
Mucoid morphotype variation in Burkholderia multivorans:
Role of a Two-Component Regulatory System and a LysR
Regulator
Ana Rita Martins Guerreiro
Thesis to obtain the Master of Science Degree in
Biotechnology
Supervisors: Doctor Leonilde de Fátima Morais Moreira
Doctor Inês Nunes Silva
Examination Comitee:
Chairperson: Doctor Arsénio do Carmo Sales Mendes Fialho
Supervisor: Doctor Leonilde de Fátima Morais Moreira
Members of the Committee: Doctor Dalila Madeira Nascimento Mil-Homens
December, 2015

Mucoid morphotype variation in Burkholderia multivorans:
Role of a Two-Component Regulatory System and a LysR
Regulator
Ana Rita Martins Guerreiro
Thesis to obtain the Master of Science Degree in
Biotechnology
Supervisors: Doctor Leonilde de Fátima Morais Moreira
Doctor Inês Nunes Silva
Examination Comitee:
Chairperson: Doctor Arsénio do Carmo Sales Mendes Fialho
Supervisor: Doctor Leonilde de Fátima Morais Moreira
Members of the Committee: Doctor Dalila Madeira Nascimento Mil-Homens
December, 2015

i
ACKNOWLEDGMENTS
This work would not be possible without the help and support of several important people and
entities.
First, I would like to address a special acknowledgment to my supervisor Prof. Leonilde
Moreira for the opportunity to work in her lab, for all the support and advices during my research.
I would also like to address a special acknowledgment to my co-supervisor Doctor Inês Nunes
Silva for the constant support in the lab, for the encouragement to follow my own ideas and for her
patience and dedication in all stages of this work.
I would also thank to Marcelo Ramires and Inês Correia for their contribution in this work,
availability and dedication.
I also thank to every member of the Biological Sciences Research Group (BSRG) that directly
or indirectly contributed to this work and to my integration.
Financial support by FCT (contract: PTDC/QUI-BIQ/118260/2010) is gratefully acknowledge.
Finally, I would like to say a special thanks to my family and friends for always being there for
me and for giving me all the support, trust and motivation essential for the development of this work.

ii
RESUMO
As bactérias do complexo Burkholderia cepacia (Bcc) são um grupo de microrganismos
geneticamente relacionados, capazes de induzir infecções crónicas em pacientes com fibrose quística
(FQ). Durante as infecções crónicas com bactérias do Bcc ocorrem variações do fenótipo mucoso
para não-mucoso, estando os dois fenótipos associados a diferentes propriedades fenotípicas e à
expressão diferencial de vários genes que codificam reguladores de transcrição, incluindo um
regulador transcripcional do tipo LysR (LTTR), codificado pelo gene Bmul_2557. Neste trabalho, foi
desenvolvida uma estratégia para a complementação do mutante isogénico ΔBmul_2557::dhfR de
Burkholderia multivorans ATCC 17616 e realizada uma caracterização fenotípica da estirpe
selvagem, do mutante de eliminação isogénico e da estirpe mutante complementada. Quando se
realizou um estudo para avaliar a produção de exopolissacárido (EPS) em meio contendo manitol,
observou-se que todas as estirpes produzem níveis semelhantes, excluindo um papel directo deste
regulador na regulação da biossíntese de EPS. Quando foi testado o crescimento das estirpes em
meios contendo diferentes fontes de carbono, observou-se que na presença de D-glucose, ocorre a
lise celular da estirpe B. multivorans ATCC 17616, bem como uma acidificação irreversível do meio
de cultura, enquanto que no mutante de eliminação este efeito não é observado. A identificação dos
metabolitos presentes no sobrenadante das culturas revelou a acumulação de ácido 2-ketoglucónico
e D-lactato na estirpe selvagem, metabolitos resultantes do consumo da glucose através da via
oxidativa e da conversão do excesso de piruvato em D-lactato, respectivamente. De modo a verificar
se a lise celular, na presença de glucose, também ocorre noutras estirpes de Burkholderia, as
estirpes B. multivorans D2095, B. contaminans IST408 e B. multivorans HI2229 também foram
testadas. Os resultados mostraram que as estirpes B. multivorans HI2229 e B. multivorans D2095
apresentam um perfil semelhante à B. multivorans ATCC 17616, no entanto, na estirpe B.
contaminans IST408 não ocorreu lise celular, sugerindo que este fenómeno pode ser específico da
estirpe/espécie. Também foi investigada a relevância de um regulador de resposta do tipo OmpR na
variação do fenótipo mucoso. Um plasmídeo contendo o gene ompR foi mobilizado, por conjugação
triparental, para variantes não mucosos da estirpe B. multivorans. O fenótipo mucoso foi restaurado
em todas as estirpes testadas, sugerindo que o fenótipo não-mucoso se deve a mutações neste
gene. A sequenciação do genoma de 5 desses variantes não mucosos confirmou a presença de
mutações no gene ompR, afectando quer o local de ligação ao DNA, quer o local de ligação ao
substrato por parte desse regulador. Em suma, a análise funcional de reguladores do tipo LysR e
OmpR contribui para a compreensão das características metabólicas/virulência do Bcc, direccionando
os estudos futuros sobre os mecanismos moleculares da variação de fenótipo nestas bactérias.
PALAVRAS-CHAVE
Burkholderia multivorans; Regulador transcripcional do tipo LysR; Metabolismo da glucose; Variação
de morfotipo mucoso; Sistemas reguladores de dois componentes

iii
ABSTRACT
Bacteria from Burkholderia cepacia complex (Bcc) is a group of genetically related
microorganisms causing chronic infections in patients with Cystic Fibrosis (CF). During chronic
infection with Bcc, mucoid-to-nonmucoid morphotype variation occurs, with the two morphotypes
exhibiting different phenotypic properties and consistent differential expression of several genes
encoding transcriptional regulators, including a LysR-type transcriptional regulator (LTTR)-encoded by
Bmul_2557 gene. Here we developed a strategy for the complementation of the ΔBmul_2557::dhfR
isogenic mutant of Burkholderia multivorans ATCC 17616 and performed a phenotypic
characterization of the wild-type, isogenic deletion mutant and complemented strain.
Exopolysaccharide production assessed in mannitol-containing medium showed similar levels in all
tested strains, excluding a direct role of this regulator in EPS biosynthesis regulation. Growth using
different carbon sources was assessed and cell lysis of the B. multivorans ATCC 17616 in the
presence of D-glucose was observed as well as an irreversible culture supernatant acidification,
whereas in the deletion mutant this effect was not seen. Identification of metabolites present in culture
supernatants revealed the accumulation of 2-ketogluconic acid and D-lactate in wild-type culture,
metabolites resulting from glucose consumption through the oxidative pathway and from the
conversion of excess pyruvate into D-lactate, respectively. To ascertain if cell lysis in the presence of
glucose also occurs in other Burkholderia strains, we cultured B. multivorans D2095, B. contaminans
IST408 and B. multivorans HI2229. Results showed that B. multivorans HI2229 and B. multivorans
D2095 presented a similar profile of B. multivorans ATCC 17616, however, in B. contaminans IST408
cell lysis did not occur, which suggest that this phenomenon can be strain/species specific. The
relevance of an OmpR-like response regulator in Burkholderia mucoid morphotype variation was also
studied. A plasmid containing the ompR gene was mobilized into nonmucoid B. multivorans variants
by triparental mattings. In all tested strains, the mucoid phenotype was restored, suggesting that
nonmucoid phenotype is due to mutations in this gene. Whole-genome sequencing of 5 nonmucoid
variants confirmed the presence of mutation in the ompR-like gene affecting either the DNA-binding
site on the substrate-binding site of this regulator. Overall, the functional analysis of the LysR-type and
OmpR-type regulators contribute to the understanding of metabolic / virulence traits in Bcc and will
help in directing future studies on the molecular mechanisms of morphotype variation in these
bacteria.
KEYWORDS
Burkholderia multivorans; LysR-type transcriptional regulator; Glucose metabolism; Mucoid
morphotype variation; Two component regulatory systems

iv
ABBREVIATIONS
2-KG – 2-keto Gluconic Acid
5-KG - 5-keto Gluconic Acid
AHL - N-Acyl Homoserine Lactones
Bcc – Burkholderia cepacia complex
BLAST – Basic Local Alignment Search Tool
bp – Base Pair
Cm – Chloramphenicol
CmR – Chloramphenicol Resistance
CF – Cystic Fibrosis
CFTR – Cystic Fibrosis Transmembrane Conductance Regulator
CFU – Colony Forming Unit
CGD - Chronic Granulomatous Disease
dNTPS – Deoxyribonucleoside Triphosphate
EPS – Exopolysaccharide
HK – Histidine Kinase
HPLC – High Performance Liquid Chromatography
HTH - Helix-Turn-Helix Domain
IPTG – Isopropyl β-D-1-thiogalactopyranoside
Km – Kanamycin
KmR – Kanamycin Resistance
LB – Lennox Broth
LDH – Lactate Dehydrogenase
LPS – Lipopolysaccharide
LTTR – LysR-Type Transcriptional Regulator

v
NCBI – National Center for Biotechnology Information
OD – Optical Density
PBP2 – Periplasmic Binding Protein Type2 Superfamily
PCR – Polymerase Chain Reaction
QS – Quorum sensing
rpm – Rotations per minute
rRNA – Ribosomal RNA
RI - Refractive Index Detector
RR – Response Regulator
SM – S Medium with Mannitol
SGal – S Medium with Galactose
SMan – S Medium with Mannose
SSuc – S medium with Sucrose
TCS – Two Component Regulatory System
TpR – Trimethoprim Resistance
UV – Vis – Ultra Violet Visible Spectroscopy
X-gal – 5-bromo-4-chloro-3-indolyl-β-D-galactopyranoside
YEM – Yeast Extract Mannitol Medium

vi
TABLE OF CONTENTS
ACKNOWLEDGMENTS .......................................................................................................................... i
RESUMO ................................................................................................................................................. ii
PALAVRAS-CHAVE ............................................................................................................................... ii
ABSTRACT ............................................................................................................................................ iii
KEYWORDS .......................................................................................................................................... iii
ABBREVIATIONS .................................................................................................................................. iv
TABLE OF CONTENTS ......................................................................................................................... vi
LIST OF FIGURES ............................................................................................................................... viii
LIST OF TABLES ................................................................................................................................... x
1. Introduction .................................................................................................................................... 1
1.1 Overview of the genus Burkholderia.................................................................................... 1
1.1.1 Diversity ........................................................................................................................... 1
1.1.2 Burkholderia cepacia complex and Cystic Fibrosis ......................................................... 2
1.1.3 Burkholderia Virulence Factors ....................................................................................... 4
1.1.4 Mucoid-to-non mucoid phenotypic variation .................................................................... 6
1.2 LysR-type Transcriptional Regulators ................................................................................. 8
1.2.1 General Structure and Functions..................................................................................... 8
1.2.2 Role of LysR-type transcriptional regulators in morphotype variation ............................. 9
1.3 Two Component Regulatory Systems ............................................................................... 10
1.3.1 General Structure and Functions................................................................................... 10
1.3.2 Role of a EnvZ/OmpR-like TCS in mucoid morphotype variation in B. multivorans ..... 13
1.4 Objectives .......................................................................................................................... 14
2. Materials and Methods ................................................................................................................ 15
2.1 Bacterial strains, plasmids and culture conditions ............................................................. 15
2.2 DNA manipulation .............................................................................................................. 16
2.3 Phenotypic Characterization of Burkholderia strains......................................................... 16
2.4 Exopolysaccharide quantification ...................................................................................... 17
2.5 HPLC analysis ................................................................................................................... 17
2.6 Triparental conjugation ...................................................................................................... 18
2.7 Construction of the ompR gene replacement vector with Gateway-compatible allelic
exchange system ............................................................................................................... 18
2.8 In silico analysis of nucleotide and amino acid sequences ............................................... 18
2.9 Genome sequence determination and detection of SNPs and indel mutations ................ 19
3. Results .......................................................................................................................................... 20
3.1 Role of Bmul_2557 LysR transcriptional regulator ............................................................ 20

vii
3.1.1 Characterization of the Bmul_2557 gene and homologs through computation tools .... 20
3.1.2 Complementation of B. multivorans ATCC 17616 Bmul_2557 gene deletion mutant .. 24
3.1.3 Role of Bmul_2557 LysR regulator in cepacian biosynthesis ....................................... 25
3.1.4 Growth of the LysR mutant in the presence of glucose or mannitol .............................. 25
3.1.5 Analysis of extracellular metabolites by HPLC .............................................................. 28
3.1.6 Role of Bmul_2557 transcriptional regulator in the metabolism of other sugars ........... 29
3.1.7 Growth behavior of different Bcc isolates using glucose as carbon
source……………………………………………………………………………………….….34
3.2 Role of a OmpR-like response regulator ........................................................................... 36
3.2.1 Complementation of nonmucoid B. multivorans variants with an ompR-containing
plasmid …………….. ..................................................................................................... 36
3.2.2 Strategy to obtain the B. multivorans D2095 deletion mutant in ompR gene ………….38
4. Discussion.................................................................................................................................... 41
5. References ................................................................................................................................... 45
6. Appendix ...................................................................................................................................... 52

viii
LIST OF FIGURES
Figure 1.1 – Genetic organization of bce-I and bce-II clusters of genes encoding proteins involved in
cepacian biosynthesis………………………………………………………………………………………….. 6
Figure 1.2 – Biosynthesis of the exopolysaccharide cepacian by Burkholderia…………….………….. 7
Figure 1.3 – General mode of action for two-component signal transduction…………….……..…….. 10
Figure 2.1 – Glucose, galactose, 2-KG acid and D-lactate standard curves determined by HPLC…. 17
Figure 3.1 – Putative conserved domains for Bmul_2557 encoded LTTR of B. multivorans ATCC
17616 as obtained at NCBI conserved domains database…………………………….…….……...….... 20
Figure 3.2 – Phylogenetic three comprising 29 Burkholderia homologues of Bmul_2557 aligned by
clustal X and sorted by tree view………………………………………………….…………….…….…….. 21
Figure 3.3 – Genomic organization of B. multivorans ATCC 17616, B. multivorans D2095, B. cepacia
GG4, B. xenovorans LB400 and B. phytofirmans PsJN……………………………..……….…….…….. 22
Figure 3.4 – Multiple Sequence Alignment of Bmul_2557 homologs and 3D-structure as predicted, by
I-TASSER………………………………………………………………………………………….…….…….. 23
Figure 3.5 – Cloning strategy to obtain pARG015-1 plasmid……………………………………………. 24
Figure 3.6 – Exolysaccharide production by B. multivorans ATCC 17616, ΔBmul_2557::dhfR mutant
and ΔBmul_2557::dhfR + pARG015-1 in the presence of mannitol (SM medium) and in glucose (S
medium) at 37ºC………………………………………………………….……………………….……..….... 25
Figure 3.7 – Growth curves of B. multivorans ATCC 17616, ΔBmul_2557::dhfR mutant and
ΔBmul_2557::dhfR + pARG015-1 in SM and S medium at 37ºC……………...………………….....….. 26
Figure 3.8 – Number of viable cells of B. multivorans ATCC 17616, ΔBmul_2557::dhfR mutant and
ΔBmul_2557::dhfR + pARG015-1 in SM and S medium at 37ºC …………..……………….……....….. 26
Figure 3.9 – Culture supernatant pH values of B. multivorans ATCC 17616, ΔBmul_2557::dhfR
mutant and ΔBmul_2557::dhfR + pARG015-1 in SM and S medium at 37ºC…………...……………... 26
Figure 3.10 – Alternative pathways of 6-phosphogluconate formation……………………………….... 27
Figure 3.11 – 2-keto gluconic acid and D-lactate production by B. multivorans ATCC 17616,
∆Bmul_2557::dhfR mutant and ΔBmul_2557::dhfR + pARG015-1 in S medium, at 37ºC…..….…….. 28
Figure 3.12 – Glucose consumption by B. multivorans ATCC 17616, ΔBmul_2557::dhfR mutant and
ΔBmul_2557::dhfR + pARG015-1 in the presence of glucose (S medium) and in the presence of
mannitol (SM medium) at 37ºC………………………………………………………………….…….…….. 29

ix
Figure 3.13 – Growth curves, number of viable cells and culture supernatant pH values of B.
multivorans ATCC 17616 and ΔBmul_2557::dhfR mutant in medium supplemented with D-galactose
at 37ºC...............................................................................................................………….…….... 30
Figure 3.14 – Leloir pathway…………………………………………………………………….……....….. 30
Figure 3.15 – 2-keto gluconic acid production and galactose consumption of B. multivorans ATCC
17616 and ΔBmul_2557::dhfR mutant in SGal medium, at 37ºC………………………………………... 31
Figure 3.16 – Growth curves, number of viable cells and culture supernatant pH values of B.
multivorans ATCC 17616 and ΔBmul_2557::dhfR mutant in medium supplemented with D-mannose at
37ºC…………………………………………………………………………………………………….......….. 32
Figure 3.17 – D-mannose metabolism......................................................................................... 32
Figure 3.18 – Growth curves, number of viable cells and culture supernatant pH values of B.
multivorans ATCC 17616 and ΔBmul_2557::dhfR mutant in SSuc medium at 37ºC ……………..….. 33
Figure 3.19 – Sucrose hydrolysis into glucose and fructose…………………………………………..... 34
Figure 3.20 – Growth curves, viable cells and culture supernatant pH values for B. multivorans
D2095, B. multivorans HI229 and B. contaminans IST408, in S medium at
37ºC…………………………………………………………………………………………….….……...….... 34
Figure 3.21 – 2-keto gluconic acid production, D-lactate production and glucose consumption for B.
multivorans D2095 and B. multivorans HI229, in S medium at 37ºC………………….….………....….. 35
Figure 3.22 – Colony morphologies of B. multivorans NMV121 and NMV124 variants, in selective
YEM medium after 72 hours at 30ºC, complemented with pBBR1 vector or pLM014-5………......….. 37
Figure 3.23 – Construction of the gene replacement vector pENTRPEX18Tp-SceI pheS-ompR, BP
clonase reaction and transformation of B. multivorans D2095........................................................ 38
Figure 3.24 – Agarose gel electrophoresis of the PCR amplified DNA fragments located upstream and
downstream of the ompR-like encoding gene, from B. multivorans D2095…………………….......….. 39
Figure 3.25 – Transformation of B. multivorans D2095 with the expression vector pDAI-SceI-pheS
and second homologous recombination event…………………………………………………………….. 40

x
LIST OF TABLES
Table 1.1 – Burkholderia cepacia complex species and sources………………………...….….……….. 3
Table 1.2 – Burkholderia virulence factors………………………….………………………………..……… 4
Table 2.1 – Bacterial strains used in this study.............................................................................…... 15
Table 2.2 – Plasmids used in this study........................................................................................…... 16
Table 3.1 – Bacterial strains used in this study…………………………………………………………..... 36
Table 3.2 – Sequence results of some nonmucoid variants that restored EPS production upon
complementation with the ompR-containing plasmid……………………………………….…………...... 37
Supplementary table 1 – Oligonucleotide primers used in this study................................................. 52

