
TOXICOLOGY AND APPLIED PHARMACOLOGY 97,480-488 (1989)
Genotoxicity of N-Nitrosochlordiazepoxide in Cultured Mammalian Cells’
GIOVANNIBRAMBILLA,*"LUIGIROBBIANO,*ANTONIETTAMARTELLI,*ENRICACAJELLI,* ALESSANDRAALLAVENA.*ANDMAUROMAZZEI**
*Institutes ofPharmacology and OJ **Pharmaceutical Sciences, University ofGenoa, I- 16132 Genoa. Italy
Received May I-7, 1988; accepted October 3. 1988
Genotoxicity of iliNitrosochlordiazepoxide in Cultured Mammalian Cells. BRAMBILLA. G..
ROBBIANO, L., MARTELLI. A., CAJELLI, E., ALLAVENA, A., AND MAZZEI. M. (I 989). Toxicol. Appi. Pharmacol. 97,480-488. Chlordiazepoxide. a benzodiazepine derivative commonly used
for the treatment of anxiety. was found to react with sodium nitrite in HCI aqueous solution yielding, at pH ranging from 0.5 to 5. N-nitrosochlordiazepoxide (NO-CDE). In the absence of
a metabolic activation system. a dose-dependent frequency of DNA single-strand breaks was revealed by the alkaline elution technique in V79 cellsexposed to subtoxic NO-CDE concentra- tions ranging from 33 to 330 PM. DNA lesions were only partially repaired within 48 hr. and
their promutagenic character was demonstrated by the induction of 6-thioguanine resistance in the same cells. The genotoxicity of NO-CDE was confirmed by results obtained in metabolically
competent primary cultures of both rat and human hepatocytes. which displayed similar dose- related amounts of DNA fragmentation and of DNA repair synthesis after treatment with con- centrations ranging from 33 to 1000 PM. In conditions similar to those which might occur in
the stomach of a patient taking chlordiazepoxide the concentration of NO-CDE in the reaction mixture (50 FM) was of the same order as the concentrations found to induce a genotoxic effect in cultured mammalian cells. 2’ 1989 Academic press. IIIC
The possibility that carcinogenic N-nitroso compounds are formed in vim from the reac- tion of drugs containing amino groups with nitrite present in the gastric environment has been demonstrated in a large number of ani- mal experiments (Mirvish, 1975: Lijinsky and Taylor. 1977; IARC, 1980a,b). Such in- teraction should be favored by the acidity of human gastric juice. A series of data recently summarized by Bartsch and Montesano (1984) unequivocally gives proof that N-ni-
’ This work was supported by Consiglio Nazionale delle Ricerche Special Projects “Oncology” (Contract 86.00326.44), “Preventive and Rehabilitative Medicine”
(Contract 86.01877.56), and by funds of the Public In- struction Ministry (Italy).
’ To whom correspondence should be addressed at: Is-
tituto di Farmacologia dell’Universiti, Viale Benedetto XV. 2, I- 16 I32-Cienova, Italia.
troso compounds are generated in the human body, even after ingestion of levels of precur- sors considered to be normal, and provides evidence that exposure to nitrosamines will lead to an increased probability of cancer in humans, Consequently, the need of verifying the risk to human health arising from the in- take of nitrosatable drugs has become evident in recent years (IARC, 1980a: Brambilla, 1985).
The following experimental findings prompted us to further investigate the forma- tion and the genotoxicity of the nitrosation products of chlordiazepoxide, a benzodiaze- pine derivative used for the treatment of anx- iety that is widely prescribed (Blackwell, 1973). Walser cf al. (1974) first reported that treatment of chlordiazepoxide with sodium nitrite yields in acetic acid a nitrosoamidine, i.e.. 2-N-nitrosochlordiazepoxide (NO-CDE),
0041-008X/89 $3.00 Copyright 8 1989 by Academic Press, Inc. All rights of reproduction in any form resewed.
480

N-NITROSOCHLORDIAZEPOXIDE GENOTOXICITY IN MAMMALIAN CELLS 481
and in hydrochloric acid a dihydroquinazo- line. Induction by NO-CDE of mitotic gene conversion in S. cerevisiae was observed in a host-mediated assay by Krafft and Onken (1976). Preda et al. (1976) demonstrated the presence of the N-nitroso derivative in the stomach and in the small bowel of rats fed with chlordiazepoxide and nitrite, and Lijin- sky and Taylor ( 1977) detected in similar ex- perimental conditions a marginally signifi- cant increase in the incidence of nervous sys- tem tumors. Induction of mutation was observed by Andrews et al. (1980) in five S. typhimurium strains after exposure to the chlordiazepoxide nitrosation mixture. The mutagenic effect was more evident by the ab- sence rather than by the presence of a meta- bolic activation system, as subsequently re- ported by Takeda and Kanaya (198 1). Fi- nally, DNA fragmentation was detected (Brambilla et al., 1985) in CHO cells after ex- posure to an -0.2 mh4 concentration of chlordiazepoxide nitrosation products.
Recently Mazzei et al. (unpublished data) found that the reaction of chlordiazepoxide with nitrite yields NO-CDE, not only when performed in acetic acid as reported by Walser et al. ( 1974) but also when carried out in aqueous HCl solution. The measurement by HPLC analysis of NO-CDE formation at pH ranging from 0.5 to 5.0 revealed that the yield reaches its maximum at pH 3.5. This yield was approx 55% of the theoretical one when 40 mg of chlordiazepoxide hydrochlo- ride was reacted with 20 mg of NaNOz in 10 ml of HCl (pH 3.5) at 37°C for 1 hr in the dark.
The present work was undertaken to evalu- ate the genotoxicity of NO-CDE in some short-term in vitro assays on mammalian cells.
