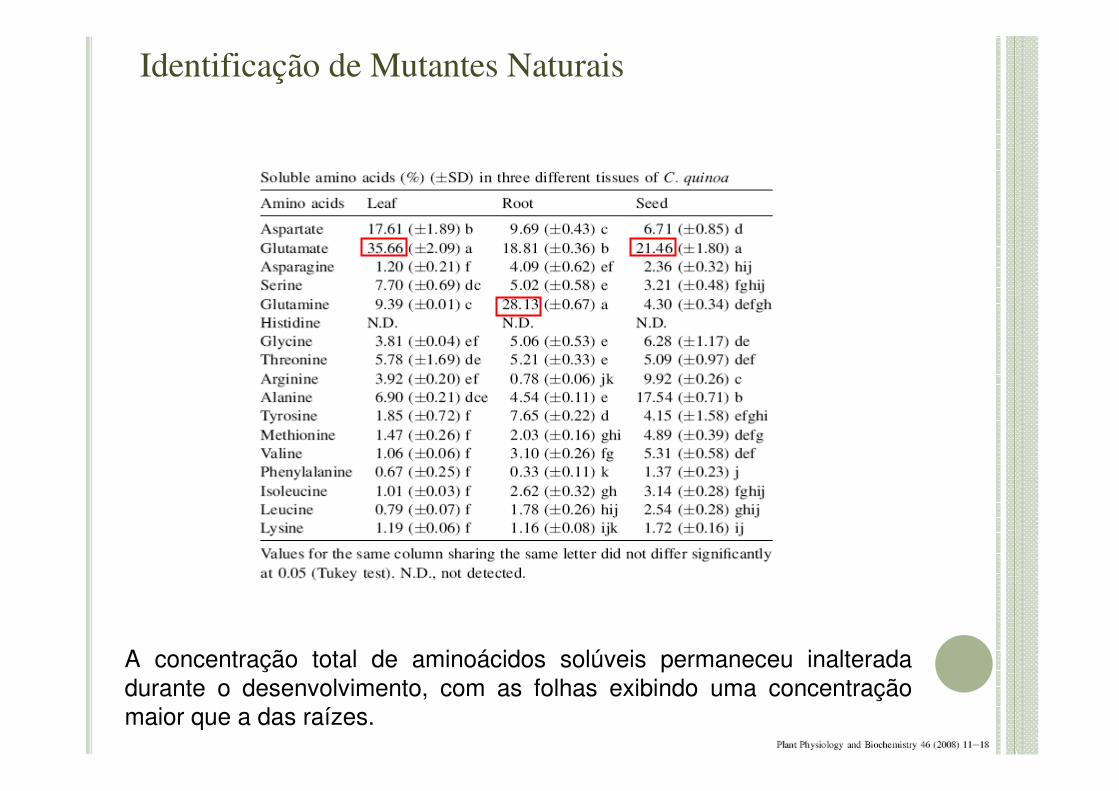
A concentração total de aminoácidos solúveis permaneceu inalterada durante o desenvolvimento, com as folhas exibindo uma concentração maior que a das raízes.
Identificação de Mutantes Naturais