Introduction _____________________________
1
1. Introduction
1.1 Overview of the genus Burkholderia
1.1.1 Diversity
Burkholderia genus is a group of bacteria belonging to the beta subdivision of Proteobacteria,
which was created in 1992 to accommodate the former rRNA group II pseudomonads (Yabuuchi et al.,
1992). Members of the Burkholderia genus are Gram-negative bacteria which occupy a wide range of
ecological niches such as soil, water, rhizosphere, and can also be found in close association with
plants and animals, due to their ability to interact as symbiotic or pathogenic organisms. Some of
those species are also known as opportunistic pathogens in humans (Coenye and Vandamme 2003).
The first isolate of the genus was Burkholderia cepacia (at the time classified as Pseudomonas
cepacia) and was described by William Burkholder, in 1950, as the phytopathogen responsible for
causing sour skin rot, a bacterial soft-rotting disease of onions (Burkholder, 1950). Over the years,
many other bacterial species have been described as belonging to this genus, or due to molecular
taxonomic studies, have been transferred to or from other genera (Vandamme and Dawyndt 2011).
Given this fact, the genus Burkholderia currently comprises around 100 species
(http://www.bacterio.net/burkholderia.html).
Burkholderia spp. have been the subject of intensive studies in the scientific community
because of the versatility of their constituents (O’Sullivan and Mahenthiralingam 2005). Traditionally,
members of Burkholderia genus were linked to pathogenicity in plants, triggering processes like rot of
onions, rice grains and seedlings. Some specimens belonging to this category are Burkholderia
caryophylli, Burkholderia glumae and Burkholderia andropogonis. However, Burkholderia species are
extremely versatile and are able to colonize very different ecological niches, being primary pathogens
in humans and horses, as is the case of Burkholderia mallei and Burkholderia pseudomallei. The first
one causes glanders in horses and several other animal species. Human infections caused by this
organism are rare although exceptionally few organisms are needed for human infection. B.
pseudomallei causes melioidosis, a disease similar to glanders. However, in contrast with B. mallei,
melioidosis is more common than glanders, affecting animals and humans with more frequency
(Compant et al. 2008).
In recent years, a growing number of Burkholderia strains and species have already been
described with the ability to establish non-pathogenic interactions with plants. While most of them do
not have an assigned biological role (Burkholderia glathei, Burkholderia graminis and Burkholderia
phenazinium), others, like Burkholderia vietnamiensis, Burkholderia xenovorans, and Burkholderia
phymatum, play important symbiotic relationships with plants, contributing to their growth promotion,
through better water management and improving atmospheric nitrogen fixation, by colonizing the
roots, leaves and stems (Coenye and Vandamme 2003; Compant et al. 2008). In certain Burkholderia
strains, genes encoding enzymes involved in metabolism of diverse carbon sources have been found,
which has prompted a growing interest in using them in fields like agriculture and also in biocontrol

Introduction _____________________________
2
and bioremediation processes (reviewed in O’Sullivan and Mahenthiralingam 2005). However, the
application of Burkholderia spp. in these fields, have to be accompanied with a stringent assessment
of the potential risks, due to the pathogenic role of some species/isolates, as mentioned above, and
also because there is no clear distinction between environmental and clinical strains
(Mahenthiralingam et al. 2008) .
Taking into account the heterogeneity of the Burkholderia genus, several efforts have been
made in order to group these species into smaller groups, with higher specificity. In 1997, Vandamme
and collaborators, using a taxonomic approach based on multiple tests, known as polyphasic
taxonomy, divided Burkholderia strains commonly isolated from cystic fibrosis (CF) patients infections
into five genomovars. In the subsequent years, more genomovars were created, resulting in a total of
ten. Genomovars were associated with different levels of virulence and patient-to-patient
transmissibility. After some time, genomovars began to be called species, since they were composed
by phenotypically similar isolates that were genetically distinct. The collective of these species was
named Burkholderia cepacia complex (Bcc), and comprises a group of species that are opportunistic
pathogens, causing lung infections in cystic fibrosis and immune compromised patients
(Mahenthiralingam et al. 2005). The Burkholderia cepacia complex currently comprises 18 closely
related species presenting moderate levels of DNA-DNA hybridization and high similarities between
16S ribosomal DNA sequences (Coenye et al. 2001; Peeters et al. 2013).
1.1.2 Burkholderia cepacia complex and Cystic Fibrosis
Bacteria belonging to Bcc are important pathogens that can chronically colonize the airways
of cystic fibrosis patients. CF is an autosomal recessive genetic disorder that renders affected
individuals susceptible to chronic and ultimately fatal lung infections (Gibson et al. 2003). This is the
most common lethal inherited genetic disease among Caucasians, affecting about 70,000 people
worldwide (http://www.cftrscience.com/epidemiology). The genetic defect underlying this disease
results from mutations in the Cystic Fibrosis Transmembrane Conductance Regulator (CFTR) gene,
which leads to a disrupted chloride channel. Individuals with this irregularity present a defective
regulation of chloride and sodium ions in epithelial cells. Thus, several organs are affected, especially
the lungs, which result in defective mucociliar clearance of bacterial pathogens, predisposing the
individuals to respiratory infections (Govan and Deretic 1996). The airways of CF patients may be a
reservoir to several pathogenic organisms, including Pseudomonas aeruginosa, Staphylococcus
aureus, Achromobacter xylosoxidans and Bcc bacteria. CF patients are also susceptible to
colonization by other pathogens, as Pandorae genus, Stenotrophomonas maltophilia and non-
tuberculous Mycobacteria (Lipuma 2010).
Bcc causes infections in only about 3.5% of CF patients worldwide, however these infections
are particularly problematic. One of the reasons results from the fact that Bcc infections outcome
ranges from asymptomatic infections, with little or no impact on lung function, to rapid deterioration,
that in the worst cases can lead to septicemia and death, being unpredictable (reviewed in Leitão et al.
2010). The most serious diagnosis, called “cepacia syndrome” is associated with chronic infection and
exacerbation episodes, resulting in necrotizing pneumonia and bacteremia (Isles et al. 1984). Besides

Introduction _____________________________
3
this, Bcc bacteria not only have the capability to be transmitted among people by social contact, but
they also have intrinsic resistance to currently available antimicrobial therapies. Taking this into
account, many efforts have been done to follow strict guidelines to avoid any contact between Bcc-
infected and noninfected patients (Lipuma et al. 2001). Also, the prevalence of Bcc species varies
geographically and regionally, which makes the diagnosis and treatment extremely difficult. B.
cenocepacia and B. multivorans have been the most predominant species among CF patients,
representing around 90% of all Bcc infections in the world. This makes B. cenocepacia and B.
multivorans the most well studied species among all Burkholderia species (reviewed in Drevinek and
Mahenthiralingam, 2010). Typically, the development of a chronic pulmonary infection involves a
single Bcc strain. However, it has been described some cases where prolonged co-infection with two
distinct strains occur, and also the replacement of an initial infecting strain with another one, during the
course of chronic lung infection (Lipuma 2010). Strains from Bcc present large genomes, usually with
three chromosomes, and many of them contain also plasmids. Their gene content ranges from 5,500
to 7,900 genes (Holden et al. 2009), explaining their extraordinary metabolic versatility and allowing
their adaptation to a wide range of environments (reviewed in Sousa et al. 2011).
Bacteria belonging to Bcc have been isolated from both environmental reservoirs and clinical
sources, and these have proven to be important opportunistic pathogens in cystic fibrosis, chronic
granulomatous disease and immunocompromised patients (Baldwin et al. 2007; Leitão et al. 2010)
(Table 1.1).
Table 1.1 - Burkholderia cepacia complex species and sources (adapted from Silva, 2012, phD).
Bcc species Sources
References Natural Environment Clinical Environment
B. cepacia Rhizosphere, soil, plant, river
water
Human (CF and non-CF)
medical soultion contaminant
(Yabuuchi et al. 1992)
(Vandamme et al. 1997)
B. multivorans Rhizosphere, soil, plant, river
water Human (CF and non-CF) (Vandamme et al. 1997)
B. cenocepacia
Rhizosphere, soil, plant, river
water, industrial contaminant
Human (CF and non-CF) (Vandamme et al. 1997)
(Vandamme et al. 2003)
B. stabilis Rhizosphere Human (CF and non-CF)
medical devices contaminant
Vandamme et al. 1997)
(Vandamme et al. 2000)
B.vietnamiensis Rhizosphere, soil, plant, river
water, industrial contaminant Human (CF and non-CF)
(Gillis et al. 1995) (Vandamme et
al. 1997)
B. dolosa Rhizosphere Human (CF) (Coenye et al. 2001)
(Vermis et al. 2004)
B. ambifaria Rhizosphere, soil Human (CF) (Coenye et al. 2001)
B. anthina Rhizosphere, soil, plant, river
water
Human (CF) medical devices
contaminant (Vandamme et al. 2002)
B. pyrrocinia Rhizosphere, soil, plant, river
water Human (CF and non-CF)
(Viallard et al. 1992)
(Vandamme et al. 1997)
(Vandamme et al. 2002)
B. ubonensis Soil Human (non-CF) nosocomial
infection
(Yabuuchi et al. 2000)
(Vanlaere et al. 2008)

Introduction _____________________________
4
Table 1.1 - Burkholderia cepacia complex species and sources (adapted from Silva, 2012, phD) (cont)
Intensive research has been made not only in the epidemiology field, but also in the discovery
of novel therapeutic targets and in the elucidation of the mechanisms underlying infections, host-
pathogen interactions and virulence factors involved in Bcc colonization.
1.1.3 Burkholderia Virulence Factors
During the course of an infection, Bcc bacteria produce a wide range of virulence factors that
play determinant roles. Nevertheless, the progress on the development of new therapeutic agents is
still limited. However, many efforts are being made to discover molecular mechanisms underlying the
virulence factors and determinants of Bcc, which can be the key to develop strategies to combat
infections caused by these bacteria (reviewed in Leitão et al. 2010 and Mahenthiralingam et al. 2005).
Different strategies have been designed to identify pathogenicity-related genes from Bcc bacteria,
including systematic gene-by-gene inactivation and high-throughput sequencing (reviewed in Sousa et
al. 2011). Some of the virulence factors employed by Bcc bacteria during the course of the infection
are listed in table 1.2.
Table 1.2 - Burkholderia virulence factors
Bcc species Sources
References Natural Environment Clinical Environment
B. latens _ Human (CF) (Vanlaere et al. 2008)
B. diffusa Soil, water Human (CF and non-CF) (Vanlaere et al. 2008)
B. arboris Rhizosphere, soil, river water,
industrial contaminant Human (CF and non-CF) (Vanlaere et al. 2008)
B. seminalis Rhizosphere, soil, plant Human (CF and non-CF) (Vanlaere et al. 2008)
B. metallica _ Human (CF) (Vanlaere et al. 2008)
B. contaminans Soil, sea water, animal Human (CF and non-CF)
medical devices (Vanlaere et al. 2009)
B. lata Soil, river water, plant Human (CF and non-CF) (Vanlaere et al. 2009)
B. pseudomultivorans Rhizosphere Human (CF and non-CF) (Peeters et al. 2013)
Virulence Factor Description References
Flagella
- Composed by flagelin
- Facilitates penetration through the host epithelial cell barriers and to the onset of
systemic spread of the organism
- Involved in host inflammatory responses
- Required for signaling through Tol-like receptor and virulence
(Tomich et al. 2002)
(Chua et al. 2003)
(Urban et al. 2004)
Cable Pilli and
22 kDa Adhesin
- Large peritrichous organelles expressed on the surface of bacterial pathogens
- Required for full adherence to epithelial cells and to mucins
- 22 kDa Adhesin mediates binding to human respiratory epithelium
- Involved in transmigration of bacteria across epithelia cell
(Sajjan et al. 1995)
(Sajjan et al. 2002)
(Tomich and Mohr 2003)

Introduction _____________________________
5
Table 1.2 - Burkholderia virulence factors (cont.).
Virulence Factor Description References
Lipopolysaccharide
(LPS)
- Large surface components, composed of a lipid A, a core polysaccharide and O-
antigen
-Important in resistance to antimicrobial peptides against cationic antibiotics, like
polymyxin B and melittin
- A thrisaccharide repeating unit of the O-antigen tested as a vaccine component
- A considerable variety of O-antigen, making difficult to design the corresponding
vaccines
(Dubiel and Goldberg 2003)
(Loutet et al. 2006)
Exopolysaccharide
(EPS)
- Extracellular high-molecular weight sugar-based polymers
- Role in bacterial adaptation to stress conditions
- Involved in protection from the host immune system and harmful compounds
- Ability to inhibit neutrophil chemotaxis
- Scavenge reactive oxygen species
- Interfere with phagocytosis
- Reduce the antimicrobial activity of antimicrobial peptides (AMPs)
- Contributes to biofilm formation
- Most common EPS produced is cepacian
(Chung et al. 2003)
(Ferreira et al. 2011)
(Ferreira et al. 2010)
(Bylund et al. 2006)
(Herasimenka et al. 2005)
(Cescutti et al. 2000)
Extracellular
Enzymes
- Lipases, Metalloproteases and Serine proteases
- ZmpA and ZmpB metalloproteases are active against members of the host immune
system (neutrophil α-1 proteinase inhibitor and γ-interferon)
- HtrA serine protease present ability to use ferritin as an iron source
- A hemolysin induce hemolysis of erythrocytes
(Corbett 2003)
(McClean and Callaghan
2009)
(Hutchison, Poxton, and
Govan 1998)
Secretion
Systems
- Secretion of effector molecules that disrupt host cellular processes
- Type I, type II, type III, type IV, type V and type VI secretion systems
- Involved in close host-pathogen interaction
- Allow translocation of trimeric autotransporter adhesins
- Disruption of host cells actin cytoskeleton
(Loutet and Valvano 2010)
(Mil-Homens and Fialho 2011)
(Rosales-Reyes et al. 2012)
Siderophores
- Low-molecular-weight iron-chelating compounds
- Pyochelin, salicylic acid, cepabactin, cepaciacheline and ornibactin
- Involved in free iron uptake from surrounding environment
(Bevivino et al. 1994)
(Meyer et al. 1995)
(Visser et al. 2004)
Biofilms
- Complex, multicellular bacterial communities composed of extracellular DNA,
proteins and exopolysaccharides wherein bacteria live in a sessile lifestyle
- Protection against environmental insults and host immune system defenses
- Provide increased resistance to antibiotics
- Biofilm formation is affected by EPSs production, motility, and iron availability
(Conway et al. 2002)
(Huber et al. 2002)
(Van Acker et al. 2013)
(Ferreira et al. 2011)
Resistance to
Antibiotics and
Oxidative Stress
- Intrinsically resistance to most of the clinically available antimicrobials
- Efflux pumps that remove antibiotics from the cell
- Changes in the cell envelope that reduce the membrane permeability - Production
of enzymes able to degrade antibiotics, as β-lactamases
- Resistance against reactive oxygen species (ROS)
- Catalase, catalase-peroxidase, and superoxide dismutase (SOD) activities
(Nzula 2002)
(Gibson et al. 2003)
(Leitão et al. 2008)
Quorum Sensing
- Cell density dependent communication system utilized by Gram-negative bacteria
for the coordination of gene expression
- Mediated by N-Acyl Homoserine Lactones (AHL)
- Allows regulation of wide range of different processes related with virulence
- Can occur within a single bacterial species as well as between diverse species
- Several QS systems: CepIR, BvIR and CcIR and different types of signals
(Fuqua et al. 1994)
(Lewenza et al.1999)
(Malott and Sokol 2007)

Introduction _____________________________
6
1.1.4 Mucoid-to-non mucoid phenotypic variation
Within the CF lung, colonizing bacteria is faced with adverse conditions, like high osmolarity,
heterogeneous distribution of oxygen and nutrients, high concentration of antimicrobials, and constant
challenge by the host immune defenses. These factors exert a selective pressure in colonizing
bacteria and are thought to be the driving force of microevolution during their persistence in the CF
lung (Lyczak et al. 2002; Döring et al. 2011). Genotypic and phenotypic variation are common
phenomena during the course of an infection, and have been described in many pathogens including
P. aeruginosa, the major pathogen colonizing the CF lung, where most phenotypic conversions occur
from the nonmucoid-to-mucoid morphotype, characterized by the overproduction of the
exopolysaccharide (EPS) alginate (Govan et al.1996). In Bcc bacteria, phenotype transitions in
isolates recovered from CF patients have also been reported. However, most of them were from
mucoid-to-nonmucoid morphotype (Zlosnik et al. 2008) and nonmucoid isolates have been associated
with a poorer clinical outcome (Zlosnik et al. 2011). A hypothesis derived from these observations is
that while the mucoid phenotype may be associated to persistence in CF lung, nonmucoid isolates
may be associated to increased disease severity. EPS may be required for colonization in the early
stages of infection and persistence in the CF lung, but once bacteria have colonized the lung, the EPS
production may no longer be necessary and can be shutdown, since it represents a great spending of
energy to the cell (Silva et al. 2011).
The most abundant EPS produced by Bcc bacteria, is cepacian. This exopolysaccharide has
been identified in both clinical and environmental isolates of Bcc bacteria (Ferreira et al., 2010) and
the characterization of its chemical structure and composition showed that it is composed of a
branched acetylated heptasaccharide repeat unit with D-glucose, D-rhamnose, D-mannose, D-
galactose and D-glucuronic acid, in the ratio 1:1:1:3:1, respectively and linked by (1→3) glyosidic
bonds. Genes involved in cepacian biosynthesis are located within bce-I and bce-II clusters (Figure
1.1). The biosynthesis process starts with the synthesis of the nucleotide sugar precursors, necessary
for the repeat unit formation, and is catalyzed by isomerases, mutases, epimerases, among other
enzymes (Figure 1.2) (Ferreira et al., 2010). This step is followed by the assembly of the repeat-units
by the sequential addition of sugars to an isoprenoid lipid by dedicated glycosyltransferases. In the
last step the polymerization and export of the polysaccharide to the extracellular environment is
carried out by a multienzyme complex that includes a repeat-unit translocase, a polysaccharide
polymerase, and an outer membrane protein, among others (Moreira et al. 2003; Ferreira et al.,
2010).
Figure 1.1 - Genetic organization of bce-I and bce-II clusters of genes encoding proteins involved in cepacian biosynthesis (Adapted from Ferreira et al., 2011).■ Nucleotide sugar biosynthesis; ■ Glycosyltransferase; ■ Polymerization/Export; ■Acyltransferase; ■ Unknown function. The two clusters are separated approximately by 155-314 kb in the genome of most Burkholderia species.