METHODS
Chemicals. NO-CDE was synthesized by Mazzei as de- scribed by Walser et al. (1974). In brief, over a period of 30 min NaNOz (0.26 mol) was added in small portions to a stirred solution of chlordiazepoxide (0.20 mol) in
400 ml of glacial acetic acid, and the reaction mixture was left at room temperature with stirring for another 30 min. The precipitate was recrystallized from ether/ hexane. NO-CDE (98-99% pure: 75% yield) was the only product obtained (melting point 158-160°C dec.). Kept at -20°C as a solid, NO-CDE was found to be chemically stable for 6 months. In aqueous solution, under condi- tions corresponding to those of the experiments carried out to evaluate its biological activity, NO-CDE was found to be stable for 24 hr, as revealed by HPLC analy- sis. N-Nitrosodimethylamine (DMN) and tetraethylam- monium hydroxide were purchased from E. Merck (Darmstadt, FRG): N-nitroso-N-methylurea (NMU) from Serva Feinbiochemica (Heidelberg. FRG); N-lauro- ylsarcosine sodium salt (Sarkosyl), 6-thioguanine. and collagenase type I from Sigma Chemical Co. (St. Louis. MO): a-modified minimum essential medium (a- MEM). Williams’ medium E (WME) and fetal calf serum from Flow Laboratories, Milan, Italy; [meth$3H]thymi- dine (sp act 23-25 Ci/mmol) from the Radiochemical Centre (Amersham, UK). All other chemicals, reagent grade, were obtained from E. Merck (Darmstadt, FRG).
Cell culture and treatment. V79 cells (NIH. Coriell In- stitute for Medical Research, Camden. NJ), an estab- lished line from Chinese hamster lung, were cultured in a-MEM supplemented with 5% fetal calf serum. Di- methyl sulfoxide was used as a solvent for NO-CDE and was added at the same concentration in control cultures.
For the determination of cytotoxicity. 6 X lo5 cells were inoculated in 25cm’ plastic flaks and after 24 hr were exposed to the test compound for 60 min in serum- free medium. Cell viability was measured immediately after exposure either by trypan blue dye exclusion or by cloning efficiency. In the trypan blue assay, at least 1000 cells/sample were examined. In the colony formation as- say, cells were trypsinized at the end of treatment and inoculated in plastic dishes (200 cells/dish); after incuba- tion for 7 days, colonies were fixed, stained. and counted.
For the assay of DNA-damaging activity, 24 hr after seeding V79 cells were labeled with 0.1 &i/ml of [meth- yl-3H]thymidine for 24 hr. Cells were then exposed to the test compound for I hr in nonradioactive serum-free medium and assayed for DNA fragmentation either im- mediately or after incubation in complete medium at 37°C for 24 and 48 hr.
Rat hepatocytes were isolated from Sprague-Dawley male albino rats (200-250 g) by collagenase perfusion, as described by Williams (1977). Human hepatocyte sus- pensions were prepared from fragments of human liver discarded during the course of prescribed surgery essen- tially according to Strom el al. (1982). Case 1 was a 60- year-old male who underwent surgery for hepatic metas- tases of a colon carcinoma. Case 2 was a 57-year-old male who underwent surgery for an echinococcus hydatid cyst. The proportion of viable cells after perfusion, as measured by trypan blue exclusion, was 77% in Case 1 and 75% in Case 2.

482 BRAMBILLA ET AL.
Isolated rat or human hepatocytes were suspended in
WME supplemented with 10% fetal calf serum and gen- tamycin (50 pg/ml), and aliquots of these suspensions were plated in plastic dishes as follows: 2 X IO6 cells in
60-mm uncoated dishes for DNA fragmentation assay,
and I X IO6 cells in 35-mm dishes coated with rat tail collagen for determination of cell viability and the un-
scheduled DNA synthesis (UDS). After attachment for 3 hr. cells were incubated for 20 hr with the test compound in serum-free medium. DMN was used as the positive
control to verify the capability of hepatocytes to activate
procarcinogens. DNA fragmentation and UDS were measured immediately after exposure to the test com-
pound. The choice for the hepatocytes of a 24-hr expo- sure, instead of 1 hr as for V79 cells, was dictated by the
opportunity to adhere to the routinely employed proto- col developed by Williams et a/. (1982) for the hepatocyte
primary culture/DNA repair assay. and thus allow a meaningful comparison with the amounts of unsched- uled DNA synthesis elicited by other N-nitroso com-
pounds. It is obvious that this choice. while making it
possible to obtain some information on the influence of the length of treatment, at the same time. makes a com-
parison of the results provided by the two cell types difficult.
Determirlation qf DNkdamaging activity. In both
V79 cells and rat or human hepatocytes the presence of DNA fragmentation (single-strand breaks. and/or alkali-
labile sites) was evaluated by the alkaline elution tech- nique (Kohn et al., 1976). as previously reported (Bram-
billa e/ al.. 1978). The cell suspension (- IO6 cells) was deposited onto a Millipore filter (mixed esters of cellu-
lose: 25-mm diameter, 5-hrn pore size), washed with cold Merchant’s solution (NaCI 0.14 M, KC1 2.7 mM.