Lisina nas concentrações de 1 e 5 mM inibiram a atividade de AK em 19,5% e 23,8% respectivamente.
Treonina nas concentrações de 1 e 5 mM inibiram a atividade de AK em 45,2% e 49,7%, respectivamente.
Lys +Thr nas concentrações de 1 e 5 mM inibiram a atividade de AK 37,6% e 47,6%, respectivamente.
Controle lisina treonina metionina S-2-aminoetil L-cisteina
Sementes com 20 DAP
Identificação de Mutantes Naturais
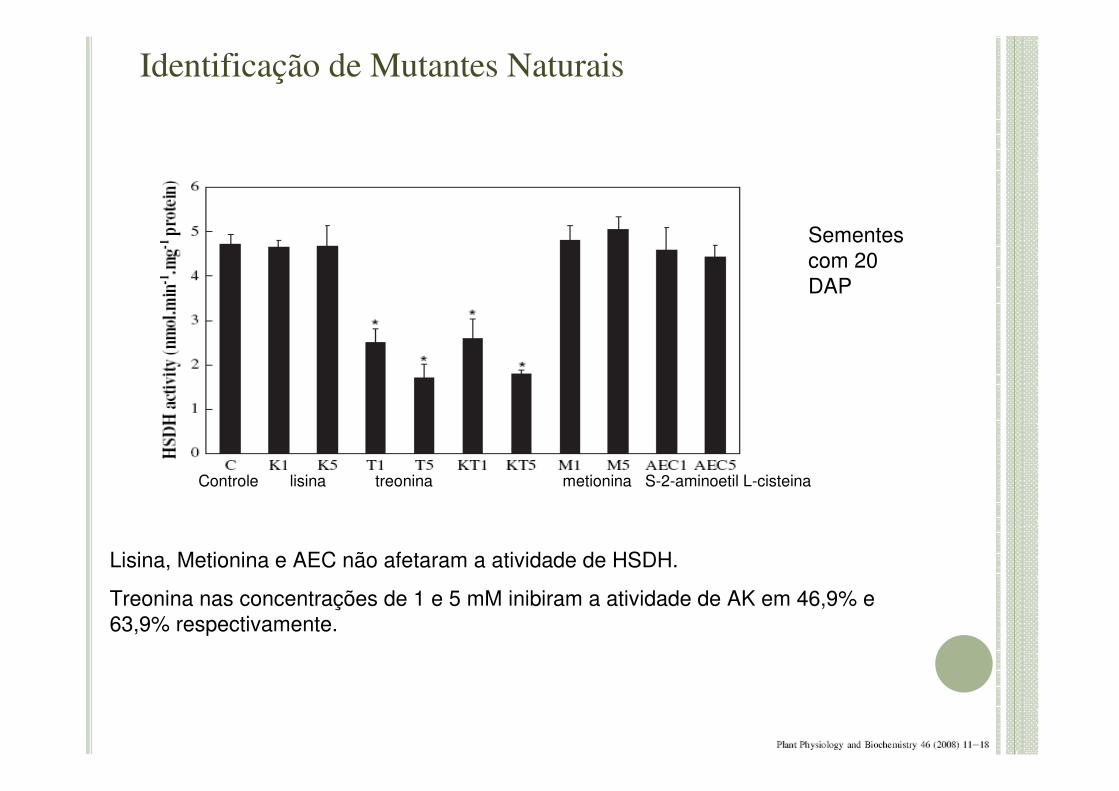
Lisina, Metionina e AEC não afetaram a atividade de HSDH.
Treonina nas concentrações de 1 e 5 mM inibiram a atividade de AK em 46,9% e 63,9% respectivamente.
Sementes com 20 DAP
Controle lisina treonina metionina S-2-aminoetil L-cisteina
Identificação de Mutantes Naturais

A adição de lisina na mistura com treonina, nas concentrações de 1 e 5 mM induziu uma forte inibição da atividade de DHDPS, 65,3% e 69,9%, respectivamente.
AEC nas concentrações de 1 e 5 mM inibiram a atividade de DHDPS em 28,7%e 60,1%, respectivamente.
A treonina e a metionina não alteraram a atividade de DHPS
Sementes com 20 DAP
Controle lisina treonina metionina S-2-aminoetil L-cisteina
Identificação de Mutantes Naturais

• Conclusões
-Presença de duas isoenzimas de AK
-Maior concentração de treonina nas sementes de Quinoa
-Presença de duas isoenzimas de HSDH
-Uma isoenzima de DHDPS altamente sensível à inibição por lisina
-Alta concentração de lisina (alta síntese de lisina e acumulo na forma solúvel)
Identificação de Mutantes Naturais

� O estudo dos mutantes apresentando alterações na sensibilidade das enzimas AK e DHDPS aos aminoácidos lisina, treonina e ao AEC, contribuíram para a compreensão dos mecanismos regulatórios envolvidos na via metabólica do ácido aspártico, principalmente aos relacionados à regulação das enzimas chave da síntese de lisina AK e DHDPS. O uso de plantas transgênicas se revelou como uma importante metodologia para o estudo desta via metabólica.
Produção de Plantas Transgênicas