Introduction _____________________________
7
Figure 1.2 - Biosynthesis of the exopolysaccharide cepacian by Burkholderia (Adapted from Ferreira et al., 2011). Repeat-units
of the polymer are assembled on a lipid carrier, in the cytoplasmic side of inner membrane in a reaction initiated by the BceB enzyme and continued by the other glycosyltransferases BceG, BceH, BceJ, BceK, and BceR and putative acyltransferases BceO, BceS and BceU. The lipid-linked repeat-units are translocated across the inner membrane by the putative BceQ membrane protein. Polymerization occurs at the periplasmic face of the inner membrane and is dependent on BceI. Wzy-dependent polymerization/export requires the activity of the BceF tyrosine kinase. BceD is a protein tyrosine phosphatase enzyme responsible for dephosphorylating BceF. BceE forms a channel structure for export of EPS chains to the outside. BceP, putatively involved in polysaccharide degradation, is depicted as associated to the outer membrane, as one of its possible locations. Glc - glucose; GlcA - glucuronic acid; Gal - galactose; Rha - rhamnose; Man - mannose; Fru - fructose; GDP - guanosine-5’-diphosphate; UDP - uridine-5’-diphosphate; PGM - phosphoglucomutase; UGP - UDP-glucose pyrophosphorylase; UGD - UDP-glucose dehydrogenase; UGE - UDP-glucose epimerase; PGI - phosphoglucose isomerase; PMI - phosphomannose isomerase; PMM - phosphomannomutase; GMP - GDP-D-mannose pyrophosphorylase; GRS - GDP-rhamnose synthase; ATP - adenosine-5’- triphosphate; ADP - adenosine-5’- diphosphate; Y
P - phosphorylated tyrosine residue;
Pi - inorganic phosphate; IM - inner membrane; OM - outer membrane; PL - peptidoglycan layer.
Although genes involved in cepacian biosynthesis are already known, the genes responsible
for the regulation of EPS production have not been identified, and more studies are needed to better
understand this virulence factor. Conway et collaborators (2004), have shown that besides the
possible shutting down of EPS production, during the infection, other phenotypic alterations occur,
such as resistance to antibiotics, biofilm formation, motility and lower toxin production. Mucoid-to-
nonmucoid morphotype variation was also seen to occur in vitro and to be triggered by stress
conditions with relevance in the CF airways such as, the presence of antibiotics, osmotic and oxidative
stresses, nutrient starvation and microaerophilia (Silva et al. 2013).
Regarding the discovery of the regulatory mechanisms by which Bcc bacteria undergo
mucoid-to-nonmucoid variation, global transcriptomic profiling studies have been carried out in our
laboratory. In a study performed by Silva et al (2011), the expression profiles of two sequential B.
multivorans isolates, from a chronically infected CF patient, displaying different morphotypes (mucoid
D2095 and nonmucoid D2214) were analyzed. The results showed decrease expression of genes
related to virulence in the nonmucoid isolate, and also lower motility and lower virulence in the acute
model of infection Galleria mellonella, but increased biofilm formation. Nonmucoid D2214 also
presents decreased expression of genes involved in carbon and nitrogen metabolism, which may
reflect lower metabolic needs of this isolate, possibly caused by lack of EPS production, but also by
the attenuation of pathways not required for survival. This was consistent with the fact that isolate
D2214 presented higher survival rate in minimal medium, compared with D2095. In a further study,

Introduction _____________________________
8
transcriptomic profiles of mucoid B. multivorans D2095 and B. multivorans ATCC 17616 were
compared with the ones from nonmucoid variants obtained in vitro for each strain (Tavares, 2012,
MSc Thesis). The analysis of the three transcriptomic data sets showed consistent decreased
expression in the nonmucoid strains of gene Bmul_2557, from B. multivorans ATCC 17616, encoding
a transcriptional regulator. This transcriptional regulator is from the LysR family of transcriptional
regulators (LTTR) which includes members involved in virulence and metabolic processes.
In a different attempt to identify genes involved in the regulation of mucoid-to-nonmucoid
phenotype variation, whole-genome sequencing of nonmucoid variants of B. multivorans D2095
obtained in vitro (Silva et al, 2013) revealed that about 90% have mutations in an open reading frame
encoding a response regulator (RR) of a two-component system (TCS), being annotated as OmpR-
like (unpublished results). Taking this into account, the study of this TCS and its involvement in
regulation of phase variation is of major importance to understand morphotype variation in Bcc.
Despite our knowledge on the relevance of mucoid-to-nonmucoid variation in Bcc, the molecular
mechanisms triggering mucoid morphotype variation still need to be investigated to discover new
therapeutic targets. Therefore, taking into account what has been mentioned above, the study of this
LysR-type regulator and TCS will enlighten regulation of metabolic and/or virulence traits in Bcc and
may help to understand the molecular mechanisms of morphotype variation in these bacteria.
1.2 LysR-type Transcriptional Regulators
1.2.1 General Structure and Functions
The LysR-type transcriptional regulator (LTTR) family is a group of transcriptional regulators
that are highly conserved and abundant amongst bacteria, presenting functional orthologues in
Archaea and eukaryotic organisms. LTTRs orthologues retain their structure and function, which
suggests their role in regulation of similar genes with functions involved in metabolism, cell division,
quorum sensing, virulence, motility, nitrogen fixation, oxidative stress responses, toxin production,
attachment and secretion (reviewed in Maddocks and Oyston 2008; Schell, 1993). LTTRs present a
structure with an N-terminal DNA-binding helix-turn-helix motif and a C-terminal coinducer-binding
domain. Residues 20–80 are the most highly conserved, and are directly involved with DNA
interaction. On the other hand, there is little conservation at the amino acid level for the C-terminus of
LTTRs (Maddocks and Oyston 2008). Originally, LTTRs were described as transcriptional activators of
a single transcribed gene. However, nowadays it is known that these proteins can act as global
transcriptional regulators, acting not only as activators, but also as repressors of single and operonic
genes (O’Grady et al. 2011).
The best characterized protein of LTTR family is LysR and it has been the subject of many
studies. However, LysR family has continued to increase in size, with other regulators being described
(Maddocks and Oyston 2008). One of them was described in Neisseria meningitidis, a human
pathogen that causes septicemia and meningitis. N. meningitidis adheres to the host in two steps: first,
bacteria attach to the target cells surface, in a process mediated by a pilus. After that, bacteria come
into close contact with cells (intimate adhesion). Gene crgA encodes a transcriptional regulator of the

Introduction _____________________________
9
LysR family, CrgA, and the insertional inactivation of this gene decreases adhesion of bacteria to
epithelial cells, especially during intimate adhesion. This suggests that crgA gene has a role in
biosynthesis of the capsule and pili, and also in the regulation of genes involved in intimate adhesion
to host cells (Deghmane et al. 2002).
Another member of LTTR family that has been described is AphB. This protein, together with
AphA, a member of a new regulators family, is involved in the expression of the Vibrio cholerae
virulence cascade. AphA together with AphB, activates transcription by binding to tcpPH promoter,
which activates the expression of the virulence cascade. Mutations in AphA cause reduction in the
expression of tcpPH promoter and prevent binding to DNA (Kovacikova et al. 2004). When this
mutation is induced, it is also observed that AphB rescue some mutants, suggesting its capacity to
binding directly to DNA, and stabilize defective proteins. On the other hand, mutations in AphB, which
lead to disruption of AphB-DNA interaction, result in prevention of the virulence cascade. In this case
AphA does not have the ability to rescue the mutants. This fact, along with the observation that
increased amounts of AphB can compensate for the loss of AphA, whereas increased amounts of
AphA cannot compensate for the loss of AphB, suggest that although both AphA and AphB are
required for tcpPH expression, AphB is probably the primary activator and AphA plays an accessory
role (Kovacikova et al. 2004).
1.2.2 Role of LysR-type transcriptional regulators in morphotype
variation
Colony morphotype variation has been reported in many bacteria species, such as Vibrio
parahaemolyticus, Pseudomonas aeruginosa and Actinobacillus actinomycetemcomitans, being
associated with some behavioral changes, including motility, biofilm formation, resistance to
antibiotics, chlorine, and osmotic or oxidative stresses. However, these altered phenotypes are not
only related with colony morphology changes, but also occur as a result from a mutation in some
transcriptional regulators (O’Grady et al. 2011).
Previous studies showed that B.cenocepacia C1394 switches its colony morphotype changing
from matte to shiny mucoid colony that exhibits hyperpiliation and a higher EPS production (Bernier et
al. 2008).To better understand this phenomenon, Bernier and collaborators (2008) conducted a study
in other Bcc strain, B. cenocepacia K56-2, which is unable to produce cepacian. This strain showed a
variation from rough to shiny colony on agar medium after shaken or static incubation in liquid culture
medium (Subramoni et al. 2011). The shiny variants typically showed absence of extracellular matrix
and a reduced biofilm formation, higher persistence, and lower virulence in a chronic agar bead model
of respiratory infection, than the original isolate (O’Grady et al. 2011). Through transposon
mutagenesis, gene BCAS0225, which encodes ShvR, a LysR-type transcriptional regulator, was
identified as responsible for colony morphotype variation in this B.cenocepacia strain. Mutants in shiny
variant regulator (shvR) gene were shiny, defective in biofilm formation and show an absence of
extracellular matrix. Furthermore, wild-type matte phenotype was restored after complementation of
shiny variants with intact BCAS0225, highlighting the role of this gene in phenotype variation (Bernier
et al. 2008). TEM micrographs revealed the presence of a similar extracellular matrix, surrounding

Introduction _____________________________
10
bacteria in the rough colonies, that was absent in the shiny variants and a BCAS0225 mutant,
suggesting that ShvR regulator is involved in the biosynthesis or production of this extracellular matrix
(Bernier et al. 2008). However, the nature of this extracellular matrix is unknown and more studies are
needed to determine the function of ShvR and mechanisms underlying this colony morphotype
variation .
1.3 Two Component Regulatory Systems
1.3.1 General Structure and Functions
Two-component regulatory systems (TCSs) are sophisticated signal transduction devices,
serving as a stimulus-response coupling mechanism that allow organisms to sense and respond to
changes in many different environmental conditions (reviewed in Beier and Gross 2006). Over the
years, researchers have found such systems in all domains: Eubacteria, Archaea and also Eukarya.
However the abundance in each domain differs substantially, having been identified more TCSs in
prokaryotes than eukaryotes (Stock et al. 2000).
TCSs are typically composed by a membrane-bound histidine kinase (HK), which senses a
specific environmental stimulus, and a corresponding response regulator protein (RR), located in the
cytoplasm, that mediates the cellular response (Figure 1.3) (Beier and Gross 2006). The sensor HK,
typically a transmembrane protein, has an extracytoplasmic domain, which acts as a sensor, detecting
changes in the environment. When this happens, there is activation of intrinsic histidine kinase activity
that phosphorylates a histidine residue in the cytoplasmic domain (transmitter module). The sensor HK
then transfers the phosphate to a conserved aspartate residue in the N-terminal end (receiver domain)
of the RR protein. Response regulator receiver domain phosphorylation induces a conformational
change in the regulator (output domain). The output domain of most of the known response regulators
typically binds target DNA sequences; although some output domains are involved in protein–protein
interactions (Foster and Spector 2002).
Figure 1.3 - General mode of action for two-component signal transduction (Adapted from Foster and Spector 2002). Upon
detection of an appropriate signal, autophosphorylation occurs at a conserved His residue of the HK, followed by phosphoryl
group transfer to an Asp residue of the RR. A typical function for the RR is gene regulation.

Introduction _____________________________
11
It is also important refer that phosphorylation of the RR does not necessarily lead to a
downstream response, as might be assumed. HK may keep the response regulator in phosphorylated
state, to prevent it from triggering a response, being the dephosphorylated regulator the species
responsible for subsequent events (Foster and Spector 2002). Between the signals that are believed
to be detected by the TCSs are chemical and physical parameters, including ions, temperature, pH,
oxygen pressure, osmolarity, auto inducer compounds, the redox state of electron carriers, and the
contact with host cells (Beier and Gross 2006).
Until now, a high number of TCSs have been identified in bacterial genomes, highlighting the
impact of these systems on environmental adaptation of bacteria. The number of TCSs is related with
bacteria genome size and metabolic versatility, with larger genomes and organisms that colonize
different environments tending to encode more TCSs than bacteria with small genomes and living in a
uniform habitat (Beier and Gross 2006). In Escherichia coli, sense and response to changes in
environmental conditions occur through TCSs, including changes in medium osmolarity.
OmpF and OmpC are porin proteins located in the outer membrane that allow polar molecules to
cross this barrier, changing their amounts in response to medium osmolarity (Foster and Spector
2002). In low osmolarity medium, OmpF is present in higher amounts than OmpC in bacterial cell
membrane. Otherwise, in high osmolarity media, OmpC is more abundant than OmpF. Regulation of
genes codifying for these proteins is mediated by envZ and ompR genes, which codify EnvZ/OmpR
proteins, members of a TCS. EnvZ is a transmembrane protein that acts as a sensor, suffering
autophosphorylation in a histidine residue in response to increase of medium osmolarity.
Subsequently, the phosphoryl group is transferred to the response regulator (OmpR), which increases
the level of phosphorylated OmpR (OmpR-P) in the cell. At low osmolarity, OmpR-P binds to ompF
promoter sites (with high affinity), activating the transcription of ompF, and subsequently increasing
OmpF levels. When an increase in media osmolarity occurs, OmpR-P binds to ompC promoter sites
(with low affinity), which result in OmpC activation. Besides osmolarity, the EnvZ-OmpR system is
responsible for other functions in cellular physiology, including flagellar expression, cell division, fatty
acid transport, microcin synthesis, curly fibers and acid tolerance, showing the versatility of this TCS
system (Foster and Spector 2002).
The regulation systems for Staphylococcus aureus virulence properties have also been well
characterized, and involve an interaction of different TCSs and additional regulators to control
expression of virulence factors at different stages of infection (Beier and Gross 2006). The regulation
of S. aureus virulence, involving the AgrA-AgrC TCS, which responds to cell-density and controls the
transcription of the regulatory RNA III, as well as three additional TCSs (SaeR-SaeS, SsrA- SsrB and
ArlR-ArlS) has been described, showing different levels of complexity in two-component signaling
(Beier and Gross 2006).
In contrast to S.aureus, that shows integration of various systems into regulatory networks,
Bordetella pertussis presents a TCS (BvgA-BvgS) that appears to be the master regulator of their
virulence, controlling all known virulence traits. BvgA is a 23-kDa DNA-binding response regulator,
whereas BvgS is a 135-kDa transmembrane sensor kinase. Bordetellae bacteria can exist in three
distinctive phenotypes: Bvg+, Bvg
i and Bvg
-. Bacteria in the Bvg
+ phase, the virulent phase, express

Introduction _____________________________
12
several virulence factors, including toxins such as pertactin, and pertussis. During the Bvg- phase, the
avirulent phase, the majority of virulence factors is down regulated, whereas other genes, for example
those that are required for motility, are up regulated. Some virulence genes, including those encoding
fimbriae, are expressed also in a Bvg intermediate phase, known as Bvgi. BvgA-BvgS control at least
four different classes of genes: those that are expressed maximally only in the Bvg+ phase and are
known as late Bvg-activated genes; those that are expressed exclusively in the Bvgi phase; those that
are only expressed in the Bvg- phase; and those that are expressed maximally in both the Bvg
+ and
Bvgi phase, known as early Bvg-activated genes (Mattoo and Cherry 2005). Bordetellae can be forced
into a particular phenotypic phase by growing them under specific conditions. When grown at 37oC in
the relative absence of MgSO4 or nicotinic acid, bacteria grow in the Bvg+ phase. When the bacteria
are grown either at 25oC or at 37
oC in the presence of ≥ 40 mM MgSO4, Bordetellae display the Bvg
-
or Bvgi phenotype, respectively (Weiss and Melton 1993). The role of these distinct phenotypic phases
in the pathogenicity of the Bordetellae has been a subject of extensive research (Beier and Gross
2006).
Other TCS described as being involved in phenotype variation in Gram-negative bacteria is
GacS/GacA, wherein GacS is the sensor kinase and GacA is the response regulator. This regulatory
system belongs to the FixJ family of transcriptional regulators, is present in a wide variety of Gram-
negative bacteria, and controls the production of secondary metabolites and extracellular enzymes
involved in virulence of plant and animal pathogenic bacteria, biocontrol of soil borne plant diseases,
ecological fitness, or tolerance to stress (Heeb and Haas 2001). It has been reported that the
GacA/GacS system is subject to the accumulation of mutations in several Pseudomonas spp.
Pseudomonas strain PCL1171 exhibits a high frequency of reversible phase variation between a thick,
opaque phase I and a thin, translucent phase II colony (Van den Broek et al. 2005). Mutations of the
gacA and gacS genes locked the bacteria in a phase II phenotype. The complementation of mutants
with the gacA and gacS genes, restored all mutants to a phase I phenotype, showing that
spontaneous mutation of the GacA/GacS TCS forms the basis of the switch from phase I to phase II in
these bacteria (Broek et al. 2005).
In Burkholderia, only one TCS was described in detail, and is called AtsR/AtsT, wherein AtsR
is a membrane-bound hybrid sensor kinase and AtsT is a cytosolic response regulator. The AtsR/AtsT
system is a major global regulator of B. cenocepacia pathogenicity and regulates negatively quorum
sensing and virulence factors such as biofilm formation, type VI-secretion and protein secretion
(Khodai-Kalaki et al. 2013). It was observed that in the absence of atsR, the expression of cepIR and
cciIR quorum-sensing is up-regulated and mediates early and increased N-acylhomoserine lactone
production, which suggests an AtsR role in controlling virulence gene expression by modulating the
timing of quorum sensing signaling (Aubert et al. 2008). AtsR was also seen to repress the expression
of virulence genes (Aubert et al. 2008). The inactivation of atsR in B. cenocepacia leads to increased
biofilm formation, adherence to polystyrene and lung epithelial cells, extracellular protease secretion,
and expression of a type VI secretion system. It was demonstrated, by in vitro phosphorylation, that
histidine 245 and aspartic acid 536 are conserved sites of phosphorylation in AtsR. His-245 residue is
absolutely essential for initiation of signal transduction and its autophosphorylation is crucial for AtsR

Introduction _____________________________
13
function in vitro and in vivo. The Asp-536, on the other hand, plays a role in modulating the stability of
phosphorylated AtsR (Khodai-Kalaki et al. 2013). In short, AtsR phosphorylation has a significant
biological relevance as a global virulence regulator, modulating the expression of proteases through
AtsT. However, more experiments are needed to identify the genes that are specifically controlled by
AtsT, as well as the environmental signals that trigger activation.
Due to their versatility in sensing diverse intracellular and extracellular signals and their
variable modular architecture, TCSs are convenient devices for the regulation of the expression of
virulence properties. However, despite the detailed knowledge about the phosphorylation-based signal
transduction mechanism itself, little information is available about the molecular basis for this type of
signaling in bacterial virulence, and more studies are needed to understand TCSs, particularly in Bcc
bacteria.
1.3.2 Role of a EnvZ/OmpR-like TCS in mucoid morphotype variation in
B. multivorans
Mucoid-to-nonmucoid morphotype variation due to the presence or absence of cepacian
biosynthesis has been demonstrated for several Burkholderia cepacia complex species exposed to
subinhibitory concentrations of antibiotics or prolonged stationary phase (Silva et al. 2013). Among the
tested strains, the clinical CF isolate B. multivorans D2095, showed the highest frequency of
nonmucoid colonies arising from the original mucoid ones and was chosen to investigate the
molecular basis of this morphotype variation. The approach followed was to determine the genome
sequence of the parental B. multivorans D2095 and 10 nonmucoid variants obtained under several
stress conditions imposed to the parental strain. Comparative genomic analysis revealed that the
majority of the nonmucoid variants harbored mutations (singe nucleotide polymorphism (SNP), indels
and frameshift mutations) in a gene encoding a response regulator of a TCS annotated as
EnvZ/OmpR-like. Furthermore, trans complementation of these D2095-derived nonmucoid variants
with the wild-type copy of the ompR-like gene restored cepacian biosynthesis and the mucoid
morphotype (Silva et al., unpublished data).