KHZP04 1.47 mM, Na2HP0, 8.1 mM, NazEDTA 0.53 mM; pH 7.6). and lysed at room temperature (20-22°C)
with 0.2% sodium lauroylsarcosinate (Sarkosyl). 2 M NaCl, and 20 mM NazEDTA (pH 10). Single-stranded
DNA was eluted from the filter in the dark, at a pump speed ofO.13 ml/mitt. with 13 ml of60 mM tetraethylam-
monium hydroxyde and 20 mM NazEDTA (pH 12.3). Eluted fractions werecollectedat IO-min intervals. In the
experiments performed with V79 cells. the DNA content ofthe IO eluted fractions and that remaining on the filter
was dosed radiometrically by liquid scintillation count- ing. In the experiments performed with rat or human he-
patocytes the DNA content was determined by a pre- viously described microfluorimetric procedure (Bram- billa ef al.. 1978). Data are expressed as the percentage of
DNA eluted from the filter which gives direct informa- tion about the experimental results obtained. The aver-
age elution rate over controls (k; ~ &) is also reported: K, is the elution rate of DNA from treated cells and & the elution rate from corresponding controls. K values
were calculated from the equation K = (-In FR)/T: where FR is the fraction of DNA retained on the filter and I’is the eluting volume (I 3 ml). As a first approxima-
tion K is directly proportional to the frequency of DNA
breaks (Kohn et al., 1976). DNA repair te,st. The procedure used to evaluate the
induction of DNA repair synthesis was essentially that
described by Williams (1977). Rat or human hepatocyte primary cultures were simultaneously exposed to NO-
CDE and [meth$3H]thymidine (10 &i/ml). After 20 hr incubation the cultures were washed twice with 0.9% NaCl and treated with 1% sodium citrate solution for 10
min. The cells were fixed in acetic acid:ethanol (1:3) for three IO-min changes. A section of the air-dried culture
dish bottom was cut out, glued to a microscope slide. dipped in Kodak NTB-2 emulsion, and exposed for 7 days at 4°C. Autoradiographs were developed and stained with May-Gruenwald-Giemsa. Autoradio-
graphic nuclear grains were counted in 50 consecutive cells of each slide. The highest cytoplasmic background
count for each cell was subtracted from the nuclear count to obtain the net nuclear grains due to DNA repair syn-
thesis. The data are expressed as the means of the 100 net nuclear counts obtained from two autoradiographs f SD. When the increase in the average number of net
nuclear grains exceeded five as compared with the corre- sponding control. DNA repair induction was estimated
to be positive. Determination c?f’mutagenic artivii~: The quantitative
assay for induced frequency of 6-thioguanine-resistant (TG’) cells was a modification (Cajelli et al.. 1987) ofthat described by Myhr and Di Paolo (1978). The duration of
exposure to the test chemical was 1 hr. At the end of the h-day period of mutant expression, the fraction of TG’ cells in the population was determined from IO dishes
( 100 mm) by counting mutant colonies 6-7 days after seeding 3.5 X IO5 cells per dish. For selection of mutants,
h-thioguanine was added 2 hr after seeding to give a final concentration of 10 pg/ml.
RESULTS
Because of the low solubility, the maximal concentration of NO-CDE which could be tested was 1 mM. Data of Table 1 show that a dose-dependent reduction of V79 cells plat- ing efficiency was produced by 1 -hr exposure to concentrations ranging from 10 to 330 PM. In contrast, when measured by trypan blue exclusion, cell survival was practically un- affected at the same concentrations and only minimally decreased at 1 mM. Taking into account that with plating efficiency what is measured is the effect on the capacity of cell
N-NITROSOCHLORDIAZEPOXIDE GENOTOXICITY IN MAMMALIAN CELLS 483
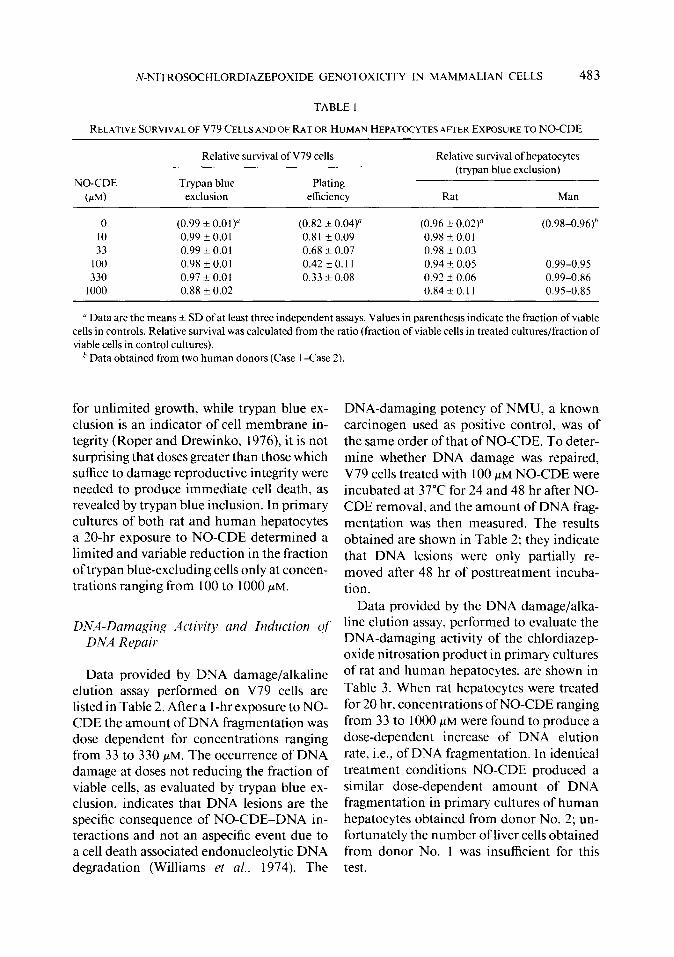
TABLE 1
RELATIVE SURVIVAL OF V79 CELLS AND OF RAT OR HUMAN HEPATOCYTES AFTER EXPOSURE TO NO-CDE
NO-CDE (N)
Relative survival of V79 cells
Trypan blue Plating exclusion efficiency
Relative survival of hepatocytes (trypan blue exclusion)
Rat Man
0 (0.99 f 0.01) (0.82 * 0.04) (0.96 k 0.02)” (0.98-0.96)h 10 0.99 -e 0.0 1 0.81 F 0.09 0.98 f 0.01 33 0.99 + 0.0 1 0.68 -+ 0.07 0.98 k 0.03
100 0.98 f 0.01 0.42t0.11 0.94 k 0.05 0.99-0.95 330 0.97 Ik 0.01 0.33 + 0.08 0.92 +- 0.06 0.99-0.86
1000 0.88 + 0.02 0.84 zk 0.11 0.95-0.85
’ Data are the means f SD of at least three independent assays. Values in parenthesis indicate the fraction of viable cells in controls. Relative survival was calculated from the ratio (fraction of viable cells in treated cultures/fraction of viable cells in control cultures).