• Objetivo
Aumentar o acumulo de lisina livre combinando duas vias de transformação de milho.
-CordapA (Corynebacterium glutamicum) gene com expressão resistente ao feedback de lisina.
-Supressão do gene LKR/SDH
Produção de Plantas Transgênicas
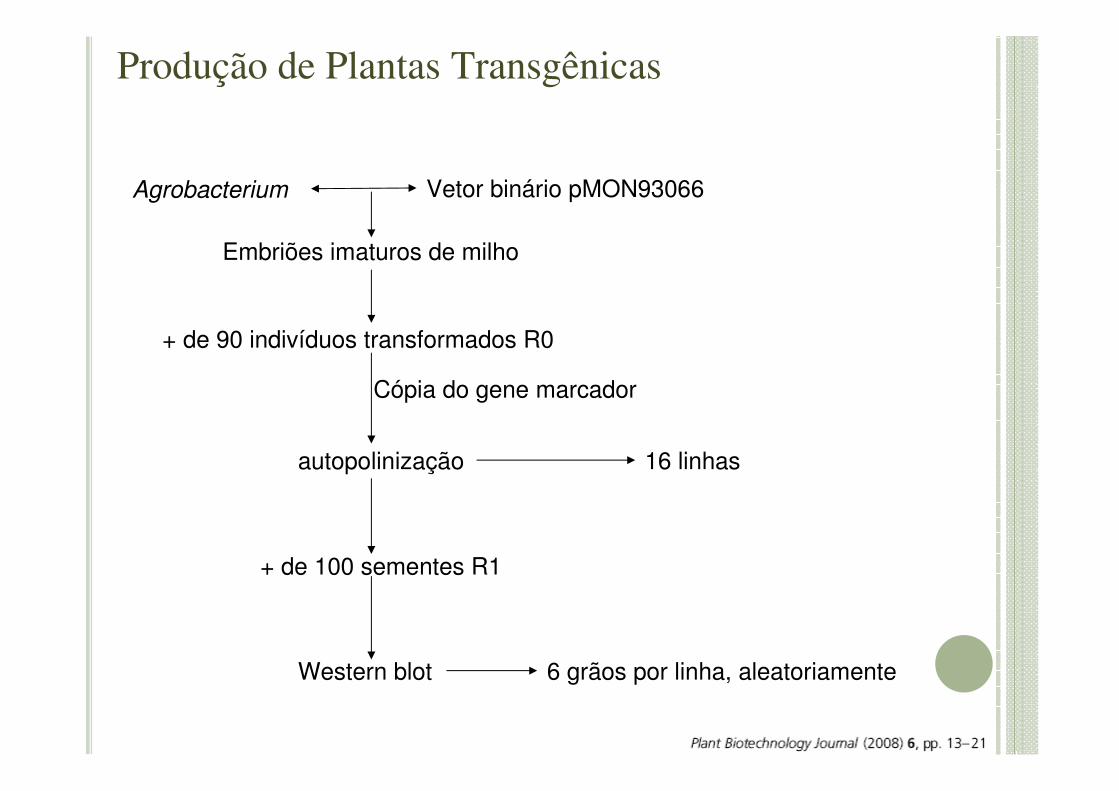
Embriões imaturos de milho
Agrobacterium Vetor binário pMON93066
+ de 90 indivíduos transformados R0
Cópia do gene marcador
autopolinização 16 linhas
+ de 100 sementes R1
Western blot 6 grãos por linha, aleatoriamente
Produção de Plantas Transgênicas

• Western blot
A maioria das linhas 11/16 apresentaram expressão de CordapA e redução de LKR/SDH (a).
Duas linhas mostraram expressão do CordapA, mas sem redução de LKR/SDH (b).
Uma linha não apresentou expressão de CordapA, mas redução de LKR/SDH (c).
Duas linhas não apresentaram nem expressão, nem redução.
baa c
Produção de Plantas Transgênicas

Produção de Plantas Transgênicas

A análise de lisina livre em sementes maduras da R2, demonstrou que não ocorre acumulo de lisina nos grãos maduros de milho, quando transformados com apenas o gene CordapA.
Na transformação M115160, na qual a expressão de CordapA não foi significativa, o acumulo de lisina foi ~ 2x menor quando comparado com o acumulo nas transformações com expressão do CordapA e redução do LKR/SDH.
M – transformãção com os genes CordapA e LKR/SDH
S – transformação com o gene CordapA
Produção de Plantas Transgênicas

• Conclusão
-Nova metodologia para aumentar a concentração de lisina em grãos de milho maduro.
-Transgene com CordapA e a supressão do gene LKR/SDH
-Constatou-se um aumento no acumulo de lisina solúvel de 2x e de 3x no total de lisina nas sementes transgênicas.
Produção de Plantas Transgênicas

CONSIDERAÇÕES FINAIS
Algumas enzimas secundárias da via metabólica do aspartato ainda não tiveram sua regulação esclarecida por completo;
Entretanto, para as principais enzimas reguladoras da vai ou para aquelas que são alvo da ação de herbicidas, grandes avanços científicos foram alcançados;
Já foram clonados e expressos em E. coli diversos genes das enzimas da via metabólica, com isto foi possível a obtenção da estrutura cristalizada das enzimas, assim como o estudo de seu mecanismo de reação.

CONSIDERAÇÕES FINAIS
Espécies transgênicas de cereais e leguminosas, contendo o aumento ou redução da atividade de uma enzima tem demonstrado que a regulação da via pode ser dramaticamente alterada.
A meta de produzir linhagens comerciais viáveis com uma maior qualidade protéica, aumentando o conteúdo de aminoácidos essenciais nas sementes ainda tem que ser alcançada.

OBRIGADA!