Introduction _____________________________
14
1.4 Objectives
Morphotype variation is a phenomenon that has been described in CF patient’s airways. In CF
lungs, bacterial pathogens are challenged with a harsh environment that leads to the emergence of
phenotypic variants (Lyczak et al., 2002). Mucoid-to-nonmucoid morphotype variation was also seen
to occur in vitro and to be triggered by stress conditions with relevance in the CF airways (Silva et al.,
2013). Comparative genomics and gene expression studies have identified the consistent reduction of
the expression of several virulence factors in nonmucoid isolates, relatively to the mucoid ones (Silva
et al., 2013). Between these genes, is a gene encoding a transcriptional regulator belonging to the
LysR family of transcriptional regulators (LTTRs) and also a gene encoding a response regulator from
a two-component system (TCS), OmpR (unpublished results). Therefore, the aim of this work was to
evaluate the contribution of these two regulators to exopolysaccharide biosynthesis and the mucoid-
to-nonmucoid morphotype variation. To achieve that, we report the study of the Bmul_2557 gene,
encoding the LTTR that showed consistent decreased expression in Burkholderia nonmucoid variants,
in comparison with the respective mucoid wild-type. A strategy to complement a previously
constructed deletion mutant for this gene was developed and phenotypic assays of B. multivorans
ATCC 17616, its isogenic deletion mutant and complemented strain were envisaged, in order to
assess the influence of this regulator at carbohydrate metabolism level. Furthermore, computational
analysis to characterize orthologous of this regulator in Burkholderia and studying the degree of
conservation in other Bcc were also planned. A second part of this work will be focused on the study
of the OmpR-like protein, a response regulator from the EnvZ/OmpR TCS. As mutations in OmpR
encoding gene were also found in nonmucoid variants obtained in vitro, we also aim to comprehend
the relevance of this RR in mucoid-to-nonmucoid morphotype variation. A plasmid containing ompR
gene will be mobilized to nonmucoid variants derived in vitro from several mucoid B. multivorans
clinical isolates. The aim is to observe if after triparental mating, complementation of the nonmucoid
Burkholderia variant with the ompR gene, occurs, and in this case, if the original mucoid phenotype
will be restored. In addition, a novel strategy to construct a deletion mutant in ompR gene to allow
further understanding of the influence of this regulator in mucoid morphotype variation in B.
multivorans is planned.

Materials and Methods _____________________________
15
2. Materials and Methods
2.1 Bacterial strains, plasmids and culture conditions
The bacterial strains and plasmids used in this study are described in Table 2.1 and Table 2.2,
respectively. E. coli was grown at 37oC in Lennox Broth (LB) supplemented with kanamycin (50
µg/ml) or chloramphenicol (25 µg/ml) when required to maintain selective pressure. Burkholderia
strains were grown in LB or in S (Richau et al. 2000), SM, SGal, SMan or SSuc media (12.5 g/l
Na2HPO4.2H2O, 3 g/l KH2PO4, 1 g/l K2SO4, 1 g/l NaCl, 1 g/l Yeast Extract, 1 g/l Casamino acids, 20
g/l of glucose (in S medium), mannitol (in SM medium), galactose (in SGal medium), mannose (in
SMan medium) or sucrose (in SSuc medium), at 37°C. The deletion mutant in Bmul_2557 gene in B.
multivorans ATCC 17616 was grown in LB supplemented with trimethoprim (100 μg/ml). The
deletion mutant in Bmul_2557 gene complemented with pARG015-1 was grown in LB supplemented
with chloramphenicol (200 μg/ml).
Table 2.1 - Bacterial strains used in this study.
Strain Relevant Characteristics Source/ Reference
B. multivorans ATCC 17616 Soil isolate, USA (Vandamme et al. 1997)
B. multivorans ATCC 17616 ΔBmul_2557::dhfR
B. multivorans ATCC 17616 deletion mutant in Bmul_2557 gene; Tp
R
(Silva et al, unpublished)
B. multivorans ATCC 17616 ΔBmul_2557::dhfR + pARG015-1
B. multivorans ATCC 17616 ΔBmul_2557::dhfR complemented with Bmul_2557 gene, its
promoter region, and also Bmul_2558 gene; TpR,
CmR
This study
B. multivorans D2095 CF clinical isolate, Canada, EPS+
(Zlosnik et al, 2008)(Silva et al. 2011)
B. contaminans IST408 CF clinical isolate, Portugal, EPS+ (Richau et al. 2000)
B. multivorans HI229 Soil isolate, USA (Vandamme et al. 1997)
B. multivorans NMV121 Nonmucoid variant obtained in vitro from an
original CF clinical isolate BM4; EPS-
(Silva et al, 2013)
B. multivorans NMV122 Nonmucoid variant obtained in vitro from an
original CF clinical isolate BM4; EPS-
(Silva et al, 2013)
B. multivorans NMV123 Nonmucoid variant obtained in vitro from an
original CF clinical isolate BM4; EPS-
(Silva et al, 2013)
B. multivorans NMV124 Nonmucoid variant obtained in vitro from an
original CF clinical isolate BM6; EPS-
(Silva et al, 2013)
B. multivorans NMV126 Nonmucoid variant obtained in vitro from an
original CF clinical isolate BM6; EPS-
(Silva et al, 2013)
B. multivorans NMV127 Nonmucoid variant obtained in vitro from an
original CF clinical isolate BM6; EPS-
(Silva et al, 2013)
B. multivorans NMV129 Nonmucoid variant obtained in vitro from an
original CF clinical isolate BM7; EPS-
(Silva et al, 2013)
B. multivorans NMV130 Nonmucoid variant obtained in vitro from an
original CF clinical isolate BM9; EPS-
(Silva et al, 2013)
B. multivorans NMV131 Nonmucoid variant obtained in vitro from an
original CF clinical isolate BM9; EPS-
(Silva et al, 2013)
B. multivorans NMV132 Nonmucoid variant obtained in vitro from an
original CF clinical isolate BM9; EPS-
(Silva et al, 2013)
B. multivorans NMV133 Nonmucoid variant obtained in vitro from an
original CF clinical isolate BM9; EPS-
(Silva et al, 2013)
Escherichia coli DH5α recA1, lacUi69, Φ80dlacZΔM15 Gibco BRL

Materials and Methods _____________________________
16
Table 2.2 - Plasmids used in this study.
2.2 DNA manipulation
Genomic DNA from Burkholderia was extracted and purified using the DNeasy blood and
tissue kit (Qiagen) following the manufacturers’ recommendations. A fragment of 2.7 kb of B.
multivorans ATCC 17616 genome containing the promoter region of Bmul_2557, Bmul_2557 and
Bmul_2558 genes was amplified by polymerase chain reaction (PCR) using P1 and P2 primer
sequences (Supplementary Table 1) under the following conditions: 5 min at 95ºC; 34 cycles of 30
seconds at 95ºC, 45 seconds at 57ºC and 1 min at 72ºC; followed by an additional extension step at
72ºC for 7 min. PCR reaction mixture included 1 ng/µl of B. multivorans ATCC 17616 DNA, 200 µM
dNTPs, 0.5 pmol/µl of each primer oligonucleotide, 1.5 µM Mg2SO4 and 2U of Taq DNA polymerase
(Citomed). Amplification product was separated by 0.8% (w/v) agarose gel electrophoresis at 10
V/cm. For amplified DNA purification, ZymoResearch gel extraction kit was used. Plasmid DNA was
extracted and purified using ZymoResearch Miniprep kit following the manufacturers’
recommendations. Plasmid and amplified DNA were restricted with HindIII and DNA ligation of the
amplified DNA to pBBR1-MCS was performed using standard protocols (Sambrook, 2001). E. coli
DH5α cells were transformed by classic tranformation and grown for 1 hour at 37ºC before plating in
selective media supplemented with 0.1 mM X-gal and 0.1 mM IPTG. The originated plasmid,
pARG015-1 (pBBR1MCS containing the promoter region of Bmul_2557, Bmul_2557 and Bmul_2558
genes) was confirmed by DNA sequence determination. pARG015-1 plasmid was mobilized into B.
multivorans ATCC 17616 ΔBmul_2557::dhfR mutant, by triparental conjugation using plasmid
pRK2013 as helper. Transformants were selected in LB plates supplemented with 100 µg/ml
ampicillin and 200 µg/ml chloramphenicol.
2.3 Phenotypic Characterization of Burkholderia strains
To assess growth of Burkholderia strains, triplicate cultures were grown in 100 ml of S, SM,
SGal, SMan or SSuc media (initial OD640nm of 0.1) at 37ºC, with orbital agitation at 250 rpm for 7
days. To measure pH, 2 ml aliquots were taken at each day, centrifuged at 13,000 g for 5 min to
remove cells and the pH of the supernatant was registered. At the same time, aliquots of 100 μl were
collected and serially diluted with NaCl 0.9%, spread onto the surface of LB plates, and incubated at
37ºC for 2 days. Plates were examined with respect to the number of colony forming units (CFU). 1-
Plasmid Relevant Characteristics Source/ Reference
pBBR1MCS
Cm
R, 4.7kb, broad-host-range cloning vector (Kovach et al. 1994)
pLM014-5 pBBR1MCS derivative containing 1.2-kb HindIII/XbaI fragment with ompR coding
and promoter region Silva et al, unpublished
pRK2013 Tra
+ Mob
+ (RK2) Km::Tn7 ColEl origin, helper
plasmid, KmR
(Figurski and Helinski, 1979)
pARG015-1 pBBR1MCS containing the promoter region of
Bmul_2557, Bmul_2557 and Bmul_2558 genes
This study

Materials and Methods _____________________________
17
ml aliquots of cell culture were also taken at each day, centrifuged at 13,000 g for 5 min and
supernatants were stored at -20ºC before high pressure liquid chromatography (HPLC) analysis.
2.4 Exopolysaccharide quantification
The amount of EPS was assessed based on the dry weight of the ethanol-precipitated
polysaccharide recovered from triplicates of 100 ml culture samples (supplemented with NaCl (0.1%
w/v) of the different strains grown in liquid SM and S medium at 30°C, with orbital agitation at 250
rpm for 7 days.
2.5 HPLC analysis
Cell free supernatants of Burkholderia cultures were diluted 1:5 factor with mobile phase
(H2SO4 5 mM). Standard solutions of D-glucose, D-galactose, 2-keto gluconic acid, 5-keto gluconic
acid and D-lactate were prepared, in the following concentrations: 50 mM, 25 mM, 5 mM, 2.5 mM
and 0.5 mM for 2-keto gluconic and 5-keto gluconic acids; 200 mM, 100 mM, 50 mM, 10 mM, 5 mM
and 1 mM for D-glucose and D-galactose, and 100 mM, 50 mM, 10 mM, 5 mM and 1 mM for D-
lactate (Figure 2.1). HPLC was performed using an Aminex HPX-87H column (BioRad) at 65ºC
using 5mM H2SO4 at a constant flow of 0.6 ml/min. For a qualitative analysis, the retention times of
peaks obtained for each sample, in UV-Vis and RI detectors, were compared with those obtained for
standard samples, to identify the compound produced by B. multivorans strains during growth.
A
B
C
D
Figure 2.1 – D-Glucose (A), D-galactose (B), 2-KG acid (C) and D-lactate (D) standard curves determined by HPLC.

Materials and Methods _____________________________
18
2.6 Triparental conjugation
Nonmucoid B. multivorans nonmucoid variants (Table 2.1) were complemented with pLM014-
5 or pBBR1MCS as negative control, by triparental conjugation using pRK2013 helper plasmid.
Transformants were selected in EPS-producing Yeast Extract Mannitol (YEM) plates (Zlosnik et al.,
2008), supplemented with 100 µg/ml ampicillin and 200 µg/ml chloramphenicol.
2.7 Construction of the ompR gene replacement vector with
Gateway-compatible allelic exchange system
To generate ompR mutants in B. multivorans strains, the upstream fragment of ompR gene
was amplified using the primers P3 and P4 (Supplementary Table 1) and the downstream fragment
of ompR gene was amplified using the primers P5 and P6. Both fragments were amplified using Pfu-
X High-Fidelity DNA polymerase (Jena Bioscience) according to the manufacturer’s instructions and
the following thermal cycling conditions: 98 °C for 2 min; 30 cycles of 98 °C for 20 sec, 57 °C for 20
sec (ompR upstream region) or 54ºC for 20 sec (ompR downstream region), and 72 °C for 1 min; a
final extension step of 72 °C for 7 min. The PCR fragments were separated by 0.8% (w/v) agarose
gel electrophoresis at 10 V/cm, purified with ZymoResearch gel extraction kit and their
concentrations and quality were estimated using an UV spectrophotometer (ND-1000 UV-Vis,
NanoDrop Technologies, USA). The up- and downstream fragments were fused together and
amplified using the primers P7 and P8 (Supplementary Table 1) in splicing-by-overlap extension
PCR to generate the ompR mutant allele. Equal amounts (50 ng) of each up- and downstream
fragments and the other components of the PCR reaction except the primers P7 and P8 were mixed.
The PCR reaction will be carried out using the following thermal cycling conditions: 98 °C for 2 min;
10 cycles of 98 °C for 15 sec, 58 °C for 30 sec and 72 °C for 1 min; and a final extension step of 72
°C for 1 min. The final extension step will be paused at 30 sec, the primers GW-attB1 and GW-attB2
will be added, and the thermal cycling continued with 27 cycles of 98 °C for 15 sec, 64 °C for 30 sec
and 72 °C for 2 min; and a final extension step of 72 °C for 7 min. The PCR product will be
separated by 0.8% (w/v) agarose gel electrophoresis at 10 V/cm.
2.8 In silico analysis of nucleotide and amino acid sequences
The BLAST algoritm (Altschul et al. 1997) was used to compare sequences of the deduced
amino acids to database sequences available at the National Center for Biotechnology Information
(NCBI). Alignments were performed using the program CLUSTALW (Thompson et al. 1994).
Molecular evolutionary relationships between Bmul_2557 homologues were examined by the
neighbor joining method of tree construction. Alignment of these proteins was performed with
clustalX (1.81). Phylogenetic tree was displayed by Tree View 4.5.

Materials and Methods _____________________________
19
2.9 Genome sequence determination and detection of SNPs and
indel mutations
Genomic DNA from the 5 B. multivorans nonmucoid variants was extracted and purified using
the DNeasy blood and tissue kit (Qiagen). DNA samples were processed according to Illumina
instructions and sequenced at Instituto Gulbenkian da Ciência. Raw paired-end reads were filtered
based on Phread quality scores and adapter contamination and ambiguous nucleotides trimmed off.
These datasets were mapped against reference draft genome sequence of B. multivorans D2095
(Silva et al., 2015) using Bowtie2 v2.2.4 and samtools and detection of mutations was carried out
using Breseq v0.26.1. Predicted mutations were inspected for average, mutation frequency and
presence in forward and reverse reads using Geneious v6.1.8 (Kearse et al. 2012).

Results _____________________________
20
3. Results
3.1 Role of Bmul_2557 LysR transcriptional regulator
3.1.1 Characterization of the Bmul_2557 gene and homologs through
computation tools
Bmul_2557 gene encodes a LysR-type transcriptional regulator (LTTR) that has shown
consistent decreased expression in nonmucoid Burkholderia isolates and variants compared with the
respective mucoid ones (Tavares, 2012, MSc Thesis). In general, LTTRs can regulate the
transcription of genes involved in metabolism, quorum sensing, virulence, motility, toxin production,
attachment and secretion (reviewed in Maddocks & Oyston, 2008; Schell, 1993). However, as a LTTR,
Bmul_2557 or its homologs has not yet been characterized.
To better understand the role and importance of this LTTR, a bioinformatic characterization of
this protein was performed. By accessing the NCBI (National Center for Biotechnology Information)
site it was possible to observe that the LTTR encoded by Bmul_2557 gene of B. multivorans ATCC
17616 is a cytoplasmic protein, composed by 327 amino acids, with two highly conserved domains:
the HTH_1 domain and the PBP2 CrgA-like domain (figure 3.1). HTH_1 domain functions as
sequence-specific DNA-binding domain. PBP2 CrgA-like domain includes the substrate binding
domain of LysR-type transcriptional regulator CrgA, from Neisseria meningitidis, and its related
homologs. In contrast to the tetrameric form of other LTTRs, CrgA-like regulators form a subclass of
the LTTRs that function as octamers. The structural topology of this substrate-binding domain is most
similar to that of the type 2 periplasmic binding proteins (PBP2), which are responsible for the uptake
of a variety of substrates such as phosphate, sulfate, polysaccharides, lysine/arginine/ornithine, and
histidine.
Figure 3.1 – Putative conserved domains for Bmul_2557 encoded LTTR of B. multivorans ATCC 17616 as obtained at NCBI
conserved domains database.
To ascertain if this LTTR is present in other Burkholderia strains, a local alignment based on
protein sequences (BLASTp) was perfomed against the Burkholderia cepacia complex (taxid:87882)
and the Burkholderia genus (taxid:119060). This alignment tool compares the LTTR amino acid
sequence with amino acid sequences present in the selected groups and calculates the statistical
significance of matches. It was observed that the majority of the sequences produce alignments with
high coverage relatively to the query sequence, which indicates that this regulator is conserved and
may be important for other Burkholderia species. A phylogenetic tree based on a multiple sequence

Results _____________________________
21
alignment between the amino acid sequence from Bmul_2557 and its homologs from the Burkholderia
genus is depicted in figure 3.2. Protein Bmul_2557 of B. multivorans clusters with the representatives
from other Bcc species. The closest non-Bcc representatives include the phytopathogen B. glumae
and the animal pathogens B. mallei and B. pseudomallei. All the other strains from environmental
sources clustered a part (Figure 3.2).
Figure 3.2 – Phylogenetic three comprising 29 Burkholderia homologues of Bmul_2557 aligned by clustal X and sorted by tree
view. The Bmul_2557 homologues included are: B. multivorans ATCC 17616 (Bmul_2557); B. dolosa AU0158 (gi|124895726);
B. lata (gi|499670367); B. contaminans FFH2055 (gi|816522179); B. vietnamiensis (gi|780384099); B. ambifaria AMMD
(gi|115280748); B. cepacia GG4 (gi|402248647); B. jiangsuensis (gi|737523741); B. zhejiangensis (gi|736451165); B. glathei
(gi|837911553); B. sordidicola (gi|737595895); B. phytophirmans (gi|187715317); B. heleia (gi|754903957); B. nodosa
(gi|737538926 partial); B. ferrariae (gi|754982598 partial); B. acidipaludis (gi|654280604); B. bannensis (gi|754940653 partial);
B. oxyphila (gi|759568007 partial); B. sacchari (gi|737554315 partial); B. caribensis (gi|738045076); B. phymatum
(gi|501370240); B. thailandensis (gi|492895054); B. oklahomensis (gi|497802638); B. mallei (gi|752608015); B. pseudomallei
group (gi|497791808); B. glumae (gi|738020558); B. pyrricinia (gi|515902743); B. ubonensis (gi|497777334); B. cenocepacia
DDS 22E-1 (gi|685647057);
To infer about the genomic organization and search for orthologues of Bmul_2557 LTTR in
other Burkholderia strains, the Burkholderia genome database was used
(http://www.burkholderia.com/). This site functions as a resource for updated annotations and
comparative genomic analyses. By doing the search for orthologues of Bmul_2557 in other
Burkholderia strains and the comparison of the genomic loci containing the gene it was observed that
the genomic organization for the majority of the Burkholderia strains is very similar (Figure 3.3).