’ Data obtained from two human donors (Case 1 -Case 2).
for unlimited growth, while trypan blue ex- DNA-damaging potency of NMU, a known clusion is an indicator of cell membrane in- carcinogen used as positive control, was of tegrity (Roper and Drewinko, 1976) it is not the same order of that of NO-CDE. To deter- surprising that doses greater than those which mine whether DNA damage was repaired, suffice to damage reproductive integrity were V79 cells treated with 100 PM NO-CDE were needed to produce immediate cell death, as incubated at 37°C for 24 and 48 hr after NO- revealed by trypan blue inclusion. In primary CDE removal, and the amount of DNA frag- cultures of both rat and human hepatocytes mentation was then measured. The results a 20-hr exposure to NO-CDE determined a obtained are shown in Table 2; they indicate limited and variable reduction in the fraction that DNA lesions were only partially re- of trypan blue-excluding cells only at concen- moved after 48 hr of posttreatment incuba- trations ranging from 100 to 1000 PM. tion.
DNA-Damaging Activity and Induction Qf DNA Repair
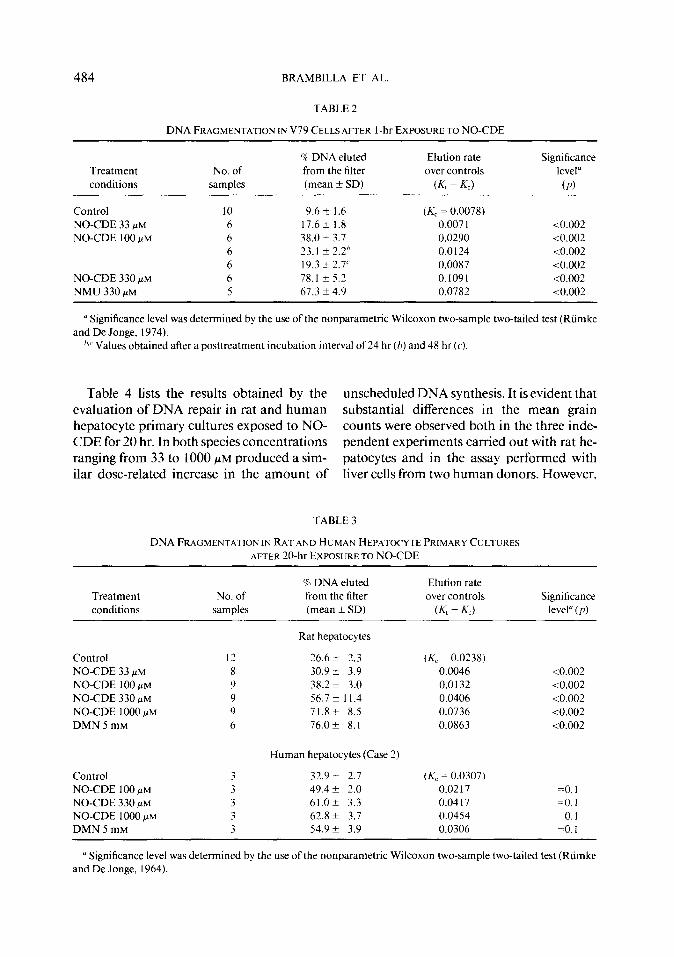
Data provided by DNA damage/alkaline elution assay performed on V79 cells are listed in Table 2. After a 1 -hr exposure to NO- CDE the amount of DNA fragmentation was dose dependent for concentrations ranging from 33 to 330 PM. The occurrence of DNA damage at doses not reducing the fraction of viable cells, as evaluated by trypan blue ex- clusion, indicates that DNA lesions are the specific consequence of NO-CDE-DNA in- teractions and not an aspecific event due to a cell death associated endonucleolytic DNA degradation (Williams ef al., 1974). The
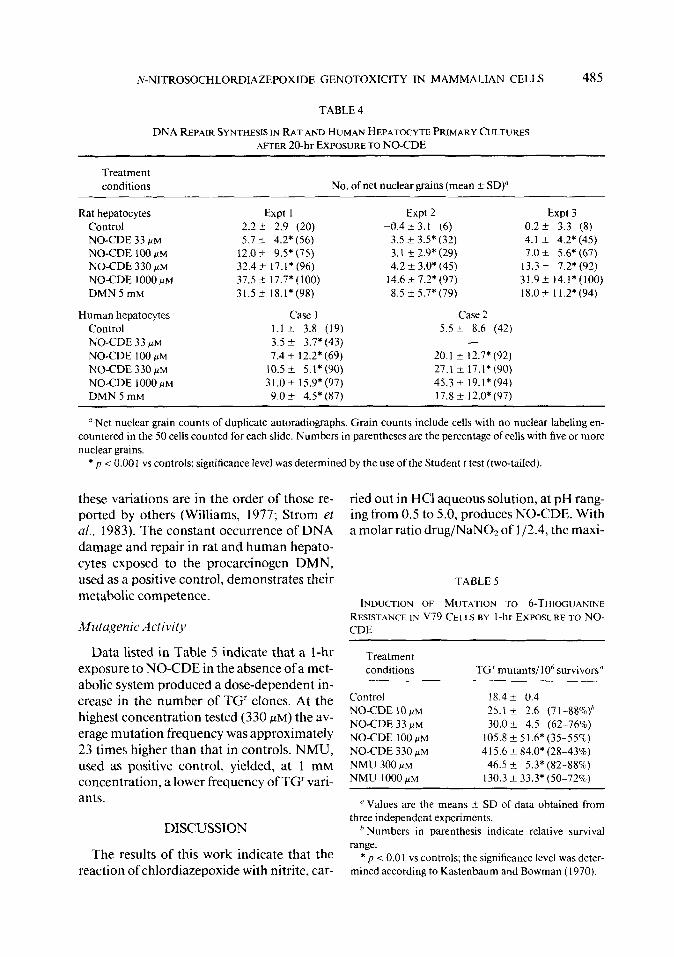
Data provided by the DNA damage/alka- line elution assay, performed to evaluate the DNA-damaging activity of the chlordiazep- oxide nitrosation product in primary cultures of rat and human hepatocytes. are shown in Table 3. When rat hepatocytes were treated for 20 hr. concentrations of NO-CDE ranging from 33 to 1000 PM were found to produce a dose-dependent increase of DNA elution rate, i.e., of DNA fragmentation. In identical treatment conditions NO-CDE produced a similar dose-dependent amount of DNA fragmentation in primary cultures of human hepatocytes obtained from donor No. 2: un- fortunately the number of liver cells obtained from donor No. 1 was insufficient for this test.
484 BRAMBILLA ET AL.
TABLE 2
DNA FRAGMENTATION IN V79 CELLS AFTER I-hr EXPOSURE TO NO-CDE
Treatment conditions
No. of samples
% DNA eluted
from the filter (mean + SD)
El&on rate
over controls
(Kt - K-c)
Significance
level”
(P)
Control 10 9.6 k I .6 (A; = 0.0078) NO-CDE 33 pM 6 17.6 k 1.8 0.007 1 to.002 NO-CDE 100 PM 6 38.02 3.1 0.0290 <0.002
6 23.1 f 2.2" 0.0124 -co.002 6 19.3 k 2.7' 0.0087 <0.002
NO-CDE 330 PM 6 78.1 + 5.2 0.1091 -co.002 NMU 330 PM 5 67.3 ?I 4.9 0.0782 <0.002
a Significance level was determined by the use of the nonparametric Wilcoxon two-sample two-tailed test (Riimke and De Jonge, 1974).
‘.’ Values obtained after a posttreatment incubation interval of 24 hr (h) and 48 hr (c‘).
Table 4 lists the results obtained by the unscheduled DNA synthesis. It is evident that evaluation of DNA repair in rat and human substantial differences in the mean grain hepatocyte primary cultures exposed to NO- counts were observed both in the three inde- CDE for 20 hr. In both species concentrations pendent experiments carried out with rat he- ranging from 33 to 1000 PM produced a sim- patocytes and in the assay performed with ilar dose-related increase in the amount of liver cells from two human donors. However,
TABLE 3
DNA FRAGMENTATION IN RAT AND HUMAN HEPAT~CYTE PRIMARY CULTURES
AFTER IO-hr EXPOSURE TO NO-CDE
Treatment conditions
No. of samples
‘3% DNA eluted
from the filter (mean *SD)
Elution rate
over controls
K - Kc)
Significance level” ( p)
Control NO-CDE 33 yM
NO-CDE 100 FM NO-CDE 330 PM
NO-CDE 1000 PM DMN 5 mM
Control
NO-CDE 100 PM NO-CDE 330 pM NO-CDE 1000 PM DMN 5 mM
Rat hepatocytes
26.6 + 2.3 (Kc ~0.0238) 30.9 i 3.9 0.0046
38.2 f 3.0 0.0132 56.7 A I 1.4 0.0406 71.8* 8.5 0.0736
76.0+ 8.1 0.0863
Human hepatocytes (Case 2)
32.9 i- 2.7 (Kc= 0.0307) 49.4 f 2.0 0.0217 61.0t 3.3 0.04 I7 62.8 f 3.7 0.0454 54.9 f 3.9 0.0306
-co.002 -co.002 <0.002 <0.002 -co.002
=0.1 =o. 1 =O.l =o. 1
a Significance level was determined by the use of the nonparametric Wilcoxon two-sample Iwo-tailed test (Riimke and De Jonge, 1964).
N-NITROSDCHLORDIAZEPOXIDE GENOTOXICITY IN MAMMALIAN CELLS 485
TABLE 4
DNA REPAIR SYNTHESIS IN RAT AND HUMAN HEPATWYTE PRIMARY CULTURES AFTER 20-hr EXPOSURETO NO-CDE
Treatment conditions No. of net nuclear grains (mean t SD)
Rat hepatocytes Expt I Control 2.2 f 2.9 (20) NO-CDE 33 /.LM 5.7 k 4.2* (56)
NO-CDE 100 /.LM 12.0 It 9.5* (75)
NO-CDE 330 +M 32.41 17.1*(96) NO-CDE 1000 #M 37.5 2 17.7* (100)
DMN 5 mM 31.5 2 l&1*(98)
Human hepatocytes Case 1 Control 1.1 + 3.8 (19) NO-CDE 33 fiM 3.5 t 3.7* (43) NO-CDE 100 /.LM 7.4 f 12.2* (69)
NO-CDE 330 HIM 10.5 ? 5.1* (90) NO-CDE 1000 fiM 31.0, 15.9*(97) DMN 5 mM 9.0 * 4.5* (87)
Expt 2 Expt 3 -0.4 + 3. I (6) 0.2 i 3.3 (8)
3.5 +- 3.5* (32) 4.1 f 4.2* (45) 3.1 r 2.9* (29) 7.0 2 5.6* (67)
4.2 k 3.0* (45) 13.3 + 7.2* (92) 14.6 _t 7.2* (97) 31.9* 14.1*(100) 8.5 F 5.7* (79) 18.0+ 11.2*(94)
Case 2 5.5 ?I 8.6 (42)
-
20.1 + 12.7* (92) 27.1 i 17.1* (90) 45.3 f 19.1* (94) 17.8 + 12.0* (97)
a Net nuclear grain counts of duplicate autoradiographs. Grain counts include cells with no nuclear labeling en- countered in the 50 cells counted for each slide. Numbers in parentheses are the percentage of cells with five or more nuclear grains.