Results _____________________________
22
Figure 3.3 – Genomic organization of B. multivorans ATCC 17616, B. multivorans D2095, B. cepacia GG4, B. xenovorans
LB400 and B. phytofirmans PsJN. The colors represent different subcellular localizations: ■ – cytoplasm; ■ – cytoplasmic
membrane; ■ – unknown localization; ■ – extracellular; ■ – outer membrane; The numbers represent the types of proteins: 1-
LTTR; 2- Dehydrogenase; 3- Reductase; 4- Major facilitator transporter; 5- Hypothetical protein; 6- AraC family transcriptional
regulator; 7- Aminotransferase; 8- Arabinose transporter permease; 9- FAD-dependent oxidoreductase; 10- 2-dehydropantoate
2-reductase; 11- Aldolase II superfamily protein; 12- RND efflux system membrane fusion protein; 13- OmpC family outer
membrane protein;14- Phosphonoacetaldehyde hydrolase; 15- 2-aminoethylphosphonate – pyruvate transaminase; 16-Short-
chain dehydrogenase/reductase SDR; 17- Diguanylate cyclase.
In B. multivorans ATCC 17616, gene Bmul_2557 is flanked upstream by a putative AraC
family transcriptional regulator encoding gene transcribed in the opposite orientation. Downstream in
the same orientation is located Bmul_2558 gene encoding a putative D-isomer specific 2-hydroxyacid
dehydrogenase (lactate dehydrogenase). As Bmul_2557 and Bmul_2558 genes are separated by 33
nucleotides it is possible that they are in the same operon. In many Bcc strains, such as B.
multivorans D2095 and B. cepacia GG4, a similar genetic organization was also seen. Contrastingly,

Results _____________________________
23
in other Burkholderia strains that do not belong to Bcc, such as B. xenovorans LB400 and B.
phytofirmans PsJN, a different organization of this locus was seen. In the first case, the genes located
downstream of the ortologue, Bxe_B1227, encode a FAD-dependent oxidoreductase, an hypothetical
protein, a 2-dehydropantoate 2-reductase and an aldolase II superfamily protein. In B. phytofirmans
PsJN, downstream to the ortologue, Bphyt_2140, are located a phosphonoacetaldehyde hydrolase, a
2-aminoethylphosphonate-pyruvate transaminase, another LTTR and a short-chain
dehydrogenase/reductase SDR.
To infer about the protein structure of Bmul_2557 LTTR, we resort to I-TASSER,
a bioinformatics method for predicting three-dimensional structure model of protein molecules from
amino acid sequences. It detects structure templates from the Protein Data Bank by a technique
called fold recognition (or threading). The full-length structure models are constructed by reassembling
structural fragments from threading templates using replica exchange Monte Carlo simulations. The
threading templates used by I-TASSER for predicting Bmul_2557 model structure were the CrgA
LysR-type transcriptional regulator from Neisseria meningitidis, the AphB LTTR from Vibrio cholerae
virulence activator and the probable transcriptional regulator PA0477 from Pseudomonas aeruginosa.
The model in figure 3.4 shows the N-terminal α-helix rich regions involved in DNA binding, while the C-
terminal region is mostly β-strand.
We also performed a multiple alignment with CrgA from Neisseria meningitidis, AphB from
Vibrio cholerae and several Burkholderia homologs. Through this multiple alignment we observed that
the N-terminal region is more conserved than the C-terminal, which was expected because the N-
terminal is responsible for DNA-binding while the C-terminal is related with substrate binding, which is
dissimilar between these regulators (figure 3.4).
Figure 3.4 – Multiple Sequence Alignment of Bmul_2557 homologs and 3D-structure as predicted by I-TASSER.

Results _____________________________
24
3.1.2 Complementation of B. multivorans ATCC 17616 Bmul_2557 gene
deletion mutant
To better understand the role of the LysR protein encoded by Bmul_2557 gene, a deletion
mutant was previously obtained and named ΔBmul_2557::dhfR (Tavares, 2012, MSc Thesis). In an
initial strategy to complement this deletion mutant the Bmul_2557 gene was cloned into pBBR1MCS
and expressed from the bce promoter that directs the expression of the bce genes responsible for
cepacian biosynthesis. The introduction of this plasmid into ΔBmul_2557::dhfR mutant did not restore
phenotypes displayed by the mutant (described in the section below). Two possible explanations for
this result are the inefficient expression of Bmul_2557 from the bce promotor, or not only Bmul_2557
has to be expressed but also Bmul_2558. As these two genes may be in an operonic structure,
replacing of Bmul_2557 by the dhfR cassette might have affected the expression of Bmul_2558,
explaining our results. To overcome that, a strategy to obtain another plasmid for complementation of
B. multivorans ATCC 17616 ΔBmul_2557::dhfR mutant was designed (Figure 3.5), and included the
Bmul_2557 gene, its promoter region, and also gene Bmul_2558 cloned into pBBR1MCS cloning
vector.
Figure 3.5 – Cloning strategy to obtain pARG015-1 plasmid. Plasmid pBBRMCS-1 serves as backbone of the expression vector
pARG015-1.
A fragment of 2.7 kb was amplified by PCR from genomic DNA of B. multivorans ATCC
17616. The ligation between the amplified DNA product and pBBR1MCS DNA was performed (section
2 Material & Methods) and the resulting plasmid, pARG015-1, was then introduced into E.coli DH5α
cells using the heat shock method (classic transformation) and the transformants were selected. After
that, it was performed a triparental conjugation where E.coli DH5α was used as donor strain of the
pARG015-1 plasmid to B. multivorans ATCC 17616 ΔBmul_2557::dhfR. Transconjugants were
selected and mutant complementation was confirmed by phenotypic assays.

Results _____________________________
25
3.1.3 Role of Bmul_2557 LysR regulator in cepacian biosynthesis
Since gene Bmul_2557 displayed decreased expression in nonmucoid isolates (Tavares,
2012, MSc Thesis), we postulated this gene could be involved in the expression of the mucoid
phenotype. To test this hypothesis, the wild-type B. multivorans ATCC 17616, the ΔBmul_2557::dhfR
deletion mutant and the complemented mutant harboring pARG015-1 were grown in polysaccharide
producing medium supplemented with glucose or mannitol. In mannitol containing medium, all strains
produced between 10-12 g/l of cepacian after 48 hours and that value remained approximately
constant for the next two days (Figure 3.6 A). Regarding growth in glucose containing medium, only
the ΔBmul_2557::dhfR deletion mutant produced cepacian (Figure 3.6 B). Although the same amount
(~12 g/l) as in mannitol containing medium was reached, detection of this exopolysaccharide in the
culture supernatant was delayed by 48 hours. From this experiment we could conclude that
Bmul_2557 transcriptional regulator did not have a significant influence in cepacian biosynthesis in
mannitol supplemented medium as the wild-type, ΔBmul_2557 deletion mutant and the complemented
mutant produced similar levels. Nevertheless, the result obtained in the presence of glucose was
unexpected since, in a Bmul_2557 gene background, these strains were unable to produce any
polysaccharide and we could clearly see cell lysis. Only in the absence of the Bmul_2557 gene was
possible to obtain cepacian production. This observation prompts us to investigate the effect of several
sugars, and glucose in particular, in growth properties of the strains under study.
A B
Figure 3.6 – Exopolysaccharide production by B. multivorans ATCC 17616 (▲), ΔBmul_2557::dhfR mutant (Δ) and
ΔBmul_2557::dhfR + pARG015-1 (▲) in the presence of D-mannitol (SM medium) (A) and D-glucose (S medium) (B) at 37ºC.
3.1.4 Growth of the LysR mutant in the presence of glucose or mannitol
To assess growth of B. multivorans ATCC 17616, ΔBmul_2557::dhfR, and ΔBmul_2557::dhfR
+ pARG015-1, cultures were inoculated in S medium (glucose as carbon-source) and SM medium
(mannitol as carbon source) during 7 days, and the OD640nm, CFUs and pH of the culture supernatant
were registered (Figures 3.7; 3.8 and 3.9).

Results _____________________________
26
A
B
Figure 3.7– Growth curves of B. multivorans ATCC 17616 (▲), ΔBmul_2557::dhfR mutant (Δ) and ΔBmul_2557::dhfR +
pARG015-1 (▲) in SM (A) and S medium (B) at 37ºC. The data are based on mean values from the results of three
independent cell cultures.
A
B
Figure 3.8 – Number of viable cells of B. multivorans ATCC 17616 (▲), ΔBmul_2557::dhfR mutant (Δ) and ΔBmul_2557::dhfR
+ pARG015-1 (▲) in SM (A) and S medium (B) at 37ºC. The data are based on mean values from the results of three
independent cell cultures.
A
B
Figure 3.9 – Culture supernatant pH values of B. multivorans ATCC 17616 (▲), ΔBmul_2557::dhfR mutant (Δ) and
ΔBmul_2557::dhfR + pARG015-1 (▲) in SM (A) and S medium (B) at 37ºC. The data are based on mean values from the
results of three independent cell cultures.
In SM medium with mannitol as carbon source, all strains presented similar growth as
measured by OD640nm, or by assessing CFUs/ml (Figure 3.7 A and Figure 3.8 A) and similar culture
supernatant pH values (Figure 3.9 A). However, in S medium, with glucose as carbon source, several
differences were observed while reaching stationary phase. In wild-type B. multivorans ATCC 17616
and complemented mutant strain, both OD640nm (Figure 3.7 B) and CFUs/ml (Figure 3.8 B) decreased

Results _____________________________
27
as consequence of bacterial lysis. This effect was not seen, however, for the mutant strain, as the
OD640nm is maintained as well as the CFU number. Regarding the pH of culture supernatant, whilst in
the wild-type and in the complemented strain, the supernatant pH decreases until 3.5 and 3.6 by 48
hours of growth, respectively, in the mutant, the pH decreases until reaching a minimum of 5.1 and
thereafter, occurs a progressive neutralization of the culture medium until initial pH of 7.0, by the 7th
day of culture (Figure 3.9 B).
Given these results, is possible to observe that the mutant complementation appears to have
occurred efficiently with pARG015-1, which results in the restoration of the original phenotype in S
medium. Besides this, in contrast with what happens in SM medium, where all strains presented
similar growth, in S medium with glucose as carbon source, differences in growth were observed. In S
medium, in wild-type and complemented mutant cultures, cell lysis was observed, as suggested by the
decrease in the OD640nm. In the deletion mutant, however, this effect was not seen, and no cell lysis
occurred, suggesting that Bmul_2557-encoded LysR has a negative effect on cell growth in the
presence of glucose, probably being related with glucose metabolism.
Figure 3.10 – Alternative pathways of 6-
phosphogluconate formation (adapted from
(Sage et al. 1990). Direct oxidative or
phosphorylative pathways of glucose
utilization. Abbreviations: PP, periplasm; CP,
cytoplasm; Fru, fructose; Glu, glucose; Gna,
gluconic acid; 2-KG, 2-ketogluconic acid;
KGP, 2-keto-6-phosphogluconate; 6PGA, 6-
phosphogluconate; KDPG, 2-keto-3-deoxy-6-
phosphogluconate; GAP, glyceraldehyde-3-
phosphate; Mtl, mannitol; Pyr, pyruvate; FBP,
fructose-l, 6-bisphosphate; F6P, fructose-6-
phosphate; G6P, glucose-6-phosphate; Frk,
fructokinase; Gcd, glucose dehydrogenase;
Gad, gluconate dehydrogenase; Kgt,
transport system for 2-KG; Kgk, 2-ketoglucono- kinase; Kgr, KGP reductase, Gct, glucose transport system; GIk, glucokinase;
Zwf, G6P dehydrogenase; Edd, 6PGA dehydratase; Eda, KDPG aldolase; Fba, FBP aldolase; Fbp, fructose-l,6-bisphosphatase;
Pgi, phospfhoglucoisomerase.
The consumption of glucose occurs via two routes, the direct oxidative and the
phosphorylative pathways, that converge at 6-phosphogluconate (6PGA), which is metabolized to
produce the end metabolite pyruvate (Figure 3.10) (Allenza and Lessie, 1982). It is possible that when
B. multivorans ATCC 17616 wild-type strain grow with glucose as carbon source, the favored pathway
is the oxidative which leads to the formation of acids, like gluconic and 2-ketogluconic acids, causing
culture medium pH lowering that results in bacteria failure. Contrastingly, in the ΔBmul_2557::dhfR
mutant, the culture medium pH decrease until similar values of those obtained for the wild-type, but in
this case a restoration to pH initial values was observed. An hypothesis that explains these
observations is that only in the wild-type may occur the production of other acidic compounds,
resulting in bacteria death.

Results _____________________________
28
3.1.5 Analysis of extracellular metabolites by HPLC
To identify differences in metabolite composition of bacterial cultures supernatant of B.
multivorans ATCC 17616, B. multivorans ATCC 17616 ΔBmul_2557::dhfR mutant, and
ΔBmul_2557::dhfR + pARG015-1, both at qualitative and quantitative level, we performed HPLC
analysis of culture supernatants. By doing this, we aimed to identify and quantify the compound(s)
resulting from B. multivorans ATCC 17616 metabolism that may be responsible for further culture
acidification upon growth with glucose as carbon source, as well as carbon source (glucose)
consumption. During growth in S medium, aliquots of culture supernatant of the wild-type strain,
mutant strain and the complemented mutant were processed as described in Materials & Methods. As
standards, the metabolites 2-KG and 5-KG were chosen because they are products of glucose
metabolism through the oxidative pathway and D-lactate is the possible product of Bmul_2558 activity
and therefore, relevant in this study. For a qualitative analysis, the retention times of peaks obtained
for each sample were compared with those obtained for standard samples (Figure 2.1, section 2,
Materials & Methods), to identify the compound produced by the strains during growth. By analyzing
the chromatograms of the wild-type, mutant and complemented mutant supernatants in S medium
over time, the only differences were two peaks with retention times of 7.8 and 12.7 minutes, in the UV-
Vis and RI chromatogram, respectively. The area of the first peak was seen to increase in the wild-
type and complemented strains’ supernatant and remained stable in the following days whereas in the
mutant the peak area decreased after the second day of growth. Under the same conditions, the
retention time of the 2-KG was 7.8 minutes, which is indicative of the production of this acid during the
growth in S medium. The second peak at 12.7 minutes was present in the wild-type and
complemented mutant, but absent from the ΔBmul_2557::dhfR mutant. The retention time of D-lactate
used as standard was also 12.7 minutes. Glucose consumption was evaluated by assessing the
evolution of peak area with a retention time of 9.23 minutes in the RI detector.
A
B
Figure 3.11 – 2-keto gluconic acid (A) and D-lactate (B) production by B. multivorans ATCC 17616 (▲), ∆Bmul_2557::dhfR
mutant (Δ) and ΔBmul_2557::dhfR + pARG015-1 (▲) in S medium, at 37ºC. The data are based on mean values from the
results of three independent cell cultures.
In figure 3.11 A is represented the quantification of 2-KG in S medium, in B. multivorans ATCC
17616, its isogenic ΔBmul_2557::dhfR mutant and the complemented mutant strain. It is possible to

Results _____________________________
29
observe that B. multivorans ATCC 17616 reached 117 mM of 2-KG by the second day of growth, and
from there the concentration of this metabolite remains unchanged. In contrast, the ΔBmul_2557::dhfR
mutant reached 134 mM of 2-KG by the second day of growth but instead, a successive decrease in
the production of 2-KG occurred, presenting in the 6th day a practically null concentration.
Consumption of this acidic metabolite leads to a pH recovery by the mutant strain to physiological
levels. In contrast, the wild-type and complemented mutant strain, maintain 2-KG acid levels very high,
indicating no consumption. It is also interesting to note that the complemented mutant strain showed
concentration levels of 2-KG higher than the wild-type, suggesting faster consumption of glucose.
Regarding D-lactic acid production and accumulation in the culture supernatant, the wild-type strain
showed the highest concentration already at 24 hours of growth (Figure 3.11 B). Interestingly, from the
ΔBmul_2557::dhfR deletion mutant supernatant was not detected D-lactic acid, while the
complemented mutant showing D-lactic acid accumulation although at lower concentration than the
wild-type strain. Taking into acount glucose consumption (Figure 3.12), it is possible to observe that
the wild-type strain takes longer to metabolize the sugar, which is consistent with the lower 2-KG
accumulation (Figure 3.11 A). In contrast, both mutant and complemented mutant strain degrade
glucose faster, presenting in the second day of growth very low concentration of this metabolite and
higher levels of 2-KG.
Figure 3.12 – Glucose consumption of B. multivorans ATCC 17616 (▲), ΔBmul_2557::dhfR mutant (Δ) and ΔBmul_2557::dhfR
+ pARG015-1 (▲) in S medium, at 37ºC. The data are based on mean values from the results of three independent cell
cultivations.
3.1.6 Role of Bmul_2557 transcriptional regulator in the metabolism of
other sugars
To evaluate the effect of other sugars in B. multivorans ATCC 17616 growth and the effect of
Bmul_2557 LysR transcriptional regulator in controlling those sugars metabolism, the wild-type strain
and its isogenic mutant were cultured in S medium without glucose, supplemented with the following
carbon sources: D-galactose (SGal), D-mannose (SMan), and sucrose (SSuc). Growth parameters
such as OD640nm, CFUs and culture medium pH were registered for several days.
In SGal medium, with galactose as carbon source, both strains presented similar growth as
measured by OD640nm or by assessing CFUs, (Figure 3.13 A, Figure 3.13 B) and similar culture
supernatant pH values (Figure 3.13 C).