* p f 0.001 vs controls; significance level was determined by the use ofthe Student I test (two-tailed).
these variations are in the order of those re- ported by others (Williams, 1977; Strom et al., 1983). The constant occurrence of DNA damage and repair in rat and human hepato- cytes exposed to the procarcinogen DMN, used as a positive control, demonstrates their metabolic competence.
Mutagenic Activity
Data listed in Table 5 indicate that a I-hr exposure to NO-CDE in the absence of a met- abolic system produced a dose-dependent in- crease in the number of TG’ clones. At the highest concentration tested (330 PM) the av- erage mutation frequency was approximately 23 times higher than that in controls. NMU, used as positive control, yielded, at 1 mM concentration, a lower frequency of TG’ vari- ants.
DISCUSSION
The results of this work indicate that the reaction of chlordiazepoxide with nitrite, car-
ried out in HCl aqueous solution, at pH rang- ing from 0.5 to 5.0, produces NO-CDE. With a molar ratio drug/NaNO* of l/2.4, the maxi-
TABLE 5
INDUCTION 0F MUTATION pro 6-THIOGUANINE RESISTANCE IN V79 CELLS BY I-hr EXPOSURE TO NO- CDE
Treatment conditions TG’ mutants/ lo6 survivors”
Control 18.4 t 0.4 NO-CDE 10 ).LM 25.1 zk 2.6 (71-88Yio)”
NO-CDE 33 /.tM 30.0 t 4.5 (62-7610) NO-CDE 100 PM 105.8 + 51.6* (35-55s) NO-CDE 330 pM 415.6 + 84.0* (28-4310)
NMU 300 &LM 46.5 rt_ 5.3* (82-88%>) NMU 1000 LLM 130.3 + 33.3* (50-72s)
u Values are the means + SD of data obtained from three independent experiments.
’ Numbers in parenthesis indicate relative survival range.
* p < 0.01 vs controls; the significance level was deter- mined according to Kastenbaum and Bowman ( 1970).

486 BRAMBILLA ET AL.
ma1 yield of NO-CDE (55% of the theoretical one) was obtained at pH 3.5.
NO-CDE concentrations ranging from 33 to 1000 pM were found to produce dose-de- pendent genotoxic effects in some short-term tests routinely employed to evaluate the po- tential carcinogenic activity of chemical agents. DNA single-strand breaks were ob- served in V79 cells; the repair of these lesions was largely incomplete after 4X hr, and their promutagenic character was demonstrated by the induction of mutation to TG’ in the same cells. Both DNA fragmentation and un- scheduled DNA synthesis were detected in rat hepatocytes, and it is worth noting that NO- CDE exerted similar effects in primary cul- tures of liver cells obtained from two human donors. In contrast, any evidence of geno- toxic effects (results not shown in the tables) was absent in parallel assays carried out with the parent compound chlordiazepoxide up to the maximal soluble concentration (1 mM).
Since a satisfactory correlation has been found to exist between the tumorigenic activ- ity and the capabilities of inducing DNA frag- mentation (Swenberg et al.. 1976: Sina et [II., 1983) DNA repair (Williams, 1977; Wil- liams et a/., 1982) and somatic mutations (Bartsch et al., 1980; Bradley ct a/., 198 1 ), our findings, which are in good agreement with the previously acquired evidence of NO-CDE genotoxicity (see Introduction), contribute to support the hypothesis that this N-nitroso compound may initiate a carcinogenic pro- cess. This hypothesis is strengthened by the comparison of the genotoxic potency of NO- CDE with those of the known carcinogens NMU and DMN. In fact, the potency of NO- CDE was of the same order of that of NMU in assays performed on V79 cells and greater than that of DMN in rat and human hepato- cytes. Moreover. the DNA-damaging and mutagenic activities exerted by NO-CDE on V79 cells in the absence of a metabolic sys- tem indicate that it behaves as an activation- independent agent, and in turn this suggests that it may act directly as an ultimate carcino-
gen in the site of its production, i.e., at the level of gastric mucosa.
The usual therapeutic dosage of chlordiaz- epoxide is 25 mg/day, administered in di- vided doses without observing a temporal re- lation with meals. The concentration of ni- trite in human gastric juice has been found to vary from 0 to 3.2 PM in fasting healthy volunteers (Raisfeld-Danse and Chen, 1983) and to depend on gastric pH in patients with stomach pathology, ranging from 1.7 to 4.6 pM at pH < 3 and from 30 to 300 pM at pH > 5 (Kyrtopoulos et al.. 1985). High nitrite concentrations are normally present in the achlorhydric stomach with the highest level reported being 600 yM (Tannenbaum et al.. 1979). After a meal the gastric concentration of nitrite seems to depend essentially on the salivary contribution. which in turn is related to the nitrate and nitrite concentration of the diet (Klein el a/., 1978). For U .S. residents the estimated amounts of nitrate and nitrite in- gested daily correspond to 99.8 and 1 1.2 mg, respectively (Tannenbaum et al., 1977). Wal- ters ( 1980) demonstrated that after consump- tion of meals containing 15.9 mg of nitrite the gastric juice nitrite concentration was raised to about 300 pM. The reaction of 25 mg of chlordiazepoxide with 2 mg of NaN02, car- ried out at 37°C in 40 ml of HCl solution (pH 3.5) yielded about 0.6 mg of NO-CDE (2.5% ofthe theoretical yield), which corresponds to an -50 PM concentration of this N-nitroso derivative in the reaction mixture. Therefore, in conditions very similar to the possible gas- tric environment of a patient taking chlordi- azepoxide, NO-CDE is formed in a concen- tration of the order of those found to induce DNA damage and mutations in mammalian cells. In this respect, it is noteworthy that DMN. which is about five times less potent than NO-CDE in producing DNA damage and repair in both rat and human hepato- cytes, was shown to induce rat liver cell carci- nomas in a dose-related frequency when ad- ministered chronically with the diet in dos- ages ranging from 0.2 to 5 mg/kg of body weight (Terracini et al.. 1967).

IV-NITROSOCHLORDIAZEPOXIDE GENOTOXICITY IN MAMMALIAN CELLS 487
Nitrosation reactions depend on the rela- tive concentrations of substrate, catalysts, and inhibitors and are influenced by the in- homogeneity of the stomach contents, by pH and by other factors. Thus our data do not allow any prediction of NO-CDE yields in pa- tients ingesting chlordiazepoxide. However, they suggest that at least in some individuals NO-CDE might be formed in amounts sus- ceptible of inducing genotoxic effects. This might occur, for example, in patients treated with gastric antacids and/or H,-blocking agents which should enhance chlordiazepox- ide nitrosation either directly by raising the pH of gastric environment to values favoring NO-CDE formation or indirectly by allowing the development of nitrate-reducing bacterial species (Reed et al., 198 1; Kyrtopoulos et al., 1985). A short-term screening study of 6257 users of chlordiazepoxide, covering a follow- up period that was, at most, 7; years from the first recorded dispensing of the drug, showed no evidence of excess of any cancer (Fried- man and Ury, 1980). However, taking into account that the latency period for cancer de- velopment after exposure to carcinogens is often measured in decades from first expo- sure, the results of this analysis should be con- sidered inadequate to provide a definitive proof of noncarcinogenicity in humans.
REFERENCES
ANDREWS. A. W., FORNWALD, J. A., AND LIJINSKY. W. ( 1980). Nitrosation and mutagenicity of some amine
drugs. Tovicol. Appl. Phurmacol. 52,237-244. BARTSCH, H., MALAVEILLE, C.. CAMUS, A.-M., MAR-
TEL-PLANCHE. G.. BRUN, G., HAUTEFEUILLE, A., SA-
BADIE, N.. BARBIN, A.. KUROKI, T.. DREVON, C., PIC- COLI, C., AND MONTESANO, R. (1980). Validation and comparative studies on 180 chemicals with S. typhi- murium strains and V79 Chinese hamster cells in the presence of various metabolizing systems. Mzkzt. Res. 76, l-50.
BARTSCH, H., AND MONTESANO, R. (1984). Relevance
of nitrosamines to human cancer. Curcinogenesis 5, 1381-1393.
BLACKWELL. B. (1973). Psychotropic drugs in use today.
J. ‘4mer. Med. .4ssoc. 225, 1637- 164 1.
BRADLEY, M. 0.. BHUYAN. B., FRANCIS, M. C.. LANGENBACH, R., PETERSON, A., AND HUBERMAN. E. (198 1). Mutagenesis by chemical agents in V79 Chi-
nese hamster cells: A review and analysis of the litera- ture. A report of the Gene-Tox Program. Mutat. Res. 87,81-142.
BRAMBILLA, G. (1985). Genotoxic effects of drug/nitrite
interaction products: Evidence for the need of risk as- sessment. Pharmacol. Re.T. Commun. 17,307-321.
BRAMBILLA, G., CAJELLI, E., FINOLLO. R., MAURA. A..
PINO. A., AND ROBBIANO, L. (1985). Formation of DNA-damaging nitroso compounds by interaction of drugs with nitrite. A preliminary screening for detect-
ing potentially hazardous drugs. J. To-UCO/. Environ. Health 15, 1-24.
BRAMBILLA. G., CAVANNA, M., PARODI. S., SCIAB,& L.,
PINO, A., AND ROBBIANO, L. (1978). DNA damage in liver, colon, stomach, lung and kidney of BALB/c mice treated with I .2-dimethylhydrazine. Int. J. Can- cer 22, 174- 180.
CAJELLI, E.. CANONERO. R., MARTELLI, A., AND BRAM- BILLA. G. (1987). Methylglyoxal-induced mutation to 6-thioguanine resistance in V79 cells. Mutat. Res. 190,
47-50.
FRIEDMAN, G. D., AND URY, H. K. (1980). Initial screening for carcinogenicity ofcommonly used drugs.
J. Natl. Cancer Insr. 65,723-733. International Agency for Research on Cancer (IARC)
(1980a). IARC Monographs on the Evaluation of the Carcinogenic Risk of Chemicals IO Humans, Vol. 24, Some Pharmaceutical Drugs, pp. 297-314. IARC, Lyon, France.
International Agency for Research on Cancer (IARC)
( 1980b). N-Nitroso Compounds: Analysis, Formation. and Occurrence. IARC Scientific Publication No. 3 1. IARC, Lyon, France.
KASTENBAUM. M. A., AND BOWMAN, K. 0. (1970). Ta-
bles for determining the statistical significance of mu- tation frequencies. Mutat. Res. 9.527-549.
KLEIN. D., GACONNET, N., POULLAIN, B., AND DERBY, G. ( 1978). Possibility of nitrosation during human di- gestion. I. Evaluation of precursor and affecting factors
in rlivo. II. Effect of food intake on the reaction. In L4RC Scientzjk Publications (E. A. Walker, M. Cas- tegnaro, L. Gricciute. and R. E. Lyle. Eds.). Vol. 19.
pp. 485-494. IARC. Lyon, France.
KOHN. K. W.. ERICKSON, L. C.. EWIG, R. A. G.. AND FRIEDMAN, C. A. (I 976). Fractionation of DNA from mammalian cells byalkalineelution. Biochemistry 15, 4629-4637.
KRAFF~, S., AND ONKEN, A. (1976). Induction of mi- totic gene conversion in Saccharomvces cerevisiae with chlordiazepoxide and its N-nitroso derivative in a host-mediated assay. Mutat. Res. 34,333-336.