Results _____________________________
1
A
B C
Figure 3.13 – Growth curves (A), number of viable cells (B) and culture supernatant pH values (C) of B. multivorans ATCC
17616 (▲) and ΔBmul_2557::dhfR mutant (Δ) in medium supplemented with D-galactose at 37ºC. The data are based on mean
values from the results of three independent cell cultures.
Galactose can be metabolized by the oxidative pathway or the Leloir pathway. The profiles
obtained for growth in medium with galactose as carbon source can be explained with its metabolism
through the Leloir pathway (Figure 3.14).
Figure 3.14 – Leloir pathway (adapted from Frey 1996). The process begins with the initial phosphorylation of galactose to
galactose-1-phosphate by the action of galactokinase. Galactose-1-phosphate is transformed into glucose-1-P in the presence
of galactose-1-P uridylyltransferase and UDP-glucose, which acts as cofactor .
Galactose can be converted into glucose-1-P through the Leloir pathway, and after that,
glucose 1-P can be transformed in glucose-6-P, by the transfer of the phosphate group of the
galactokinase
α-D-galactose galactose 1-phosphate
UDP-glucose
Galactose 1-
phosphate
uridyltransferase
UDP-galactose
glucose 1-phosphate

Results _____________________________
31
glucose from the 1' to the 6' position, by phosphoglucomutase (Maxwell et al., 1962). Glucose-6-P is
an intermediate of the phosporylative pathway, which means that galactose can be metabolized via
this pathway.
To corroborate these results, the supernatant recovered during growth in SGal medium was
also analyzed by HPLC. For a qualitative analysis, the retention times of peaks obtained for each
sample were compared with those obtained for standard samples. In SGal medium, the wild-type and
ΔBmul_2557::dhfR mutant showed a peak with a retention time of 7.8 minutes in the UV-vis detector
corresponding to 2-KG production, and a peak with a retention time of 9.81 minutes in the RI detector,
corresponding to the carbon source used in SGal medium. No peak corresponding to D-lactic acid
was detected under these conditions.
A
B
Figure 3.15 – 2-keto gluconic acid production (A) and galactose consumption (B) of B. multivorans ATCC 17616 (▲) and
ΔBmul_2557::dhfR mutant (Δ) in SGal medium, at 37ºC. The data are based on mean values from the results of three
independent cell cultures.
In figure 3.15 A is represented the production of 2-KG acid in SGal medium, for B. multivorans
ATCC 17617 and its isogenic ΔBmul_2557::dhfR mutant. It is possible to observe that the wild-type
and mutant strains presented similar profiles, however while in the mutant 2-KG acid levels are
relatively constant (~40 mM) until the 4th day of growth, in the wild-type strain occured a slight increase
until that day (~60 mM). From that moment, both strains presented a decrease in the amount of 2-KG
acid. Relatively to galactose consumption (Figure 3.15 B), the profiles are once more similar and no
significant differences were observed between the two strains, corroborating the results described
above. These results can be explained if we take into account what occurs in other organisms, like
E.coli, where galactose can be metabolized through the De Ley-Dourodoff pathway. In this pathway,
D-galactose is oxidized to galactonate, which is converted into 2-keto galactonic acid (Lessie and
Phibbs 1984). This acid, similar to 2-KG, can be detected in the HPLC. Although the peak observed
can be due to this compound, we did not run this pure standard to assign its nature. Nevertheless, our
data suggests that D-galactose metabolism in B. multivorans ATCC 17616 involves simultaneously
the phosphorylative and oxidative pathways.

Results _____________________________
32
In SMan medium, with mannose as carbon source, no significant differences were found
between the wild-type and mutant strain, which is observed through the growth curves (Figure 3.16 A),
viable cells (Figure 3.16 B) and culture supernatant pH (Figure 3.16 C).
A
B
C
Figure 3.16 – Growth curves (A), number of viable cells (B) and culture supernatant pH values (C) of B. multivorans ATCC
17616 (▲) and ΔBmul_2557::dhfR mutant (Δ) in medium supplemented with D-mannose at 37ºC. The data are based on mean
values from the results of three independent cell cultures.
The profiles obtained with mannose can be explained by this sugar metabolism (Figure 3.17).
Mannose appeared to be metabolized via two routes: one involved the intracellular conversion to
glucose by mannose epimerase and the other one is the conversion to fructose by mannose
isomerase (Lessie and Phibbs 1984; Allenza et al., 1990). So, the bacteria can use the oxidative and
phosporylative pathway to metabolize this sugar. HPLC analysis was not performed in this experiment
impairing our conclusion of which one is predominantly being used.
Figure 3.17 –D-mannose metabolism. Mannose is converted in mannose 6-phosphate, by hexokinase. Then, mannose 6-
phosphate isomerase converts mannose 6-phosphate into fructose 6-phosphate (Adapted from (Allenza et al. 1990).
Mannose
Hexokinase
Mannose 6-phosphate
Mannose 6-
phosphate
isomerase Frutose 6-phosphate

Results _____________________________
33
When B. multivorans ATCC 17616 and ΔBmul_2557::dhfR were grown with sucrose as
carbon source (SSuc), the results were similar to the other carbon sources such as mannose and
galactose. However some differences were found between wild-type and mutant strain in the growth
curves, where the wild-type showed a higher growth than the mutated strain (Figure 3.18 A).
Regarding the number of viable cells (Figure 3.18 B), both strains presented similar values and no
significant differences were observed. However, while the mutant strain maintained the pH
approximately constant all over the time, the wild-type strain decreases the pH until the first day of
growth and from there occurs a recovering to the initial values (Figure 3.18 C).
A
B
C
Figure 3.18 – Growth curves (A), number of viable cells (B) and pH values (C) of B. multivorans ATCC 17616 (▲) and
ΔBmul_2557::dhfR mutant (Δ) in SSuc medium at 37ºC. The data are based on mean values from the results of three
independent cell cultures.
The profiles obtained during sucrose metabolism can be explained taken into account the
composition of this sugar. Sucrose is a disaccharide that is hydrolyzed in its two sugars, glucose and
fructose by invertase (Figure 3.19) (Combes and Monsan 1983). In Burkholderia, invertase is probably
an intracellular enzyme and, for sucrose to be degraded, it must enter the cell. In this situation,
glucose utilization does not result in strong pH variation due to secretion of acidic metabolites.

Results _____________________________
34
Figure 3.19 – Sucrose hydrolysis into glucose and fructose. The reaction is catalyzed by invertase.
3.1.7 Growth behavior of different Bcc isolates using glucose as carbon
source
To evaluate if the effect of D-glucose utilization in cell lysis seen for B. multivorans ATCC 17616
occurs in other Bcc isolates, B. multivorans D2095, B. multivorans HI2229 and B. contaminans IST408
were also cultured in S medium using glucose as carbon source, and OD640nm (Figure 3.20 A), CFUs
(Figure 3.20 B) and culture medium pH (Figure 3.20 C) were registered.
A
B
C
Figure 3.20 – Growth curves (A), viable cells (B) and culture supernatant pH values (C) for B. multivorans D2095 ( ), B. ●
multivorans HI229 (□) and B. contaminans IST408 ( ), in S medium at 37ºC. The data are based on mean values from the
results of three independent cell cultivations.
D-Glucose D-Fructose

Results _____________________________
35
In B. contaminans IST408 we did not observed significant cell lysis, as cultures reached the
stationary phase maintaining similar OD640nm and CFUs levels (Figure 3.20 A and Figure 3.20 B).
However, in B. multivorans D2095 and B. multivorans HI229, as happened with B. multivorans ATCC
17616, cell lysis occured. It was also interesting to observe that culture supernatant pH values for B.
contaminans IST408 decreased upon reaching stationary phase, by 48 hours of growth, reaching a
minimum of 5.9. However, contrastingly to the other isolates, in B. contaminans IST408, a recovery to
neutral values of pH was seen (Figure 3.20 C). This suggests that an unbalanced utilization of glucose
may be strain-specific or even species-specific because it only happens in B. multivorans isolates.
Nevertheless, more strains needed to be tested, including environmental strains of other Bcc and non-
Bcc species.
To complement the results, it was also performed an HPLC analysis of the supernatants of B.
multivorans D2095 and B. multivorans HI2229, cultured in S medium. The results demonstrated that
2-KG acid (Figure 3.21 A) and D-lactic acid (Figure 3.21 B) production and glucose consumption
(Figure 3.21 C) is similar in both strains, which corroborate the results for B. multivorans ATCC 17616
described previously.
A
B
C
Figure 3.21 – 2-keto gluconic acid production (A), D-lactate production (B) and D-glucose consumption (C) for B. multivorans
D2095 ( ) and B. multivorans HI229 (□), in S medium at 37ºC. The data are based on mean values from the results of three ●
independent cell cultivations.

Results _____________________________
36
3.2 Role of a OmpR-like response regulator
3.2.1 Complementation of nonmucoid B. multivorans variants with an
ompR-containing plasmid
Nonmucoid variants of B. multivorans D2095 were previously obtained in vitro (Silva et al,
2013). Whole-genome sequencing of those nonmucoid variants revealed that about 90% have
mutations in an open reading frame (BMD20_11660) encoding an OmpR-like response regulator (RR)
from a two-component system (TCS) (unpublished results). To assess whether mutations in this
ompR-like gene were specifically occurring in the nonmucoid variants from the D2095 clinical isolates
or were extended to other isolates, we have chosen four other isolates recovered from the same CF
patient. These isolates, named B. multivorans BM4, BM6, BM7 and BM9 were exposed to prolonged
stationary phase (21 days at 42ºC in SM medium). Several nonmucoid colonies were obtained and
several were kept for further analysis (Table 3.1).
Table 3.1 - Bacterial strains used in this study.
Isolate Nonmucoid variants
Complementation of the mucoid morphotype
with pLM014-5
B. multivorans BM4
NMV121
NMV122
NMV123
Yes
Yes
Yes
B. multivorans BM6
NMV124
NMV126
NMV127
Yes
Yes
Yes
B. multivorans BM7 NMV129 Yes
B. multivorans BM9
NMV130
NMV131
NMV132
NMV133
Yes
Yes
Yes
Yes
To evaluate the relevance of this RR in the nonmucoid morphotype displayed by the variants,
the ompR gene and its promoter region cloned into pBBR1MCS (pLM014-5) was mobilized into these
variants by triparental mating. The aim was to observe if after triparental mating, the nonmucoid
Burkholderia variant restored exopolysaccharide production. After the complementation with the ompR
containing plasmid pLM014-5, all of them restored the mucoid phenotype (Table 3.1). Examples of
complementation with the empty vector or with the ompR gene are shown in figure 3.22. These results
suggest that the nonmucoid phenotype is due, most likely, to mutations in ompR gene.

Results _____________________________
37
A pBBR1MCS pLM014-5
B pBBR1MCS pLM014-5
Figure 3.22 –Colony morphologies of B. multivorans NMV121 (A) and NMV124 (B) variants, in selective YEM medium after 72
hours at 30ºC, complemented with pBBR1 vector or pLM014-5.
Whole-genome sequencing of five nonmucoid variants that restored EPS production upon
complementation with the ompR-containing plasmid was performed, in order to confirm the presence
of mutations in ompR gene. Sequencing results (Table 3.2) confirmed that all of them have mutations
in the ompR-like gene. Nonmucoid variants NMV124 and NMV126 derived from B. multivorans BM6
had a SNP mutation leading to a non-synonymous substitution of Y233N in the OmpR-like protein
(Table 3.2). NMV129 derived from BM7 CF clinical isolate had a three nucleotide deletion in the ompR
gene coding sequence, affecting the amino acid at position 11 and two additional mutations in the
gene encoding histidinol dehydrogenase and in another gene encoding a HlyD family secretion
protein. Finally, variants NMV128 and NMV130 derived from B. multivorans BM9 had a frameshift
mutation in the ompR gene due to an insertion of two nucleotides at position corresponding to amino
acid 173 (Table 3.2). The deletion affecting one amino acid maps to the receiver domain, while the
substitution of Y233N is in the DNA binding domain.
Table 3.2 – Whole genome sequence results of some nonmucoid variants that restored EPS production upon complementation with the ompR-containing plasmid.
Clinical Isolate
Nonmucoid variant
Seq. id Position Mutation Annotation Gene Description
BM6 NMV 124 JFHP01000005 320516 A→T Y233N (TAC→AAC) BMD20_11660 OmpR-like RR
NMV 126 JFHP01000005 320516 A→T Y233N (TAC→AAC) BMD20_11660 OmpR-like RR
BM7 NMV 129
JFHP01000003 574640 Δ12 bp coding (860-871/1050
nt) BMD20_07790
HlyD family secretion protein
JFHP01000005 321182 (CGA)3→2 coding (29-31/726 nt) BMD20_11660 OmpR-like RR
JFHP01000014 90842 G→T G391C (GGC→TGC) BMD20_28895 Histidinol
dehydrogenase
BM9 NMV 128 JFHP01000005 320694 +CC coding (519/726 nt) BMD20_11660 OmpR-like RR
NMV 130 JFHP01000005 320694 +CC Coding (519/726 nt) BMD20_11660 OmpR-like RR

Results _____________________________
38
3.2.2 Strategy to obtain the B. multivorans D2095 deletion mutant in
ompR gene
To study the relevance of the OmpR-like regulator in morphotype variation, a novel strategy to
knock-out that gene in B. multivorans D2095 was initiated (Figures 3.23 and 3.25). The aim is to
obtain a deletion mutant to do the functional analysis of this regulator. With this strategy it is possible
to produce a deletion mutant without a selection marker, which is a major advantage because allows,
in a cleaner way, the study of OmpR in phenotype variation and impact in virulence.
Figure 3.23 – Construction of the gene replacement vector pENTRPEX18Tp-SceI pheS-ompR, BP clonase reaction and
transformation of B. multivorans D2095 (Adapted from Fazli et al. 2015). Green balloon represent E.coli DH5α cells and orange
ballons represent B. multivorans D2095.
pDONRPEX18Tp-SceI-pheS

Results _____________________________
39
Figure 3.23 shows the first part of the process. Firstly, a fragment of the upstream (985 bp)
and downstream (1099 bp) regions of the ompR-like gene was amplified by PCR, using primers P3/P4
and P5/P6, respectively (supplementary table 1, Appendix) (Figure 3.24). As non-specific amplification
occurred, the two fragments were purified from the agarose gel and quantified, to obtain the desired
concentration. The next step is the Gateway PCR, where by using primers P7 and P8, the upstream
(L’) and downstream (R’) regions are fused together to generate the ompR mutant allele. Amplification
of this fragment was attempted a few times, but with no amplicon being detected. Further optimization
on this step is required, but was not yet done. When this amplicon is obtained, it will be recombined
into pDONRPEX18Tp-SceI-pheS plasmid using BP clonase reaction. The product will be transferred
to competent E. coli DH5α cells and mobilized to B. multivorans D2095, by triparental conjugation. The
transformants will be selected in LB-agar plates containing trimethoprim and screened by colony PCR
to verify the insertion of the ompR gene flanking regions.
Figure 3.24 - Agarose gel electrophoresis of the PCR amplified DNA fragments located upstream and downstream of the
ompR-like encoding gene, from B. multivorans D2095. L, denotes 1 kb DNA ladder; OmpR Left, denote de fragment located
upstream of ompR encoding gene; and OmpR Right, denotes the fragment located downstream of ompR encoding gene.
Figure 3.25 depicts the second part of Gateway strategy. A single positive merodiploid clone
will be transformed with pDAI-SceI-pheS by triparental mating. The I-SceI endonuclease expressed
from the plasmid induces a break in the double DNA strand, stimulating the second homologous
recombination event that can happen in different locations, generating the desired gene deletion or the
wild-type allele, which is easily identified by PCR.
OmpR Left L
OmpR Right

Results _____________________________
40
Figure 3.25 – Transformation of B. multivorans D2095 with the expression vector pDAI-SceI-pheS and second homologous
recombination event (Adapted from Fazli et al. 2015). The green balloons represent E.coli DH5α cells and the orange ones
represent B. multivorans D2095.

Discussion _____________________________
41
4. Discussion
Bacteria belonging to the Burkholderia cepacia complex are important opportunistic pathogens
that cause severe infections in cystic fibrosis patients (reviewed in O’Sullivan and Mahenthiralingam
2005). Within the CF lung, colonizing bacteria is faced with adverse conditions, like high osmolarity,
heterogeneous distribution of oxygen and nutrients, high concentration of antimicrobials, and constant
challenge by the host immune defenses. These factors exert a selective pressure in colonizing
bacteria, and are thought to be the driving force of microevolution during their persistence in the CF
lung which results in chronic infections that leads, in many cases, to poorer clinical outcomes of the
infected patients (Lyczak et al. 2002; Döring et al. 2011). Genotypic and phenotypic variation are a
common phenomenon during the course of an infection, and have been described in many pathogens
including P. aeruginosa, where most phenotypic conversions occur from the nonmucoid to mucoid
morphotype (Govan et al.1996). In Bcc bacteria, phenotype transitions in isolates recovered from CF
patients have also been reported. However, contrastingly to P. aeruginosa, little is known about the
adaptive traits and mechanisms underlying this phenomenon. Regarding the discovery of the
regulatory mechanisms by which Bcc bacteria undergo mucoid-to-nonmucoid variation, global
transcriptomic profiling studies have been carried out in our laboratory. Comparative genomics and
gene expression studies have identified the consistent reduction of the expression of several virulence
factors in nonmucoid isolates, relatively to mucoid ones (Silva et al., 2013). Between these genes, was
a gene encoding a transcriptional regulator belonging to the LysR family of transcriptional regulators
(LTTRs) and also a gene encoding a response regulator from a two-component regulatory system
(TCS), from the OmpR family (unpublished results).
In this work, we report the study of gene Bmul_2557, the LysR-type transcriptional regulator
that showed consistent decreased expression in B. multivorans nonmucoid variant, in comparison with
the respective mucoid wild-type. A strategy to complement the deletion mutant for this gene was
developed and phenotypic assays of B. multivorans ATCC 17616, its isogenic deletion mutant and
complemented strain were performed, in order to study the influence of this regulator on EPS
production regulation and morphotype variation. Looking at cepacian biosynthesis, the strains reached
the same levels, suggesting that this regulator is not directly involved in the regulation of mucoid
phenotype. However, other traits potentially different such as virulence in Galleria mellonella acute
model of infection, biofilm formation and siderophore production were tested previously (unpublished
results). None of the phenotypes tested showed significant differences between B. multivorans ATCC
17616 and ΔBmul_2557::dhfR mutant, which indicates that, in contrast to what happens in N.
meningitidis and V. cholerae, where the LTTRs described are related with bacteria virulence
(Deghmane et al. 2002, Kovacikova et al. 2004), this LTTR is not involved in that.
Discarding a putative direct influence in the regulation of well-known virulence factors, we
hipothesize a role for this LTTR in carbon metabolism regulation. In S. aureus is described an LTTR,
CidR, that regulates the cidABC operon, which is involved in murein hydrolase activity and antibiotic
sensitivity. The cidR gene product positively regulates the transcription of cidABC in the presence of
acetic acid, produced as a result of the metabolism of glucose. The upregulation of cidABC expression

Discussion _____________________________
42
resulted in increased murein hydrolase activity produced by these cells (Yang et al. 2005). In
Pseudomonas sp. strain ADP, a LTTR called AtzR regulates the atzDEF operon in order to control de
nitrogen concentration. Expression of atzDEF is induced by cyanuric acid, the substrate of the
pathway, and repressed by nitrogen. AtzR is required for both nitrogen and cyanuric acid dependent
control (Porrúa et al. 2007). In our work, the ability of B. multivorans ATCC 17616 and
ΔBmul_2557::dhfR mutant to metabolize different carbon sources was compared. When growing with
mannitol, galactose, sucrose or mannose as carbon source, all strains presented similar growth.
However, in medium with glucose as carbon source, whilst in wild-type B. multivorans ATCC 17616
and complemented strain bacterial lysis occurred, in the mutant strain, this phenomenon did not
happened, suggesting that Bmul_2557-encoded LysR and the downstream lactate dehydrogenase
encoding gene, have a deleterious effect on cell survival. The consumption of glucose occurs via two
routes, the direct oxidative and the phosphorylative pathways, that converge at 6-phosphogluconate
(6PGA), which is metabolized to produce in the end pyruvate (Allenza and Lessie, 1982). It is possible
that when B. multivorans ATCC 17616 wild-type strain grows in S medium with glucose as carbon
source, the favored pathway is the oxidative which leads to the formation of acids, like gluconic and 2-
ketogluconic acids, causing culture medium pH lowering. Additionally, D-lactic acid is also being
produced and secreted to the culture medium, further decreasing the pH value to critical levels which
lead to cell death and lysis. Contrastingly, the ΔBmul_2557::dhfR mutant also oxidize D-glucose to 2-
keto gluconic acid with the concomitant lowering of the culture medium pH, but the values never
decrease that much, and cells recover to the medium pH to neutral levels. This behavior might have to
do with the absence of D-lactic acid in the mutant culture supernatant.
Based on the obtained data we propose that glucose metabolism in B. multivorans ATCC
17616 leads to excess of pyruvate formation. Among the different routes for pyruvate utilization is the
one leading to D-lactic acid production. Is then possible that production of D-lactate dehydrogenase
(encoded by Bmul_2558) is dependent on the induction of the LysR transcriptional regulator (encoded
by Bmul_2557) through binding of pyruvate or even the end product, D-lactate. In the absence of
Bmul_2557 gene, there is no expression of Bmul_2558 gene and therefore no D-lactic acid is being
produced. If we try to unravel the reason why bacteria produce such high concentrations of D-lactate,
probably responsible for its lysis, we must think about the natural habitat in which it was found. B.
multivorans ATCC 17616 is a soil isolated, that probably has to compete with other organisms to
survive. The D-lactate can be produced in order to be dissipated to the surrounding environment,
functioning as a toxic compound for organisms potentially threatening these bacteria. In a closed
environment such as laboratorial flask-growth, dispersion of D-lactate does not occur leading to
bacterial death. In line with our model are results from gene expression of Bmul_2558 obtained in S
medium at 22 hours of growth. Under these conditions the expression of this gene was 2-fold
decreased in the mutant when compared to the wild-type strain (unpublished results).
Glucose seems to be the only sugar resulting in strong pH acidification and B. multivorans
ATCC 17616 cell death. Although we expected the same behavior in the presence of D-galactose this
was not observed. Indeed, the culture medium pH decreased to approximately 5.3 for the wild-type
and Bmul_2557 mutant, but it recovered to neutral values for both strains. Possible explanations are

Discussion _____________________________
43
the simultaneous use of the phosporylative and oxidative pathways or a more efficient metabolization
of the carbon intermediates, so that no significant amount of D-lactate is produced. D-mannose
metabolism runs through the phosphorylative pathway and only a slight decrease of the culture
medium pH was observed. Sucrose catabolism results in D-glucose and D-fructose, which could lead
to the previously observed D-glucose effect. Nevertheless, no significant differences in growth were
observed for the wild-type and mutant strain. This can be explained if sucrose hydrolysis occurs
intracellularly due to the action of an intracellular invertase. In that situation, the most likely route for
glucose metabolisms would be the phosphorylative pathway.
When a comparison was made between B. multivorans ATCC 17616, and other clinical
isolates, two from the same species, B. multivorans D2095 and B. multivorans HI2229, and one from
another species, B. contaminans IST408, it was observed that B. multivorans HI2229 and B.
multivorans D2095 presented a similar profile of B. multivorans ATCC 17616. In contrast, in B.
contaminans IST408 cell lysis did not occur, which suggests that this phenomenon may be
strain/species specific. Nevertheless, more strains needed to be tested, including environmental
strains of other Bcc and non-Bcc species. Bmul_2557 homologs might have an important role in
Burkholderia metabolic pathways. Experimental evolution studies leading to biofilm life-style
adaptation by Burkholderia cenocepacia HI2424 grown in D-galactose rich medium showed that one
of the early mutated genes was indeed a Bmul_2557 homologue. These mutations include the
deletion of amino acids 38 and 39 and the substitution of L40 by V, both located in the DNA-binding
site (Traverse et al. 2013). These authors suggest these mutations may be specifically adaptive in
biofilms by directing metabolic resources towards sessile growth.
In a second part of this work, we studied the relevance of an OmpR-like regulator from a TCS,
in Burkholderia morphotype variation. A plasmid containing the ompR gene was mobilized into
nonmucoid B. multivorans variants obtained from 4 different clinic isolates by triparental mating. In all
of the cases, the mucoid phenotype was restored, confirming that nonmucoid phenotype is due to
mutations in this gene. Whole-genome sequencing of five nonmucoid variants was performed with the
sequencing results indicating that all have mutations in the ompR gene. To complement that, a novel
strategy to knock-out the ompR gene in B. multivorans D2095 was initiated. The aim was to prepare a
nonmucoid deletion mutant to proceed studies envisaging the understanding of bce genes expression
and regulon determination. With this strategy it is possible to produce a deletion mutant without a
selection marker, which is a major advantage because allows, in a cleaner way, the study of OmpR in
phenotype variation and impact in virulence. This strategy is being performed and, until now, the main
“bottleneck” is the optimization of the Gateway PCR, when the PCRs resultants from amplification of
the upstream and downstream regions of the ompR gene are fused together, to generate the ompR
mutant allele.
In conclusion, in this work we found that the Bmul_2557 encoding LTTR acts as a positive
regulator, transcriptionally regulating gene Bmul_2558 encoding a D-lactate dehydrogenase. Although
the conversion of pyruvate into D-lactic acid by D-lactate dehydrogenase could be an extra pathway
for cells deeding with excess of carbon source, in the case of B. multivorans, it turns to be lethal since
due to medium acidification, cells lyse. Regarding this part of the work, in future, we intend to measure

Discussion _____________________________
44
the enzymatic activity of 2-ketogluconokinase an enzyme involved in the oxidative pathway, as well as
the enzymatic activity of D-lactate dehydrogenase, both in the B. multivorans ATCC 17616 and
∆Bmul_2557::dhfR, in order to verify if is this hypothesis is correct. Also, if the activity of those proteins
is altered, and knowing that Bmul_2557 is a transcription regulator, we also plan to assess transcript
levels of genes encoding the respective enzymes by RT-PCR. In the second part of this work, the
results suggest that mutations in the ompR gene are a mechanism to mucoid-to-nonmucoid switch,
and occur in several isolates irrespective of genetic background. Thus, the study of TCS and their
involvement in regulation of phase variation, and specifically of the OmpR-like RR is of major
importance to understand morphotype variation in Bcc. Therefore, more studies of this LysR-type
regulator and OmpR will enlighten regulation of metabolic and/or virulence traits in Bcc and may help
to understand the biology of this organism.

References _____________________________
45
5. References
Allenza, P., & Lessie, T. G. 1982. Pseudomonas cepacia mutants blocked in the Entner- Doudoroff Pathway. Journal Of Bacteriology,150(3), pp.1340–1347.
Allenza, P., Morrell, M. J., & Detroy, R. W. (1990). Conversion of mannose to fructose by immobilized mannose isomerase from Pseudomonas cepacia. Applied Biochemistry and Biotechnology, 24(1), pp.171-182.
Altschul, S. F., Madden, T. L., Schäffer, A. A., Zhang, J., Zhang, Z., Miller, W., & Lipman, D. J. (1997). Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Research, 25(17), pp. 3389-3402.
Aubert, D. F., Flannagan, R. S., & Valvano, M. A. (2008). A novel sensor kinase-response regulator hybrid controls biofilm formation and type VI secretion system activity in Burkholderia cenocepacia. Infection and Immunity,76(5), pp.1979-1991.
Baldwin, A., Mahenthiralingam, E., Drevinek, P., Vandamme, P., Govan, J. R., Waine, D. J., Dowson, C. G. (2007). Environmental cepacia complex isolates in human infections. Emerging Infectious Diseases,13(3), pp.458–461.
Beier, D. & Gross, R., (2006). Regulation of bacterial virulence by two-component systems. Current Opinion In Microbiology, 9(2), pp.143–52.
Bernier, S.P., Nguyen, D.T. & Sokol, P. (2008). A LysR-type transcriptional regulator in Burkholderia cenocepacia influences colony morphology and virulence. Infection and Immunity, 76(1), pp.38–47.
Bevivino, A., Tabacchioni, S., Chiarini, L., Carusi, M. V., Del Gallo, M., & Visca, P. (1994). Phenotypic comparison between rhizosphere and clinical isolates of Burkholderia cepacia. Microbiology, 140(5), pp.1069-1077.
Bunch, P. K., Mat-Jan, F., Lee, N., & Clark, D. P. (1997). The IdhA gene encoding the fermentative lactate dehydrogenase of Escherichia coli.Microbiology, 143(1), pp. 187-195.
Chua, K.L., Chan, Y.Y. & Gan, Y.H. (2003). Flagella are virulence determinants of Burkholderia pseudomallei. Infection and Immunity, 71(4), pp.1622–1629.
Chung, J. W., Altman, E., Beveridge, T. J., & Speert, D. P. (2003). Colonial morphology of Burkholderia cepacia complex genomovar III: implications in exopolysaccharide production, pilus expression, and persistence in the mouse.Infection and immunity, 71(2), pp.904-909.
Coenye, T., Vandamme, P., Govan, J. R. W., Lipuma, J., & Coenye, T. O. M. (2001). Taxonomy and identification of the Burkholderia cepacia complex. Journal of Clinical Microbiology, 39(10), pp. 3427–3436
Coenye, T. & Vandamme, P., (2003). Diversity and significance of Burkholderia species occupying diverse ecological niches. Environmental Microbiology, 5(9), pp.719–729.
Combes, D., & Monsan, P. (1983). Sucrose hydrolysis by invertase. Characterization of products and substrate inhibition. Carbohydrate research,117, pp. 215-228.

References _____________________________
46
Compant, S., Nowak, J., Coenye, T., Clément, C., & Barka, E. A. (2008). Diversity and occurrence of Burkholderia spp. in the natural environment.FEMS Microbiology Reviews, 32(4), pp.607-626.
Conway, B.A.D., Venu, V. & Speert, D.P, (2002). Biofilm formation and acyl homoserine lactone production in the Burkholderia cepacia complex. Journal of Bacteriology, 184(20), pp.5678–5685.
Conway, B. A. D., Chu, K. K., Bylund, J., Altman, E., & Speert, D. P. (2004). Production of exopolysaccharide by Burkholderia cenocepacia. Results in altered cell-surface interactions and altered bacterial clearance in mice. Journal of Infectious Diseases, 190(5), pp.957-966.
Corbett, C.R., (2003). An extracellular zinc metalloprotease gene of Burkholderia cepacia. Microbiology, 149(8), pp.2263–2271.
Deghmane, A. E., Giorgini, D., Larribe, M., Alonso, J. M., & Taha, M. K. (2002). Down‐regulation of pili and capsule of Neisseria meningitidis upon contact with epithelial cells is mediated by CrgA regulatory protein. Molecular microbiology, 43(6), pp.1555-1564.
Döring, G., Parameswaran, I.G. & Murphy, T.F., (2011). Differential adaptation of microbial pathogens to airways of patients with cystic fibrosis and chronic obstructive pulmonary disease. FEMS Microbiology Reviews, 35(1), pp.124–46.
Drevinek, P. & Mahenthiralingam, E., (2010). Burkholderia cenocepacia in cystic fibrosis : epidemiology and molecular mechanisms of virulence. Clinical Microbiology and Infection, 16(7), pp.821-830.
Eberl, L., (2006). Quorum sensing in the genus Burkholderia. International Journal of Medical Microbiology : IJMM, 296(2-3), pp.103–10.
Fazli, Mustafa, Joe J. Harrison, Michela Gambino, Michael Givskov, and Tim Tolker-Nielsen. (2015). In-Frame and Unmarked Gene Deletions in Burkholderia cenocepacia via an Allelic Exchange System Compatible with Gateway Technology. Applied and Environmental Microbiology, 81(11), pp. 3623–30.
Ferreira, A. S., Leitão, J. H., Silva, I. N., Pinheiro, P. F., Sousa, S., Ramos, C. G., & Moreira, L. M. (2010). Distribution of cepacian biosynthesis genes among environmental and clinical Burkholderia strains and role of cepacian exopolysaccharide in resistance to stress conditions. Applied and Environmental Microbiology, 76(2), pp. 441–50.
Ferreira, A. S., Silva, I. N., Oliveira, V. H., Cunha, R., & Moreira, L. M. (2011). Insights into the role of extracellular polysaccharides in Burkholderia adaptation to different environments. Frontiers in Cellular and Infection Microbiology: 16. PMC. Web. 12 Nov. 2015.
Figurski, D. H., & Helinski, D. R. (1979). Replication of an origin-containing derivative of plasmid RK2 dependent on a plasmid function provided in trans. Biochemistry, 76(4), pp.1648–1652.
Foster, J. W., Moat, A. G., Spector, M. P. (2002). Microbial Physiology, (4th ed.). New York: Wiley-Liss
Frey, Perry A. (1996). The Leloir Pathway : A Mechanistic Imperative for Three Enzymes to Change the Stereochemical Configuration of a Single Carbon in Galactose. FASEB journal : official publication of the Federation of American Societies for Experimental Biology 10(4), pp. 461–70.
Fuqua, W.C., Winans, S.C. & Greenberg, E.P., (1994). Quorum sensing in bacteria : the LuxR-LuxI family of cell density-responsive transcriptional regulators, Journal of Bacteriology, 176(2), pp.269–275.

References _____________________________
47
Gibson, R.L., Burns, J.L. & Ramsey, B.W., (2003). Pathophysiology and management of pulmonary infections in cystic fibrosis. American Journal of Respiratory and Critical Care Medicine, 168(8), pp.918–51.
Gillis, M., Van Van, T., Bardin, R., Goor, M., Hebbar, P., Willems, A., ... & Fernandez, M. P. (1995). Polyphasic taxonomy in the genus Burkholderia leading to an emended description of the genus and proposition of Burkholderia vietnamiensis sp. nov. for N2-fixing isolates from rice in Vietnam. International Journal of Systematic Bacteriology, 45(2), pp.274-289.
Govan, J.R. & Deretic, V., (1996). Microbial pathogenesis in cystic fibrosis : mucoid Pseudomonas aeruginosa and Burkholderia cepacia. , Microbiological Reviews, 60(3), pp.539–574.
Heeb, S. & Haas, D., (2001). Regulatory roles of the GacS/GacA two-component system in plant-associated and other gram-negative bacteria. Molecular Plant-Microbe Interactions : MPMI, 14(12), pp.1351–63.
Holden, M. T., Seth-Smith, H. M., Crossman, L. C., Sebaihia, M., Bentley, S. D., Cerdeño-Tárraga, A. M., ... & Parkhill, J. (2009). The genome of Burkholderia cenocepacia J2315, an epidemic pathogen of cystic fibrosis patients. Journal of Bacteriology, 191(1), pp.261-277.
Huber, B., Riedel, K., Köthe, M., Givskov, M., Molin, S., & Eberl, L. (2002). Genetic analysis of functions involved in the late stages of biofilm development in Burkholderia cepacia H111. Molecular Microbiology, 46(2), pp.411-426.
Hutchison, M.L., Poxton, I.A.N.R. & Govan, J.R.W., (1998). Burkholderia cepacia produces a hemolysin that is capable of inducing apoptosis and degranulation of mammalian phagocytes. Infection and Immunity, 66(5), pp.2033–2039.
Isles, A., Maclusky, I., Corey, M., Gold, R., Prober, C., Fleming, P., & Levison, H. (1984). Pseudomonas cepacia infection in cystic fibrosis: an emerging problem. The Journal of Pediatrics, 104(2), pp.206-210.
Kearse, M., Moir, R., Wilson, A., Stones-Havas, S., Cheung, M., Sturrock, S., ... & Drummond, A. (2012). Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics, 28(12), pp. 1647-1649.
Khodai-Kalaki, M., Aubert, D.F. & Valvano, M., (2013). Characterization of the AtsR hybrid sensor kinase phosphorelay pathway and identification of its response regulator in Burkholderia cenocepacia. The Journal of Biological Chemistry, 288(42), pp.30473–84.
Kovacikova, G., Lin, W. & Skorupski, K., (2004). Vibrio cholerae AphA uses a novel mechanism for virulence gene activation that involves interaction with the LysR-type regulator AphB at the tcpPH promoter. Molecular Microbiology, 53(1), pp.129–142.
Leitão, J. H., Sousa, S. A., Ferreira, A. S., Ramos, C. G., Silva, I. N., & Moreira, L. M. (2010). Pathogenicity, virulence factors, and strategies to fight against Burkholderia cepacia complex pathogens and related species. Applied Microbiology and Biotechnology, 87(1), pp31-40.
Leitão, J. H., Sousa, S. A., Cunha, M. V., Salgado, M. J., Melo-Cristino, J., Barreto, M. C., & Sá-Correia, I. (2008). Variation of the antimicrobial susceptibility profiles of Burkholderia cepacia complex clonal isolates obtained from chronically infected cystic fibrosis patients: a five-year survey in the major Portuguese treatment center. European Journal of Clinical Microbiology & Infectious Diseases, 27(11), pp.1101-1111.
Lessie, T. G. and P. V Phibbs. (1984). Alternative pathways of carbohydrate utilization in pseudomonads. Annual Review of Microbiology 38, pp. 359–87.

References _____________________________
48
Lewenza, S., Conway, B. & Greenberg, E.P., (1999). Quorum Sensing in Burkholderia cepacia : Identification of the LuxRI Homologs CepRI. Journal of Bacteriology, 181(3), pp.748–756.
Lipuma, J. J., Spilker, T., Gill, L. H., Campbell III, P. W., Liu, L., & Mahenthiralingam, E. (2001). Disproportionate distribution of Burkholderia cepacia complex species and transmissibility markers in cystic fibrosis. American Journal of Respiratory and Critical Care Medicine, 164(1), pp.92-96.
Lipuma, J.J., (2010). The changing microbial epidemiology in cystic fibrosis. Clinical Microbiology Reviews, 23(2), pp.299–323.
Loutet, S. A., Flannagan, R. S., Kooi, C., Sokol, P. A., & Valvano, M. A. (2006). A complete lipopolysaccharide inner core oligosaccharide is required for resistance of Burkholderia cenocepacia to antimicrobial peptides and bacterial survival in vivo. Journal of Bacteriology, 188(6), pp.2073-2080.
Loutet, S. A., & Valvano, M. A. (2010). A decade of Burkholderia cenocepacia virulence determinant research. Infection and Immunity, 78(10), pp.4088-4100.
Liu, S. (2003). Practical Implications of Lactate and Pyruvate Metabolism by Lactic Acid Bacteria in Food and Beverage Fermentations. International Journal of Food Microbiology, 83(2), pp.115–31.
Lyczak, J.B., Cannon, C.L. & Pier, G.B., (2002). Lung Infections Associated with cystic fibrosis. Clinal Microbiology Reviews.15(2), pp.194–222.
Maddocks, S.E. & Oyston, P.C.F., (2008). Structure and function of the LysR-type transcriptional regulator (LTTR) family proteins. Microbiology, 154(12), pp.3609–3623.
Mahenthiralingam, E., Baldwin, A. & Dowson, C.G., (2008). Burkholderia cepacia complex bacteria: opportunistic pathogens with important natural biology. Journal of Applied Microbiology, 104(6), pp.1539–51.
Mahenthiralingam, E., Urban, T. a & Goldberg, J.B., (2005). The multifarious, multireplicon Burkholderia cepacia complex. Nature Reviews. Microbiology, 3(2), pp.144–156.
Malott, R.J. & Sokol, P., (2007). Expression of the bviIR and cepIR quorum-sensing systems of Burkholderia vietnamiensis. Journal of Bacteriology, 189(8), pp.3006–16.
Mattoo, S. & Cherry, J.D.,(2005). Molecular pathogenesis, epidemiology, and clinical manifestations of respiratory infections due to Bordetella pertussis and other Bordetella subspecies. Clinical Microbiology Reviews, 18(2), pp.326–82.
Maxwell, E., K. Kurahashi, and H. M. Kalckar. (1962). Enzymes of the Leloir Pathway. Methods Enzymol. 1036(5), pp. 174–89.
Meyer, J. M., Van Van, T., Stintzi, A., Berge, O., & Winkelmann, G. (1995). Ornibactin production and transport properties in strains of Burkholderia vietnamiensis and Burkholderia cepacia (formerly Pseudomonas cepacia).Biometals, 8(4), pp.309-317.
Moreira, L. M., Videira, P. A., Sousa, S. A., Leitao, J. H., Cunha, M. V., & Sá-Correia, I. (2003). Identification and physical organization of the gene cluster involved in the biosynthesis of Burkholderia cepacia complex exopolysaccharide. Biochemical and Biophysical Research Communications,312(2), pp.323-333.