KYRTOPOULOS, S. A., DASKALAKIS, G.. LEGAKIS, N. I., KONIDARIS, N., PSARROU, E.. BONATSOS, G.. Go- LEMATIS. B.. LAKIOTIS, G., BLIOURAS. N.. AND OUT-

488 BRAMBILLA ET AL.
RAM, J. R. (1985). Studies in gastric carcinogenesis. II. Absence of elevated concentrations of N-nitroso com- pounds in the gastric juice of Greek hypochlorhydric individuals. Carcinogenesis 6, 1135-l 140.
LIJINSKY, W., AND TAYLOR, H. W. (1977). Nitrosa- mines and their precursors in foods. In “Origins OfHu- lnan Cancer” (H. H. Hiatt, J. D. Watson, and J. A. Winstein, Eds.). pp. 1579-1580. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY.
MIRVISH, S. S. (1975). Formation of N-nitroso com- pounds: Chemistry, kinetics and in vivo occurrence. Toxicol. Appl. Pharmacol. 31,325-35 1.
MYHR, B. C., AND DI PAOLO, J. A. (1978). Mutagenesis by N-acetoxy-2-acetyl aminofluorene of Chinese ham- ster V79 cells is unaffected by caffeine. Chem. Biol. In- teract. 21, l-18.
PREDA. N., POPA, L.. AND GALEA. V. (1976). N-Nitroso compound formation by chlordiazepoxide and nitrite interaction in vitro and in vivo: Protective action of ascorbic acid. In Environmental N-Nitroso Com- pounds Analysis and Formation, IARC Scientific Pub- lication No. 14. pp. 301-3 14. IARC, Lyon, France.
RAISFELD-DANSE, I. H., AND CHEN, J. (1983). Drug in- teractions. III. Formation of nitrosamines from thera- peutic drugs. Formation, mutagenic properties and safety assessment of propranolol hydrochloride with respect to the intragastric formation of N-nitrosopro- pranolol under conditions found in patients. J. Phar- macol. Exp. Ther. 225,7 13-7 19.
REED, P. I.. HAINES, K., SMITH, P. L. R., AND HOUSE. F. R. (1981). Effect of cimetidine on gastric juice N- nitrosamine concentration. Lancet 2,553-555.
ROPER, P. R.. AND DREWINKO, B. (1976). Comparison of in vitro methods to determine drug-induced cell le- thality. Cancer Res. 36,2 182-2 188.
R~~MKE, C. L.. AND DE JONGE, H. (1964). Design, statis- tical analysis and interpretation. In Evaluation qfDrug Activities: Pharmacometrics (D. R. Laurence and A. L. Bacharach, Eds.). Vol. 1, pp. 80-82. Academic Press, New York.
SINA, J. F., BEAN, C. L., DYSART, G. R., TAYLOR, V. I.. AND BRADLEY, M. 0. (1983). Evaluation of the alka- line elution/rat hepatocyte assay as a predictor ofcarci- nogenic/mutagenic potential. Mutat. Res. 113, 357-
391. STROM, S. C.. JIRTLE. R. L., JONES, R. S.. NOVICKI,
D. L., ROSENBERG, M. R.. NOVOTNY, A., IRONS, G..
MCLAIN, J. R., AND MICHALOPOULOS, G. (1982). Iso- lation, culture, and transplantation of human hepato- cytes. J. Natl. Cancer Inst. 68,77 l-778.
STROM, S. C., NOVICKI, D. L.. NOVOTNY, A., JIRTLE, R. L., AND MICHALOPOULOS, G. (1983). Human hep- atocyte-mediated mutagenesis and DNA repair activ- ity. Carcinogenesis 4,683-686.
SWENBERG. J. A., PETZOLD, G. L., AND HARBACK, P. R. (1976). In vitro DNA damage/alkaline elution assay for predicting carcinogenic potential. Biochem. Bio- ph.vs. Res. Commun. 12,732-738.
TAKEDA. Y., AND KANAYA, H. (1981). Formation of ni- troso compounds and mutagens from tranquilizers by drug/nitrite interaction. Cancer Lett. 12,8 l-86.
TANNENBAUM, S. R., ARCHER. M. C.. WISHNOK, J. S., CORREA. P.. CUELLO, C., AND HAENSZEL, W. (1977). Nitrate and etiology ofgastric cancer. In Origins ofHu- man Cancer (H. H. Hiatt, J. D. Watson, and J. A. Winstein. Eds.), pp. 1609- 1625. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY.
TANNENBAUM, S. R.. MORAN, D., RAND, W., CUELLO, C., AND CORREA, P. (1979). Gastric cancer in Colum- bia. IV. Nitrite and other ions in gastric contents of residents from high-risk region. J. Natl. Cancer Inst. 62,9- 12.
TERRACINI, B., MAGEE. P. N., AND BARNES, J. M. (1967). Hepatic pathology in rats on low dietary levels of dimethylnitrosamine. Brit. J. Cuncer 21,559-565.
WALSER. A., FRYER. R. I., STERNBACH, L. H.. AND ARCHER, M. ( 1974). Quinazolines and 1,4-benzodiaz- epines. LXV ( I ) Some transformations of chlordiazep- oxide. J. Heterocycl. Chem. 11,6 19-62 1.
WALTERS, C. L. (1980). The exposure of humans to ni- trite. Oncology31,289-296.
WILLIAMS. G. M. (1977). The detection of chemical car- cinogens by unscheduled DNA synthesis in rat liver primary cell cultures. Cancer Res. 37, 1845- 185 1,
WILLIAMS, G. M., LASPIA, M. F.. AND DUNKEL, V. C. (1982). Reliability of the hepatocyte primary culture/ DNA repair test in testing of coded carcinogens and noncarcinogens. Mutat. Res. 97,359-370.
WILLIAMS, J. R., LITTLE, J. B., AND SHIPLEY, W. U. ( 1974). Association of mammalian cell death with a specific endonucleolytic degradation of DNA. Nature (London) 252,754-755.