References _____________________________
49
Mil-Homens, D., & Fialho, A. M. (2011). Trimeric Autotransporter Adhesins in Members of the Burkholderia Cepacia Complex: A Multifunctional Family of Proteins Implicated in Virulence. Frontiers in Cellular and Infection Microbiology, 1, 13.
Mil-Homens, D. & Fialho, A.M., (2012). A BCAM0223 mutant of Burkholderia cenocepacia is deficient in hemagglutination, serum resistance, adhesion to epithelial cells and virulence. PloS one, 7(7), pp.e41747.
Nzula, S., (2002). Influence of taxonomic status on the in vitro antimicrobial susceptibility of the Burkholderia cepacia complex. Journal of Antimicrobial Chemotherapy, 50(2), pp.265–269.
O'Grady, E. P., Nguyen, D. T., Weisskopf, L., Eberl, L., & Sokol, P. A. (2011). The Burkholderia cenocepacia LysR-type transcriptional regulator ShvR influences expression of quorum-sensing, protease, type II secretion, and afc genes. Journal of Bacteriology, 193(1), pp.163-176.
O’Sullivan, L. & Mahenthiralingam, E., (2005). Biotechnological potential within the genus Burkholderia. Letters in Applied Microbiology, 41(1), pp.8–11.
Peeters, C., Zlosnik, J. E., Spilker, T., Hird, T. J., LiPuma, J. J., & Vandamme, P. (2013). Burkholderia pseudomultivorans sp. nov., a novel Burkholderia cepacia complex species from human respiratory samples and the rhizosphere. Systematic and Applied Microbiology, 36(7), pp.483-489.
Porrúa, Odil, Manuel García-Jaramillo, Eduardo Santero, and Fernando Govantes. (2007). The LysR-Type Regulator AtzR Binding Site: DNA Sequences Involved in Activation, Repression and Cyanuric Acid-Dependent Repositioning. Molecular microbiology 66(2), pp. 410–27.
Richau, J. A., Leitão, J. H., Correia, M., Lito, L., Salgado, M. J., Barreto, C., Sá-Correia, I. (2000). Molecular Typing and exopolysaccharide biosynthesis of Burkholderia cepacia Isolates from a portuguese cystic fibrosis center. Journal of Clinical Microbiology, 38(4), pp.1651–1655
Rosales‐Reyes, R., Skeldon, A. M., Aubert, D. F., & Valvano, M. A. (2012). The Type VI secretion system of Burkholderia cenocepacia affects multiple Rho family GTPases disrupting the actin cytoskeleton and the assembly of NADPH oxidase complex in macrophages. Cellular Microbiology, 14(2), pp.255-273.
Sage, A., Linker, A., Evans, L. R., & Lessie, T. G. (1990). Hexose phosphate metabolism and exopolysaccharide formation inPseudomonas cepacia. Current Microbiology, 20(3), pp.191-198.
Sajjan, U.S., Sun, L.I. & Goldstein, R., (1995). Cable ( Cbl ) type II pili of cystic fibrosis-associated Burkholderia (Pseudomonas ) cepacia : Nucleotide sequence of the cblA major subunit pilin gene and novel morphology of the assembled appendage fibers. Journal of Bacteriology, 177(4), pp.1030–1038.
Sambrook, J., & Russell, D. W. (2001). Molecular cloning: a laboratory manual CSHL Press.
Schell, M. A. (1993). Molecular biology of the LysR family of transcriptional regutlators. Annual Review of Microbiology. 47, pp.597–626.
Silva, I. N., Ferreira, A. S., Becker, J. D., Zlosnik, J. E., Speert, D. P., He, J., ... & Moreira, L. M. (2011). Mucoid morphotype variation of Burkholderia multivorans during chronic cystic fibrosis lung infection is correlated with changes in metabolism, motility, biofilm formation and virulence. Microbiology,157(11), pp.3124-3137.
Silva, I.N. (2012). Comparative genomics and transcriptomics to study mucoid morphotype variation in Burkholderia cepacia complex clinical isolates. phD Thesis, Instituto Superior Técnico, UTL, Lisbon, 2012.

References _____________________________
50
Silva, I. N., Tavares, A. C., Ferreira, A. S., & Moreira, L. M. (2013). Stress conditions triggering mucoid morphotype variation in Burkholderia species and effect on virulence in Galleria mellonella and biofilm formation in vitro. PLoS ONE, 8(12), pp. 82522.
Silva, I. N., Santos, P. M., & Moreira, L. M. (2015). Draft genome sequences of two Burkholderia multivorans sequential isolates from a chronic lung infection of a cystic fibrosis patient. Genome announcements, 3(1), e01531-14
Sousa, S., Ramos, C.G. & Leitão, J.H., (2011). Burkholderia cepacia complex: Emerging multihost pathogens equipped with a wide range of virulence factors and determinants. International Journal of Microbiology, 2011.
Stock, A.M., Robinson, V.L. & Goudreau, P.N., (2000). Two component signal transduction. Annual Review of Biochemistry, 69, pp.183–215.
Subramoni, S., Nguyen, D.T. & Sokol, P., (2011). Burkholderia cenocepacia ShvR-regulated genes that influence colony morphology, biofilm formation, and virulence. Infection and Immunity, 79(8), pp.2984–97.
Tavares, A. F. C. (2012). Biological relevance of mucoid vs. nonmucoid morphotype variation by Burkholderia cepacia complex. MSc Thesis, Instituto Superior Técnico, UTL, Lisbon, 2012.
Thomas, T. D., Turner, K. W., & Crow, V. L. (1980). Galactose fermentation by Streptococcus lactis and Streptococcus cremoris: pathways, products, and regulation. Journal of Bacteriology, 144(2), pp. 672-682.
Tomich, M., Herfst, C. A., Golden, J. W., & Mohr, C. D. (2002). Role of flagella in host cell invasion by Burkholderia cepacia. Infection and Immunity, 70(4), pp.1799-1806.
Tomich, M. & Mohr, C.D., (2003). Adherence and autoaggregation phenotypes of a Burkholderia cenocepacia cable pilus mutant. FEMS Microbiology Letters, 228(2), pp.287–297.
Thompson, J. D., Higgins, D. G., & Gibson, T. J. (1994). CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice.Nucleic acids research, 22(22), pp. 4673-4680.
Traverse, C. C., Mayo-Smith, L. M., Poltak, S. R., & Cooper, V. S. (2013). Tangled bank of experimentally evolved Burkholderia biofilms reflects selection during chronic infections. Proceedings of the National Academy of Sciences,110(3), E250-E259
De Torrontegui, G., Diaz, R., & Canovas, J. L. (1976). The uptake of 2-ketogluconate by Pseudomonas putida. Archives of microbiology, 110(1), pp. 43-48.
Urban, T. A., Griffith, A., Torok, A. M., Smolkin, M. E., Burns, J. L., & Goldberg, J. B. (2004). Contribution of Burkholderia cenocepacia flagella to infectivity and inflammation. Infection and Immunity, 72(9), pp.5126-5134.
Van Acker, H., Sass, A., Bazzini, S., De Roy, K., Udine, C., Messiaen, T., ... & Coenye, T. (2013). Biofilm-grown Burkholderia cepacia complex cells survive antibiotic treatment by avoiding production of reactive oxygen species. PLoS One, 8(3), e58943.
Van Den Broek, D., Chin-A-Woeng, T. F., Bloemberg, G. V., & Lugtenberg, B. J. (2005). Molecular nature of spontaneous modifications in gacS which cause colony phase variation in Pseudomonas sp. strain PCL1171. Journal of Bacteriology, 187(2), pp.593-600.
Vandamme, P., Henry, D., Coenye, T., Nzula, S., Vancanneyt, M., LiPuma, J. J., ... & Mahenthiralingam, E. (2002). Burkholderia anthina sp. nov. and Burkholderia pyrrocinia, two

References _____________________________
51
additional Burkholderia cepacia complex bacteria, may confound results of new molecular diagnostic tools. FEMS Immunology & Medical Microbiology, 33(2), pp.143-149.
Vandamme, P., Mahenthiralingam, E., Holmes, B., Coenye, T., Hoste, B., De Vos, P., ... & Speert, D. P. (2000). Identification and Population Structure of Burkholderia stabilis sp. nov.(formerly Burkholderia cepacia Genomovar IV).Journal of Clinical Microbiology, 38(3), pp.1042-1047.
Vandamme, P., Holmes, B., Vancanneyt, M., Coenye, T., Hoste, B., Coopman, R., ... & Govan, J. R. W. (1997). Occurrence of Multiple Genomovars of Burkholderia cepacia in Cystic Fibrosis Patients and Proposal of Burkholderia multivorans sp. nov. International Journal of Systematic Bacteriology, 47(4), pp.1188-1200.
Vandamme, P. & Dawyndt, P., (2011). Classification and identification of the Burkholderia cepacia complex: Past, present and future. Systematic and Applied Microbiology, 34(2), pp.87–95.
Vanlaere, E., LiPuma, J. J., Baldwin, A., Henry, D., De Brandt, E., Mahenthiralingam, E., ... & Vandamme, P. (2008). Burkholderia latens sp. nov., Burkholderia diffusa sp. nov., Burkholderia arboris sp. nov., Burkholderia seminalis sp. nov. and Burkholderia metallica sp. nov., novel species within the Burkholderia cepacia complex. International Journal of Systematic and Evolutionary Microbiology, 58(7), pp.1580-1590.
Vanlaere, E., Baldwin, A., Gevers, D., Henry, D., De Brandt, E., LiPuma, J. J., ... & Vandamme, P. (2009). Taxon K, a complex within the Burkholderia cepacia complex, comprises at least two novel species, Burkholderia contaminans sp. nov. and Burkholderia lata sp. nov. International Journal of Systematic and Evolutionary Microbiology, 59(1), pp.102-111.
Vermis, K., (2004). Proposal to accommodate Burkholderia cepacia genomovar VI as Burkholderia dolosa sp. nov. International Journal of Systematic and Evolutionary Microbiology, 54(3), pp.689–691.
Viallard, V., Poirier, I., Cournoyer, B., Haurat, J., Wiebkin, S., Ophel-Keller, K., & Balandreau, J. (1998). Burkholderia graminis sp. nov., a rhizospheric Burkholderia species, and reassessment of [Pseudomonas] phenazinium,[Pseudomonas] pyrrocinia and [Pseudomonas] glathei as Burkholderia.International Journal of Systematic Bacteriology, 48(2), pp.549-563.
Vinion-Dubiel, A.D. & Goldberg, J.B., (2003). Lipopolysaccharide of Burkholderia cepacia complex. Journal of Endotoxin Research, 9(4), pp.201–13.
Visser, M. B., Majumdar, S., Hani, E., & Sokol, P. A. (2004). Importance of the ornibactin and pyochelin siderophore transport systems in Burkholderia cenocepacia lung infections. Infection and Immunity, 72(5), pp.2850-2857.
Weiss, A.A.N.N. & Melton, A.R., (1993). Bordetella pertussis. Infection and Immunity, 61(3), pp.807–815.
Yabuuchi, E., Kawamura, Y., Ezaki, T., Ikedo, M., Dejsirilert, S., Fujiwara, N., ... & Kobayashi, K. (2000). Burkholderia uboniae Sp. Nov., l‐Arabinose‐Assimilating but Different from Burkholderia thailandensis and Burkholderia vietnamiensis. Microbiology and Immunology, 44(4), pp.307-317.
Yabuuchi, E., Kosako, Y., Oyaizu, H., Yano, I., Hotta, H., Hashimoto, Y., ... & Arakawa, M. (1992). Proposal of Burkholderia gen. nov. and transfer of seven species of the genus Pseudomonas homology group II to the new genus, with the type species Burkholderia cepacia (Palleroni and Holmes 1981) comb. nov.Microbiology and Immunology, 36(12), pp.1251-1275.
Yang, S., Rice, K. C., Brown, R. J., Patton, T. G., Liou, L. E., Park, Y. H., & Bayles, K. W. (2005). A LysR-Type regulator , CidR , is required for induction of the Staphylococcus aureus cidABC operon. Journal of Bacteriology, 187(17), pp.5893–5900.

References _____________________________
52
Zlosnik, J. E., Hird, T. J., Fraenkel, M. C., Moreira, L. M., Henry, D. A., & Speert, D. P. (2008). Differential mucoid exopolysaccharide production by members of the Burkholderia cepacia complex. Journal of Clinical Microbiology,46(4), pp.1470-1473.
Zlosnik, J. E., Costa, P. S., Brant, R., Mori, P. Y., Hird, T. J., Fraenkel, M. C., ... & Speert, D. P. (2011). Mucoid and nonmucoid Burkholderia cepacia complex bacteria in cystic fibrosis infections. American Journal of Respiratory and Critical Care Medicine, 183(1), pp.67-72.
(http://www.bacterio.net/burkholderia.html), consulted in November 24, 2015
(http://www.cftrscience.com/epidemiology), consulted in October 15, 2015
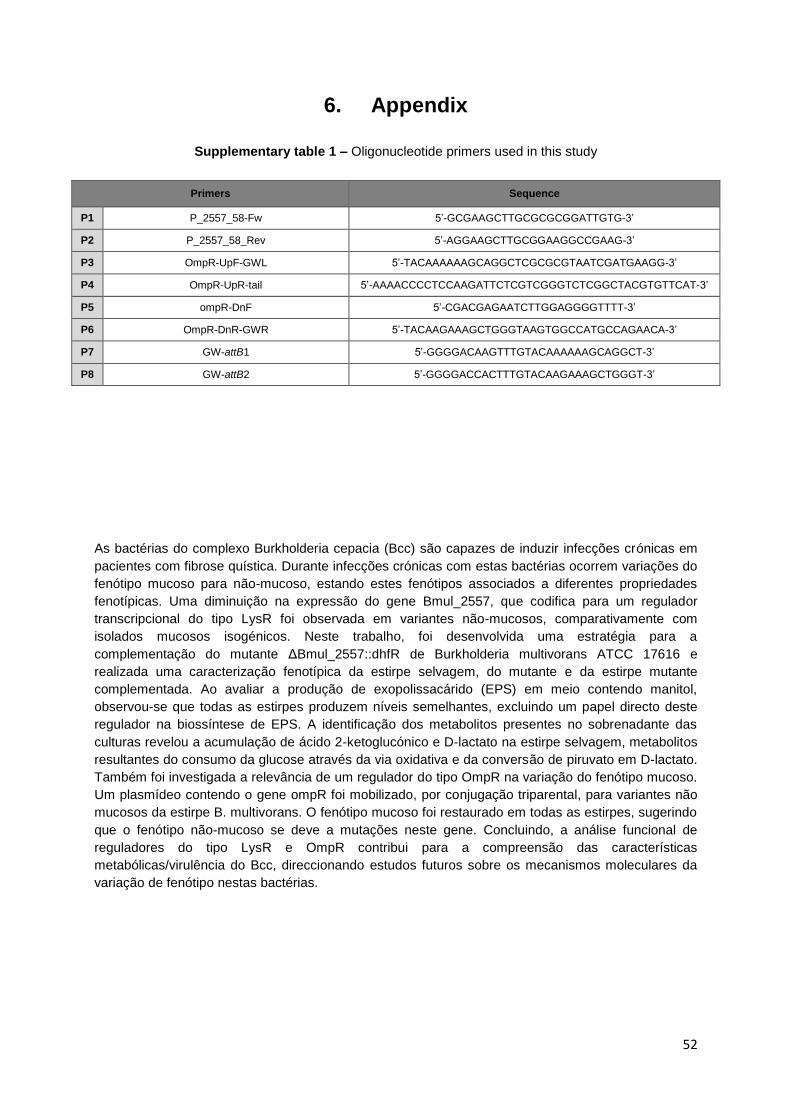
52
6. Appendix
Supplementary table 1 – Oligonucleotide primers used in this study
As bactérias do complexo Burkholderia cepacia (Bcc) são capazes de induzir infecções crónicas em
pacientes com fibrose quística. Durante infecções crónicas com estas bactérias ocorrem variações do
fenótipo mucoso para não-mucoso, estando estes fenótipos associados a diferentes propriedades
fenotípicas. Uma diminuição na expressão do gene Bmul_2557, que codifica para um regulador
transcripcional do tipo LysR foi observada em variantes não-mucosos, comparativamente com
isolados mucosos isogénicos. Neste trabalho, foi desenvolvida uma estratégia para a
complementação do mutante ΔBmul_2557::dhfR de Burkholderia multivorans ATCC 17616 e
realizada uma caracterização fenotípica da estirpe selvagem, do mutante e da estirpe mutante
complementada. Ao avaliar a produção de exopolissacárido (EPS) em meio contendo manitol,
observou-se que todas as estirpes produzem níveis semelhantes, excluindo um papel directo deste
regulador na biossíntese de EPS. A identificação dos metabolitos presentes no sobrenadante das
culturas revelou a acumulação de ácido 2-ketoglucónico e D-lactato na estirpe selvagem, metabolitos
resultantes do consumo da glucose através da via oxidativa e da conversão de piruvato em D-lactato.
Também foi investigada a relevância de um regulador do tipo OmpR na variação do fenótipo mucoso.
Um plasmídeo contendo o gene ompR foi mobilizado, por conjugação triparental, para variantes não
mucosos da estirpe B. multivorans. O fenótipo mucoso foi restaurado em todas as estirpes, sugerindo
que o fenótipo não-mucoso se deve a mutações neste gene. Concluindo, a análise funcional de
reguladores do tipo LysR e OmpR contribui para a compreensão das características
metabólicas/virulência do Bcc, direccionando estudos futuros sobre os mecanismos moleculares da
variação de fenótipo nestas bactérias.
Primers Sequence
P1 P_2557_58-Fw 5’-GCGAAGCTTGCGCGCGGATTGTG-3’
P2 P_2557_58_Rev 5’-AGGAAGCTTGCGGAAGGCCGAAG-3’
P3 OmpR-UpF-GWL 5’-TACAAAAAAGCAGGCTCGCGCGTAATCGATGAAGG-3’
P4 OmpR-UpR-tail 5’-AAAACCCCTCCAAGATTCTCGTCGGGTCTCGGCTACGTGTTCAT-3’
P5 ompR-DnF 5’-CGACGAGAATCTTGGAGGGGTTTT-3’
P6 OmpR-DnR-GWR 5’-TACAAGAAAGCTGGGTAAGTGGCCATGCCAGAACA-3’
P7 GW-attB1 5’-GGGGACAAGTTTGTACAAAAAAGCAGGCT-3’
P8 GW-attB2 5’-GGGGACCACTTTGTACAAGAAAGCTGGGT-3’







